Page 1
ORIGINAL PAPER
Hydrolysis of fungal and plant cell walls by enzymatic complexesfrom cultures of Fusarium isolates with different aggressivenessto rye (Secale cereale)
Jolanta Jaroszuk-Sciseł • Ewa Kurek
Received: 22 December 2011 / Revised: 25 January 2012 / Accepted: 6 February 2012 / Published online: 3 March 2012
� Springer-Verlag 2012
Abstract The efficiency of hydrolysis of fungal (Fusar-
ium spp.) cell wall and rye root cell wall by crude enzy-
matic complexes from (42-day-old) cultures of three
F. culmorum isolates, a plant growth–promoting rhizosphere
isolate (PGPF) DEMFc2, a deleterious rhizosphere isolate
(DRMO) DEMFc5, and a pathogenic isolate DEMFc37, as
well as two other, pathogenic isolates belonging to
F. oxysporum and F. graminearum species was studied. In the
enzymatic complexes originating from the Fusarium spp.
cultures, the activities of the following cell wall–degrading
enzymes were identified: glucanases, chitinases, xylanases,
endocellulases, exocellulases, pectinases, and polygalac-
turonases. The preparation originating from a culture of the
PGPF isolate was the least efficient in plant cell wall
(PCW) hydrolysis. There were no significant differences in
the efficiency of PCW hydrolysis between preparations
from cultures of the DRMO and the pathogenic isolates.
PGPF was the most efficient in liberating reducing sugars
and N-acetylglucosamine (GlcNAc) from fungal cell walls
(FCW). Xylanase activities of the enzymatic complexes
were strongly positively (R [ ?0.9) correlated with their
efficiency in hydrolyzing PCW, whereas chitinase activi-
ties were correlated with the efficiency in FCW hydrolysis.
Keywords Efficiency of cell wall hydrolysis � PGPF �DRMO � Pathogenic fungi � Xylanase activity �Chitinase activity
Introduction
The activities of lytic enzymes hydrolyzing cell wall
polysaccharides (CWDEs) of fungi as well as plants
increase substantially in aging culture filtrate of filamen-
tous fungi (Alfonso et al. 1995b; Douaiher et al. 2007;
Jaroszuk-Sciseł et al. 2011).
The lytic enzymes released from microorganisms could
be involved in the control of fungal growth under specific
conditions via various mechanisms (Boothby and Magreola
1984). Fungal cell wall (FCW) degrading enzymes are
probably involved in the penetration and infection of plants
(Annis and Goodwin 1997; Keon et al. 1987). CWDE
secreted by a pathogen may diffuse in the apoplast and
react with host cell wall constituents, thereby facilitating
pathogen penetration through the loosened host cell walls
and middle lamella matrices (Annis and Goodwin 1997;
Carpita and Gibeaut 1993). CWDE synthesized during
plant infection by F. culmorum (Kang and Buchenauer
2000) include cellulases, xylanases, and pectinases.
The plant cell wall (PCW) is thought to be a dynamic
and metabolically active structure that functions as a
potential barrier to the penetration and spread of pathogens.
It is at the same time, a substrate for extracellular enzymes
secreted by pathogens (Akimitsu et al. 2004; Annis and
Goodwin 1997) and a reservoir of signal molecules regu-
lating or activating plant defense responses to microbial
attack (Degefu et al. 1995; Hatsch et al. 2006). Fungal
glycosidases such as b-galactosidases, xylosidases, and
arabinosidases release sugar moieties that can be used by a
Communicated by Erko Stackebrandt.
J. Jaroszuk-Sciseł (&) � E. Kurek
Department of Environmental Microbiology,
Institute of Microbiology and Biotechnology,
Maria Curie-Skłodowska University,
Akademicka St. 19, 20-033 Lublin, Poland
e-mail: [email protected]
E. Kurek
e-mail: [email protected]
123
Arch Microbiol (2012) 194:653–665
DOI 10.1007/s00203-012-0803-4
Page 2
fungus as a nutritional source during its growth through
plant tissues (Akimitsu et al. 2004). It has been demon-
strated that cereal pathogens produce more xylanase
activity than other cell wall–degrading enzymes that use
cereal cell walls as a C source (Apel et al. 1993; Degefu
et al. 2001; Giesbert et al. 1998; Lalaoui et al. 2000; Phalip
et al. 2009; Wanjiru et al. 2002; Wu et al. 1995, 2006). In
graminaceous crops and grasses, arabino-xylan accounts
for up to 40% of the walls (Belien et al. 2006; Carpita
1996; Hatsch et al. 2006; Labavitch and Ray 1978).
Earlier studies (Jaroszuk-Sciseł et al. 2008) on three
Fusarium culmorum isolates originating from the rhizo-
sphere of cereal plants indicated that their interactions with
rye plants were quite different from one another. Inocula-
tion of seedlings with isolate DEMFc2 resulted in a 20%
increase in shoot fresh weight, whereas DEMFc5 and
DEMFc37 caused more than a 20% and a 38% reduction in
this parameter and fusariosis symptoms in 14-day-old
plants, respectively. The two rhizosphere isolates (DEM-
Fc2 and DEMFc5) colonized the epidermis and the cortex
but were not found in vessels, while the pathogen colonized
all three layers of root cells. The number of DEMFc37
colony forming units (CFU) isolated from plant tissues was
much higher than for the rhizosphere isolates in spite of the
same number of macroconidia used as inoculum. The CFU
number of the pathogenic isolate originating from the
interior of roots of plants pre-colonized with the rhizo-
spheric isolates was as low as 10% of the number of
pathogenic strain CFU isolated from plants inoculated with
the pathogen alone. A study of in vitro interactions
between the rhizosphere isolates and the pathogen suggests
that changes in plant colonization by the pathogen
observed in the presence of the rhizosphere isolates were
not connected with inhibition of the growth of the former
by a direct action of the latter.
Cytological studies of root sections 4 days after inocu-
lation indicated the presence of wall appositions, which are
symptoms of a defense reaction in plant cells, associated with
a fungal infection (Jaroszuk-Sciseł et al. 2008). This obser-
vation suggested that the 24-h interval between inoculation
with a rhizosphere strain and the introduction of the pathogen
may be sufficient to start a cascade of plant reactions
involved in the induced systemic reaction. Probably, chitin
oligosaccharides released by CWDE activities could be
effective elicitors of plant resistance responses (Furman-
Matarasso et al. 1999; Montesano et al. 2003).
Recently, complexes of CWDE have been found in
supernatants of autolyzing cultures of F. culmorum strains,
and the activities of particular enzymes have been deter-
mined (Jaroszuk-Sciseł et al. 2011).
The aim of the present study was to find answer whether
the efficiency of degradation of cell walls of plants (rye
roots) and fungi by enzymatic complexes originating from
supernatants of rhizosphere (plant growth promoting
[PGPF] and deleterious [DRMO]) and pathogenic Fusar-
ium spp. cultures could be related to differences in the type
of fungus–plant interactions.
Materials and methods
Fusarium spp. isolates
Five Fusarium spp. isolates belonging to three species were
used in this study: three Fusarium culmorum (W.G. Smith)
isolates with distinct effects on rye growth—two non-
pathogenic rhizospheric isolates (DEMFc2 and DEMFc5)
and one pathogenic isolate (DEMFc37) (Jaroszuk-Sciseł
et al. 2008), as well as two additional pathogenic isolates
provoking fusarium wilt symptoms and a 30% reduction in
shoot fresh weight—a Fusarium graminearum Schwabe
(teleomorph: Gibberella zeae Schweinitz) isolate (DEM-
Fc36) (Kurek and Jaroszuk 1997) and a pathogenic
Fusarium oxysporum Schlechtendal isolate (DEMFc38)
(Wolska-Mitaszko et al. 2007). All these Fusarium spp.
isolates are deposited at the Department of Environmental
Microbiology’s Fungal Collection (DEM), Maria Curie-
Skłodowska University in Lublin, Poland, and in Centra-
albureau voor Schimmelcultures Collections (CBS)
(accessed 16/07/06), P.O. Box 85167, NL-3508 AD
Utrecht, The Netherlands, http://www.cbs.knaw.nl/database/
index.htm. Nucleotide sequences of the isolates were depos-
ited in the NCBI GenBank.
The two nonpathogenic rhizosphere isolates, the PGPF
isolate DEMFc2 (CBS 120098, NCBI accession number
for nucleotide sequence DQ453700) and the DRMO isolate
DEMFc5 (CBS 120101, DQ450880), had been isolated
from the rhizosphere of healthy rye (Secale cereale L.)
grown in a field in the Lublin region (lat. 51.1500N and
long. 22.3400E) in Poland at the tillering stage (March) of
rye growth. The pathogenic isolates F. culmorum DEM-
Fc37 (CBS 120103, DQ450878), F. graminearum DEM-
Fc36 (CBS 120102, DQ453701), and F. oxysporum
DEMFc38 (CBS 120104, DQ450879) had been isolated
from winter wheat (Triticum aestivum L.) plants with
severe fusariosis symptoms.
