Human Biology 534 Introduction to Virology and Human Retroviruses 26/27 March 2001 James I. Mullins, Ph.D., Professor and Chairman, Department of Microbiology, [email protected] Spring Quarter, 2001 - PowerPoint PPT Presentation
Human Biology 534 Introduction to Virology and Human Retroviruses 26/27 March 2001 James I. Mullins, Ph.D., Professor and Chairman, Department of Microbiology, [email protected]Spring Quarter, 2001 Copies of lectures can be downloaded (pdf) from: http:// ubik . microbiol . washington . edu /Index.html KEYWORDS are in bolded red
Transcript
Human Biology 534
Introduction to Virology and Human Retroviruses
26/27 March 2001
James I. Mullins, Ph.D., Professor and Chairman, Department of Microbiology, [email protected]
Spring Quarter, 2001
Copies of lectures can be downloaded (pdf) from: http://ubik.microbiol.washington.edu/Index.html
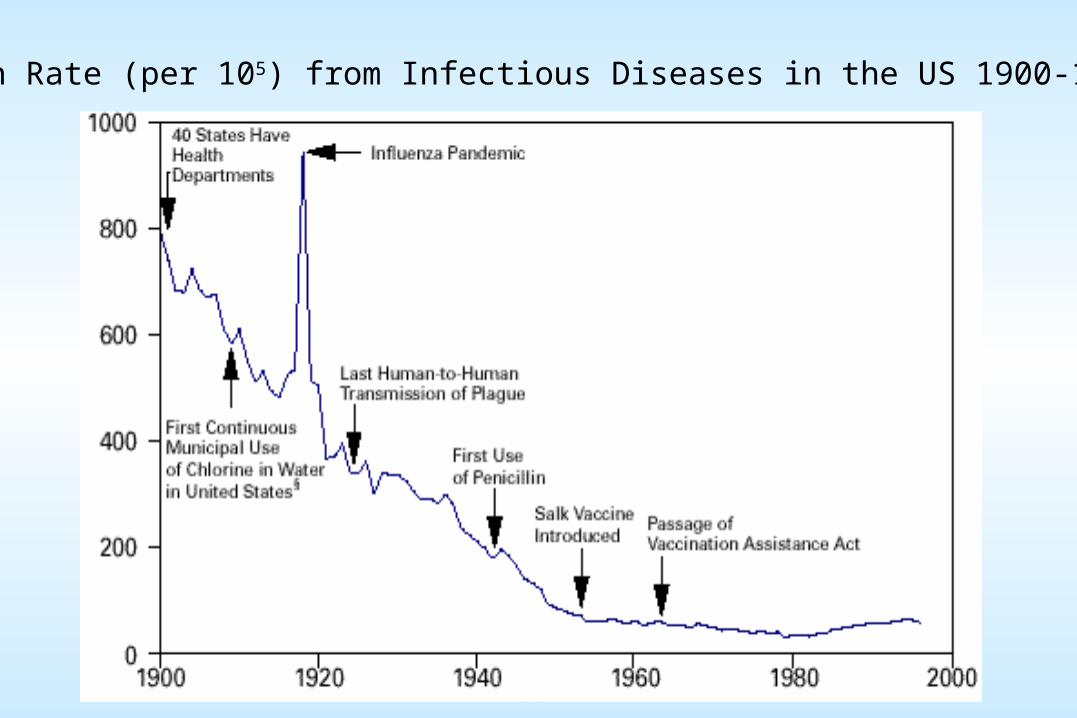
Death Rate (per 105) from Infectious Diseases in the US 1900-1996
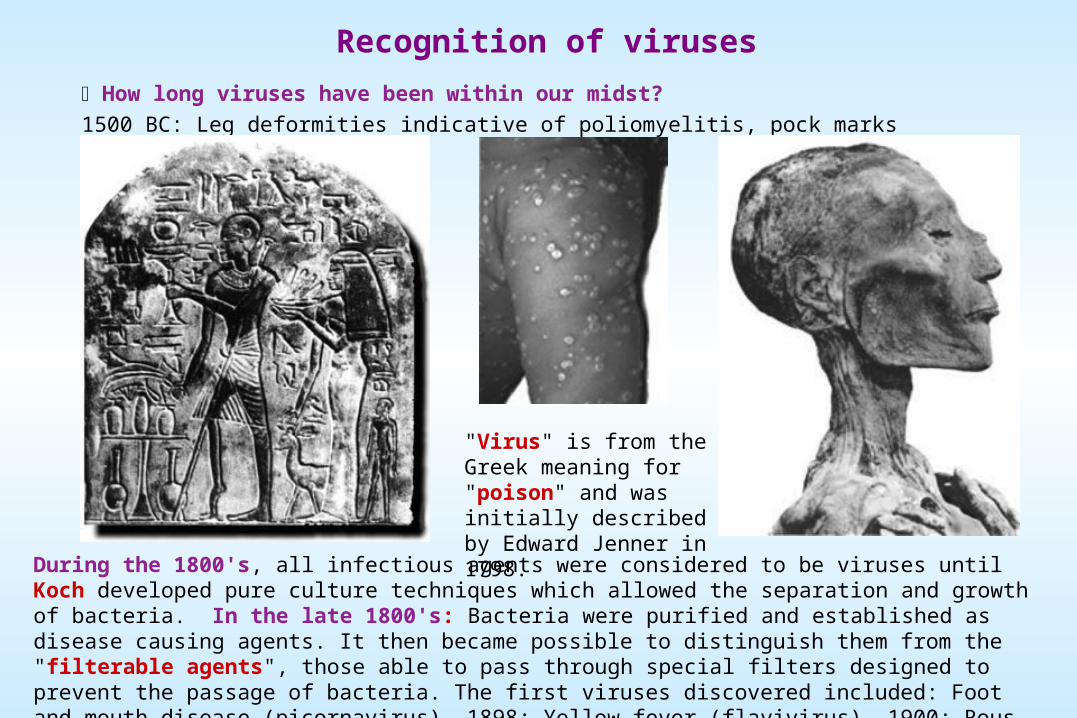
Recognition of viruses
How long viruses have been within our midst?
1500 BC: Leg deformities indicative of poliomyelitis, pock marks indicative of smallpox.
During the 1800's, all infectious agents were considered to be viruses until Koch developed pure culture techniques which allowed the separation and growth of bacteria. In the late 1800's: Bacteria were purified and established as disease causing agents. It then became possible to distinguish them from the "filterable agents", those able to pass through special filters designed to prevent the passage of bacteria. The first viruses discovered included: Foot and mouth disease (picornavirus), 1898; Yellow fever (flavivirus), 1900; Rous sarcoma virus (oncogenic retrovirus), 1906
"Virus" is from the Greek meaning for "poison" and was initially described by Edward Jenner in 1798.
Viral diseases have played a major role in human history:
Over the past 1000 years: Smallpox and measles were brought to North and South America by early European explorers/conquerers. These diseases, for which the native American populations had no acquired (partial) immunity, killed large fractions of the populations, and were a major factor in the decimation of these societies.
Over the past 100 years: A newly emerged strain of influenza killed 20 million people in 1918-1919 in the immediate aftermath of World War I (It was the most lethal combatant of the war). A decade later, polio became one of the most feared infections of children and young adults (Franklin D. Roosevelt, the U.S. President throughout the Depression and World War II, had polio).
As the last century entered its final 20 years, a new >90% lethal virus, HIV, spread rapidly around the world via body fluid transmission.
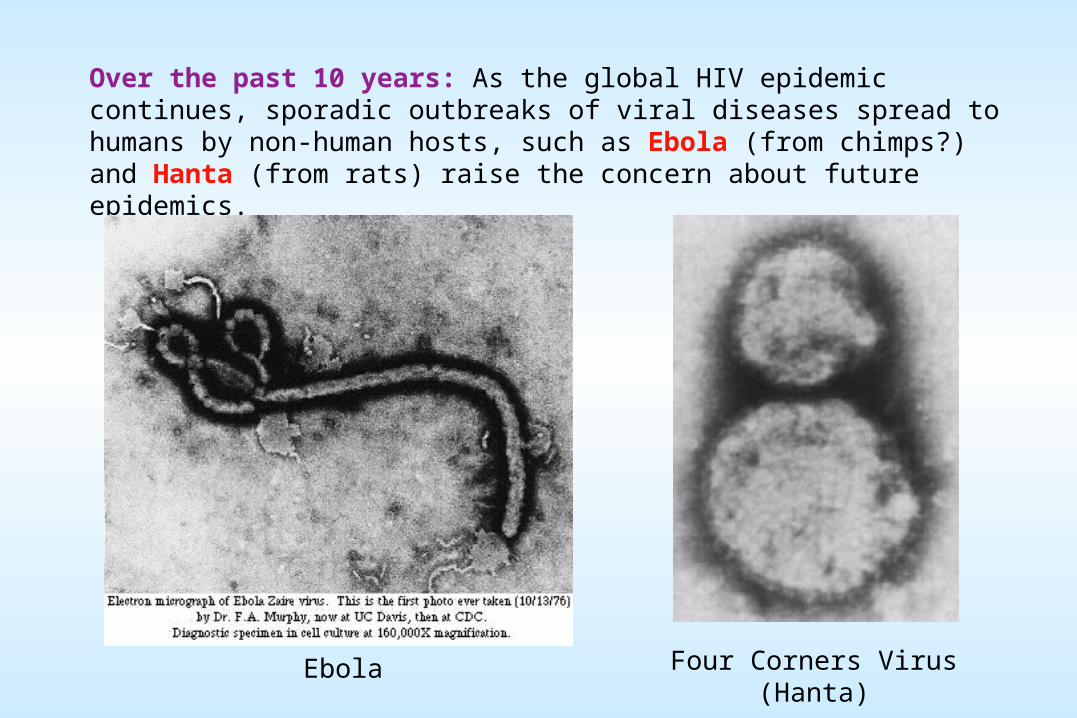
Over the past 10 years: As the global HIV epidemic continues, sporadic outbreaks of viral diseases spread to humans by non-human hosts, such as Ebola (from chimps?) and Hanta (from rats) raise the concern about future epidemics.
Four Corners Virus (Hanta)Ebola
THE DISCIPLINE OF VIROLOGY
The study of virology inherently involves a merging together of what has traditionally been thought of as two separate "kinds" of science: basic and applied. We want to figure out how viruses are transmitted, how they replicate, and how the host organism responds. We also want to figure out how to prevent transmission, how to interfere with virus replication, and how we can confer immunity on the host. The "applied" follows from, and is dependent upon, the "basic" in a quite direct way. Virology as it is studied today, is therefore an outgrowth of research in:
Infectious diseases - which provides recognition of viral pathogens and their sequelae, and
Molecular Biology- because of the usefulness of viruses as probes of cellular metabolic processes and as vectors with potential for gene therapy.
Mouse primary spleen cells transduced with a GFP (green fluorescent protein) -retrovirus vector
WHERE WE STAND IN 2001: PREVENTING • CONTROLING • CURING VIRAL DISEASES
Smallpox: effective vaccine; this is the only viral disease that has been wiped out worldwide
Measles: effective vaccine since 1963; this disease could be eliminated with a world-wide effort
Influenza: effective strain-specific vaccine, but new variant strains emerge periodically
Polio: effective vaccine; will soon be the second viral disease wiped out
HIV: no vaccine; effective drugs, but they are costly and toxic, plus resistant strains appear. World-wide spread continues via intimate contact. 50 million infected thus far
Ebola: no vaccine; important host species unknown (found recently in chimps and rodents); outbreaks controllable because people die quickly and human-human transmission is via blood
Hanta: no vaccine; rodent host; easy transmission to humans, but outbreaks controllable
We also share the world, and our bodies, with viruses that cause hepatitis, respiratory disease, mononucleosis, diarrhea, genital warts, genital herpes, and some forms of cancer
WHAT IS A VIRUS?
Viruses may be defined as acellular organisms whose genomes consist of nucleic acid, and which obligately replicate inside host cells using host metabolic machinery to different extents, to form a pool of components which assemble into particles called Virions.
A virus differs from a cell in three fundamental ways:i A virus usually has only a single type of nucleic acid serving as its genetic material. This can be single or double stranded DNA or RNA;ii Viruses contain no enzymes of energy metabolism, thus cannot make ATP;iii Viruses do not encode sufficient enzymatic machinery to synthesize their component macromolecules, specifically, no protein synthesis machinery.
Viruses cannot be grown on sterile media, but require the presence of specific host cells.
