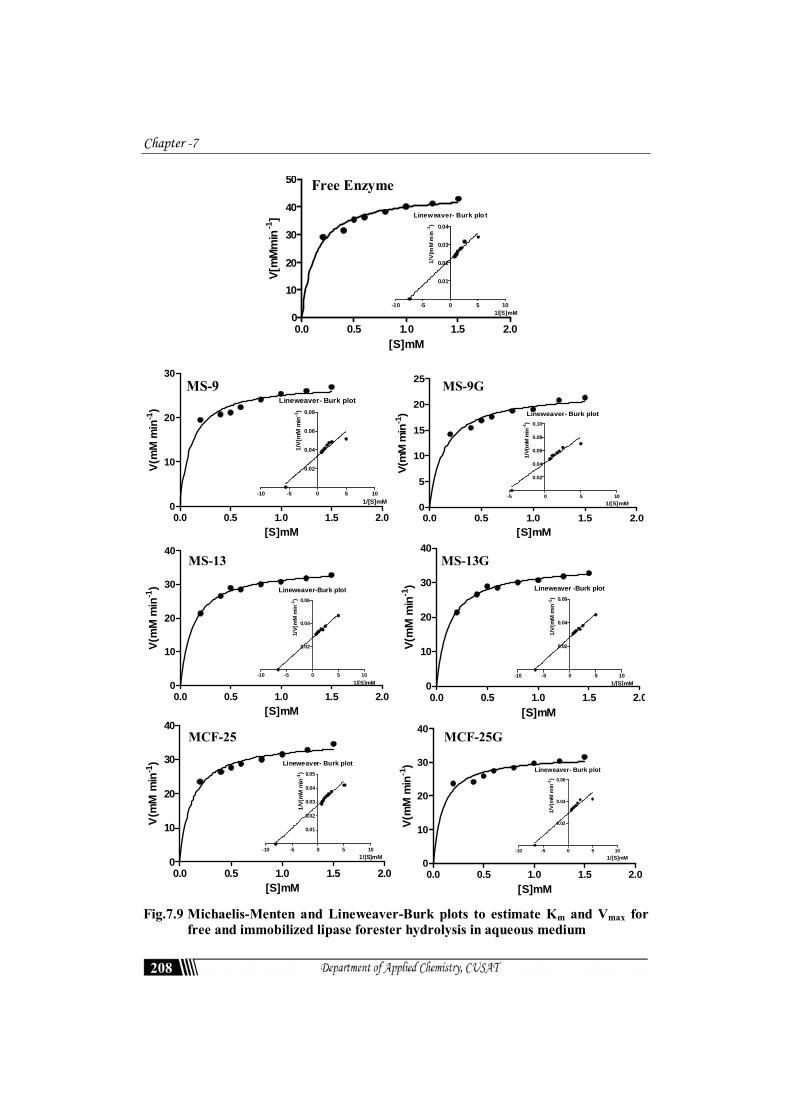
Hydrolysis of Ester by Lipase Immobilized on Mesosilica 185 HYDROLYSIS OF ESTER BY LIPASE IMMOBILIZED ON MESOSILICA Lipases from Candida Rugosa have many excellent characteristics and they are widely used for preparative purposes for organic synthesis in many industrial applications and scientific research projects. One aim of the research dealing with enzymes in aqueous media is to determine the optimal conditions for a specific application. Very few studies have been devoted to direct comparison of lipase activity in aqueous and organic media. The features of support enable the immobilization of lipase via strong interactions enhancing the stability of immobilized enzyme molecules. Biocompatible mesoporous silica nanoparticle supports have been used for biocatalysis in various applications owing to their long-term durability and efficiency. In this study, the hydrolysis rate of p-nitrophenyl palmitate in a batch reactor with Candida rugosa lipase was investigated. The conditions such as p-PNPP concentration, temperature and pH were evaluated to achieve the optimum reaction conditions for both free and immobilized lipase. Esterasic activity is used to test a rapid assessment of lipase activity. 7.1 Introduction Lipases (EC 3.1.1.3) (triacylglycerol acylhydrolases, glycerol ester hydrolases) are serine hydrolases which under physiological conditions catalyze the hydrolysis of ester bonds in the molecules of triacylglycerols to glycerol and free fatty acids. They also catalyze a variety of synthesis reactions under reduced aqueous conditions (e.g., esterification, transesterification,
Transcript
Hydrolysis of Ester by Lipase Immobilized on Mesosilica
Lipases from Candida Rugosa have many excellent characteristics and they are widely used for preparative purposes for organic synthesis in many industrial applications and scientific research projects. One aim of the research dealing with enzymes in aqueous media is to determine the optimal conditions for a specific application. Very few studies have been devoted to direct comparison of lipase activity in aqueous and organic media. The features of support enable the immobilization of lipase via strong interactions enhancing the stability of immobilized enzyme molecules. Biocompatible mesoporous silica nanoparticle supports have been used for biocatalysis in various applications owing to their long-term durability and efficiency. In this study, the hydrolysis rate of p-nitrophenyl palmitate in a batch reactor with Candida rugosa lipase was investigated. The conditions such as p-PNPP concentration, temperature and pH were evaluated to achieve the optimum reaction conditions for both free and immobilized lipase. Esterasic activity is used to test a rapid assessment of lipase activity.
