Hydrocortisone effect on hyaluronate synthesis in aself-assembled human dermal equivalentMadhura Deshpande1, Suzanne Papp2, Lana Schaffer2 and Tara Pouyani1*1Department of Pharmaceutical Sciences, Bouvé College of Health Sciences, Northeastern University, Boston, MA, USA2DNA Array Core Facility, Scripps Research Institute, La Jolla, CA, USA
Glucocorticoids are used extensively in the treatment ofskin diseases (Murphy and Reich, 2011; Crowley, 2010;Zivanovic et al., 2011). They have been reported in varioussystems to reduce the tissue content of glycosaminoglycans(GAGs), especially hyaluronic acid (hyaluronate,hyaluronan, HA), in some cases inducing dermal atrophy,leading to thinning of the skin (Lubach et al., 1989;Hoffmann et al., 1998; Averbeck et al., 2010; Gebhardtet al., 2010). Their many anti-inflammatory effects are at-tributed to the suppression of T cell proliferation,
suppression of pro-inflammatory cytokines and thedownregulation of certain adhesion molecules (Stuhlmeierand Pollaschek, 2004). The effects of glucocorticoids ondermal fibroblasts in cell culture have also been studied,leading to the conclusion that suppression of hyaluronatesynthase 2 (HAS2) is responsible for inhibition of HAsynthesis (Zhang et al., 2000). Addition of glucocorticoidsto cultured human skin fibroblasts was seen to depress therate of cell proliferation (Ponec et al., 1979). Hydrocortisone(HC), specifically, has also been shown to reduce the rate ofcollagen synthesis in fibroblasts derived from normal humandermis (Russell et al., 1989; Booth et al., 1982).
Human dermal matrix is a self-assembled dermal equiva-lent produced under serum-free conditions and without theaid of a three-dimensional (3D) scaffold. Its preparationand characterization have been previously described in
*Correspondence to: T. Pouyani, Department of PharmaceuticalSciences, Bouvé College of Health Sciences, Northeastern University,Boston, MA 02115, USA. E-mail: [email protected]
JOURNAL OF TISSUE ENGINEERING AND REGENERATIVE MEDICINE RESEARCH ARTICLEJ Tissue Eng Regen Med (2013)Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/term.1809
detail (Pouyani et al., 2009, 2012). It bears strong biochem-ical and morphological resemblance to the human dermis.One of its outstanding characteristics is that it contains highamounts of hyaluronic acid (Table 1). Hyaluronate is animportant multifaceted molecule that plays an importantrole in a number of biological processes (Toole, 2004;Wanget al., 2011; Prestwich, 2011; Toole and Slomiany, 2008).The high HA content of the human dermal matrix maycontribute to rapid wound closure and enhanced vasculari-zation, and may lead to minimization of scarring during thewound healing process (Stern and Maibach, 2008; Jianget al., 2007). Because human dermal matrix is grown andmatures under completely serum-free conditions, it offersan excellent opportunity for the use of different concentra-tions of exogenous compounds to alter its extracellularmatrix composition.
Given the importance of hydrocortisone in dermatolog-ical therapy, we designed experiments to test the effects ofvarying concentrations of hydrocortisone on the matura-tion in culture of the human dermal matrix over a21 day period. Our objectives in designing these experi-ments were two-fold: first, we sought to prepare dermalequivalents with varying amounts of hyaluronate andcharacterize these self-assembled dermal skin constructs;second, we aimed to decipher mechanistically the actionsof the hormone hydrocortisone on HA biosynthesis in thehuman dermal matrix, as a model for the actions of hydro-cortisone on the human dermis. To accomplish thesegoals, we cultured neonatal dermal fibroblasts in the pres-ence of varying amounts of hydrocortisone, using the pre-viously described technique of self-assembly (Pouyaniet al., 2009) under completely serum-free conditions andwithout the interference of a 3D scaffold. The resultanthuman dermal matrices were characterized. We observedthat hydrocortisone exerts both a stimulatory and an inhibi-tory effect on hyaluronate synthesis in the human dermalmatrices, based on the concentration used. We observed nochange in other glycosaminoglycan synthesis at lower and in-termediate concentrations; however, some GAG inhibitionwas observed at higher concentrations of hydrocortisone.We chose the human dermal matrix cultured under a higherconcentration of hydrocortisone (500× HC=200μg/mlHC), where significant inhibition of HA was observed, andconducted a differential gene array analysis in comparisonto the construct cultured under physiological levels of HC(normal, 0.4g/μml HC). Differential expression of 43 geneswas observed; of these, 25 genes were upregulated and 18were downregulated.
2. Materials and methods
2.1. Production of human dermal equivalents
Neonatal human dermal fibroblasts (ATCC; passage 7)were harvested at confluence and seeded at high density(3 million cells/4.52 cm2) on a porous polycarbonatemembrane in a transwell format (Corning, Big Flats, NY,USA) for 21days in the presence of serum-free medium.Specifically, the formulation contains the following ingredi-ents: a base 3:1 mixture of Dulbecco’s modified Eagle’s me-dium (DMEM; high-glucose formulation withoutL-glutamine) and Ham’s F12 medium (Sigma-Aldrich, St.Louis, MO, USA) supplemented with 4mML-glutamax(Invitrogen, Carlsbad, CA, USA), 5 ng/ml epidermal growthfactor (Millipore, Billerica, MA, USA), 0.4μg/mlhydrocortisone (Sigma-Aldrich), 1×10–4M ethanolamine(Sigma-Aldrich), 1×10–4M O-phosphoryl-ethanolamine(Sigma-Aldrich), 5μg/ml insulin (Sigma-Aldrich), 5μg/mltransferrin (Sigma-Aldrich), 20pM triiodothyronine,6.78ng/ml selenium, 50μg/ml L-ascorbic acid-2-phosphate(Asc-2-P; Wako Chemicals, Richmond, VA, USA), 0.2μg/ml L-proline and 0.1μg/ml glycine. The lower chamberwas fed 3ml and the upper chamber 2ml of mediumthroughout the 21day culture period, with mediumchanges every 2–3days. Hydrocortisone in varying concen-trations was added to the medium, using 95% ethanol asa co-solvent.
