Brain Research Buktin, Vol. 28, pp. 107-l 13. 0 Pergamon press plc, 1991. Printed in the U.S.A 0361-9230/92 $5.00 + .oO Hypothalamic GABAergic Mechanism Involved in Respiratory Response to Hypercapnia CLAUDIA A. PEANO, CASEY A. SHONIS, GLENN H. DILLON AND TONY G. WALDROP’ Department of Physiology and Biophysics, College of Medicine, University of Illinois, Urbana, IL 61801 Received 3 1 January 1991 PEANO, C. A., C. A. SHOMS, G. H. DILLON AND T. G. WALDROP. ~ypot~lamic GABAergic mechanism involved in ~espi~u~o~ response ?o ~ype~c~pnju. BRAIN RES BULL 28(l) 107-l 13, 1992.-Previous studies have demons~a~ that supra- pontine areas of the brain modulate the respiratory responses to hypoxia and hy~~apnia. The purpose of the present study was to determine if neurons in the posterior hypothalamus are responsible for this modulation. The respiratory (monitored from diaphrag- matic activity) and cardiovascular responses to hypoxia and to hypercapnia were examined in anesthetized rats before and after microinjection of a GABA synthesis inhibitor (3-mercaptopropionic acid, 3-MP) into the posterior hypothalamus. Unilateral micro- injection of 3-MP into the posterior hypothalamus elicited an increase in minute diaphragmatic activity ( + 54.9 f 15.8%), arterial pressure (10.5 z!z 3.2 mmHg) and heart rate (26.8 2 10.7 min-‘) after a delay of 15-20 minutes. The respiratory responses to hypercapnia but not the cardiovascular responses were greatly accentuated after hypothalamic microinjections of 3-mercaptopropi- onic acid. In contrast, none of the responses (increases in diaphragmatic activity and heart rate; fall in arterial pressure) elicited by hypoxia were altered after microinjections of the GABA synthesis inhibitor into the posterior hy~~alamus. These findings indi- cate that a GABAergic inhibition of posterior hy~~~a~c neurons modulates the respiratory response to hypercapnia. Arterial pressure Diaphragmatic activity GABA Hypothalamus Rat Respiration THE posterior hypothalamus exerts a strong influence upon the respiratory and cardiovascular systems. Electrical stimulation of the posterior hypothalamus elicits increases in arterial pressure, heart rate and respiratory output (7, 11, 12, 24). Recent studies have indicated that these responses result from activation of cell bodies located in the posterior hy~th~~us (1, 7, 25, 27). The posterior hypothalamus is also known to modulate sev- eral cardiovascular and respiratory reflexes. Numerous studies have shown that activation of posterior hypothalamic neurons blunts the heart rate response to baroreceptor stimulation (10,20). A previous study from this laboratory demonstrated that this ef- fect involves a GABAergic mechanism (1). Other reflex re- sponses are also modulated by hy~~~a~c structures. Both Redgate (18) and Rom~iuk et al. (19) have reported that the posterior h~~~~us modulates the He~ng-Breuer reflex. Several studies have suggested that suprapontine structures are involved in the respiratory responses elicited by hypoxia and hypercapnia (2, 8, 16, 21). However, most of these studies did not locate the actual neuroanatomical structure(s) which modu- late these respiratory responses. A recent study from this labora- tory demonstrated that the posterior hypothalamus may be a site that exerts an effect upon the respiratory response to hypercap- nia in anesthetized cats (23). Mic~inj~tion of GABA antago- nists into the posterior hy~~~~us accentuated the phrenic nerve responses to ventilating the cats with incremental increases in PCO,. However, this study did not determine if the respira- tory response to hypoxia was also altered. The purpose of the present study was to examine the effects of manipulating GABAergic activity in the posterior hypotbala- mus upon the respiratory response to hypoxia and to hypercap- nia in the anesthetized rat. The findings of this study support and expand the results of the prior study which indicated a role of this h~~~~ic region in m~ulating the response to hy- percapnia (23). However, a GABAergic mechanism in this re- gion does not appear to exert an effect upon the respiratory response to hypoxia in the anesthetized rat. METHOD Twenty-one male rats (14 Wistar-Kyoto and 7 Sprague-Daw- ley) weighing between 190 and 286 grams (2342 16.8 g) were studied. All rats were. anesthetized with an in~~~tone~ injec- tion of u-chloralose (65 mgkg) and urethane (800 mg/kg). Ad- ditional anesthetic was given if indicated by a withdrawal reflex to pinching a hindpaw. A catheter was placed into the trachea and all rats breathed spontaneously throughout the experiment through a tube placed over the tracheal cannula; the rats breathed a hyperoxic gas (PO,>300 torr) during control conditions. Catheters were placed into an external jugular vein and a common carotid artery for ad~nis~ation of editions anesthetic and me~~ement of arte- rial pressure, respectively. Heart rate was derived from the arte- rial pressure signal with a biotechometer (Gould). Body temperature was measured with a rectal probe and maintained at 36.538°C with a heating pad and a radiant heat lamp. Arterial blood sam- ples (0.25 ml) were taken during each experimental condition in ‘Requests for reprints should be addressed to Tony G. Waldrop, Ph.D., Department of Physiology and Biophysics, University of Illinois, Burrill Hall, 407 South Goodwin Avenue, Urbana, IL 61801. 524 107