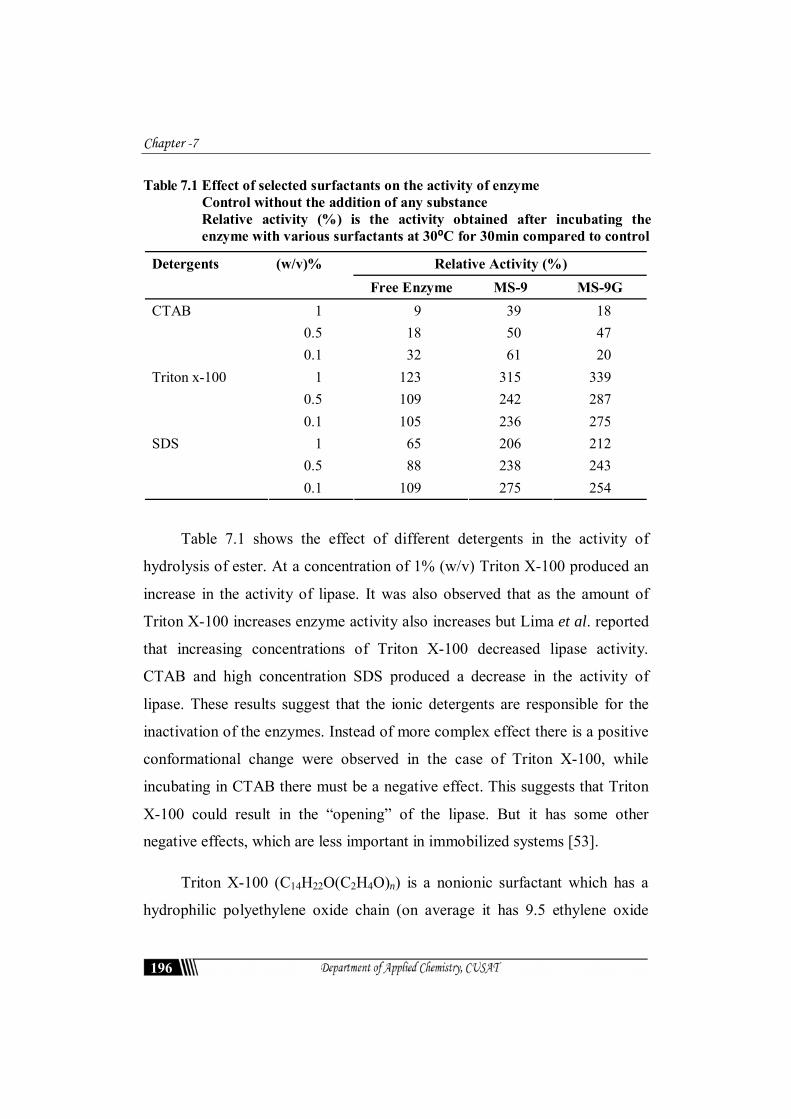
Table 7.1 shows the effect of different detergents in the activity of
hydrolysis of ester. At a concentration of 1% (w/v) Triton X-100 produced an
increase in the activity of lipase. It was also observed that as the amount of
Triton X-100 increases enzyme activity also increases but Lima et al. reported
that increasing concentrations of Triton X-100 decreased lipase activity.