2.2. Cell count procedure
Human dermal matrices were digested with collagenaseIV (Sigma-Aldrich) for 1 h at 37oC in collagenase premix(120ml PBS, 14ml 2.5% trypsin and 1ml 0.45% glucose)at the end of the 21 day culture period. Serum-containingmedium was added to the digest to stop further enzymaticactivity. The cells were diluted 1:1 with trypan blue(Sigma-Aldrich) and counted using a haemocytometer.
2.3. Determination of total collagen
Total collagen was quantified using the hydroxyprolineassay. The constructs were placed in 6 N hydrochloric acid(HCl) in a sealed glass tube and hydrolysed for 18h at110oC (Brown et al., 2001). At this point all the collagenand proteins present were degraded to the componentamino acids. After completion of hydrolysis, the hydroly-sates were partially neutralized with 5 N NaOH andcentrifuged to sediment particulate debris. The superna-tants were transferred to fresh tubes for further analysis.The standards, blanks and unknowns (with appropriatedilutions) were pipetted in triplicate, in 50μl aliquots, ontoa 96-well plate. Oxidizing solution (100μl) containingchloramine T (Sigma-Aldrich) was added to each well,except those designated as blanks; to those wells oxidizingbuffer solution without chloramine Twas added. The platewas gently shaken (on a laboratory rotator/shaker) for
Table 1. Hyaluronic acid content of human dermal matrix com-pared to adult human skin, neonatal foreskin and fetal skin
*HA content is reported as μg HA/mg wet weight of tissue.$Results for HA content were obtained from a competitiveinhibition ELISA and are reported±SEM, n=3.
5min at room temperature. Ehrlich’s reagent (100μl) wasadded to each well with thorough mixing. The plate wascovered with an adhesive plate seal and incubated in awater bath or oven at 60oC for 45min. The absorbancewas then read at 570nm. The hydroxyproline values(expressed in μg/ml) were determined directly from thestandard curve. In order to convert the hydroxyprolinevalues to the total amount of collagen present in theconstructs, a conversion factor of 7.46 was used. The finaldata were expressed as μg collagen/mg wet weight oftissue, or μg collagen/construct.
2.4. Hyaluronic acid inhibition ELISA assay
Human dermal matrix samples were digested with0.5mg/ml Proteinase K (Invitrogen) overnight at 60oC.The digested mixture was boiled for 10min to inactivatethe protease, spun down and the supernatant transferredto a fresh tube. ELISA plates (Immulon4, Thermo FisherScientific, Waltham, MA, USA) were prepared by coatingovernight with a 100μg/ml solution of sodium hyaluronate(Genzyme, Cambridge, MA, USA) in sodium bicarbonatebuffer, pH9.2. The plate was then washed with 0.1%bovine serum albumin (BSA) in phosphate-buffered sa-line (PBS) and blocked with 5% BSA for 6 h. Samplesto be tested were appropriately diluted with 1% BSAand 100 μl of each were added to individual wells. Asolution of biotinylated HA-binding protein (1:1000 di-lution; Associates of Cape Cod, East Falmouth, MA,USA) was added to the samples and allowed toequilibriate at 4oC overnight. The wells were washedwith 0.05% Tween/PBS and avidin–horseradish peroxi-dase (1:5000; Vector Laboratories, Burlingame, CA,USA) was added to each well and incubated at 37oCfor 1 h. The plates were washed, the substrate ABTS(Kirkegaard and Perry Laboratories, Gaithersburg, MD,USA) was added to each well and the colour was allowedto develop for 10min. The reaction was stopped by the ad-dition of 1% SDS to each well. The plates were read in amicrotitre plate reader at 410nm.
2.5. Glycosaminoglycan (GAG) disaccharideanalysis
Proteinase K-digested samples of human dermal matriceswere initially digested with 20 U/ml hyaluronidase SD(Associates of Cape Cod) in 100mM ammonium acetate,pH7.00, for 2 h at 37oC, followed by chondroitinase ABC(Associates of Cape Cod) digestion for 3 h. The digestsand disaccharide standards (Associates of Cape Cod) weresubjected to reductive amination and conjugation with 2-aminoacridone (AMAC; Invitrogen) to produce fluorescentGAG disaccharides. The labelled GAGs were subjected topolyacrylamide gel electrophoresis (PAGE) and imagedusing a ChemiDoc XRS imager (BioRad, Hercules, CA,USA) to obtain the demonstrated profiles. For quantifica-tion, AMAC-labelled glucose (Sigma-Aldrich) was used as
a standard, Quantity One software v. 4.6.1 (BioRad) wasused to determine various GAG disaccharide (ΔDi-HA,ΔDi-4S, ΔDi-6S, ΔDi-4,6S) values.
2.6. Histological processing
Samples to be analysed were fixed overnight in 10%phosphate-buffered formalin (Thermofisher Scientific,Pittsburgh, PA, USA). They were subsequently transferredto vials containing 70% ethanol prior to embedding andprocessing. The samples were embedded into paraffinblocks, sectioned at 6μm and stained with haematoxylinand eosin (H&E), imaged and photographed at×400,using a Westover Micromaster I microscope equipped witha digital camera.
2.7. Transmission electron microscopy (TEM)
Human dermal matrices (4.52 cm2) were fixed overnight at4oC in 2.5% glutaraldehyde in 0.1M sodium cacodylatebuffer, pH7.4 (Electron Microscopy Sciences, Hatfield, PA,USA), washed in 0.1M cacodylate buffer and post-fixedwith 1% osmium tetroxide (OsO4)/1.5% potassium ferrocy-anide (KFeCN6) for 1 h, washed inwater three times and in-cubated in 1% aqueous uranyl acetate for 1 h, followed bytwo washes in water and subsequent dehydration in gradesof alcohol (10min each; 50%, 70%, 90%, 2×10min100%). The samples were then put in propyleneoxide for1 h and infiltrated overnight in a 1:1 mixture ofpropyleneoxide and TAAB Epon (Marivac Canada, St.Laurent, Canada). The following day the samples were em-bedded in TAAB Epon and polymerized at 60oC for 48h.Ultrathin sections (about 60nm) were cut on a ReichertUltracut-S microtome, picked up on to copper grids stainedwith lead citrate and examined in a Jeol 1200EX transmis-sion electron microscope or a TecnaiG2 Spirit BioTWIN.Images were recorded using an AMT 2k CCD camera.