The isolates were stored on rose bengal and strepto-
mycin (Martin 1950) 2% agar slants [(glucose, 10.0 g;
proteose peptone, 5.0 g; KH2PO4, 1.0 g; MgSO4, 0.5 g;
agar, 15.0 g; streptomycin, 30.0 mg; rose bengal, 30.0 mg
in 1,000 mL of H2O distilled water (DW)] at 4�C.
Preparation of macroconidia
Macroconidia for the preparation of inocula were obtained
from a culture grown on Martin liquid medium without
654 Arch Microbiol (2012) 194:653–665
123
Page 3
rose bengal dye and were prepared as described by
Jaroszuk-Sciseł et al. (2008, 2009). The isolates were culti-
vated in the darkness at 20�C and 60% of relative humidity
in an Innova 4900 growth chamber (New Brunswick
Scientific Co Inc., Edison, NJ, USA) at 120 rpm for 7 days,
and then, the cultures were filtered through 5 layers of
sterile gauze (cotton). Supernatants were collected, and the
macroconidia were pelleted by centrifugation (Beckman
J2-HS) at 10,000g for 15 min. The macroconidia were
washed three times by suspension in sterile DW (SDW)
and vortexed vigorously. Then, the numbers of macroco-
nidia in suspension were determined in a hemocytometer,
using an LM Nikon Eclipse E200 light microscope, and
adjusted by dilution to the desired concentration (density).
Preparation of mycelium
Mycelium biomass was obtained from 5-day cultures of the
Fusarium spp. isolates in 250 mL liquid Martin medium
without rose bengal dye in 1,000-mL Erlenmeyer flasks
shaken at 120 rpm in an Innova 4900 growth chamber. Liquid
Martin medium was inoculated with 1 9 105 CFU mL-1 of
macroconidia. The obtained biomass was separated by cen-
trifugation at 10,0009g and washed with SDW. The centri-
fugation was repeated five times. Next, the fungal mycelium
was frozen in liquid nitrogen, dried by lyophilization using
FreeZone 6 l (Labconco Co, Kansas City, Missouri, USA),
and stored for use in the preparation of FCW material.
Preparation of fungal cell walls (FCW)
Lyophilized five-day-old mycelia were used for the prep-
aration of FCW material according to the procedure
described by Gomez-Miranda et al. (1990) and recom-
mended by Alfonso et al. (1992). The mycelia were dis-
rupted at 4�C in an Ultra-Turax Ika�-Werke T18
homogenizer at 16,000 rpm for 10 min at 1-min intervals
and then centrifuged (Beckman J2-HS) at 10,0009g. The
pellet was frozen in liquid nitrogen, ground in a mortar, and
washed 5 times by centrifugation with SDW. Then, the
fungal mycelia were disintegrated by ultrasonic (Sonicator
XL2020, Misonix Inc, New Yersey, USA) treatment (15
intervals per 1 min at 150 W) and washed by centrifuga-
tion (10 min at 10,0009g) alternately with phosphate
buffer (50 mM, pH 6.5) and NaCl (1 M). This process was
repeated until the cell walls were free of cytoplasmic
material (protein-free), as seen under the light microscope
after Coomassie staining (i.e., until disappearance of
absorbance at 260–280 nm). Next, the cell walls were
washed ten times with SDW to eliminate NaCl. The
obtained FCW were dried by lyophilization and stored at
4�C. The FCW were (1) added to the medium as a carbon
source, (2) used as a substrate to determine the ability of
crude enzymatic preparations to hydrolyze FCW, and (3)
used in the next step to obtain FCW fractions.
Fractionation of FCW fractions
Fungal cell walls fractions were obtained using the chem-
ical method (Alfonso et al. 1995b; Gomez-Miranda et al.
1984, 1990; Leal et al. 1992) of extraction with 1 N NaOH
(Fig. 1). The following fractions were obtained: (a) (F1S)
galacto-manno-glucan (alkali soluble and water soluble at
20�C); (b) (F1I) glucan weakly associated with chitin,
precipitated with ethanol (alkali soluble at 20�C and water
insoluble); (c) (F3) glucan strongly associated with chitin
(alkali soluble at 70�C and water insoluble, obtained after
precipitation with ethanol; and (d) (F4) glucan-chitin
(alkali insoluble at 70�C and water insoluble).
Plant material
Seeds of rye (Secale cereale L. cv Dankowskie Złote pur-
chased from ‘‘Danko’’ Company Choryn–Racot, Poland)
were used in the experiments. They were surface-sterilized
according to the following procedure: the seeds were washed
in tap water for 30 min, then soaked for 10 min in a solution
of HgCl2 (0.1%, w/v), and next soaked for 10 min in a
solution of H2O2 (30%, v/v). After these treatments, they
were rinsed ten times in SDW. The seeds were then trans-
ferred into sterile Petri dishes (25 seeds per dish) and soaked
with 2 mL SDW and subsequently germinated for 5 days in
the darkness at 20�C and 60% of relative humidity in an
Innova 4900 growth chamber. Rye roots were separated from
the tops. The roots were five times washed with SDW with
agitation at 150 rpm, frozen in liquid nitrogen, lyophilized,
and stored at 4�C for use in the preparation of PCW.
Preparation of PCW
Protein- and lipid-free PCW was obtained from roots of
5-day-old rye seedlings (Cohen et al. 1998). The roots were
placed in a mixture of methanol and chloroform (2:1, v/v),
mixed at 150 rpm for 3 days at room temperature, and then
washed 10 times with SDW during 1 day. In the next step,
PCW was obtained using the method described above for
the preparation of FCW. PCW was used (1) in culture
medium as a carbon source or (2) as a substrate for the
determination of the ability of crude enzymatic prepara-
tions to hydrolyze PCW.
Crude enzyme preparations used to degrade PCW
and FCW
Crude enzyme preparations were obtained from 42-day-old
autolyzing culture filtrate liquids of Fusarium spp.
Arch Microbiol (2012) 194:653–665 655
123
Page 4
cultivated in 1,000 mL Erlenmeyer flasks with 200 mL of
modified liquid synthetic Reyes and Byrde (1973) medium
[KH2PO4, 1.0 g; MgSO4 � 7H2O, 0.5 g; KCl, 0.5 g;
(NH4)2SO4, 0.5 g; 1.0 mL of microelements solution
(Na2B4O7 � 10H2O, 100.0 mg; CuSO4 � 5H2O, 10.0 mg;
FeSO4 � 7H2O, 50.0 mg; MnSO4 � 5H2O, 10.8 mg;
(NH4)6Mo7O24 � 4H2O, 10.0 mg; ZnSO4 � 7H2O, 70.0 mg
in 1,000.0 mL of DW) and streptomycin, 30.0 mg in
1,000.0 mL of DW] supplemented with 0.5 mg PCW or
FCW per mL (0.05%, w/v) as described by Jaroszuk-Sciseł
et al. (2011). After inoculation with 100 lL of macroco-
nidia suspensions (2 9 108 macroconidia per mL), the
fungi were cultivated in the dark at 20�C and 60% relative
humidity in an Innova 4900 growth chamber under
stationary conditions for 42 days. The fungal biomass was
separated by filtration through sterile filter paper and the
filtrate liquids were additionally centrifuged (10,0009g for
10 min). Then, the supernatants were dialyzed three times
for 24 h against distilled water using dialysis tubing with a
3.5 kDa limit, dried by lyophilization, stored at -20�C and
used as crude enzyme preparations to study the efficiency
of PCW and FCW hydrolysis. The enzyme activities of the
crude enzyme preparations were periodically predeter-
mined and found to be stable for several months. Imme-
diately before their use in cell wall hydrolysis experiments
fivefold concentrated crude enzyme preparations were
obtained by solution of lyophilized culture supernatants
powders (7.5 mg mL-1) in phosphate (0.06 M) buffers
Cell wall material (freeze-dry lyophylizate)
Extraction with 1N NaOH at 200C
Supernatant Insoluble residue
Extraction with 1N NaOH at 700C
Centrifugation Centrifugation
Dialysis against water
Precipitation with EtOH (v/v)
Supernatant Precipitate Supernatant Insoluble residue
-(1,5)- galacto-manno-
glucan
F1S
fraction alkali (20oC) soluble,
water soluble
/ -(1,3)- glucan
weakly associated with chitin
F1I
fraction alkali (20oC) soluble,
water insoluble
F3
-(1,3)- glucan
strongly associated with chitin
fraction alkali (70oC) soluble,
water insoluble
F4
-(1,3)- glucan-chitin
complex
fraction alkali (70oC)
insoluble, water insoluble
Freeze-drying lyophylization
Freeze-drying lyophylization
Freeze-drying lyophylization
Freeze-drying lyophylization
Dialysis against water
Dialysis against water
Precipitation with EtOH (v/v)
βα /βαβ β
Fig. 1 Fractionation procedure of cell wall material (according to Gomez-Miranda et al. 1984, 1990; Alfonso et al. 1995b)
656 Arch Microbiol (2012) 194:653–665
123
Page 5
Ta
ble
1A
ctiv
itie
s(U
mL
-1)
of
enzy
mes
(glu
can
ase,
chit
inas
e,x
yla
nas
e,en
do
cell
ula
se,
exo
cell
ula
se,
pec
tin
ase,
po
lig
alac
tou
ron
ase)
inth
eso
luti
on
so
fcr
ud
een
zym
atic
pre
par
atio
ns
ori
gin
atin
gfr
om
42
-day
Fu
sari
um
spp
.(F
.cu
lmo
rum
iso
late
sD
EM
Fc2
,D
EM
Fc5
,an
dD
EM
Fc3
7,
F.
