Nutrients and photosynthetic pigments were measured over a north–south transect of the East/Japan Sea(EJS), which includes various oceanographic environments such as the cold and warm-water masses, sub-polar fronts, and eddies in the surface ocean. Sampling was conducted in May 2004, July 2005, and October2005. The N:P ratios were low (b4) for high chlorophyll a areas owing to the active Redfieldian consumptionof N and P from waters with a low N:P ratio (approximately 13) in the entire EJS. The compositions ofphotosynthetic pigments were used for identifying phytoplankton groups at a class level using the CHEMTAXprogram. It is notable that cyanobacteria (40–60%) dominated the phytoplankton community in the frontalzone in 2004 and formed approximately 10–50% of the community over the entire surface layer in 2005. Thisdominance of cyanobacteria appears to be associated with the seawater conditions of low N:P ratio andoptimum temperature. As such, the relative amount of prymnesiophytes increased with a decrease in the N:Pratio in the upper 200 m of the EJS. In contrast, more preferentially in high N:P ratio areas, diatoms (40–80%)dominated the phytoplankton community inMay 2004 because of the spring bloom, and pelagophytes (20–60%)dominated the community in July andOctober 2005. Therefore, theEJS seems to provide an ideal environment forstudying the physical and chemical factors that control the community structures of phytoplankton.
The East/Japan Sea (EJS) includes several oceanographic environ-ments with varying water mass properties, such as warmwater in thesouthern half, cold water in the northern half, a well-defined sub-polar front, and many eddies (Fig. 1). The warmwater originates fromthe subtropical Kuroshio current that enters through the Tsushima/Korea Strait (depth: 150 m), and the cold water originates from theLiman current that flows south along the eastern coast. Theconfluence of the warm and cold currents forms the sub-polar frontpositioned at around 40°N. Further, the EJS is one of the most eddy-rich regions in the world (Morimoto et al., 2000). In particular, thewarm eddies play an important role in spreading the southern coastalsurface water to the interior region of the EJS (Isoda, 1994).
In general, the warm water is oligotrophic and has high salinity(N34.5), while the cold water is rich in oxygen and has low salinity(33.9–34.1) (Gong and Son, 1982). The ratio of the total nitrogen tothe total phosphorus concentration in the EJS is approximately 11(Yanagi, 2002), and the ratio of the dissolved inorganic nitrogen (DIN)to the dissolved inorganic phosphorus (DIP) is approximately 13
(Talley et al., 2004). These ratios are considerably lower than theRedfield ratio (16), suggesting that the primary production of thephytoplankton could be limited by nitrogen.
Although nitrogen is almost depleted in the EJS, the EJS has one ofthe most highly productive marine environments (Jenkins, 2008),with a primary production rate of approximately 200gC m−2 yr−1 asmeasured by sediment trap experiments (Hong, 1998). On the basis ofhelium isotope tracers, new production in the southern EJS wasreported to be ∼64gC m−2 yr−1 (Hahm and Kim, 2001), close to thatin the western North Pacific (∼50gC m−2 yr−1; Goes et al., 2000).From satellite image data analyses, it was found that chlorophyll aconcentrations were generally high in spring and fall (Kim et al., 2000;Yamada et al., 2004; Yoo and Kim, 2004); the highest chlorophyll aconcentrations were observed in the sub-polar frontal area (Parket al., 1991). Chlorophyll a concentrations and plankton abundancewere also affected by meso-scale physical features such as upwelling,warm eddies, and the sub-polar front in the Ulleung Basin that islocated in the southwestern part of the EJS (Kang et al., 2004).Seasonal, yearly, and decadal changes in biomass and communitystructure in the EJS have been found to be associated with climatevariability (Lee et al., 2009), and often with Pacific Decadal Oscillation(Liu and Chai, 2009). Recently, Jo et al. (2007) showed that the springbloom in the EJS was considerably enhanced by episodic dust inputs,especially when these dust inputs were accompanied by precipitation.However, basin-scale observational studies on phytoplankton com-position have not yet been conducted in the EJS.
