Page 1
•
\
, . '. " 1
•
"
__ .. - _f~
\;:;';î_ -:-'., ~,_":""~_::'::""'.",:t,/'~::';~:",: i, ',u _. _~~'; ..--
l '
THE COMPETENCE OF LYMPHOID ORGANS DURING ThE COURSE OF
EXPERIMENTAL TRICHINELLOSIS
by
Ory8ia M. Ulczak ...'
A ~h.~i ... ub~t.d to the Pacult~ of Graduat.
Studies and Research in partial f,ul filment of the requirements for the deqree of
Master o( Science
In8ti tute of Parasi toloqy - Montreal
January 1979
'~I ___ ._o_r_Y_S'f_a_M_~·_Ul czak __ 1_97_9 _____ J,
l '
, "
Page 2
", "
()
l'
'''.
;.10
d ,
~_'
\ \. " ~i~ ~ tr~
I~
" "1
G)
.... /
i ~
"SUI)SOA ted short, ti tle 1
\ IMMtINOCOMPl::Tr;NCE OF 'LYMPHOID
IN TRICHI~r;LLOSIS
,
.. "
r
",-l
" / 1
/
•
ORGANS
, ,
,
\ '
': ..
..... _._--...... ~~"
~. ' ' .. ~
' __ ':" __ oi~; .. l _____ .... _' ~1_f;""':1.:_":_iÙ~~1~?~:/0;\r(,,,,f,~{(;;-;,
----
c;
/
Page 3
~!I;'
--'~
/
C)
" ... , ~,'
\
ABSTRACT
Low and moder~t •. infections with the nematode'" Trichinell~ . ,
,.pirali. i)l4ue. a luppre •• ion ol the immunologieal m' •• pons. of '
-:' '~~~ ~~o.;' • The e Uec;:t ,of th" infe~~ion on' the' inteqA ty o~ t~" • lymphoiè1 system was studied by adoptive transfer int~ \ '
inununo~o9~~all~-dep;i ved anima1s and ~y. !oU vi tro celi~ eU'l ~ur~ .. Thymui cella from model:"ately-infeeted A/J mie" are
to re.tore immunologieal campete'nee to deprivèd syngen 10 " ,
reciplents than normal oells. Bone marrow éèlla from oder
. infected mice, on the other hand, enhr:'neil the oapaqi ty \ of
, ~ecipient animals to relpond to lipop~ly,a~charide <LP$). , • 1
,capaci ty of Bepri vad recipient mice to ;espond to shae '
erythrocytea ls eventua~ly restorea after the tranafer! of
spleen cells from, infected animals, it seamac
, tharefore, that > 1
, • immunoaupprassion ls- a 'tranalent phenomenon. Trichine1la
" .. ,,,
) .
spiralis may s'urvive in,the infected animal by a dose-dependef\t· , ~
, , '1
Alteration of the eapacit,{ ~f the l-ymehoid system to reapo~d to
immunoloq ica1 stimuli. 0 1
,~ -, .
1.
l ,1 /
"-,
:
" <:1
~) ,'1
~-~- -
;il , '
l' .... , ,
tt <,
- -
\
Page 4
c. , ;r
,
"- ~ ~ ,
1 1
, ,~
\,"i .~~J
li ,
ri~ r .\ J: ~' 'l
,. I~
, l .. il;\ ~ l " ," ,} ~, J, \\ <
1 9, , 1
'-
" '
1
\J
r'
).
'1. . '.
"
ABREGE
\ • infections modls'r'el et baises avec le n~atode
,
~==~l;.,;;l~a spiralis induisent une suppression de la rlponse ~
L'effet de l' infeotton. sur l ',intêqri t' "
.y.tA el lymphoide a êtê ~tudHle par transfer adoptif c:hez . ----
et par cultute cellulaln
souris A/J infect.e.
sont moins aptes 1 restorer la comp'tence immuno
rêciplents syngênes dêpourv':ll, que lesont le,s
rmalea, Pa,r contre, les cellules de ,la moelle d'os
BOUriS modlrêment infectAes augmentent la capaci tG
ts a r'pondre au lipopolysaccharide (LPS) , Chez
à'pourvus, la r'action immunologique aux qlobules _ ...
'roug B de m uton 8e normal.iie 'ventuellement, a la Buite d'un
traJsfer de cel{~l~'S splênioques provenant à' animaux infectll. J o ~
il semble one que l' immUnQ8Uppreuion Boit un phAnomlne . \.
transitoir\, De plus, l '.importance de l'effet d'une infection
par!. s.,piralis Bur la comp'tence immunologique est lffSe a la dose inocul'e et pourrait àonc influencer la survie du
\/ parasite. 1
"
'.
Page 5
",
, 1 .' , '
'l' 1
" , .!
• : ,.' , ,r'~ 'r', ':~~ ,~}1,'"' 't 'l"~ ...... # . ~ . . , , ,
*)",4_"_"'~""~"~~~~~u .. ).,..,' ." ... , .... " ......... ' ...... ~ ~ .-
.'
» l, 1\ ,l
PQRPOSE '1'1' L'
\ ~ ~
. The purpose of thi. ~tudyfl;was to investigate the'
effect of infections with Trichin'ella spiralis on' the ~immuno-
competence' of the primary and secondary lymphoid organs of'
mica, to determine where an inun,unOl'OgiC~l defect could be , '
located which leads to immunosuppression. In addition, it was ~ \ .
of interest to determine whethet: inununosuppression is a
function 0(, and determined by, the presence of the parasite,
or whether it is due to a prOlonged.effect because of a damage
by the parasite of the lymphoid org~ns of the hoat.. The effects ./ J'
of different inoculating doses on the phenomenon of immuno-
aUPP'resliion and the response at different stages of the l ''';
infection were a180 subjects of this investigation. Ultimately,
'it was hoped to obta'in some inaight into the mechanism whieh
indueea ~mmunosuppression in"trichinellosis.
1
iv
'"
Page 6
1
.. • ~>'
l' . ,)
'-'
c
. \
"
,
ACKNOWLEDGEMENTS
r <:
) l would like 'to express my appreciation and thanks to:
Dr. C. E. Tapner, my supervisor, for his advice,
encouragement and willinqness te he'lp i,n. every possible way.
throughout' the course of this study, l '
Dr. Esmail Ghadirian, for hie invaluable assistance in
many of the adoptive transfer stl,1dies, and especially for his
.!, encouragement and uAdetstandinq,
, Dr. de Voe, ,Chairman of Microbiology and Immunology, "
\MCGill university, for the use of the gamma r-adiation facilities,
Dr. J.E. Tanner, Department of Microbiology, Macdonald
College, for the donation of E. coli E145 lipopolysaccharide, #'t - --
Dr. Hai,-Choo Smith and Mrs. Klara Gyapay for their ve~y
helpful and expert technical assistance, 11'\ Mr. Michael Sukhdeo for his grea t help in the photographing
and printing of graphs, ""'\
Misses Carole St. Charles and Louise Mfner for their
help in translating the abstract, ,
" all the professors and students at the Institute for their
interest and constructive cri·ticisms of my werk,
Dr. Croll for his geneul he~p,
Mrs. Hall and _Mary La Duke for their able administrative
assistance,
Mr. G. Bingham for his patience and good care of the
animals,
Page 7
o.
~ .... •
" 7.
f· ... '.
~: '\
r, a ~' ~ ~ ~ l, l< ~,
~ \ ~l
i. L,
1: f' t , " ~ 1
, .'~
'1
" \\' t ~
k " <",
" ,
t , ~
J' >,' .. Î'
~l (-
~ ,~ ~
r , " ~r~ ( ç ,
t .' " f li
0,
'.t ", j
.... 'and finalry, a very special thanka to Rohani bta Ahmad
for her great moral aupport and for bein.9' such a dear friand.
The finaneial asaiatanQe of the Natural'Scienc •• and
.En9in.e~in9 Council, Grant-A 4954 ia a180 appreclated.
.. (
)
'Ii
"
. vi
\ \ \
l ' , \
\ \
, ' .~
, ~,
. , . • , ",
>~
J
•
fi
,
('
'~, -.----- .. -.. ---.-~~ _________________________ .... ! .. s ....... ÎIiIIIII"' __ l1li
Page 8
Ct
f.
)
o
/
LIST OF CONTENTS
Paqe
Title page ••••.•••••••••• , ••••• • ~ .................... . i . Ab.traot ...... ~.. . . . • . . . • . . . . . • . . . . . . . . . . . . . . . . . . . . . . 11
1
Abrege. . . . . . . . . . . . . . . . . . . . . . . . . < ...... , .' •••••••••••••• iii
Purpoa8 .•......•...••..•....••. ,Il •• -.-. - •• , ••••••••••••• iv J Acknowledqementa. • • • • . • • • • • • • • • . • • • • • • . • • • • . • • • • • • • • v
Liat of Con tenta ••••••••• " •••• ". • • • • • • • • • • • • • • • • • • • • • vii ,
Liat of FiqureB ••••• ~.. •••••••••••••••••• ••••••••••• x
Liat of Tables....................................... xii
Liat of Abbreviationl •••••••••.••••••.•••••••.••...• xiii t?
LITERATURE REVIEW •••••••••••• J •••••••••••••••••••••• l
1. Morphology and Life Cycle of Trichinella
apira11s ................... , .................... . l !
2. Evasion of the immune re.pon.e by para.ites ••••• 3
A. Immuno8upp~e.8ion and Immunopo~entiation
in Trichinella .piralia inf.ction •••••••• : •• , 4 1
B. Machaniama of Immunoauppr ••• ion ••••••••••••• 13
3. Imm.un i ty ............................. ' .......... . 17
A. Acquirad Immunity ta Trichinella spiralis ••• 17
B. The Role of Humoral Factora in the Immunity "\ .., '
to Trichinella .eiralia •••••••.••••••••••••• 20
C. The Ro1e of Cellular Factora in.the Immunity
ta Trichinell •• pirali...................... 25 f _
D. Mecha~i.m of Acquired Immunity •••••••••••••• 29
MATlRtALS AND MB'l'HODS. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • ' 34
A. Para.ite •••••••• 0:' •••••••••••••••••••••••••• 34
vii
Page 9
o
.-
o
B • H08 t. . . . . . ....... ~ . . . . .. ' . , .... · ............ · · .
C ~ Infeotive M~cle Larvee.! •••••.••••.••••••••
D. counting,ôf Infective Larv •• and Inf.étion
l4
34
of Mic.e ••• , ,'" ••••••••.••.•.••••...••••••••.. "1.... 35 1 ,.... ' - " E. Thymectomy , ••••••••••• .! •••••• " ••••••• , ••••••
F. l~radiation •••• \ ••••. \ .............. • • . · • • • .•. ~
G. Collection and Transfer of Thymus ,and Spleen ..
lS
36
,Cella .................. , ••••• "_LI~ ••• ~ •.••• , • l,' 36 , 1
H. Collection and Transfer of Bone
Calla .........•......• ' ........ .
I. An ti gens and I~un'i za tion. '.' • • • •• • ~ J • • :-. • • • 1
J. Plaque-forminq Cell A •• ay •••••••••••••••• : •• - - \
, 1 Marbrook Çell Culture Chambers •••••••••••••• K.
l7 ·
l8
38
39
L. Sta tiltical Analysis ••••••••• .' •.• -. • • • • • • • • • • 40
• RESULTe ••••••• ~. • • • • • • • • • • • • • • • • • • • • • • • • • .. • • • • • • • • • • • 42
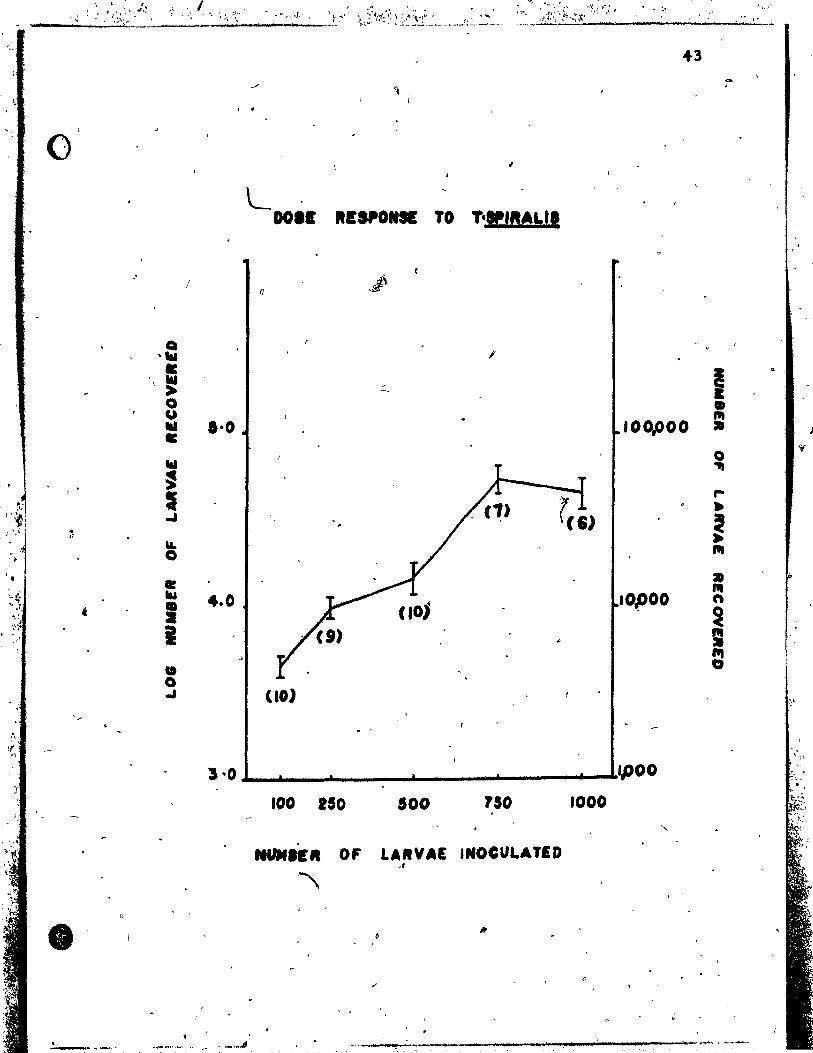
'Exe,riment 11 The level of in,fection in A/J mica
fOllowing inoculation with different d08es of
infective larvae •••••••••••• ,;. ••••.• ••••••••••• 42
Experiment 2: The abil~ty of A!J mice inoculated ( -
vith 100 or 500 larva. to r •• pond to immunization
with aheep erythrocytea. The t-ime courae of thil o
re.pon ............. .,e •••••••• ( ............. • • •• , • • • •
ExP.riment~: Reconstitution of immunologicelly· ,... ,
dagcived recipient. vith normal Iynqeneic thymu~
and bon. marrow cella ••••••••••••••••••• < ~ . . . . . . 41 "
IImri_nt ! 1 The l_unocœpet.nce of the thyaui
of infected A/J .1c •••• ~ ••••• t •••••••• ~ •••••••••
viii
, .
Page 10
!
"
o
~
, \ i
Exper iman t ! 1 '. J "" The immunooompetence of bone marrow
oella from, infected A/J mic ••••••••••••••••• ' ••••• ) (
51
The reconstitution of immunologically • 1
depriVled recipients- by spleen oells trom ,nonna'l . .... ''tif •
donora .......... 1 • ••••••••••••••• ". • • • • • • • • • • • • • •• .. S4 > o;} ,
Experiment 1& The irnmunooompetence- of the spleen of • 1 .. ù, ..,
infect~d A/J mice ••••••••••• ,. •••••• j. •••••••• ' ••• 1. 56 q
}I-
AI 'l'he assay of competence 6 daya
after cell transfe!!'-••••••••••
B. The assay of co~petence 30 daya ~
after cell transfer ••••••••.•
C, The effect of thymectomy •••••
O. The effect of large numbera of
adopti va oella ••• ",.: ••••••• ,.
E. The a~say of competence in o -
Marbrook ohambers in vi tro ••• '-
56
ù58
60
62
64
DISCUSSION ......................... t ................... t • 66
t ~ BIBLIOGRA.PHY ••••••••••••••••••••••••••••••••••.•••••. 86
..,. '" .. <:
, . -, "-
, ..
1
, 1 1
,"
j
\
Page 11
"
1 1
P'
~ -
, i
,'-
l,'r l
.. , "
LIST OF FIGURES
Figure i: The mean log ~umber of muscle larvae
• recoveré~ from ~/J mice JO days after , l ,
inoculation with diffe~ent doses of'infec-' .
r
, ,.
,Page
l ti ve !. spiralis > larvae ••••. ti ••••••••••• feQ ••••• ' ••• 43
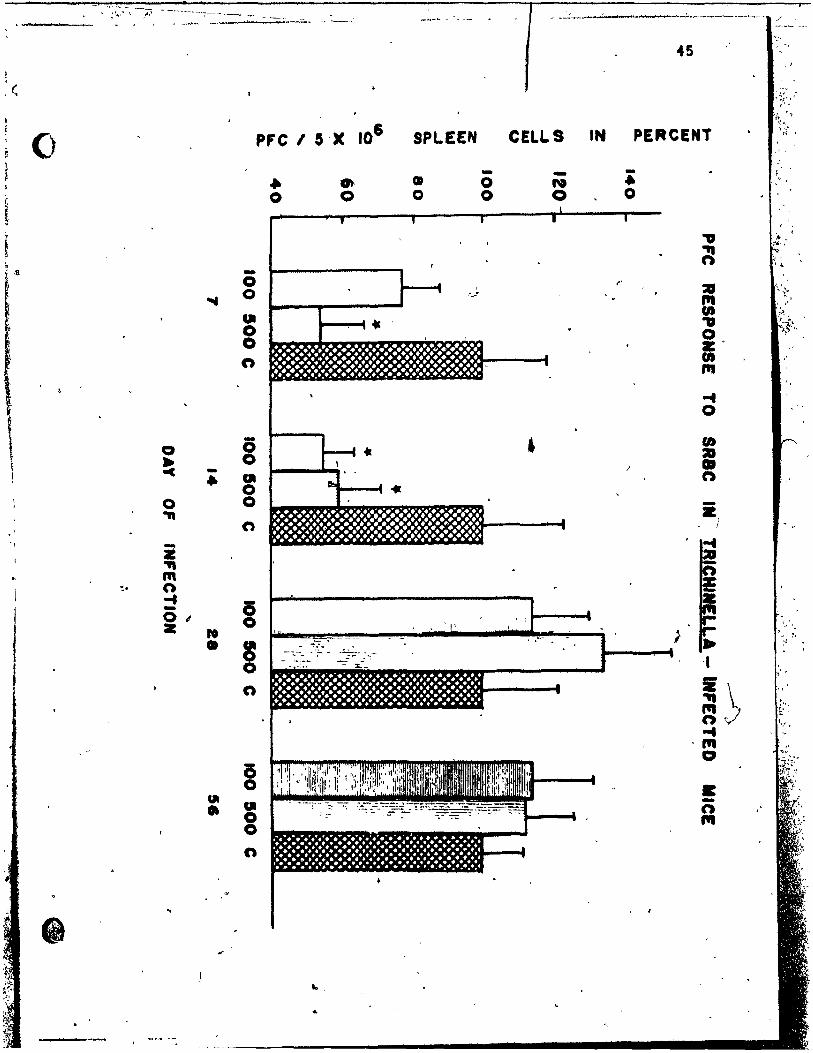
Figure ~: -~~C respon
r ,different ti
SRaC of A/J mice at
"" inocu~~t~o~ with 100 ~~
~.!.!:.~~-.&J:~r-ac .................. ~ ••• ' ••• 4·5
Figure 1: The'PFC response ta SRaC of eprived recipi
ents re~onstituted with normal,bone marrow.
cell~, and thymus cells from either,normal
, 1
• Figu[e !:
donors or from donors previously inoculat~d , .
with 500' larvae ••• , ............................... ~
TQe PFC response te LPS of depriv~d recipi-)
ents reconstituted with normal bone marrOW
cells'or bone marrow cells from dOnors Q \ • 1
previously inoculated wf,th 100 or 500 larvae ..... 52
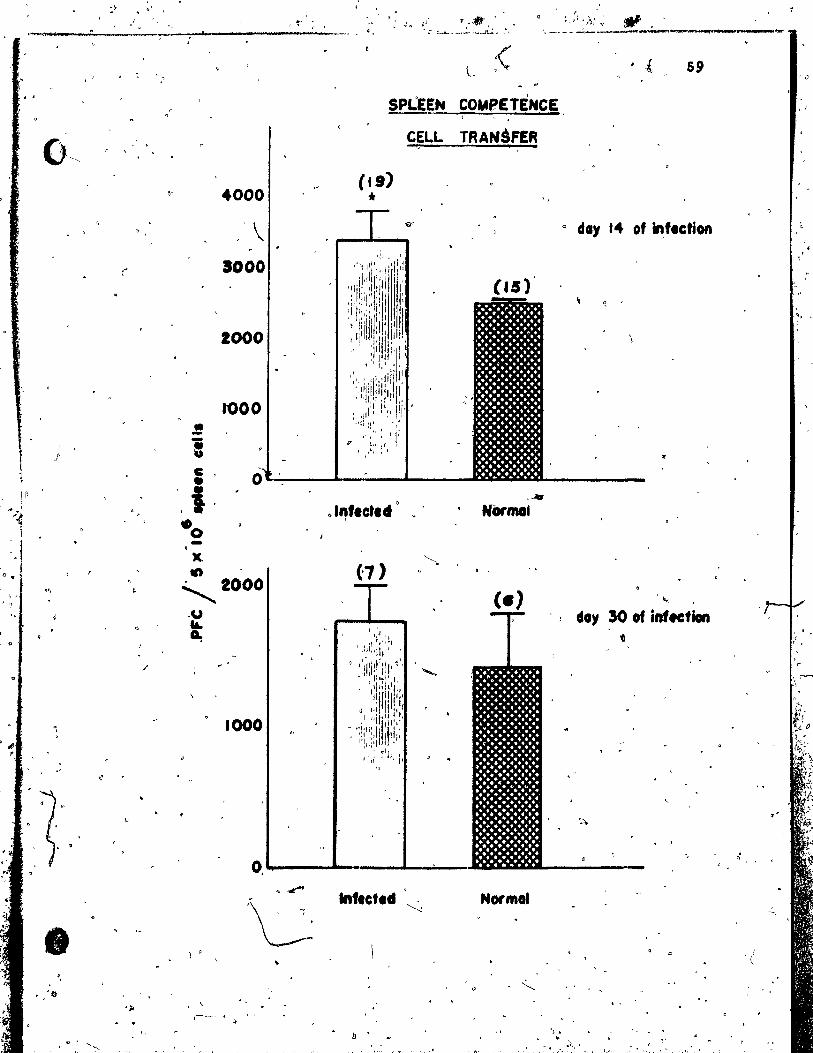
Figure 1: Reconst~tution of deprived recipients by,
normal spleen cells ••••••••••••••• ' •••. ' ••• ',' •• '.: • ss
Figure!: the PFe response to S~C of deprived recipi
ents 6 days after aaoptive transfer of normal > 1.
;> spleen cells or sple~n cells from donors
previously inoculated with 100 or 500 larvae •••• 57 l ,.,. ..
Figure 7: The PFC response ta SRBCC5'f recipients JO 1,
day. after ce'll transfer of normal spleen
x " ; ,J'.
L'
.' "
Page 12
o
....
_" .~L.&4 L ____ mLt &_ t~
cells or spleen cells from donors previously ) inoculated wi th 500 larvae ••••••• " ••••••••••••••• 59
1 \. "
Figure 8: The PFC response to SReC in deprived (thymec~ - ~
tomized) recipients reconstituted with norma~
spleen cells or spleen cells tram don~rs , .;j
previously inoculated with 100 larvae.: ••••••••• 61
F.igure 9: The PFC resjt~nse ta SRBC of depri ved recipients
Figure !.Q.:
/' .
reconstituted wit~ làrqe' number of normal
spleen cells or spleen cells from mice inoculated
wi th 500 larvae •• ~ ••••••••••••••••••••••••••••• 63 "
The PFC responsé !!l vitro of normal sp-Ieen (
cells or splee~ cells from inf~cted micè.: ••••• 65
, .1
J "
. ". - .'
Page 13
o
.-.,
" ; , '.'
~' ':;1(
f; )" " ~~ .,' " , ,~ ..