gra
min
earu
mis
ola
teD
EM
Fc3
6,
and
F.
oxy
spo
rum
iso
late
DE
MF
c38
)cu
ltu
res
gro
wn
in
the
pre
sen
ceo
f:p
lan
tce
llw
all—
PC
W(a
)o
rfu
ng
alce
llw
all—
FC
W(b
)as
C-s
ou
rces
atac
idic
and
alk
alin
ep
Hin
reac
tin
gm
ixtu
re
En
zym
esp
Hin
reac
tin
gm
ixtu
re
Aci
dic
Alk
alin
e
Iso
late
sIs
ola
tes
DE
MF
c2D
EM
Fc5
DE
MF
c37
DE
MF
c36
DE
MF
c38
DE
MF
c2D
EM
Fc5
DE
MF
c37
DE
MF
c36
DE
MF
c38
a Glu
can
ase
1,1
64
.00
aB5
65
.30
bD
87
7.0
0aC
43
.55
eE2
,60
4.3
0aA
58
3.6
0aB
30
3.4
0b
C2
15
.80
bD
8.3
0eE
3,1
34
.10
dA
Ch
itin
ase
60
.35
dC
21
8.6
5d
A9
8.0
5d
B6
1.9
0d
C2
18
.35
fA3
4.8
5fB
42
.55
eA2
0.1
5eC
20
.55
cC3
6.5
5eB
Xy
lan
ase
63
.40
dE
1,6
50
.20
aA8
19
.25
bB
13
9.1
0b
D4
86
.85
cC4
7.2
5eD
1,6
24
.35
aA7
43
.80
aB1
24
.20
aC2
3.4
5fE
En
do
cell
ula
se3
8.0
5eE
11
4.2
0eB
96
.80
dC
75
.65
cD3
16
.75
eA1
7.3
0g
D7
3.3
0d
B6
5.3
5d
C1
5.3
5d
D2
88
.35
cA
Ex
oce
llu
lase
38
.60
eC6
3.6
5fB
32
.95
eC2
5.2
0fD
12
5.3
5g
A1
34
.60
dA
77
.85
dB
21
.90
eC1
3.8
5d
D7
4.2
0d
B
Pec
tin
ase
78
1.0
0b
A5
61
.35
bC
18
7.5
0cE
22
1.7
5aD
67
7.5
5b
B4
54
.75
bA
31
3.6
bC
14
3.8
5cD
13
1.2
0aD
41
6.1
5b
B
Po
lig
alac
tou
ron
ase
25
9.9
5cC
29
9.4
0cB
18
4.4
5cD
13
9.6
5b
E3
70
.00
dA
24
4.4
0cA
B2
27
.25
cB1
47
.10
cC1
06
.80
bD
26
7.7
5cA
b Glu
can
ase
61
4.8
5aA
55
1.4
0aB
37
4.2
5aC
37
0.1
0aC
53
2.5
5b
B3
31
.85
aD3
77
.05
dC
31
2.4
0aD
90
1.9
5aA
51
8.3
5aB
Ch
itin
ase
12
7.7
5b
A2
0.4
5eB
11
.25
gC
6.3
5g
D1
15
.60
eA1
3.0
0d
B9
.90
fC1
0.0
0eC
4.9
0fD
86
.00
eA
Xy
lan
ase
11
5.9
5cD
57
9.4
5aB
12
4.8
0cE
29
4.8
0b
C1
,00
0.2
5aA
16
.25
cD1
58
.15
bB
18
.05
dD
66
.00
dC
40
7.4
5b
A
En
do
cell
ula
se1
5.2
5d
E2
7.1
5d
D3
2.6
5eC
71
.60
eB2
35
.40
cA0
.00
eD2
3.2
0eB
19
.70
dB
14
.90
eC1
86
.85
cA
Ex
oce
llu
lase
12
.25
eC1
3.5
5fC
15
.95
fB1
0.8
0fD
29
.75
fA1
3.0
5d
B3
2.3
5d
A2
.10
fD4
.45
fC3
8.1
5fA
Pec
tin
ase
11
8.0
0b
cD1
80
.40
cB2
02
.70
bA
18
3.7
0cB
14
1.4
5d
C6
2.0
5b
C1
28
.25
cB1
78
.20
bA
16
1.5
5b
A1
35
.95
dB
Po
lig
alac
tou
ron
ase
0.0
0fC
21
7.5
5b
A9
0.3
5d
B9
6.2
0d
B2
34
.55
cA0
.00
eE1
60
.95
bA
77
.25
cD9
4.2
0cC
14
2.8
5d
B
Mea
nv
alu
esw
ith
inth
eli
ne
for
each
pH
ran
ge
foll
ow
edb
yd
iffe
ren
tca
pit
alle
tter
sar
esi
gn
ifica
ntl
yd
iffe
ren
tac
cord
ing
toth
ele
ast
sig
nifi
can
td
iffe
ren
ce(L
SD
)te
st(p
\0
.05
).M
ean
val
ues
wit
hin
the
colu
mn
foll
ow
edb
yd
iffe
ren
tsm
all
lett
ers
are
sig
nifi
can
tly
dif
fere
nt
acco
rdin
gto
the
leas
tsi
gn
ifica
nt
dif
fere
nce
(LS
D)
test
(p\
0.0
5)
Arch Microbiol (2012) 194:653–665 657
123
Page 6
(pH 5.6 or pH 8.8) with 0.02% (w/v) sodium azide. These
solutions were then centrifuged for 5 min at 14,0009g and
sterilized by filtration through a 0.22 lm filter (FP 030/3
Sigma Aldrich, St. Louis, Missouri, USA).
The suspensions of the crude enzyme preparations were
used to determine the activities of b-1,3-glucanase (EC
3.2.1.6), chitinase (EC 3.2.1.14), xylanase (EC 3.2.1.8),
endocellulase (EC 3.2.1.4), exocellulase (EC 3.2.1.91),
pectinase (EC 3.2.1.15), and polygalacturonase (EC
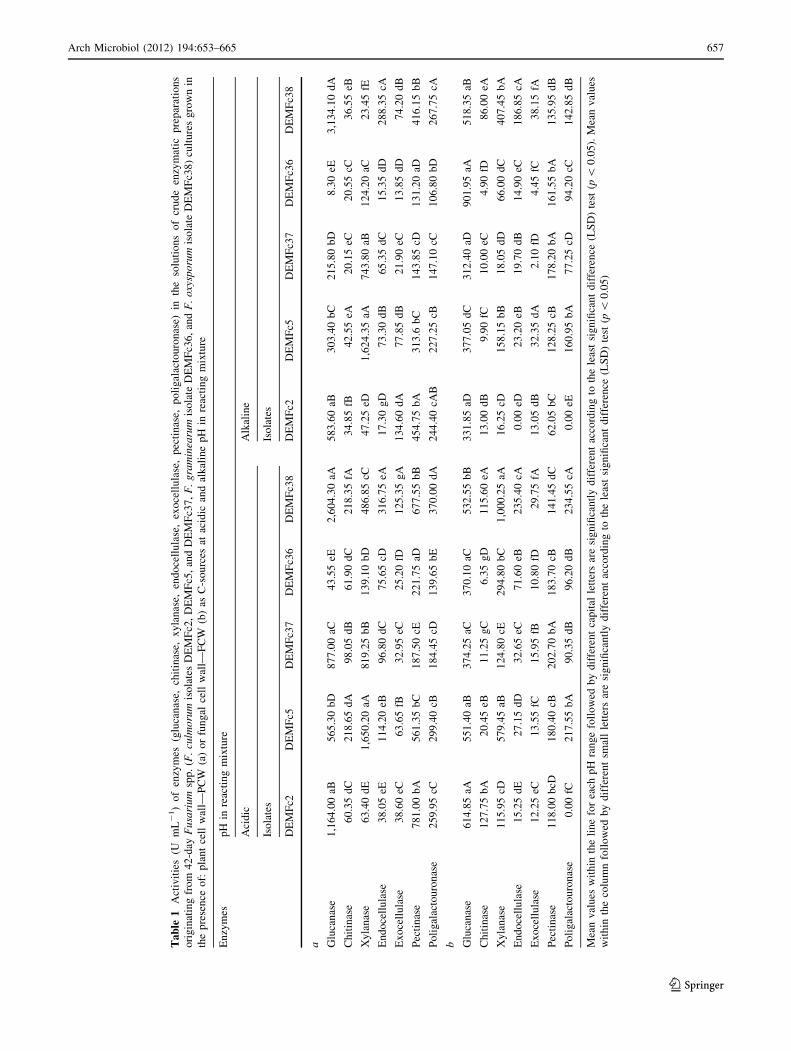
3.2.1.15) (Table 1a, b) by methods described in detail by
Jaroszuk-Sciseł et al. (2011).