CTAB and high concentration SDS produced a decrease in the activity of
lipase. These results suggest that the ionic detergents are responsible for the
inactivation of the enzymes. Instead of more complex effect there is a positive
conformational change were observed in the case of Triton X-100, while
incubating in CTAB there must be a negative effect. This suggests that Triton
X-100 could result in the “opening” of the lipase. But it has some other
negative effects, which are less important in immobilized systems [53].
Triton X-100 (C14H22O(C2H4O)n) is a nonionic surfactant which has a
hydrophilic polyethylene oxide chain (on average it has 9.5 ethylene oxide
Hydrolysis of Ester by Lipase Immobilized on Mesosilica
197
units) and an aromatic hydrocarbon lipophilic or hydrophobic group. Free
lipase increased the activity 5% at 0.1% Triton X-100 concentration, while
with 0.5% the activity increased to 9% and with 1% the behaviour was quite
different. Addition of 1% Triton X-100 enhanced enzyme activity more than
200% for immobilized systems.
CTAB caused a decrease in the activity even at the lowest concentration
(0.01%). At 1% CTAB concentration the relative activity was around 8% for free
enzyme 0.1% CTAB produced a slight increment in enzyme activity, although at
higher concentrations it produced a decrease in enzyme activity. Similar reports
were observed for lipase immobilized on cyanogen bromide [44].
SDS at 0.1(w/v %) slightly stimulated the free enzyme activity (9%) but
significant activities were observed with immobilized systems. Incubation of
the enzyme in the presence of these detergents (CTAB) suggested that they
could inactivate immobilized lipase. Immobilized system was more resistant to
this inactivation due to the higher stability of this immobilization method.
Most nonionic detergents, such as Triton X-100 and Tween 20 slightly
increased the lipase activity while SDS completely inhibited [45].
7.3.6 Effect of various chemicals on the activity and stability
Cofactors are not required for lipase activity but divalent cations such as
calcium generally stimulate the activity. It has been postulated that this is based
on the formulation of calcium salts of long-chain fatty acids [46]. The lipase
activity is inhibited drastically by Co2+, Ni2+, Hg2+ and Sn2+; and is slightly
inhibited by Zn2+, Mg2+ and EDTA [47]. In H. lanuginosa S-38, sulphahydryl-
reducing agents like dithiothreitol did not alter the enzyme activity but did render
it more susceptible to heat inactivation. Inactivation is accelerated by the addition
of urea. Reducing compounds (cysteine, 2-mercaptoethanol), chelating agents
Chapter -7
198
(EDTA, o-phenanthroline) and thiol group inhibitors (p-chloro mercuric benzoate,
monoiodoacetate) did not show a detectable effect on lipase Lipase is not a
metallo-enzyme and it does not require either free -SH group or an intact S–S
bridge for its activity.
To evaluate activity and the stability of free and immobilized enzymes, they
were incubated with 5 mL of various chemical solutions at 30⁰C for 30 min at
1 mM concentrations. The relative lipase activity after incubation was determined
under standard assay conditions of p-NPP as substrate and the results expressed as
percentage of the activity obtained without the chemical agent.
