Hyalella armata (Crustacea, Amphipoda, Hyalellidae) and the description of arelated new species from Lake TiticacaExequiel R. González1, Charles O. Coleman2
1 Depto. de Biología Marina, Universidad Católica del Norte, Casilla 117, Coquimbo, Chile, [email protected]
2 Museum für Naturkunde Berlin, Humboldt-Universität, Invalidenstraße 43, D-10115 Berlin, Germany;e-mail: [email protected]
Received 13 March 2002 ⋅ Accepted 12 August 2002
Abstract
Hyalella armata (Faxon, 1876) was considered for a long time to be a monotypic species. The peculiar morphology, very distinct from otherspecies in Lake Titicaca, was not analysed in detail after the original description. However, the present authors’ study of an extensive col-lection from The Natural History Museum in London has revealed two morphologically distinct species. We herein redescribe H. armata anddescribe the new species H. longispina.
Key words: Crustacea, Amphipoda, Hyalella, Lake Titicaca, taxonomy
Introduction
The distinctive fauna of amphipod crustaceans of thegenus Hyalella Smith, 1874 in Lake Titicaca hasbeen extensively sampled, but not properly studied.The first work by Faxon (1876), based on thematerial collected by A. Agassiz and S. W.Garthman in 1875, identified seven endemic and onenot endemic species. Later, Chevreux (1904, 1907)described four species collected by Dr. Neveu-Lemaire during the Mission de Crequi Montfort etSenegal de la Grange to Lake Titicaca. Chevreuxalso redescribed two of Faxon’s species. Two moremajor sampling efforts were made in the Lake. In1937, G. I. Crawford collected extensively in theLake during the Percy Sladen Trust Expedition anddeposited the material of about 20,000 specimens inthe Natural History Museum of London. More than15,000 specimens were collected by C. Dejoux ofORSTOM, France (Dejoux 1992). The present studydeals with two of the spiny forms of Hyalellapresent in the Lake, H. armata (Faxon, 1876) andthe new species H. longispina.
Material and methods
The following material was studied (see Fig. 14):
Hyalella armata
From the collection of C. Dejoux: Lago Menor(material illustrated), Gulf of Puno, Gulf of Desa-guadero, Chuquito, Juli, between Taquili andAmantane. From the Crawford collection, NaturalHistory Museum, London (numbers in brackets =specimens): Coata Bay, GIC 253, 23 m (196); CoataBay, GIC 286, 12-19m (15); Coata Bay, GIC 275,30m (30); Coata Bay, GIC 296, 19m (12); CoataBay, GIC 321, 1m (1); Coata Bay, GIC 283, 12-15m(4); Coata Bay, GIC 304, 23m (19); Coata Bay, GIC268, 30 m (16); Coata Bay, GIC 291, 19m (201);Coata Bay, GIC 263, 30 m (204); Coata Bay, GIC160a (2); Coata Bay, GIC 282 (1); Isla Suana, P. 46,GIC 993/1a 14.2-14.6m (3); without location, GIC170a, 6m (1), GIC 160a (2).
Material deposited in the Museum of Compara-tive Zoology (Harvard University) by A. Agassizand S. W. Garthman, and identified by Faxon, wasalso examined.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 2
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
Hyalella longispina n. sp.
Holotype male, 10 mm; PERU, Lake Titicaca,Choccocoyo Bay, St. P 6, GIC 481, 11-23 m, leg.Crawford, 25.VI.1937. Paratypes (collecting data asholotype): allotype female, 9 mm; 106 males, 6-10mm; 93 females, 5-10 mm. All deposited at TheNatural History Museum, London (holotype:2002.311; allotype: 2002.312; other paratypes:2002.313-322). Additional material (in brackets =number of specimens) from the Crawford collection,Natural History Museum, London: Taman, GIC1327, 36-38 m (7); Taman, GIC 1323, St. P67, 56-61 m (29); Taman, GIC 256/3, 15-30m (3); Taman,GIC 1325/1 St. P67, 56-61 m (3); Patón, GIC 850,St. P. 19, 25-34m (5); Patón, GIC 843 P. 17, 25-34m(5); Siripata Bay, GIC 1005/1 P52, 12.3-16.2m (21);Molinopampa, GIC 881, P. 24,19-27m (1);Molinopampa, GIC 884, P. 28, 25m (8);Molinopampa, GIC 890, P. 28, 25m (4);Ancoraimes, P. 20, GIC 854/1, 12-24m (3);Ancoraimes, P20, GIC 855/2, 12-24m (1);Capachica, GIC 333, 20m (3), Choccocoyo, GIG468 (478?) (5).