:"J ,,1
,~ l"~f""
;:,
j! ,-
a n
''''~1 -
'. , ~1 1':. '.) ( ''J ,1 'l'
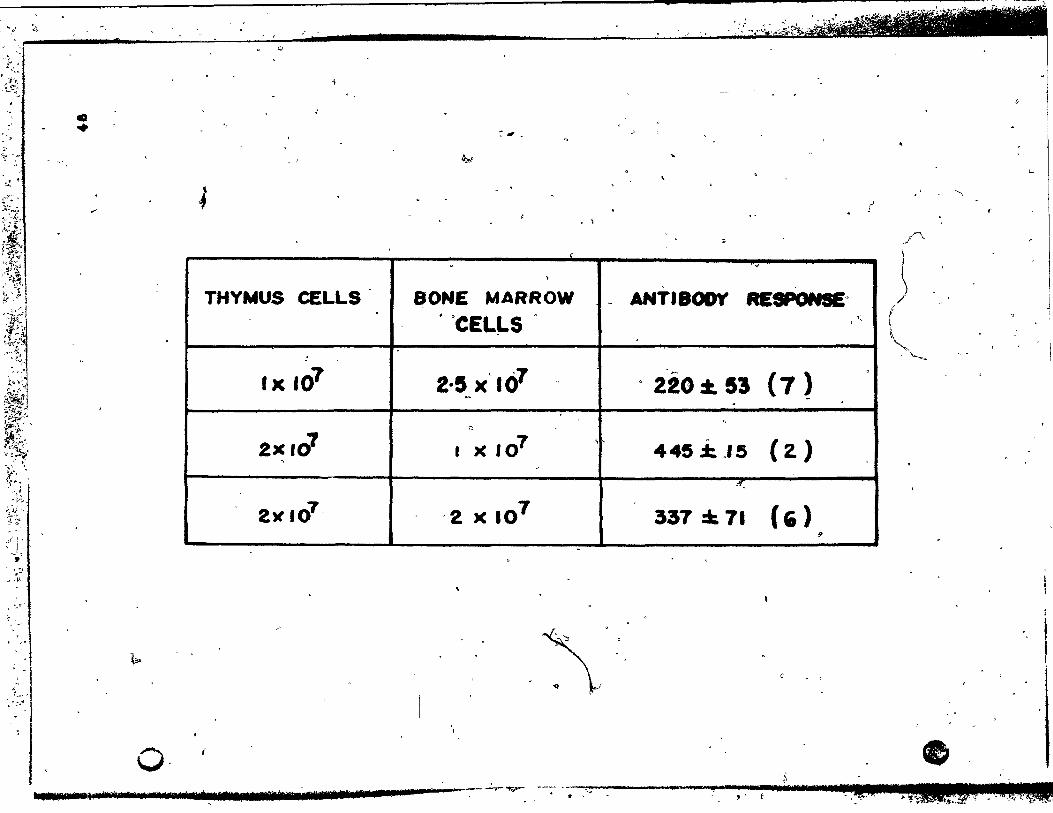
Table 1:
"
/
LIST OF TABLES
The PFC response of deprived recipients~.'
reconstituted wlth different combinations
of normal thymus cells and bone marrow
Page
cells ........... . 1 • ••••• " • ,. ••••••••••••••••••••• 48
"
"
xil , ,
,(.Jo.' ~ •.
, ~'
,
, ,,. ....
Page 14
_. __ . --~.~ ".
o BCG
Con A
GVH
19
JBE
LPS
PFC
PRA
PVP
1 SRBC , 1
1
1 j
1
o
LIST OF ABBREVIAT10NS
Bacille Calmette-Gu'rin
Concanavalin A
Graft veraus Hos~
Immunoqlobulin
Japanesese B Encephalitis
Lipopolysaccharide
Plaque-forming Cell
phytohaemaq,glutinin
polyvinylpyrrolidone
Sheep Red Blood Cells " <
..
~'" •
xiii
; .1"
J
r r
Page 15
()
\ '
,. ',à_"
(
1
LITERATURE REVIEW ,
1
1. Morphology and Life Cycle.of Trichinella spiralis
Thé parasite used rn these studies,
(OWen, 1835) billiet, 1895, is' unique among' e parasitic
nematodes in that all the stages of its life cycle occur within
ft single hast (Gould, 1970). The discovery of this parasite ls
attributed ta James Paget, à medical student who described this
worm in 1&35 in the muscles of a human cadaver.:- Sév~ral)weeks
later, Richard Owen,' haviag been informed ôf Paget's findings,.
found aimi~ar e~idence and named the paras'ite Trichina spiralis.
Prior usa~e of the~enus resulted in a change in the generic
name of this nematode to Trichinella spiralis (Steele and
Arambulo, 1975).
According to the classification of the Nemâtoda by
chitwood (1974), !,'.spiralis ,ia assigned to the superfarnily
Trichuroidea, Railliet, ~895, in the class Aphasmidia; this
nematode possesses no, 1ateral pits (believed to be sense organs)
at the caudal extremity. Trichinella spiralis has a very long,
fine eaophagus embedded, for most of its length, in a column of -'
glandular cells called stichocytes. ln the female, -the vulva
.opena n~ar the middle of the esophageal region, the anterior
part of the uterus 18 crowded with erobryos, whereas the posterior
part contains developing e9g8. The female 18 ovoviV~parou8"gi~ing
birth to live y~ung. The mal •• are eharaeterized'by th. ,
pre •• nce of a pair ot conieal appendage. at the ~osterior end;
Page 16
"
/
. ~. , ._""_ .. -....-,. ... ~ .. 1
2
and possess no sp~cu1e (Chandler and Read,1 1961).
This parasite has adapted to a very broad range of
vertebrate reservoir hosts: swine, cats, dogs, and many wild , ' .
animaIs, including fox, wolf, bear, polar bear and marine 1
mammals and rats (Be~~nsorr, 1975): the occurre~ce of trichi
neIIos~s is'wo~ldwide but variable in preval~ (SteeIe/and
Arambulo, 1975). Trichinella spiralis is maintained and •
transmitted in nature by two kinds of cycles: the synanthropic
cycle which leads to human infections, particularly that
between pigs and pigs and, the natural, or sylvatic', cycle which
is maintained by wi1d carnivores. In the latter cycle,
transmiss~ is regarded simply as incidentai to natural
interactions and the intraspecific behavior of various wild
animaIs.
The life cycle of !.spiralis begins upon the ingestion of
the ;aw, or insuffiéiently cotled, ~flesh of animals containing
viable encysted larvae. ~pon reaching the stomach, the larvee
are released from their cysts in the striated muscle tissue by
the enzymatic action of the gastric juices. Molting, growth and
maturation to apults takes place in the small intestine very
soon after th~ parasites enter the gut (intestinal phase).
Foilowing copulation, the gravid females deposit the new-born
larvae directly in the ·tissu~ of the small intestine. The
new-born larvae enter the lymph or blood v~8sels in the
intestinal wall and are thU8 disaeminated throughout the body {l-ifr'
(migratinq phas.). After pa •• ing into the mUlculature with
the blood or lymph, each larva~ quickly penetra tel a Itriated ) ,i
Page 17
( f ' .. \
o
.'
" ,
"
lit... ~.~ - - ~ .... ~ , -" ..,... ~ ....... -~ 0 ~ --..,- ~...- ~J .... ~- ,~
3
muscle cel1, grows rap~ly and becomes sexua11y diffefentiated
(ear1y sedentary phase), the muscle larvae t~eh roll , •
themse1ves into a spiral and the enclosing muscle cells
contrib'ute to the formation of characteristic 1emon-shaped cyats , ,
whose long aXé~ are paralle1 with muscle fibres, these larvae
become infective about 17-18 days after ~hey have entered the
muscle fibere. After ency~tation, no further deve10pment ~akes il\ place until the infected flesh is eaten by a susceptible animal
(late sedentary phase) (Chandler and Read, 1961, Gou1d, 1970,
Muller, 1975: Madsen, 1976) ~ ,
2. Evasion of the Immune Response ,by Parasites
,
Despite th~ battery of responses that parasite infections
can elicit, parasites survive in their natura1 hosts, often for
considerable periode of time: it is obvious, therefore, that j
there must be weIl develope~ mechanisms by whiéh the parasite
can evade the harmfu1 consequences of the hostls immune response ,
to the invasion. A variety of mechanisms has . pe'en proposeà
which' cou~d enable parasites to survive immunological rejection
by the host: antigenic variation in the protozoa, the acquisition,
of host-like antigens by schistoB'omes, the pro'duction of , 4'
anti-complementary factors by cestodes, encystment 'and the intra-
cellular habitat of certain parasites, and the formation of
Ogi1vie and Wi~son, 1976). Immunosuppression ia 9't another of
the mechani.ma, "Y which a parasite may .~. the effects of the
immune re.pons. of the hOlt,' thi. phenomenon h •• b •• n reported
Page 18
\ \ . \
4
-to oceur in experirnental protozoal infections (Clinton !! al,
1969, Greenwood !! a1~ 19711 Sengers et al, 1971: Mansfield and -- q ,
Wallace, 1974, Hudson ~ al, 1976: 8rown et al, 1977, Reed et
al.,1977J Strickland and Saylea, 1971), as weIl as in helminth
infections (Shimp et al, 1975: Cranda11 and Crandall, 1976;
Araujo et !l, 1977).
)
A. Immunosuppression and Immunopotentiation in !.spira1is
Infections
In laboratory animals, the immunosuppression caused by
experimental tr1ehinel10sis has been demonetrated numerous
times. Kilharn and Oliver (196i) reported that rats inoculated
with Trichinella spiralis in doses below those resulting in
" apparent illn,esB, became tRore susceptible than did normal rats
to cripplinq and death induced by encephalomyocarditis (EMC) 1
virus. Similarly, thè prornoting ~ffect of trichinellosis on the
disease of the central nervous system (CNS) caused by Japanese
B encephalitis (JBE) virus has aiso been demonstrated (Wa1ton
and Gleiser, 1966; Cypess ~ al, 1973; Lubiniecki !!!l, 1974a).
The unpubli~hed work of Walton and Gleiser (1966) has also shown
increased mortality and decreased survival time in mice infected
with Trichinella spiralis f~11owin9 per~pheral challenge with \
.eastern equine encephalitis (EE!) virus and I1heus virus, in
comparison to non-parasitized mice. In addition, trichinelloais '\
a1so increasès the 8u8cePtib~rnice and rabbits to vaocinia
virus (Chimishkyan and Ovumyan, 1975). The mechaniama respon
lible for the enhancinq affect of !.spirali~ ~n virQ8 infedtion
- . )
Page 19
i i
1 j. .
.~ .. ""~""-~ ... _~... ~_ 1 ,
(J
----0
" , . t ., -''"-~-' 'r~''" ,- -
5
q7 ... have not yet been elucidated: however, there is sorne speculation
that th~ facilitation of virus establishment~i. due to factors
other than immunosuppression by the parasite. Mice challenged
with JBE virus at various intervals after inoculation with
!.spiralis showedl a depressed antibody response to the virus,
when the virus was given on or before day 28 of the parasite
,infection (Cypess et al, 1973; Lubiniecki et al, 1974a,
Lubiniecki and Cypess, 1975a). ' However, increasea Busceptibi1ity
to virus infection in these parasitized mice was demonstra'ted
~ntil the fourteenth dày fo110wing !.spira1is ~no~ulation but
susceptibility to the virus was comp1ete1y abrogated by the
twenty-first day (Cypess et al, 1973; Lubiniecki et al, 1974a).
Moreover, several workers indicate that he1minths, particular1y
migrating he1minthic larvae, can, through non-specifie tra~atic 1
effects, either transmit or aggrava te infection with micro
organisms (Shope, 1943, Mochizuki et !!, 1954, Wescott and Todd,
1966, Woodruff, 1968). E~~ virus has been demonstrated in hig~ ~
titres in sk'ele'tal muscles that contained migràting and . / ..,..;:'
developing !.spira1is larvae, while no virus was found in
corresponding musc1eaV'taken from control rats infected with the
virus alone <filham and Oliver, 1961). In addition, JBE virus.
replicated quickly to high titres in the brains of mice infected
with !.spir,a1is, in comparison to mice given JBE virus a10ne -(Lubiniecki !1 .!!, 1974a).
\
Cases have recently been presented of human patients with
oral Iquamous ca11 carcinoma and Trichine11a'.Piralil larvae in
o
J
Page 20
1 , t
i
,,-,1If01 lit. 4 ___ "" .. '-\.I4ol.. .... c:~
o
'" l
'i!I'!r'I'.', "1"-....
" .. '".
the a.sooiated soft tissues. These oases may only reprèaent
6
, , "
an interesting phenomenon of two dissimilar disease processes ,
oCGurring simultaneous1y: however, the occurrence ls_ 80 extreme1y
rare that a potent!al interrelat!on8hip should be considered
(Bruée, 1975).
Experimental trichinellosis in laboratory animals has", also . been shown ta be aocompanied by functional changes in the humoraI
system. Studiea on the synergistic interaction between JBE . virus and Trichinella spiralis have shown that mice infected
with !.spiralis manifested 10wer titred primary and,secondary
complement-fixing and neutralizing antibody responses to JBE
virus, (Cypess ~ al, 1913:- Lubiniecki 'et al, 1974a; Lubiniecki
and Cypess, 1915a), non-lethally irradiated !.spira1is had no
effect 'on the subsequent' antibody production to JB! virus .'
(CypeBs!!!l,1973). !.spira1is has also been shown to exert
an immunoBuppressive effect in mice on the production of haemag
g1utinating,antibody to vaccinia virus (Chimishkyan and Ovumyan,
1915) •
VariouB workers have shawn a marked reduction in the res-
ponsiveness of mice infected with !.spira1is to another common-ly
used unre1ated antigen, heter01ogous erythrocytes. - Immunosup
pre.sion of the primary haemag91utinating antibody response to
aheep red blood cells (SRec) has been demonstrated in the serum
of infeéted mice (Faubert and Tanner, 1971; Barriga, 1915;
Lublnlecki and cype.a, 1915a; Chimishkyan ~!!' 1976, Jon.s
~!l. 197', Ljunvstr6m and'Hu1dt. 1977)1 lmMunoBuppr ••• lon
J1
Page 21
~ " ,.
in tr~chine1losi~~as ,
by the plaque-forming
7
also be~n demonstrat~d at the B ce11 ~ev • .J:... cel1 (PFe) Assay (~ubiniecki and Cy~ss,
1975a, Chimishkyan ~ al, 1976, Faube~t, 1976, Jone~ !!!l, 1976,
Ljungstr6m and H~1dt, 1977) and by the rosette-forming ce11
• (RrC) assay, {Faubert and Tanner, 1974aL__ lnununosu2Eression
appears to aUect both IgG (Lubiniecki and Cypess, 1915a,
Ljungstr6m and Huldt, 1977) and IgM ~laque-forminq cells
(Lubi~iecki and Cypess, 1975af Faubert, 1976: Jones!!!l, 1976,
Ljungstr6m and Huldt, 1977), a's weIl as serum antibody of the
IgG (Lubiniecki and Cypess, 1975a; Jones et al, 1916) and IgM
classes . (Jones et al, 1976, LjungstrlSm and Huldt, 1917). " ~. -- : v
Some experiments have, however, failed to demonstrate Any
significant reduction in IgM serum antibody response to foreign
erythrocytes as -'a result of Trichinella spiralis infections , ~
(Lubiniecki!1 al, 1974b, Lubiniecki and Cypess, 1975a).
!.spiralis has been shown ta have little effect on (and may
actually increase) the antibady response tO,thymus-independent
antigens such as polyviny1pyrrolidone (PVP) (Barriga, 1975:
LjungstrOm and Huldt, 1977) and ~he Vi antigen of Salmonella
typhi (Chimishkyan et al, 1976).
Recently, Jones et !! (1976) have demonstrated that
cultures of splenocytes from mice infected with !.spiralis
developed fewer antibody-forming -cells th an did control
cultures after subsequent immunization in vitro with SRBC.
The addition of the supernatant fluide of cultures of apleen
cella from infected ~i~e aignificantly auppre ••• d thè re.pons.
Page 22
l \ ;
. '
o
, . ' ,,'
8
!!! vitro of normal sp1enocytes to SRBC (Jones !!.!? al '0 1976). ~
Moreover 1 extracts of !.spiralis 1arvae (Faubert and Tanner,
1974a, Barriga, 1975), as well as the serum from animals
infected with !. spira1is ,(Faubert and Tanner, 1971, 1974a), j
depressed the in vivo humoral response to these erythrocytea " ')
when given intraperitoneally prio~ ta challenge ~h ~R8C.
The experimenta1 infection of mice with !.spiralis
has also been demonstrated to induce a state of relative
unresponsiveness to pas'sive cutaneOUB anaphylaxis (PCA)
reactions ta hen avalbumin (Munaz and 'Cole, 1977). Rather than ,--
being due to an irnrnunosuppressive effect of the parasite,' it
was SU9gested that the unresponsiveness of infected mice to this
antigen was due to an increase in the production of IgE during
infection which competitively b10cked the attachment to the maat
cells of albumin-specific IgE molecules (Munoz and Cole, 1977).
The effect of the parasite on the cellular arm of the,
immune response has also been investigated. Studies on the re
jection of al1ografts, an assay used to measure the efficiency Cl
of effector cells of the cell-mediated immune response in vivo, 1 --
have demonstrated a prolonged survival of allogeneic grafts in
mice infected with !.spiralis (Svet-M01davsky et al, 1969,1970f
Chernyakttovskaya !! al, 1972 ~ Faubert and Tanner, 19751
Chimishkyan !! al, 1916; LjungstrOm, 1916: Lj,unqstrtsm and Huldt f
1977). This delay in the rejection p~ocess was more ~ronbunced
in èhose mice which had larqer numbers of !.82ir&11. larvae in
the muscle (Chernyakhovakaya et al, 1971, 1972). Furth.~re, --
l '
" ,
Page 23
\ f'
l J
1 fI , f .'\. 1
~ fI
t , ~t ~~
." l,
;;: If :\
" t, " "
~ .. t ,\
'î ~ (\, "" •
1 \
(~
Ljungstram and Hu1dt (1977) '-have shown that the delay in the
rejection of a110qrafts was Most pron~unéed in miee 9rafte~
in the first:. week of the infection, a time which eoincidea wi th ,
maximum depletion of the thymus (Ljungstr~, 1976), on the other \.
hand, Chimishkyan ~ al (1976) obtained maximum prolongation of
rejeètion of skin allografts onlY on the twenty- fou~th day of
the inJ:ction. Svet-Moldavsky and colleagues (1970) also demon-
strated a rnarked suppression of the second set reaetion in the ......... ' .. ,"
rejection of skin alloqraftâ in mice infected with !.spiralis.
A significantly de1ayed rejection of grafta has also bet!fn
observed in, mice re-inoculatad with !..apiralis larvae: allografts
were, however, rejected ear1c1,er by the re-inoeulated animals .... ~ .... "
than by mica that bore only il primary infection' (Ljungstram and
Huldt, 1977). Graft rejeetion was also delayed in mi.ce inocula
ted wi th ir6diated larvae and in mice in which the paras! te ..
infection had been terminated at the adult stage after treatment ..- , ~
with the anthelminthic methyridine (Ljungst\:~m and HUldt, 1977). . " \ ,. )
Interestingly, the skin allograftB survived longer in mice whose ,~
fnfection was curtailed by methyridi~e, than in mice inocula ted' 1
with irradiated larvae1 following suoh ino,cula, the parasite t
devélops through the intestinal stage but produces no progeny to
infect the muscles. However, alloqrafts in methyridine-treated
infècted mice were rejected earlier than those in m!ce with a
normàl infèction (LjunqstrlSm and Huldt, 1977). Studies by
Faubert and Tanner (1975) have indicated -that treatment of normal
mica by an intraperi toneal injection of th. serum of infect.cS
animals was all~ effective in .uppr ••• ing tran.plant.tion i .. unlty •
, --
, 0
Page 24
"
.'
,"
10 J
UsJng another !!!. ~ ass~y td st~dy the effect of
T:8piralis inf~ction on cy~otoxic T-lymphocytes, ChLmishkyan
!j: !l r (1976)'· have demonstrated a sharply supprJsséd capacity of ~ ~ -
spleen cells from inlected mice to induce -Il- graft-versus-host f .. ,
(GVH) reaction. Cypess et al (l974b'), investigating t'he effect -- , , ~
of !.spiralis infection on the development of cell-mediated,
delayed-type hypersensitivity (DTH) to ~acillus Calmette-GuErin
(BCG), observed ..,that infection wi th T. spiralis produced an .
,., initial suppression of ., ~,
t~e response qp old tùberculin (OT)
,
-"'*' ' ta BCG-treated "con troIs. In vitro , ' \ -
't) challenge, ,in contrast
~/ •
studies by,Chernyakhovskaya ~ !!.! (1972) of cytotoxicity for
target cells have shown that lymph node cells from mice i.nfected ~ :J
with !.spiralis did not destroy murine embryonic fibroblast
ce~ls in tissue culture.
, A recent_study of the effect of the parasite on the proli-
fera,tive capacity of cells using in.vitro.transformation 'tech
niques has shoV(1 tha t the response of spleen cella from infected
animaIs ta lipopolysaccharide (LPS) was essentially normal while ,
the blastoge~ic response to,T-lymphocyte mitogens phytohaemag-~
glutinin. (l'HA) an'd concanavalin A (Con A) was suppressed (Jones 1·
et al, 1976))\ Blasè6genic studies by Tanner, Lim and Faubert
,1978) have suggasted that:. in moderate\ infections iilduced by an
inoculum of 50_0 larvae, the leucocytes l~~he secondary lymphoid--J' , , •
organs (the spleen and lymph nodes) are'insensitive to a stimula-1
ting action of Con A and LPS, whereas an enhanced response ta
these mi togens could be sean in lover infections induced by an
?
Page 25
1
i
i 1 1 ;
, ,
.......... _------------------- , "
< 1
Il
inocuhun of 100 larvae. However, Lubiniecki and Cypess (19'75a) . failed to demoJ)strate any signif~cânt differencè in the .ability.
of spleen' cells trom infected mice to undergo blast formation-.
a-fter st~mulation by PHA •
. "
Some authors have been unable to,detect immunosuppression ,
in animaIs' infected with Trichinella spiralis cr have found a
concomitant e~~d res~stance to ~on-related microQrganisms
a: d tumor cells., ~Studies by Weatherly (1970) show~d that a
infect'ion wi th, T" spiralis in female mice increaseJ! , v
time, partly due to a markedly reduced incidence > '
mtmma~y tumors in, the infected animaIs. Lubiniecki lA
j' ,
and Cypess (1975b).,. have demonstrated that !.spiralis infection
28 days' 'prior to inoculation" with the ascitic tumor, sarcoma 180,
produced a ~tatistically significant increase in the·length of ,
both the incubation time of the tumor and the survival time of
the infected mi ce • However, no effect of T" spiralis on the
reproduc~ion ~f Na~séher leukemi~ virus has been observè'di the ~-~ ,
virus ti~res ~n the mice infeéted with tne parasite ~erê
identical in "thqse mice without the nematode (Trubcheninova ~ f
al, 1975). In addition, T.spiralis Dinfèction has increased the 1
resistance ta both intravenous and intrâperitoneal i culation .. . .
of Lister+~ monocytogenès (CYP'9ss,!i al, 1974a, , ... c ,,~ i
elfilvate"d the rate of clearance of colloidal from the
blood (Cypess !.!:. al, 1973, 1974c). Other studies have 'shown
\ \ /1 \ :
. ,
that immunoPQtentiation of delayed-typ~ hypersensitivity to BCG
either before (Molinari
"
'.
Page 26
f t
t "
.J J;:
f ;-~ ~
~ ~
1 " t ~ ~ f
1 f
·~) -
~ -____ .--____ 0' ___ ._._, _ •• " ,_." _______ ~~[:'.'~ ·.i?f~·:;~~:}(' ',~ " _ .... ~ .... :....~ ... ~.:_., ' --
1
()
\.
o
12
~ !l, 1914) or after sensitization (Cypess !! al, 1974b)
with viable ,and heat killed bacteria (Molinari and Cypess, 1975).
It has also been shawn that experimental.!.spira1is infectionSJ
can inhibit the parasitemia of concurrent infection with o /
Trypanosoma equiperdum and Trypanosoma lewisi in rats .
(Meerovitch and Ackerman, 1974). ; slgnificant1y lO,nger survival , ,
times of T. equiperdum have been reported in a~mals infected 1
" with Trichi~lla spiralis (Wagner and Nembhard, 1916). Campbell
et !!. (1.976) hav failed to show any slgnlficant reductiQn of ,
the lethal effec T.cruzi in mica by concurrent trichinel-
10sis.
While pr10r infection with T.spiralis did not influence
the numbers of schistosomes deve10ping ~rom a moderately light
inoculum (50 cercariae), they did reduce the numbers which
developed from an inocul~ of 200 cercariae (Jachowski"tëlnd
Bengham, 1961). The natural resistance of mice to Hymenolepis
nana var. fraterna was increased when they were simultaneous1y , '
!noculated with 200-or 400 larvae of T.spiralis, but not when
the dose of larvae waS increased to 800 (Larsh and Campbell, ,
1952)., Rats which had been immunize~ by several ~ncr~asing
inocula of !.spiralis larvae, 'and' subsequentlY challenged with
infective'Strongyloides ratti larvae, harbored fewer ~.ratti
adults at ,necropsy than did unimmunized con troIs (Kazacos, 1976)'.
Increased protection has also been found to Ascaris suum infection o
(Matov and Kamburov, 1968) and to Nippostrongylus brasiliens s
infection (Ka2aco., 1975) in rata infected vith !.8piralis. /
u
. , "
Page 27
'l
i f ,. i
1 f t f·
r f
, , ,
, ,,:,.'"''''*~ ____ , __ ,_._. ___ ~_v__ ~ __ " .'-----_ .... _. -- ,; !. :~,.:.-. ___ -"-~ .. '~\; • ~_: ,:" ~ "~~.i!1~~_,_, ~~ __ ~~~
"
o
,
13
B. Mechanisms of Immunosuppression
Various hypotheses have been proposed to explain ~he
immunosuppression induced by Trichinella spiralis, but the
precise mechan!sm(s) which ia responsible for this phe~omenon
has yet to be elucidated. It has' been suggested by
~vet-Mo1davsky and his co1leagues (1969, 1970) that !.spiralis
may produce inununosuppress! ve substances which depress the
host' st immune mechanism and, thus, permit the survi val ~f the
parasite. Such immunosuppressive substances wou1d probably be
produced most actively at those stages of the life cycle of the 1 '
helminth when development is particularly vigorou$, e.g. during
the mig~ation or the larvae'. Certain lines of evidence support
the hypothesis of the generation of suppressor substances during
infection: treatment of normal animaIs with the serum of syngeneic
infected animaIs inhibitëd the ability of the recipients to form -~ .r
antibodies to sheep erythrocytes (Faub~r't and Tanner, '1971) and
to re'ject skin allografts (Faubert and TanneJ;;, 1974a}. The o
serum of mice infected with !.spiralis has also been shown to be (.
leucoagglutinating and cytotoxic, specifically against lymphoLd
cells (Faubert and Tanner, 1975): Tanner.and Faimert (1974)
demonstrated that !.spiralis larvae contai~ ~ubstances wh~ch are
able to enhance or to supprese muscle invasion. The purified , .
fraction from the larvae wh!ch enhanced infection a1so contained
substances which agglutinated and kille~ lymph node cells;
..... interestingly, very few antigens could be discerned ~n this . . '" particular fraction. On the other hand, treatman,t vith the
fJ "
\
Page 28
f '1
1:
~ Ji
1 ~.
t f
\
.1 1
1 t
,r
. ,
t'
o
14
fraction which contained the majority of the antigens against l' qf'
which antibodies are' formed during infection protected against
the parasite (Tanner and Faubert, 1974)." That the immunosup
pressive substances could be of parasite origin has been "
) supported by Faubert (1976) who has demonstrated that newborn
larvae .ètre capableo of producing soluble substances that
interfered with the PFC response of normal spleen cells. In
addition, a correlation exist.s between the level of trichinel-
10sis and the survival tirne of allografts (Chernyakhovskaya
et al, 1971, 1972). - - "
Alternatively, the suppressive factor could be of host 1
origin sinoe the host cou1d produce specifie autoantibbdies . against immunogens released froM tissue damaged during the
development of the (Faubert and Tanner, 1975). No
studies have been to determine the selectivity of the
r(s) found in the serum of i~fected
animaIs. It COU.uu.,....s:;It:n that this factor (5) seleçtively and
immunosuppressive
/
directly affects effector lymphOcytes, particularly thyrnus-
derived cells, only at a certain stage of their differentiation. .. , . On the other hand, tpis' factor (s) could suppress the irnmuno
logical response indirectly by enhancing suppressor T-cel1
function, or by, àltering macrophage function .