Transcript
Brain Research Buktin, Vol. 28, pp. 107-l 13. 0 Pergamon press plc, 1991. Printed in the U.S.A 0361-9230/92 $5.00 + .oO
Hypothalamic GABAergic Mechanism Involved in Respiratory Response to Hypercapnia
CLAUDIA A. PEANO, CASEY A. SHONIS, GLENN H. DILLON AND TONY G. WALDROP’
Department of Physiology and Biophysics, College of Medicine, University of Illinois, Urbana, IL 61801
Received 3 1 January 1991
PEANO, C. A., C. A. SHOMS, G. H. DILLON AND T. G. WALDROP. ~ypot~lamic GABAergic mechanism involved in ~espi~u~o~ response ?o ~ype~c~pnju. BRAIN RES BULL 28(l) 107-l 13, 1992.-Previous studies have demons~a~ that supra- pontine areas of the brain modulate the respiratory responses to hypoxia and hy~~apnia. The purpose of the present study was to determine if neurons in the posterior hypothalamus are responsible for this modulation. The respiratory (monitored from diaphrag- matic activity) and cardiovascular responses to hypoxia and to hypercapnia were examined in anesthetized rats before and after microinjection of a GABA synthesis inhibitor (3-mercaptopropionic acid, 3-MP) into the posterior hypothalamus. Unilateral micro- injection of 3-MP into the posterior hypothalamus elicited an increase in minute diaphragmatic activity ( + 54.9 f 15.8%), arterial pressure (10.5 z!z 3.2 mmHg) and heart rate (26.8 2 10.7 min-‘) after a delay of 15-20 minutes. The respiratory responses to hypercapnia but not the cardiovascular responses were greatly accentuated after hypothalamic microinjections of 3-mercaptopropi- onic acid. In contrast, none of the responses (increases in diaphragmatic activity and heart rate; fall in arterial pressure) elicited by hypoxia were altered after microinjections of the GABA synthesis inhibitor into the posterior hy~~alamus. These findings indi- cate that a GABAergic inhibition of posterior hy~~~a~c neurons modulates the respiratory response to hypercapnia.
Arterial pressure Diaphragmatic activity GABA Hypothalamus Rat Respiration
THE posterior hypothalamus exerts a strong influence upon the respiratory and cardiovascular systems. Electrical stimulation of the posterior hypothalamus elicits increases in arterial pressure, heart rate and respiratory output (7, 11, 12, 24). Recent studies have indicated that these responses result from activation of cell bodies located in the posterior hy~th~~us (1, 7, 25, 27).
The posterior hypothalamus is also known to modulate sev- eral cardiovascular and respiratory reflexes. Numerous studies have shown that activation of posterior hypothalamic neurons blunts the heart rate response to baroreceptor stimulation (10,20). A previous study from this laboratory demonstrated that this ef- fect involves a GABAergic mechanism (1). Other reflex re- sponses are also modulated by hy~~~a~c structures. Both Redgate (18) and Rom~iuk et al. (19) have reported that the posterior h~~~~us modulates the He~ng-Breuer reflex.
Several studies have suggested that suprapontine structures are involved in the respiratory responses elicited by hypoxia and hypercapnia (2, 8, 16, 21). However, most of these studies did not locate the actual neuroanatomical structure(s) which modu- late these respiratory responses. A recent study from this labora- tory demonstrated that the posterior hypothalamus may be a site that exerts an effect upon the respiratory response to hypercap- nia in anesthetized cats (23). Mic~inj~tion of GABA antago- nists into the posterior hy~~~~us accentuated the phrenic nerve responses to ventilating the cats with incremental increases in PCO,. However, this study did not determine if the respira- tory response to hypoxia was also altered.
The purpose of the present study was to examine the effects
of manipulating GABAergic activity in the posterior hypotbala- mus upon the respiratory response to hypoxia and to hypercap- nia in the anesthetized rat. The findings of this study support and expand the results of the prior study which indicated a role of this h~~~~ic region in m~ulating the response to hy- percapnia (23). However, a GABAergic mechanism in this re- gion does not appear to exert an effect upon the respiratory response to hypoxia in the anesthetized rat.
METHOD
Twenty-one male rats (14 Wistar-Kyoto and 7 Sprague-Daw- ley) weighing between 190 and 286 grams (2342 16.8 g) were studied. All rats were. anesthetized with an in~~~tone~ injec- tion of u-chloralose (65 mgkg) and urethane (800 mg/kg). Ad- ditional anesthetic was given if indicated by a withdrawal reflex to pinching a hindpaw.
A catheter was placed into the trachea and all rats breathed spontaneously throughout the experiment through a tube placed over the tracheal cannula; the rats breathed a hyperoxic gas (PO,>300 torr) during control conditions. Catheters were placed into an external jugular vein and a common carotid artery for ad~nis~ation of editions anesthetic and me~~ement of arte- rial pressure, respectively. Heart rate was derived from the arte- rial pressure signal with a biotechometer (Gould). Body temperature was measured with a rectal probe and maintained at 36.538°C with a heating pad and a radiant heat lamp. Arterial blood sam- ples (0.25 ml) were taken during each experimental condition in
‘Requests for reprints should be addressed to Tony G. Waldrop, Ph.D., Department of Physiology and Biophysics, University of Illinois, Burrill Hall, 407 South Goodwin Avenue, Urbana, IL 61801.