Viruses are distinguished from other obligate parasites, some of which are even simpler than viruses:
MYCOPLASMA: Small bacterium that grows only in complex medium or attached to eucaryotic cells.
CHLAMYDIA: Obligate intracellular bacterial parasite which depends on eucaryotic cell for energy.
PROTOZOA: Obligate intracellular parasite of eucaryotes that replicate within eucaryotic cells.
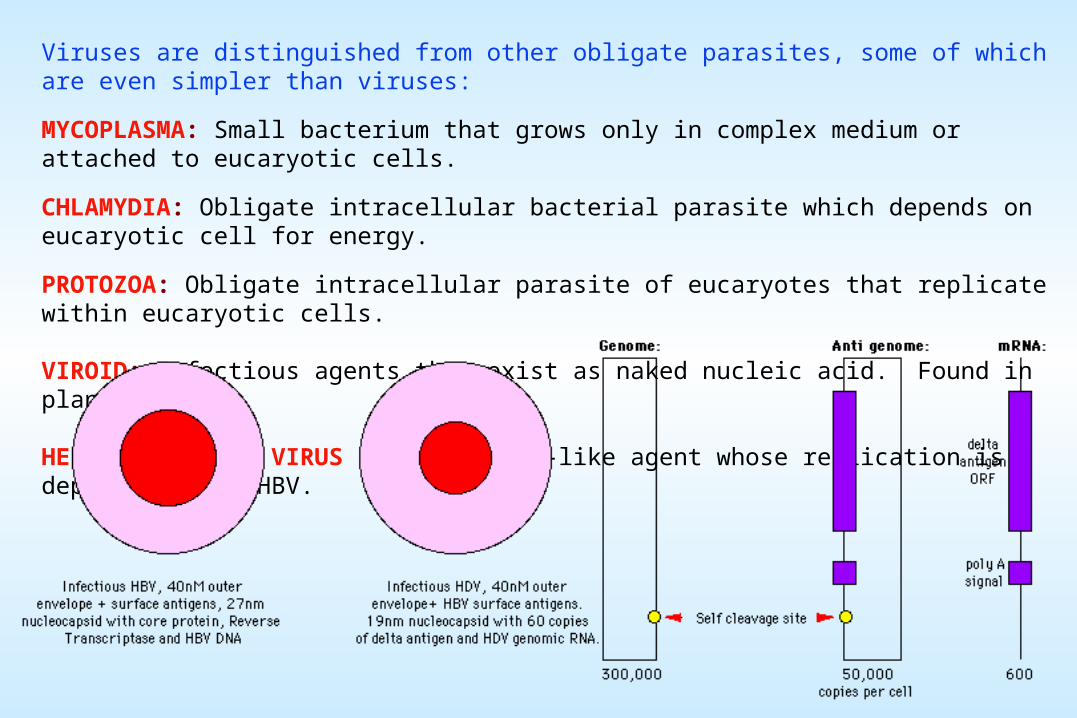
VIROID: Infectious agents that exist as naked nucleic acid. Found in plants.
HEPATITIS DELTA VIRUS (HDV): Viroid-like agent whose replication is dependent upon HBV.
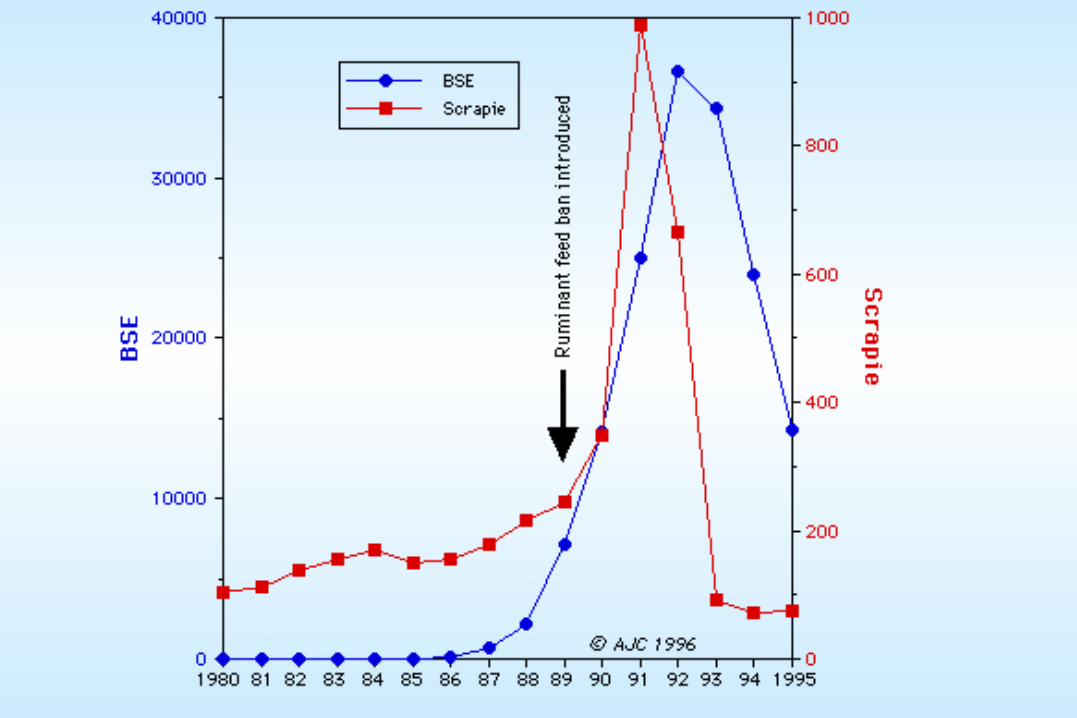
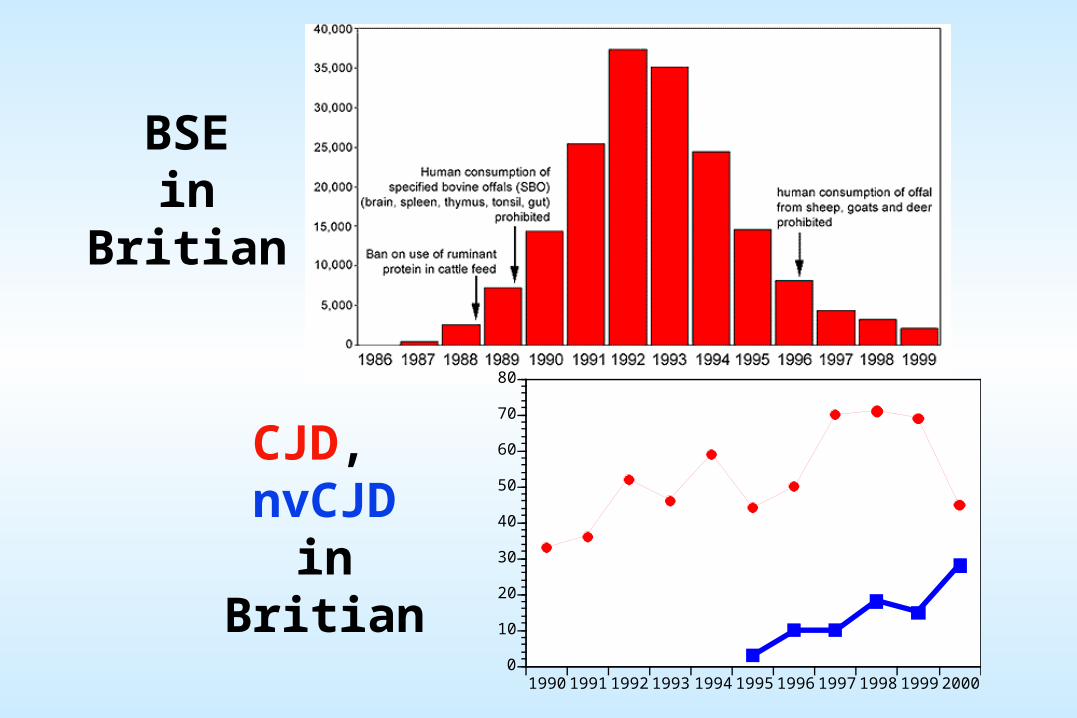
PRION (proteinacious infectious agent): Hypothesized identity of the unconventional slow viruses (such as the Kuru and Scrapie agents). No nucleic acid is known to be required for prion function. They are thought by many to consist solely of protein and perhaps lipids.
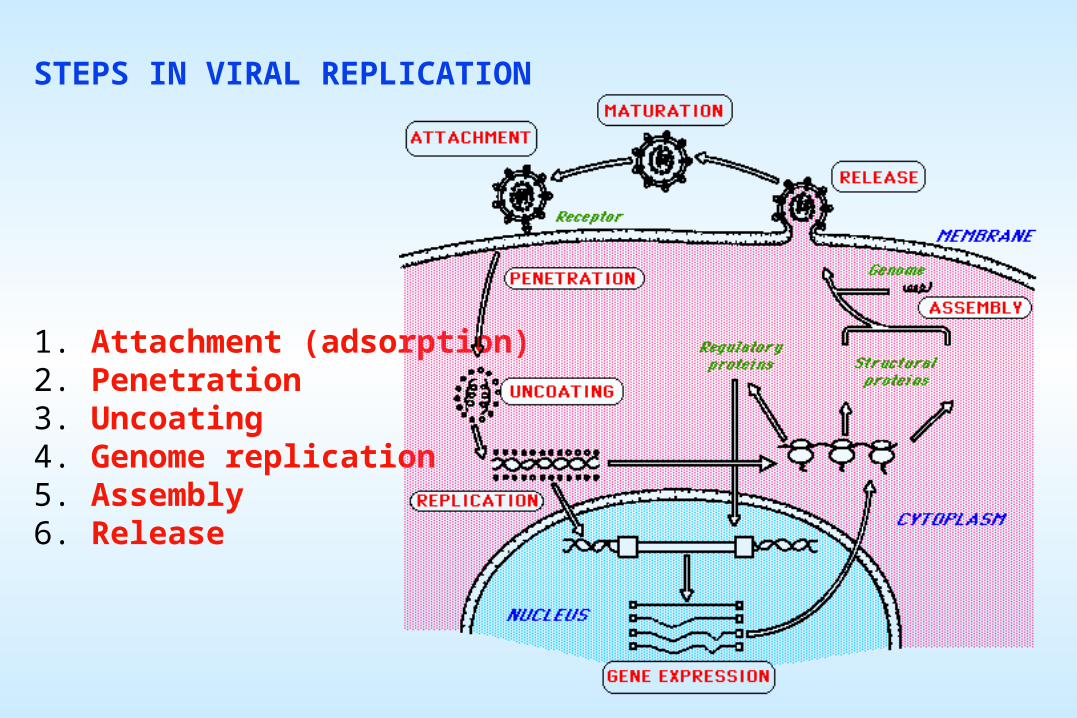
The first steps in the replication process for all viruses involve the attachment of the virus particle (the virion) onto the cell surface and then the entry of at least a portion of the particle (including the genome) into the cytoplasm. Both of these steps are specific, and are the main reason for the species-specificity of viruses.
For all viruses, some aspect of membrane disruption must occur to achieve entry. For enveloped viruses, this almost always involves membrane fusion between the viral envelope and a cell membrane (either the plasma membrane or an internal vesicle membrane). For non-enveloped virions, the viral surface protein(s) usually play a role in causing some very localized membrane disruption.
The details of attachment and entry for many viruses are still somewhat unclear, but there are some viruses for which the process has been studied in depth.