The activities of acidic and alkaline b-1,3-glucanase,
xylanase, endocellulase, exocellulase, polygalacturonase, and
pectinase were assayed according to the Alfonso et al. (1992)
method by estimating the liberation of reducing substances
during hydrolysis of a 0.5% (w/v) solution of laminarin from
Laminaria digitata (L-9634 Sigma Aldrich, St. Louis,
Missouri, USA); a 1% (w/v) suspension of xylan from oat spelts
(Sigma X-0627); a 0.7% (w/v) solution of carboxymethyl
cellulose-CMC (Sigma C-5678); a 1% (w/v) suspension of
Avicel/Sigmacell� type20 cellulose (Sigma S-3504); a 0.1%
(w/v) solution of polygalacturonic acid (Sigma 81325); and a
0.1% (w/v) solution of pectin from apple (Sigma P-8471),
respectively. The reacting mixtures for these activity assays
contained 1 mL solutions of appropriate crude enzyme
preparations and 1 mL of appropriate substrate solutions or
substrate suspensions in phosphate buffers (pH 5.6 or 8.8).
Supernatants after thermal inactivation (10 min in 100�C)
were used as controls. The reacting mixtures were incubated at
37�C for 3 h with gentle agitation (80 rpm). Immediately after
incubation, soluble reducing sugars released from these sub-
strates were measured by the Nelson (1944) and Somogyi
(1945) method modified by Hope and Burns (1987). Absor-
bance was measured at 520 nm within 20 min. For pectinase
and polygalacturonase activity measurement, D-galacturonic
acid (Fluka 73960, St. Louis, Missouri, USA) was used as a
standard and for the measurements of the other enzyme
activities glucose was the standard. Enzyme activities
expressed as U mL-1 of supernatant were calculated from the
linear portion of the activity curves. One unit of activity was
defined as the amount causing the liberation of 1 lg of
reducing sugars (glucose or galacturonic acid) equivalent
min-1 under the above-mentioned conditions and at a corre-
sponding pH.
The activities of acidic and alkaline chitinases were
determined according to the Rossner (1991) method rec-
ommended by Alef and Nannipieri (1995). Chitinase
activity was assayed by estimating the N-acetyl-D-gluco-
samine (GlcNAc) equivalents liberated after hydrolyzing
the colloidal chitin suspension. The reacting mixture was
composed of 1 mL solutions of lyophilized culture super-
natants in phosphate buffers (pH 5.6 or 8.8) and 1 mL of
1% (w/v) colloidal chitin suspension in phosphate buffers
(pH 5.6 or 8.8). Thermally inactivated (10 min in 100�C)
supernatant was a control. These reacting mixtures were
incubated at 37�C for 3 h with gentle agitation (80 rpm).
The reaction was terminated by GlcNAc extraction with
2 M KCl, and then the reacting mixture was filtered. Glc-
NAc was determined in a reaction with saturated sodium
borate and p-dimethylaminobenzaldehyde (DMBA).
Absorbance was measured at 585 nm within 20 min. The
standard was Sigma A-8625 GlcNAc. Chitinase activity
expressed as U mL-1 of supernatant was calculated from
the linear portion of the activity curves. One unit of activity
was defined as the amount causing the liberation of 1 lg of
GlcNAc equivalent from chitin min-1 under the above-
mentioned conditions and at a corresponding pH.
Hydrolysis of FCW and PCW by crude enzyme
preparations
FCW and PCW degradation was studied according to the
method described by Alfonso et al. (1992). The PCW and
FCW suspensions (2 mg mL-1) in phosphate (0.06 M)
buffers (pH 5.6 or pH 8.8) with 0.02% (w/v) sodium azide
were prepared by ultrasonic disintegration (15 intervals per
1 min at 150 W) and then sterilized by autoclaving and
sterile washing with SDW by centrifugation (10 min at
10,0009g). Two mL of this suspension was incubated with
2 mL of an appropriate crude enzyme preparation solution
(7.5 mg mL-1) at 37�C during 4 days. After 24, 48, 72, and
96 h of incubation, the reacting mixtures were filtered and
additionally centrifuged. Immediately after filtration and
centrifugation, liberation of reducing sugars during PCW
and FCW hydrolysis was measured using methods descri-
bed by Nelson (1944) and Somogyi (1945), and GlcNAc
liberation during FCW hydrolysis was measured according
to Rodriguez-Kabana et al. (1983) and Rossner (1991).
Appropriate controls were included: (1) the crude enzyme
preparation solution after thermal inactivation (10 min in
100�C) and (2) PCW and FCW suspensions.
Statistical analysis
Statistical analysis of data was performed on three repli-
cates from each treatment. Standard deviations (shown as
deviation bars) were determined using Microsoft� Excel
2000 (Microsoft Corp., Redmond, Washington, USA)
Analysis of variance ANOVA was used to determine sig-
nificance of differences between the individual enzyme
activities in the crude enzymatic preparation. Data were
analyzed using STATISTICA 8.0 (StatSoft Inc., Tulsa,
Oklahoma., USA) (Armitage and Berry 1987). Mean sep-
aration was accomplished using the least significant dif-
ference (LSD) test. Significance was evaluated at p \ 0.05
for all tests.
658 Arch Microbiol (2012) 194:653–665
123
Page 7
The Pearson correlation coefficient (R) and the linear
regression coefficient (R2) were determined (using Micro-
soft� Excel 2000) to show the direction and strength of the
relationship between the activities of the particular
enzymes in crude preparations from the cultures of
Fusarium spp. isolates and the efficiency of these prepa-
rations in cell wall hydrolysis. The efficiency was expres-
sed as the amounts of reducing sugars released from PCW
and the amounts of reducing sugars and GlcNAc released
from FCW.
Results and discussion
Activities of CWDE in culture supernatants of F. culmo-
rum isolates grown in media with PCW or FCW as C
sources
Lytic enzymes, with activities hydrolyzing the polymers
building PCW and FCW, released by fungi as a result of
autolysis of their mycelia and secretion during their growth,
are involved both in the colonization and penetration of plant
tissues by pathogenic and endophytic fungi and in the lib-
eration of plant defense elicitors (Ebel and Casio 1994;
Montesano et al. 2003; Shibuya and Minami 2001).
For hydrolysis of PCW as well as FCW to be efficient
and complete, synergistic action of several enzymes is
required (Alfonso et al. 1995a; Collins et al. 2005; Lalaoui
et al. 2000; Perez-Leblic et al. 1982; Reyes et al. 1977).
The activities of the hydrolases present in the crude
preparations obtained from 42-day culture supernatants of
Fusarium isolates grown in a medium containing purified
rye root cell walls (PCW) and in a medium containing
purified Fusarium isolate own cell walls (FCW) as C
sources are shown in Table 1a, and b, respectively.
Activities of seven CWDE (glucanases, chitinases, xylan-
ases, endocellulases, exocellulases, pectinases, and poli-
galacturonases) were detected in solutions of crude
enzymes preparations originated from autolyzing 42-day
cultures of three Fusarium culmorum isolates differently
affecting rye growth as well as in preparations obtained
from cultures of F. graminearum and F. oxysporum. The
activities of glucanases, chitinases, xylanases, and pectin-
ases measured in those solutions were higher than those of
the other enzymes tested and were also higher in acidic
reacting mixtures than in alkaline ones.
These enzyme activities in 14-, 28- and 42-day-old
culture supernatants of these three F. culmorum grown in
media containing one of the four C sources (PCW, FCW,
glucose, and chitin) were described in the previous Jar-
oszuk-Sciseł et al. (2011) paper; however, the values from
one period only (maximal from those three periods) were
presented. The results of those earlier studies indicate that
these 42-day-old autolyzing cultures are a very good source
of CWDE enzymes, and a majority of the enzymatic
activities determined were maximal exactly on day 42.
Alfonso et al. (1995a, b) have obtained similar results
(describing maximal autolysis in 60-day cultures).
A comparison of CWDE activities in solutions of crude
ezymes preparations originated from supernatants of cul-
tures containing PCW and FCW as C sources showed that
only the activity of polygalacturonases was enhanced in all
of the cultures tested when FCW was replaced with PCW.
In the preparations obtained from supernatant of PGPF
F. culmorum DEMFc2 grown in a culture medium containing
FCW as a C source, the highest activities were measured for
acidic glucanases and chitinases. However, the activity of
glucanases was higher in the preparations originated from
culture supernatant of this isolate grown in the medium con-
taining PCW than in the medium containing FCW as a C
source (Table 1a, b).
The activity of acidic xylanase in the solutions of crude
enzymes preparations originated from cultures of DRMO
DEMFc5 and pathogenic DEMFc37 was significantly
higher when the isolates were grown in the medium con-
taining PCW as a C source than when they were grown in
the presence of FCW as a C source.