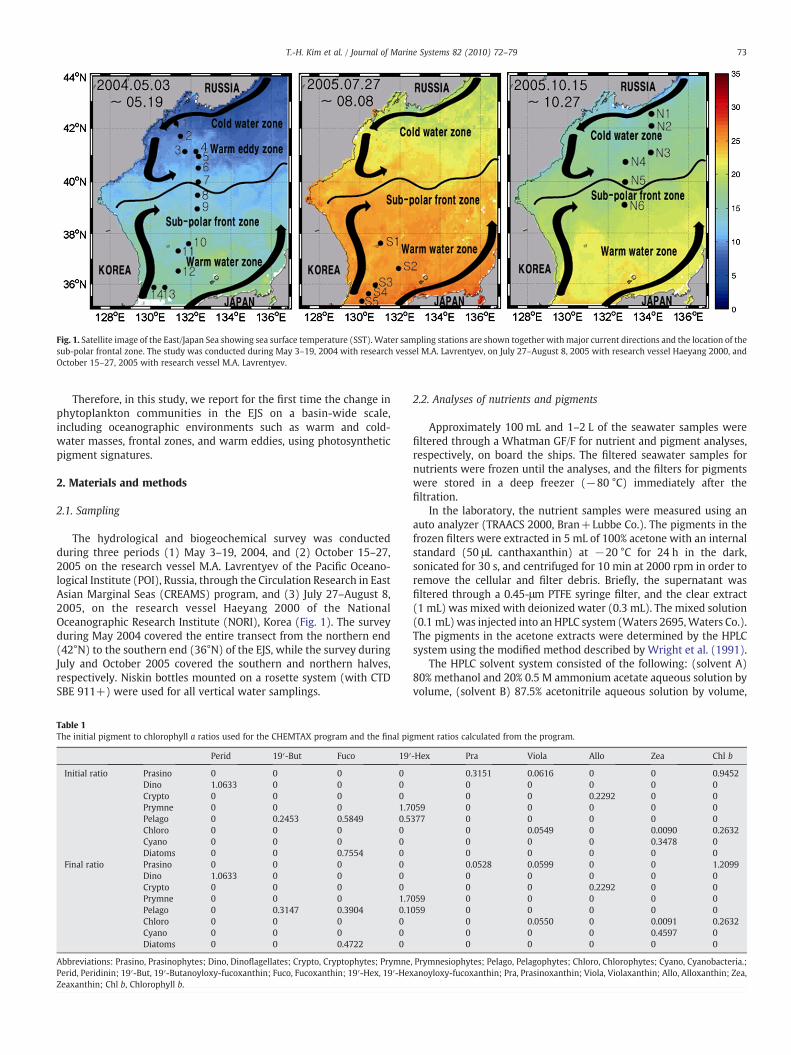
Fig. 1. Satellite image of the East/Japan Sea showing sea surface temperature (SST). Water sampling stations are shown together with major current directions and the location of thesub-polar frontal zone. The study was conducted during May 3–19, 2004 with research vessel M.A. Lavrentyev, on July 27–August 8, 2005 with research vessel Haeyang 2000, andOctober 15–27, 2005 with research vessel M.A. Lavrentyev.
73T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
Therefore, in this study, we report for the first time the change inphytoplankton communities in the EJS on a basin-wide scale,including oceanographic environments such as warm and cold-water masses, frontal zones, and warm eddies, using photosyntheticpigment signatures.
2. Materials and methods
2.1. Sampling
The hydrological and biogeochemical survey was conductedduring three periods (1) May 3–19, 2004, and (2) October 15–27,2005 on the research vessel M.A. Lavrentyev of the Pacific Oceano-logical Institute (POI), Russia, through the Circulation Research in EastAsian Marginal Seas (CREAMS) program, and (3) July 27–August 8,2005, on the research vessel Haeyang 2000 of the NationalOceanographic Research Institute (NORI), Korea (Fig. 1). The surveyduring May 2004 covered the entire transect from the northern end(42°N) to the southern end (36°N) of the EJS, while the survey duringJuly and October 2005 covered the southern and northern halves,respectively. Niskin bottles mounted on a rosette system (with CTDSBE 911+) were used for all vertical water samplings.
Table 1The initial pigment to chlorophyll a ratios used for the CHEMTAX program and the final pig
Approximately 100 mL and 1–2 L of the seawater samples werefiltered through a Whatman GF/F for nutrient and pigment analyses,respectively, on board the ships. The filtered seawater samples fornutrients were frozen until the analyses, and the filters for pigmentswere stored in a deep freezer (−80 °C) immediately after thefiltration.
In the laboratory, the nutrient samples were measured using anauto analyzer (TRAACS 2000, Bran+Lubbe Co.). The pigments in thefrozen filters were extracted in 5 mL of 100% acetone with an internalstandard (50 μL canthaxanthin) at −20 °C for 24 h in the dark,sonicated for 30 s, and centrifuged for 10 min at 2000 rpm in order toremove the cellular and filter debris. Briefly, the supernatant wasfiltered through a 0.45-μm PTFE syringe filter, and the clear extract(1 mL) was mixed with deionized water (0.3 mL). The mixed solution(0.1 mL)was injected into an HPLC system (Waters 2695,Waters Co.).The pigments in the acetone extracts were determined by the HPLCsystem using the modified method described by Wright et al. (1991).