2.8. Measurement of tissue thickness
The thicknesses of H&E-stained cross-sections of humandermal matrices were measured by digital image analysis,using Micron imaging software (one sample, different fieldof view). The measurements are made across the sections,perpendicular to the base of the construct. The reportedthickness is a mean of 10 measurements± standard errorof the mean (SEM).
2.9. Statistical analyses
Statistical analysis of the data was done using one-wayANOVA, with equal variances assumed. Multiple compari-sons were done by the Bonferroni method. StatisticalPackage for Social Sciences (SPSS) software was used forconducting the one-way ANOVA significance tests. Thesignificance level (α) used was 0.05. Probability values of
Hydrocortisone effect on hyaluronan synthesis in a dermal equivalent
p< 0.05 were considered statistically significant. Experi-ments were repeated three times, and data are expressedas mean±SEM unless otherwise indicated.
2.10. Total RNA extraction
Total RNA was extracted from human dermal matrix usingthe Qiagen RNeasy® Fibrous Tissue Midi Kit (Qiagen,Valencia, CA, USA). Dermal matrices were placed in 15mlcentrifuge tubes with 3ml RLT buffer (provided in the kit)and 10μl β-mercaptoethanol (Sigma-Aldrich) and homoge-nized using a TissueRuptor homogenizer (Qiagen) for1min. The homogenized tissue was treated with ProteinaseK for 15min at 55oC and subjected to centrifugation at3500× g for 5min at 20–25oC. The supernatant wastransferred to a new 15ml centrifuge tube and 0.5 volumesof ethanol (Sigma-Aldrich) was added to the lysate. Thesample was then transferred to an RNeasyMidi spin columnplaced in a 15ml collection tube, 3ml at a time, and eachtime centrifuged for 5min at 3500× g. The flow-throughwas discarded. The column membrane was washed withRW1 wash buffer and centrifuged. DNAse digestion wascarried out by addition of 160μl DNase I incubation mix(20μl DNase I stock solution in 140μl RDD buffer) to thespin column membrane for 18min at room temperature.The column was washed with RW1 buffer and centrifugedfor 5min at 3500× g. Two other washes were also carriedout with RPE buffer. The RNA was eluted by adding150μl RNAse-free water twice, directly to the spin columnmembrane, and centrifuged each time for 3min at3500× g. The RNA samples were monitored using a 2100Agilent Bioanalyser, consistently demonstrating high-quality RNA, as evidenced by the 28S:18S ratio ofapproximately 2.
2.11. Gene array analysis
The GlycoV4 oligonucleotide array is a custom GeneChip(Affymetrix, Santa Clara, CA, USA), designed for theConsortium for Functional Glycomics (http://www.functionalglycomics.org/static/consortium/consortium.shtml;a complete description and annotation for the GlycoV4array is available at: http://www.functionalglycomics.org/static/consortium/resources.shtml). The GlycoV4focused array includes probes for ~1260 human probe-IDsand ~1200 mouse probe-IDs related to glycogenes. Thisarray does not contain mismatched probes.
Data normalization was performed using RMA Express1.0, with quantile normalization, median polish andbackground adjustment (Bolstad et al., 2003; Irizarryet al., 2003).
The Limma package in the R software was used to findtranscripts with differential expression (Benjamini andHochberg, 1995; Smyth, 2004). The fold changes andstandard errors were estimated by fitting a linear modelfor each gene, and empirical Bayes smoothing was appliedto the standard errors. Results are presented between two
or more experimental conditions as a fold change inexpression level, the moderated t statistic, the p valueand the adjusted p value. The adjusted p value is the pvalue adjusted for multiple testing using Benjamini andHochberg’s method. The transcripts identified as differen-tially expressed were those with adjusted p value< 0.10and fold change> 1.4.
The heat map (Figure 5) was generated using the dChipprogram (www.dChip.org). Red indicates increasedexpression and blue indicates decreased expression relativeto the mean transcript expression value.
3. Results
3.1. Preparation of self-assembled humandermal matrices
Neonatal human dermal fibroblasts were cultured in astate of ’super-confluence’ on a polycarbonate membrane
Figure 1. H&E-stained sections of human dermalmatrices grown inthe presence of varying concentrations of hydrocortisone. Thenormal construct is grown in the presence of 0.4μg/ml HC; otherconcentrations are stated as a multiple of this base value (shown tothe left of the H&E sections); thicknesses of constructs are shownto the right: *p<0.05 for comparison with normal constructthickness, using one-way ANOVA at α=0.05
under completely serum-free conditions, as previously de-scribed (Pouyani et al., 2009). These cells were culturedfor 21 days, during which they were fed every 2–3 dayswith medium supplemented with growth factors, hor-mones and vitamins. In order to assess the effect of hydro-cortisone on HA/GAG and extracellular matrix synthesis,varying concentrations of HC, as multiples of the basevalue (0.4 μg/ml), were added to the culture mediumfrom the initial time of culture until maturation 21 dayslater. Specifically, the following multiples of the normalconstruct were chosen: 30× (12μg/ml); 60× (24μg/ml);200× (80μg/ml); 500× (200μg/ml); 750× (300μg/ml); and 1000× (400μg/ml). After the 21day cultureperiod, a substantial extracellular matrix had formed andthe human dermalmatriceswere peeled off the polycarbon-ate membrane and analysed.