. A second possible rnechanism which could explain ,i.mmunosup-
pression is an alteration or a deteri~r'tion of macrophage -'
activity. Tanner et al (1978) have demonstrated that Trichinella
spiralis infections deplete the numbers of Adherent cella in the
\
Page 29
l,
1 1
()
o
" ,1!
15
spleen, most marked1y on the fourteenth day of infection.
Despite this dep1etion, ,however, the~average r~t~' of the 'clear
ance of carbon partic1es is increased significantly (Cypess ~
al, 1973, 1974c), a1though a decreased humoral re.ponse to JBE~
virus has a1so been shown at this time (Cypess et al, 19:73; ,
Lubiniecki ~ ai, 1974a; Lubiniecki and Cypess, 1975a).
However, the phagocytic index assayed by carbon clearance, tests
on1y the ability of phagocytes to ingest particles but not
their ability to,degrade them intracellu1ar1y, nor,does,it
test for the abi1i ty of these leucocytes to participate in an , ~
irnrnuno1ogica1 reaction. No studies ~ve yet;. been done to
measure the, function of the intracel.1ular mechanisms of macro-
phages from mice infected wi th !.spiralis (Le. bactericida1
activity) •
It had been reported by Faubert and Tanner (1974a) that
bone marrow cells from infected mice were inefficient in
-!!:$ reconstibuting a humoral responsiveness to sheep erythrocytes
\ in mice ~imrnuno109i~al1y deprived by thymectomy and irradiation. ~,
"
, This observation indicates that suppression could a1so be ·due
to a deficiency of -the precursor lymphoid cells which arise
from the bone marrgw. This deficiency may be due to an alt;,er
ation in the function, or to an acbla1 ';eduction in the number ;::.."'JItiD
of stem ce11s. The leucocytotoxic factor described by Faubert
and' Tanner (1975) may a1so play a similar role in influencing
the funcÙon of stem ce11s.
One group of inveatigators has reported that the bnune
Page 30
_~.".w _______ ~ __ .. ~ ___ _
1
1 f
f
, , - '. ... ~_ .. -.. ....:...,..~--...... -_. _~_w __ ._ ........... _ ......... I-.... _H .... ". , ... _._ ... '_""."w __ ~l't~ /
16
" suppression induced by infection with Trichinella spiralis is
consistent with the features which have been described fo,
antigenic competition (Lubiniecki et al, 1974bl Lubiniecki and
Cypeaa, 1975a). Immunosuppression iri trlchinellosis has been
demonstrated to be.route dependent (Lubiniecki ~!!, 1974b),
thymus-dependant cell populations are'implicated in the
integrity of the response of the. host to t.spiralis (Wal1s et !l,
1973; Ruitenbarg and Steerenberg, 1974: Ljungstrôm and 1
~uitenberg, 1976)1 infection with this parasite impairs a number
of T-cell functions (Barriga, 19751 Ljungstrôm and Huldt, 1977).
A study by Jones ~ al (1976) haà suggested that
T-suppressor cella may be responsible for the observed immuno
suppression induced by!.spira1is. These authors'have shown \ . that splenocytes from infected mice actively suppressed the ~
, vitrQ response of normal cells to SRBC, and that this in vitro
suppression was abolished by lysis with anti-thy l antiserum and
enhanc~d ,by lysis with qrti-immunoglObUlin serum (Jones !1 !l, 1976) . ,
The dynamics of the infection in an animal doee not involve
directly only 1ymphoid ce11s but also various other organ systems
as well, each of which has i ts own phys,io.logy ~ ThuB, a parasi te ;. ~ ,,1 -...... ,' - f"
draws on"the nutrient supply of the host and dauses hormonal
changes which can alter the immune status of the animai. The , /
non-immunological aspects of the infected state should also be
considered, not only beinq involved in immunosuppresslon (Hud.on~ ,
1973), but also as an Integral part of the complex mechanism
which produces disea8e.
Page 31
," '1' , ,
- - -.., ~-> ..... ~-----~_ .... ---..... _- • 1
17
<: 3. Immunity
~'-~--~-'~~~ During the early studies on helminths, most irnrnunoloqists
l'
o ,
)
and parasitoloqists eonsidered that the larger animal parasites
would not stimulate any demonstrable acquired immunity which
would inhibit the development of or kill the parasite. In
general, recovery and elimination of infection we~e presumed to , ,
be largely an expression of old age and death of the parasites
(Taliaferro, 1940). Earlier investigators were rather skeptical
of the possibility that infection by Tric~ine11a spiralis might
confer Any imm~nity against reinfection, sinee several cases of
reinfection with !.spiralis had been demonstrated (Romanovitch,
19121 Schwar·tz, 1917, Hall 1t.nd Wigdor, 1918). However, investi
gators began to collect evidence that infection with sorne
'metazoan parasites can prevent superinfection, and that recovery
from infection resul t~ in a greater or lesser acquired immunity .
to subsequent infection (Taliaferro, 1940). Ducas in 1921 was
the first to demonstrate experimentally that A partial immunity
to reinfection with T. spira1is had been estab1ished in rats
which had recovered from a previous infection wi th this parasite.
These findings were later confirmed in rats (McCoy, 1931, 1935;
Bachman and Oliver-Gonzalez, 1936: Fischthal, 1943), in mice
(Culbertson, 1942a), in quinea pigs (Roth, 1939) and, to a lesser extent, in hogs (Bachman anq Rodriquez~M01ina, 1933).
, A. Acquired Immunity to Trichinel1a spiralis
'-In ACt! vely immunized hosta, there may be or lIIay not be an
Immediate 1088 from the intestine of a large proportion of
Page 32
(' " '" f r ~ ,.
t ~,
:< ~ ft 'S-
i li!
! i, \ , • 1
---""._--.. /._~~ ____ ~_~._ __ .. J 'j ...... __ • ...1.,.0.0., .. -_ .., ........ l'il ,~. 1 -::,:' .. • ,
.1.... ~,~ ............ "' .... ~ ~ •• ~"...... ...,.,..._~ .. ~ __ • ~ .~ _
()
o
18
larvae of the c~a11enging dose within a few hours f0110wing
infection. McCoy (1940)--demonstrated that in hyperimmune rats "
considerable' numbers of larvae from the challe!lging dose appeared
in the feces as early as three hours after feeding, ,and that
the majcSrity of larvae, f~d were e1iminated eight to eüjhteen p ,
hours later. Moreover, such larvae wer~'stil1 viable and
infective when fed to non-inunune rats (McCoy, 1940): Rappaport
and Wells (1951) o,n the other hand, fai1ed to demonstrate Any
grea t initiaI 10ss of larvae from the challenging dose ~n
reinfected rats·.
The larvae in the challenge inoculum which do succeed in
estab1ishing themse1ves in the intestine develop within an
immune host to produce adulte which are significantly. stunted,
particularly the femalee (Rappaport and Wells, 1951: Semrad and 1
Coors, 1951: Shikhobalova, 1952; Larsh and Race, 1954).
Furthermore, such females con~ain fewer 1arvae ln their uteri
than those Irom controls qiven only the cha11enging infection
Rappaport and Wells, 1951J Semrad and Coors, 1951: Larsh and
Race, 19~4). Moreover, these adulte are expelled sooner from
the intestine than those in the respective controls (Ducas,
1921; McCoy, 1931, 1935, 1940: Bachman and Oliver-Gonzalez,
1936; Roth, 1939; Cu1bertson, 1942a: Fischtha1, 1943: Rappaport
and Wells, 1951; Wakelin and Ll,oyd, 1976). The inhibitory
effect of the immune state on the female is further reflected
by the significant1y reduced numbers of muscle larvae which 1
" the se parasites produce (McCoy, 1931; Bachman and Rodrigutlz-Molina, ..
Page 33
t
19
1933, Culbertson, 1942a; Fiachthal, 1943; Rappaport and Wells,
1~51; Shikhobalova, 19~2).
A certain time interval following a single immuni~in9
d~e of larvae ia necessary for immunity to develop aqainst
the challenqing infection. Fischthal (1943) demonstrated that
approxima'tely 14 days, but not 7 days, was adequate for rats '"
to develop immunity against a challenginq dose fol10wing a
single immunizing dose (80, 160, 320, or 64,0 1arvae) •
Rappaport and Wells (19~1) sbowed that yields of adu~t worms
were somewhat lower in mice reinocu1ated 35 da ys and 3.5 months '"'1
following the suri! of the, initial infection, but not when the
animals were chal1enged at 10 d~ys.
There is a180 an" indication that immunity wanes gradua11y
with time. McCoy (1931) ahowed that two rats lost their '
immunity to the parasite 14 weeks and 10 months after the
·initial infection. Semrad and Coors (1951)' demonatrated that
effecta of il'lll\un.tty were greatelt in thoae animals which W8re
challenged 20 days fo1lowin9 inoculation, as indicated by
the 9rowth of the adul t famale, an4 the numbers 'of e9gs and
larvae preaent, by the thirtieth or fifty-fourth day, however,
the protected ltate had begun ~"dec1ine. ,4
Shikhobalova (1952)
reported that immunity in mica, measured by the length of <
Trichinella spiralis adults, was more intense at 12 to 30 days
than at 60 day' after initial expo8ure.
Page 34
20
, . The deqree of immunity obtained a1so appears to be
dependent upon the size of the immunizinq and challenging ,
infeètions. Fischthal (1943) d~onstrated that ~mmunity " ,
produced in rats against a challenge infection of 10, 000 larvae
was more complete when a higher (320 or 640. larvae) rather than
a low (80 or 160 larvae) irnmunizing inoculum was used. Rappaport
and Wells (1951) dernonstrated that, following a challenge dose
of 800 larvae, adult worm counts during the course of the
intestinal phase did not show any significant differences ,',
~
between animafs ini tially inoculated wi th 100 larvae and
.;
control animals; when mice were reinoculated w!t::h 300 larvae,
howewr, a significant loss of adult worms was seen beginning
'in the second week of the infection. , B. The Role of Humoral Factors in the Immunity to Tricrhinella
spiralis lJ
Metazoan parasi tes do not have the sarne enormous reproduc
tive capacity as the protozoans; however, because of their
si~e and metaboli'c activity, they êstablish themselves as an ,
important and an abundant source of antigens (Capron ~ al, 1977).
Most of the somatic constituents and physiologica+ endproducts
of the metazoans are potentially imm"i1ogenic in the host and
are, thus, effective in stimulating the host's antibody-forming
mechanisms (Ta11aferro, 1940).
During a ~.spiralrs infection, the gamma globu1in fraction ,
of lerwn has been shawn to 1ncre ••• aignificant1y (Wright and
Page 35
()
.'
21
~Oliver-Gonzalez, 1943; crandall and Moore, 1968) and la
associated with the anti-parasite antibodies (Mauss, 1941).
Changes in the levels of the immunoglobulins during the course 1
t of a !.spira1is infection have been demonstrated by a number
of different'serologieal methods (Bachman, 1929: Kagan and "
Bargai, 19567 Cranda11 and Moore, 1968; Crandall and Crandall,
1972; Ruitenberg and Duyzings, 1972; Ottesen ~ Ali 1975; \
Vernes ~!!, 1975b: LjungstrOm and Ruitenberg, 1976: Perrudet
Bado€: et al, 1976; Matossian et al,' 1977), as' well as by immuno·
histologieal studies of the lymphatic tissue (Crandall et al,
1967: Karmanska an~ Kozar, 1969: Crandall and Crarrdall, 1972;
Ruitenberg and Duyzings, 1972; LjungstrOm and R~itenberg, 1976:
Tanner et al, in preparation). In these\iatter studies, the
immunoqlobulin on the surface of the eel1s has not been identi
fied as specifie anti-Trichine11a spira1is antibodies: in sorne
cases it has been assumed that the increases in immunog1obu1in
leve la ',ia a direct consequence of the para~i te infection , (Ruitenberg and Duyzings, 1972). It appears, however, that the
large increase~ in immunoglobulins (humoral, as weIl as ce1l
bound), as a res~lt of a parasite infection may not be due
801e1y to the productio~ of specifie antibody (Houba and Ailiaon,
1966; Smithers, 19677 Houba et al, 1969; Freeman et !l, 1970:
~udson !!!l, 1976). It has been generally assumed ~n parasito
logy that the production of circulating antibodiea and protective
lmmunity are synonymous; it has, however, se1dom been possible
to correlate an -àntibody reaponae with protective immunity in
J
Page 36
1 1 ,
1 1 •
i J
1 l
f
o
22
parasite infections (Soulsby, 1962; Smithers, 1967: Tanner, 1968;
Sinclair, 1970). Neverthe1ess, it ls probab1y true that anti
bodies play an important role in imm~nity in trichinellosis
(Larsh,1963).
Antibodies have been shawn ta form precipitates around
the ·orifices of worms in vftro following their incubation in
immune serum (Mauss, 1940b, 1941; 01iver-Gonza1ez, 1940, 1941;
Offutt, 1941:" Roth, 1941; Chute, 1956: JacKson, 1959). It ia ,tif
J
accepted that antibodies interfere with the metabolism, stunt
the growth, reduce the reproductive potentia1 and cause
physica1 damage ta the worms in vitro (Wake1in, 1976). and it - .,
has been assumed that such effects a1so occur in vivo. Worms
examlned with the e1ectron microscope have shown degenerative "
changes during the later stages of infection (Love et al, 1~76)
and prote~ precipitates have been observed deep in the mucosa ~J .
of immune mice after a challenge infection with !.spira1is
(Larsh and Race, 1954). It is a1so known that precipitin titres ,? ...
in mice increase progressive1y after each of four stimulating
infections with 200 1arvae, given at 21 day intervals (Hendricks, 'f
1950). Moreover, exposure to homologous immune serum in vitro J
has been shown to have de1eterious effects on the larvae
since this treatment reduces the infectivity of 1arvae of
!.spiralis in uninfected animaIs (Mauss, 194,Ob, 1941; Kwan, 1971).
A number of workers have succeeded in transferring immunity, )
as evide~ced by an accelerated worm expulsion, by transferrinq
.erum trom immune ta naive animaIs (Culbartlon and Xaplan', 19381
"
1
Page 37
..
, '
! !
,(f
23
Oliver-Gonza.1ez, 19417 Cu1bertson, 1942b; Hendricks, 1953; MUls
and Kent, 1~65). Other, investigators have, however-, failed to
demonstrate accelerated worm expulsion using this sarne treatment .
(McCoy and Bond, 1941: Larsh!! al, 1964a, 1970b: Larsh, 1967a:
Denharn, 1969; Gore !! a1,~1970, 1974). 1hese conflicting
results might be explained by the" work of Oliver-Gonzalez (1941)
who demonstrated specifie anti":adult and anti-1arva antibodies
in the serum of infected animals: the leveI of these two antibody
fract~ons in the serum varied at differ.ent times durinq the
course of the infection. Thus, conflictinq resu1 ts of the
protective effect of serum transfer could have been due to the .
time after infection that the serum was drawn and ,the amount
and the specifici ty<tof the antibodies i t con tained. A transfer
ofù immunity against Trichinel1a spiralis has also been demon-~
strated from mother t? Offs~!ing in bot]) rats and mice (Mauss'
1940a; Culbertson, 1943; Duckett et!b, 1~72; Perry, 1974); ~his .(
resistance:,was apparently transmitted post-natally via the milk"
and not in utero.
~tudies on parabiotic rats wi th a common blood circulation
have also provided evidence that antibod,ies can play a role in
immunity to !.spiralis (Zaiman, 1953; Zaiman and Rubel, 1953;
Zaiman et al, 1953; Boyd and Petersen, 1954; Zaiman et al, 1955a, -- -- 1-
1955b). The results of these experiments have indicated that
the "uninfected" mate of the infected rat was more resistant to
infection with !ospiralb than control animals al'ld, fhere:fore, ,
atrongly auqq •• ts the tranafer of ~rot.ctiV8 hum,pral,lntibc;)CUIUI.
J '
o
Page 38
()
1 i
f
1
o
24
Parenteral injections of. living, heat-killed or dried
powdered larvae have a1so been ~hown to prod~ce an artificial
immunity in recipient animais (McCoy, 1935: Culbertson, 1942a; . ,
Dorin, 1946; Despommier !! al, 1971); excretion and secretion
pIlOducts of the ~orm can also stimulate sorne protection
(Campbell, 1955; Chute, 1956), indirect1y suggesting that
immuni ty oin trichinellosis is a generalized Phenomen~ Larsh
(1~53) showed th~t mice previously immunized intracecal\Y with
,!.spiralis harbored aboout the sama 'number of adults from a later
oral challenge as infected con troIs given an immunizing infection 1
.1) orally. Since the intracecally immunized mlce had no chance
to develop local immunity in the proximal half of the small
intestine, but yet showed the same degree of immunity as those
mice orally - infected, the role for humoral factors fn immuni ty
was again indicated (~arsh, 1953).
In considering the role of""humoral factors, one cannot r • c-/"~
overlook the possibili~y that those antibodies which produce
irnrnediate (humoral) hypersensitivity (IgE) may play an important
ro1e in the immunity demonstrated. It is apparent from many
studies tha t !.spiralis can act as a powerfu1 inducer of homo-
cytotropic or reaginic antibodies (Sadun et al, 1968: Catty" --\
1969.; R.ivera-grtiz and NusS~~eig, 1976; Perrudet-Badoux!! al,
1976). The ~sequent subcutaneous i~~culation of metaboliç. , or somatic antigens 'of this parasite i'iito mice act,ivelY sensi- J
tizèd to !..spiralis by Infection or by, the injec;tion ~f somatic
antigens caUlel marked local reactions which are characterized A
il J
• J
.~l
. , , " .
,- .
Page 39
j
·1 1 1
t 'f
, ~ 1 l .... _~'" ___ .... -=-__ ..... ___ . _____ _
J
..
25
o
i \ by mast cell disruption (Bri,ggs, 1963). Br;i.ggs and DeGiusti
, ' 'Il-
(1966) sh<?wed that~ hypers~nsitivity reactions to intravenously
'adroinistered antigen could be elicited in mice before ~he eli'Co
mination' of intestinal worms; thus) host sens,itivity could be
tempdfâlIy associated ~i~h worm expulsion • ;
.,. The observation 'that strains of mice.showing wide differen~
ces 1'n their abili ty to form aritibody ar~ e'!:}yally infected by
!.spiralis, 'as judged by ~the number of muscle Ia,rvae, may , ,
suggest that humoral ~nt~bodies {and specifically anaphylactic , ' .
• ntibodies) are not primarily responsible' for the mechanism .' / w~ich protects against reinfection (~errudet-Badoux et al, 1975).
, " ---However, one ~~nnot exclude the possibility that antibodies
~
play a role"in protection, only in cooperation w~th the cellular ,
arm of the immune response.
c. The R9le of cellul~~Factors in the to Trichinella . \
spira1is .} ,
The ~articiPat.it~c of a -é~11-~diated immune r.esponse to tne -~
parasite
y, - . , in the control of triçhinelrosis was ~nitially
" suggested by observations of the intestinal inflammatory
to challenge infections in mice .(Larsh and Race, ~954; Larsh
!!, 1956)., it was iater shown. tha~ lymphoid cells frqm infect.ed·
animaIs conferred immunity when transferred int~ 'normal recipi~ _ , ~ . ' , ... ent animaIs' (Larsh et al,·1964a, 1964b, 1966; Gare et al, 1970, -.-- -- ,
L, • 1 ~ 1 ';1
1914; Love et al, 1976~ Wakèlin and Lloyd, 1976). Moreover, t.he
patte:rn of ,int:e'tinal inflannati~n, including thè 41ar infil .. JI' "- '. ,-0
f
... ..
Page 40
/
(j
j,
,. c
... -_._,----_ ... ~~-_ ...... _~-
1 26 , 1
tration in the 'tissues of the "recipients, was similar to ~ha~ odescribed in various earlier studies in challenge infections
(Larsh and Race, 1954; Larsh ~!l, 1956); the inflammatory
response was initiated ear1ier, developed more rapidly and
was more severe in mice given cells from infected donors than ,
in infected animaIs which had not received the lymphoid ce11 ~o
transplant (Larsh et,a1, 1966; Larsh, 1967a). Lymphbid cells -- -from animaIs infected with !.spiralis were a1so able to trans-
. fer delayed hypersènsitivity to normal recipients, as measured
by the macrophage inhibition Assay (Piotrowski, 19,75) ..
Sensitization with a crude saline extract of. !~spira1is
larvae produced effects that caused'a significant expuIsion of
worms after challenge (Larsh et al,_ 1970a, 1972, 1974) J, spleen
cells trom such treated mic~ were responsibJ for a simi1ar It 0
effect when transferred into subsequent1y-in ~cted syngeneic . ,"'-animals (Larsh et al, 1969, 1970a, 1970b, 1972). Tiss~e
o
sections obtained from mice sensitized with a crude saline
extract of larvae revealed an infilt~ation in which mononuclear ,
cells pr~dominated,~ pheno~non characteric of a de1ayed
hypersensitivity response (Ki~, 1966aJ Larsh ~ al, 1974).
Ani~a1s sensitized with the'crude,saline exttact a1so demon
strated ,a t~.pical de1ayed skin ~on against an !ntradermally
introduced'test antiqen; this hypersensitive state could be
transferred to normal" recipients by lymphoid cells from sensi-~ ',.
ti zed donors,. in the absense of antibodies (Kim, 1966a, 19606b:
" Kim ~!l, ~967, 1970). Histopatholoqic studies have demon-
Page 41
o
, t !
',' ;"
, .~
o
'" . ,
, J,
--------_.~
27 l '
strated that the transfer~f spleen cells from sénsitized donors
, to normal syngeneic reeipients also resulted in an earlier and
more acute deqreë of inflammation after ehallenqe (Larsh!i al,
1974). Further evidence t&at delayed hypersensitivity develops in ~ .
animaIs infeeted with Trichinella spiralis, as weIl as in
immunized animaIs, is given by the fact that stimulated lympho-o '
cytes of infected animaIs release factors which inhibit the
migration of macrophages, a response in ,vitro that parallels
delayed hypersensitivity reactions in vivo (Cypess and Larsh"
1970'; Cypess et !!' 1971; Kozar 'and Plotrowski, 1971; Kozar ~
al, 1971b; Stefanski and Ma1czewski, 1972; Vernes et al', 1975a).
, "
Using th~ lymphocyte transformat~~"~ and
Crandall (1972) and o~sen !i al (1975) have detected antigen
specifie cellular rea'tivity in the spleens, lymph nodes and
Peyer's patches of mice and rats during infection. Immunohisto
logical studies in experimental trichinellosis have also' shawn
a morphologie expression of the cell-mediated immune response
in an increase in the number of pyroQinophilic blast ceils in , ~
the thymus-dependent areas of lymphatic ,tissue (Ruitenberg and
Duyzings, 1972).
Additional evidenee supporting the functional role of
cell-mediated immunity to !.spiralis has also been provided by
thé results~f immunosuppressive treatments, Congenitally
athymie (nude) mice demonstrate an impaired ability to eliminate
. adu~t worms fram thé gut (Ruitenberg ~d St~renberg, 1974),
.. Go.ra !! al (1970 , . 1974) haVe ob'tained a delayeci expulsion of
Page 42
"
, ' -~"""-----' ____ .. _~_ r _"~ _~ ____ ~ ____ .-. • ___ _
28
intestinal worms, as well as higher n~ers of muscle larvae,
in neonata,lly thymectornized rats 0 0 ' On the other hand, Kozar et -~
!! (1969) and Faubert and Tanner (1974b) did not observe any
changes in the number of muscle larvae in mice that had besn
thymec'tomizeà in the neonatal period ,or when they we,re adulte
However, the latter authors noticed that the 1ymph nodes of
~ymec:tomized, infected animaIs did not increase in size as
as they do in non-thyrnectornized infected mice~ they conc1uded
that the enlargement of lymph nodes durlng trichine1losis was . , ~
thymus-dependent tFauber,t and Tanner, 1974b). Thymectomized, \\
irradiated, bone-marrow redonstituted mice can show increased • J
numbe~s of muscle larvae, P\O~ab~y rêfle~ng the,persistence
of adults in ~he gut beyond the time of their normal existencel
the inflammatory response in these animaIs was a1so negligible,
as compared to con troIs (Wal1s ~!!, 1973).' Gore et al (1970,
1974) prolonqed the surviva1 of worms in the intestine and
increased the l'lumber of 'larvae rècovered by treating rats with
anti~ymPhd~te serum (ALS), similar but more striking eff~cts •
. were demonstrated in rats which had been thymectomized prior to
treatment with ALS lGore ~ al, 1974). Antithymocyte'terum
(ATS) treatmènt was also effec1t.i"~ ;n retarding the expulsion "-
of a~u1t worms from the intestine and in increasing the yield "
of muscle' larvae (I<9zar ~~, 1971a; Machnicka, 1972; Ruiten
berq et al., 1974). Finally, no differences have been found -- . in the morpholoqy of thymus-independent areas in both intact
and T-c~ll deprived rntce; however, differences have been found
1.'0 the thymUS-depen_de~ areas durinq the course of trichinelloais (.
o .'