The HPLC solvent system consisted of the following: (solvent A)80% methanol and 20% 0.5 M ammonium acetate aqueous solution byvolume, (solvent B) 87.5% acetonitrile aqueous solution by volume,
74 T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
and (solvent C) ethyl acetate. Each 100 mL of solvent A and solvent Bincluded 0.01 g of butylated hydroxy toluene (BHT) as an antioxidant(Latasa and Bidigare, 1998). The binary linear gradient (min, solvent A%,solvent B%, solvent C%) was (0, 90, 10, 0), (11, 0, 100, 0), (23, 0, 75, 25),(30, 0, 30, 70), (36, 0, 10, 90), and (40, 90, 10, 0). The flow rate was 0.5–1.0 mLmin−1. The authentic standards for chlorophyll a and otherpigments were obtained from Sigma (UK) and DHI (Institute for Waterand Environment, Denmark), respectively. Pigments in the organicsolvents were identified and quantified using documented extinctioncoefficients (Jeffrey, 1997).
2.3. Estimating phytoplankton community structures using a CHEMTAXprogram
HPLC pigment data were processed using CHEMTAX, a factoriza-tion matrix program, in order to estimate the contribution of differentalgal classes to the total chlorophyll a. The CHEMTAX program is aneffective tool for estimating phytoplankton community structures
Fig. 2. Horizontal and vertical variations in temperature, salinity, and nutrients along
(Suzuki et al., 2002). On the basis of the pigment contents, weidentified seven algal categories in our study: diatoms, dinoflagellates,cyanobacteria, prymnesiophytes, pelagophytes, cryptophytes, andprasinophytes. The initial pigment to chlorophyll a ratio for eachmarker used in the CHEMTAX calculation was obtained from theSouthern Ocean data (Mackey et al., 1996; Table 1). The CHEMTAXresults are influenced by the value of the initial pigment ratios used inthe program. Since the cellular pigment composition varies as afunction of light or nutrient conditions (Latasa and Berdalet, 1994;Goericke and Montaya, 1998; Mackey et al., 1998), different initialpigment ratio matrices from the same environment were generallyused (Lohrenz et al., 2003). In order to address this issue, wecompared our CHEMTAX results (using Mackey et al., 1998) with theresults obtained by using two different pigment ratio sets from Suzukiet al. (2002) and Furuya et al. (2003). The comparison showed thatthe abundances of the major algal groups (prasinophytes, diatoms,cyanobacteria, and pelagophytes) in the EJS varied within a range ofapproximately 15%, depending on which initial ratios were chosen.
the transect line of the East/Japan Sea in May 2004 and in July and October 2005.
Fig. 3. Relationship between chlorophyll a concentrations and (A) DIN concentrationsand (B) N:P ratios within the upper 200-m layer in the East/Japan Sea.
75T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
Although the uncertainty was higher for the other phytoplankton(b20%), the horizontal and vertical distribution patterns for algal groupswere almost the same (r2N0.97) for different initial values. Therefore, thehorizontal and vertical comparisons of each phytoplankton communityin our data have, in fact, rather small uncertainties. However, in order toobtainmore accurate CHEMTAX results, class-specific pigment composi-tions from the same environment need to be acquired throughmicroscopic observations (Letelier et al., 1993; Andersen et al., 1996;Mackey et al., 1996; Jeffrey, 1997; Mackey et al., 1998).
3. Results and discussion
3.1. Hydrographic and biogeochemical variations
Temperatures in the upper 100 m layer ranged from 2 to 10 °C inthe northern part of the EJS in May 2004 and October 2005, and from10 to 20 °C in the southern part in May 2004 and July 2005 (Fig. 2).The mixed layer depth ranged from 10 to 50 m in May 2004, with athicker layer in the warm-water zone. In 2005, the mixed layer depthwas approximately 15 m in July and 30 m in October. Temperatureswerewithin a narrow range (1–4 °C) between 100 and 200 m. Salinityranged from 34.0 to 34.6 in the upper 200 m layer through the entiretransect in 2004 and 2005, with slightly lower values (33.6 – 34.0) inthe upper 50 m layer in 2005. The occurrence of a warm eddystructure was apparent at 41°N within the upper 200 m layer in 2004.The eddy exhibited higher temperatures (4–16 °C) with a uniformsalinity (34.10–34.15) in the upper 200 m layer (Fig. 2).
The concentrations of nutrients increased sharply at depths belowthe bottom of themixed layer. The nitrate concentrations ranged from0.1 to 0.8 μM in themixed layer and were less than 17 μM in the upper100-m layer in both years. The nitrite concentrations were consider-ably lower (b0.2 μM) than the nitrate concentrations in general, withrelatively high values at a depth of ∼50 m, right below themixed layer(Fig. 2). The DIP concentrations were less than 0.2 μM in the mixedlayer and less than 1.2 μM in the upper 100-m layer in both years. Thesilicate (DSi) concentrations in the mixed layer ranged from 0.9 to7.7 μM in 2004 and from 1.6 to 3.1 μM in 2005, and theywere less than25 μM in the upper 100 m layer in both years (Fig. 2).