The animals were transferred into glycerol anddrawn with a camera lucida on a Leica Wild M8dissecting microscope. Specimens were dissected,appendages and mouthparts were transferred ontoslides in glycerol and drawn under a Leica DMLBlight microscope using a camera lucida. Morphome-tric measurements were taken with a dissectingmicroscope and a camera lucida calibrated with astage micrometer. The results were calculated andplotted with Microsoft Excel using the linearregression mode.
Type material. Museum of Comparative Zoology,Harvard University, Cambridge, Mass.
Type locality. Achacache, Lake Titicaca, BOLIVIA.
Description of male. Size 9.5 mm. Body with aposterior dorsal transverse ridge on every segment(Fig. 1a). Epimeral plate 1 round, 2 straight, 3acuminate (Fig. 1b).
Head smaller than first two thoracic segments,typically gammaridean, rostrum absent. Eyes pig-mented, medium, round, located behind insertion ofantenna 1.
Antenna 1 (Fig. 4d) shorter than antenna 2,slightly longer than peduncle of antenna 2; pedunclelonger than head, articles 1 and 2 subequal in length,article 3 shorter than article 1, and same length asarticle 2; flagellum of 12 articles, same as peduncle,basal article not elongated.
Antenna 2 (Fig. 4e) less than half body length;peduncle slender, longer than head, article 4 shorterthan article 5, setal groups on article 4 and 5abundant; flagellum of 13 articles, longer thanarticle 5, basal article elongated. Upper lip (labrum)wider than long, entire, ventral margin slightlyrounded to truncate.
Basic amphipodan mandible (in the sense ofWatling 1993); incisor toothed (Fig. 4g); left laciniamobilis with five teeth; setae row on left mandiblewith three main setae without accessory setae, rightmandible with two main setae without accessorysetae; molar large, cylindrical and triturative,accessory seta absent. Lower lip (Fig. 4c) outerlobes rounded without notches or excavations,mandibular projection of outer lobes round.
Maxilla 1 (Fig. 2a, c) palp uniarticulate, longerthan wide, reaching more than half the distancebetween base of palp and tip of setae on outer plate,distal setae strong; inner plate slender, smaller thanouter plate, with two strong and pappose apicalsetae; outer plate with nine stout and serrate setae.Maxilla 2 (Fig. 3b) inner plate subequal in lengthand width to outer plate, one strong pappose seta oninner margin, outer and inner plates with abundantsetules.
Maxilliped (Fig. 3a) inner plates apically trunca-ted, with three conate setae, pappose setae apicallyand medially; outer plates larger than inner plates,apically truncated, apical, medial and facial setaesimple; palp longer than outer plate, four articles;article 2 wider than long, medial margin with longsimple setae; article 3 outer distal face with severallong simple setae, inner distal face with longplumose setae, inner distal margin with long setae,outer margin with one or two strong and longplumose setae; dactylus unguiform, shorter thanthird article, distal setae simple and shorter than nail,inner border without setae, distal nail present.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 3
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
Coxae 1 to 4 increasing in size, apicallyacuminate (Fig. 1a). Coxa 4 (Fig. 3e) much deeperthan 1 to 3 and at right angle to sagittal plane ofbody, wing-like. Coxa 1 subequal to 2 and 3. Coxa 3(Fig. 4b) narrower than 4. Coxa 4 deeper than wide,excavated posteriorly. Coxa 5 (Fig. 3f) posteriorlobe deeper than anterior. Coxa 6 anterior lobesmall.
Gnathopod 1 (Fig. 1d) subchelate; carpus widerthan long, subequal in length and as wide aspropodus, with strong and wide posterior lobe,produced and forming a scoop-like structure, open tothe inside, inner face with four to seven papposesetae, border pectinate and with several papposesetae; propodus (Fig. 4a) length less than two timesmaximum width (quadrangular), hatchet-shaped,with no setae on anterior border, inner face withmore than ten pappose setae, without smalltriangular setae, distoposterior and distoanteriorborders without setose scales, palm slope slightlyoblique, margin convex, posterior distal corner withrobust setae; dactylus claw-like, no setae on innercurvature, congruent with palm.