524
107
PEANO, SHONIS, DILLON AND WALDROP
RESPIRATORY loo r FREQUENCY
37 r , 1 PCO, I
,
(torr)
0 -.- L I
Ik------
ARTERIAL 120
90
RESPIRATORY loo FREQUENCY
(mit?‘) 50
INTEGRATED DIAPHRAGMATIC
EMG ACTIVIN
37
PC02
(ton)
0 i
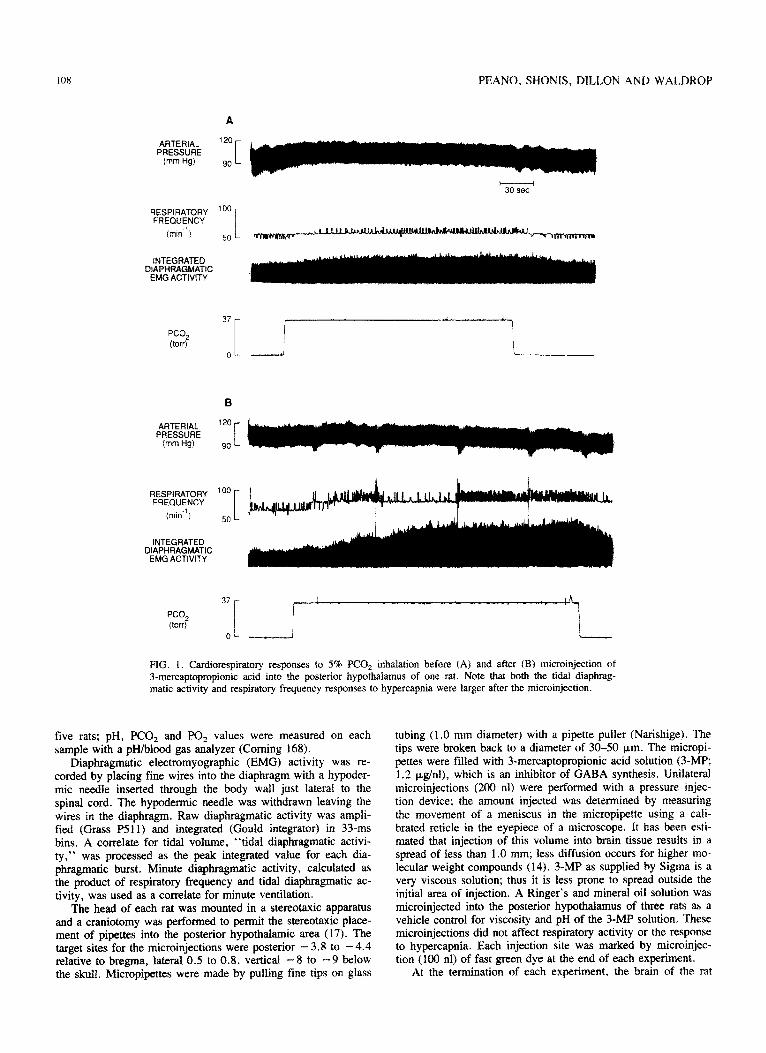
FIG. 1. Cardiorespiratory responses to 5% PCO, inhalation before (A) and after (B) microinjection of 3-mercaptopropionic acid into the posterior hypothalamus of one rat. Note that both the tidal diaphrag- matic activity and respiratory frequency responses to hypercapnia were larger after the microinjection.
five rats; pH, PCO, and PO, values were measured on each sample with a p&I/blood gas analyzer (Coming 168).
Diaphragmatic electromyographic (EMG) activity was re- corded by placing fine wires into the diaphragm with a hypoder- mic needle inserted through the body wall just lateral to the spinal cord. The hypodermic needle was withdrawn leaving the wires in the diaphragm. Raw diap~agmati~ activity was ampli- fied (Grass P511) and integrated (Gould in~grator) in 33-ms bins. A correlate for tidal volume, “tidal diaphragmatic activi- ty,” was processed as the peak integrated value for each dia- phragmatic burst, Minute diaphragmatic activity, calculated as the product of respiratory frequency and tidal diaphragmatic ac- tivity, was used as a correlate for minute ventilation.
The head of each rat was mounted in a stereotaxic apparatus and a craniotomy was performed to permit the stereotaxic place- ment of pipettes into the posterior hypothalamic area (17). The target sites for the microinjections were posterior - 3.8 to - 4.4 relative to bregma, lateral 0.5 to 0.8, vertical -8 to -9 below the skull. Micropipettes were made by pulling fine tips on glass
tubing (1 .O mm diameter) with a pipette puller (Narishige). The tips were broken back to a diameter of 30-50 pm. The micropi- pettes were filled with 3-mercaptopropionic acid solution (3-MP, 1.2 pg/nl), which is an inhibitor of GABA synthesis. Unilateral microinjections (ZOO nl) were performed with a pressure injec- tion device; the amount injected was determined by measuring the movement of a meniscus in the micropi~tte using a cali- brated reticle in the eyepiece of a microscope. It has been esti- mated that injection of this volume into brain tissue results in a spread of less than 1 .O mm; less diffusion occurs for higher mo- lecular weight compounds (14). 3-MP as supplied by Sigma is a very viscous solution; thus it is less prone to spread outside the initial area of injection. A Ringer’s and mineral oil solution was microinjected into the posterior hypothalamus of three rats as a vehicle control for viscosity and pH of the 3-MP solution. These microinjections did not affect respiratory activity or the response to hypercapnia. Each injection site was marked by microinjec- tion (100 nl) of fast green dye at the end of each experiment.