We will take a look today at three of these: Picornaviruses (poliovirus and "common cold" rhinoviruses), Influenza virus and HIV (retrovirus)
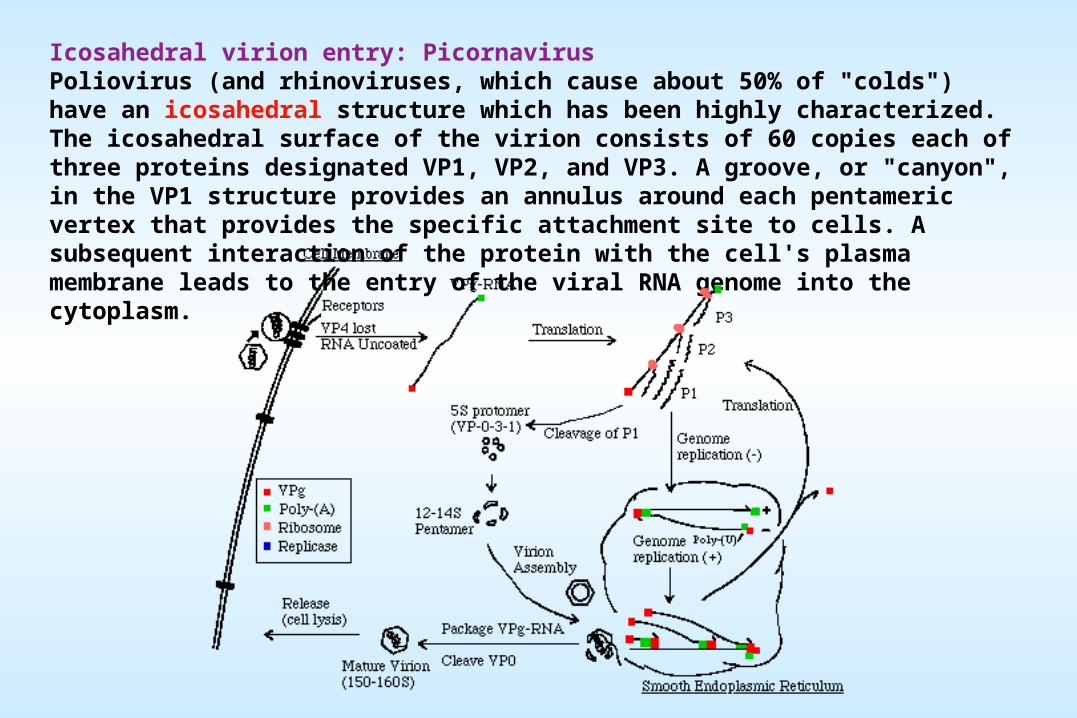
Icosahedral virion entry: PicornavirusPoliovirus (and rhinoviruses, which cause about 50% of "colds") have an icosahedral structure which has been highly characterized. The icosahedral surface of the virion consists of 60 copies each of three proteins designated VP1, VP2, and VP3. A groove, or "canyon", in the VP1 structure provides an annulus around each pentameric vertex that provides the specific attachment site to cells. A subsequent interaction of the protein with the cell's plasma membrane leads to the entry of the viral RNA genome into the cytoplasm.
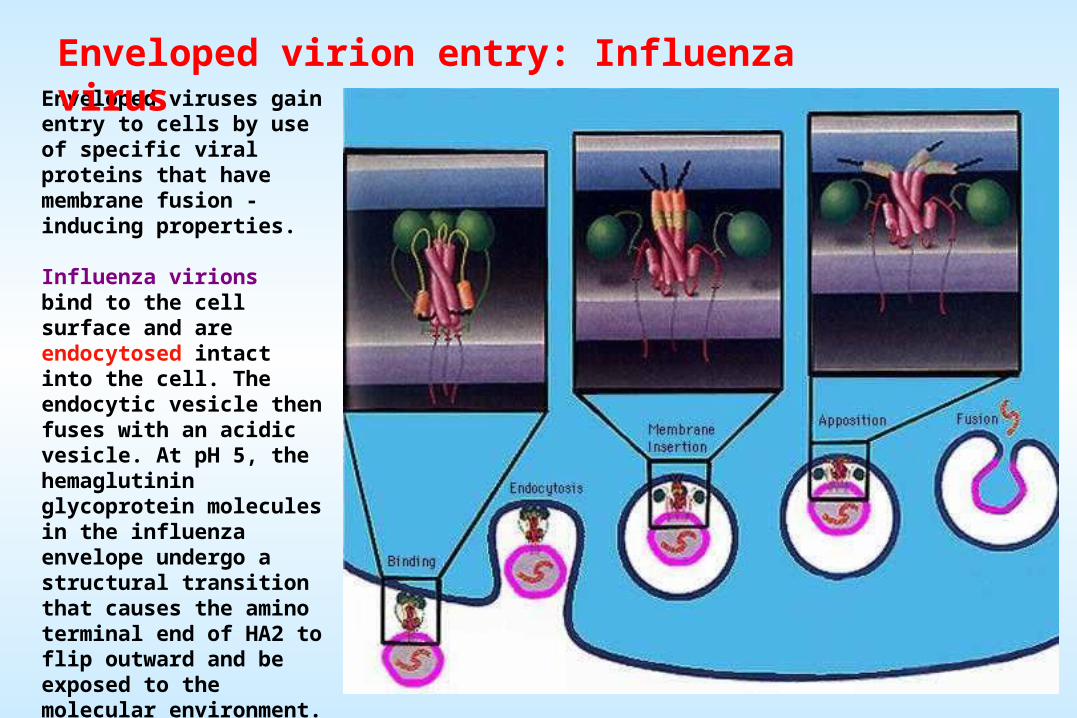
Enveloped viruses gain entry to cells by use of specific viral proteins that have membrane fusion - inducing properties.
Influenza virions bind to the cell surface and are endocytosed intact into the cell. The endocytic vesicle then fuses with an acidic vesicle. At pH 5, the hemaglutinin glycoprotein molecules in the influenza envelope undergo a structural transition that causes the amino terminal end of HA2 to flip outward and be exposed to the molecular environment. This highly hydrophobic segment interacts with the vesicle membrane and causes fusion. This fusion event dumps the viral genome into the cell's cytoplasm.
Enveloped virion entry: Influenza virus
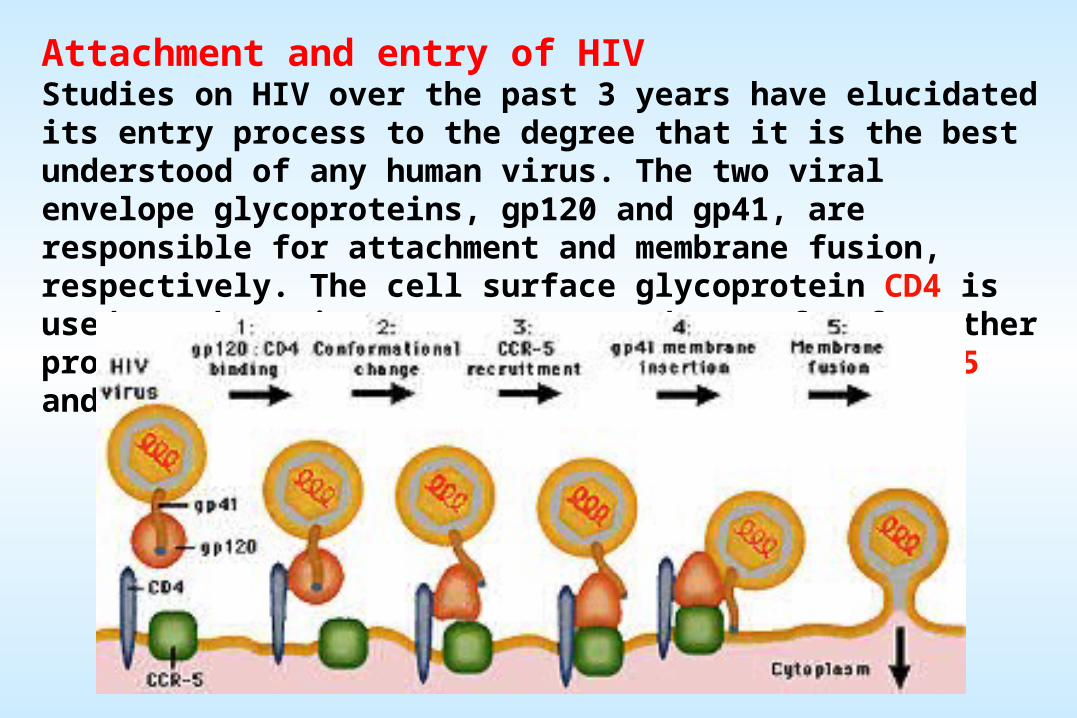
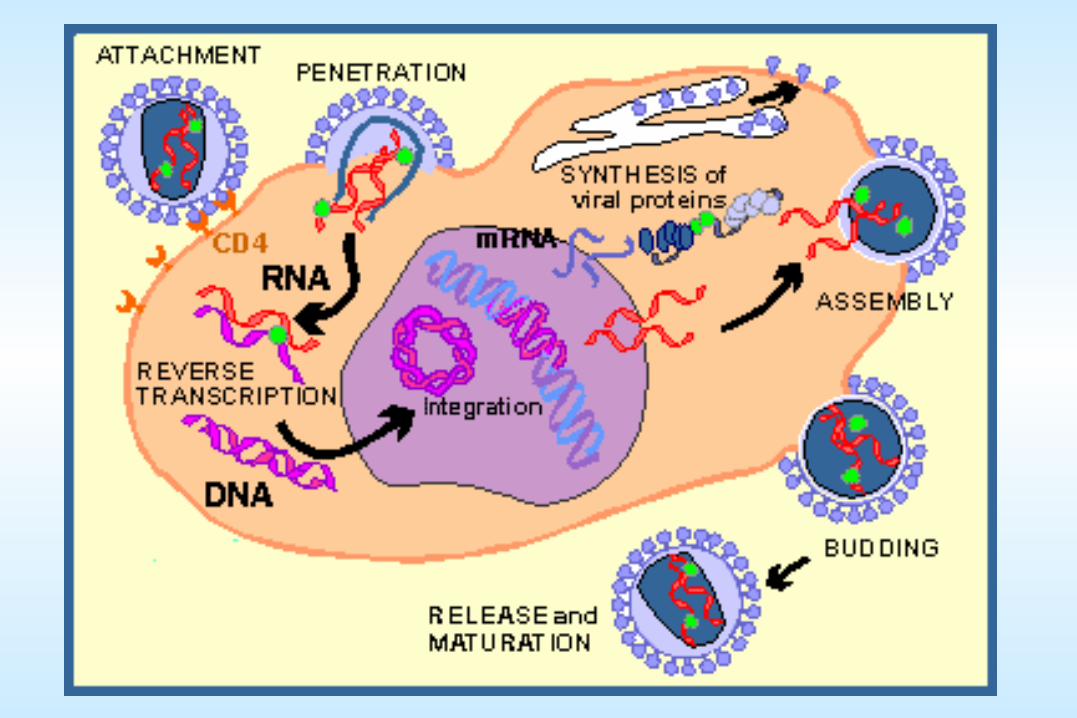
Attachment and entry of HIV Studies on HIV over the past 3 years have elucidated its entry process to the degree that it is the best understood of any human virus. The two viral envelope glycoproteins, gp120 and gp41, are responsible for attachment and membrane fusion, respectively. The cell surface glycoprotein CD4 is used as the primary receptor, and one of a few other proteins are used as co-receptors (typically CCR5 and CXCR4).
Information Flow in cells:DNA to DNA (Replication)
DNA to RNA (Transcription)RNA to Protein (Translation)
The Central Dogma
QuickTime™ and aGIF decompressor
are needed to see this picture.