The high xylanase activity in the crude enzyme prepa-
rations from the pathogenic Fusarium strains grown in the
medium with rye cell wall is not surprising since in
graminaceous monocotyledonous plants, the primary and
secondary cell walls consist mainly of hemicellulose and
xylan (Belien et al. 2006; Carpita 1996; Hatsch et al. 2006;
Labavitch and Ray 1978).
There are reports suggesting that endoxylanases might
be pathogenicity factors for phytopathogenic microorgan-
isms. Indirect evidence for their role in pathogenesis has
been obtained by analyzing the production of cell wall-
degrading enzymes upon infection or by examining the
effect of purified endoxylanase on plant cells and infected
tissues (Belien et al. 2006; Brito et al. 2006; Polizeli et al.
2005). For instance, bacterial strains of Erwinia chrysant-
hemi that infect maize have been found to secrete more
endoxylanase activity than strains that infect only dicoty-
ledonous plants (Braun and Rodrigues 1993). Similarly,
fungal pathogens, such as Septoria nodorum, that infect
graminaceous monocotyledons have been reported to
secrete more xylan-degrading enzymes than pectinases
(Cooper et al. 1988; Lehtinen 1993). Fusarium-infected
barley has also been found to contain considerable amounts
of endoxylanase (Schwarz et al. 2002). Since xylans rep-
resent a large proportion of the hemicellulosic fraction of
cereal cell wall matrices, xylan-degrading enzymes are
expected to be important components of the offensive
arsenal of cereal pathogens (Cooper et al. 1988; Wanjiru
et al. 2002) and may have a role similar to that of pectic
enzymes in the infection of dicotyledons.
Arch Microbiol (2012) 194:653–665 659
123
Page 8
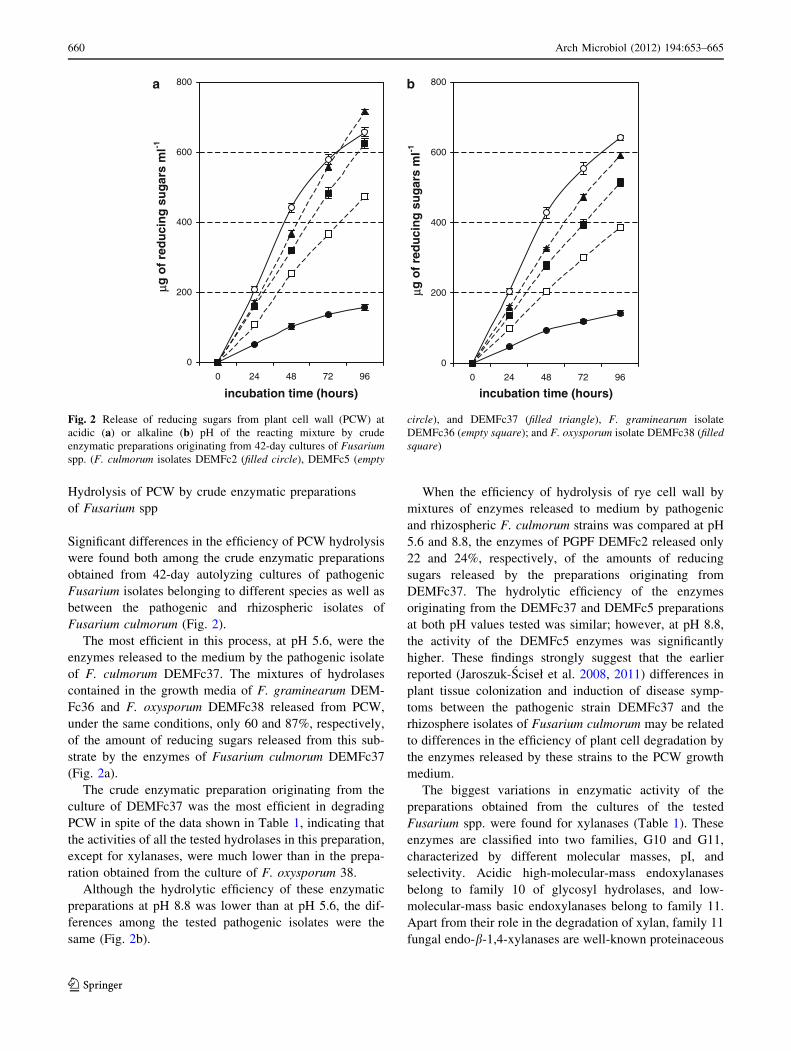
Hydrolysis of PCW by crude enzymatic preparations
of Fusarium spp
Significant differences in the efficiency of PCW hydrolysis
were found both among the crude enzymatic preparations
obtained from 42-day autolyzing cultures of pathogenic
Fusarium isolates belonging to different species as well as
between the pathogenic and rhizospheric isolates of
Fusarium culmorum (Fig. 2).
The most efficient in this process, at pH 5.6, were the
enzymes released to the medium by the pathogenic isolate
of F. culmorum DEMFc37. The mixtures of hydrolases
contained in the growth media of F. graminearum DEM-
Fc36 and F. oxysporum DEMFc38 released from PCW,
under the same conditions, only 60 and 87%, respectively,
of the amount of reducing sugars released from this sub-
strate by the enzymes of Fusarium culmorum DEMFc37
(Fig. 2a).
The crude enzymatic preparation originating from the
culture of DEMFc37 was the most efficient in degrading
PCW in spite of the data shown in Table 1, indicating that
the activities of all the tested hydrolases in this preparation,
except for xylanases, were much lower than in the prepa-
ration obtained from the culture of F. oxysporum 38.
Although the hydrolytic efficiency of these enzymatic
preparations at pH 8.8 was lower than at pH 5.6, the dif-
ferences among the tested pathogenic isolates were the
same (Fig. 2b).
When the efficiency of hydrolysis of rye cell wall by
mixtures of enzymes released to medium by pathogenic
and rhizospheric F. culmorum strains was compared at pH
5.6 and 8.8, the enzymes of PGPF DEMFc2 released only
22 and 24%, respectively, of the amounts of reducing
sugars released by the preparations originating from
DEMFc37. The hydrolytic efficiency of the enzymes
originating from the DEMFc37 and DEMFc5 preparations
at both pH values tested was similar; however, at pH 8.8,
the activity of the DEMFc5 enzymes was significantly
higher. These findings strongly suggest that the earlier
reported (Jaroszuk-Sciseł et al. 2008, 2011) differences in
plant tissue colonization and induction of disease symp-
toms between the pathogenic strain DEMFc37 and the
rhizosphere isolates of Fusarium culmorum may be related
to differences in the efficiency of plant cell degradation by
the enzymes released by these strains to the PCW growth
medium.
The biggest variations in enzymatic activity of the
preparations obtained from the cultures of the tested
Fusarium spp. were found for xylanases (Table 1). These
enzymes are classified into two families, G10 and G11,
characterized by different molecular masses, pI, and
selectivity. Acidic high-molecular-mass endoxylanases
belong to family 10 of glycosyl hydrolases, and low-
molecular-mass basic endoxylanases belong to family 11.
Apart from their role in the degradation of xylan, family 11
fungal endo-b-1,4-xylanases are well-known proteinaceous
0
200
400
600
800
incubation time (hours)
μg o
f re
du
cin
g s
ug
ars
ml-1
a
0
200
400
600
800
0 24 48 72 96 0 24 48 72 96
incubation time (hours)
μg o
f re
du
cin
g s
ug
ars
ml-1
b
Fig. 2 Release of reducing sugars from plant cell wall (PCW) at
acidic (a) or alkaline (b) pH of the reacting mixture by crude
enzymatic preparations originating from 42-day cultures of Fusariumspp. (F. culmorum isolates DEMFc2 (filled circle), DEMFc5 (empty
circle), and DEMFc37 (filled triangle), F. graminearum isolate
DEMFc36 (empty square); and F. oxysporum isolate DEMFc38 (filledsquare)
660 Arch Microbiol (2012) 194:653–665
123
Page 9
elicitors of defense response reactions in plants in a way
that is independent of their enzymatic activity (Enkerli
et al. 1999). The activity of alkaline xylanases in prepa-
rations obtained from Fusarium spp. cultures was usually
lower than that of acidic ones (Table 1).
The obtained results suggest that xylanase activity may
play a significant role in PCW degradation and pathogen-
esis. Experimental evidence demonstrating the requirement
for a single endoxylanase for virulence has been provided
by Brito et al. (2006), who showed that deletion of xyn11A
from Botrytis cinerea strongly affected its ability to infect
tomato leaves and grape berries. The appearance of sec-
ondary lesions on tomato leaves was delayed, and the
average size of those infections, that were actually
spreading, was reduced by more than 70%.