3.2. Histological analysis of humandermal matrices
Human dermal matrices cultured under varying concentra-tions of hydrocortisone were formalin-fixed and paraffin-embedded sections were subjected to H&E staining.H&E-stained sections of all human dermalmatrices demon-strated the presence of fibroblasts dispersed in a densecollagenous matrix (Figure 1). The normal human dermalmatrix had a thickness of 281.3±13.01μm. The 30× and60× human dermal matrices showed an enhancedthickness of 323.6±4.50 and 329.6±4.08μm, respec-tively. The 200× human dermal matrix showed a returnto a thickness similar to that of the normal construct,290.6±6.76μm. Human dermal matrices cultured underhigher concentrations of hydrocortisone showed a
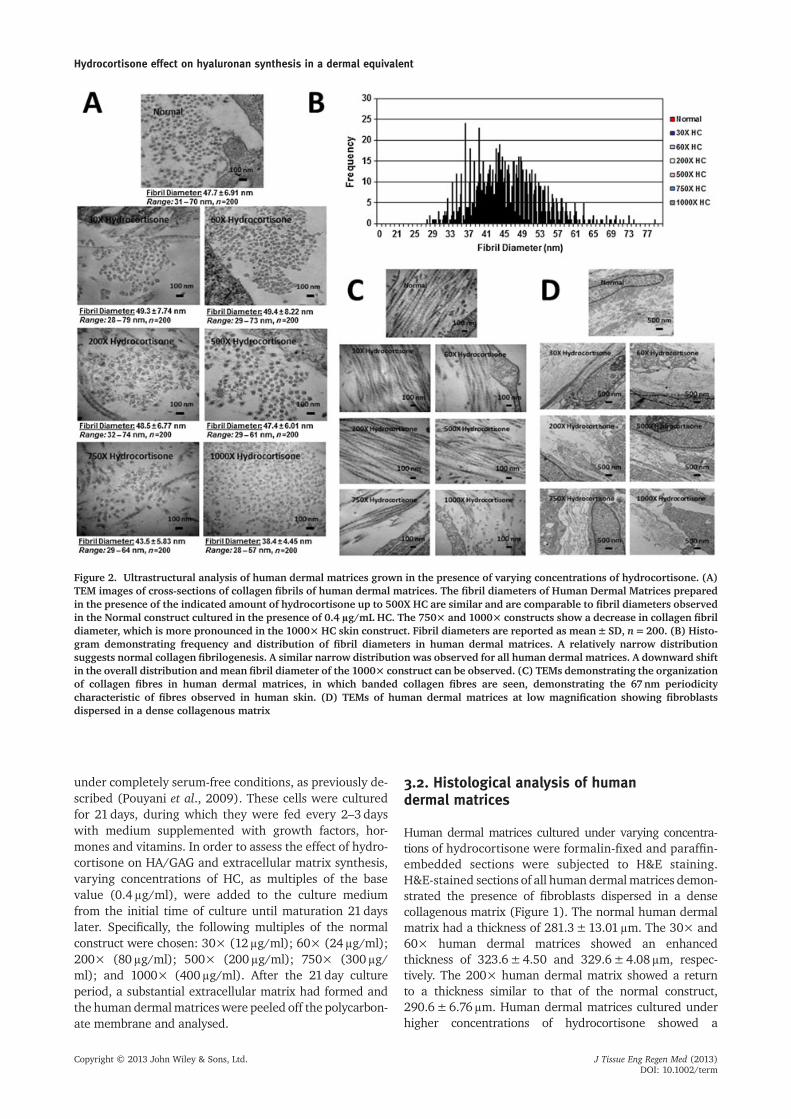
Figure 2. Ultrastructural analysis of human dermal matrices grown in the presence of varying concentrations of hydrocortisone. (A)TEM images of cross-sections of collagen fibrils of human dermal matrices. The fibril diameters of Human Dermal Matrices preparedin the presence of the indicated amount of hydrocortisone up to 500X HC are similar and are comparable to fibril diameters observedin the Normal construct cultured in the presence of 0.4 μg/mL HC. The 750× and 1000× constructs show a decrease in collagen fibrildiameter, which is more pronounced in the 1000× HC skin construct. Fibril diameters are reported as mean±SD, n=200. (B) Histo-gram demonstrating frequency and distribution of fibril diameters in human dermal matrices. A relatively narrow distributionsuggests normal collagen fibrilogenesis. A similar narrow distribution was observed for all human dermal matrices. A downward shiftin the overall distribution andmean fibril diameter of the 1000× construct can be observed. (C) TEMs demonstrating the organizationof collagen fibres in human dermal matrices, in which banded collagen fibres are seen, demonstrating the 67nm periodicitycharacteristic of fibres observed in human skin. (D) TEMs of human dermal matrices at low magnification showing fibroblastsdispersed in a dense collagenous matrix
Hydrocortisone effect on hyaluronan synthesis in a dermal equivalent
progressive thinning: 119.9±4.63μm for the 500×construct and 61.3±2.69μm for the 750× construct. The1000× human dermal matrix was composed of only a fewcell layers with a thickness of 15.5±1.12μm (Figure 1).
3.3. Ultrastructural analysis of humandermal matrices
The self-assembled human dermal matrices prepared in thepresence of varying amounts of hydrocortisonewere subjectedto ultrastructural analysis by transmission electronmicroscopy(TEM). Analysis of cross-sections of collagen fibril diametersdemonstrated that therewas no appreciable difference infibrildiameter in the 30×, 60×, 200× and 500× constructs ascompared to the normal construct containing physiologicallevels of hydrocortisone (0.4g/ml; Figure 2A). The 750×human dermal matrix showed a small decrease in collagenfibril diameter at 43.5±5.83nm, compared to the normalconstruct with a fibril diameter of 47.7±6.91nm. The1000× human dermal matrix showed an appreciable declinein collagen fibril diameter, at 38.4±4.45nm, compared tothe normal construct (Figure 2A). The statistical distributionof the collagen fibril diameters was studied; all constructsshowed a relatively tight distribution, suggesting normal colla-gen fibrillogenesis (Figure 2B). At higher magnifications, allhuman dermal matrices grown in the presence of hydrocorti-sone demonstrated the presence of longitudinal collagen fi-bres exhibiting the quarter-staggered 67nm periodicitycharacteristic of fibres in human skin (Figure 2C)(Fleischmajer et al., 1990). This arrangement of collagen fibreswas also seen in thenormal humandermalmatrix (Figure2C).TEM images of the human dermal matrices at lower magnifi-cations (Figure 2D) showed the presence offibroblasts embed-ded in a collagenous matrix analogous to that observed in thenormal human dermal matrix (Figure 2D).