At the termination of each experiment, the brain of the rat
HYPERCAPNIA AND THE HYPOTHALAMUS 109
A. RESPIRATORY RESPONSE TO HYPERCAPNIA
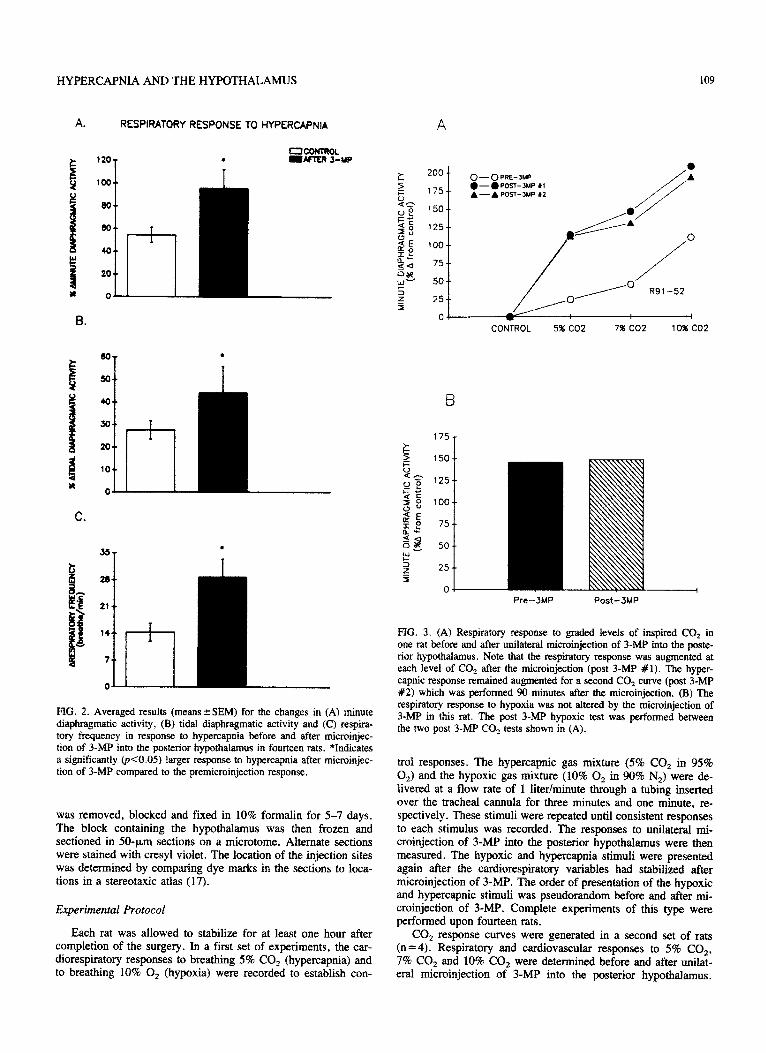
FIG. 2. Averaged results (meanskSEM) for the changes in (A) minute diaphragm&c activity, (B) tidal diaphragmatic activity and (C) respira- tory frequency in response to hypercapnia before and after microinjec- tion of 3-MP into the posterior hy~~~~us in fourteen rats. *Indicates a significantly QKO.05) larger response to hypercapnia after microinjec- tion of 3-MP compared to the premicroinjection response.
was removed, blocked and fixed in 10% formalin for 5-7 days. The block containing the hypothalamus was then frozen and sectioned in 50-pm sections on a microtome. Alternate sections were stained with cresyl violet. The location of the injection sites was determined by comparing dye marks in the sections to loca- tions in a stereotaxic atlas (17).
Experimental Protocol
Each rat was allowed to stabilize for at least one hour after completion of the surgery. In a fist set of experiments, the car- diorespiratory responses to breathing 5% CO, (hypercapnia) and to breathing 10% 0, (hypoxia) were recorded to establish con-
O-OPRE-JMP .-@POsT-3MP 11
A- A POsl-SAP I?.
ot I
CONTROL 5% co2 7% co2 10% co2
FIG. 3. (A) Respiratory response to graded levels of inspired CO, in one rat before and after uniiateml microinjection of 3-MP into the poste- rior hypothalamus. Note that the respiratory response was augmented at each level of CO, after the microinjection (post 3-MP #l). The hyper- capnic response remained augmented for a second CO1 curve (post 3-MP #2) which was performed 90 minutes after the microinjection. (B) The respiratory response to hypoxia was not altered by the microinjection of 3-Mp in this rat. The post 3-Mp hypoxic test was performed between the two post 34%’ CO, tests shown in (A).
trol responses. The hypercapnic gas mixture (5% CO2 in 95% 0,) and the hypoxic gas mixture (10% 0, in 90% N,) were de- livered at a flow rate of 1 liter/minute through a tubing inserted over the tracheal cannula for three minutes and one minute, re- spectively. These stimuli were repeated until consistent responses to each stimulus was recorded. The responses to unilateral mi- croinjection of 3-MP into the posterior hypothalamus were then measured. The hypoxic and hypercapnia stimuli were presented again after the c~dioresp~ato~ variables had stabilized after microinjection of 3-MP. The order of presentation of the hypoxic and hypercapnic stimuli was pseudorandom before and after mi- croinjection of 3-MP. Complete experiments of this type were performed upon fourteen rats.