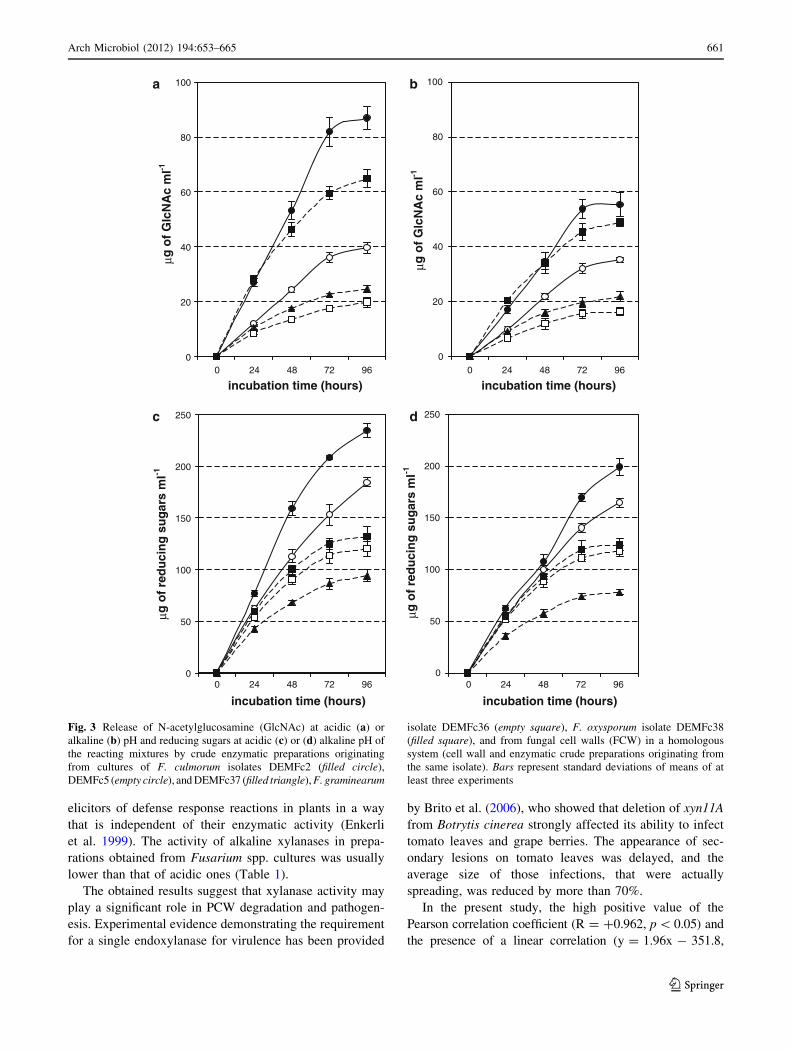
In the present study, the high positive value of the
Pearson correlation coefficient (R = ?0.962, p \ 0.05) and
the presence of a linear correlation (y = 1.96x - 351.8,
0
20
40
60
80
100
incubation time (hours)
μg o
f G
lcN
Ac
ml-1
a
0
20
40
60
80
100
0 24 48 72 960 24 48 72 96
0 24 48 72 96 0 24 48 72 96
incubation time (hours)
μg o
f G
lcN
Ac
ml-1
b
0
50
100
150
200
250
incubation time (hours)
μg o
f re
du
cin
g s
ug
ars
ml-1
c
0
50
100
150
200
250
incubation time (hours)
μg o
f re
du
cin
g s
ug
ars
ml-1
d
Fig. 3 Release of N-acetylglucosamine (GlcNAc) at acidic (a) or
alkaline (b) pH and reducing sugars at acidic (c) or (d) alkaline pH of
the reacting mixtures by crude enzymatic preparations originating
from cultures of F. culmorum isolates DEMFc2 (filled circle),
DEMFc5 (empty circle), and DEMFc37 (filled triangle), F. graminearum
isolate DEMFc36 (empty square), F. oxysporum isolate DEMFc38
(filled square), and from fungal cell walls (FCW) in a homologous
system (cell wall and enzymatic crude preparations originating from
the same isolate). Bars represent standard deviations of means of at
least three experiments
Arch Microbiol (2012) 194:653–665 661
123
Page 10
R2 = 0.926, p \ 0.05) pointed to a close dependence
between the ability of the crude preparations originating
from the cultures of the tested Fusarium strains to
reduce the sugars released from PCW and their xylanase
activity.
Hydrolysis of FCW by Fusarium spp. crude enzymatic
preparations
The hydrolytic efficiency of enzymatic complexes was
tested in homologous (enzymes and cell wall substrates
0
20
40
60
80
100μg
of
Glc
NA
c m
l-1FCW of DEMFc2a
0
20
40
60
80
100FCW of DEMFc5
0
20
40
60
80
100FCW of DEMFc36
0
20
40
60
80
100
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
incubation time (hours)
FCW of DEMFc38
0
20
40
60
80
100FCW of DEMFc37
0
50
100
150
200
250
mg
of
red
uci
ng
su
gar
s m
l-1
b
0
50
100
150
200
250
0
50
100
150
200
250
0
50
100
150
200
250
0 24 48 72 960 24 48 72 960 24 48 72 960 24 48 72 960 24 48 72 96
0 24 48 72 960 24 48 72 960 24 48 72 960 24 48 72 960 24 48 72 96
0
50
100
150
200
250
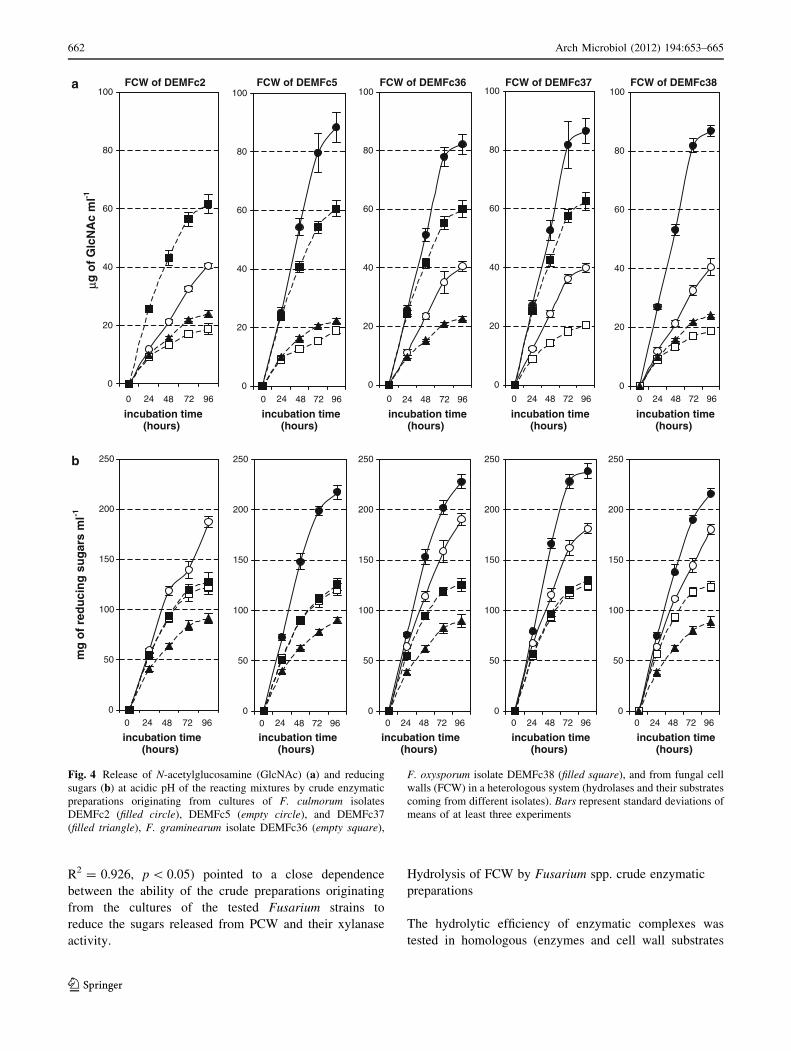
Fig. 4 Release of N-acetylglucosamine (GlcNAc) (a) and reducing
sugars (b) at acidic pH of the reacting mixtures by crude enzymatic
preparations originating from cultures of F. culmorum isolates
DEMFc2 (filled circle), DEMFc5 (empty circle), and DEMFc37
(filled triangle), F. graminearum isolate DEMFc36 (empty square),
F. oxysporum isolate DEMFc38 (filled square), and from fungal cell
walls (FCW) in a heterologous system (hydrolases and their substrates
coming from different isolates). Bars represent standard deviations of
means of at least three experiments
662 Arch Microbiol (2012) 194:653–665
123
Page 11
originating from the same isolate) (Fig. 3a, b) and heter-
ologous (Fig. 4a, b) (hydrolases and their substrates orig-
inating from different isolates) systems.
The activities of hydrolases contained in crude prepa-
rations obtained from the supernatants of fungal cultures
grown in media with their own cell walls as C sources are
shown in Table 1b.
In homologous systems, F. oxysporum DEMFc38 was
found to be the most efficient of the tested pathogenic
strains at both pH values (5.6 and 8.8). The efficiencies of
release of reducing sugars in the homologous system of
F. graminearum DEMFc36 at pH 5.6 and 8.8 were 91 and
95%, respectively, of those of F. oxysporum (Fig. 3b).
However, there was a much larger difference in the
amounts of GlcNAc released in these two homologous
systems (30.6% at pH 5.6 and 24% at pH 8.8) (Fig. 3a).