Page 43
i l, 1
> '
"'MoI;~ ... ~",... .. _ ... ~~ ______ ~_ ~,ri _____ ,_.~ ... _. __ • ____ •.
Cl
. ,
\
29
(Ljungstram ~nd Ruitenberg, 1976),
D. Mechanism of Acquired Immuni ty to"!. spira1'is
1richine11a spira1is infection in 1aboratory animais has • (. 10 d ' been extens1vely useo as a model to stu y the development of .
immunity to nematode parasites but, Qespite considerable study,
the mechanism by which specifie immunity causes the expulsion 1 ),
of adult worms remains undefined.
.. During earlier studies on trichine11os1s, the hypothesis
1
of local and general'immunity received the greatest attention.
According to the hypothe,sil;l of local immuni ty developed by '~ -
Chandler (1939), intestinal responses played the most important
and dominant role- in functional protection; circulating antibody •
response was relegated to a re1atively unimportant role. Other
investigators, however, be1ievèd that the mechanism of irnmunity ,
rested primari1y on humoral factors, with secondary cellular . cooperationr this latter was' the hypothesis of general irnrnunity
proposed by Ta1iaferro (1940). At this same time, others pos
tulated othat re'sistance was chiefly mechani~a1' or allergie r
(McCoy, 1940: Culbertson, 1942), although this concept remained
relatively unnoticed. Sorne years later, Larsh' (1953) was able
to suppo~t the evidence which indicated a role for general
immunity in trichinel1osis, as postulated by Taliaferro.
Detailed studies of the cellular reactions in the anterior
ha1f of the small intestine convinced Larsh (Larsh and Race, 1954) "" ,
that the expulsion of adult ~.spirali8 from the ~ut was mediated
"',' .
. '
"
A."
" ';~ ..
Page 44
"-... ,- « ........... ~ .... ~ ~_~ ........... _~ _ ... .>' _ , ............... _, ................ _~ ~ ...... "i~",_"
30
by an acute inf1ammatory response. This inf1ammatory response
cou1d be Buppressed by cortisone and X-irradiation,' with a
concomitant persistance of the adult worm burden (Coker, 1956;
Yarinsky, 1962; Larsh ~ al, 1952), the inflammatory response
varied in intensity"in accôrdance with the degree of acquired
immunityoPfesent (Larsh and Ra~e, 1954; Larsh!1 al, 1959, 1962).
Larsh (1963) then suggested that a specific antigen-antibody
reaction triggered indirect effects of a panmucosal cellular ~- 1
f ' respons~; a1though both h~ral and cellular factors were re-
quired, the actual elimination was due to the cellular response.
A significant ad~ance in the study of immunity to trichinel-
losis was made when Larsh and coworkers demonstrated the passive
transfer of protection with lymphoid cells from previously
infected miee (tarsh et al, 1964a, 1964b, 1966). Evidence then
began to accumulate which indicated that delayed (cellular)
hypersensitivity d~ve1oped in infected and in immunized animals
(see above). These observations have led to the hypothesi~ that'
a specifie delayed hypersensitivity response between antigen- ,
sensitive T-eells and'antigen results in tissue in jury, followed
by an immunologically non-specifie inflammatory reaetion to
the in jury (Larsh and Race, 1975). Larsh has also hypothes~ed
that pharmaeologically-act~ve substances may also be invo1ved
in the reaetion and that acute inflammation is accompanied by . ~ .
an unfavorable environment which is harmful to resident para-..Jtf
sites and cause 'their withdrawa1 (Larsh, 1967a, 1968; Larsh and
Race, 1975). Larsh has expanded his concepts to sU9geat that
/'
Page 45
\.
t , ~, ! 1
J '" t t , f .r, '1 "
t t , • " ! t
)
l !
1
1 (
î
()
. G' "1 • ..
4pSCtc • ,(
31
the chain of events he visualizes in the control of triJhinel
losis a1so plays a prominent role in the immunit~o other
tissue invading parasites (Larsh, 1967b: Larsh and 'weatherlY, ,
19741 Larsh and Race, 1975). Despommier and Wostmann (1969) ,
have speculated that, as adult female worms approach a maximum
production of larvae, the hostts immune m~chanisms limit the
parasitels nutrition and thereby cause t~e worm to resorb part
of itself in order to continue the production of its progeny.
With increasing levels of immunity as the infection progresses,
"stunted" worms are forced to relin_quish their hold on the host ,\ \1
and are, thus eliminated.
;
Althouqh antibodies of different subclasses and antibody-
•
conta!ning cells have been demonstra~d in experimental trichine1-
10sis, it is generally agreed that humoral factors do not play
an important role in the expulsion of adult worms from the
intestinal tract (Denham, 1969; Gore et al, 19701 Larsh, 1967a; fer --
Stank~ewicz and Weatherly, 1975). However, the relative ease
with which immediate hypersensitivity (antibody-mediated) reac
tions can be induc~d in animals and the p~esence of hi~tamine
d and serotonin in the sera and tissues of infected animals'
(Briggs and Degiusti, 1970) c1early demon~trate that humoral
immunity must be, somehow, involved in the total immune response.
In fact, Catty (1969),rworking with infected guinea pigs, has
suggested that resistance in trichinellosis may have 'a stronq
anaphylactic involvement which results fram the dévelopBent of .
a long-term aensitizinq antibody with biologieal and phyaieo-
.,
Page 46
,', i
1 . i.
1
l
0
" .
. /
\
#'t •
. _ .. _------....... _--32
chemical properties analogous to the reagin of humans and the " "
reagin-like antibody of'rats. .. .
It has been shown that little'or no intestinal mast ce11 . ,
response accompanies !.spiralis in conge?ital1y athymie (nude)
mice (Ruitenberg and Elg~rsma, 1976) and in mice'treated with
ALS (Karmanska'et al, 1973): in both o~ these cases there i6
a significantly-prolonged suryival of adults in the intestine
(Kozar et al, 1971; Màchnicka, 1972; Ruitenberg et al, 1974).
Karmanska ~ al (1971) express the opinion that biogenic amines
\ liberated from mast cells influence the expulsion of parasites ;.
from the intestine since a pronounced peak mast cell response
<loccurs in the mucous membrane of the intestine at the time of
the exit of adult worms. L (,
Moreover, the transfer of bot~mesenteric lymph node cells ~ 1
and serum brings about a marked acceleration of worm expulsionJ
when the lymphoid cells and the serum are given separate1y, a
significant degree of immunity can not be transferred (Wake1in .
ana Lloyd, 1976). Furthe~more, sinee antibody-mediated damage 1 ..
to worms has been demonstrated (~ve et al, 1976), it seems .'
1J.kely· that ce1ls "and antibodies may work together to expel . 1
!.spiralis. Further evidence of the joint action of humoral
and cellular clements hlay, perhapR, he provided by the followinq:
niridazole, a potent suppressor of cell-mediated immunologieal
responses, fails to alter the numbers of adult worms ln the
intestine of experimentally indtcteQ mice (~rove and Warren,
1976), indicating that the elimination of adult worma fr~the
Page 47
-
o
. ~
~
"
, \~ ,\
.. '
33
intestine may require intact humoral and ~ellular immunological.
mechanisms. Moreover, since the immune response te !.spiralis
is complex, it'is more probable that a11ergic inflammation
(Larsh and Race, 1975), antibody-mediated werm damage (Love et •
al, 1916) and other immunologically mediated anti-worm affects
act in concert (Wakelin and Lloyd, 1976) .
. .
\,
.' 'l,
,
.'
Page 48
, ! 1
~. ;,1 \ j _ ,
t- " ---------- \' <. ;-_:_~~ ~.::.' -_!'" ;-. ,:. ~~.:..- .. ;.... •. ,.
(l
34
MATE RIALS AND METRODS
A. Parasite
The strain of Trichinella spiralis used in this study was
isolated many years ago from an infected pig. It has been G:\
maintained since .then exclusively, with periodic passage, as
stock infections in Sprague-Dawley rats.
B. Host
Young adûlt 15-20 gram female AIJ mice, purchased from
the Jackson Laboratories, Bar Harbor, Maine, served as the
syngeneic donor as well as the principal animal host throughout
the course of this study. The mice were maintained in groups
of six in 33 cm x 1S cm x 13 cm plastic cages and fed commercial ,:: "9
1ab chow ad libitum. A total of 800 A/J mice, both donor and
recipient animaIs, were used in this study •.
C. Infectiv~ Muscle Larvae
The infective muscle larvae were recovered from donor rats
by pepsin digestio~ (Tanner, 1968). The carcass, decapitated,
eviscerated and skinned was digested, after homogenization in a 11
Waring blender, in a solution of 0.3% pepsin (w/v) in 3000 ml
of tap watcr containing 0.4% hydrochloric acid (v/v). Diqcstion
was' done at 37°C for approximately four hours in a four-litre "
Erlenme~er flask; the digestion ~ixture was fiitered througp
cheesecloth into a 2000-ml Erlenmeyer'flasK and the larvae
allowed to· sediment.. for half an hour. 'the larvee were then
. ,
Page 49
~...,.~ .. __ .-
1 i ,
.~.--.2--... ----. '. '. ,
" .
1
3S
. callected by aspi1ation. walihed three Urnes with phosphate
buffered saline (pH 7.2} and c6'unted.
D. Counting of Infective Larvae and Infection of ·the Mice· \
After washing the freshly-digested larvae three t{mes ,
in buffered saline in a 15 ml test tube, a magnetic stirring
bar was then .placed inside the tube and 'the larvae stirred until
a homogenous suspension was obtained. Further dilutions were
made from this stock until a'suitable suspension was obtained
for counting. The counting'was done in a McMaster nematode
egg-counting chamber (Hawksley Ltd., London); the dilution to
be counted was kept under constant agitation during counting.
Ten ,samples from the final suspension were coùnted; these ,
counts were then averaged and the number of larvae Fecovered
from the infected ani~al calculated, taking int~ ~ccount the
dilution of the worms and the volume inside the chamber.
After counting, the larvae were diapensed in individual
volumes calculated to contain the appropriate inoculum for o •
each animal: the mice were inoculated with the parasite on the
sa~e day of digestion. The animaIs were inoculated by mouth . ---~,;),
without anesthesia ~hrough a blunt 19G stainless steel needle
fixed ta a tuberculin syringe.
E • Thy!!'Cc tomy
The mice ta be thymectamized were anaesthetized by an po
intraperitoneal injection of 10\ Nembutal (Abbot~ taboratori •• t \
. :1
Page 50
j
1
J , ,
-... _ .. _~-~ .. --
o
1;\ \.JI
. ----_ ..... .- ..... _. ._---,-_.-
Montreal) 'at a dose of 0.1 ml/, 10 gram body weight and thèn
placed supine on a cork board. The four limbs were stretched
and immobilized with adhesive tape; ,the head of the aniC1 was
Q held bac::k by a rubber band. The sl:tin of 'the cerv,.ical area waB
swabbed with 70t alc9hol and then an incision w~s made along 1
the midline from the. top of the ',sternum anterior~ for about
l cm. The trachea was exposed by incising thé surrounding
membrane which was then retracted. A plunted 0.5 cm 0.0.
glass cannula attached to a motor-driven a~pirator, waB
inserted Along the trachea ~nder the clavicles into the
mediastinum until the tip came in contact wi th the thymus; ~
the gland was gently loosened from itSi bed and the .suctton WBS
then increased until both lobes were sucked through the tube.
The wound was then closed wi th one or two surgieal clips 1
(Autoclips 9nun, C.l'(lY Adams Ine., New Jersey) or by suturing it
with 5-0 silk (Ethicon Sutures Ltd., Ontario).
F. Irradia tion
Irradiation was done two or three weeks following thymec
tomy with a Co~alt-60 source in an Atomic Energy of 'Canada
'!Gamma Celi 220" at a dose rate of approximately 131,650 rads
per hour. A lethal dos~ of 850 rads of total body irradiation . ,. ~
, , used in these studies was obtained by sh~eldin9 the animals
from the source vith a 70\ lead attenuator.
G. Collectipn ~ Tran.fer 2! Thyau. ~. Spleen Cella
The thyau8 and the .pleen vére ~ved ••• ptic.Uy frOil
l.
.)-1
, \
Page 51
..... -., ~.- _1l" u ........ __ , ~ ___ ... _~_ ~ __ ..,. __ ~ _ ..... ______ .-.. ___ ,
infected and normal donor mice. 'The thymuf was blotted dry 'on'
a sterile gauze pad and the adhering non-thymic tissue anèi
o mediastinal lymph nodes care.fully removed. Cell suspensions
were prepared by passinq each o~gan through a sterile stainl~ss
steel wi~e cloth (80 mesh4,~l6 gauge) with a sterile glass
pestle into a 5 cm Petri dish colltaininq ice-cold sterile , ,
RPMI-1640 medium which contained 10% heat-inactf'vated fetal ,
-:palf serum, 100 U/ ml' penicill~n, 10.0 mg/ ml s treptomye in, 1 " U/ml heparin, Hepes and sodium bicarbonate. Cell suspensions
o
from homologous animals kept in ap ice bath were then pooled
and an aliquot "as taken to determine the number of viable -
cells by counting the leucocytes" in a NeUbauer haemaeytometer , c
in the presence ot 0.2\ ~ypan blue; o~ly<> those cells from
\
which the dye was èxcluded were counted. The pooled écell , 'r' suspension. were wa8~ed once' and reau_nded te a concentration \1 of, ,1 x 107 cells/ ml. Immunologica11y deprived recipient mice
~
were injected intravenously in the caudal vein wi th th. appropriate cell sUàpensiop ~n a total volume of 0.2 ml. •
• . H. Collection!!!.2. 'l'ransfer 2!. !Sm!. .Ma .. r:;.;;r ... ow.- Cella,
l:nfected and normal A/J mice were kill~ and their femur.~
removed., the epiphyses ware cut off asept,ically with soi'slors " ,
and approximately 1.0 ml of ice-C'o~d sterile, complete RPMl:-~640
medium (aee above) wa, forced throucJh, the shank of the bone by
Dleana of a tuber~ulin syrinve ~itted with a 26G neecUe. t'he
bone marrow cell. troll normal or infected dOnor8# kept in an Jf.I- . 0 ". -"
ice ba.th, ,~. poole<! and"a 0.1 ml aliquot vas taken ,to detez:1D.ine
'~;.t ,,' .. ' >'
1\
!_ .. "L "'t; :'f:>:, ~:-.u:_ '.,-,~.~.,ÜL':tii __ :
Page 52
'1
i f
1
,---....JR, ___ .
f3
0'
38
Il the viable cell count by trypan ·blue exclusion. Concentration
of the cells was adjusted so that l x 10 7 viable bone marrow 1
- cells were injected either alone or with thymus cells into t.he, ,. ,
tail veins of thymectomized i'rradiated mice in a total volume .y
of 0.2 ml.
-? " 1. Antigens and lmmunization {'SI
~hëeQ red blood cells (SRBC), stored in sterile Alsever's 1
solution at 40 C, were washed three times in sterile normal 1 .
saline (O.8sit and resuspended for intraperitoneal injection
to a. concentration of 1 x 108 cells/ 0.5 ml. '.
" . d\ d . . h' . 14 ï The T-cell l.n epen ent antl.gen Escherl.c l.a coll. E 5 i' ,
lipopolysaccharide (LPS) obtained by phenol-water extraction
(courtesy of Dr.J.E.Tanner, pepartment of Microbiology) was (
coupled onto SRBC by incubatinq, for 30 minutes at 3.7oe, one
volUJrié of packed' erythrocytes wi th fi ve volumes of LPS in a 1
concentration of 1 mg/ ml normal saline: before incubation with
SRB~, the ~PS had been boiled for 60 minutes to destroy the , \
toxic lipid ~. The LPS-coa ted SRBC suspension was washed three
times with sterile saline and _diluted to a concentration of ... ~.
1 x 10 8 cell~1 O. 5 ml prior~ intraperi toneal injection i..Ç" . recipient mice.
~.
" 'J
Plaque-formin9 S!!! AS8~Y The method \1sed for assayinq the nl.8ber of antibOdy-foraù.nq ,
cella in thé apleene of mica iRlftunized vith SItBC or LPS4a •
••• entially c1e.cribe~by Cunningham and S •• nberg (196.). Spleen
<
Page 53
, '
~ 1
f l f. r. >,
r
1 r ~ , 1 \,
1 1
i
cr
"-
' .
. ,
, . ( ,l'
-~-..... ~---- .. ~--_ .. _--- --~ ---- -------~ .... ..,..-- ... ~;.....,.ft'- .... ~-~,.... '. '
39
, L
cell suspensions in' the domp1ete. RPMI-1640 medium were prepared'
and counted as described above. ' The number of cells was
adjustep t,a 5 x 107 CÉüls/ ml .and then 0.1 ml of this 8uspén8~on ~
,wa.s mixed in àn ice bath with 0.4' ml of a 20% dilution, in the
complete culture medium, of guinea pig complement (BB~, Becton,
Dickinson and Company 1 Maryland) and 0.1 ml of a 20% suspension ~ ,
in medium of sheep red blood ce-l!s. The mixture was then used
to fill' 5'-or ~ Cunningham chanmers prepa:r:ed by joining 'two
75 mm x 25 mm slides (Ingram ~nd 'Bell Ltd., Montreal) with
double-sided "Scotch" tape. The filled chambers ~ere then
se.aledowith heated paraffin ~nd incubated at 37°C for 60 minutes.
The lj.umber of hemorytic plaques was deter~ined by counting the
areas of discrete hemolysis in the SRBC substrate by macroscopic
ëxamination under indirect fluorescent light ag~inst a black,
background. This ass;;ty. wa~ always dorte on 1!he four th day
following inmunization of th~ anlma1s with the ~rythrocyte anti-.
gen) since anti-i~unoglobuiin serum was not used. in the, assay, c: (J -' 6
only ('direct, (IgM) plaques were de.termined.' . '
1(. Marbrook Cell Culture Chambers
This cell culture system used to study an tibodY responsea , &
'. !!! vi tro ~aa essen tially tha t deve loped and", described by
Marbrook (1967). Spl~en cell suspensions·were pr~pàred and .. . counted as above. Plastic-capped 2.S cm x 6 cm- high. scintilla- ' .. tion via ls were used to qons~ruct the Marbrook chambers used in
these experim,e'iits. 0 A noIe large enouC]h te hold a 1.3 cm
@ ':' diameter x 7 CIft hi9h~er tube was made in the cap of the vial.
8/it!'
"
.\
"
Page 54
1
f
,. ..
---"_.-.... -._~ ....... -----... _- g...- .......... ,...., .... --oyd»---_ .. _- . ---
o ·'1
(J
o
40
A dialysis membrane was stretehed aeross "the end of the inner
.tube and held in' place with a "Silastic" non-toxie rubber band
and trimmed: the oPPosite open end of the tube was loosely
covered with a plastic cap and the vial and th~ tube' were ,,"----
assembled to produce a culture vessel consisting of a relatively
smaU inner chamber in a largèr outér container. Fo11owing .'
autoclaving, tHe chan:tbers were -dried and coo1ed and the outer
chamber was filled \ aseptically with the complete RPMI-1640
culture medium (see above) to a level just above the bo~tom of
" the dialysis membrane-covered lower end of ,the' inner chamber.
0.5 ml of the s:pleen cel1 suspenB'ion, ~ontaining 5 x 10 6 ce1ls, ,
and 0.1 ml of a 0.2% sase suspension were then aseptically ~
pipetted into the inner tube. The chambers were then incuba ted
for fç'uJ;" days at 37°C in a· humidified, 5% CO~ atmo~PheJre.
An tibody formation to the antigen was determined usinq the
elaq4-e-forming cell assay (see .above) .' /
L. Statistical Analysis
The "Student' s" t-distribution was, used to analyzé the "----
dat-a of a11 experiments. This test was used to relate thé ,
d~ta obtaincd from untreated controls to those obtained from
treated groups. The data wau considered statlstically signifi
_cal!.t if the "t" valué was above the five percent level (Steel
and Torrie, 1960).
Since the ra"qe of the data sometimes giVEts., unnecessary , ,}
wight to extraneoull values in an •• timate of di.penion, the o
Q test was used to eliatinate those value. which failed ta pass
,,.
Page 55
i
t l
!
t
r 1
"
41
the screening test (Dean and Dixon, 1951). The Q statistic,
applicable to ten ~ fewer in4ependent observations, 18 defined
as (x2- x 1)/ w where (x2-xl ) is the distance of a doubtfu1
observation from its nearest neighbor, and w ia the range. If o
the calculated 0 value exceeds the tabulatéd values, then the
questi(mable observation may be rejected with 90\ ·confidence'.
/ ' ..
.1
\.
Page 56
o
/'
~ , 'r-.", . ", . '~' ,- "'110.,: '-!l \ 1 ~~i \, ,~ ...
~ .... - ~ ___ ... .....:.....:;.II ~ __ .:: ___ --. ~-:.~,_""....;,_' :._'._._; _. _._ • .-___ ....... __ • __ _
42
RESULTS
Experiment 1: The' level of infection in A/J mice followinq o
inoculation with different dQs •• of infective
larvae . .. -ro determine the characfferistlcs of the ~ost-parasi te
1
relë.!-tionship between Trichinella spiralis and the syngeneic line
of A/J mice used in this study, a dose response curve was Qb
tained. Fifty female A/J mice were ai vided into five groups and
treated as follows: one group was 'inoc~lated orally with 100 .. " c viable muscle larvae per mouse," the second group received 250
4arvae, the third qroup 500 larvae,. and~the fourth and fifth } 0
groups, 750 and 1000 larvae, re'spectively. AlI the mice were
, inoculated on the same day and wi th the same poo~ of, larvae
recovered from stock infections. Thirty days after inoculation,
the mice were killed and the levèl of parasieism was determined
bf c9unting tHe number Qf infecti ve musc~e larvae recovered
from each mO,use fOllowing peptic digestion. The data were not . pooled.
" The results iri Figure 1 illustrate that, generally, th~
intensity of muscle igvasion in mice J:>ears a direct rela~ionship
to the number of larvae inoculated. The number of muscle larvae
recovered increased logarithmically as the number of larvae "-
, inoculated was incre,aBe~ . ---. This increase was observed up~ to the
dose of 750 larvae:. ~he inocul,tion of 1000 larvae did not
sighificantly incr~a8e the levaI of para.itism above that ob
tained vith 750 larvaé. In lIlOre detail, the n\8ber of ausele t
Page 57
)
.'
.., . ,
(,
,
/ . [
Fiqure 1. 2iPe mean loq number of muscle larvae ' recovered fP'om A/J mice 30 claya after inoculation wi th
different doses' of infective !.spiralis larvae. lite nUmbers in the parentheses indicate the number of mica used to obtain the Mean value of larvae recovered at
each levei of infection, and each bar representJl- tw1 standard érrors •
/
. • .' '> l,. ..
•
. '. P" , .'
. ,
Page 58
~. ,l, (l~i
) ~~ \'
,f
1
" , , ;f ..
C
• tf
4l
." . -,-
.'
Q ~ ... a ....
'> 0 U III II:
III
~ 1 ~
... 0
~ lU • a i
" 0 ...
+3 ...
-" '1 .
1 •
,
L 0011 IES'O ... TO T·.,"'Ab··
1 --1» ...
1-0 IOopOO ,. 0
~ .,. r-
• ! l'II
,. 4.0 .opOO
fil ft
~ g ~
... 0
(l0)
3-0~ __________ ~ ______ ~ ______ ~~OO
100 250 500 750 .000
"""IEII OF LAIIYA! .NOCULATID .f
"
,_......:-______________ - ..... 'T--~' ,----,
Page 59
.; 1
{" !
.'
"
---1!'f\\'W ___ • __ ~_ ..._ __ _
'.
, l
44
larvae recovered following the' inoculation of 100 and 500
larvae were significantly different (p ( o. OS) from each other;
the number of 1arvae recovered ,following inoculation with 500
and 750 muscle larvae were also significantly different (p ( 0.05)
from each other. There was, however, no significant difference , .
between the number of parasites obtained fr'Om mice f?llowing ''''
inoculation oL250 and 500 larvaé.
The experiments described below are based on the res
ponses given ,by A/J mice, inoculated with 100 ()r 500 larvae
sinci' the ~ormer dose prod?ced a relativel~ light inf~ction , ~-
and the latter gave moderate levels. Lar.ger inocula were not
studied because patholoqical changes induced by high par~sJ. te
\,burdens mi9ht, ~b8eure ~e interpretation of immunologie.l
. phenomena.
/ r
Experiment 2: The abi1ity of A/J mice inoculated with 100 or
500 !.spiralis larva~ to respond to immtinization
wi th sbeep. erythrocytes. The time course of
this response.
An experiment was done to establish whether low ... and
moderate levels of trichinellosis reduce the ability of the
synqeneic line of A/J mice to respond te sheep erythrocytes ~
(SRSe), and to determine when this immunosuppression occurs.