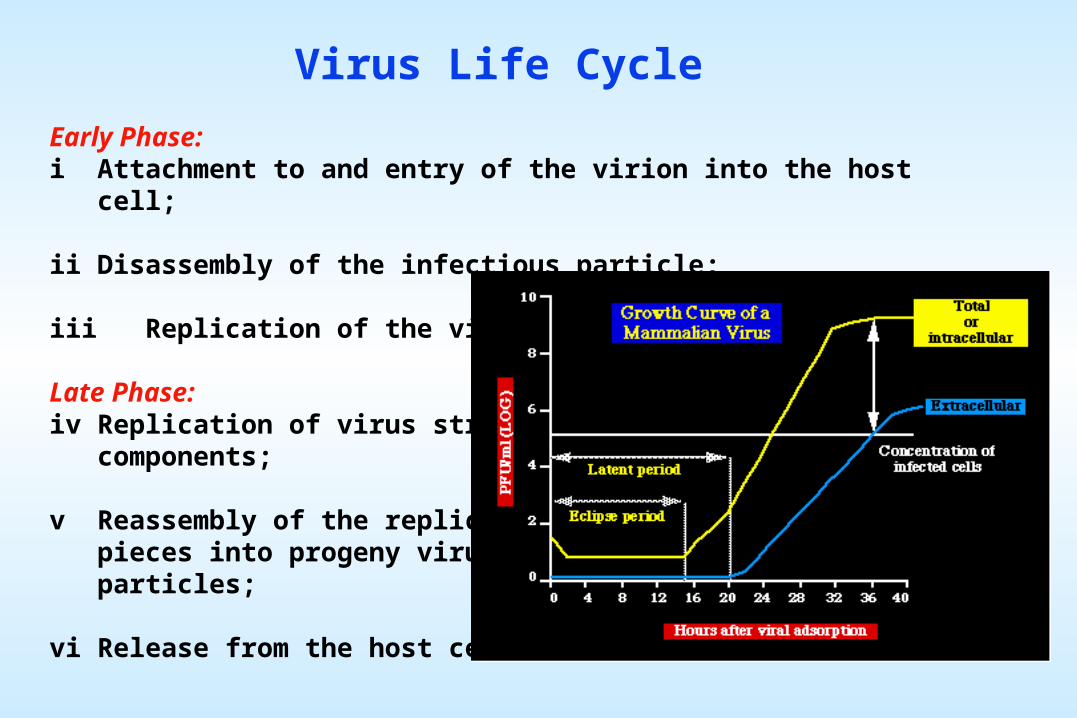
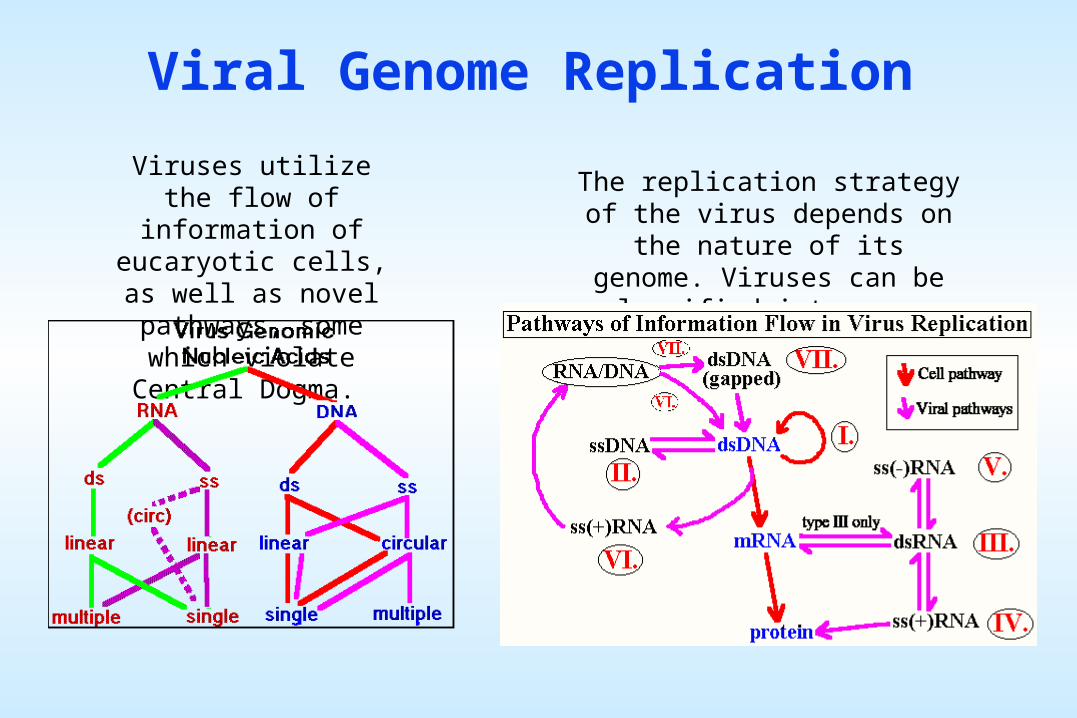
Viral Genome Replication
Viruses utilize the flow of information of eucaryotic
cells, as well as novel pathways, some which
violate Central Dogma.
The replication strategy of the virus depends on the nature of its
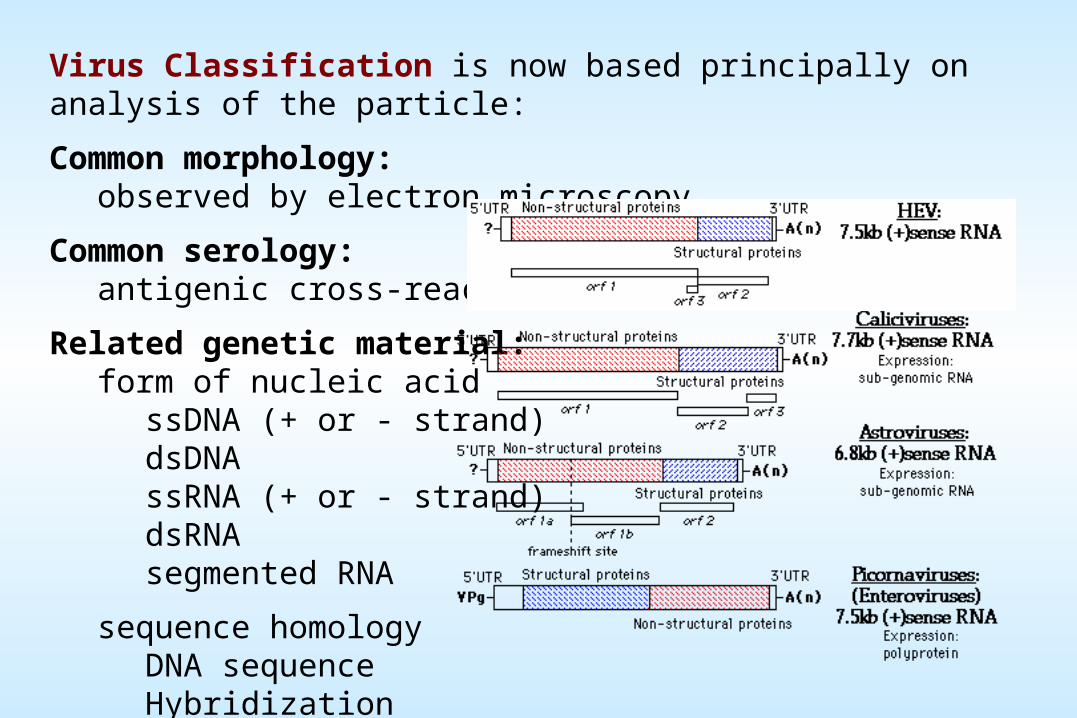
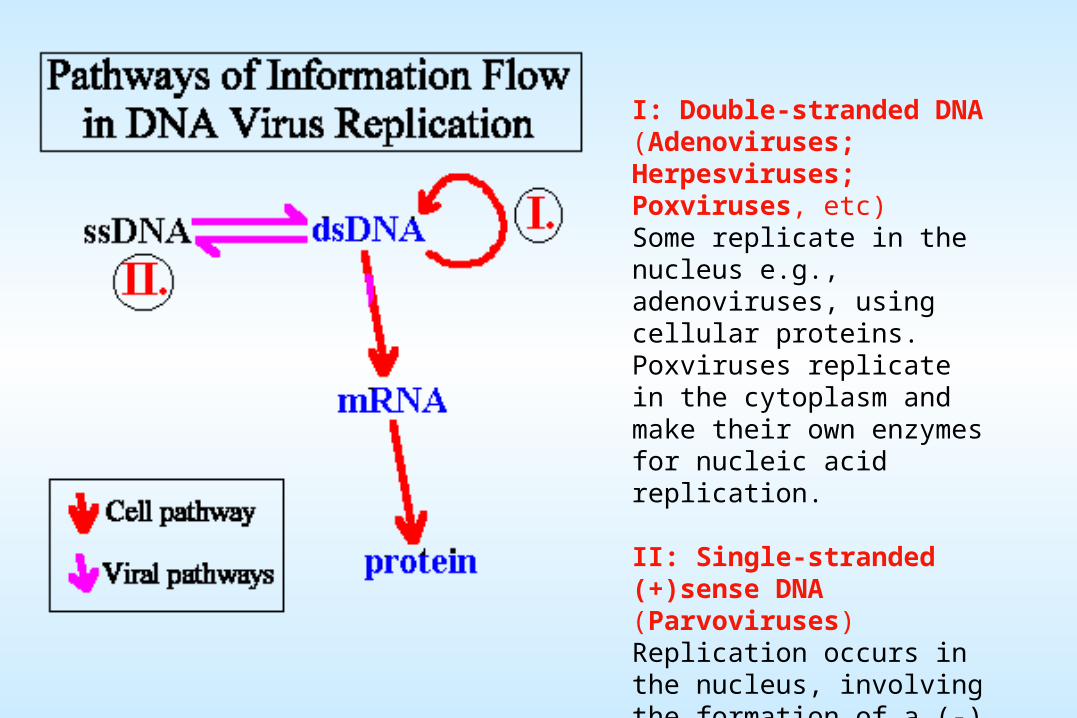
genome. Viruses can be classified into seven (arbitrary) groups:
I: Double-stranded DNA (Adenoviruses; Herpesviruses; Poxviruses, etc)Some replicate in the nucleus e.g., adenoviruses, using cellular proteins. Poxviruses replicate in the cytoplasm and make their own enzymes for nucleic acid replication.
II: Single-stranded (+)sense DNA (Parvoviruses)Replication occurs in the nucleus, involving the formation of a (-) sense strand, which serves as a template for (+) strand RNA and DNA synthesis.
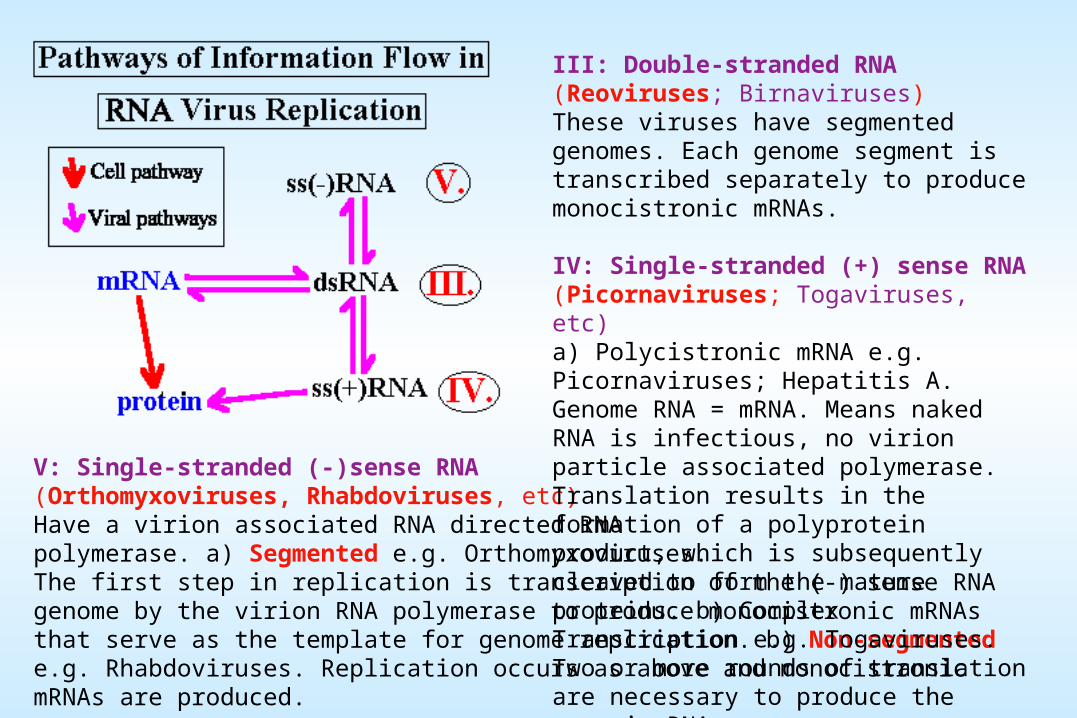
V: Single-stranded (-)sense RNA (Orthomyxoviruses, Rhabdoviruses, etc)Have a virion associated RNA directed RNA polymerase. a) Segmented e.g. Orthomyxoviruses. The first step in replication is transcription of the (-) sense RNA genome by the virion RNA polymerase to produce monocistronic mRNAs that serve as the template for genome replication. b) Non-segmented e.g. Rhabdoviruses. Replication occurs as above and monocistronic mRNAs are produced.
III: Double-stranded RNA (Reoviruses; Birnaviruses)These viruses have segmented genomes. Each genome segment is transcribed separately to produce monocistronic mRNAs.
IV: Single-stranded (+) sense RNA (Picornaviruses; Togaviruses, etc)a) Polycistronic mRNA e.g. Picornaviruses; Hepatitis A. Genome RNA = mRNA. Means naked RNA is infectious, no virion particle associated polymerase. Translation results in the formation of a polyprotein product, which is subsequently cleaved to form the mature proteins. b) Complex Transcription e.g. Togaviruses. Two or more rounds of translation are necessary to produce the genomic RNA.