Gnathopod 2 (Fig. 1c) subchelate; basis hindmargin with group of seven or more setae; meruswith less than seven setae on posterior margin,postero-distal margin straight, distal corner rounded;carpus posterior lobe elongate, produced betweenmerus and propodus, border pectinate with severalpappose setae; propodus triangular, distoposteriorand distoanterior borders without setose scales, palmlonger than posterior margin, slope oblique, marginstraight, several medium-size setae, anterior edgewith a small process, posterior distal corner withweak setae, and with change of slope; dactylus claw-like, congruent with palm, no setae on innercurvature.
Peraeopods 3 to 7 simple. Peraeopods 3 and 4merus and carpus posterior margins each with fourhind marginal clusters of long setae; propodusposterior margin with two to four groups of setae;dactylus half length of propodus. Peraeopods 5 to 7all similar in structure and slightly longersuccessively; dactylus more than half length ofpropodus. Peraeopod 5 (Fig. 3f) subequal toperaeopod 4, basis posterior lobe deeper than wide,smaller than posterior lobe of peraeopod 7, meruswith two hind marginal setae, proximal and distalsetae subequal. Peraeopod 6 (Fig. 2b) longer thanperaeopod 4, basis posterior lobe deeper than wide,similar to posterior lobe of peraeopod 5, smaller thanposterior lobe of peraeopod 7. Peraeopod 7 (Fig. 2e)
subequal to peraeopod 6, basis posterior lobe widerthan deep.
Pleopods not modified; peduncle slender; longestramus longer than peduncle.
Uropod 1 (Fig. 2f) longer than uropod 2; pedunclelonger than rami; rami subequal; inner ramus withtwo dorsal and six distal setae, two of them longer,male without curved setae on inner side of theramus; outer ramus with one dorsal seta, and threedistal setae; peduncle setation present.
Uropod 2 (Fig. 3d) rami subequal; inner ramuswith two dorsal and three distal setae; outer ramuswithout dorsal and four distal setae; pedunclesetation present.
Uropod 3 (Fig. 2d) same length as urosomite 3,shorter than peduncle of uropod 1, shorter thanpeduncle of uropod 2; peduncle globose, wider thanramus, with two strong distal and two marginalsetae; inner ramus absent; outer ramus uniarticulate,shorter than peduncle, basal width more than twotimes tip of ramus, with three simple apical slendersetae, and one conate seta.
Telson (Fig. 3c) wider than long, entire, apicallyrounded, with more than two short simple setae,arranged symmetrically on the apical margin.
Coxal gills sac-like, on segments 2 to 6. Ventrola-teral sternal gills tubular, present on segments 3 to 7.
Characters of female that differ from male. Size8.8 mm. Antenna 1 flagellum with eight articles.Antenna 2 flagellum with ten articles. Gnathopod 1(Fig. 5a, b) similar in size and of same shape asgnathopod 2; similar to male gnathopod 1 in size andshape. Gnathopod 2 (Fig. 5c, d) smaller than malegnathopod 2 and different in shape, propodus lengthless than two times maximum width, normallysubchelate, palm transverse.
Habitat. Freshwater, epigean, normally more than10 m depth.
Distribution. Endemic to Lake Titicaca, Peru andBolivia.
Hyalella longispina n. sp. (Figs 6-11)
Type material. See above section on material andmethods.
Description of male holotype. Head (Fig. 6) longerthan peraeonite 1, rostrum absent. Eyes circular.Peraeonites and pleonites 1-3 with transverse roun-ded elevations near posterior margin. Epimera 1-3with straight posterior margin, epimeral plate 1rounded ventrally, plate 2 with rounded posteroven-
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 4
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
tral angle, plate 3 pointed posteroventrally.Urosomite 1 longest; urosomite 2 rather short, withurosomite 3 attached to the posterodorsal margin;urosomite 3 surpassed by the telson.