However, in the subsurface eddy zone in 2004, nitrite concentra-tions (2.1 and 2.3 μM for stations 4 and 5 at 50 m) were similar to orhigher than nitrate concentrations (2.5 and 0.77 μM for stations 4 and5 at 50 m), indicating the active regeneration of nutrients from theconverging surface organic matter in this warm eddy. Since unusuallyhigh nitrite concentrations were observed for the two stations at thesame depth, these high values could not be attributed to analyticalartifacts. At this depth, nitrate concentrationswere slightly lower thanthose obtained at other stations, thereby showing general increasingtrends for DIN and DIP with depth for the eddy stations (Fig. 2).
The N:P ratios in themixed layer were less than 11 in 2004 and lessthan 6 in 2005, and those below 100 m were all approximately 13during the study periods; these values were considerably lower thanthe Redfield ratio (16). The very low N:P ratios in the mixed layercould be due to the active Redfieldian consumption of N and P sincethe initial N:P ratio in the entire EJS was approximately 13 (Talleyet al., 2004). The DIN concentrations decreased exponentially asbiological production, inferred from chlorophyll a concentrations,increased (Fig. 3A). However, DIP could not be fully consumed,causing a sharp increase in the N:P ratio depending on the increase inthe biological utilizations (Fig. 3B). This trend indicates that the EJS isa strongly N-limited environment (Fig. 2).
The lower N:P ratios (12) in the upper EJS are in sharp contrast tothose in the Mediterranean Sea (22–29; Krom et al., 1991), anotherlarge marginal sea in the Atlantic. Yanagi (2002) suggested activedenitrification in the EJS leading to lower N:P ratios. However, theselower N:P ratios could be due to the inflow of lower N:P ratio watersthrough the Korea Strait that connects the EJS and the East China Sea
(ECS), rather than due to the strong denitrification in this highly oxicenvironment (DO: 210–380 μM). Onitsuka et al. (2009) found N:Pratios of approximately 12 in the Korea Strait, and Zhang et al. (2007)found unusually low N:P ratios (b10), in association with denitrifi-cation, in themiddle shelf region over the ECS. However, more studiesare necessary to reveal the cause of these lower N:P ratios in the EJS.
3.2. Hydrographic variation in phytoplankton community
The chlorophyll a concentrations, a phytoplankton biomass index,were relatively higher in the upper 100 m layer of the cold-water(20–1000 ng L−1) and the frontal zones (10–1200 ng L−1) in 2004and of the frontal zone (20–470 ng L−1) in 2005 (Fig. 4). Thehighest concentrations (50–1200 ng L−1) of chlorophyll a werefound in the eddy zone in 2004, which could be due to either theconvergence of the organic matter in the cyclonic eddies or theoutcropping of highly productive water masses. Similarly, Moonet al. (1998) reported higher chlorophyll a concentrations in thefrontal zone in summer.
The concentrations of chlorophyll b, a marker pigment of greenalgae including prasinophytes, were highest in the frontal zone(b280 ng L−1 in 2004, and b90 ng L−1 in 2005). They were mostlyhigher in the cold-water zone than in the warm-water zone in bothyears (Fig. 4). The contributions of prasinophytes to the total biomassof phytoplankton were from 10% to 35% in the upper 100 m layer,except for the eddy zone in 2004 (May), and from 5% to 15% in 2005(Fig. 5; July andOctober). In general, the contribution of prasinophytesis about 15% in the open ocean, with higher contributions in spring(Guillou et al., 2004). The contributions of prasinophytes in the EJS
Fig. 4. Horizontal and vertical variations in each marker pigment along the transect line of the East/Japan Sea in May 2004 and in July and October 2005. The variations inprasinoxanthin and violaxanthin are not shown since these concentrations were less than 1 ng/L.
76 T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
were slightly lower than those in theMediterranean Sea (25%) and theAntarctica (33%).
The concentrations of fucoxanthin, a marker pigment of diatoms,were higher in the warm-water zone (20–400 ng L−1) than those inthe cold-water zone (10–250 ng L−1) in the upper 100 m layer in2004. The highest concentrations of fucoxanthin were found in theeddy zone (30–630 ng L−1) in 2004. In 2005, the highest fucoxanthinconcentrations were observed in the upper 100-m layer of the warm-water zone (10–195 ng L−1) (Fig. 4). In general, the diatombiomass inthe EJS is higher in the southern zone than in the northern zone of theEJS (Ohwada and Ogawa, 1966; Kuroda, 1987). Diatoms constitutedabout 40% and 15% of the phytoplankton community in the surfacelayer (0–20 m) in 2004 and 2005, respectively, with the highestproportions (∼80%) in the eddy zone (stations 4 and 5) and thewarm-water zone (stations 10 and 11) in 2004. Similarly, higher proportionsof diatoms were found in a warm eddy occurring in the subtropicalNorth Pacific (Brown et al., 2008). In the deeper layer (20–100 m), theproportion of diatoms increased by about 30% as compared to the
surface layer (Fig. 5). The higher chlorophyll a concentrations anddiatom proportions in May 2004 could be associated with the springbloom in the EJS, occurring in April and May in general (Kim et al.,2000).