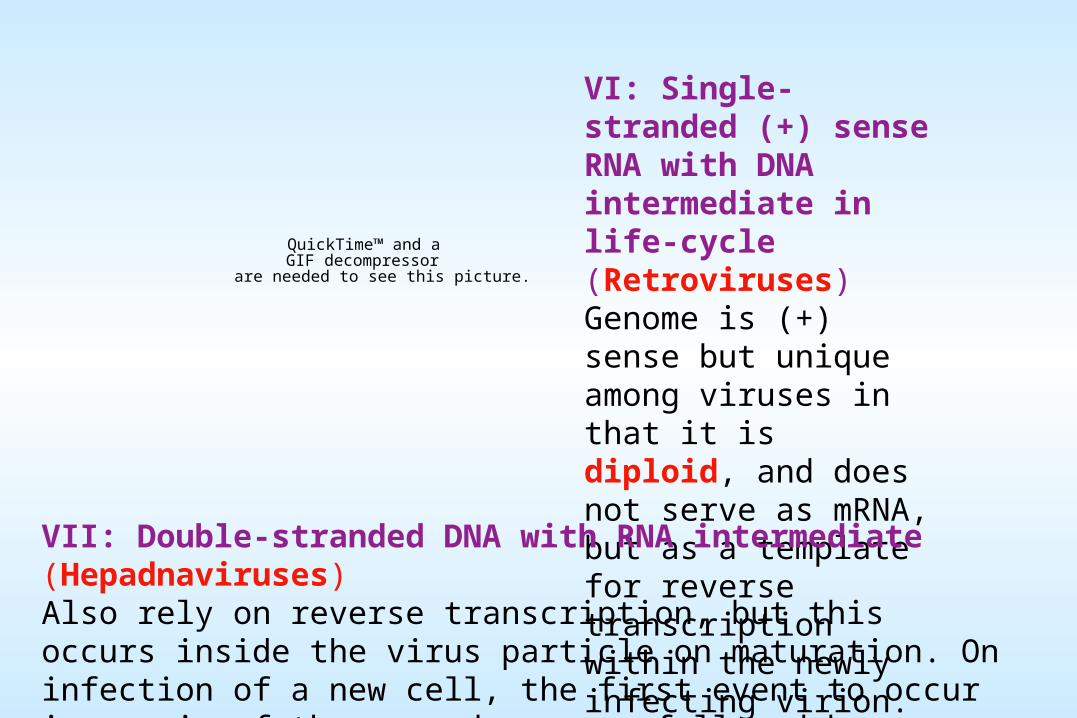
VI: Single-stranded (+) sense RNA with DNA intermediate in life-cycle (Retroviruses)Genome is (+) sense but unique among viruses in that it is diploid, and does not serve as mRNA, but as a template for reverse transcription within the newly infecting virion.
VII: Double-stranded DNA with RNA intermediate (Hepadnaviruses)Also rely on reverse transcription, but this occurs inside the virus particle on maturation. On infection of a new cell, the first event to occur is repair of the gapped genome, followed by transcription.
QuickTime™ and aGIF decompressor
are needed to see this picture.
Naked icosahedral viruses - Preassembled capsomers are joined to form empty capsids (procapsid) which are the precursors of virions.
They are released from infected cells in different ways.
Poliovirus is rapidly released, with death and lysis of infected cells.
DNA viruses tend to mature in the nucleus tend to accumulate within infected cells over a long period and are released when the cell undergoes autolysis, and in some cases, may be extruded without lysis.
MATURATION AND RELEASE Maturation proceeds differently for naked, enveloped, and complex viruses
Enveloped Viruses - Viral proteins are first associated with the nucleic acid to form the nucleocapsid, which is then surrounded by an envelope. In nucleocapsid formation, the proteins are all synthesized on cytoplasmic polysomes and are rapidly assembled into capsid components. In envelope assembly, virus-specified envelope proteins go directly to the appropriate cell membrane (the plasma membrane, the ER, the Golgi apparatus), displacinghost proteins. In contrast, the carbohydrates and the lipids are produced by the host cell. The viral envelope has the lipid constitution of the membrane where its assembly takes place (eg. the plasma membrane for orthomyxoviruses and paramyxoviruses, the nuclear membrane for herpesviruses [on right]). A given virus will differ in its lipids and carbohydrates when grown in different cells, with consequent differences in physical, biological, and antigenic properties.
VIRAL PATHOGENESIS
Results from:
Transmission to a new host
Replication and spread within the host(a function of viral tropism)
Cell damage and dysfunction(can be mediated by the virus or by immune defense mechanisms)
Disease symptoms and abnormal laboratory test values
KOCH'S POSTULATES
[Proof of etiology for infectious agents]
1) The organism must always be found in the diseased animal but not in healthy ones
2) The organism must be isolated from diseased animals and grown in pure culture away from the animal
3) The organism located in pure culture must initiate and reproduce the disease when reinoculated into susceptible animals
4) The organism should be reisolated from the experimentally infected animals
Although not all of these criteria can be met when evaluating the etiology of human disease, and some etiologic agents still cannot be cultured, these general principles guide the establishment of etiology of all infectious agents
West Nile virusTransmission:By mosquitoes that have fed on infected birds. In 10-14 days the virus reaches the salivary gland and can be transmitted to humans and animals.Host:At least 17 native bird species, including the house sparrow. Once infected they can pass the virus to mosquitos for up to 5 days.Carrier:Several species can carry the virus, Culex pipiens is most common carrier in the NE US. They take 3-4 blood feedings in their 2-3 weeks life span.Incidental hosts:Humans and horses can be infected but most people do not become sick. Ordinary human contact will not spread the virus.
Viral PathogenesisViral pathogenesis is an abnormal situation of no value to the virus - the vast majority of virus infections are sub-clinical, i.e. asymptomatic.. For pathogenic viruses, there are a number of critical stages in replication which determine the nature of the disease they produce:
1) Entry into the HostThe first stage in any virus infection, irrespective of whether the virus is pathogenic or not. In the case of pathogenic infections, the site of entry can influence the disease symptoms produced. Infection can occur via:
* Skin - dead cells, therefore cannot support virus replication. Most viruses which infect via the skin require a breach in the physical integrity of this effective barrier, e.g. cuts or abrasions. Many viruses employ vectors, e.g. ticks, mosquitos or vampire bats to breach the barrier. * Respiratory tract - In contrast to skin, the respiratory tract and all other mucosal surfaces possess sophisticated immune defence mechanisms, as well as non-specific inhibitory mechanisms (cilliated epithelium, mucus secretion, lower temperature) which viruses must overcome. * Gastrointestinal tract - a hostile environment; gastric acid, bile salts, etc * Genitourinary tract - relatively less hostile than the above, but less frequently exposed to extraneous viruses (?) * Conjunctiva - an exposed site and relatively unprotected
2) Primary ReplicationHaving gained entry to a potential host, the virus must initiate an infection by entering a susceptible cell. This frequently determines whether the infection will remain localized at the site of entry or spread to become a systemic infection, e.g:
Apart from direct cell-cell contact, there are 2 main mechanisms for spread throughout the host:
* via the bloodstream * via the nervous system
Virus may get into the bloodstream by direct inoculation - e.g. Arthropod vectors, blood transfusion or I.V. drug abuse. The virus may travel free in the plasma (Togaviruses, Enteroviruses), or in association with red cells (Orbiviruses), platelets (HSV), lymphocytes (EBV, CMV) or monocytes (Lentiviruses). Primary viremia usually proceeds and is necessary for spread to the blood stream, followed by more generalized, higher titer secondary viremia as the virus reaches other target tissues or replicates directly in blood cells.
As above, spread to nervous system is preceded by primary viremia. In some cases, spread occurs directly by contact with neurons at the primary site of infection, in other cases via the bloodstream. Once in peripheral nerves, the virus can spread to the CNS by axonal transport along neurons (classic - HSV). Viruses can cross synaptic junctions since these frequently contain virus receptors, allowing the virus to jump from one cell to another.
4) Cell/Tissue DamageViruses may replicate widely throughout the body without any disease symptoms if they do not cause significant cell damage or death.
Retroviruses do not generally cause cell death, being released from the cell by budding rather than by cell lysis, and cause persistent infections, even being passed vertically to offspring if they infect the germ line.
Conversely, Picornaviruses cause lysis and death of the cells in which they replicate, leading to fever and increased mucus secretion in the case of Rhinoviruses, paralysis or death (usually due to respiratory failure) for Poliovirus.
5) Cell/Tissue TropismTropism - the ability of a virus to replicate in particular cells or tissues - is controlled partly by the route of infection but largely by the interaction of a virus attachment protein (V.A.P.) with a specific receptor molecule on the surface of a cell, and has considerable effect on pathogenesis. Many V.A.P.'s and virus receptors are now known.
6) Persistence vs. ClearanceThe eventual outcome of any virus infection depends on a balance between two processes:
i) Persistence:Long term persistence of virus results from two main mechanisms:
a) Regulation of lytic potentialThe strategy followed is the continued survival of a critical number of virus infected cells - sufficient to continue the infection without killing the host.
* For viruses which do not usually kill the cells in which they replicate, this is not usually a problem, hence these viruses tend naturally to cause persistent infections, e.g. Retroviruses. * For viruses which undergo lytic infection, e.g. Herpesviruses, it is necessary to develop mechanisms which restrict virus gene expression, and consequently, cell damage.
b) Evasion of immune surveillance - Includes:
* antigenic variation * immune tolerance, causing a reduced response to an antigen, may be due to genetic factors, pre-natal infection, molecular mimicry * restricted gene expression * down-regulation of MHC class I expression, resulting in lack of recognition of infected cells e.g. Adenoviruses * down-regulation of accessory molecules involved in immune recognition e.g. LFA-3 and ICAM-1 by EBV. * infection of immunocompromised sites within the body e.g. HSV in sensory ganglia in the CNS * direct infection of the cells of the immune system itself e.g. Herpes viruses, Retroviruses (HIV) - often resulting in immunosuppression.
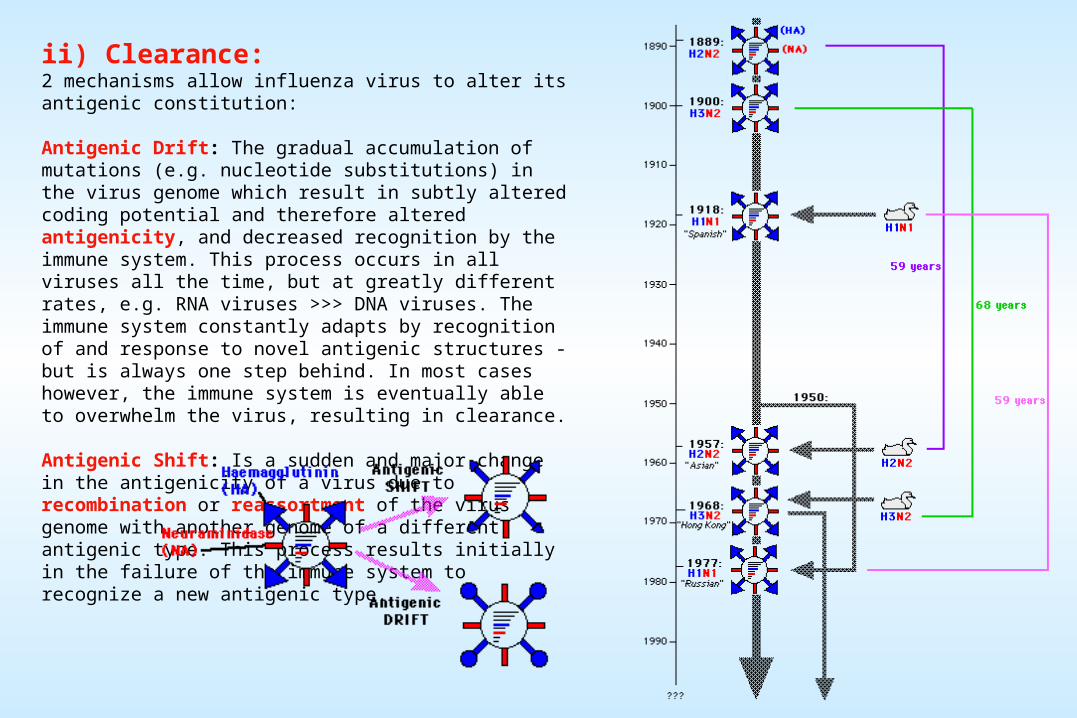
ii) Clearance:2 mechanisms allow influenza virus to alter its antigenic constitution:
Antigenic Drift: The gradual accumulation of mutations (e.g. nucleotide substitutions) in the virus genome which result in subtly altered coding potential and therefore altered antigenicity, and decreased recognition by the immune system. This process occurs in all viruses all the time, but at greatly different rates, e.g. RNA viruses >>> DNA viruses. The immune system constantly adapts by recognition of and response to novel antigenic structures - but is always one step behind. In most cases however, the immune system is eventually able to overwhelm the virus, resulting in clearance.