'l'wenty four mice were orally inoculated with 100 arvae and an
equa1 number was inoculated with 500 larvae of iralisJ
aixteen uninoculated mica aerved ... controle. On 'the 7th, 14th,
28th and 56th day of the infection, .ix aie. lnoculatad wlth
Page 60
t •
·"1 . ..
.' r 1 ~
-::::rzc:;-~--~
,"
--
Figure 2. The plaque-f?rming c~ll (~FC) reaponae to
sheep èry~hrocytes of A/J mice at different times
after inoculation~ith 100 or 500 T. spiralis larvae.
The PFe ;esponse is expresaed as a 7percentage of the
normal '(control) re.ponee. Bach hiatogram represents
either six infected animals or four controls, each bar
re~reaents one standard error,. .The asterisks deno~e , .a significant difference (p(O .05) of the experimental
• group to the control group.
t
'l
l ')
(
\
" .
f :_ ••
1 • ,1 ~I l"· ;
, A.. ~"
;. ~I ~'!! ~~" J l.
1"' ...... {o!
o
. -
'.
Page 61
~-' ... --~--.. ," .. t~ ~ ,ol" ----
1 1 ,C
~ Cl '1
~ , ;: ~ • . ~
t 6 ~ .u t-
i
\ 0
1 ~ -c
0 -w1
., -Z
"" nt :' n ... -o~ z
" •
,~---.... - -._---..;..-- - --.~
....
-•
JO •
-0 0
ca 0 0 n
-g ca 0 0
n
0 0
S 0
n
ë o UI o o ft
• o
----~- -
.. o
.-
•
~- ... --
..
SPLEEN
CI o
c-..:
-o o
45
CELL S IN PERCENT
-JO o
-... o
."
"" n
" ~
'" en ." 0 Z· Ut
'" ... 0
Ut :a • n
-z J
-t
' 1
i ... ,1
I~ -t .'" 0
K -n ....
"'.:::' , ., , '
"
, ,,,. " '
1",',
Page 62
.\. 1
~
1 1
-1
.'
r 1
.,
- . . , --_._--,.- ------_ .. __ ... --""".,........ ~-~-46
, . . either 100 or 500 larvae and four uninoculated contr(l)!s were
immunized by an intraperitoneal injection of i~108 SRaC.
These four days were deliberately chOsen to represent the
intestinal' stage (7 days), the migrating stage (14 days), a",d
the ear'ly and late sedentary st~ges (28' days and '56 days; respec-.)'
. tively) of the' infection. Four days after immunizâtion with the
erythrocytes, the spleens of th~f: infected and control mice
were removed and the number of ~ntibody-prod~cing dir~ct plaque
formin~ cells was determined.
The res~~"'h~ in Figure 2 demonstrate that miée
inoculated wi th, 500 larvae responded significan tIy less well
(p ( 0.05) to the antigenic stimulation of sheep red blood cells
at day seven of infection than did normal uninfected mice; this
significantly reduced (p:( 0.05) antibody response ta SRBC was
a180 observed on the 14th day of infection in mlce inoculated
with 500 larvae. On the other hand, those mice inoculated with
100 larvae showed' a normal antlbody resPQnse to SRBC on the 7th'
day of the infection, but a significantly diminished response
at 14 days {p ( 0.051, ,in comparison to uninfected control mice. o <
The antibody responses to sheep erythrocytes were normal in'
mice inoculated wiJ:h ei ther 100 or 500 1arvae on the. 28th and
56th day of the infection •
As immunosuppression"occurred in the A/J strain of mioe
vith low and moderate trichinellosis, the following series o~
experiment. vere deaigned to" det.~ne "hether infecti~n ha. a
direct effect on the i_unoloqlcal competence of the cella of
Page 63
\ 1
"
"
"
" " , .... -.~_ ....... __ ..... __ .. _. -- ." " . ..,.,. . ~ ','"' ,,',
~ ~ -- -_ ..... _~ •. _--- - --( +-- -- --~ .... _-_ .. ---~----... _---
47
the primary and secondary 1ymphoid organs. The immunologlca1
competencè of the lymphoid organs of mice infected with Trichi
ne1la spiralis jas tested in an envi~onrnent free of the parasite
in immurtologically unrespopaive synqeneic re~ipients. Lymphoid ~ .-
cells were tranaplanted at different stages of infection from
A/J mice with low and moderate trichinellosis to lethal1y
irradiated and/or thy,mectomized uninfected recipients. The
immunQlogical competence to respond to sheep eryth,rocytes of the
animaIs who ~ad received cells from infected donors was then ',--
compared to that of mice which had received normal cells."
Experiment ~ Cy
3: Reconstit~tion of immunologically-depriv~d recipi-ents with normal synqeneic thymus and bone marrow cells.
As the induction of anti~ody formation by many immunogens !
requirea the specifie interaction of both Band T cells, a
study was first done to determine the optimum combination of
thymus cells and bone marrow cells that would give the beat
immunological reconstitution in 'adoptive transfer. For this
purpose, ten syngeneic adult thymectomized, lethally irradiated
mice were injected with l x 10 7 thymus cells and 2.5 x 10'
bone \.,
rnarrow cells; thirteen mice received 2 x 10 7 thymus cells and
1 x 10 7 cells: and eigb.~ rrtice recei ved 2 x 10.1 thymus cells and
2 x 10 7 bone rnarrow cells from normal 'uninfected syng~neic donors.
One month following reconstitution, aIl the recipients vere
immu~ized with SRBC and the antibOdy response vas deterDdned
by countinq the (DOber of plaque-forminq cella in the apleena'
~! i 1
Page 64
"\ '.,.~ >:! \
• '4' -rot", " , • A ~.1
• 1 </ .! ~I
••
",
1
,J
Table 1. The number of plaque-forming cells to SRBC per 5 x 106 splenocytes, in thymectomized, irradiated recipienta reconstituted with different combinations of no~l thymus cella and bone marrow cells. Eaeh value listed is the mean plaque-for;minq-cell response of the number of mice in parentheses, followed by the standard error.
•
,"
" , '. , \ ~' '. • ,..!, • _a~'
.~. l '~{: ..::' : ·'C··:iS~;~~::~~;~~\, .. /< "
Page 65
,
","/ :.~ "-,
d 1 T~ . / -,
Q ',', -"~;,,~;,%T.:},~
'ç., 1(,," ~
, ~t ~ ~ r .... ,.,.
,}~lJ .., ~""'1I ~ .~ , <>
>"j' l , 'j,'
, .", .--, .~
.0
>
~t ~~~ ~ , ' •• >1
410 ...
"
~
:- ., ~
!w ,
1 r' o ,
/\.
Il) 1 X 107 1 . 1 ~.
THYMUS CELLS BONE MARROW , 'CELJ,.S 0
. ANTI800Y RESPONSE"
,;
2.5 )('107 o 220:ta 53 (7)
" 2" 101
1 )( 107 44!5 ;1; .• 5 ( 2. )
2lt 107 , '2)(107 337 :l: 71 ( , ) 6'
,
.~"
CtN'b '.e ~- ""V~,..- _ ... -~ -~ ·"4 ............... ~:::. .... ~ .............. ~ ...... ~ ....... "., .. ----~-' tt "'lin r p. . '1 é.nelr«Wr .. Y .. ' M' d' ' un .' III .. _ .. _ MWta'WH ..... ," , ' IllE \1
~\ o· •
"
Page 66
i
1 f
1 J
" \
\ 49
four days la ter.
The results shawn in Tabole_l, ,ind~ate that the highest ,
antibody response was obtained in recipi nts reconsti tuted 1
with 2 x 10 7 thym~ cells and l x 107 bone ~rrow cells, where-~ ., .
as the lowest response was induced in those mice injected with .,
1 x 107 thymus, cells and 0 2.5 x 10 7 bone marrow cells. On the
basis of these results, a combinati~n of 2 x 10 7. thymus cells
and 1 x 107 bone "'marrow cells was used in 'il study .. of the immuno-
competence, of adoptively-transferred syngeneic thymus cells
from mice inoculated with !.spiralis.
Exp~riment 4: The immunocompetence of the thymus of infected
A/J mice.
, The immunocompetence of the thymus cells Qf infected
animals 'lias measured by their abili ty to cooperate in elici tin.g o
a h~oral response to the T-dependent sheep erythrocyte' antigen
(SRBC) after transfer 'into syngeneic immunodeprived recilifents.
Thymus cells from infeéted mice were transferred into thyllÎÉ!c:""~
tomized, lethally irradiatéd recipients: bone marrow cell~s frem
normal uninfected mice were given simultaneously to provide a
source of immunocompetent antibody-producing cells, as weIl as
to protect the animaIs from the effects of hematopoiètic
due to irradiation. l'
Twènty seven, thymectomized,
reconstituted vith 1 x 107 normal
irradiated recipients were ~ . bone marrow cells, thirteen
.. of" these animals ,Jiéce! wd- 2 x 107 thymus cella bken frOID
0
\ ~.
~. ~
.'
. :~
Page 67
~ " ~' ,,'. ,\,' ., j' f' " ... ...;1, _, ,,~. 1 ~ "''\).: .:: ...... _ ....... m_ ..... _. ____ ' ..:.'~~ ___ ~ __ ......... ,......:_ i- ~" t
J ~ ~ III
.j~ }::: 1
Ji '" ~. ' I~: ... ;' -.~_~~~,~ .. .:.:',~.~;,~=,.:_...;, _'1"' ___ "' _____ '_'_'_
)
. ï
0'
• 1
"
"L- ' ." , l' •
-... ' .
Flgure 3. The plaque-forming cell respOnse to SRBC
o"f thymectomized, irradiated recipiente re~onstituted 7, 2 ' ,-," 7
with 1 x lQ normal bone rnarrow cells, and 2 x 10 thymus 'cells trom either nôrmal mice or from mice - '
inoculated with 500 larvae 14 days previously .. Immunizatiori ,with SRBC was done 30 days after celi
transfer. The nwnp~r of" animaIs in each group is '
ind;cated wi thin the parentheses p and eaéh bar represents one 'standard error. A s'ignificant difference
.~
at ,the five perce~t levél ie ill~àt~ated by the asterisk. f
"', \ \
0\
\'
..
, . ...----'- ,\
",
, ,
, '
•
\
., ' , 1
Page 68
600 '-
., 500 ..1 ..
..1
..," , U
• 400 -... ... ,
..1 CL fit 300 -• 0 -
)( 200 '-
1 0 i t
" f
100 ... U "-..
~, 0
" ' . " ,',
,~
• J '
-_iiSQloLc •• 4"'_ 1 ~~
, 50
"
THYMUS COMPETENCE
14 DAYS Of INFECTION
(2.) wf
-'"
(3) If.
T Ill'
1,1 : , "
,H
tll 1 , ;1' 1
dd,! i' : ',II !.I
illlllll!':!i 1: 1 t 1ft: :!Ill 1 I! :". T 'II l' j Pi, l, 11 ,: l , " '1 "I,I!" ,II
1'1: i' l' 0 1 ' .! 1
<1
'NFEC1~O NORMAL
DOMOR OF THYMUS· C'ELL;S 0,
'. \
--_._---_ ... ~. " ..
".~ ,
O!
" ,
" ;; 0
. " ,
'.' . ' : ,il:
Page 69
, : !
i
1 l ! i
. ' \ ' J .... ~. ____ ~_ ~_ ...... ___ • ___ _ ----"'-,----- --_ .... - "-' - .. _ .... _____ • _. """SOV ... _,.. ............ '" .... ,( ,...,; j"" __ -
\ 1
51
uninf~cted donors while fourteen animaIs received the same
number of thymus cells from donors inoculated with 500 Trichinella
spiralis larvae fourteen days previously. The recipients were
immunized intraperitoneally with 1 x 108 sheep erythrocytes o
thirty day's after cel1 transfer and four days after immunization t .
the number of spleen PFC was determ~ned. The results (Figure 3)
indicate that, thymus cells from infected animaIs were signifi~
cantly less capable than thymus cells from normal animaIs in
restoring the immunological response to ,SRBC in immunologically-, '
unrespons~ve recipients {p (0.05). This experiment suggests
that the reduced ability of mice to respond to SRBC fourteen'
days after the inoculation of !.spiralis larvae ls p~obably due,
at lea,st ln part, to a reduced immunocompetence o,f the' thymus
cells. Unfortunately, the n~er of surviving ~ce used in the
ebnstruction of Figure 3 was' small; lt is presumed that t~e •
~ortality in the recipient,mice was due to embolization 9f
clwnps of thymus cells in the lungs, whlch is oftcn el\countered ,
followlng intravenous inject~on.·
Experiment 5: Theo immunocompetence of bone marrow cells from
infected A/J mice.
To determlne the effect of trichinellosis on the bone
marrpw, ~he antibody response to the T-independent 1mmunogen
,E.coli lipopolysaccharlde (LPS) was used as the assay of the "'-- o
i~unological competence of these cel1s. Nine thymectomized, ~ -7----
irradiated recipients vere r~con.tituted with 1 x 10 boRe
marrow celll frOil noonaal ' aiee and a total of .eventeen .lee
'1
"
Page 70
o '-
/
" ,
'Figure 4. The mean' log plaque-formi'ng cell response ,
of thymectomized, irradiated recipients to LPS one month ,aft~r cel1 transfer of 1 x 10 7 b~ne marrow cel1s
from normal donors or fram donor anim~~~ inQculated with either 100 ~r 500 1arvae 7 days or 14 d~s previously. Each histogram represents the mean log PFC response of the number of animaIs indicated wi~hin
parentheses, and each bar represents onè7 standard error. '""'
Statistical significance i8 demanstrated by the a8~erisk
(p(O.OS).
/
"
"
•
Page 71
',:1 , ~:
:'1 , 1 , l' ;..::
-f~'7j ~,;:.r"1 :,'<<;', ." ·,i·'
!. ~t ~~ . ' ,!"f'~"
; " . ;\ ~ .' . "
, .
":{ "
-, , 1 '.. 1
~
"
l ~" ". "
1 ~ II'
j ".~.O r &&1 C,.)
co 0 -)( ,. II)
~
u 1·0 LI. 0-
1 e:» 0 i.J
~I 0
o ""
,'i: BONE (3) ..
.(,
1 (4)
T· (3)
, ';1 _ 1 1 1
..,
III l . ':'- 1 lIIl
100 500 C
-7°, . "
DAY
~
" .. _~~,._-'- ,-.....,..;.~I<""-,..- -
~
1 MARRO" COMPETENCE (
...----. .
-----, ~'
\,
.:. L (1) (7)
),
( -- ..,:..-
-,' .. "
1 -'-~"
~ :t ~, ~DOOOOOCI . ~. , --- -- .. j -:.. -' ,1
-
~ -1.
, "-
...". 100 500 ' C
" ( 14 OF INFECTlÔN, . ,
J:
~
'~ --~---- • f 'II Pl ln q' q'Q r ... • . as Cl 3 4. A!.l! 1 .. ,< 1
Page 72
!
\ \
-1
o
o
____ ....... __ ................ _J ""'_~" .... _-......,..._~_ .... \_ "
53
was r~constituted with ~one marrow cell~ from mice which had .
be~n inoculate~w~t.h either 100 or 500 !.spiralis larvae 7 days
or-14 days·previously.- One month after the transfer of the r
,cells, the .recipients were immunized with !.22!! LPS coup~ed
ta chicken red blood cells; t~e direct plaque-forming spleen
assay was then done using target cells of sheep red b100d cells .
coupled with LPS. The alternation 'in the carrier (of LPS) and ,
the target erythrocyte was done to measure only the antibody
response to LP& and not agai~st the red celle
The results presented in Figure "4 indicate that bone
marrow cells from animaIs infected for 7 days with 500 larvae -
have a'significantly enhanced capacity to restore immunological
competence to "the T-independent LPS immunogen.in thymectomized,
irradiated recipients (p < 0.05) as compare~ to: bone ·~rrow cella- from
normal animaIs. However, at 14 days of infection the bone .
marrow cells of animaIs infected with 500 larvae are no mo~e
efficient in restoring an antibody response to LPS ip deprived
recipients. than normal bone marrow cells. The capacity of
bone marrow cells to restore Gompetence i9 not, apparently,
affected by an infection initiated 7 or 14 days earlier by 100 .~ ,
larvaeJ the number of direct spleen LPS plaque-forrnin~ cells
vas not significantly different from those receiving bone marrow
cel1s from uninfected mtée. Control thymectomized, irradiated .
mice that had not'been transplanted with bone marrow cells died
within ten days of the radiation treatmebt.
Page 73
t , • t
!
f 1
1 _ ' ; , " ~
--" . , ---.... _~----_...:......-_-~~-._-- __ ._._-L..;...,. ___ ~_. __ ...... .. _~ ~._06~ .... ~ ______
" Ex~riment 6: 'The reconstitution of immunologically deprived
recipients by spleen cells from normal donors. 1
( . gate the eff~ t of trichinellosis on the immunological com-
S4
The f~l owing series of studies were desi~ned to investi·
petence of a seconda~y lymphoid organ, the spleen. A preliminary
study was first done to determine the kirietics of the reconsti
tution of immunologically deprived animaIs by normal spleen
~ells. Fdt this purpose, eighteen 1ethally irradiated recipients 7 '
were given 2 x l~ spleen cells from normal uninfected donors
:ithin an hour of the irradiation; these animaIs were subse~ , 8
quentIy immunized with 1 x 10 sheep erythrocytes 3, 6, 9, 16,
23, or 30 days after the transfer of 'the lymphoid oe11s& The
antibody production to SRBC was measured by the plaque-fprming ,
ceL! assay four days later.
In Figure 5 one can clearly see two peaks of reconstitu
tion: . one at 6 days and a second one 30 days after the immuni
zation with sheep erythrocytes. In more detail, when,antigen
was given 3 days following cell transfert very few plaque-forming , J .
cells were observed in the reconstituted recipients. Subse-
quently, the number of plaque-forming cells in the recipient
animaIs increased quantitatively, r~achin'g a peak when antigenic
stimulation with SRBC was given on .the si~th day after cell ~
tr~nsfer. A decline ih immunological competence then followed li
until the slxteenth ~ay following cell transfer: thereafter,
th~ competence lncreased rapidly until the:twenty-third~IY'
COmpetence, as meaeured by the number of PlaqÛé-forainq cella,
(), rellained constant until the last obaervation on day 30. 'rh •.
.1
.'
'.
Page 74
, ,
'.
.. Figure 5.r The PFC response of irradiated recipient.s
reconstituted with 2 x 107 normal spleen cells af~er immunization with 'SRaC at different times after cell
transfer.
. ,
, 1
.- '
',' , f'
. .. '
i , p
r
, .. '
" .:: ~ :'~:J: . ~ , ~~ , 1-' "
_.~Îl~~ (~ _____ ~_~_,~. __ ~~~----,-~~_~' ",--" ,,-' _~ ___ _
,"
Page 75
55 ',' ' , -.. 6
0 ,Fe /5 x 10 SPLEEH CElLS
~
N ~, .. Qt 0 0 0 0 0 0 0 0
,; ~
CIl
0
~ .. " "II
0 r r
~,o Z ~ ... -- Z , G') Q
'" , Z 1 n 'i "' . ~ 1= -.1 'CIt ..
.1 -4
~ :0
- ,. "
~ z CI)
z "'" c", :0
:0 . lit n 0 '. N
Z '" Z
'" en
,Z .. -... c: ... -0 a
"" CIl i '0
'~
• , - .. ::'('1- D
\ ' .
'!I
Page 76
1.
" , ; .. ' 1 - '~' -...... - • \, '" ilil."'
; J~.~: ,~~~~:~~.:--f· . ~ , , __ ~_~ ___ ............".~ cL I>L ........... ,. iii --_ ...... _------ .. --------- --_ ... ----~--". ~ • Jo,'
o
./
156
antfbo4y response illustrated by the first peak wa~ appro.xi
mately one-third o( that observed in the second peak. ,
These results suggest that the first phase of competence
in the recipient an~mals was probably due to the transferred
cells themselves. The second, higher peak of ,plaque-forming .....
cells may have been due to the progeny of donor cells, or"to
a population that included many of the recipient's own cells.
On the basis of ·these results and conclusion, it was decided
to immunize reconstituted recipients 6 days following cell
transfer in 'the study of the competence of the spleen of
infected animals that follows in Experiment 7A.
EXp!riment 7: The immunocompetence of the spleen of infec,ted" A/J mice. A. The Assay of competence 6 days
. after cell t»ansfe,r _>
Figure 6 i~lustrates the PFC response of lethally irrÀdi-
ated normal recipient uhce reconstituted, as in Experiment 6..,. -[
with either 2 x 10 7 spleen cells from normal uninfected donors
or with 2 x 107 spleen cells from animaIs inoculated with 100 ~
or 'soo larvae.7, 14 or 28 days previously. The recipients were
immunized with SRBC 6 days a(ter the adoptive transfer of cells.
The results (Figuré 6) indi~ate that spleen cells from .
animaIs inoculated with 100 larvae 7 days previously were just
as able as normal cells to\~econstitute immunologic.l competence .
to the T-dependent SRBC antigen in lethally irradiated recipienta •
. Hove~r, 14 days followinq inoculation vith 100 larv.e, ,apleen
Page 77
1 1 1 .1.
1 ! -.... dl
., l ,
, 1
'1
~
•
1 -
. , Figure' 6. The P?C -response of irraaiated recipients
reconstituted wi~h ; x 107 'normal spleen cells or, - 2 x 107- spleen cells from, mice inoeulated witk 100 or -'. 500 larvae 7, 14 or 28 days previously. The sasc -
antigen was given 6 ~ay~ after cell transfer. Significance at the five percent lavel ls.denoted by the asterisk,
" . the number of animaIs in each grqup is found in brackets, and each bar represents one standard error.
, 0
. \
t:'.J ,"
0,
,
~ '.
:'
.," •
Page 78
" ~
"
~ ,~.
l: ~ ,. 'rr
l ; ~
~ \ i 1
< ,
.' .j 1
'1
0
....
..
N •
o o
Ut o o
n
-8
S 0
n
o o
Ut g
ft
o
c
N o ,
f-t ..
-
-~
.. o T
-= =
PFe 1 s-x .. o 1
.
-, . ....,
ta o 1
,
'"
CELLS
o o 1
'H; "'-J
.
..... < ....,
::::;::: ;::: ::::;:;:;::::: ::;:.:.:.:::::.:.::: :!:!::::::::::::::::::::::::: =:::;:::;:;:;:;
N o 1
D
, {
0
.. o 1
. J
~~~~~~~~~;~;~;~;~;~;~;~;~;~;~;mm~mmm~m~;~;;;~~~;~~~;~~~~~~~~~~~~~f~~~~~~~;~ ......
--------------~ ~
."
"'"' ..... ----... 1~
-~ --.
",
n f'II rra
51
ft o • '" ~ III Z n III
.... '-j.,
.. -
Page 79
,
"
" /
,--_. --~_. ---58
cells of the infe~,ed mice were significantly less competent in
restoring the anti-SRBC response in deprived recipients th an -
were normal spleen cells (p<O.OS). By the 28th day of the
infection, the abili ty of the spleen ceUs from animaIs ino-. - , culated with 100 larvae ta reconstitute competence in i'rradiated
recipients was again equal to that ~f normal splèen cells. oc
On the other hand, although splenocyt~s from animaIs
inoculated,with 500 larvae showeda trend ta an enhanced capacity
for reconstit~tionat 7 days of the infection, and a reduced
capapity for restoration of an anti-SRBC response i~ irradiated
mice 14 and 28 days after inoculation, no s'tatistically signi
ficant dtfferences were demonstrated in these cases.
B. The assay of competence (of spleen cells) 30
days after cell ;transfer. il u
'Theoprevious experiment (7A) has shown that the ~doptively
transferred cells from infected animaIs show sorne evidence of
damage from the infection which reduces their abili ty to restorlf .
immunological competence in deprived syngeneic rec~pients. An
experiment was then done to determine ,the effect of infection
wi th !. spiralis on the competence of spleen cells "30 days after ,"
adoptive ,transfer when, "presumably, there has been a prolifera- "
tion of' the initial transferred population 'and/or a flrecr~it~
ment8l ' from the tecipient' s l~phocytes. As in the previous .'
e~periment (7~), lethally. irradiated recipients were reconsti
tuted with 2 x' 107 spleen cella from eithèr noraal uninfected
o ,.
Page 80
a
... :;, S 1
,10
" . ~----~
" .
-/
:1-
Figure 7. The me an PFC response of irradiated ~ecipients reconstituted with 2 X=10 7 spleen cel1s from normal donors
" III ...
or from donors inoculated wi~lt 500 la~vae of T .sRiralis . 14 days or 30 days previously. Immuni~ation with SRBC .
was dohe ,30 days after celf transfer. Figures within parentheses indlcate the a'urnber of an'imals in each qroup, and each bar indicates one standard error. Slqnificance. at the five percent ',level ls shawn by the asteriskl' ...
- ,
" -, 0
"
,0
..
Page 81
.'
. ,
(
J
in 1
• ","",1
'~ ; l
o ..
", , , . '.\
/
" ,
.' "
l . .
• '.
, ,
4000
1000
2000
1'000 • -i c 0' • • 1
·0 -, X C)
,2000
·u .... A,
-.
1000
0,
î '
. ,
_, -.61!""
:< SPLIgtt. COMPETENCE
1 '.
CELL TRANSFER
(II) • T ~.
(-, ) --
tftftct.d ' . ~
(15)
.... tformal
Normal
" .
, i 59 • 0
. (
o day 14 of in,flction
' ..
'" day 30 of i"'.ction 11
, '
../' .'