Antenna 1 (Fig. 6b) peduncular article ratios:1:1:0.67; flagellum damaged, at least 10 articles.Antenna 2 (Fig. 8d) peduncular article 1 massive,deeply incorporated in lateral head shield, articles 3-5 successively longer; 10 flagellar articles. Labrum(Fig. 6d) truncate, with hair-like setae close to apicalmargin. Mandible with 5-dentate incisor on the rightside (4-dentate on left); lacinia mobilis with 2 strongteeth and a few smaller denticles on the rightmandible (3 teeth on left); setal row consisting of 2long setae and a row of shorter setae; pars molarisridged with stout seta on the anterior face. Lower lip(hypopharynx) (Fig. 6f) with wide rounded lobesand short rounded mandibular processes. Maxilla 1palp uniarticulate, not reaching distal margin ofouter plate, with 1 terminal seta; outer plate with 9strongly serrate apical spiniform setae; inner plateslender with 2 apical pappose setae. Maxilla 2 (Fig.8b) inner plate slightly wider and shorter than outer,with one stout long medial seta and rows of apicalsetae shorter than those on outer plate. Maxilliped(Fig. 8c) inner plate with 3 apical robust setae; outerplate longer than inner plate, relatively slender; palparticle 2 medially produced; article 3 ovoid; article 4slender, with apical nail. Gnathopod 1 (Fig. 8a, e)coxa with strongly tapering and pointed process,directed lateroventrally (Fig. 7); basis slightly con-cave anteromarginally, convex posteroventrally;ischium subquadrate with anteromarginal notch;anterior margin of merus with articulation of carpus;carpus with setose posteromarginal rounded lobe;propodus expanded distally, two groups of setae onthe outer face, medial face with stout setae; palmwith setae of different length and one posteriorspine-like seta; dactylus stout and only weaklycurved. Gnathopod 2 (Fig. 9d) coxa directed moreventrally than those of peraeopods 1 and 3, shapesimilar to gnathopod 1 but longer (142 %), widerproximally; basis with anterior straight margin,posteriorly slightly convex with some marginalsetae; ischium subquadrate, anterior marginexpanded distally; merus subrectangular, posterodi-stally angular; carpus shorter than that of gnathopod1, with projecting, relatively narrow carpal lobe,guarding more than half length of the posteriormargin of propodus, with some terminal short setaeand a row of setae on the medial face; propodusmassive, expanded distally, palm oblique, with twoshort spine-like setae at posterior angle, palm lined
with short setae; dactylus weakly curved withmicrotrichs on inner curvature. Peraeopod 3 coxa(Fig. 9b) similar to that of gnathopods, but anteriorlydeeply excavate; basis with anterior straight margin,posteromarginally sinuous; ischium as for gnathopod2; merus slightly expanded distally and slightlydrawn out anterodistally; merus to propodus sube-qual in length, successively narrower; dactylus stoutwith apical nail. Peraeopod 4 (Fig. 9a) coxa withextremely elongate pointed process, directedlaterally, wing-like (Fig. 7), span from left to rightprocess apex longer than total body length (Fig. 12),additional short subacute process on posteriormargin close to body articulation of coxa; basis todactylus as for peraeopod 3, except for slightlylonger carpus and dactylus. Peraeopod 5 (Fig. 10a)coxa wider than long, bilobed, anterior lobeventrally rounded and slightly shorter than narrowerposterior lobe; basis tapering distally, weaklyrounded anteromarginally with groups of setae,posterior margin slightly sinuous, apical region withweak depression; ischium longer than wide, withstraight anterior margin and weak excavationposteriorly; merus somewhat expanded distally,drawn out posterodistally, with apical group ofsetae; carpus 1.12 x merus; propodus 1.28 x merus,dactylus only weakly curved. Peraeopod 6 (Fig. 10b)coxa bilobed, anterior lobe very short, angular; basissimilar in shape to that of peraeopod 5, but widerand longer; ischium as for peraeopod 5; merus topropodus similar in shape, much longer than those ofperaeopod 5, relative lengths 1.00:1.26:1.32, dac-tylus slightly longer than that of peraeopod 5.Peraeopod 7 (Fig. 10d) shorter than peraeopod 6,coxa smallest, rounded anteroventrally, posteriormargin rather straight; basis much wider than thoseof peraeopods 5-6, ovoid, lobate posteroventrally,groups of setae on anterior margin; shape of ischiumto dactylus as for peraeopod 6. Pleopods (Fig. 10c)normal. Uropod 1 (Fig. 11f) peduncle longer thaninner ramus, outer ramus slightly shortened;peduncle and rami sparsely setose, rami withterminal setae of different length. Uropod 2 (Fig.11c) peduncle as long as inner ramus, outer ramusslightly shortened compared to inner, with terminalsetae only. Uropod 3 reduced in length, pedunclelonger than ramus, only one ramus present, withsome terminal setae. Telson wider than long, entirewith some marginal short setae (Fig. 11g).