3.4. Analysis of hyaluronic acid and collagencontent
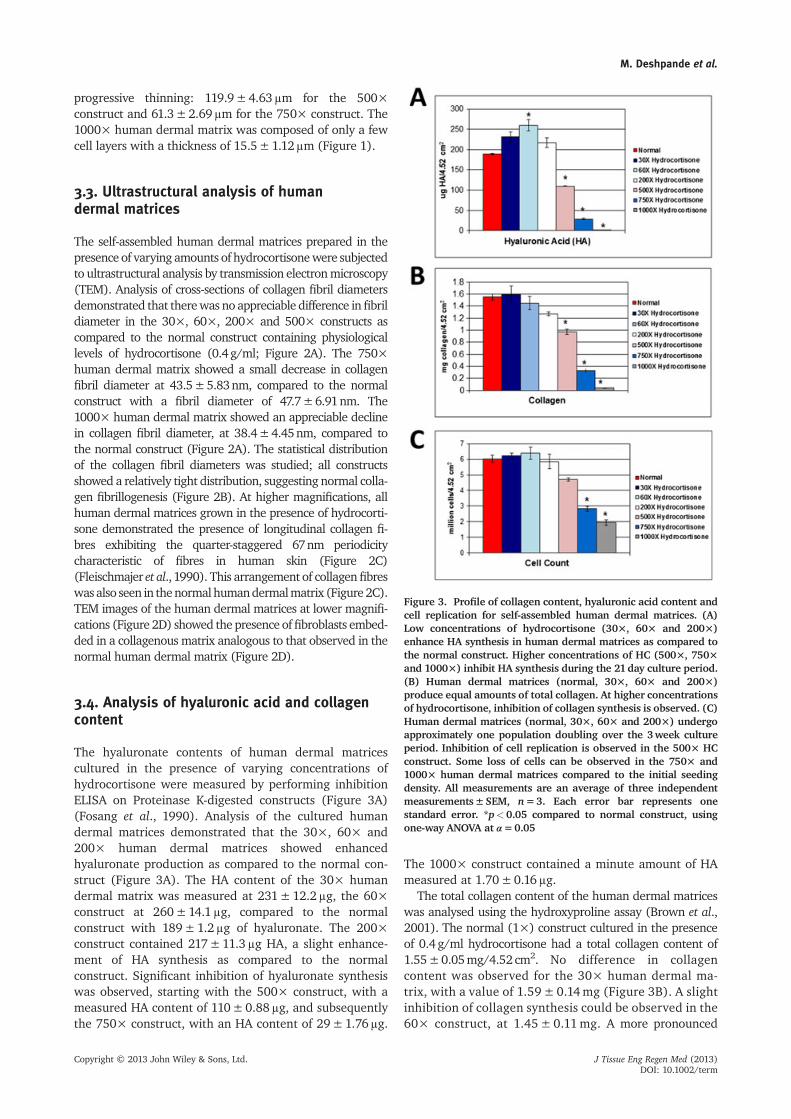
The hyaluronate contents of human dermal matricescultured in the presence of varying concentrations ofhydrocortisone were measured by performing inhibitionELISA on Proteinase K-digested constructs (Figure 3A)(Fosang et al., 1990). Analysis of the cultured humandermal matrices demonstrated that the 30×, 60× and200× human dermal matrices showed enhancedhyaluronate production as compared to the normal con-struct (Figure 3A). The HA content of the 30× humandermal matrix was measured at 231±12.2μg, the 60×construct at 260±14.1μg, compared to the normalconstruct with 189±1.2μg of hyaluronate. The 200×construct contained 217±11.3μg HA, a slight enhance-ment of HA synthesis as compared to the normalconstruct. Significant inhibition of hyaluronate synthesiswas observed, starting with the 500× construct, with ameasured HA content of 110±0.88μg, and subsequentlythe 750× construct, with an HA content of 29±1.76μg.
The 1000× construct contained a minute amount of HAmeasured at 1.70±0.16μg.
The total collagen content of the human dermal matriceswas analysed using the hydroxyproline assay (Brown et al.,2001). The normal (1×) construct cultured in the presenceof 0.4g/ml hydrocortisone had a total collagen content of1.55±0.05mg/4.52 cm2. No difference in collagencontent was observed for the 30× human dermal ma-trix, with a value of 1.59± 0.14mg (Figure 3B). A slightinhibition of collagen synthesis could be observed in the60× construct, at 1.45± 0.11mg. A more pronounced
Figure 3. Profile of collagen content, hyaluronic acid content andcell replication for self-assembled human dermal matrices. (A)Low concentrations of hydrocortisone (30×, 60× and 200×)enhance HA synthesis in human dermal matrices as compared tothe normal construct. Higher concentrations of HC (500×, 750×and 1000×) inhibit HA synthesis during the 21day culture period.(B) Human dermal matrices (normal, 30×, 60× and 200×)produce equal amounts of total collagen. At higher concentrationsof hydrocortisone, inhibition of collagen synthesis is observed. (C)Human dermal matrices (normal, 30×, 60× and 200×) undergoapproximately one population doubling over the 3week cultureperiod. Inhibition of cell replication is observed in the 500× HCconstruct. Some loss of cells can be observed in the 750× and1000× human dermal matrices compared to the initial seedingdensity. All measurements are an average of three independentmeasurements±SEM, n=3. Each error bar represents onestandard error. *p<0.05 compared to normal construct, usingone-way ANOVA at α=0.05
inhibition was observed with the 200× human dermalmatrix, with a collagen content of 1.27±0.03mg/con-struct. Higher levels of collagen synthesis inhibition were ob-served with the 500× construct at 0.974±0.043mg. The750× and 1000× human dermal matrices had0.329±0.024 and 0.04±0.004mg collagen/4.52 cm2,respectively, and showed the highest levels of dermalthinning.
3.5. Cell replication analysis
Previous research had indicated that cultured neonataldermal fibroblasts of the human dermal matrix (normal)undergo one population doubling during a 21day cultureperiod (Pouyani et al., 2009). The effects of different con-centrations of hydrocortisone on cell replication were in-vestigated. All constructs were uniformly seeded with 3million cells/4.52 cm2 area. At the end of the cultureperiod, the normal construct had 6.03±0.23 millioncells. We observed that after 21 days, the 30× and 60×human dermal matrices showed a very slight increase incell number compared to the normal construct
(Figure 3C); the 30× and 60× dermal constructs had6.23±0.18 and 6.39±0.4 million cells, respectively. Aslight inhibition was observed with the 200× construct,with a cell count of 5.86±0.46 million at the end of the21 day culture period. A higher inhibition of cell replica-tion was seen with the 500× human dermal matrix, witha final cell count of 4.71±0.09 million. The 750× and1000× constructs showed the presence of fewer cells,with 2.83±0.15 and 1.95±0.17 million cells, respec-tively, at the end of the 21 day culture period, comparedto the initial seeding density.