Page 82
i ~ j
,()
, u
, ,
- 60
donors, or fram mice inoculated with 500 !.spiralis larvae
14 or 30 days previously. The 1 x 108 ~~c 'antigen immuniza
tion was given one month following cell transfer~d immunolo-
gical competence, was measured by the PFC Assay. 1
The reaul ta - showh in., Figure 7 indicate that spleen cells>
taken from animaIs 14 days ,after an inoculation of 500 larvae . .
have a significantly enhanced tpapacity to restore a PFC' res-e
ponse to SRBC in irradiated reçipients '~30 days after tI'ansfer
in to depr i ved syngeneic recipients (p < 0 • O~). sp~een pelIs
from animaIs infected for 30 days, however, are not signifi
cantly better than normal cells in restoring competence, when
the PFC, responsë is determined 30 days after ceU transfer.
C. - The Assay of competence (Of spleen ce~ls) 30
day~ after cell transfer. The effect of 0
thymectomy on' the immunocompetence of spleen .-...-'
cells. ,
In "the previous_ adoptive transfers of spleen cells
(Experiments 7A and 7B) l, thymectomy was not perfo,rmed in the ~
recipient prior to lethal irradiation.\ Spleen cells vere
transplanted into thymectomized, l~thally 'lrradiate~ recipients '. .
to determine whether the presence of the thymus had Any ef,fect • 6
on the number ~1raque-forming cells produced in the recipient 7 • "
after immuniz~tion with SRBC •. Five thymecto~ized, ir~adiated
. " . 7 recipients were given· 1. x 10 normal Il'pleen cells and severt
similarly treated recipients vere qiven l x 10? sple~n cells ", ,
trom donots inoeulàted vith 100 Trichinella spiralia larvae
..
\. J. --
" . , . 1
Page 83
1· ·1 ~
·1 ·1 , • i .
1
o
fi ' , ",'
~I
..
.'
. r
•
Figure 8. Mean PFC'response of thymeetomized, irradiated
recipients, to SRBC' 30 "daysfJafter cel1 transfer of either 7' ,Q.
1 x 10 normal' spleen cells or the sarne number of spleen
,cells fro~ do~oros iftoculated wi ~h lo~arva~ '14 dâys previously. The number of animaIs in' ~ach group are
within pare"ntheses, and each bar ~epresents one standard
error.
)'':-
0'
-'\1-.'
" , o
D , ,
o
" .
. . -'
Ut •
\ \) ~
. .
. " • ~ "T, j ... ~ • ~'y '. ,.;
. '., . .. ;.;};'§.-è~~;;.·.i~;~~,;i/(
.;
Page 84
~ ,
-"
-~-_._,---"-,,, - --_ ... _,.._-_. ----
o
/
'" 50
• 40 --1 1 "1 30
C8' 0
te 1ft 20
" u t
o . , o .
-. , '
----~----.----~-----,- --. -
(1)
mIEN ,COMPETENCI
CELL TRANSfEB
r
(S)
. , fnltcted
..
)
: ... ~--~
"-..
"
-. ,
Page 85
:>
1 1 1
'.'
) ~-_ ... _--~--_ .. _ .~~ ______ ---..L~~_~_~~-_
14 days previous1y. Immunization wi th S,RBe was done one month
fo1lowing cell transfer and immuno1ogical competence, was' , measured in the recipients by the PFC assay.
62
1 Although it appears from th'e results presented in Figure 8
that the PFe response in recipie~ts which received spleen cells
from mice "infected for 14 days wi th' 100 larvae was enhanced in
comparison to that by normal spleen cells, no statistically'
significant difference couldo-be demonstrated • . ' ,....,
D. The assay of competence (of spleen cells) 30
days after ce11 transfer. The effect of
adoptive spleen cell number on the immuno
competence.
In the previous adoptive transfers ofospleen cells
(Experiments 7A, 7B and 7e), 1 x 107 and 2 x 107 spleen cells
vere us_ed. It was of interest to determine. -whether the
demonstration of competence would be determined by the number <
of spleen cells used in the adoptive transfèr. For this purpose, , 1
. , ,
ten thymectomized, lethally irradiated,recipients receive~ ,J
o 5 x 107 spleen cells from normal uninfected donors and an E{qual "
. number, received spleE7n cells 'fram mice which ·h~d been inoculated
with 500 !.sP:lràlis.. larva~ 14 day~ previously. The recipients
were i~unized with SRBC 30 days after C?ell transfer ànd
antibody ~roduction w~s measur~d by the PFC assay.
" From the results presented in Figure 9, recipi~ntà
reconstituteô vlth 5 x 107 .pl .... cella fra- IÛce 14 daY8' ~fter
"
Page 86
,1
-,
, .,
~ r /
\.
)
Figure 9. Mean PFC reapon$e of thymectomized, irradiat~d reçipients teceiving 5 ~ 101 spleen cells, from eith,r normal animaIs or trom animals inoculated with 500 larvae • 14 days previously. SRBC antiqen w~s given one month after cell transfer. Bach bar represents one standard' error, and th~' number of ani~ls in ea~h group is within
J' the parenthes.es.
, ' <>
1
" ,
"
, ,''-
, '. "
. , •••
Page 87
r
t C>
.1 t'
"
1
.400
.200
t '" rt,
\ 1000 l "
• -.. u
, 1 eoo ~ li )
1
" 600 -,'.
)C "- .,.,
"-,U 400 "-.. 200 "
~
o ...
. ;.-
. , . , ."
- " .,. ~ -~----"--'--~ ... - ~ -,
SPLEEN COMPETENΠ-CELL TRANSFER
't
,
, . 2.) .... -
, ,
(1-)
0
,
. filllll 1111111
~
'.11111
, 111111
. -:
. /
Infect'" 0 .....
1D l,
. ,
0
/
,
.
.-' ..
, .
"
}
. ... ,
~(r'~ "2-. 1
.
!J
-r
"
0
63
1
"
, , '
Page 88
--------------- ---~~ --
1
. .1
'. , . v __ _ . ____ .. - -~ ... -----_ .... _-~,-~- --. --_ .. _ .... -
64
inoculation with 500 larvae appe~r to respond less weIl to
" SRaC than thos~ recipients reconsti tuted wi th the -same npmber
of normal spleen cells 30 da.ys after, cell transfér. "However, no
aignificant difference' could be demonstrated.
E. The assay of competence (of spleen cells)
.measu~ed by primary stimuI,ation wt th sheep
erythrocytes in v1tro in Marbrook c~ambers. 6
A final study of the competence of spleen cells' from .. '.
infected animaIs jas done in the Marbrook chamber !.!!: vitro
culture system. 5 x 106 ., spleen cella from mice inoculated 1
with . ei ther 100 or 500 Trichinella spiralis larvae 7, 14 or -
28 days previously, ·were incubated wi th the SRaC antigen; four
days after incubatiO{1 at 370 C in an atmosph~re of 5% CO2 ;'
the number of plaque ... fGJrming cells was-,-'_~4etermined. Spleen
cells fram nomal uninfected mice $erved as con troIs • Each
assay w~s repli,ca ted three to eight times, depending on the
·nu.mber of cells available. b '\
l " ...
The resulta of this experiment presented in Figure 10~
dernonstràte a significant marked suppressio~ of the ·n~er of
"p'l~que-form:J.ng cella to SRBC in Marbroo~ chambers containing
spleen cells from animaIs infec ted for 7 days (p < O. OS) and , . 14 days (p < 0.01) following i~9.culation with SOO !.spiralis
·larvae. No significa~t differences were dernonstrable between
thoae cultures which c06t,ined spleen cells from anima~s ino-\..
cu~ated with 100 larvae and control culture. of spleen cella
froa normal uninfectéd .Iliee. , iii
. ,
"
Page 89
" 1 !
·1' .r :P
\ .. t1
i ~. J 1
~
1 /
!
I~
Figure 10. The mean plaque--forming cell response in vi,tro' of '
normal cellS and cells"from animais'inoculated ~i~100 or 500 lar':ae 1 d8j'S, 14 days or 28 days previously.
'The double asterisk denotes a highly s(gni.ficant . . difference (P(O.Ol) betwee.n the experimental and the'
• control group. The PFe response ls reporte4 as a per-centaqe of the control .One standard error i9 sh'own' by . the bars,. and the number of rep1icates for eacl:t. histo-
gram ar~ shawn in pa~entheses.
o ".
-,
, .
, .
•
. \
J
o
Page 90
, \ .---_______ • ________ 1
0 ----- - PFe
'- IN
lit ... Gt 0 0 0
of
il
-0 0 QI
'., ""8 n
:t • < "" - 4 0 ",.. ~l ,. a '1:,·
.If • 0 l, , Z Ut
~- "-8 ...... (') -4 n
'i ~-0
, ;;. -g ,;; w
:1 ,
n
" " 0
8-· . ,
..
" (~:..; , ,.,'i - _ .... -.... --' - ~ -- -... -
6 .
1 , X 10 CELLS PERCENT
• 0 fil) 0 0 0
..... ft -
:~ ....... ~
.. , --_._--_._---
----
-------
-._.,. 0
"" W' ..,;
-..fa --• • :II •• :II • o· 0-~
n
l' :1
~ <>
..,., J -
~--
--------------------------~----- ---~--
"
"
,
Page 91
•
t '/1 ' ' i !
e •
..
, DISCUSSIQN
. '
In recent years the phenomenon of immunoffbppression has ,J
been weIl documented in both protozoal an~ helminth infections ')
(Clinton et al, 19691 Greenwood et al, 1971; Sengers et al,
1971; Mansfield and Wallace, 1974; Shimp ~ al, 1975; Crandall
and Crandall,' 1976; Hudson ~t al, 1976; Araujo et al, 1977: -- ..,...--!'
66
Brown'~ a~, 1977: Reed et !l, 1977: Stric~land and Say1es, 1977) • .. In trichine11osis, immunosuppression has been.demonstrated by
o an enhanced susceptibility to virus infections (Kilham and ., Oli~r,. ~96l: Wa1ton and Gleiser, 1966; Cypess et al,\ 1973; • Lubiniecki et al, 1974a; Ghimishkyan and Ovumyan, 1975), by a
-, -pro1onged survival o"f allografts (Svet-Moldavsky et laI," 1969,
" --. . 7 ' 1970; Chernyakhovskaya et al, 1972; Faubert and Tanner, 1975:
'" CI J - • .., ~
Chimishkyan ~ al, 1916: LjungstrOm, 1976: LjungstrOm and Huldt,
1977), and by ~he impairment of humoral and cell-mediated
immun' responses'~o various antigens (Faubert and' Tanner, 1971;
che~nyakhovskaya et al', 1972; Cypess et al, 1973, 19~4b; Chimish
kyan et al, 19761 Faubèrt and Tanner, 1974a: Lubiniecki et al, -- ,. " 1974a; Barriga, 1~75; Chimishkyan and Ovumy~, 1975; Lubiniecki
and Cypess, 1975a, 1975b: Faubert,. 1976; Jones ~ a1, 1976~ ,
LjungstrOm and Huldt, 1977). - In spite of the fact' that in'fected
animaIs are less able to respond to hetero1ogous immunogens,
humoral and cel1-mediated immune rcsponses are observ~d during
T.spira1is infections (Baehman, 1929; Kagan and Bargai, 1956;
KarmansJéla and Ko,ar, 1962; Crandall et !!, 1967, Crandall. and
....
~ . \ 1\ ,
.
" ~, :::
.'
Page 92
1 (
! f \
" 1 !i:'
~: ~ 1
-,
'----------- " -______ J-..-.~_ .. __ _
• ... f,",;
Moore, 1968; Cranda1l 'and Crandall, 1972; Ruitenberg and , \ 0
Duyzings, 1972; Ottesen et ~l, ~97S;. Vernes et ai, 1975b;
Ltlung~trOm and Ruitenberg, 1976; P~rrudet-Badoux et al~ 1976; . , --Matossi~n !! al" 1977; Tanner et al, in preparation). However,
despite these responses elicited in the host by the parasite,
Trichinella spira1is sti1'l survives in the, infected animal. The
paradoxical phenomena of irnmunosuppression 'and irnrnunological
response is probab1y a ba1anced mechanism which permits the
survi val of T .spira1is, and the primacy of one or the other
response determines the fate and the ïn~ity of the infection. 1(
'The,purpose of this study vas t9 'in~stigate'the affect'of .
infections with !.spira1is on the immunocompetence of the
primary and secondary lymphoid organs of mice, to determine ,
67
where an immunological defect could be lo'cated which 1eads to ~." .
imrnunqsuppression. In addition, i t was of interest to determine
whether immunosuppression is a function of, and determined by,
the presence of the parasite, or whether it is due to a .li
prolonged effect due to a damage by the' parasite of the lymphoid
organs of hosto The effects of different doses of inoculation
on' the phenomenon of immunosuppression 'and the response at
different stages of the infection' were a1so sub~ects of this
investigation. U1timate1y, it was hoped tQ obtain'some insight
~ 'the mechanism which induces immunosuppress~on in trichinel- 0
losis. The competence- of the lymphoid cells of infeeted mica
was determined by their abLlit~ ~éconstitutc immunologieal
competence after adoptive tr~nsfer into immunologically unreapon-
Page 93
., Q • ~_·,"_tif __________________ ~ __________ __ . ' - ---~---t-~-~-"'-"''''-''·''''
o
()
/
sive syngeneic ~/J mice.
'c:,
The results of the· preliminary study of th~, level of ~ ,
infection pfoduced, by different inoculating doses of Trichinella
spiralis in syngeneic A/J mice dëm?nstrated that the number 1 •
of muscle larvae recovered increased in direct relation to "
the inct:ease in the 'inoculating dose '(Figure 1). These results
;ponfirm those obtained in a similar study done in Swiss albino
;' mice by Tanner ~ al (l978). In,comparing our results to those
of Tanner et al (1~78), A)J mice appear to be mor~ susceptible 0- -
to infection with T.spiralis, since more muscle larvae were
recovered from A/J rmice than from Swiss mice after th~ame
'inoculating doses up tq a dose of 750 larvae. Beyond tl1at dose,
the slope of the 'recovery of muscle lc)rvae from Swiss mice
(Tann,er et al, 1978) increases markedly, whereas the slope of
68
the recovery from A/J "mice fell after an inoculum of 1000 larvae.
Probably,'becau~e they were more suscept~ble to infection, 40%
of the A/J mice died in this last group of animaIs, most of
tQe animaIs dying?about JO'days after inoculation. The flat-
..---' tening of the recovery curve of the A/J mice was very likely , (
due to the 1055 of a good proportion of the an~mals inoculated
wi th l'OOO larvae.· While considerable mortali ty was observed
in A/J mice'after inoculation with the higher doses, no such , •
mortality was observed in Swiss mi~e after inoculatioR (Tanner
!!. al, 19·78-). The greater suscept~bility of A/J mice to infec- /
t;ion with !.spiralia and the differen~es in the latterDpart of ,
the dose re8ponse curve between A/J mi~ and SViss lIice IÙght
l '
\
"'"'
Page 94
l
• f
, "
v
--~------- ----~- _:. --.. -- -- ______ • _~. _..,.--..:~ k
be explained by genetic differences between the two strains. ,
, For the purpose of further experimentl; a low dose of'
100 Iarvae and a moderate dose of 500 Ulrvae we're used!, 'A1til
hhe larvae ;ecovèr~d'fOi~ing inoculation of 250 lar~ae an~ 500 1arvae were not statistical1y different, the latter dOS~ was qhosen so that the Most intense immunepatho1ogica1 respè " . ()
could be e1icited wïth the' least possible i.njury te the m~" s
hosto The study of thé; response of A/J mice infect~d With:,1
Trichinella spiralis to immunizatibn with sheep erythrocyte
demonstrated an impai~nt and a marked reduction in the
responsiveness of mice (Figure 2)~ in' confirmation of the 1
J 1
" of other workers (Fâubert and Tanner, 1971, 1974a; Barriga,,~
Cyeess , 1975b; Chimishkyan ~ al, -197~1~ et al, 1976; Lfun9st~m and Huldt, l"
1975: Lubiniecki and d '1
Faubert~ 1976; Jones
Immunos~ppresslon to :-T-dependent a,.nti9~~ has' been again. ~ demonstrated' and this 'study confirms reports of immunosupp
" occurring at the cell leve1, measured by the p1aque-for .. u.u .... ",.
cells (PFC) ~ssay (Lqbiniecki and ·Cypess,' 197~b: Ch~mis
et al, ]976; Faubert, 1976; Jones et al, 1976: Ljûogstr&m ,
Huldt, 1977). Às no anti-immunoglobu1in serum was used'in
PFC assay, direct (IgM) plaques were ctetected~ confirming ~Q ,~ •
v
results of others (Lubinrècki and Cypess, 1975ta; Faubel:t, ~
Jones .=.! al, 01976; Ljungstr8m and Huldt" 1977). The 1
of" this study also agree ·vith ear1ier repor~s that
pression is a tràns-i-ent phenomenon (Faubert, 1916;" Ljunqs . ~ .. . . , o. and Huldt, 1977), and that ~n is 'lnainly correlated with
)
Page 95
, 1
"
.'
r
( ~"'~,.. ........... --.. _ ... ---~... .. ~ ... ,
. . "\ migrat~ng phase of inf~etion (Faubert, 1976) sinee the maximum
impairrnen t of the respçmse to sheep {erythrocytes 'occurs on' .the
fourteent:h day''pf the infection.
Most of the workers who have reported the immunosuppreB8io~
, of' the ~mti-sheep erythrocyte re.§ponse -have used an inoculating
,~~.'do8e of 200 '!.SPiraliS 'larv:"e ,or' more (Faubert and ~anner. 197i.
,0 ô 1974'a1 "Lubiniecki and Cypess, 197~b; Chimishkyan!!::. !l', 1976;
F~ubert, 1976; Jones et al, 1976; Ljungstr6m and Hu1dt, 1977). tJ
In our study, however, inocu1a~ion' with as few aB 100 larvae
resulted in a s~gn1f1cant (p< 0.(5) reduction in the r~spori
slveness of mice onRthe fourteenth day of the infection.
Significant suppression of antibody response in ipfected mice . '
W8S not demqnstrable earlier t'han 20 da ys after the inoculation . ,
qf 200 or 500 1arvae in the studies by Jones" et al (1976) 'ant
Ljun9strÔ~ a~d H\J.1dt ("1977), ~respectiv~lY. 'I;~;: study .we ! '" obtained S19~i'fi~~n,." (p (O. OS) immunosuppression as early as
the seven,th day 0o/the infection after. inocu1~tinCJ the moderate f
-dose of 500 larvae. Our observations are in agreement.. with
those of Faubert (197~) and Lubiniecki and Cypess (1974a) who J '
',demonstrated signiflcant suppre~sion of antibody response a't
14 çiays o,f infection. T~e results of this study de)n~nstrate '\
tha t th~ 'higher dose of larvae prodUces tin ~ ear lier 'evidence
of the immunoauppreased state (Figure 2). An~ interest'tng
point ia that the inténaity of immunoauppression obtalned was
not correlated wi th theJ leve1 of ~ec~tion, the re~uction of
blood ceU aritibOd~ responsivenesl was Itati,atical1y anti-st\eep red "
'.
"k
1 l ,
Page 96
=====::::::.......;...;:.:..;:,.-,----==~...:;..----------·-------· .. T .. --;----·----'--,f'-~-;---------------
1
1 , 1
1 ~
/ \
} 6 " • fi
,', i
() ,
,.
, . .. ~
72
Adul t thymectomy> folo1owed by letbal total body irradia
tion, ~BsentiallY el"iminates lnost of the lymphoid êells in ani
maIs, and, thus, severely impairs immunoloqical responsiveness a ~
(Miller et al, 19611 Arnaaon !! !!, 1~62; Cro •• ~ al, 1964" '\ ,
Humphrey et al, 196411 Fahey et al" 1965) • "-Mitchell and Miller . -- --(i968) showed tha't aduft ,thymectomized and irradiated mi~e had ~
to be repopulated with bo~h thymus and bone marrow cells before ,J •
1
they could produce a reeponse ta sheep red b100d cells (S·RBC).
Thymus and bone marrow éells transferred into the sarne recipi~
ent Act synergistically in the restoration of the immune respan
siveness to SRBC (Claman!1 al, 1966a), and this response.is
lin!i!arly yrCl.ated tQ :?e number of thymus ana bone marrow cells
in the mixture (Claman ~ al, 1966b). 1 Furthermore, the full
reatoration 'of ar antibody response to SRSC in immunologically
unresponsive mice requires viable, intact T-cells (Mitchell and
Miller, 196~r. The""results of our study on th~ reconatit4tion
of immunologically-depri ved mice wi th thymus and bone marrow , L
cells support tltose of Claman ~ al (1966b) and Mitchell and
Miller. (1968) since the greatest antibody response to SRaC was
observed in those récipients reconst!tuted with' the hiqhest
ratio Qf thylnuS cells to bone marrow cells (Table 1).
The study of the immunocompetence of the thymus cells of .. infected animaIs (Figure 3) demonstrates that thymus celle from
animals infected vith 500 larvae for 14 days we~e eonaiderably , ~
less competent than normal cells (p<.O.05) in reatQrinq an ' . .
antibody reaponse ,to SRBC in immunoloqic:ally'depr1vecl Cthymecto-
Page 97
, .
mized irradiated') mice' wben transferred simultaneously with
normal bone marr~w cells,. The infection with !.spiralis may
have 'l5uppressed the helper or antigen-bindinq funct~on of the . . \ '
thymua cell it.elf or the proliferation of thymocyt •• or the
ability of thymus cells to cooperate with macrophage à and ~
B- èe1ls in the induction of an P antibody reaponse. A proq-
73
<l ,.
ressi ve -109s of thymus weight and thymocytes has been demonstra-1
ted during infection with !.spiralis in mice (Ljung,strOm, 1976,'
Ljungs.tIlBm and Hu1dt, 1917r Tanner ~ al, 1979). Histoloqical, ,"
examination of the thymus of infected animals shows a cortical . . , .
depletioh of thymocytes (Ljungstr6m, 1976; Ljungstr6m and Huldt,
1977) accompanied by a siqnificant' 10s8 of t'heta antigen-bearinq 1>
cells t'rom t~e ~gan and a reduced ability of thyrnocytea to
respond to Con A mitog~n (Tanner!i!l, 1979). These observa- ,
• tions indicate that the thymus loses potentia11y immunocompetent
T-ce11s during the course of infection and, thus, an inab!lity
of the thymus of infected mice to provide enough T-cel1 helper 1
functi~n, may simply reflect the loss of immunocompetent celt.
from the thymus, rather than sorne effect of !.spiralis infection
on T-cell function.
/ 1
Faubert and Tanner (1974a) reportea. that bone marrow cella
from mica infected with 500 Trichinella.spirâlis larvae for 30
days were considerably less efficient than nor"mal cells in . ' , '
re"toring the humoral re.ponse of immunologically deprived
.yngeneic' animale to SIBC. Moreover, \he ~eneic bone marrow
Crom infected animale vas, dso probably Vneffi~i.nt in re8~oring
. - ~
Page 98
..
J
1
1.
1.
,-
f
V 6
J. "
~ < il -,J
, '1
. '" - . ~" ': " ,l u' ") ... '" w' ~ \ t, (~, J
, •• 1 . ," . , • ,', • ", • 1
....... __ /_~ ~_ w~ ........ .JO! ___ -.,. ' .. ~ .. _ .. _-,-..--~ ........ ~~~--~",,-:
hematopoietic function sinee a considérable n~ber of animala
reatored wi th (rells' from infected animale died during the
~our8e .of the' experiment. 'l'he bon~ marrow ls not fn1Y ~he
. souroe of. immunocompetent' and hematopJ.t1cltem caUI. (AbcSou , ' l , "
and Richter, 1970) but a180 the site of the' production and
74
differentiation of newly:.forme<l surface immunogloD':!lin-bearing,
lymphocytes (Osmond ~nd Nossal, 1~741 Ryser and vassallifj197 . • 5{ f
A. study was inltiated to re-examine the e'fféets of linfec io~ with T.spiralis on the inununocomp~tence of the bone marro
using' the thymu~-independent ! .. ~ lipopolysacchari~e (LPS)
antigen (Andersson and Blomgren, 1971). Inoculation Of A/J "
mice with 100 larvae had no effect On the capacibY., of bone
'~row cells to ,estore respons:L,veness to LPS in immuno1ogically-. ,
deprived (thymectomized, irradiated) miee. A significantly l '
(~<O.05) enhanced abild.ty to reato,re a1)' anti-LPS reaponse " .
was demonstrated ~y bone mar~ow cellii--~imals 7 daY8 a,fter
inoculation with_SOO larvae (Figure 4). These reSUltB are in 1
genera1 aqreement with tho8e of Barriga (197S~, Chimiahkyan et Q
.!! (1976) and LjungstrM and Huldt' (1977) ," who showed that
"
i~ctions with :t.spirali! had little effect on t~e humoral
re"'nae ta thymua-indepandènt \'ntigena sucb aa PVP and LPS.
, i ,
& significantly enhanced reaponle of bone marrow calls to "LPS
i~nimala inoculated with moderate' doses of Trichinell", apira'lls
has not béen 'demonatrated previously and a~qgeât* that rela
tively heavy in(ect.iona may activate the immunologieal potential
.of bone ~rrow ce 17;1. 'rhe mort. lit y reported by Faubert: and. _'*
'l'anner (l9Ha) .... not _ ..... 4 in thb Itudy, perh.p~ r .. \.
1
Page 99
"
/,1
-,.