The efficiency of release of reducing sugars in the
homologous system of pathogenic F. culmorum DEMc37 at
both pH values was the lowest among the tested pathogens.
Among the homologous systems, the rhizosphere isolate
of F. culmorum DEMFc2 (PGPF) was the most efficient in
the hydrolysis of FCW at both pH values tested. The
amount of reducing sugars released at pH 5.6 accounted for
177% of the amount released in the most efficient homol-
ogous system of the pathogenic F. oxysporum isolate.
When the amounts of reducing sugars released from the
same substrates by hydrolases of the various individual
pathogens (heterologous system) were compared, no sig-
nificant differences were found between the efficiencies of
the enzymes originating from the cultures of DEMFc36
and DEMFc38. Much less efficient in this process were the
enzymes present in the supernatant of the autolyzed culture
of the isolate DEMFc37 (Fig. 3b).
When the efficiency of Fusarium cell wall hydrolysis in
homologous (Fig. 3a, b) and heterologous systems
(Fig. 4a, b) was compared, no significant differences were
found in the amounts of reducing sugars and GlcNAc
released in the homologous system of the rhizosphere
isolate DEMFc2 and the heterologous systems consisting
of hydrolases of DEMFc2 and cell walls of the pathogenic
strains F. culmorum DEM Fc37 or F. graminearum
DEMFc36 or F. oxysporum DEMFc38 at pH 5.6 and 8.8.
The results presented here indicate that the efficiency of
FCW degradation by the individual strains is mainly rela-
ted to the activities of two hydrolases released by them to
the medium: glucanases and chitinases. In general, glu-
canase activities in the crude preparations used to hydro-
lyze FCW were about 10 times higher than chitinase
activities, and the amounts of reducing sugars released by
glucanases from FCW were higher than the amounts of
GlcNAc (Table 1; Figs. 3a, b, 4a, b). There are reports
showing that effective degradation of FCW is catalyzed by
the synergistic action of chitinases releasing GlcNAc and
b-1,3-glucanases releasing reducing sugars (Adams 2004;
Alfonso et al. 1992, 1995a, b; Isaak and Gokhale 1982;
Skujins et al. 1965; Lahoz et al. 1976; Santamaria et al.
1995). The highest activities of chitinases and b-1,3-glu-
canases were found in the preparation from the culture of
the PGPF strain DEMFc2. This strain also released the
highest amounts of GlcNAc and reducing sugars from
FCW (Table 1; Figs. 3a, b, 4a, b). By contrast, Lynch et al.
(1985) found that culture filtrates of F. tricinctum having
maximum b-glucanase activity but no chitinase activity
were efficient in liberating protoplasts from the young
mycelium of the same species and in lysing F. oxysporum
mycelium. A study by Perez-Leblic et al. (1982) also
indicated that b-1,3-glucanase was the only enzyme present
at high levels in all of the extensively autolyzed cultures
(Neurospora crassa, Botrytis cinerea, Polystictus versi-
color, Aspergillus nidulans, Schizophyllum commune,
Aspergillus niger and Mucor mucedo) those authors tested.
The highest effectiveness of DEMFc2 in degrading
FCW can explain the protective effect of pre-inoculation of
rye with this isolate against infection by the pathogenic
strain DEMFc37 (Jaroszuk-Sciseł et al. 2008). This is
because one product of chitin degradation by the PGPF
isolate is an oligosaccharide with elicitor activity which
could induce plant resistance (Shibuya and Minami 2001).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%DEMFc2 DEMFc5 DEMFc37
% o
f F
CW
dry
wei
gh
t
GCH G1 G2 GGM
Fig. 5 Cell wall fractions of the particular Fusarium culmorumisolates: (GCH) – b-(1,3)-glucan–chitin complex fraction—F4;
(G1) – fraction of a/b-(1,3)-glucan weakly associated with chitin—
F1I; (G2) – fraction of a/b-(1,3)-glucan strongly associated with
chitin—F3; (GGM) – b-(1,5)-galacto-manno-glucan fraction—F1S
Arch Microbiol (2012) 194:653–665 663
123
Page 12
When the efficiency of FCW hydrolysis in homologous
and heterologous systems was compared, the obtained
results (Figs. 3a, b, 4a, b) suggested that the degree of
FCW hydrolysis was affected by the composition and
activities of enzymatic compounds present in the reacting
mixture rather than the origin of the FCW used. Santamaria
et al. (1995) also found that the level of efficiency of
F. oxysporum sp. lycopersici cell wall lysis was determined
by the origin of the hydrolases used.
The highest positive Pearson correlation coefficient value
(R = ?0.977) and a linear correlation (y = 2.87x - 43.584,
R2 = 0.955, p \ 0.05) was found between chitinase
activity in enzymatic preparations obtained from culture
supernatants of the Fusarium spp. strains and the efficiency
of FCW hydrolysis in homologous and heterologous
systems.
Chemical fractionation of the tested Fusarium spp. cell
walls indicated that there were no significant differences in
the contents of glucochitin (GCH), glucanases weakly
(F1I) and strongly (F3) associated with chitin, and galacto-
manno-glucan (F1S) fractions between the tested rhizo-
sphere and pathogenic Fusarium culmorum strains (Fig. 5).
The chemical composition of the fungal cell wall is species
specific but also varies depending on the development and
morphological stage as a result of aging (Bowman and Free
2006; Ruiz-Herrera 1991).
Acknowledgments The scientific research was financed from sci-
ence funding resources as a personal research project no. N N310
441338 in the years 2010–2013.
References
Adams DJ (2004) Fungal cell wall chitinases and glucanases.
Microbiology 150:2029–2035
Akimitsu K, Isshiki A, Ohtani K, Yamamoto H, Eshel D, Prusky D
(2004) Sugars and pH: a clue to the regulation of fungal cell
wall-degrading enzymes in plants. Physiol Mol Plant Pathol
65:271–275
Alef K, Nannipieri P (1995) Methods in applied soil microbiology and
biochemistry. Academic Press, New York, USA, pp 360–361
Alfonso C, del Amo F, Nuero OM, Reyes F (1992) Physiological and
biochemical studies on Fusarium oxysporum f. sp. lycopersicirace 2 for its biocontrol by nonpathogenic fungi. FEMS
Microbiol Lett 99:169–174
Alfonso C, Nuero OM, Santamaria F, Reyes F (1995a) Purification of
a heat-stable chitin deacetylase from Aspergillus nidulans and its
role in cell wall degradation. Curr Microb 30:49–55
Alfonso C, Santamaria F, Nuero OM, Prleto A, Leal JA, Reyes F
(1995b) Biochemical studies on the cell wall degradation of
Fusarium oxysporum f. sp. Iycopersici race 2 by its own lytic
enzymes for its biocontrol. Lett Appl Microbiol 20:105–109
Annis SL, Goodwin PH (1997) Recent advances in the molecular
genetics of plant cell wall-degrading enzymes produced by plant
pathogenic fungi. Eur J Plant Pathol 103:1–14
Apel PC, Panaccione DG, Holden FR, Walton JD (1993) Cloning and
targeted gene disruption of XYL1, a beta 1,4-xylanase gene from
the maize pathogen Cochliobolus carbonum. Mol Plant Microbe
Interact 6:467–473
Armitage P, Berry G (1987) Statistical methods in medical research.
Blackwell Science Publications, Oxford
Belien T, Van Campenhout S, Robben J, Volckaert G (2006)
Microbial endoxylanases: effective weapons to breach the plant
cell wall barrier or, rather, triggers of plant defense systems?