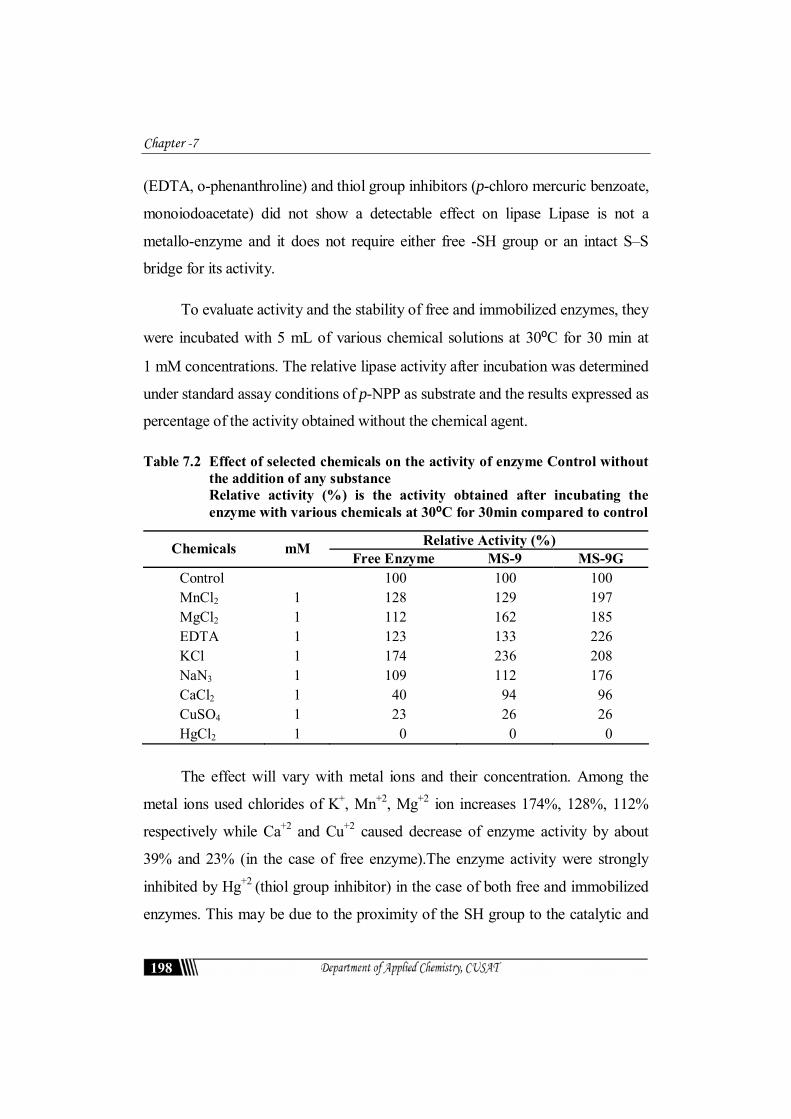
Table 7.2 Effect of selected chemicals on the activity of enzyme Control without the addition of any substance Relative activity (%) is the activity obtained after incubating the enzyme with various chemicals at 30⁰C for 30min compared to control
Relative Activity (%) Chemicals mM Free Enzyme MS-9 MS-9G
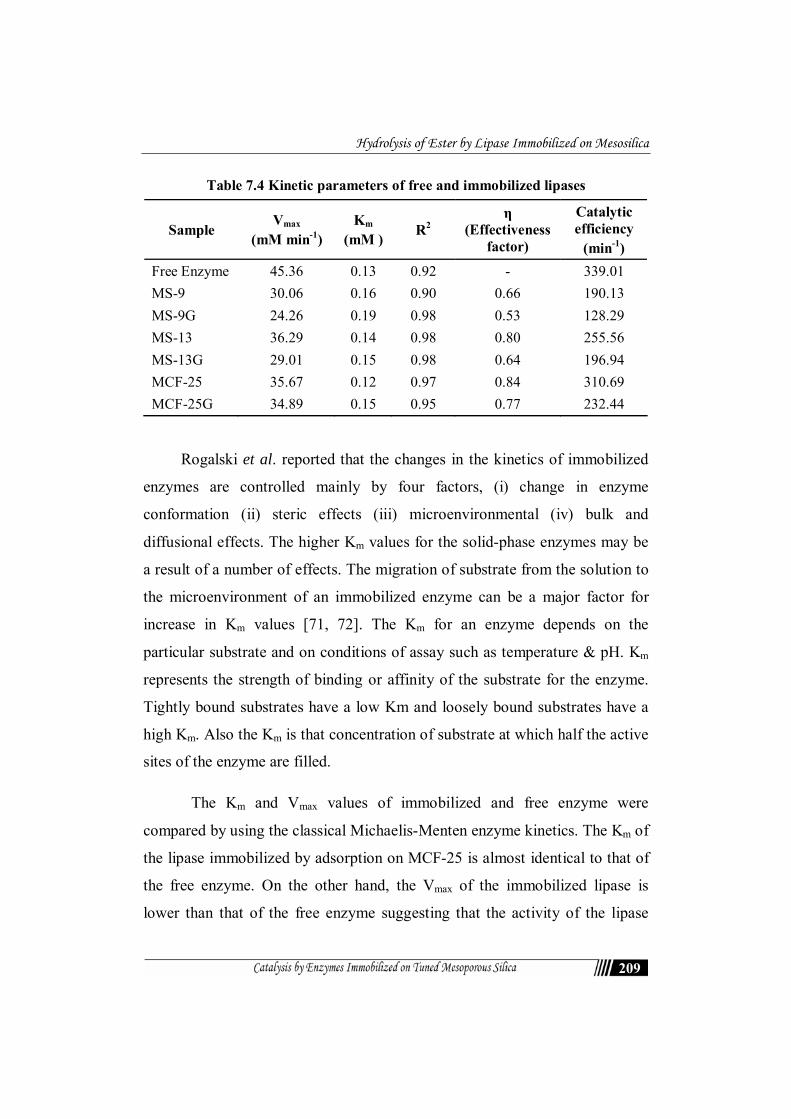
The effect will vary with metal ions and their concentration. Among the
metal ions used chlorides of K+, Mn+2, Mg+2 ion increases 174%, 128%, 112%
respectively while Ca+2 and Cu+2 caused decrease of enzyme activity by about
39% and 23% (in the case of free enzyme).The enzyme activity were strongly
inhibited by Hg+2 (thiol group inhibitor) in the case of both free and immobilized
enzymes. This may be due to the proximity of the SH group to the catalytic and
Hydrolysis of Ester by Lipase Immobilized on Mesosilica
199
interfacial binding site but spacially remote from the catalytic site. This may have
induced the marked loss of activity [30, 48]. The catalytic triad of lipases has been
recognized to consist of Ser, His, and Glu or Asp [49]. Thus the bulky Hg2+ group
might cause steric interference to the approach of the substrate to the active site.
NaN3 at levels of incorporation of 1mM activated free CR Lipase and covalently
bound lipase on to MS-9G by 8% and 76% respectively. EDTA (1 mM) activated
the free enzyme, adsorbed lipase and covalently bound lipase by 22%, 33% and
126% respectively.
Treatment with Cu2+ at both 1 and 10mM significantly inhibited the
activity of grey mullet lipase with only 19% residual activity after 30 min
incubation at 10 mM. This was similar to the findings of Choo et al [50] and
Aryee et al [51], but different from what Sztajer et al [52] and Lima et al [53]
reported for Penicillium simplicissimum lipase (4.7%) and Penicillium
aurantiogriseum lipase (31%) by 1mM Cu2+.
7.3.7 Effect of incubating medium on the activity and stability of lipase
Enzymes have been described to function in non aqueous media although,
in at least the majority of cases a finite level of water associated with the protein
must be present to retain conformational integrity and there by activity. Many