Antigenic Shift: Is a sudden and major change in the antigenicity of a virus due to recombination or reassortment of the virus genome with another genome of a different antigenic type. This process results initially in the failure of the immune system to recognize a new antigenic type.
Determinants of cell damage and dysfunction
1. Direct destruction of cells by virusPoliovirus - poliomyelitis Herpes simplex - cold soresRotavirus - diarrhea HIV - AIDS
2. Immune mediated destruction of virus-infected cells (CTL and Ab)Hepatitis B - hepatitis Dengue - Dengue shockMeasles - post-measles encephalitis & SSPE
3. Indirect effector mechanismsInfluenza - interleukins and interferons releasedRhinovirus - kinin release, vascular changesRespiratory syncytial - IgE Ab-mediated effectsHepatitis B and C - Ab:Ag complex deposition
4. Virus-encoded immune alterationAdenovirus - E3 alters class I MHC expressionVaccinia (and other poxviruses) - TNF analogueEpstein-Barr - Interleukin 10
5. Allow access to other pathogens
Subtle effects of viruses
1. Chronic/persistent infectionLong term viral growth without disease, more important in immunocompromised hostsRetroviruses, herpesviruses, hepatitis B, adenoviruses
2. Latent infectionLifelong presence of viral genome with potential for reactivationRetroviruses, papillomaviruses, herpesviruses
3. Abortive infectionVirus persists with partial replicationMeasles: subacute sclerosing panencephalitis
Explanations for the lack of replication in non-permissive cells:
• No attachment to cell(no receptor)• Virus can enter but not uncoat• Virus can uncoat but not express genes (mRNA or protein)• Virus can express genes but produce no particles• Progeny particles form but fail to mature• Mature particles accumulate in the cell but are not released• Released viruses are not infectious for other cells
LATENCY
When virus infection fails to result in an immediate production of progeny. Rather, the virus enters a latent state in which the viral genome may become incorporated into the host's genome or maintained as an extrachromosomal element.
Latent viral genomes are passively replicated along with host chromosomes. Not all viral genes remain silent during latency and there may be a perceptible change in the phenotype of the cell. The viral genome may become reactivated, potentially up to decades after the initial infection.
Herpes, papilloma, Hepadnaviruses: Latent forms exist as plasmids (nuclear, extrachromosomal element)
Virus: Site of latent infection:Herpes simplex - sensory neuronsVaricella-zoster - dorsal root ganglionEpstein-Barr - B lymphocytesPapilloma - basal epitheliume.g. Development of "cold sores" by herpes viruses after the host is exposed to large doses of unfiltered sunlight.
Retroviruses:Latent forms exist as "proviruses" integrated into the host chromosome
HTLV 1 - T cellsHIV - CD4+ T cells, macrophages
Latency can be associated with tumorigenic transformation of the cell. Infrequently, latent viral nucleic acid may be passed not only to the progeny of the cells within the host, but also through successive generations in the germ line.
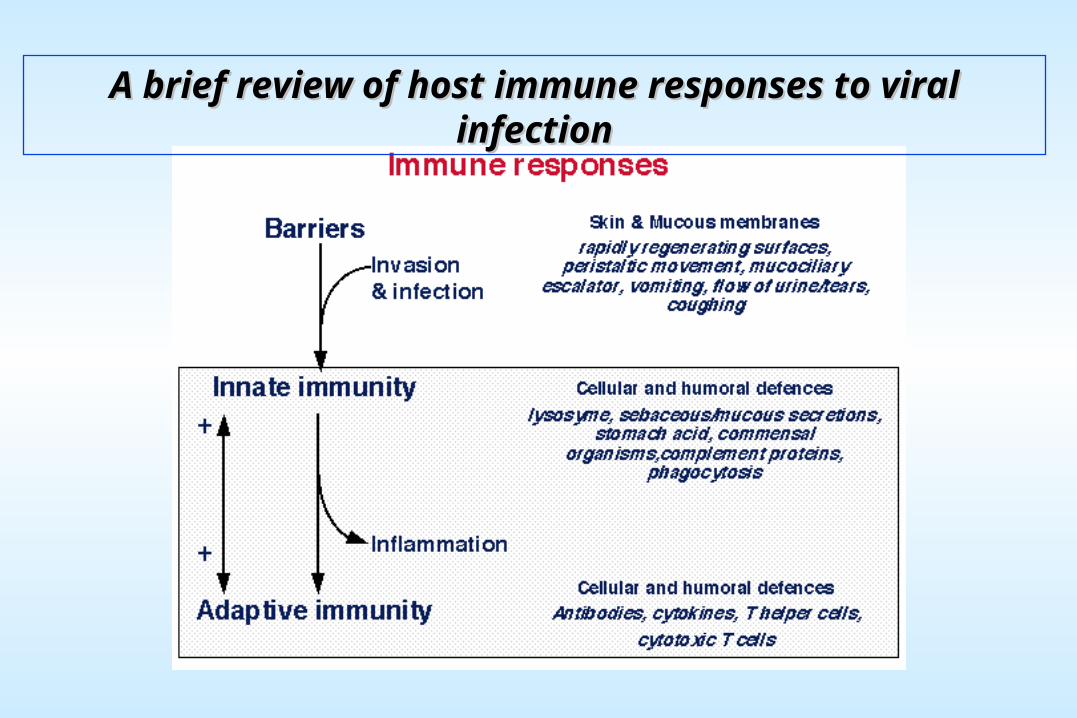
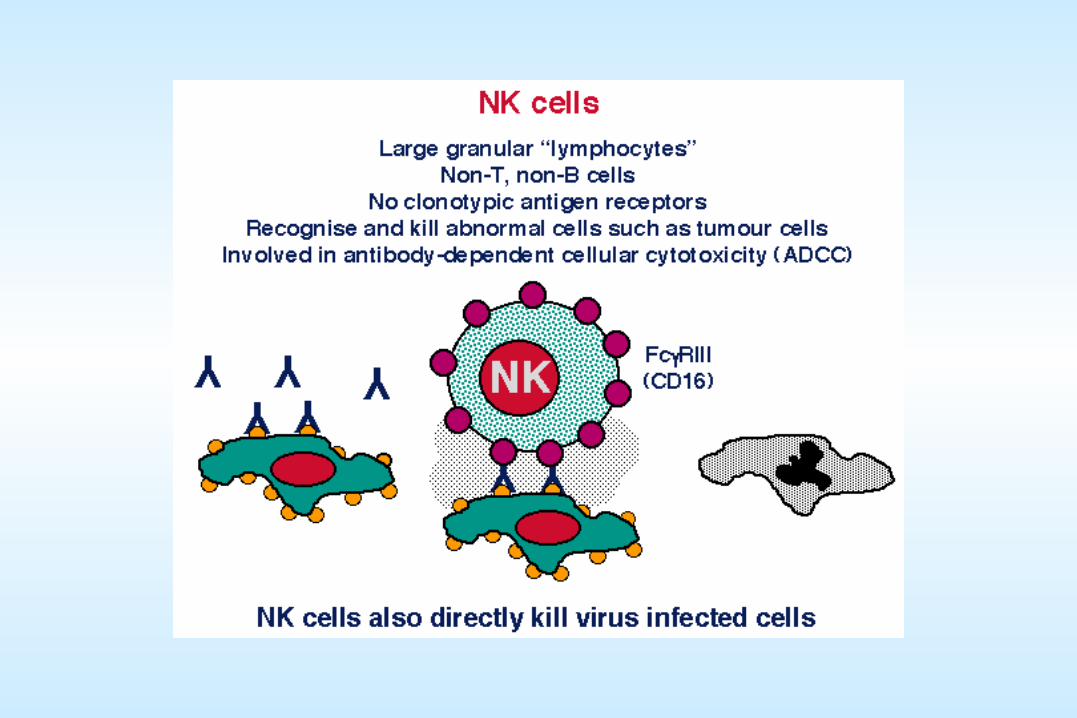
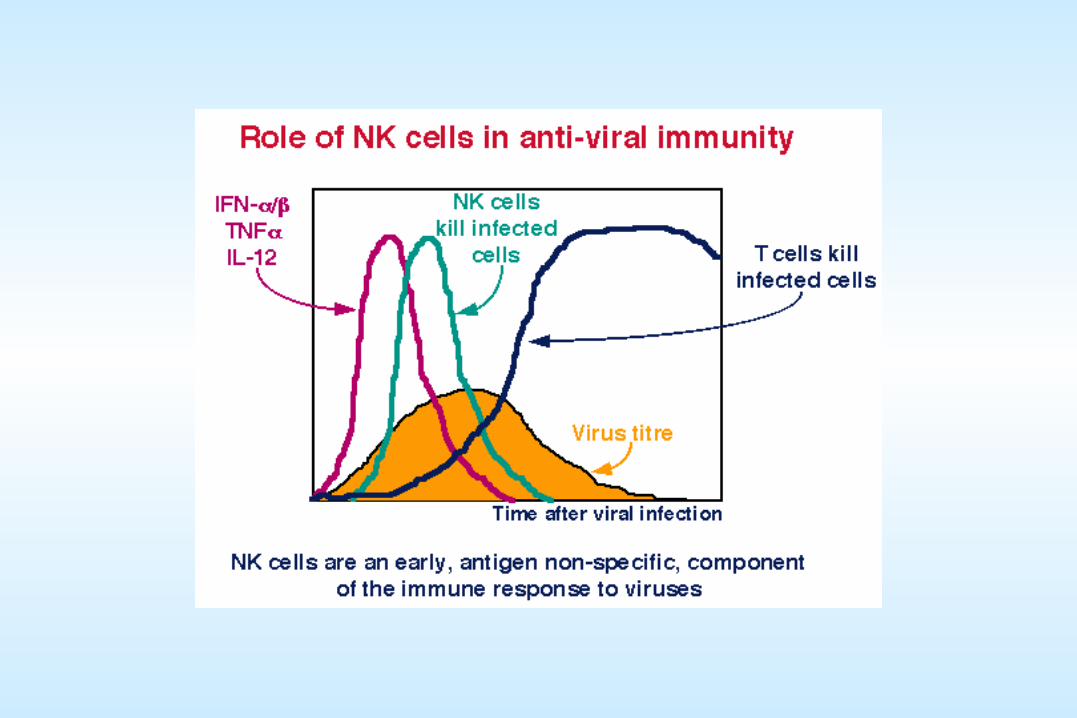
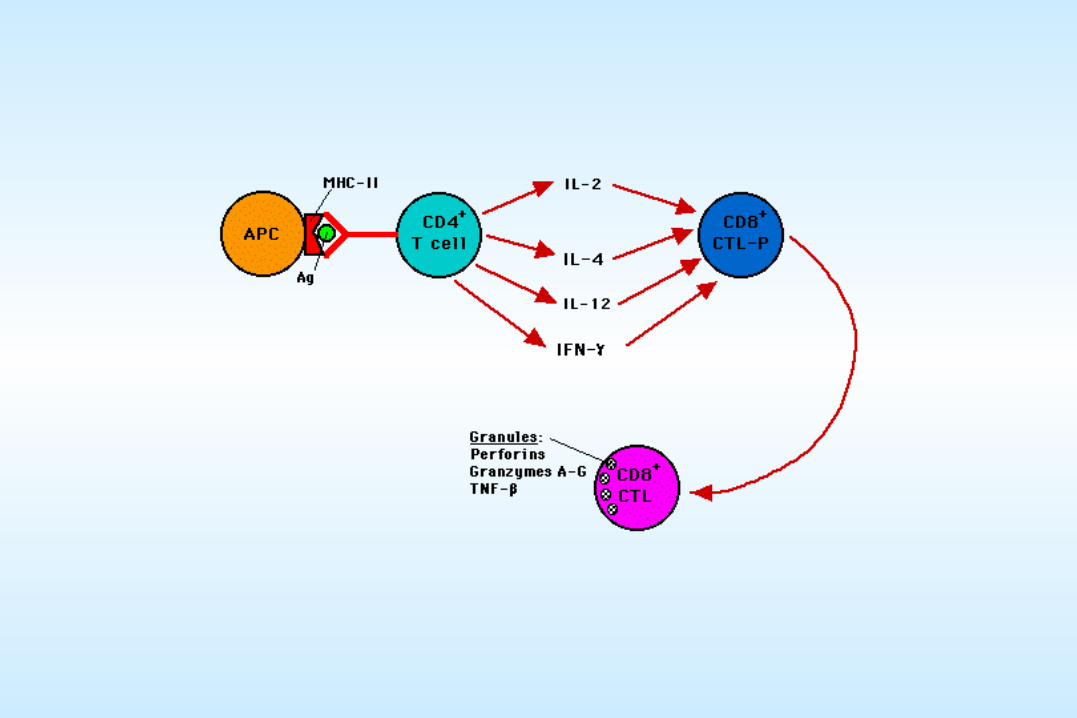
A brief review of host immune responses to viral infectionA brief review of host immune responses to viral infection
QuickTime™ and aVideo decompressor
are needed to see this picture.