CO, response curves were generated in a second set of rats (n =4). Respiratory and cardiovascular responses to 5% CO,, 7% CO, and 10% CO, were determined before and after unilat- eral microinjection of 3-MP into the posterior hypothalamus.
I IO PEANO. SHONIS, DILLON AND WALDROP
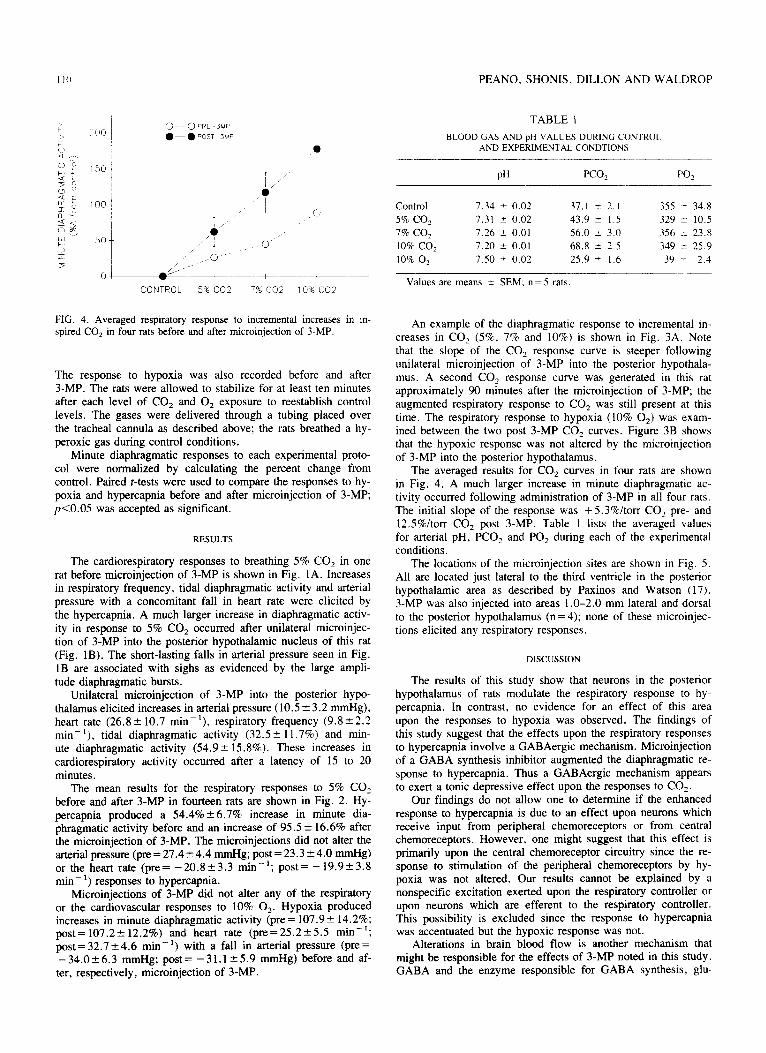
TABLE 1 BLOOD GAS AND pH VALUES DURING CONTROL.
AND EXPERIMENTAL CONDTIONS ._ ___ -___._
” ? t -’
150 T
4 c i ,,’
PH PCO, PO, I 0 * <: Zt I P 1004 ,t I
f i /
Control 7.34 k 0.02 37.1 r 2.1 35s i 34.8
$S I / /’ 5% co, 7.31 2 0.02 43.9 + 1.5 329 I 10.5
\I 0 k IT t :” 50 /' ’ ! 770 CO? 7.26 0.01 56.0 3.0 356 23.X 0’
2 , 10% co2 7.20 2 0.01 68.8 k 2.5 349 ir 25.9
5 ,,,o--- 100/c o2 7.50 f 0.02 25.9 IT 1.6 39 t 2.4 ,’ ,
01 =+, _----+------- -+--- ----,--+ ___ __
CONTROL 5% co2 7% co2 lO%CO? Values are means t SEM; n = 5 rats.
FIG. 4. Averaged respiratory response to incremental increases in in- spired CO, in four rats before and after microinjection of 3-MP.
The response to hypoxia was also recorded before and after 3-MP. The rats were allowed to stabilize for at least ten minutes after each level of CO, and 0, exposure to reestablish control levels. The gases were delivered through a tubing placed over the tracheal cannula as described above; the rats breathed a hy- peroxic gas during control conditions.
Minute diaphragmatic responses to each experimental proto- col were normalized by calculating the percent change from control. Paired t-tests were used to compare the responses to hy- poxia and hypercapnia before and after microinjection of 3-MP; pcO.05 was accepted as significant.