. :" _UIf_~,,_J_. __ .................. _ ..... _ ~ _ ............ _. ______ ,
" ,0
)t
l "
" a }arqer nwnbér of bone marrow celle were 'used for reconsti
tution in th~s investigation. The 'resultB of the bone marrow
competence study ~ndioate that t~e parasite has a differ.nt
effeet on the immunologic.l function of bone marrow cell.,
depending on the level of infection. It May be that, at the
higher ',level of. infect~ ! .. spiralis may have a mitogenic ,
effeot on the bone marrow (the proliferation of thls tissue
dur!n; trlchineliosis has never b en studied). ,It ls possible
75
that the enhanced respons!veness thymus':' independen t an tigens o
May contribute to the mechanism of nosuppression observed q.
tn the parasitized animal by, perhaps, enhancing m.chanis~s
such as antigenic competition·'(Tanner; 1974) a?d defleeting the
inununological response away fram the "funetional" antigens of
the parasite. . \
,The next organ stud!ed to determine whether infeotion
with !.spiralis affects its immunologieal comp~tenee was the
spleen, a secondary lymphoid"organ., Impairment of the're.ponse , .
of infecte.d mica to the T-dependent antigen, SRaC, -hild been
demonstrated in spleàn cel1s by the direct plaque-forming· oéll r
assay (Figure 2). Tanner!!!! (1978) have dem~nstrated that,
concomitant with,the 108S of leucocytes and theta antigen
bearing,cells from the thymus, there is an inorease in the -
number of,cells bearing theetheta Marker in the spleen in mice " -- ,~ ,
infected wi~h !.spiralis. 'However, t~mmunooompetenee of the p . ' "
spleen cel18 ftom i~feeted animals was ques~ionable since thes • ., \ \. ,
cel,la w,re incapable'\ of re.ponding to mi togenio atimulation by
Page 100
o \
\
L,. 1
.J . . , J'l ,
l '
\ 0
"
.i
"
~, " f
} ,
76
Con A (Jonua et al, .l976a Tanner-et al, 1978) and PRA (Jones et -...... --,- ,--. !l, 1976). Spleen cella in infections indueed wit~ 500 larv~e.
1 •
showed an i~palrment',in lI'\itogenic reaponae to LPS (Tanner !! a~,
. 1978), wh.r.a. a nonnal re.ponse hal be.n demonltrated by oella , .
front anima~B inoculated with 200 larvae (Jones ~ !l, '1976). On
t.he other hand, in infèot.iona obt.ained aft.er inoculation with ~ \
100 !.apirali's larvae, spleen cells demonstrated an inore'ased
oapaeity to reapond to l)oth LPS and C~n A (Tannér et al, 1978) • .. ~ " --
Chimishkyan et al (1976) have reported that the spleen cella of , --
(
miee inoculated with 200 larvae are .!h~rply suppre,ssed in their
. eapacity t.O induee a GVH reaction.
In an" adoptivè ceU transfer system, spleen cells from Jl
mice inoculated witht
100 Trichinell. spiralis larvae 14 da}\a
previously were ~ignificantly (p< O.OS) less ~ompetent' than. ~ .
normal spleen cells in r~storin9 t~e immunolQqical response 1 1 ( • l'
to SRBC in immunologically unresponsive mice (Figure 6). This
o~servation is in agreement with that observed in the intact . \.. ' , \
animal infected \tIith the same number of !.spiralis larvee and -
immunized with SRBC (Figure 2). However, at the hiqher level cl. ~ - 1, 1
of infection obtained after inoculati'ng 500 laivae no signifi-
cant differences \tIere shown in thé reconstitutive ability of
cells from normal or infected mice. A trend for an enhanced
eapacit.y of spleen cells from mice infected for 7 days with 500 /
lar,vae to transfer 'competence to SRaC and' a trend for a auppre •• ed
ability to tran.fer compet~nce to BRBC af~er 14 and 28 days of. L
"infection were' evident (Figure 6)... The }\ab.ence of si9l\ificant ~
Page 101
J ' ,
JlJ r i:
~ /
, ,
" .", l,i ': ./,:~~e,\' :', '~, ,.', ;}~ ;~~ Z:tri:L~~~" .~~~,:.,:"
"
" ,~", ........ -..... ...... ~ ... ,~' --~ "" ' \ 77
\ li diffe~enoea in the level of rec~n.titUtion afteÎ the tranafer of
< J '
these,latter spl!en cella ia due to the large variation obtained
in the anti ... SRBC antibody reapon ef ofethe control groupa (which
reaeived no~.l cell. when the ex arimental ~roup. vere 'tran.fer
red wi th cells from day 14 and 28 days of infection), "'and in the
experimental group of' dey 7. It 1 uncl~er how ie could be that
SPlen~CYte~ rep,~rted' to be ~nable 1 tOe r~spond tq mi togen (Tanner "'" ... [ !!.!l, 1978,) after t~e ,inocull.tior o~ mic~ with 500 larvae, could
be immunocompetent in a_rj!lBP--'IDSaJo a spe(:ific ant,igen, particu-, ,
larly one that requires the cooperation of different cell Bub-" . ,
populations (Claman !1!l, 1966a; Mitchell and Miller, 1968)
unless response to mitogena 1a not al1laya related to immunologi-. ,
cal competence. The resulta of this experiment aljO'su9gest ~ , t ~ ,
that immunosuppression ia not directly related to the presenoe , '
,)Î0f' the.rpàr~~it~ since the' effect that 'the- parasite' has on the
-\ e immunocol'llpetence of the spleen cells remains, aft~r th~~ls , ' ,i" t
e.~ • {
ha~ been rémoved to a "cléan" ,env1ronrnent for at least 10 aays
(Figure 6). A p~t of interest is. that a trend tow~rds an 1
enhanced response to SRaC of spleen cells taken on the 7th day )
after inoculat.ing 500 larvae (rigur~ 6), was paJallei, t.Ve
enhanced respon~e of bone marrow cells at the same ttme of infec-e
J '
tion after the inoêulatl-on of the ',~ame 1
J '
number of larvae "(FigUre{,
'1 fr.om mice infected ·for 14 da ys after te, -Spleen cells taJçen
inoculation of 500 larvae and immUnized with~SRBC-.re immunoaup-.' \1 ..
---presÎed in the reapon.~ to the erythrocyte antiqen (Piqure 2) and
th.là cel~8 a180 Ihow a trend to an inapaired capacity to. ralto1.
,
, .
, , 'N;. ,.. 1
Page 102
o
1 , ,
78
. , itnmune respollJiven.se 1 ~o' BRaC 10 daya after adoptive tranÎfer
" ,
(Figur~ 6). ' Spleen: ct118 fram 8ii1î~i~riY infected mlc. show _ l "" 1 l'if""./
an enhanced ability td, restore immunologiçal c;:ompetence ta SRBC ~ , :}
(p< 0.05)" when· the itradiated recJ.pient. were te.te4 on. mon.~h _ -.../~
after cell transfer (F.igure 7) ~ Recip1en~a of spleen' cella from '
m~ce infected for 30 days" after inoculation witp 500 larvae, on
the other h,and, responded normally to immunization with SRBC one " 1
mOrY~h after ceU transfer (Figure '7). The results of ~i9ure 7 .' ~ ,
confirm that immunosupp~ession ia a transient phenomenon (Faubert~ , '
1976, Ljungstrem and HUldt, 1977: Figure 2) and that cells that·
behaved as inununosuppréssed cella probably eventually reaume
their normal function. The resulta (Fi9ur~ 7) a1ao seem to ./
indicate that sorne polyclonal activation \ is present. "
In th~ experimel'\t which ia illustrated by Figure 7, the 1
. thymus wes not removed priar to irradiation. Experimenta by
Radiah and Basch (1975) indicated the existence of 4' pool of .
l ,
intrathymic radioresistant precursor cells which ~re capable of
partial1y restorinq t~ymus fu~tion in lethally irX'adiated ani .. " . ~
mals. In our experiments the recipients, recuperated for one 'Ii:' . ' . month -before .immunization with SRBC and the pool of radioresistant
cells ,in the recipient mlly have contributed to antibody formation , .
~y donor apleen cella fram', infected animals. Thua, it would
JJave b~en more appropriate to havaI\. hacl a control vroù,p,.of ',normal
thymectom1zed miee a. r.cipientl aubsequently, ilàlftuahed wittl-SRBC. .. ~ /-- -~--
Spleeno
bella ffam-' a-n-i-1U.-l-.-i:C~4 :f~~ 14 4ay. after Inocu-
..
..
"
, .'
Page 103
\,
\ "
(
(!' 79
" 1atioh with 100 larvae- were, thereforé, transferred into thymec
tomized, letha11y-irradiated syngeneic mice; the recipients were . }- '\ .
immunhed with SRBC 30 days after cell transf~r. A slight, but
, not. .1gn1~ioln,t, enhancemant ,of the Inti-SRaC prc r-elpon.e wla ,
demonstrated in the recipients (Figure 8)', One of the conclusions
that can be drawn from -the resu1ts <lf this experiment ls that
the lmmunosuppressed state induced by infec~ion wi th !,.spiralis
18 transitory (see above). When mice were lmmunized with sheep . . erythrocytes 6 days after the adoptive transfer of spleen cells
in a previous simil8:r study (Fiqu.re 6) the recipients were immu-
nosuppressed'. (.
The fact t.hat the tran!!,fer of spleen cella 'from anima s 14 ~ ~ ,
days after inoculation with 100 larvae into irradiated recipients ~ (
showed evidence of immunosuppression (Figure 6), whereas the same
transfer into thymeetomized recipients d*d not show suppression
suggeats that thymeeto~y may have removed suppressor cells or' a
stimulus for the differentiation of suppreasor cells in the reci
pient àhimal. It has been suggested by Jones et al (1976) that ,--suppressor cel1s may be responsible for the immunosuppressio~
observed in trichinellosis. A better way of studying for the
presence of suppressor cells in our experiments would have been
to tranafer normal spleen cells- simti1taneously with spleen cells . , . from infected mice into thymectomized irradiated recipients; ,
the antibody reaponse of the recipients would have been studied
at Ilx days to observe the -phenomenon at art ear11er period fol10-
wing the trans t'er. 1 t woule! have allo been pbsiible to have gone
Page 104
1 f
1 t
... ...... ............. - , ... .1 ... " .... _ ..... I-_~,..-_ .- b,
80 .
• > . into further detai! by analyzing th~ Band T cel! populations ."
in the spleen, taken at different times of the infection, for
their suppressor cell activity.
In our f~nal study, splenocytes from mice 7 or 14 days CI
after inoculation with 500 Trichinella spiralis larvae were
si~nificantly suppressed in their primary antibody response to
sheep erythrocytes in vitro (Figure IO). These results are in
agreement with those of Jones et al (1'976) who demonstrated ~.
suppression in vitro tO SRBC of splenocytes from mice infected
"'--- with !'..spiralis. These results also parallel the immunosuppression
observed in vivo in mice infected with the sarne dose of muscle
larvae (500 laJae: Figure '2). On the other hand, the spleno
cytes of mice inoculated with 100 larvae did not show Any inability
to responœ to sheep erythro~ytes in vitro (Figure 10), although
a trend for a decreased primar,y response to SRBC was evident in ~
cells taken on the 28th day of the infection. Thus, the Marbrook
chamber selec~ively dernonstrated in vitro suppression of 'spleen " ----
cel~ to SRBC ln the higher l'evei of infection (500 larvae) but
failed tb demonstrate suppression of spleen cells in the Iower'
levei of infection (100 larvae).
The greater part of this study was based on adoptive
transfer whereby donor cells from normal mice. and mice infected
o with !.spiralia were transplanted into letha11y irradiated'--, . recipients. Radiation renders recipients immunologically inert
,
(Taliaferro !!!!, 1964; Micklem and Loutit, 1966), and thu., it
is Assumed that the donor lymphoid cella ara responaible for the
( '.
Page 105
)
! 1
81
antibody response following immunization after cell transfer
(Cochr~ne and Dixon, 196-2'-: -- F1r th!s reason, we concluded that ....
donor cells accounted for the~irst peak of' the PFe response
in depriyed,recipients reconstituted'with normal spleen cells
(Figure 5). Therefore, we a1'80 examined the effect of the para- ,. -
site on the immunocompetence of spleen cells by immupi~inq the ,/
recipient on the sixth day after cell transfer (Figure 6).
Gradually, regeneràtion of the lymphoid cell components (which
are dep1eted after radiation) oceurs" the ,extent an~ rate of
which'are dependent on the, radiation dose (Anderson et al, 1977).
Thus, the Band T cells of the!reèipie~t which have regenerated ri".:. •
will contribute to the secono peak of competence in Figure S,
and to the competence of lymphoid cells from infected animaIs , ,
(and homo1oqous controls)'30 days ~fter cell transfer: memory
ce11s from the donor transplant may also contrj.bute to the
antibody response in thèse cases.
A numbcr ?f parasite infections have been shown to be ....... , II;
L a~companied by pronounced Lmmunosuppression, and in various
infections, different m~ehaniBms ,have been proposed to explain o
the phenoménon (Terry and Smithers, 1975; Cohen, 1976: Ogilvie
and Wilson, 1976). Our studies s~qqest .that imm~nosuppress-ion " , .. , -,'
in infections with ·!.spiralis may also be due to different
mechanisms, depending on the level of infection with-tte
paruj.te. Immunosuppression W8S observed -. in A/J mice inoculated' - , . .
with either_ 100 or 500 larvae (Figure 2). Hovever, wh.n the
cella from thea. immuno.uppre.ae~ animal. were removed trom the
Page 106
o
(
~ \.,
82 , '
parasite into other in ~ and !rr vitro systems, demonstration,
of imn\\~nos~ppr.ession 'las il (}lnction of perhap's Inherent differences , .
\ . in the Assay methoQs (Figures 6, 10). In additi.on, the spleen
cella of mice wi th 10.0 and 500 ! .,seiralis laJlWae are suppressed IJ \. , ~
, (Figure 2), an. in another report (Tanne!' et ~l, 1978) at the
lower '~evel of infection' ~[.\pleen ceu: have-aiSO BhOWljan (
enhanced susceptlbili ty to stimulat~on by mi togens, whe~eas
~t the higher l,evel of. inf~ction the 'cells are unr~sponSi ve ~ • Il
mi togen. .An enhance~ response to LPS was obtained by spleen
cells taken in this study at 7 days of
tion wi th, 500 larvae, whereas a normal ~, .. ,/"
obtained at the lower level of infection (1 larvae: Figure 4).
At the hiqher level of infection (500 larvae), the weight,of ,?
the thymus is reduced (LjungstrOm, 1976; L-jW'lgstd'm and Huldt, \
1977), the cortex is depleted~ of thymocytes (-Ljungstrem, 1976;
Ljun~strOm and Huldt, 1977) and, the number of theta-ant,igen '
béaring cells are reduced, probably through emigration to the
secondary lymphoid organs (Tanner et al, 1978). It has' beèn , ..... -
sU9gested by the latter authors that the theta cells arri ving in , .
, '\.
the spleen from the thymus of infected-,animals are insensitive o
l'.. ,
to Con A: this may suggest that, due. to an overwhelming demand , 'J
for T cells in the secondara lymphold ergans "--. thymocytes do not
have sufficient t1me to filature into immunoco~~t~nt cella ~
In A/J mice infected with !.8Eiraliâ, it appea.:.ed that, ./ ,
dependt'ng on when SRBC immuniza tion va. -91 ven, the responae' te» "
the erythrocytea vaa .e~ther enhanced or dillliniahed (Figure 2).
'l" f'
, , . -
". ,
Page 107
(
•
7 . •
...
83
6
A similar observàtion has. beeh reported by HUdson .!! al (197~)
• • in trypanosomLasis, and Capron et al (197~) hi;lve demonstrated . - - '~~ l '-~ J
that, du~ing the first two weeks of infection with S'.maRsoni, , ' - -,
the lymphocyte transformation index of" S.mansoni 'antigen was , .,. :- • t,
elevated, ,but i t was deç\eased~later. oh 'several occasions o -
in t.hese exper imen:t~, trEtnds towards enhanced responsi venes! to
SRBC and LPS were demonstra ted "in O\lr studies of adoptive transfer 1
(Figures 4, 6, 7, 8) .1
"'-... ;. ,
Background plaques to SRBè or LPS were not determined at - \ ,
any t.ime in infected, non-inununized mice in our studies. If
backgro~d pl,agues had been significant1y increased, a polyclona1 ,/ "'"
activation of B cells during infection may have been indicated.
The' resul ts of anothe,r study (Tanner et !!, in preparation) suq-'
gest that pOlyc1onal activation does, in ~fact, occur in' experi-\,. , ,
mental trichine1losis. , Polycl(:>na1 B activators can both enhance ~ b
and suppress an immune response to thymus. dependen t and. thymus
independent antigens, depending on the experimen~al conditions
(Persson, 1977). Administered ~n 10'" doses, POlyclonal B cel1
activators function as thymus,;l.ndependent antigens, whereas in" w
"
•
high doses the y induce a polyclonal activation of B cella (Coutinho
and M611er; 1973: Coutinho ~!l, 1974). ln our studies', signi-
ficant or marked enhanced responses were observed mainly in ln-
fections 07ta~ned after t~e 'i~oCUlatio~ of 500 larvae (Figures 2, fi
"4-, 6, 7, 8), whf!re a substantial amount o~ parasite pr~ucts could
be released duriftg infection.
\ \
\
J,
, .. ~
'" - ~
'.
Page 108
l, .. ., . . .4_ ~_. _____ ~ __ .... 4 ...... ....,..~"_'I\""o'~'V~_ .............. _ ..... 1t._ .., . .-'.. "\, , • ~-- ........... _-.,-~-- ... - ..... --., ---~.~~-- ..... ~ Al
Qit'
' ....
1 1
..
) ()
r
84
~ ,.
,/ AlI ~e extens~ve work that haB demonstratedc immunosuppression •
in trichinell~sis, has been based 'on "the use of unrelated anti-
gens, such as he~e~O~oqoU~ erythrocytes, vi~uses, allo9ra~s and
tumor cells. However, there,is no direct ,evidence that ~ny " o •
immunosuppression oceurs to the parasi te" an·tigens. Agrea t deal
of evidence of cellul -~~
and humoral immunity durin~ !.spiralis
infections have bé âes'cribed, and i t has been assumed by Most
investigators tha~ , - es ~ speqific respon;es to the
p~rasite;" Oit has beên shawn, howeJr, that ~he antibody'-response
in a number of parasit~ iftfections ls often non-speciflc (~ouba
and A11ison, 1,966: Smithers, 1967; Houba et al, 1969: Freeman
et al, 1970r Hbdson et al, 1976) and it has al~o been difficu1t
to correlate antibody level with protection (SOUlSbY,\1962; . .. ~ "'<,
Sm~thers, 196~; Tanner, 1968; Sinclair, 1970). It would be of J
great interes,t to assess the competence of the lymphold cell,8 of,
infe.cted 1 animaIs to the parasite antigens themselves, to obtain . ,
~ a better understanding of how'Trlchinella spiralis evades the
immune response. Since immunosuppression was apparent in these
. ,
studies at least 10 days ~fter adoptive.transfer into recipients,
some permanent effect on the ce11s, either in terms of.cell function \
or cell numbers or proportions may. be implied. Suppressor cells
~ay' al-so' be implicated, in the mechanism of levasion as h~been./' ind?catedl by J~nes et al (1'976) and su,ggc~tcd b; sorne of the .
results reportcd hcrc.
Our st~dies were general and preliminary since we dealt with
spleen cells as a whole. 'However· spleefl;s, lUte other lymphoid
. ;
Page 109
Cl
"
. '
•
" ......... ..:.... ____ ._ ........ _" ....... ,~~ .. I ... '""" 1"''''·
,'" . '1 •
organs contain B cells and T cells in differen~ p~oportions, in
addit~pn to a.significant number of macrophages. It would have ,
been of interest to study the immunological competence of the ''")
different cel1 populations of the spleen during the course of the
in fection. As all 'parasi tes,' Trichinellà ',spi'ralis survives in
a p~tatively immunoc~mpetent host and, thus, the parasite must , . either subvett the protecting responses to its own ends or ev~de
, ,'--
these responses. Th' s study has been a~ atternpt to determine " ' - ,.
the nature of, sorne f the m~chanisms by which the immunological
responses
are still
this study has,unco 1
~
ressed durin~ the infection. Although there ~
questions to be answered, it is hoped th~t
sorne areas which will be profi~~ble for
further investigation. 1 i /
/ 1
/ \ /
'j "
f
85
..
.. '
• l'
... ' ~ . . -.".
'.'\~ t .,'" " .' ,i "
" t ...
,J
. , ,
, ,
,/,
Page 110
l '
o
iJ J
1
1 .1
. ' . _~ ~ ....... ___ -.t~jI~"~",~~
\ BIBLIOGRAPHY
Abdou, N.l. and M. Richter. 1970. The ro1e.of bone marrow in the immune response. Advances in Immu~logy, 12: 201-270.
Anderson, R.E., G.B. 01son, J.R. Autry, J.L. Howarth, G.M. Troup ..' .
86
and.P.H. Bartels. 197'7. Radiosensitivity of T and B lympho-
c~tes IV. Effect of whole body. irradiation upon various lymphoid tissues and numbers of recirculating lymphocytes. 1977. Journal of Immunolog~,!!!: 1191-1200.
Andersson, B. and H. Blomgren. 1971. Evidence for thymus--independent humoral antibody production in mice against' pOlyvinylpyrrolidone and E .colr lipopolys'accharide. -,-Cellular Irnrnunology, ~: 411-424. -
. Araujo, F.G., P/.M. Coelho and L.H. Pereira. 1977. Schistosoma
mansoni~ impairment of the cell-mediated irnm~ response in mice. Clinical and Experrmental Immun01ogy, 28: 289~
, "'1 291.
Arnason, B.G., B~D. Jancovic, B.H. Waksman and C. Wennersten. ~ 1962. Role of the thymus in immune reactions in rats • .
II. Suppressive effect of thymectomy at birth on reactions of delayed (cellular) hypersensitivity and the cirèulating smalt lymphocyte. Jour~al of Experimental Medicine, 116: 177-186.
Bachrnan, G.W.
niasis • ..
,\ 1929. A precipitin test in experimental trichi-Journal of preventive Medicine, 3: 465-469.
Bachman, G: W. and J. Oliver-Gonzalez. 1936. Immunization in ,
rats against Trichinella spiralis. Proceedings of the Society for Experimental Bio1ogy and Medicine, ~: 215-217.
Bachman, G.W. and R. Rodriguez-Molina. 1933. Resistanc~ ta
iRfes~ation with Irichine11a spiràlis in hoqs. American Journal of "Hygiene, 18: 266-294.
Page 111
\ , 1 , ~,
- , " ,';'
, .
Barriga, 0.0. 1975. Selective immunodepression in mice by
Trichine11a spiralis extracts and infections. Cellular 1
Immunology, 17: 306-309. \ /-0
Benenson, A.S. 1975. Control-of Communicable Diseases in Man. American Public Health Association, Washington. .. . î Boyd, E.M. and J.H. Pètersen. 1954. Parabiosis and the problem
~< • ---,,- - of transmission and immunity in rats infected with Trichi-
J nella spiralis. Anatomical Record,~: 811-812.
r--------------~g9S, N.T. 19.63.' Immunologica1 in jury of mast cells in mice
t ! actively and passively sensitized to "antigens\ from Triehi-1 \ ne11a spiralis. Journal of Infectious Diseases, 113: 22-l '~
l '
f ' 32.
Briggs, 'N.T. and D.L. DeGiusti. 1966. Generalized allergie reactions in Trichinella-infected mice: the temporal
association of host immunity and sensitivity to exogenous
antigens. American Journal of Tropical Medicine and
Hygiene, 15: 9~9-929.
o
Briggs, N.T. and D.L. DeGiusti. 1970. Increased sensitivity of Trichinella~infected mice to serotonin and histamine.
Life Sciences, ~: 797-802.
Brown, I.N., S.R. Watson and V.S. Sljivic. 1977. Antibody response in vitro of spleen cella from Plasmodium yoelii
infected mice. Infection and Immunity, 16: 456-460. • i \ • Bruce, R.A. 1975. Tr~chlnos~s associated with oral squamous
cell carcinoma: report of three cases. Journarof Oral , Surgery, 33: 136-141.
Campbell, C.H. 1~. The antigenic r~le of the excretions and secretions of Tricllinella spiralis in the production of
immunity in mica. Journal of Parasitology, 41:-483-491.
Capron, A., D. Camus, J.P. Dessaint and E'
t Boubennec-Fischer.,
1977. Altfrations de la r~ponse imm~ e au cours des \ .
infections parasitaires. Annales 9; mmünoloqie (Institut Pasteur), 128: 541-556.
. . '.1 •
87
)
Page 112
\ , , ~ " l,
~ ~
" l ~ f .. " , t' , ~ s' r % '.
r f-
t
! 1
l 1
f
()
o·
.p ',' .r r, ,
,''- • d .... : ~ ... :._~ ....... jlt.." ~~-.~~~ ... ""'fItIM:'I.~~
Catty 1 D,' 1969. 1 The irnmunology rof nematode infections: trichinosis in guinea pigs as a model. Monographs in Allergy, . Volume 5, S. Karger, Basel.
Chandler, A.S. 1939. The nature and mechanism of immunity in yarious intestinal nematode infecti~ns. ~ican Journal, of Tropical Medicine, !!: 309-317.- ~
~
Chandler, A.S. and C.P. Read. 1961. Introduction to Parasito-~. John Wiley and Sons, Inc., New York.
88
~hernyakhovskaya, I.Y., G.S. Shagiyan and G.Y. Svet-Moldavsky. 1971. Correlation between t~~' 'ffegree of Trichinelfa .
,spiralis in faction of mice and suppressi.on .of· transplanta
tion irnrnunity._ Doklady Akademii Nauk SSSR,!2!: 1490-1492. "<l
Chernyakhovskaya, l.~., G.S. Shagiyan, ,E.G',Slavina and G.Y. Svet-Moldavsky. 1972. He1minths and allotransplantation.
European Jour~l of Clinical and Bio~ogical Research, !!: 395-399.