QuickTime™ and aVideo decompressor
are needed to see this picture.
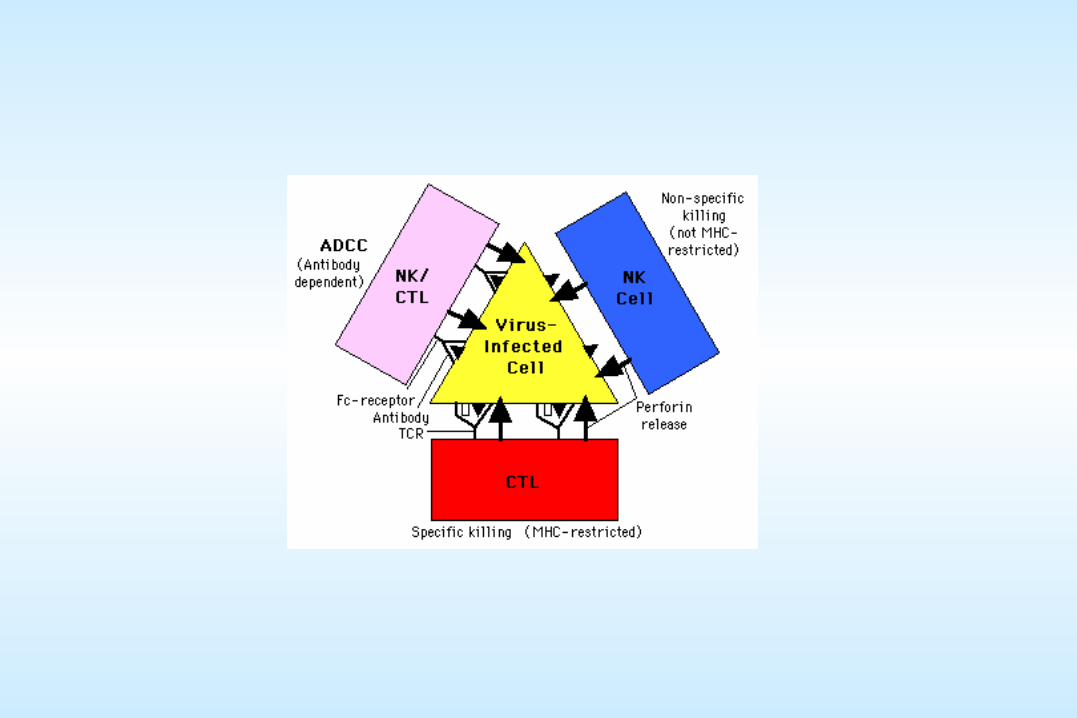
CTL vs.Virus-infected cell
Macrophage vs.Virus-infected cell
RETROVIRUSES
The first infectious agents implicated in tumors (chicken sarcomas, identified by Peyton Rous 1906)
The enormous current interest in retroviruses can be attributed to: Their etiologic role in AIDS and certain forms of cancerTheir potential utility as vectors in gene therapy, and, Due to several aspects of a unique replication mechanism and a close relationship
with the genome of its host, including that they are the only animal viruses that integrate into the host cell's genome during the normal growth cycle.
Retroviruses (and transposable elements) appear to be part of every cell's genome (bacteria, yeast, flies, mice and humans (~0.25% of the mouse genome). Some endogenous viruses can be activated to replicate and induce tumors; however, the great majority of sequences in eukaryotic genomes that are related to retroviruses seem innocuous.
In most instances retrovirus replication is non-cytopathic (AIDS is one exception)and persistent
Three retrovirus subfamilies: Oncovirus subfamily is also subdivided according to morphological criteria:
Subfamily Morph. group ExamplesOncoviruses Type A Intracisternal A Particles
(non-infectious)Type B Mouse Mammary Tumor (MMTV)
Type C Avian Sarcoma and LeukemiaMurine Sarcoma and Leukemia Human T-cell leukemia virus (HTLV)
Type D Mason Pfizer monkey (MPMV)
Lentiviruses Visna, HIV, SIV
Spumaviruses Human Foamy virus(cytopathic in vitro, no disease association)
All retroviruses have three essential genes which encode polyproteins precursors essential for virus replication:
gag (group specific antigen) gene encodes the viral matrix (MA), capsid (CA) and nucleoproteins (NC). The protease (PR) orf encodes a product that cleaves the Gag polyprotein precursor. It can be encoded as part of Gag or a Gag-Pro-Pol polyprotein, sometimes following a frame shift or a stop codon, and read-through about 5% of the time by a ribosomal frame shifting mechanism or by stop codon suppression:
The major read-through product is derived from the pol gene which encodes the RT (including RNaseH) and an integrase (IN) that is involved in provirus integration:
The envelope gene encodes the surface glycoprotein (SU) - transmembrane (TM) polyprotein.
vif
rev
env
rev
polgag
1000 2000 3000 4000 5000 6000 7000 8000 9000
nef
vpr
••
tat
••
vpu
•
tat
•
1 2 3 4 5 6 7 8 9kb
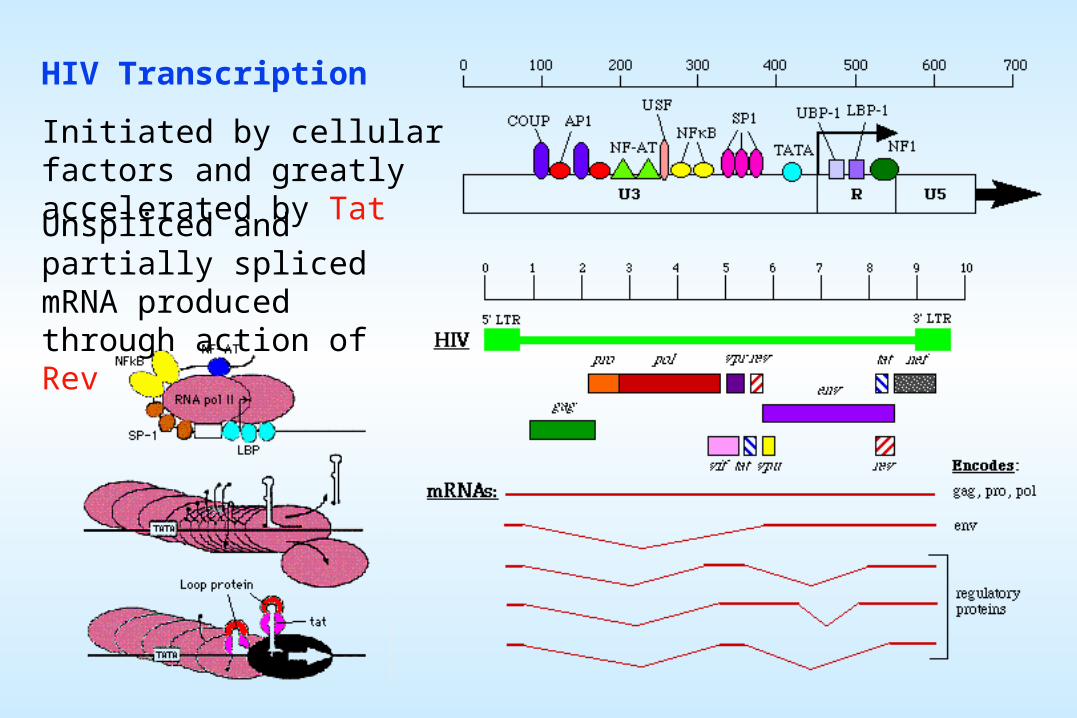
Open Reading Frame map of HIV-1:
HIV Transcription
Initiated by cellular factors and greatly accelerated by Tat
Unspliced and partially spliced mRNA produced through action of Rev
“AUXILIARY GENES”
The human oncoviruses and the lentiviruses are more complex than typical animal oncoviruses.
HTLV-1 and 2 encode regulatory sequences located between env and the 3' LTR.
The primate lentiviruses are more complex and encode an as yet poorly understood array of expression regulators encoded by “auxiliary genes” found in the central (between pol and env) and 3’ (within env and between env and U3) regions of the viral genome.
These genes are expressed from a series of differentially spliced mRNAs.
Tat and Rev are the only “essential” auxiliary genes
All auxiliary gene products modulate infectivity. Except for Nef, the additional auxiliary genes of HIV described below are expressed late in infection and are not essential for virus growth.
Nef (for negative effector) causes downregulation of the HIV receptor, CD4, from the cell surface. It may also be capable of activating resting cells and does increase virus levels in vivo, thus it may actually be a positive regulator of virus.
Nef also downregulates expression of Class I HLA molecules. This may help explain why CTL (cytotoxic T lymphocytes), that recognize peptide antigens presented by HLA molecules, are ineffective at controlling HIV infections.
Permanent disruption of the SIV nef gene by deletion results in 106 lowered levels of expression in experimentally infected monkeys. In contrast, point mutations are easily overcome by this virus.
Prior infection with the nef mutant can prevent subsequent fulminant infection with a virulent form of the virus. In Australia, a cohort of hemophiliacs infected by transfusion with a strain of HIV with multiple deletions in nef have remained asymptomatic for almost two decades. Thus, attenuated mutant forms of SIV have been explored for potential use as vaccines. However, recent results have shown both that SIV can in at least some cases “fix” these deletions, and that multiply deleted (nef plus other auxiliary genes) viruses can be fully virulent in newborns and can occasionally become virulent in adults.
Finally, the CD4 count of the Australians have begun to drop and some adult monkeys infected with the attenuated viruses have gotten disease. Hence, attenuated HIV and SIV strains can still cause disease.