3.6. Glycosaminoglycan content analysis ofhuman dermal matrices by FACE
The glycosaminoglycan (GAG) profile of the human dermalmatrices was studied by fluorophore-activated carbohydrateelectrophoresis (FACE) (Calabro et al., 2000a, 2000b). Theconstructs were digested with proteinase K in order to liber-ate the GAGs. The GAGs were then digested with hyaluroni-dase and chondroitinase ABC (CABC) to reduce them to theirdisaccharide components, subjected to reductive amination,
Figure 4. Glycosaminoglycan profiling of various human dermal matrices by FACE. (A) Image of gel demonstrating GAG profile ofhuman dermal matrices grown in the presence of varying amounts of hydrocortisone, compared with available disaccharide GAGstandards. (B) Tabulation of disaccharide content in human dermal matrices. ΔDi-HA denotes an unsaturated hyalurodisaccharide;the following are unsaturated chondrodisaccharides, the numbers indicate the position of the sulphate group on the N-acetylgalactosamine moiety, ΔDi-OS (unsulphated), ΔDi 6S, ΔDi-4S, ΔDi2S, ΔDi-4,6S, ΔDi-2,6S, ΔDi-2,4S: *p<0.05 for comparison withGAG disaccharide levels of normal construct, using one-way ANOVA at α=0.05
Hydrocortisone effect on hyaluronan synthesis in a dermal equivalent
labelled with 2-aminoacridone (AMAC) and run on a high-percentage polyacrylamide gel. The gels were imagedand the GAG contents quantified, using AMAC-labelledglucose as a standard. The fundamental GAG profiles of thehuman dermal matrices did not change in the presence ofvarying amounts of hydrocortisone (Figure 4A). The unsatu-rated hyaluro-disaccharide (ΔDi-HA) content of the humandermal matrices showed the same trend as that observedfor total high molecular weight hyaluronate; an enhance-ment was observed for the 30×, 60× and 200× HC con-structs and a subsequent decline began with the 500× HCconstruct (Figure 4B). This inhibition of ΔDi-HA becomesmore pronounced with the 750× and 1000× HC constructs
(Figure 4B). Interestingly, no significant change wasobserved in ΔDi-4S, ΔDi-6S and ΔDi-4,6S (unsaturatedchondro-disaccharides) content in the 30×, 60×, 200×and 500× human dermal matrices, as compared to thenormal construct. However, with higher levels of hydrocorti-sone in the culturemedium, inhibition ofΔDi-4S,ΔDi-6S andΔDi-4,6S was also observed in the 750× and 1000× HCconstructs (Figure 4B).
3.7. Differential gene array analysis of normaland 500× HC human dermal matrix
Having established that hydrocortisone at higher concen-trations is an effective inhibitor of hyaluronate synthesisin the human dermal matrix, we next designed prelimi-nary experiments aimed at identifying the genes involvedin this inhibition process. In order to identify these genes,differential gene array profile experiments were designed.The 500× HC construct was chosen to be compared to thenormal construct, as it showed the highest level ofhyaluronate inhibition without any significant change inother GAG synthesis. To accomplish this, we used a cDNAmi-croarray containing approximately 1260 human sequences(GlycoV4 oligonucleotide array). Using this methodology tocompare the 500× HC construct to the normal construct,43 genes were observed to be differentially expressed(Figure 5). Of these 43 genes, 25 were upregulated and 18were downregulated (Table 2). Of the downregulated genes,two were identified as being participants in the HA biosyn-thetic pathway, phosphoglucomutase 1 (PGM1) and glucosephosphate isomerase (GPI) (Figure 6). No difference inexpression of HAS (1–3) was observed.
4. Discussion
Neonatal human dermal fibroblasts were cultured in thepresence of varying concentrations of hydrocortisoneunder serum-free conditions and without the aid of anexogenous extracellular matrix (Pouyani et al., 2009).After a culture period of 21 days, the resulting self-assembled human dermal matrices were peeled off thepolycarbonate membrane and characterized. Analysis ofH&E-stained sections of the human dermal matricesshowed fibroblasts dispersed in a dense collagenousmatrix, with varying thickness depending on the concen-tration of hydrocortisone used. We observed an interest-ing correlation between the thickness of the humandermal matrices and hyaluronate content. We observedthat in constructs where hyaluronate synthesis was stimu-lated (30×, 60× and 200×), enhanced thickness of theconstructs was also observed, compared to the normalconstruct (Figure 1). In contrast, in human dermal matri-ces in which HA synthesis was inhibited (500×, 750× and1000×), a decrease in construct thickness was observed.Hyaluronate is a space-filling molecule that is largelyresponsible for tissue hydration; therefore, the variabilityin HA content may in part explain the varying thickness
Figure 5. A heat-map representation of genes differentiallyexpressed in human dermal matrices (normal and 500× HC).Red indicates increased and blue indicates decreased expressionrelative to the mean transcript expression value
observed in the dermal constructs cultured in thepresence of differing amounts of hydrocortisone (Itano,2008). These observations are consistent with previousfindings that report dermal thinning with use of relativelyhigh concentrations of hydrocortisone (Averbeck et al.,2010; Gebhardt et al., 2010). We observed some inhibitionof collagen synthesis, beginning with the 60× constructand higher levels of inhibition at higher hydrocortisoneconcentrations. Previous reports in the literature have alsoreported inhibition of collagen synthesis in human dermalfibroblasts cultured in the presence of hydrocortisone(Russell et al., 1989). Inhibition of cell replication was alsoobserved in the human dermal matrix cultured in the pres-ence of higher levels of hydrocortisone; significant inhibitionwas observed in the 500× construct. Fewer cells than the ini-tial seeding density were observed in the 750× and 1000×constructs when counted at the end of the 21day cultureperiod. This phenomenon is most likely due to loss of cellsfrom cytotoxicity resulting from the high concentrations ofhydrocortisone used.