RESULTS
The cardiorespiratory responses to breathing 5% CO, in one rat before microinjection of 3-MP is shown in Fig. 1A. Increases in respiratory frequency, tidal diaphragmatic activity and arterial pressure with a concomitant fall in heart rate were elicited by the hypercapnia. A much larger increase in diaphragmatic activ- ity in response to 5% CO, occurred after unilateral microinjec- tion of 3-MP into the posterior hypothalamic nucleus of this rat (Fig. 1B). The short-lasting falls in arterial pressure seen in Fig. 1B are associated with sighs as evidenced by the large ampli- tude diaphragmatic bursts.
Unilateral microinjection of 3-MP into the posterior hypo- thalamus elicited increases in arterial pressure (10.5 t 3.2 mmHg), heart rate (26.8? 10.7 mm’), respiratory frequency (9.8k2.2 min- ‘), tidal diaphragmatic activity (32.5 k 11.7%) and min- ute diaphragmatic activity (54.9 k 15.8%). These increases in cardiorespiratory activity occurred after a latency of 15 to 20 minutes.
The mean results for the respiratory responses to 5% CO, before and after 3-MP in fourteen rats are shown in Fig. 2. Hy- percapnia produced a 54.4% 26.7% increase in minute dia- phragmatic activity before and an increase of 95.5 f 16.6% after the microinjection of 3-MP. The microinjections did not alter the arterial pressure (pm = 27.4k4.4 mmHg; post = 23.3 k4.0 mmHg) or the heart rate (pre= -20.823.3 min-‘; post= -19.9k3.8 min _ ‘) responses to hypercapnia.
Microinjections of 3-MP did not alter any of the respiratory or the cardiovascular responses to 10% 0,. Hypoxia produced increases in minute diaphragmatic activity (pre = 107.9k 14.2%; post=107.2+-12.2%) and heart rate (pre=25.2?5.5 min-‘; post=32.7*4.6 min-‘) with a fall in arterial pressure (pre= - 34.0 k 6.3 mmHg; post = - 3 1.1 f 5.9 mmHg) before and af- ter, respectively, microinjection of 3-MP.
An example of the diaphragmatic response to incremental in- creases in CO, (5%, 7% and 10%) is shown in Fig. 3A. Note that the slope of the CO, response curve is steeper following unilateral microinjection of 3-MP into the posterior hypothala- mus. A second CO, response curve was generated in this rat approximately 90 minutes after the microinjection of 3-MP; the augmented respiratory response to CO, was still present at this time. The respiratory response to hypoxia (10% 0,) was exam- ined between the two post 3-MP CO, curves. Figure 3B shows that the hypoxic response was not altered by the microinjection of 3-MP into the posterior hypothalamus.
The averaged results for CO, curves in four rats are shown in Fig. 4. A much larger increase in minute diaphragmatic ac- tivity occurred following administration of 3-MP in all four rats. The initial slope of the response was + 5.3%/torr CO, pre- and 12.5%ltorr CO, post 3-MP. Table 1 lists the averaged values for arterial pH, PCO, and PO, during each of the experimental conditions.
The locations of the microinjection sites are shown in Fig. 5. All are located just lateral to the third ventricle in the posterior hypothalamic area as described by Paxinos and Watson (17). 3-MP was also injected into areas 1 .O-2.0 mm lateral and dorsal to the posterior hypothalamus (n=4); none of these microinjec- tions elicited any respiratory responses.
DISCUSSION
The results of this study show that neurons in the posterior hypothalamus of rats modulate the respiratory response to hy- percapnia. In contrast, no evidence for an effect of this area upon the responses to hypoxia was observed. The findings of this study suggest that the effects upon the respiratory responses to hypercapnia involve a GABAergic mechanism. Microinjection of a GABA synthesis inhibitor augmented the diaphragmatic re- sponse to hypercapnia. Thus a GABAergic mechanism appears to exert a tonic depressive effect upon the responses to CO,.
Our findings do not allow one to determine if the enhanced response to hypercapnia is due to an effect upon neurons which receive input from peripheral chemoreceptors or from central chemoreceptors. However, one might suggest that this effect is primarily upon the central chemoreceptor circuitry since the re- sponse to stimulation of the peripheral chemoreceptors by hy- poxia was not altered. Our results cannot be explained by a nonspecific excitation exerted upon the respiratory controller or upon neurons which are efferent to the respiratory controller. This possibility is excluded since the response to hypercapnia was accentuated but the hypoxic response was not.
Alterations in brain blood flow is another mechanism that might be responsible for the effects of 3-MP noted in this study. GABA and the enzyme responsible for GABA synthesis, glu-
FIG. 5. ~~ af ~~roin~cti#n sites which resuited in a~grnent~ respiratory responses to hyper- capia. F&d triaug2es denote sites at which 3-MP accentuated the respiratary response tu 5% CO,; frlied squares are sites at whieb 3-m atoned the responses to graded increments in CCr, (Jk, 7% and 10% CO& ARC= arcuate nucfeus, ~M=dorsorne~~ hy~~~~c nucleus, LV =laterai ventri- cle, MM = mediai mammillary nucleus (medial division), ML = medial mammiliary nucleus (lateral di- vision), MRe = mammi tlary recess of the tbiid ventricle. mt = rn~l~o~~~ic tract, PH = posterior h~~~~c area, SUM = suers llary nucleus, TMC =1 tubeta! ma~n~ellul~ nucleus, Abbrevia- tions and drawings adapted from Paxinos and Watson (17). ‘Rx postzxior location reiative to bregma is denoted on the lower right of Each section.