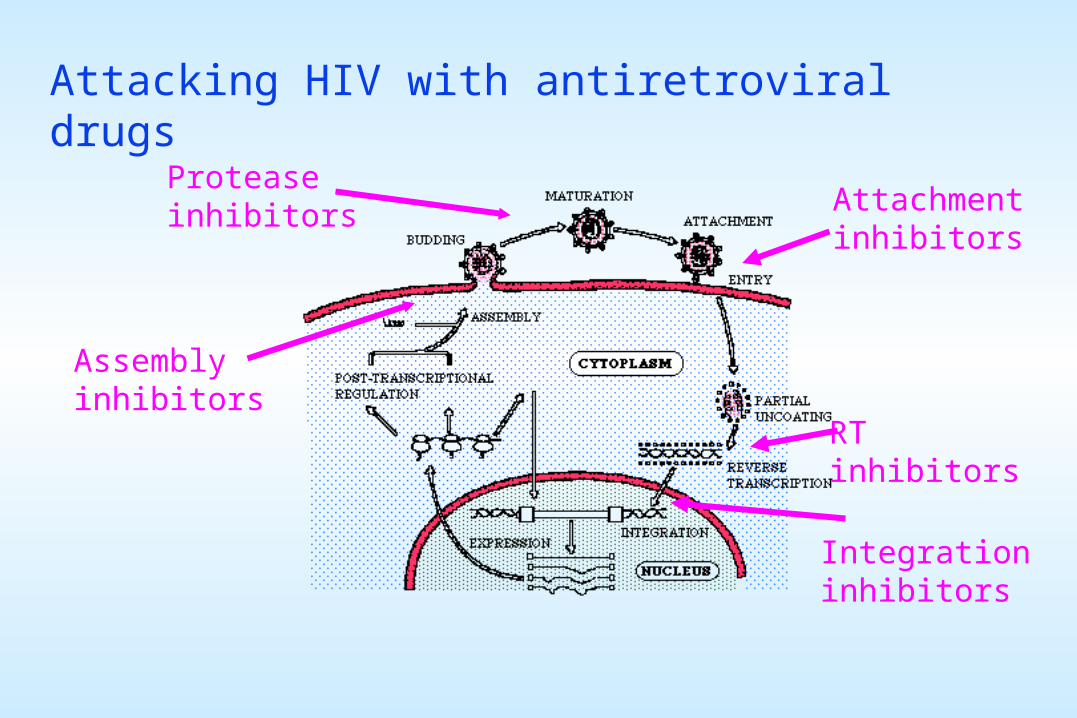
Attacking HIV with antiretroviral drugs
Attachmentinhibitors
Proteaseinhibitors
RTinhibitors
Integrationinhibitors
Assemblyinhibitors
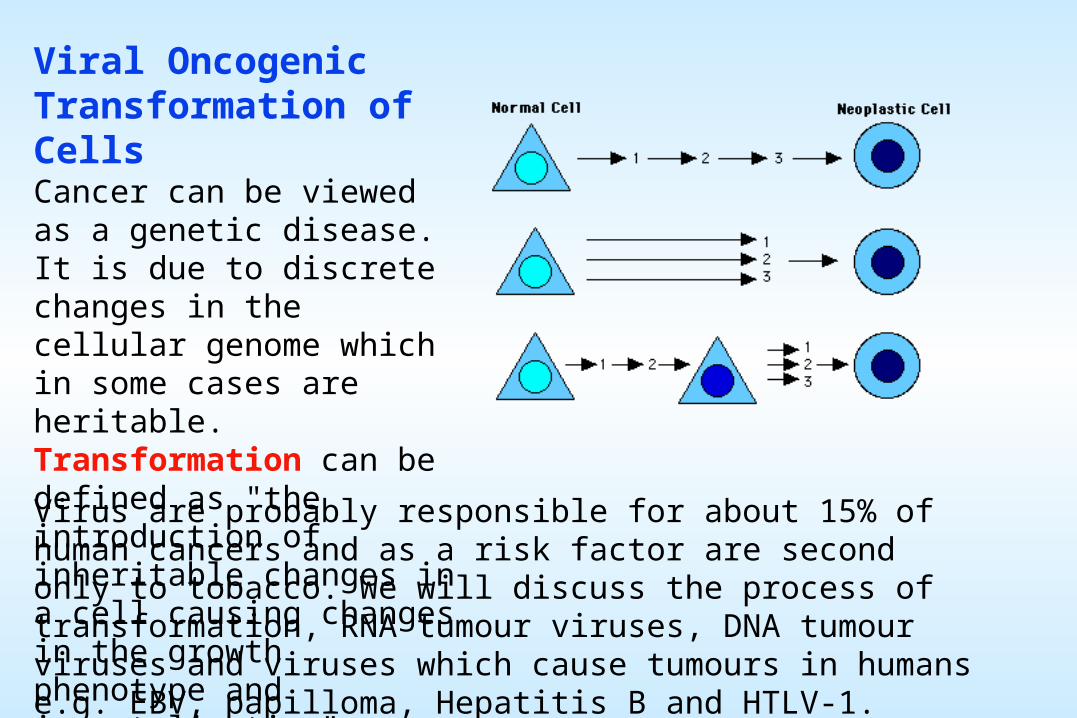
Viral Oncogenic Transformation of CellsCancer can be viewed as a genetic disease. It is due to discrete changes in the cellular genome which in some cases are heritable. Transformation can be defined as "the introduction of inheritable changes in a cell causing changes in the growth phenotype and immortalisation".
Virus are probably responsible for about 15% of human cancers and as a risk factor are second only to tobacco. We will discuss the process of transformation, RNA tumour viruses, DNA tumour viruses and viruses which cause tumours in humans e.g. EBV, papilloma, Hepatitis B and HTLV-1.
Cell Transformation
The G0/G1 boundary of the cell cycle (G0 is the resting or stationary phase, G1 is the phase in which the cell gears up for division) is a particularly important control point because this acts as a commitment to cell division. Tumor formation results from a failure of regulatory mechanisms which control this boundaryTumor cells continue to divide under circumstances in which their normal cellular counterparts do not.
Transformation of cells in culture is the in vitro counterpart of the process by which tumor induction in animals (by viruses) occurs. Transformed cells are atypical in many ways. For example:
Growth: * high/indefinite saturation density * different, usually reduced serum requirement * tumor formation when injected into animals * no contact inhibition of growth/movement
Surface: * changes in glycoproteins and glycolipids * loss of tight junctions * fetal antigen expression * increased rate of nutrient transfer * increased secretion of proteases
Intracellular: * disruption of cytoskeleton * altered amounts of signalling molecules (cyclic nucleotides, phosphoinositides)
Mechanism of Transformation
In vivo and epidemiological studies indicate that transformation is a multi-step process involving: initiation, promotion and progression
Transformation involves gene mutations, amplification of cells containing these mutations and further changes leading to transformation
How do viruses transform cells ?By subverting the normal cell control mechanisms of a critical cellular gene(s)
There is a class of cell genes which, generally speaking, promote cell replication. These are sometimes called cellular oncogenes (c-oncogenes) or proto-oncogenes. Viruses, especially retroviruses can affect the activity of these genes. Indeed retroviruses sometimes carry their own versions of these genes, called v-oncogenes.
Another class of genes suppress cell replication, these are called tumor suppressors and many viruses especially DNA viruses prevent their proper functioning.
Cell transformation by viruses is accompanied by the persistence of all or a part of the viral genome. It can also accompanied by the continual expression of a limited number of viral genes.
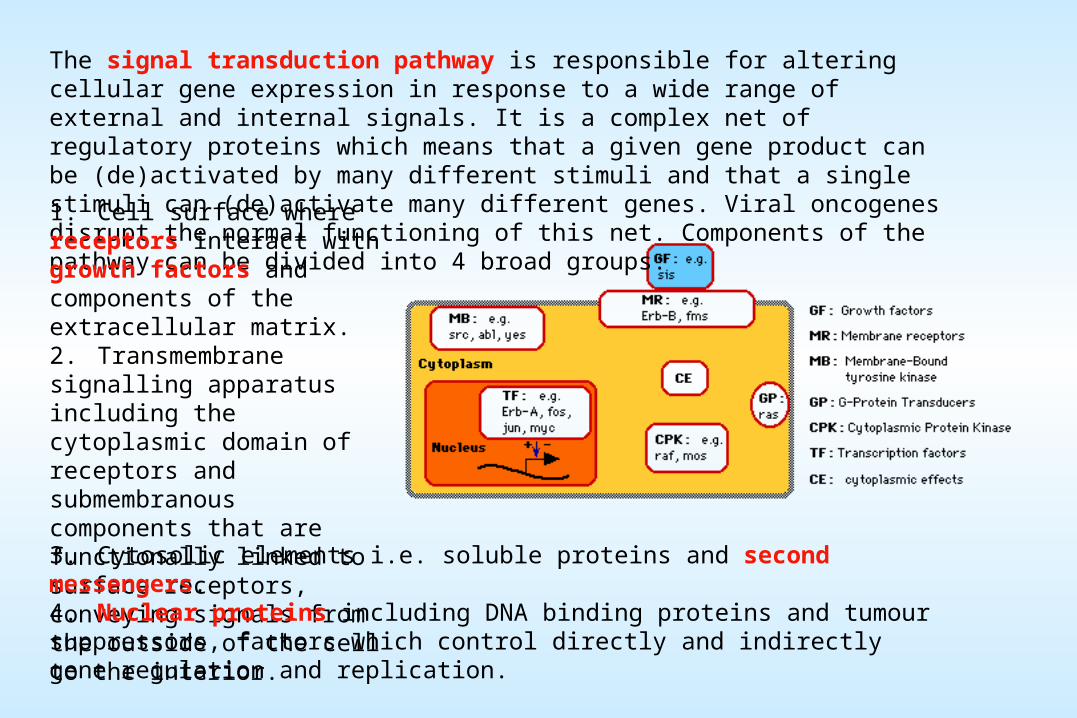
The signal transduction pathway is responsible for altering cellular gene expression in response to a wide range of external and internal signals. It is a complex net of regulatory proteins which means that a given gene product can be (de)activated by many different stimuli and that a single stimuli can (de)activate many different genes. Viral oncogenes disrupt the normal functioning of this net. Components of the pathway can be divided into 4 broad groups:1. Cell surface where receptors interact with growth factors and components of the extracellular matrix. 2. Transmembrane signalling apparatus including the cytoplasmic domain of receptors and submembranous components that are functionally linked to surface receptors, conveying signals from the outside of the cell to the interior. 3. Cytosolic elements i.e. soluble proteins and second messengers. 4. Nuclear proteins including DNA binding proteins and tumour suppressors, factors which control directly and indirectly gene regulation and replication.
RETROVIRAL ONCOGENESIS
Most retroviruses were discovered by virtue of their association with naturally occurring tumors or leukemias in animals, including humans. It soon became apparent that they could be divided into two groups according to their ability to induce tumors.
Virus Type and characteristics Examples
Acutely transforming Avian, murine and feline sarcomacause tumors in 1 to a few weekshigh efficiency of disease inductioncarry oncogenestransform cells in vitronearly all are defective
Slowly transforming Avian, murine and feline leukemiacause tumors in 6-12 months Mouse Mammary Tumorlow efficiency HTLV Ido not carry oncogenesno visible effects on cells in culturemost are replication competentsome are endogenous viruses
FAST TRANSFORMING RETROVIRUSES
High efficiency (or "acute") transforming retroviruses are usually defective, needing a helper virus to provide necessary replication functions.
Acute transforming retroviruses incorporate or transduce cellular genes, "oncogenes", which in the context of being expressed from the viral genome cause malignant growth.
Oncogene-carrying retroviruses will also transform cells in culture so that these cells will form tumors when injected into animals.
Oncogenes need not be incorporated into a virus for that virus to cause a tumor. Yet, as we study the genetic mechanisms leading to tumor induction associated with oncogene-bearing viruses, non-oncogene bearing viruses, and even tumors with no known viral etiology, a recurring theme is noted.
In virtually all cases, individual cellular genes can be identified, referred to as proto-oncogenes, which are subverted from their normal functioning, resulting in some manner in tumor formation.
This subversion occurs as a result of mutational change of coding sequences and/or alteration of their normal regulation of expression.
SLOW TRANSFORMING RETROVIRUSES
In certain retrovirus-induced tumors of chickens, proviruses were found to integrate upstream of the c-myc gene. Transcription, initiated within the viral LTR, caused enhanced expression of the c-myc gene. Thus, once again proto-oncogenes were implicated in the cause of retrovirus-induced tumors.
A partially overlapping, yet larger group of proto-oncogenes have been found to be activated by provirus insertion (without transduction).
Activation of cellular proto-oncogenes by slowly transforming viruses occurs primarily as a result of provirus insertion and is most often LTR mediated.
Different types of tumors are induced by the same virus in different host genetic backgrounds. Activation of c-myc is associated primarily with B-cell tumors in chickens and with T-cell tumors in mice and cats.
The tumorigenic potential of retroviruses is not constant. Viruses which replicate more quickly or to higher levels in the animal are generally more oncogenic, a property due in large part to LTR enhancer properties. However, other portions of the genome provide essential contributions to tumorigenicity.
TUMOR INDUCTION BY NON-PRIMATE RETROVIRUSES
Tumors induced by acutely transforming viruses are polyclonal in nature, that is, many cells lose growth control as a result of infection with an oncogene-bearing virus.
Tumors caused by slow transforming viruses are mono or possibly oligoclonal in nature, they occur as a result of an outgrowth of a single rare cell with virus integrated into a specific site near or within a proto-oncogene.
Tumor clonality therefore reflects the mechanisms of induction.
Tumor induction by HTLV-1 presents a third face:
The tumors are oligo or monoclonal cell outgrowths, yet virus integrations occurred at no preferred sites within the genome.
Furthermore, these viruses are wholly exogenous to the species they infect, hence they harbor no classic oncogenes.
Unlike animal oncoviruses, HTLV-1 encodes a transactivator, Tax, in addition to Gag, Pol and Env.
Tax transactivates expression of the viral LTR and stimulates expression of genes involved in cellular gene regulation including the interleukin 2 (IL-2) and IL-2 receptor (IL-2R) genes which are known to effect T-cell growth regulation.
These observations therefore support (albeit not convincingly to date) an autocrine model for tumor induction in which, for example, activation of IL-2 and IL-2R leads to continuous cell proliferation.
















































![Retroviruses - 2013 (FN) [Compatibility Mode]](https://static.documents.pub/doc/80x56/577cdda21a28ab9e78ad6fbf/retroviruses-2013-fn-compatibility-mode.jpg)