The concentrations of zeaxanthin, a marker pigment of cyanobac-teria, were highest (b130 ng L−1) in the surface frontal zone (0–20 m)and less than 5 ng L−1 in most of the other areas in 2004. Thezeaxanthin concentrations were relatively high (5–45 ng L−1) in thesurfacewater over the entire transect in 2005 (Fig. 4). The contributionof cyanobacteria to the total phytoplankton biomass ranged from 40%to 60% in the surface frontal zone (0–20 m) in 2004 and the surfacewater (0–20 m) in 2005. This contribution decreased with an increasein the depth to less than 10% for a layer deeper than 50 m (Fig. 5). It hasbeen documented that vertical distribution patterns for zeaxanthin:chlorophyll a ratios are controlled mainly by vertical mixing of themixed layer (Matsumoto andAndo, 2009). In general, the contributionof cyanobacteria was relatively high in low N:P ratio waters (N:Pratiob4), as shown in Fig. 6. Such a dependency was not observed for
Fig. 5. Horizontal and vertical variations in the contribution of different phytoplankton groups to the total phytoplankton biomass in May 2004 and in July and October 2005. Thevariations in chlorophytes are not shown since their contributions on the CHEMTEX results were less than 1%.
Fig. 6. Average contributions of (A) cyanobacteria and prymnesiophytes, and(B) pelagophytes and diatoms to the total phytoplankton biomass for the different rangesof N:P ratios (0–4, 4–10, and 10–16) within the upper 200-m layer in the East/Japan Sea.
77T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
pelagophytes and diatoms. In the Baltic Sea, a similar dominance ofcyanobacteria has been reported to be caused by an optimaltemperature and decreased N:P ratios (Kononen et al., 1996; Bianchiet al., 2000). It seems that cyanobacteria dominate in the EJS in thestudy seasons, except for the diatom bloom season (i.e., May 2004),because of the optimal temperature (N10 °C), low N:P ratios, andnitrogen depletion favorable for the growth of cyanobacteria (Odateet al., 1990; Le Bouteiller et al., 1992). The dominance of cyanobacteriain the frontal zone, 2004, could be due to the relatively low N:P ratiosobserved after the spring bloom.
The concentrations of 19′-butanoyloxy-fucoxanthin, a markerpigment of pelagophytes, were relatively higher in the surface cold-water zone (10–120 ng L−1) in 2004, and generally ranged from 10 to40 ng L−1 over the entire transect in 2005 (Fig. 4). The distributions ofpelagophytes, the pico-phytoplankton group, composed 10–40% of thetotal biomass of the phytoplankton community in the frontal zone andthe cold-water zone in the upper 100 m layer in 2004, and 20–60% inall the EJS stations in 2005 (Fig. 5). The contributions were relativelyhigher at greater depths. Such high contributions of pelagophytesmaybe associated with the oligotrophic features of the EJS in summer andfall, favorable for picoplanktons (Ansotegui et al., 2003). Thesecontributions are higher than those in the North Atlantic Ocean(5–30%) and the Pacific Ocean (10–30%) (Andersen et al., 1996).
The concentrations of 19′-hexanoyloxy-fucoxanthin, a marker pig-ment of prymnesiophytes,were relatively higher in the upper 20-m layerof the cold-water (30–210 ng L−1) and frontal zones (30–190 ng L−1) in2004. These concentrations generally ranged from30 to 130 ng L−1 overthe entire transect in 2005 (Fig. 4). The contribution of prymnesiophytesto the total phytoplankton biomass was almost constant (5–15%) in theentire upper EJS (Fig. 5). A large contribution of prymnesiophytes was
78 T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
observed in the case of low N:P ratio waters (Fig. 6); this observation issimilar to the observation in the case of cyanobacteria. Further, thisobservation is consistent with the results from laboratory incubationexperiments for Phaeocystis sp., one of themain prymnesiophyte species(Riegman et al., 1992). Although prymnesiophytes are one of the mostabundant nano-planktons (Andersen et al., 1996), Moon-van der Staayet al. (2000) showed a consistently larger contribution from pico-sizeprymnesiophytes (size b3 μm) in the equatorial Pacific Ocean. Thecontributions of prymnesiophytes in the EJSwere lower than those in theNorth Atlantic Ocean (40–50%) and the Pacific Ocean (30–50%)(Andersen et al., 1996).