Characters of female allotype that differ frommale holotype. Size 9 mm. Gnathopod 2 (Fig. 11a,b) resembles gnathopod 1 (Fig. 11e, d) in the wide
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 5
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
and strongly setose carpal lobe. Propodus ofgnathopod 2 very similar to that of gnathopod 1.
Habitat. Freshwater, epigean, more than 10 mdepth.
Distribution. Endemic to Lake Titicaca, Peru andBolivia.
Discussion
The high diversity of species of Hyalella in the lakeis unexpected and not well understood (Crawford etal., 1993). The amphipod fauna in Lake Titicaca hasbeen compared with the fauna of Lake Baikal andthe speciation events considered similar. However, itis important to consider that Lake Titicaca is not asold as Lake Baikal. According to Lavenu (1992), thepresent lake system on the Altiplano is derived froma more ancient system which was there since thelower Pleistocene, no more than two millions yearsago. Bousfield (1982) suggested that the endemicfauna of the lake could not be older than the lakebasin, estimated by him as originating 10 millionyears ago.
Similar to Lake Baikal also in Lake Titicacaseveral amphipod species have body processes. It isconceivable that these pointed processes act as adefense mechanism against predation as there aremore than 28 species of fish in the lake.
When we started to work on H. armata we foundstriking differences in the length of the coxalacumination within the extensive samples collectedby George Crawford. At a closer look and after amorphometric analysis of the length/width ratio ofthe animals (Fig. 13), a discontinuity between thesetwo populations became apparent. These differenceswere used to separate two distinct species. Bothspecies are very different from all hyalellids knownso far. They bear pointed coxal plates that aredirected laterally, which is unique in the Hyalellidae.
We searched for further characters, such as setalpatterns and length relations of articles ofappendages, in order to find additional morphologi-cal and morphometrical differences between thesegroups. However, we only found a large variabilityin these characters. Nevertheless we consider thenon-overlapping character clouds of the length/widthratios as indicators of genetic separation of thesepopulations. There is no geographical or obvioushabitat separation of both populations, but theyinhabit various locations in Lake Titicaca (Fig. 14).In spite of a potential contact of these populations,
there is evidently no intermediate character state ofthe length/width ratio in these populations.
The low number of morphological charactersseparating these two species perhaps reflects theirrecent origin. Without success we tried to extractDNA from the museum samples in order to getgenetic information on these populations. With freshmaterial we would like to check in a future study ifthere are more characters on the molecular level.
A comparable similarity between hyalellid popu-lations in Lake Titicaca occurs in the H. echinuscomplex we are currently working on.
Acknowledgements
Dr. Ardis Johnston from the Museum ofComparative Zoology, Harvard University, loanedFaxon’s material. This work was largely funded byUniversidad Católica del Norte, Chile, andUniversity of Maine, USA. Thanks also to Dr. C.Dejoux for providing specimens for this study. Ms.Jaqueline Meier made the drawings of H. longispina.
References
Barnard, J. L., & Barnard., C. M. (1983): FreshwaterAmphipoda of the World. vol. I. EvolutionaryPatterns, vol. II. Handbook and Bibliography. 830pp., Hayfield Associates, Mt. Vernon, Virginia.
Bousfield, E. L. (1982): Amphipoda: paleohistory.Pp. 96-100 in: McGraw-Hill Yearbook of Scienceand Technology for 1982-1983.
Bousfield, E. L. (1996): A contribution to the reclas-sification of Neotropical freshwater hyalellidamphipods (Crustacea: Gammaridea, Talitroidea).Boll. Mus. Civ. Storia Nat. Verona 20: 175-224.
Chevreux, E. (1904): Mission de Crequi-Monfort etSenegal de la Grange. Note preliminaire sur lesamphipodes recueillis par M. le Dr. Neveu-Lemaire dans le Lac Titicaca. (Julliet, 1903).Bull. Soc. Zool. Fr. 29: 131-134, 2 figures.
Chevreux, E. (1907): Les amphipodes des lacs deshauts plateaux de l’Amerique du Sud. MissionScientifique G. de Crequi-Monfort et Senegal dela Grange, Lacs Hauts Plateaux de l’Amerique duSud. 22 pp, figs 30-41.