&mic acid decarboxylase, have been isolated on cerebral blood nism, elicit blo& flow changes in the h~t~~~us or at other vessels (9). In addition, both in vitro and in viva studies have indkated that GABA and GAE%A agonists produce alterations in
brain sites. One might postulate that ~ibit~o~ of GABA syn-
vessel diameter resulting in changes in cerebral blood flow (4, thesis In the present study could have altered blood flow in a
5, 13). Thus i~bi~on d GABA synthesis in the posterior hy- region containing chemoreceptors which are stimulated by hy-
~~~arnus could, through either a humoral or neuml mecha- percapnia or a decrease in PH. These receptors might then be stimuiated by an elevated stimulus secondary to the changes in
112
blood flow. However, the results of the present study do not permit a determination of the actual mechanism responsible for the effects of 3-MP on the respiratory response to hypercapnia.
Unilateral microinjections of 3-MP into the posterior hypo- thalamus altered the respiratory response to hypercapnia but did not affect the hypoxic response. One possibility that must be ad- dressed is that this was due to an anesthetic variability during the hypercapnic stimuli compared to the hypoxia conditions. However, this is very unlikely since the order of presentation of the stimuli was pseudorandom. Moreover, the augmented hyper- capnic responses persisted for over 90 minutes in each rat; the hypoxic response was not altered at any time after the adminis- tration of 3-MP. Thus it appears likely that the altered hyper- capnic response after 3-MP was unrelated to the general anesthetic state of the rat.
Previous studies have suggested suprapontine sites modulate the respiratory response to hypercapnia. Decerebration was shown by several investigators to alter the respiratory response to changes in PCO, (2, 8, 16, 21). In contrast, one study did not note any differences before and after decerebration (15). Even though several of these studies concluded that areas in the suprapontine brain exert a stimulatory effect upon responses evoked by alter- ations in PCO,, none of these reports identified the actual brain nucleus that modulates the respiratory response to hypercapnia. The present results suggest that the posterior h~othaiamus is an involved site for m~ulation of the hypercapnic response in the anesthetized rat.
The findings of the present study support and extend the re- sults of a prior study from this laboratory. It was shown in the earlier study that the phrenic nerve response to incremental in- creases in end-tidal PCO, is accentuated after microinjections of GABA antagonists into the posterior hypothalamus of anesthe- tized cats (23). However, this prior study did not determine if the respiratory response to hypoxia is altered by hypothalamic microinjections of GABA antagonists. The present study indi- cates that such a GABAergic mechanism does not exert an in- fluence upon the hypoxic response.
Several studies have suggested that the posterior hypothala- mus modulates the respiratory responses to peripheral chemore- ceptor stimulation. Cross and Silver (3) found that the discharge of many neurons in the hypo~al~us is altered by hypoxia: similar results were obtained by Thomas and Calaresu (22), who
PEANO. SHONIS. DILLON AND WALDKOF’
recorded the response of hypothalamic neurons to stimulation oi the carotid sinus nerve. Tenney and Ou (21) also suggested that hypothalamic structures exert an effect upon the respiratory re- sponses to hypoxia. The present study did not find evidence for a role of posterior hypothalamic structures in modulating respi- ratory responses to hypoxia. However, it is still possible that other hypothalamic areas or neurotransmitters besides CABA are involved in the respiratory responses to hypoxia.
We may have underestimated the augmentation of the dia- phragmatic response to 5% CO, after 3-MP was microinjected. It has been shown that the respiratory responses to peripheral and central chemoreceptor stimulation are curvilinear. i.e.. iden- tical inputs at higher baseline levels of respiratory activity result in smaller outputs than at lower baseline levels (6). Thus one
would have expected a smaller response to hypercapnia after 3-MP since baseline diaphragmatic activity was increased after the microinjections. Thus the accentuated response may have been even greater than was observed.
The posterior hypothalamus is known to exert a strong intlu- ence upon the cardiorespiratory systems. Electrical stimulation of this area increases both cardiovascular activity and ventilation (7. Il. 13, 24). A similar effect is elicited by microinjection of GABA antagonists into the posterior hypothalamus of both rats and cats (1. 7, 25, 27). In addition, several cardiovascular and respiratory reflexes are modulated by the posterior hypothala- mus. The baroreceptor reflex, the He~ng-Breuer reflex and the reflex cardiorespiratory responses to muscular contraction are all believed to be modulated by neurons in the posterior hypothala- mus (I. 10, 18-20, 26). The present study suggests that this hy- pothalamic region also plays a role in the reflex respiratory responses to increase5 in CO,.
In summary. blockade of GABA synthesis in the posterior hypothalamus augments the respiratory response to hypercapnia but does not alter the response to hypoxia. Therefore, this hypo- thalamic area may be one location that modulates the respiratory response to CO,.