The concentrations of alloxanthin, a marker pigment of crypto-phytes, were relatively higher in the upper 20 m layer of the cold-waterzone (10–30 ng L−1) and the northern frontal zone (10–60 ng L−1) in2004. Alloxanthin concentrations were less than 20 ng L−1 over theentire transect in 2005 (Fig. 4). The distribution patterns of alloxanthinwere similar to those of 19′-hexanoyloxy-fucoxanthin and 19′-butanoyloxy-fucoxanthin. The contribution of cryptophytes, one of thesmaller nano-planktons, to the total phytoplankton biomass rangedfrom5% to 10% in the entire upper EJS in both years (Fig. 5). This result issimilar to the general contribution (b10%) of cryptophytes in the PacificOcean (Mackey et al., 1998; Suzuki et al., 2002).
The concentrations of peridinin, a marker pigment of dinoflagel-lates, were relatively higher in the warm-water zone (10–250 ng L−1
in 2004, 5–25 ng L−1 in 2005) in both years (Fig. 4). Contributions ofdinoflagellates to the total phytoplankton community were generallyless than 5% for the entire depth (0–100 m) in both years (Fig. 5);these contributions were similar to those (b5%) in the oligotrophicwaters of the North Atlantic Ocean and the Pacific Ocean (Andersenet al., 1996).
4. Conclusions
The EJS has very low N:P ratio conditions (b13) in the euphoticzone; the EJS has several oceanographic environments with consid-erably varying temperatures, such as the cold water, the warm water,and the sub-polar frontal water zones. This unique combination ofenvironments appears to provide favorable conditions for anunusually high growth of cyanobacteria, pelagophytes, and diatoms.A warm eddy observed in the northern EJS showed relatively lownutrient concentrations but a high biomass of phytoplankton. Thephytoplankton community variations induced by the hydrographicalchanges in the EJS are as follows: (1) In the warm-water zone,diatoms and prasinophytes dominated the phytoplankton communityin May 2004, while pelagophytes, diatoms, and cyanobacteriadominated the community in July and October 2005. (2) In thefrontal zone, cyanobacteria and pelagophytes dominated the phyto-plankton community in both years. (3) In the cold-water zone,diatoms, pelagophytes, and prasinophytes dominated the phyto-plankton community in May 2004, while pelagophytes and cyano-bacteria dominated the community in July and October 2005. (4) Inthe eddy zone in 2004, diatoms dominated (N70%) the phytoplanktoncommunity. More extensive observations of photosynthetic pigments,together with other biogeochemical parameters, are necessary toinvestigate the biogeochemical responses to climate changes in thisvery low N:P environment.
Acknowledgements
We would like to thank J.S. Chae for his assistance with fieldsampling and laboratory analysis. This work was supported by theMinistry of Land, Transport and Maritime Affairs as part of the KoreaEAST-1 (East Asian Seas Time-series 1) Program and by the KoreaScience & Engineering Foundation through the National ResearchLaboratory (R0A-2008-000-20055-0). THK and YWL were partially
supported by the BK21 scholarship through the School of Earth andEnvironmental Sciences, Seoul National University, Korea.
References
Andersen, R.A., Bidigare, R.R., Keller, M.D., Latasa, M., 1996. A comparison of HPLCpigment signatures and electron microscopic observations for oligotrophic watersof the North Atlantic and Pacific Oceans. Deep-Sea Res. II 43, 517–537.
Ansotegui, A., Sarobe, A., Trigueros, J.M., Urrutxuru, I., Emma, O., 2003. Size distributionof algal pigments and phytoplankton assemblages in a coastal-estuarine environ-ment: contribution of small eukaryotic algae. J. Plankton Res. 25, 341–355.
Bianchi, T.S., Engelhaupt, E., Westman, P., Andren, T., Rolff, C., Elmgren, R., 2000.Cyanobacterial blooms in the Baltic Sea: natural or human-induced? Limnol.Oceanogr. 45 (3), 716–726.
Brown, S.L., Landry, M.R., Selph, K.E., Yang, E.J., Rii, Y.M., Bidigare, R.R., 2008. Diatoms inthe desert: plankton community response to a mesoscale eddy in the subtropicalNorth Pacific. Deep-Sea Res. II 55, 1321–1333.
Furuya, K., Hayashi, M., Yabushita, Y., Ishikawa, A., 2003. Phytoplankton dynamics in theEast China Sea in spring and summer as revealed by HPLC-derived pigmentsignatures. Deep-Sea Res. II 50, 367–387.
Goericke, R., Montaya, J.P., 1998. Estimating the contribution of microalgal taxa tochlorophyll a in the field-variations of pigment ratios under nutrient- and light-limited growth. Mar. Ecol. Prog. Ser. 169, 97–112.