researchers investigated a wide variety of common solvents to see which would
support the enzyme for catalysis. In fact it seems very much like an on-off type
situation. The enzyme was either fully or near- fully active or it was not active at
all. The lipases were active in all water immiscible solvents. Conversely, the
solvents in which the enzyme was not active were all water-miscible.
Water certainly plays a very key role in maintaining conformational
integrity of proteins. Hydrophobicity is the main criteria to enhance the
activity but that is more favourable in presence of small amount of water.
Chapter -7
200
Consequently, it is proved that the water miscible solvents with decreasing
hydrophobicity and increasing polarity extract essential water from the protein.
A structural change in the protein accompanies water loss, resulting in a non-
active conformation. The stability in organic solvents is an important
characteristic of lipases. It is very interesting to determine whether the enzyme
can be used to catalyze synthetic reactions and also to predict which solvent
would be better for performing the reaction.
Lipases are diverse in their sensitivity to solvents, but there is a general
agreement that polar water-miscible solvents (−2.5 < log P < 0) are more
destabilizing than water immiscible solvents (2 < log P < 4) [54, 55].
Hydrophilic solvents exhibit a higher affinity to water and hence it is more
likely that they will strip essential water from enzyme molecules, than
hydrophobic solvents. The log P value describes the hydrophobicity of
organic solvents, where as P is the partition coefficient of the solvent
between octanol and water in a two-phase system [56, 57]. Both free and
immobilized enzymes were incubated in different solvents at 300C for 1hr
and the residual activities were measured.
Table 7.3 Effect of selected solvents on the activity of enzyme Control without the addition of any substance Relative activity (%) is the activity obtained after incubating the enzyme with various chemicals at 30⁰C for 30min compared to control
Residual Activity (%) Solvent log P Free MS-9 MS-9G
[36] J. F. Shaw, R. C. Chang, F. F. Wang, Y. Wang; J. Biotechnol. Bioeng., 35 (1990) 132
[37] S. Fadiloglu, Z. Soylemez; J. Agric. Food Chem., 46 (1998) 3411.