Crawford, G. I., Harrison, K., Lincoln, R. J. &Boxshall, G. A. (1993): An introduction to thespecies flock of amphipods (Crustacea) of LakeTiticaca. Workshop Speciation in Ancient Lakes.Evolution, Biodiversity, Conservation. Scientific
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 6
Station of the Hautes-Fagnes, Mont-Rigi,Robertsville, Belgium.
Dejoux, C., (1992): The Amphipoda. Pp. 346-356in: Dejoux, C. & Iltis, A. (eds) Lake Titicaca. ASynthesis of Limnological Knowledge. KluwerAcademic Publishers, Dordrecht, TheNetherlands.
Della Valle, A. (1893): Gammarini del Golfo diNapoli. Fauna Flora Golf. Neapel angrenz. Mee-res-Abschn., Monogr. 20: 1-948, 61 pls.
Faxon, W. (1876): Exploration of Lake Titicaca byAlexander Agassiz and S. W. Garman. IV.Crustacea. Bull. Mus. Compar. Zool. 3: 361-375,37 figs.
Lavenu, A. (1992): Origins, formation andgeological evolution. Pp. 3-15 in: Dejoux, C. &Iltis, A. (eds) Lake Titicaca. A Synthesis ofLimnological Knowledge. Kluwer AcademicPublishers, Dordrecht, The Netherlands.
Stebbing, T. R. R. (1888): Report on the Amphipodacollected by H. M. S. Challenger during the years1873-1876. Rep. Sci. Res. Voy. H. M. S.Challenger 1873-1876, Zool. 29: 1-1737, 210 pls.
Stebbing, T. R. R. (1906): Amphipoda I. Gammaridea.Das Tierreich 21: 1-806.
Watling, L. (1993): Functional morphology of theamphipod mandible. J. Nat. Hist. 27: 837-849.
Fb
ig. 1. Hyalella armata (Faxon, 1876), male 9.5 mm. a: left aspect of habitus; b: epimeral plates 1-3; c: gnathopod 2; d: gnathopod 1. Scalears: a, b = 727 µm; c, d = 192 µm.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 7
F -p
ig. 2. Hyalella armata (Faxon, 1876), male 9.5 mm. a: maxilla 1; b: peraeopod 6; c: palp of maxilla 1; d: uropod 3; e: peraeopod 7; f: urood 1. Scale bars: a, c, d = 94 µm; b, e, f = 192 µm.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 8
FS
ig. 3. Hyalella armata (Faxon, 1876), male 9.5 mm. a: left maxilliped. b: maxilla 2; c: telson; d: uropod 2; e: peraeopod 4; f: peraeopod 5.cale bars: a, b = 94 µm; c-f = 192 µm.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 9
Ffµ
ig. 4. Hyalella armata (Faxon, 1876), male 9.5 mm. a: gnathopod 1 detail; b: peraeopod 3; c: lower lip (hypopharynx); d: antenna 1, distallagellomeres omitted; e: antenna 2, distal flagellomeres omitted; f: upper lip (labrum); g: left mandible. Scale bars: a = 94 µm; b-g = 192m.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 10
FS
ig. 5. Hyalella armata (Faxon, 1876), female “f” 8.8 mm. a: gnathopod 1; b: chela of gnathopod 1; c: gnathopod 2; d: chela of gnathopod 2.cale bars: a, d = 192 µm; b, c = 94 µm.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 11