Mol Plant Microbe Interact 19:1072–1081
Boothby D, Magreola NO (1984) Production of polysaccharide
degrading enzymes by Cochliobolus sativus and Fusariumculmorum grown in liquid culture. Trans Br Mycol Soc 83:275–
280
Bowman SM, Free SJ (2006) The structure and synthesis of the fungal
cell wall. BioEssays 28:799–808
Braun EJ, Rodrigues CA (1993) Purification and properties of an
endoxylanase from a corn stalk rot strain of Erwinia chrysant-hemi. Phytopathology 83:332–338
Brito N, Espino JJ, Gonzalez C (2006) The endo-beta-1,4-xylanase
xyn11A is required for virulence in Botrytis cinerea. Mol Plant
Microbe Interact 19:25–32
Carpita NC (1996) Structure and biogenesis of the cell walls of
grasses. Annu Rev Plant Physiol Plant Mol Biol 4:445–476
Carpita NC, Gibeaut DM (1993) Structural models of primary cell
walls in flowering plant: consistency of molecular structure with
the physical properties of the walls during growth. Plant J 3:1–30
Cohen CK, Fox TC, Garvin DF, Kochian L (1998) The role of iron-
deficiency stress responses in stimulating heavy-metal transport
in plants. Plant Physiol 116:1063–1072
Collins T, Gerday C, Feller G (2005) Xylanases, xylanase families
and extremophilic xylanases. FEMS Microbiol Rev 29:3–23
Cooper RM, Longman D, Campbell A, Henry M, Lees PE (1988)
Enzymic adaptation of cereal pathogens to the monocotyledon-
ous primary wall. Physiol Mol Plant Pathol 32:33–47
Degefu Y, Fagerstrom R, Kalkkinen N (1995) Purification and partial
characterization of xylanase from fungal pathogen Helmintho-sporium furcicum (Pass). Eur J Plant Pathol 101:291–299
Degefu Y, Paulin L, Lubeck PS (2001) Cloning, sequencing and
expression of a xylanase gene from the maize pathogen
Helminthosporium turcicum. Eur J Plant Pathol 107:457–465
Douaiher M-N, Nowak E, Dumortier V, Durand R, Reignault PH,
Halama P (2007) Mycosphaerella graminicola produces a range
of cell wall-degrading enzyme activities in vitro that vary with
the carbon source. Eur J Plant Pathol 117:71–79
Ebel J, Casio EG (1994) Elicitors of plant defense responses. Int Rev
Cytol 148:1–36
Enkerli J, Felix G, Boller T (1999) The enzyme activity of fungal
xylanase is not necessary for its elicitor activity. Plant Physiol
121:391–397
Furman-Matarasso N, Cohen E, Du Q, Chejanovsky N, Hanania U,
Avni A (1999) A point mutation in the ethylene-inducing
xylanase elicitor inhibits the b-1-4-endoxylanase activity but not
the elicitation activity. Plant Physiol 121:345–351
Giesbert S, Lepping H-B, Tenberge KB, Tudzynski P (1998) The
xylanolytic system of Claviceps purpurea: cytological evidence
for secretion of xylanases in infected rye tissue and molecular
characterization of two xylanase genes. Phytopathology 88(10):
1020–1030
Gomez-Miranda B, Guettero C, Leal JA (1984) Effect of culture age
an cell wall polysaccharides of Penicillium allahabadence. Exp
Mycol 8:298–303
Gomez-Miranda B, Prieto A, Leal JA (1990) Chemical composition
and characterization of a galactomannoglucan from Gliocladiumviride. FEMS Microbiol Lett 70:331–336
Hatsch D, Phalip V, Petkovski E, Jeltsch J-M (2006) Fusariumgraminearum on plant cell wall: no fewer than 30 xylanase genes
transcribed. Biochem Biophys Res Commun 345:959–966
664 Arch Microbiol (2012) 194:653–665
123
Page 13
Hope CFA, Burns RG (1987) Activity, origins and location of
cellulase in silt loam soil. Biol Fert Soils 5:164–170
Isaak S, Gokhale AV (1982) Autolysis: a tool for protoplast
production from Aspergillus nidulans. Trans Br Mycol Soc 78:
389–394
Jaroszuk-Sciseł J, Kurek E, Winiarczyk K, Baturo A, Łukanowski A
(2008) Colonization of root tissues and protection against
fusarium wilt of rye (Secale cereale) by nonpathogenic rhizo-
sphere strains of Fusarium culmorum. Biol Control 45:297–307
Jaroszuk-Sciseł J, Kurek E, Rodzik B, Winiarczyk K (2009)
Interactions between rye (Secale cereale) root border cells
(RBCs) and pathogenic and nonpathogenic rhizosphere strains of
Fusarium culmorum. Mycol Res 113:1053–1061
Jaroszuk-Sciseł J, Kurek E, Słomka A, Janczarek M, Rodzik B (2011)
Activities of cell wall degrading enzymes in autolyzing cultures
of three Fusarium culmorum isolates: growth promoting, dele-
terious and pathogenic to rye (Secale cereale). Mycologia
103(5):929–945
Kang Z, Buchenauer H (2000) Ultrastructural and cytochemical
studies on cellulose, xylan and pectin degradation in wheat
spikes infected by Fusarium culmorum. J Phytopathol 148:
263–275
Keon JPR, Byrde RJW, Cooper RM (1987) Some aspects of fungal
enzymes that degrade plant cell walls. In: Pegg GF, Ayres PG
(eds) Fungal infection of plants. University Press, Cambridge,
pp 133–157
Kurek E, Jaroszuk J (1997) Changes in the number of Fusariumpropagules introduced to soil. Polish J Soil Sci 30:63–69
Labavitch TM, Ray PM (1978) Structure of hemicellulosic polysac-
charides of Avena sativa coleoptile cell walls. Phytochemistry
17:933–937
Lahoz R, Reyes F, Beltra R, Garcia-Tapia C (1976) Lytic enzymes in
the autolysis of filamentous fungi. Mycopathologia 60:45–49
Lalaoui F, Halama P, Dumortier V, Paul B (2000) Cell wall-
degrading enzymes produced in vitro by isolates of Phaeosp-haeria nodorum differing in aggressiveness. Plant Pathol
49:727–733
Leal JA, Gomez-Miranda B, Prieto A, Bernabe M (1992) Chemical
and structural similarities in wall polysaccharides of some
Penicillium, Eupenicillium and Aspergillus. FEMS Microbiol
Lett 90:165–168
Lehtinen U (1993) Plant cell wall degrading enzymes of Septorianodorum. Physiol Mol Plant Pathol 43:121–134
Lynch PT, Collin HA, Isaac S (1985) Use of autolytic enzyme for
isolation of protoplasts from Fusarium tricinctum hyphae. Trans
Br Mycol Soc 84:473–478
Martin JP (1950) Use of acid rose Bengal and streptomycin in the
plate methods for estimating soil fungi. Soil Sci 38:215–220
Montesano M, Brader G, Palva ET (2003) Pathogen derived elicitors:
searching for receptors in plants. Mol Plant Pathol 4:73–79
Nelson N (1944) A photometric adaptation of the Somogyi method
for the determination of glucose. J Biol Chem 153:375–380
Perez-Leblic MI, Reyes F, Martinez MJ, Lahoz R (1982) Cell wall
degradation in the autolysis of filamentous fungi. Mycopatho-
logia 80:147–155
Phalip V, Goubet F, Carapito R, Jeltsch J-M (2009) Plant cell wall
degradation with a powerful Fusarium graminearum enzymatic
arsenal. J Microbiol Biotechnol 19:573–581
Polizeli MLTM, Rizzatti ACS, Monti R, Terenzi HF, Jorge JA,
Amorim DS (2005) Xylanases from fungi: properties and
industrial applications. Appl Microbiol Biotechnol 67:577–591
Reyes F, Byrde RJW (1973) Partial purification and properties of a
p-N-acetylglucosaminidase from the fungus Sclerotinia fructi-gena. Biochem J 131:381–388
Reyes F, Lahoz R, Cornago P (1977) Autolysis of Neurospora crassain different culture conditions and release of b-N-acetylgluco-
saminidase and chitinase. Trans Br Mycol Soc 68:357–361
Rodriguez-Kabana R, Godoy G, Morgan-Jones G, Shelby RA (1983)
The determination of soil chitinase activity; conditions for assay
and ecological studies. Plant Soil 75:95–106
Rossner H (1991) Bestimmung der Chitinase-Aktivitat. In: Schinner
F, Ohlinger R, Kandeler E (eds) Bodenbiologische Arbeitsme-
thoden. Springer, Berlin, pp 66–70
Ruiz-Herrera J (1991) Biosynthesis of b-glucans in fungi. Antonie
Van Leeuwenhoek 60(2):73–81
Santamaria F, Nuero OM, Alfonso C, Prieto A, Leal JA, Reyes F
(1995) Cell wall degradation of Fusarium oxysporum f. sp.
lycopersici race 2 by lytic enzymes from different Fusariumspecies for its biocontrol. Lett Apll Microbiol 20:385–390
Schwarz PB, Jones BL, Steffenson BJ (2002) Enzymes associated
with Fusarium infection of barley. J Am Soc Brew Chem
60(3):130–134
Shibuya N, Minami E (2001) Oligosaccharide signalling for defence
responses in plant. Physiol Mol Plant Pathol 59:223–233
Skujins JJ, Potgieter HJ, Alexander M (1965) Dissolution of fungal
cell walls by a streptomycete chitinase and b-(1–3) glucanase.
Arch Biochem Biophys 111:358–364
Somogyi M (1945) A new reagent for determination of sugars. J Biol
Chem 160:61–68
Wanjiru WM, Zhensheng K, Buchenauer H (2002) Importance of cell
wall degrading enzymes produced by Fusarium graminearumduring infection of wheat heads. Eur J Plant Pathol 108:803–810
Wolska-Mitaszko B, Jaroszuk-Sciseł J, Pszeniczna K (2007) Isoforms
of trehalase and invertase of Fusarium oxysporum. Mycol Res
111:456–465
Wu SC, Kauffmann S, Darvill AG, Albersheim P (1995) Purification,
cloning and characterization of two xylanases from Magnapor-the grisea, the rice blast fungus. Mol Plant-Microbe Interact
8:506–514
Wu SC, Halley JE, Luttig C, Fernekes LM, Gutierrez-Sanchez G,
Darvill AG, Albersheim P (2006) Identification of an endobeta-
1,4-D-xylanase from Magnaporthe grisea by knockout analysis,
purification, and heterologous expression. Appl Environ Micro-
biol 72:986–993
Arch Microbiol (2012) 194:653–665 665
123