Ultrastructural analysis by TEMof human dermalmatricesproduced under varying hydrocortisone concentrationsshowed no change in fibril diameter for the 30×, 60×,200× and 500× constructs. Hydrocortisone does not appearto influence collagen fibril assembly up to the 200μg/mlused in the self-assembly of the 500× human dermal matrix.Collagen fibrilogenesis was observed to be normal, demon-strated by the relatively tight distribution of collagen fibril di-ameters in the various human dermal matrices as comparedto the normal construct. The constructs analysed at highermagnification all showed the appropriate collagen bandingpattern, even at the highest concentration of hydrocorti-sone. Similarly, at lower magnification all the humandermal matrices showed the presence of fibroblasts dis-persed in a collagenous matrix; however, a lower densityof collagen fibres was observed in the 750× and 1000×constructs. Taken together, these data suggest that hydro-cortisone had no deleterious effect on collagen fibril assem-bly in the human dermal matrices, specifically at lower andintermediate concentrations.
Table 2. Regulated genes in the 500× human dermal matrix compared to the normal construct
Gene symbol Gene name Fold change (FC) Adjusted p p
Glycosaminoglycan profiling by FACE demonstrated thesame fundamental profile for all self-assembled human der-mal matrices, suggesting that the differentiated fibroblastphenotype does not change with increasing hydrocortisoneconcentration. The hyaluronate disaccharide content, asexpected, shows an increase and subsequent decline at higherlevels of hydrocortisone. Up to a concentration of 500× HC,no appreciable change in other GAG disaccharide contentwas seen in the human dermal matrices, suggesting that bothenhancement and inhibition were indeed specific tohyaluronate. At higher concentrations of hydrocortisone(750× and 1000× constructs) this specificity was lost and in-hibition of other GAGdisaccharide contentwas also observed.
In order to attempt to identify the gene(s) involvedin hyaluronate inhibition, a differential gene arrayanalysis was conducted, in which the 500× HC(200 μg/ml) construct was compared to the normal con-struct (0.4 μg/ml). Of the 48 genes identified in thearray, specifically the 18 downregulated genes, PGM1and GPIwere known to be participants in the hyaluronatebiosynthetic pathway (Figure 6). Downregulation ofthese genes suggests that inhibition of HA synthesis mostlikely takes place early in the glycolytic pathway leadingto HA biosynthesis. Glucose phosphate isomerase isresponsible for converting glucose 6-phosphate to fructose6-phosphate, and phosphoglucomutase catalyses thetransfer of a phosphate group from the 6 position of glu-cose to the 1 position. It can also catalyse the same trans-fer reaction for glucosamine and N-acetylglucosamine,albeit at a much slower rate. It can be inferred from theseresults that hyaluronate inhibition may take place byreducing the cellular content of the precursors to both
uridine disphosphate glucuronic acid (UDP-GlcUA) anduridine diphosphate N-acetylglucosamine (UDP-GlcNAc).This in turn can lead to a decrease in the cellular pool ofUDP-GlcUA and UDP-GlcNAc, the main precursors forhyaluronate synthesis and substrates for HAS, which canthen lead to inhibition of HA synthesis. In spite of thereduced cellular levels of UDP-GlcUA and UDP-GlcNAc, nosignificant inhibition of the other GAGs was observed in the500× HC construct. This phenomenon has been previouslyobserved and may be explained by the fact that the UDP-sugars hyaluronate uses for its synthesis reside in thecytosol, whereas the chondroitin sulphates are synthesizedin the Golgi apparatus (Tammi et al., 2011; Jokela et al.,2008; Kakizaki et al., 2004). Concentrations of the UDP-sugars necessary for HA and other GAG synthesis aremuch higher in the Golgi than in the cytosol. Therefore,depletion of the cellular pool of UDP-sugars affects HAsynthesis rapidly, whereas the other GAGs synthesized in theGolgi remain protected from this substrate deficiency. Noupregulation of any of the hyaluronidases 1–3 (HYAL 1–3)was observed, suggesting that degradation/catabolism of HAwas most likely not responsible for the diminished HAquantities observed. Previously published reports (Zhanget al., 2000; Kultti et al., 2010; Averbeck et al., 2010) havesuggested that HAS2 is involved in inhibition of HA incultured monolayer dermal fibroblasts exposed tohydrocortisone. Although the present results from thedifferential gene array experiments do not show adownregulation of the HAS1–3 genes, we cannotdefinitively conclude, based on available data, that the HASenzymes (specifically HAS2) are not involved in HA inhibi-tion in this system. Future studies will focus on deciphering
Figure 6. Biosynthesis of hyaluronate. Results obtained from the differential gene array analysis by comparing the normal constructto the 500× HC construct demonstrates the downregulation of glucose phosphate isomerase and phosphoglucomutase. Red arrowsindicate the position of these enzymes in the HA biosynthetic pathway
mechanistically, in more detail, the role of the identifiedenzymes (GPI and PGM1) in HA inhibition, and specificallyon unravelling the biphasic nature of the hydrocortisoneeffect on HA synthesis in the human dermal matrix.
In conclusion, we have shown in this article that the self-assembly process can be manipulated by addition of exoge-nous compounds to form novel tissue-engineered dermalequivalentswith differing extracellularmatrix characteristics.These human dermal matrices offer the possibility of study-ing the potentially regenerative effects of hyaluronate onthewound-healing process in vivo, and provide a novelmodelsystem for deciphering the mechanisms of inhibition of vari-ous molecules in vitro (in this case inhibition of hyaluronatebiosynthesis), using differential gene array analysis.
Acknowledgements
We thank Maria Ericsson and Louise Trakimas (Harvard MedicalSchool) for their valuable assistance with transmission electronmicroscopy. We are grateful to Gilberto Hernandez (ScrippsResearch Institute) for his assistance with RNA analysis. This re-search was supported by a grant from the Charles H. Hood Foun-dation (to T.P.).
Conflict of interest
The authors have declared that there is no conflictof interest.
References
Averbeck M, Gebhardt C, Anderegg U, et al.2010; Suppression of hyaluronan synthase 2expression reflects the atrophogenic potentialof glucocorticoids. Exp Dermatol 19: 757–759.
Booth BA, Tan EM, Oikarinen A, et al. 1982;Steroid-induced dermal atrophy: effects ofglucocorticosteroids on collagen metabo-lism in human skin fibroblast cultures. Int JDermatol 21: 333–337.
Benjamini Y, Hochberg Y. 1995; Control-ling the false discovery rate: a practicaland powerful approach to multiple test-ing. J R Statist Soc B57: 289–300.