Fl
ig. 6. Hyalella longispina n. sp., holotype male, 10 mm. a: habitus; b: antenna 1; c: left mandible; d: upper lip (labrum); e: right mandible; f:ower lip (hypopharynx). Scale bars a = 1 mm; b, c, e = 100 µm; d, f = 200 µm.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 12
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
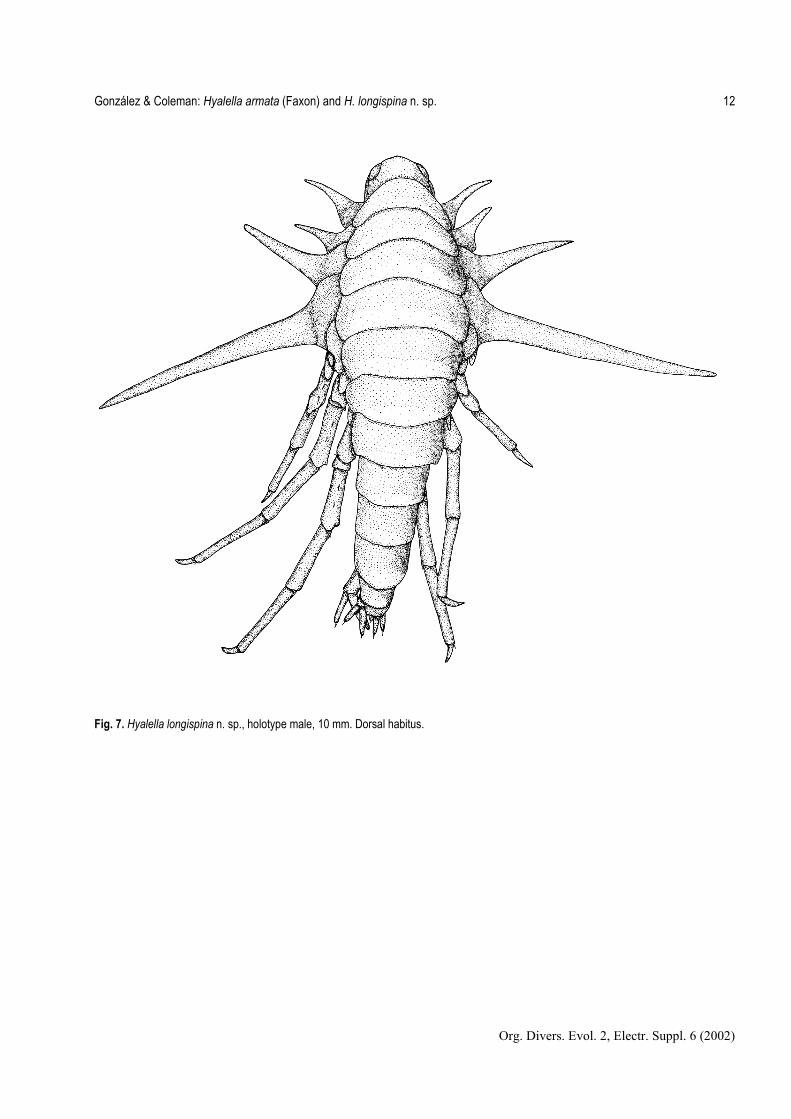
Fig. 7. Hyalella longispina n. sp., holotype male, 10 mm. Dorsal habitus.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 13
Fa
ig. 8. Hyalella longispina n. sp., holotype male, 10 mm. a: gnathopod 1 detail; b: maxilla 2; c: maxilliped, palp of right side omitted; d:ntenna 2; e: gnathopod 1; f: maxilla 1. Scale bars: a-d, f = 100 µm, e = 200 µm.
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 14
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
Fig. 9. Hyalella longispina n. sp., holotype male, 10 mm. a: peraeopod 4; b: coxa of peraeopod 3; c: basis to dactylus of peraeopod 3; d:gnathopod 2, setation of medial face of carpal lobe omitted. Scale bars: a-d = 200 µm.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 15
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 16
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
Fig. 11. Hyalella longispina n. sp., a, b, e, d; female allotype, 9 mm; c, f, g male holotype, 10 mm. a: chela of peraeopod 2; b: peraeopod 2;c: uropod 2; d: peraeopod 1; e: chela of peraeopod 1; f: uropod 1; g: uropod 3 and telson. Scale bars: a, e = 100 µm; b-d, f, g = 200 µm.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 17
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
Fig 12. Hyalella armata complex. a. H. armata, dorsal habitus aspect. b. H. armata, left lateral view. c. H. longispina n. sp.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 18
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
5
6
7
8
9
10
11
0 2 4 6 8 10 12 14
animal length
wid
th (c
oxae
4)
armata
longispina
Fig. 13. Body length vs. width for two populations of the Hyalella armata complex, n=40. The two separate clouds of dots show the distinc-tion between H. longispina n. sp. (black triangles) from Choccocoyo-Bay, GIC 481, and H. armata (grey diamonds) from Coata Bay, GIC263.
González & Coleman: Hyalella armata (Faxon) and H. longispina n. sp. 19
Org. Divers. Evol. 2, Electr. Suppl. 6 (2002)
Fig. 14. Map of Lake Titicaca with collecting sites of Hyalella armata (circles) and Hyalella longispina (diamonds).