Goes, J.I., Saino, T., Oaku, H., Ishizaka, J., Wong, C.S., Nojiri, Y., 2000. Basin scale estimatesof sea surface nitrate and new production from remotely sensed sea surfacetemperature and chlorophyll. Geophys. Res. Lett. 27, 1263–1266.
Gong, Y., Son, S.J., 1982. A study of oceanic thermal fronts in the southwestern JapanSea. Bull. Fish. Res. Dev. Agency 28, 25–54 (in Korean).
Guillou, L., Eikrem, W., Chrétiennot-Dinet, M.J., Le Gall, F., Massana, R., Romari, K.,Pedrós-Alió, C., Vaulot, D., 2004. Diversity of picoplanktonic prasinophytes assessedby direct nuclear SSU rDNA sequencing of environmental samples and novelisolates retrieved from oceanic and coastal marine ecosystems. Protist 155,193–214.
Hahm, D., Kim, K.R., 2001. An estimation of the new production in the southern East Seausing helium isotopes. J. Korean Soc. Oceanogr. 36, 19–26.
Hong, G.H., 1998. Time-series observation and sediment trap experiment. Developmentof Monitoring Technology for the wastes disposal sea areas. 3 rd KORDI Report,Ansan, pp. 500–509.
Isoda, Y., 1994. Warm eddy movements in the eastern Japan Sea. J. Oceanogr. 50, 1–15.Jeffrey, S.W., 1997. Application of pigment methods to oceanography. In: Jeffrey, S.W.,
Mantoura, R.F.C., Wright, S.W. (Eds.), Phytoplankton Pigments in Oceanography:Guidelines to Modern Methods. UNESCO Publishing, Paris.
Jenkins, W.J., 2008. The biogeochemical consequences of changing ventilation in theJapan/East Sea. Mar. Chem. 108, 137–147.
Jo, C.O., Lee, J.Y., Park, K.A., Kim, Y.H., Kim, K.R., 2007. Asian dust initiated early springbloom in the northern East/Japan Sea. Geophys. Res. Lett. 34, L05602. doi:10.1029/2006GL027395.
Kang, J.-H., Kim, W.-S., Chang, K.-I., Noh, J.-H., 2004. Distribution of plankton related tothe mesoscale physical structure within the surface mixed layer in thesouthwestern East Sea. Korea. J. Plankton Res. 26, 1515–1528.
Kim, S.W., Saitoh, S.I., Ishizaka, J., Isoda, Y., Kishino, M., 2000. Temperal and spatialvariability of phytoplankton pigment concentrations in the Japan Sea derived fromCZCS images. J. Oceanogr. 56, 527–538.
Kononen, K., Kuparinen, J., Makela, K., Laanements, J., Pavelson, J., Nommann, S., 1996.Initiation of cyanobacterial blooms in a frontal region at the entrance of the Gulf ofFinland, Baltic Sea. Limnol. Oceanogr. 41, 98–112.
Krom, M.D., Kress, N., Brenner, S., Gordon, L.I., 1991. Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea. Limnol. Oceanogr. 36, 424–432.
Kuroda, K., 1987. Chlorophyll distribution in the Kuroshio region, south of Japan.Kousuiken Note. Sora to Umi 9, 19–29.
Latasa, M., Berdalet, E., 1994. Effect of nitrogen or phosphorus starvation on pigmentcomposition of cultured Heterocapsa sp. J. Plankton Res. 16, 83–94.
Latasa, M., Bidigare, R.R., 1998. A comparison of phytoplankton populations of theArabian Sea during the spring intermonsoon and Southwest Monsoon of 1995 asdescribed by HPLC-analyzed pigments. Deep-Sea Res. II 45, 2133–2170.
Le Bouteiller, A., Blanchot, J., Rodier, M., 1992. Size distribution patterns ofphytoplankton in the western Pacific: towards a generalization for the tropicalopen ocean. Deep-Sea Res. 39, 805–823.
Lee, J.-Y., Kang, D.-J., Kim, I.-N., Rho, T., Lee, T., Kang, C.-K., Kim, K.-R., 2009. Spatial andtemporal variability in the Pelagic ecosystem of the East Sea (Sea of Japan): areview. J. Mar. Syst. 78, 288–300.
Letelier, R.M., Bidigare, R.R., Hebel, D.V., Ondrusek, M., Winn, C.D., Karl, D.M., 1993.Temporal variability of phytoplankton community structure based on pigmentanalysis. Limnol. Oceanogr. 38, 1420–1437.
Liu, G., Chai, F., 2009. Seasonal and interannual variation of physical and biologicalprocesses during 1994–2001 in the Sea of Japan/East Sea: a three-dimensionalphysical–biogeochemical modeling study. J. Mar. Syst. 78, 265–277.