Bolstad BM, Irizarry RA, Astrand M, et al.2003; A comparison of normalizationmethods for high density oligonucleotidearray data based on bias and variance. Bio-informatics 19: 185–193.
Brown S, Worsfold M, Sharp C. 2001;Microplate assay for the measurementof hydroxyproline in acid-hydrolyzed tis-sue samples. Biotechniques 30: 38–42.
Calabro A, Benavides M, Tammi M, et al.2000a; Microanalysis of enzyme digestsof hyaluronan and chondroitin/dermatansulfate by fluorophore assisted carbohy-drate electrophoresis (FACE). Glycobiology10: 273–281.
Calabro A, Hascall VC, Midura RJ. 2000b;Adaptation of FACE methodology for mi-croanalysis of total hyaluronan in chon-droitin sulfate composition from cartilage.Glycobiology 10: 283–293.
Crowley J. 2010; Scalp psoriasis: an overviewof the disease and available therapies. JDrugs Dermatol 9: 912–918.
Fleischmajer R, Perslish JS, Burgeson RE,et al. 1990; Type I and type III collagen in-teractions during fibrillogenesis. Ann NYAcad Sci 580: 161–175.
Fosang AJ, Hey NJ, Carney SL, et al. 1990; AnELISA plate-based assay for hyaluronan usingbiotinylated proteoglycan G1 domain.Matrix10: 306–313.
Gebhardt C, Averbeck M, Diedenhofen N, et al.2010; Dermal hyaluronan is rapidly reducedby topical treatment with glucocorticoids. JInvest Dermatol 130: 141–149.
Hoffmann K, Auer T, Stücker M, et al. 1998;Comparison of skin atrophy and vasocon-striction due to mometasone furoate,
methylprednisolone and hydrocortisone. JEur Acad Dermatol Venereol 10: 137–142.
Irizarry RA, Bolstad BM, Collin F, et al.2003; Summaries of Affymetrix GeneChipprobe level data. Nucleic Acids Res31: e15.
Itano N. 2008; Simple primary structure, com-plex turnover regulation and multiple rolesof hyaluronan. J Biochem 144: 131–137.
Jiang D, Liang J, Noble PW. 2007;Hyaluronan in tissue injury and repair.Annu Rev Cell Dev Biol 23: 435–461.
Jokela TA, Jauhiainen M, Auriola S, et al.2008; Mannose inhibits hyaluronan syn-thesis by down-regulation of the cellularpool of UDP-N-acetylhexosamines. J BiolChem 283: 7666–7673.
Kakizaki I, Kojima K, Takagaki K, et al.2004; A novel mechanism for theinhibition of hyaluronan biosynthesis by4-methylumbelliferone. J Biol Chem279: 33281–33289.
Kultti A, Kärnä R, Rilla K, et al. 2010; Methyl-β-cyclodextrin suppresses hyaluronan syn-thesis by down-regulation of hyaluronansynthase 2 through inhibition of akt. J BiolChem 285: 22901–22910.
Lubach D, Bensmann A, Bornemann U. 1989;Steroid-induced dermal atrophy. Investiga-tions on discontinuous application.Dermatologica 179: 67–72.
Murphy G, Reich K. 2011; In touch withpsoriasis: topical treatments and currentguidelines. J Eur Acad Dermatol VenereolSuppl 4: 3–8.
Ponec M, De Haas C, Bachra BN, et al. 1979;Effects of glucocorticosteroids on culturedhuman skin fibroblasts. III. Transient inhi-bition of cell proliferation in the earlygrowth stages and reduced susceptibilityin later growth stages. Arch Dermatol Res265: 219–227.
Pouyani T, Ronfard V, Scott PG, et al. 2009; Denovo synthesis of human dermis in vitro inthe absence of a three-dimensional scaffold.In Vitro Cell Dev Biol Anim 45: 430–441.
Pouyani T, Schaffer L, Papp S. 2012; Tissueengineered fetal dermal matrices. In VitroCell Dev Biol Anim 8: 493–506.
Prestwich GD. 2011; Hyaluronic acid-basedclinical biomaterials derived for cell and
molecule delivery in regenerative medi-cine. J Control Release 155: 193–199.
Russell SB, Trupinlf JS, Myers JC, et al. 1989;Differential glucocorticoid regulation ofcollagen mRNAs in human dermal fibro-blasts. Keloid-derived and fetal fibroblastsare refractory to down-regulation. J BiolChem 264: 13730–13735.
Smyth GK. 2004; Linear models and em-pirical Bayes methods for assessing dif-ferential expression in microarrayexperiments. Stat Appl Genet Mol Biol3(1): Article 3.
Stern R, Maibach HI. 2008; Hyaluronan inskin: aspects of aging and its pharmaco-logic modulation. Clin Dermatol 26:106–122.
Stuhlmeier KM, Pollaschek C. 2004;Glucocorticoids inhibit induced and non-induced mRNA accumulation of genesencoding hyaluronan synthases (HAS):hydrocortisone inhibits HAS1 activationby blocking the p38 mitogen-activatedprotein kinase signalling pathway. Rheu-matology 43: 164–169.
Tammi TH, Passi AG, Rilla K, et al. 2011;Transcriptional and post-translational reg-ulation of hyaluronan synthesis. FEBS J278: 1419–1428.
Toole BP. 2004; Hyaluronan: from extracel-lular glue to pericellular cue. Nat RevCancer 4: 528–539.
Toole BP, Slomiany MG. 2008; Hyaluronan: aconstitutive regulator of chemoresistanceand malignancy in cancer cells. Semin Can-cer Biol 18: 244–250.
Wang A, de la Motte C, Lauer M, et al. 2011;Hyaluronan matrices in pathobiologicalprocesses. FEBS J 278: 1412–1418.
ZhangW,Watson CE, Liu C, et al. 2000; Gluco-corticoids induce a near-total suppression ofhyaluronan synthasemRNA in dermal fibro-blasts and in osteoblasts: a molecular mech-anism contributing to organ atrophy.Biochem J 349: 91–97.
Zivanovic D, Medenica L, Tanasilovic S, et al.2011; Dexamethasone-cyclophosphamidepulse therapy in pemphigus: a review of 72cases. Am J Clin Dermatol 11: 123–129.
Hydrocortisone effect on hyaluronan synthesis in a dermal equivalent