This research was supported by NIH grant HL38726 and the Illinois Affiliate of the American Heart Association. T.G.W. is an Established Investigator of the American Heart Association. G.H.D. and C.A.S. are supported by predoctoral fellowships from AHA-Illinois Affiliate
REFERENCES
Bauer, R. M.; Vela, M. B.; Simon, T.; Waldrop, T. G. Modula- tion of the baroreceptor reflex by posterior hypothalamic neurons in cats and rats. Brain Res. Bull. 20:633-641; 1988. Cohen, M. I. Respiratory periodicity in the paralyzed, vagotomized cat: Hypocapnic polypnea. Am. J. Physiol. 206:845-854; 1964. Cross, B. A.; Silver, I. A. Electrophysiological studies on the hy- pothalamus. Br. Med. Bull. 22:2X-260, 1966. Edvinsson, L.: Larsson, B.; Skarby, T. Effect of the GABA recep- tor agonist muscimol on regional cerebral blood flow in the rat. Brain Res. 185:445-%%8; 1980. Edvinsson, L.; Krause, D. N. Pharmacological characterization of GABA receptors mediating vasodilation of cerebral arteries in vitro. Brain Res. 17389-97; 1979. Eldridge, F. L.; Gill-Kumar, P.; Millhom, D. E. Input-output rela- tionships of central neural circuits involved in respiration in cats. J. Physiol. 311:81-95; 1981. Eldridee. F. L.: Millhorn, D. E.; Kiley, J. P.; Waldrop, T. G. Stim&ion by central command of locomotion, respiration and cir- culation durine exercise. Resuir. Phvsiol. 59:3 13-337; 1985. Fink, B. R.; Katz, R.; Reinbold, H.; Schoolman, A. Suprapontine mechanisms in regulation of respiration. Am. J. Physiol. 202:217- 220; 1962. Hamel, E.; Krause, D. N.; Roberts, E. Specific cerebrovascular lo-
10.
11.
12
13
14.
15.
calization of glutamate decarboxylase activity. Brain Res. 223: 199- 204; 1981. Humphreys, P. W.; J&s, N.; McAllen, R. M. Modification of the reflex response to stimulation of carotid sinus baroreceptors during and following stimulation of the hypothalamic defence area in the cat. J. Physiol. 216:4611182; 1971. Kabat, H. Electrical stimulation of points in the forebrain and mid- brain: The resultant alterations in respiration. J. Camp. Neural. 64: 187-208; 1936. Kabat, H.; Magoun, H. W.; Ranson, S. W. Electrical stimulation of points in the forebrain and midbrain: The resultant aherations in blood pressure. Arch. Neural. Psychol. 34931-955; 1935. Kelly, P. A. T.; Faulkner, A. J.; Burrow, A. P. The effects of the GABA agonist muscimol upon blood flow in different vascular ter- ritories of the rat cortex. J. Cereb. Blood Flow Metab. 9:754-758; 1989. Mvers. R. D. Iniection of solutions into cerebral tissue: Relation beiween volume &d diffusion. Physiol. Behav. 1:171-174: 1966. Mitra. J.: Prabhakar, N. R.; Haxhiu, M.; Chemiack, N. S. Com- parison of the effects of hypemapnia on phrenic and hypoglossal ac- tivity in anesthetized decerebrate and decotticate animals. Brain Res. Bull. 17:181-187: 1986.
16. Nielson, A. M.; Bisgard, G. E.; Mitchell, G. S. Phrenic nerve re-
HYPERCAPNIA AND THE HYPOTHALAMUS
sponses to hypoxia and CO, in decerebrate dogs. Respir. Physiol. 65:267-283; 1986.
17. Paxinos, G.; Watson, C. The rat brain in stereotaxic coordinates. New York Academic Press; 1986.
18. Redgate, E. S. Hypothalamic influence on respiration. Ann. NY Acad. Sci. 109606-618; 1963.
19. Romaniuk, J. R.; Kasicki, S.; Borecka, U. The Breuer-Hering re- flex at rest and during electrically induced locomotion in the decer- ebrate cat. Acta Neurobiol. Exp. (Warsz.) 46:141-151; 1986.
20. Spyer, K. M. Neural organisation and control of the baroreceptor reflex. Rev. Physiol. Biochem. Pharmacol. 88:23-124; 1981.
21. Tenney, S. M.; Ou, L. C. Ventilator-y response of decorticate and decerebrate cats to hypoxia and CO,. Respir. Physiol. 29:81-92; 1977.
22. Thomas, M. R.; Calaresu, F. R. Responses of single units in the medial hypothalamus to electrical stimulation of the carotid sinus
nerve in the cat. Brain Res. 44:49-62; 1972. 23. Waldrop, T. G. Posterior hypothalamic modulation of the respira-
tory response to CO, in cats. Pflugers Arch. 418:7-13; 1991. 24. Waldrop, T. G.; Henderson, M. C.; Iwamoto, G. A.; Mitchell, J.
H. Regional blood flow responses to stimulation of the subthalamic locomotor region. Respir. Physiol. 64:93-102; 1986.
25. Waldrop, T. G.; Bauer, R. M.; Iwamoto, G. A. Microinjection of GABA antagonists into the posterior hypothalamus elicits locomotor movements and a cardiorespiratory activation. Brain Res. 444:84- 94; 1988.
26. Waldrop, T. G.; Mullins, D. G.; Henderson, M. C. Effects of hy- pothalamic lesions on the cardiorespiratory responses to muscular contraction. Respir. Physiol. 66:215-224; 1986.
27. Wible, J. H.; Luft, F. C.; DiMicco, J. A. Hypothalamic GABA suppresses sympathetic outflow to the cardiovascular system. Am. J. Physiol. 254:R6804687; 1988.