Lohrenz, S.E., Carroll, C.L., Weidemann, A.D., Tuel, M., 2003. Variations in phytoplanktonpigments, size structure and community composition related to wind forcing andwater mass properties on the North Carolina inner shelf. Cont. Shelf Res. 23,1447–1464.
Mackey, M.D., Mackey, D.J., Higgins, H.W., Wright, S.W., 1996. CHEMTAX — a programfor estimating class abundances from chemical markers: application to HPLCmeasurements of phytoplankton. Mar. Ecol. Prog. Ser. 144, 265–283.
79T.-H. Kim et al. / Journal of Marine Systems 82 (2010) 72–79
Mackey, D.J., Higgins, H.W., Mackey, M.D., Holdsworth, D., 1998. Algal class abundancesin the western equatorial Pacific: estimation from HPLC measurements ofchloroplast pigments using CHEMTAX. Deep-Sea Res. 45, 1441–1468.
Matsumoto, K., Ando, K., 2009. Use of cyanobacterial pigments to characterize the oceansurface mixed layer in the western Pacific warm pool. J. Mar. Syst. 75, 245–252.
Moon, C.H., Yang, S.R., Yang, H.S., Cho, H.J., Lee, S.Y., Kim, S.Y., 1998. Regenerationprocesses of nutrients in the polar front area of the East Sea: chlorophyll adistribution, new production and the vertical diffusion of nitrate. Bull. Korean Fish.Soc. 31, 259–266.
Moon-van der Staay, S.Y., Van der Staay, G.W.M., Guillou, L., Vaulot, D., Claustre, H.,Medlin, L.K., 2000. Abundance and diversity of prymnesiophytes in the picoplank-ton community from the equatorial Pacific Ocean inferred from 18 S rDNAsequences. Limnol. Oceanogr. 45, 98–109.
Morimoto, A., Yanagi, T., Kaneko, A., 2000. Eddy field in the Japan Sea derived fromsatellite altimetric data. J. Oceanogr. 56, 449–462.
Odate, T., Yanada, M., Castillo, L.V., Maita, Y., 1990. Distribution of cyanobacteria andother picophytoplankton in the western north Pacific ocean, summer 1989. J.Oceanol. Soc. Japan 46, 184–189.
Ohwada, M., Ogawa, F., 1966. Plankton in the Japan Sea. Oceno. Magaz. 18, 39–42.Onitsuka, G., Morimoto, A., Takikawa, T., Watanabe, A., Moku, M., Yoshikawa, Y., Yanagi,
T., 2009. Enhanced chlorophyll associated with island-induced cyclonic eddies inthe eatern channel of the Tsushima Straits. Estuar. Coast. Shelf Sci. 81, 401–408.
Park, J.S., Kang, C.K., An, K.H., 1991. Community structure and spatial distribution ofphytoplankton in the polar front region off the east coast of Korea in summer. Bull.Korean Fish. Soc. 24, 237–247.
Riegman, R., Noordeloos, A.A.M., Cadee, G.C., 1992. Phaeocystis blooms and eutrophi-cation of the continental coastal zones of the North Sea. Mar. Biol. 112, 479–484.
Suzuki, K., Minami, C., Liu, H., Saino, T., 2002. Temperal and spatial patterns ofchemotaxonomic algal pigments in the subarctic Pacific and the Bering Sea duringthe early summer of 1999. Deep-Sea Res. II 49, 5685–5704.
Talley, L.D., Tishchenko, P., Luchin, V., Nedashkovskiy, A., Sagalaev, S., Kang, D.J.,Warner, M., Min, D.H., 2004. Atlas of Japan (East) Sea hydrographic properties insummer, 1999. Prog. Oceanogr. 61, 277–348.
Wright, S.W., Jeffrey, S.W., Mantoura, R.F.C., Llewellyn, C.A., Bjørnland, T., Repeta, D.,Welschmeyer, N., 1991. Improved HPLCmethod for the analysis of chlorophylls andcarotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 77, 183–196.
Yamada, K., Ishizaka, J., Yoo, S., Kim, H.C., Chiba, S., 2004. Seasonal and interannualvariablilty of sea surface chlorophyll a concentration in the Japan/East Sea (JES).Prog. Oceanogr. 61, 193–211.
Yanagi, T., 2002. Water, salt, phosphorus and nitrogen budgets of the Japan Sea. J.Oceanol. Soc. Japan 58, 797–804.
Yoo, S., Kim, H.C., 2004. Suppression and enhancement of the spring bloom in thesouthern western East Sea/Japan Sea. Deep-Sea Res. II 51, 1093–1111.
Zhang, J., Liu, S.M., Ren, J.L., Wu, Y., Zhang, G.L., 2007. Nutrient gradients from theeutrophic Changjiang (Yangtze River) Estuary to the oligotrophic Kuroshio watersand reevaluation of budgets for the East China Sea Shelf. Prog. Oceanogr. 74,449–478.