[38] B. K. Yang, J. P. Chen; J. Food Sci., 59 (1994) 424.
Chapter -7
214
[39] S. Fadıloglu, Z. Soylemez, J. Agric. Food Chem., 46 (1998) 3411.
[40] J. E. Mogensen, P. Sehgal, D. E. Otzen.; Biochemistry 44 (2005) 1719.
[41] G. F. Lorente, J. M. Palomo, Z. Cabrera, R. F.Lafuente, J. M. Guisán; Biotechnol. Bioeng., 97 (2007) 242.
[42] H. J. Pignol, D. Kerfelec, B. Crenon, I. Chapus, Fontecilla-Camps; J. Biol. Chem., 271 (1996) 1807.
[43] M. P Egloff,., F. Buono, G. Verger, R. Cambillau, V. Tilbeurgh; Biochemistry., 34 (1995) 2751.
[44] G. Volpatoa, M. Filice, R. C. Rodriguesa, J. X. Heckc, J. M. Guisana, C. Mateoa, M. A. Z. Ayub; J. Mol. Catal. B: Enzym., 60 (2009) 125.
[45] K. A. Novototskaya-Vlasova, L. E. Petrovskaya, E. M. Rivkina1, D. A. Dolgikhand, M. P. Kirpichnikov, Biochemistry.,78 (2013) 385.
[46] S. E. Godfredson; Microbial Enzymes and Biotechnology (eds. Fogarty, W. M. and Kelly, E. T.), Elsevier Applied Sciences, The Netherlands, (1990) 255.
[47] S. Patkar, F. Bjorkling; Lipases: their Structure, Biochemistry and Application (eds. P. Woolley, and S. B. Petersen), Cambrdige University Press, U.K., (1994) 207.
[48] B. A. Raso, H. O. Hultin, Comp. Biochem. Physiol., 89B (1988) 671.
[49] W. Jin, U.C. Broedl, H. Monajemi, J. M. Glick, D. J. Rader, Genomics, 80 (2002) 268.
[50] D. W. Choo, T. Kurihara, T. Suzuki, K. Soda, N. Esaki; Appl. Environ. Microbiol., 64 (1998) 486.
[51] A. N. A. Aryee, B. K. Simpson, R. Villalonga, Enzyme Microb. Technol., 40 (2007) 394.
[52] H. Sztajer, H. Lunsdorf, H. Erdmann, U. Menge, R. Schmid, Biochim. Biophys. Acta., 61(1992)1124.
[53] V. M. G. Lima, N. Krieger, D. A. Mitchell, J. D. Fontana; J.Biochem. Eng., 18 (2004) 65.
Hydrolysis of Ester by Lipase Immobilized on Mesosilica
215
[54] B. Fu, P. T. Vasudevan, Energy Fuels, 23 (2009) 4105.
[55] L. M. Pera, C. M. Romero, M. D. Baigori, G. R. Castro, Food Technol. Biotechnol., 44 (2006) 247.
[56] C. Laane, S. Boeren, K. Vos, C. Beeger. Biotechnol. Bioeng., 30 (1987) 81.
[57] M.Guncheva, M. Dimitrov, D. Zhiryakova; Biochemistry, 4 (2011) 2170.
[58] P. Grochulski, L. Yunge, J. D. Schrag, F. Bouthillier, P. Smith, D. Harrison, B. Rubin, M. Cygler, J. Mol .Chem., 268 (1993) 12843.
[59] I. J. Colton, S. N. Ahmed, R. J. Kazlauskas; J. Org.,Chem., 60 (1995) 212.
[60] M. Pogorevc, H. Stecher, K. Faber; Biotechnol. Lett., 24 (2002) 857.
[61] J. C. Wu, S. S. Lee, M. M. B. Mahmood, Y. Chow, M. M. R. Talukder, W. J. Choi; J.Mol. Catal. B: Enzym., 45 (2007) 108.
[62] A. Dimitrijevic, D.Velickovic, D. Bezbradica, F. Bihelovic, R. Jankov, N. Milosavic, J. Serb. Chem. Soc., 76 (2011) 1081.
[63] S. Hazarika, P. Goswami, N. N. Dutta, A. K. Hazarika; J.Chem. Eng., 85 (2002) 61.