Chimishkyan, J<.\. land G.S. Ovumyan. 1975. 'The effec,t' of Trichinella spir411is on the susceptibili ty a·nd antibody produc-
, ~
'tion to vaccinia virus. Voprosy Virusologi~, !: 438-441.
Chimishkyan, K.L., ~.K. Shkvatsabaya, G.S. Ovumyan, V.A. Babich~v,
L.P. Trubcheninova, E.V. Sorokin a and G.Y. Svet-MoldavskY. 1976. The effect o~'Trichine11a spiralis on graft-versushost reaation, transplantat~on immunity and antibody for-, mation. Biomedicine, 25: 176-180.
Chitwood, B.G. 1974. Introduction S2 Nematoloqy. University Park Press, Baltimore.
Cnube, R.M. 1956. The dual antibody response to experimental trichinosis. Proceedings of the Helmint~ological Society of Washington, 23: 49-58.
Claman, H.N., E.A. Chaperon ~nd R.F. Triplet~. 1966a. Immunocompetence of transferred thymus-marrow cell combinations •. Journal of Immunoloqy, !l: 828-832.
, J 1 •
Page 113
1 1 1
Cl
o
.,., l\ "
," , , ,
~ _ ... " ...... ,. ..... __ ~.~J!p.._~"1>'_~-"';" #';~
89 ~
Claman, H.N., E.A'. ,Chaperon and ~.F. Trip1ett. 1966b. Thymus-\
marrow cel1 combinations. synergism in antibody production • . ,Proceedings of the Society for 'Experimental Biology and
~ ,
Medicine, 122: 1167-1171. , -
Clinton, B.A.,L.A. Stauber and N.C. Pa1czuk. 1969. Leishmania -
donovani: antibody response to chièken ova1bumin by infec-ted hamsters. Experimental Parasitology, 25: 171-180.
c _
Cochrane, C.G. and F.J. Dixon. 1962. AntibodY production by " 11 1 trans,ferred cel1~. Advances in Immunology, ~: 205-239.
Cohen, S. 1976. Survival bf parasites in the immunized hosto In: Immuno1ogy of Parasitic Infections. S. Cohen and E.H.
Sadun, editors. Blackwell Scientific Publications,'London.
, Coker, C.M. 1956. Sorne effects of cortisone in mice with ac~ired immunity to Trichinella spiralis. Journal of
y
InfectiQus Diseases, 98: 39-44.
Coutinho, A. and G. Môller. 1973. 'B cell mitogenic properties thymu~-independent ~ntigens. Nature (New Biology), 245:
12-14.
Coutinho" A., E. Gronowicz, W.W.lU1l0Ck and G. Môller. 1974. Mechanism of thymus-independent immunocyte triggering.
~itogenic activation of B cells results in specifie immune
responses. Journal of Experimental Medi@fne, 139: 74-92.
Crandall, C.A. and R.B~ Crandall. 1976. Ascaris ~:immuno-
suppression in mice during acute infection. Parasi tology, 46: 363- 372.
Expe r imen ta 1
,J '
Crandall, R.B. and C.A. Cranda11. 1972. Trichinella spira1is:
'- immunologie response to infection in mice. Experimental Parasitology, 31: 378-398. - .
, Crandall, R.B. and L.L.A. Moore. 1968. The macroglobulin
response in experimental,trichinosis. Journal of Infec·
tioua Diseases, 1181 377-385.
Crandall, R.B., J.J.Cobra and C."'. Crandall. 1967. The relative
" ,oh..-\ ~,
Page 114
.... ,,,.
, .
" .
. . ,
SM • 4 - :
., 90
proportions of 19G-, 19A- and 19M~containing cells in rabbit tissues during experimenta1 triohinosis. 1mmuno1ogy, . 12: 147-'158. '.
Cross, A~M., E. Leuchars and J.F.A.P. M;11er. 1964. Studies on the recovery of the immune response in irradiated mice thymectomized in adu1t life. Journal-of Experimental Medicine, 119: 837-850.
Culbertson, J.T. 1942a. Active immunity in mice against Trichinella spiralis. Journal. of pa!asi to,logy', 28: 197-202.
1 . Cu1bertson, J.T. 1942b. Passive transfer to Trichinel1a spiralis
in the rat. Journal of Parasitology, ~: 203-206.
Culbertson, J.T. 1943. Natural transmission of irnrnunity against Trichine11a spiralis from mother rats to their offspring. Journal of Parasi toloty, 29,: 114 .. 116.
Culbertson, J.T. and S.S. Kaplan. 1938. A study upon passive immunity in experimental trichiniasis. Parasitology, 30: 156-166.
Cunningham, A.J. and A. Szenberg.
in~the plaque t~chnique for forrning cells. 1mmuno~ogy,
1968. Further improvements detecting single antibody!,!: 599-60.1.
Cypess, R. and J.E. Larsh. 1970. The macrophage inhibition assay 0
(MIA) as an in vitro correlate of de1ayed hypersensitivity ...-(DH) in mice to Trichine11a S'piralif!·-q.rtigens. Journal of b
Paràsitology, 56(NUrnber 4, sec~o~J2): 64-65.
Cypess, R., J.E. Larsh and C. pegram.1971. Macrophage inhibition produced by qui~ea pigs aft~r sensftization with ~ larva1
~ ~ 'I} ~
antigen of Trichinella spiralis. Journal ~~'Parasitology, 57: 103-106 •
Cypess, R.', A.S. Lubiniecki and W.M. Ha,nunon. 1973. Immunosuppression and increased susceptibility to Ja~nese B ~ncephalitis virus in Trichinella~ira1is-infected mice. Proceedings of the Society for Experimental 8io109Y, !!!:
Page 115
o
1 1 •
·1 , , .cl ,
469-473.
, , . ." 91
Cypess, R.~ k.S. Lubinie~ki and D.M. Swid~a. 1974a. Oec~eased
susceptibi1ity to Listeria 'monocytogenes in mi ce after ~nfectiop with Trichinella spiralis. Infection and Immunity, !: 477-479. ~ ...
Cypess, R., J.A. Mo1i~ar', J.L; Eberso1e and A.S. Lubiniecki. , \
1914b. Immunologiéal'sequelae of Trichine11a spira1is infection in mice. II. Poten~iation of cel1-mediated . ~ . responses to BCG after infection with"Trichinel1a spira1is. Infection and Immunlty, !Q: 107-110. ,
Cypess, R., R. Zapata and D.Gitlin. 1974c.· Protection against Listeria monocytogenes in mice fo1lowing infection with Trichine11a spiralis. In: Trichine11osis: Proceedings of the Thl'rd International Conference 2!! Trichinellosis. C.W. Kim, editor. rEdu'Cational Pub1ishers,- New York. 147-152 •
Dean, R.B. and W.J. Dixon. 1951.' Simp1ified statistics for sma11 "'numbers of observations. Analytical Chemist:ç-y, ll: 636-638.
Denham, D.A. 1969. Immunity to-Tric~inel1a spira1is. IV. Passive immunity in mice. Folia Parasitologica, 16: 183-187.
Oespommier, 0.0. and B.S. Wost~ann. 1969. Trichine11a spiralis: im~une e1imination in mice. Experimental Parasitology, 24:
243-250.
Despommier, 0.0., W.C. Campbell and L.S. Blàir. 1977 • The in vitro analysis of immunity to Trichinella sEira1i's in
mice and rats. Parasito1ogy, 14: 109-119. -\
Dorin, R.P. 1946. The preparation and demonstration of an ûntÜ'l(~rum for 'l'riehir1C~llû spi ra lis. Journ"l 0 f Pûrasi to
! -109Y, ll: 8 3-8 6 •
Dueas, R. 1921. L'imriunitê da'ns la trichinose. Thllse. Jouve et Cie, Paris. ~
. ,
'[
,.' .c'
'" ., '
Page 116
1
J
i • t
~
" ~ 'i<\o , 1 ~f
_ ... ______ 1_..:..:_:1 ______ ' 7---
o
92
)! Duckett, M.G., ~.A. Denham and G.S. Ne,lson. 1972. Immunity to
TriohAnella spiralis. V. Transfer of irnmunity against . u,
the intestinal phase from mother to baby mioe. Journal , Parasitology, 58: 550-554.' ~
Fahey, J.L., W.F. Barth and W. Lloyd. 1965. Normal immuno-9 lobulins and tomized mice.
li: 663-675.
.. ' , antibody response in neonatally thymec-Journal of the National Cancer Institute,
Faubert, G.M •. 1976. D~pression of the plaque-forminq cells to sheep red blood ce1ls by the new-born larva~ of Triehinella spiralis. Irnrnunology,~: 485-489.
Faubert, G.M. and C.E. Tanner. 1971. Trichine11a spira1is: inhibition of sheep haernagglutinins in mice. Experimental Parasito1ogy, 30: 120-123.
Faubert, G.M. and C.E. Tanner, 1974a. The suppression of sheep rosette-forming cells and the inability of mouse bone marrow cells to reconstitute competence after infection with the nematode Trichine11a spiralis. Immunology, 27: SOl-SOS •
Faubert, G.M. ~nd C.E. Tanner. 1974b. En1argement of lymph nodes during infection: a preliminary histologica1 study~
'of this phenomenon. In: Trichine11osis: Proceedin~s of the Third International Conference'on Trichinellosis. - - . C.W. Kim, editor. Educationa1 Publishers, New York. 353-
W· 366.
Faubert; G.M. and C.E. Tanner. 1975. - Leucoagglutination and
cytotoxicity of the serum of infected mice and 'of extracts ~f Triehine11a spiralis larvae and the capacity of infeeteù mouse sera to prolong skin allografts. Immunology, 28: 1041-1050.
~hthal, J.H. 1943. Number of larvae and time required ta produee active immunity in rats against Trichinella spiralis.
, .
Page 117
" . ~~ ...... ---------~-_-.-.'" -
, '
, , , ~,~
Journal of Parasito1ogy, 29: 123-126.
,Freeman, T., S.R. Srnithers and G.A.T. Targett. 1970. Speci
ficity of immunoglobulin G in Rhesus monkeys infected with Schistosoma manaoni, Plasmodium knowlési, and
Tr~panosorna brucei. Journal of Infectious Diseases, 121: ,401-406. .. .
Gore, R.W., H.J. Burger and E.H. Sadun. 1970. Mechanisrns of
imrn~nity in rats infected with Trichinella spiralis.
Journal Qf Parasitology, 56 (Number 4, Section 2)': 122."
93
Gore, R.W., H:J. Burger and E.-H. Sadun. 1974. Humoral c1nd
cellular factors in the resistance of rats tO, Trichinella spiralis. In: Trichinellosis: Proceedings of the Third
International Conference on Trichinellosis. C.W. Kim,
editor. Educational Publishers, New York. 367-382.
suppression in murine malaria. l. General characteristics. < \
Clinical and Experimental Immunology, ~: 467-478.
Grove, D.I. and K.S. Warren. 1976. Effects on murine trlchinosis of niridazole, a~uppressant of cellular but not
humoral responses. Annals of Tropical Medicine and
Parasitology, 70: 449-453.
Hall, M.C. and M. Wigdor. 1918.
therapy in trichinosis.
601-609.
... An experimental study of serum
Archives of InternaI Medicine, 22:
Hendricks, J.R. 1950. The relationship between precipitin
titre and number of Trichinel1a spiralis in the intestinal
tract of mice following test infections. Journal of' Immu
nalogy, 64: 173-177.
Hendricks, J.R. 1953. Passive transfer of resistanee ta
Triehine11a spira1is in mice. Journal of Parasitology,
) [)
Page 118
f 1:
t ~ f ~.
{. )
• . " t ;.. ~,
f' ~ " î , !J" :.
f ,\ ~ !. ,-l, l
(j
o
39 (Supplement): 40-41. . Houba, V. and A.C. Allison." 1966. M-antiglobultns (rheumatoid
factor-like globulins), and other gamma globulins in rela
tion to tropical parasitic infections. Lancet i: 848-852.
Houba, V., K.N. Brown and A.C. 'Allison. 1969. Heterophile
antibodies, M-antiglobu11.ns and immunog1obulins in
experimental trypanosomiasis. Clinical and Experi~ental
Immunology, !: 113-123.
Hudson, R.J. 1973 .. _Moderated immunologie responsiveness in
parasitic,infections. Advances in Veterinary Scie~ce and
Comparative Medicine, 17: 87-117.
Hudson, K.M., C. Byner, J. Freeman and R.J.Terry. 1976.
94
Immunodepression, high IgM levels and evasion ofîthe immune
response in murine trypanosorniasis~ Nature, 264: 256-258.
Humphrey, J.H., D.M.V. Parrott and J. East. 1964. Studies on 1
globulin and antibody production in mice thymectomized at
birth. Immunology, 2= 410-439.
Jachowski, L.A. and G.A. Bengham. 1961. Influence of trichi
nosis on ·Schistosoma manso~i in mice. Journal of Parasi
tology, 47: 719.
Jackson, G.L. 1959.
nella spiralis
105: 97~11. .
Eluorescent antibody studies of Trichï
infections. Journal of Infectious Diseases,
- ~ Jones, J.F., C.A. Cranda11 and R.B. Cranda11. ,1976.
suppressi n of the primary antibody response to
erythrocytes in mice infected with Triehine11a
Cellular Immunology, 27: 102-110.
T-dependent
sheep
spiralis.
Kadish, J.L. and R.S. Baseh. 1975. Thymie rcqcncration after . lcthal irradialion: cvidcncc Cor and inlra-thymic radio-
resistant T-ce11 preeu~sor. Journal of Irnmuno10gy, 114:
452-458.
J<agan, I.G. and U. Bargai. '\ 1956. Studies on the serology of
Page 119
·_--_ ... _---
., ·r 1 f
~
f " ". ~
f ! t. .. ~ i ) 'i~
o
_. __ ._-_ .. ---,
95
trichinosis wi th ha f.'m."1gg1utination, agar diffUSiOI tes·ts
and precipitin ring tests. Journal of Parasitology, !!: 237-'245.
Karrnanska K., Z. Kozar, R. Sieniuta .and J. çzajkowska. 1971.
Behaviour of rnast cells in experimental triohinellosis in
rnice. Wiadomosci Parazyto1ogiezne, 17' 593-607.
Karmanska. K .• Z. Kozar. R •. Sieniuta 'and M ~lUgieWiZ-BUlla. 1973. The influence of antilymPhOCy(eUan? antimacrophage
serum on mast,cells in experirnen~a1 trichinellosis in rnice.
Acta Par~sito1ogi?a P01onica,' 2~: 173-182. 1
Kazacos, K. R. ~ 1975. Increased resistance in' the rat tQ Nippo-
strongy1us brazi1iensis fol1owing immunization against
Trichine1la spira1is. Veterinary Parasito1ogy, 1: 165-174.
,
r
s, K.R. 1976. Increased resistance in the rat to Strongy
loides ratti fol~owing irnrnunization against Trichine11a
( ·spira1is. Journal of Parasitology, 62: 493-494.
~lham, L. and L. Oliver. 1961. The prornoting effect ofrtri-, chinosis on encepha'lornyocarditis (Et1C) virus infectiorl
in rats. American Journal of Tropical Medicine and Hygiene,
10: 879-884.
Kim, C.W. 1966a. De1ayed hypersensitivity to 1arval antigens
of Trichinella spiralis. Journal of Infectious Diseases,
116: 208-214. ) '-
Kim, C.W. 1966b. De1ayed hypersensitivity to Trichine11a spi
ralis antigens in i~diated guinea pigs. Jour~a1 of Para
si tology t, 52: 722-726 p ~im, C.W., M.P. Jamuar and L.D.'Hamilton. ,1970. De1ayed hyper
sensitivity to Trichinella spiralis. III. Effect of
repca ted sens! ti za ti~:ms in donors' ahd recipien ts. 1 Journal of Immunology, 105: 175-186. - ,
Kim, C.W., H. Savel and L.,O. Hami 1 ton. 1967. De1ay~ hyper-.
sensitivity to Trichine11a, spiralis. 1. Tranaferlof
1 L,
• . '
~. ,
Page 120
96
t;\
delayed hypersensitivity by lymph node cells.1 Journal of Immunology, 99: 1150-1155. . ~
Kozar, Z. and R. Piotrowski. 1971.' ,Peritoneal macrophagq inhibition test in guinea pigs infected 'with Trichinella
spiral:is. Wiadomosci P~razytologiczne, 17: 609-615. ,. , .
Koza~" Z., K. Karmanska, K. Kotz and R. Sieniuta. 1971a. The influence of antil~phocytic serum (ALS) on the course of
1
trichinellosis in mice. Wiadomosci~Parazytologiczne, 17: b 541-572.
Kozar, Z., Z. Wieczorek, J. Kotz and M. Kozar. 1971b. _Oelayed . 1flf
hyp~rsensitivity and humoral antibodies in guinea pigs
sensitized with Trichinella spiralis antig~ns. Archivum } Immuno1ogiae et Therapie Experimentalis (Warszawa), 19: 491":'514.
Kozar, M., K. Karmanska, J~ Kotz, R. Sieniuta and D. Marciniec. ,~ ,
1969. Trichine1losis in thyrnectomized mice. Wiadomosci Parazytologiczne, 1~ 634-637.
Kwa~~c.K. 1971. Igr-1 and~gG and infectivity for mice. Experi-,
mental' Parasitology, 30: 82-87. -t:
Larsh, J.E. 1953. Studies; in old, mice to test the hypoth~sis, '-- .,
of locaL and genera1 immunity to Trichinella spiralis. f " ) Journal of In ect~ous D~seases, 93: 282-293.
J~
Larsh, J.E. 1963. Experimenta1'trichinosis. o
Advances-in r \
'Parasito1ogy, 1: 213-286. ,
Larsh, J.E. 196 7a. t '
The present ~nderstanding of the mechanism of inununit,Y to Trichine11a spira1is'. American Journal of
Tropical Medicine and Hygiene, 16: 123-132. ~
Larsh',J.E. 1967b. Delayed (cellular) hypersens~tivity in parasitic infections. American Journal of Tropical Medicine and Hygiene, 16: 735-745 •
. Larsh, J.E. 1968. Experimental trichinosis.o Advances in Para-
sitology, 6: 361-372.
çJ fK i 1 .. - ,.~ ________ (/";:~-_·,_~,t'!-"--'·· .. : .. ·":"Ld __ ~~~ _______ , ____ , __________ .
l
Page 121
()
)
.. '(
'i
.J.
!
1 "
• ê-
'.' i ~ \
~-~~' "'_r~_ ~"'-'''''''''''-' -..-. ~t ,l''':! ...... .-:'--~-.. L-..
91
'Larsh, J.E. and C.H. Campbell. ~952. 'The effec;:t on'the 1
natural resistance of mice, te Hymen01epis ~ var. fraterna
of a 'simultaneous infection'with Trichine1la spiralis."
Journal of Parasitology, 38 (Su~1ement): 20-21 . . La~sh, J.E. and G.J. Race. 1954.~ A histopathologic study of
. th~ anterior sm~ll intesti~e of" immunized atld non-immunized 'mice infected with Trichinella spiralis. JO,urna1 of
Infectious Diseases, 94: 262-272. -. Larsh, J.E. and G:J. Race. 1975. Allergie inflarrunation as a
h1pothesis for the expulsion of worms. Experimen~aJ
Parasito10gy, lI: 251%266,
• 1 t\ LArsh, J.E. and N.F. Weathe y. 1974. Cell-mediated irrununity .. ./ in certain parasitic infections. \ Current Topics in Micro-.
bio1pgY'apd Immunology, 67: l13~137 •
Larsh, J.E., G.J. Race and H.T. Goulson. 1959. A hisopatho1ogic
...,.. study of mice immunized with irradiated larvae of Trichi-c
nella spiralis. Journal of Infectious Diseases, 104: 156-
l63.
.~
" .
1
.~ 1
<1 1
Larsh, J.E., G.J. Race ~nd W.B. Jeffries. 1956. The association ~ - :1 p ~
in young mice of intestinal inflammation an4-the 10ss of
adult worms following an initial infection with T~ichinella
~piralis •. Journal of Infectious Di'seases, 2,2.: 63-71.
Larsh, J.E., G.J. Race and A. Yarinsky. 1962./ A histopathologic
,study iQ mice'immunized against Trichinella spiralis and
exposed.to total-body X-irradi4tion. American Journal of
Tropical Medicine and HY~'ene, Il: 633-640. , . --) .
Lars~, J.E., H.T. Goulson and N •• Weatherly. 1964a. Studies
on dela;(c~' (c~llular) hy~er~ nsitivity in mice infected
with Trichine11a spiral~~. Transfer of lymph node ce1ls. ~' id' 'l- '
Journal of the Elisha,rMitchEdl Science Society, 80: 133 ..
135.
Larsh, J.E., H.T. Goulson and N.'. Weatherly. 1964b. Studies
, ~ .-: ..
~.
.. ,
Page 122
o
.. '" .. ~ ...:.,~_'--..... _~.::.:...... .. "r<!'I~-". , ...... +'\'1.""f~ ... - ... "V"~'"
98
on de1ayed (cellular) hypersensitivity in miee infeoted ~~ Q
with Trichinella spiralis. P. Transfer of peritoneal ,\ exudate cells. "Journal of Par~si tology, 50: 496-498.
Larsh, J.E., G.J. Race, H.T. Goulson and N.F. Weatherly. " 1966. j
Studies on delayed (cellular) hypersénsitivity in mi ce infected with Trichinella spiralis.~iII. Serologie and
, ~is.topatho,logic findings in recipi~nts- given peritoneal
, " ~te cells. Journal of Parasitology, 52: 146-156.
Larsh, J.E~, G.J. 'Race~ J.H. Martil1 and N~F. W~athedY. 1974. '. Studies qn delayed (oellular) hypersensitivity in mice
• Q
infected with Trichinella spira1is. VIII. Serologie and
histepathelogic ~espDnses of recipients injected with spl~en 'cèlls, .from doners suppressed with ATS. Journal of'
'>.4. fA. t:
Parasitology, 60: 99-109., ,
Larsh, J.E., H.T. Goulson, N.F. Weatherly and E.F. Chaffee. 1969. ·~tudies on delayed (eellu1ar) hypersensitivity in mice infè~ted with Trichinella spiralis. IV.'~rtificial ~ensi
tization~of doners. ,Journal of Parasitology, 2i: 726-729.
-Larsh, J.E., H~T. Gou~on, N.F. Weatherly and E.F. Chaffee. 19lPa. Studies on delayed (cellular) hypersensitivity in
"mice infe'gced with 'Trichinella spiral!s. V. Teliits in
r~cipients
or 21 days e •
978-981.,
. , injected with donor spleen cells, l, 3, 7, 14,
o
befere infection. Journal- of Parasi tologYt, ~:
LarSh,' J.E., H.'P. Goulson; N.F. Weatherly \and .F. Chaffee • • ,1970b. Studies on° delayed (cel).ular) ypersertsitivity in mice infected wrth~ Trichinella s lis. VI.~Results in
~=-r-=;;.;;;.
racipients ~njectèd with antis spleen ,Q • ,cells. Journal of Parasitolo ,~: 1206-1209.
1 ~
Larsh, J.E., N.F. wea'therly, H.T. Goulson and R.F. Chaffee. " l~72. Studies on delayed (oellular) hypersensitivity'in'
m~ce inf~cted ,ith Trichinella' sPir~lis. VII~ Éffect of . ATS injections ?n numbers of a~lt worms recovered after
-..
.r
, .
Page 123
o \
•
\ . "'
~" '- ~, '. , .
- '-"'~-_";;"""'-""""-~""'~-'
, ...
challenge. Journal of Parasi tology, 58: 1052- 1060. ~ - 1 - ,
LjungstrlSm,: 1. 1916. graft re~ection. Infections. E.J. York. 247-253.
The effect of acute trichinosis on a1lo-h: Pathpphysfology 9!. Parasi tic 'J
Soulsby, editor. Academie Press, New
.. /'
99
Ljungstr6m, 1. and G. Huldt. 1977. Effect of experimental trichinosis on unrelated humoral and cell mediated immunity. Acta Pathologica e~ Microbio1ogica Scandinavica, Section r
C, !!2.: 131-141-
, Ljungs'tr6m, 1. and E.J. Ruitenbèrg. 1976; A comparative-study
of the immunohistological and serological response of 0
intact and T-cell üeprived mice tO,Trichinella spiralis. Clinica1 and Experimental Immun01ogy, !i: 146-156.
Love, R~J. "S.M." Ogilvie and D.J. McLaren. 1976. The immune mechanism which expe1s the intestinal stage of Trichinella spiralis from rats. Immunology, 1Q: 1-15.
Lubiniecki,' A.S. and R. Cypess. 1975a. Irnmunolocjical seqùelae Of' Trichinella spiralis inftaction in mice: ':tfect on the antibody 'responses te sheep erythrocytes and Japanese B 1-encephalitis virus. Infection and Immunity, Il: 13b6-13rl~ , ,_
Lubiniecki, A.S. an~ R. ~ypess. 1975b. Quantitative study of the effect of previous Trichirie11~ ~pir~lis infection on sarcoma 180 a~citic tumqr fôrmation in mice. Tropenmedizin und Parasitologie, 26: 329-3~4.
Lubiniecki, A.S., R. Cypess and J.P. Lucas. 1974a., Synergistie int~raction of two agents in mice: Japanese lB encephalitis
, . virus and Trichinella spiralis. American Journal of Tropical Medicine and Hygiene, 23: 235-241.
Lubiniecki, A.S., R. Cypess and J.P. Lucas. 1974b. Immune r.f!sponse to and distribution of sheep erythrocytes i Trichinella spiralis-infected mice. Tropenmedilin und
Parasitologie, n~ 345-349 •
".I,J ~
,.' '~ ~ .
tl .. ~. '"'t~;., • •
• .. ~, '
, Il' \' "', ~
;.' ....
, ,
Page 124
0, , '
,
, ~... 'lIIo" " , , { ... ~ 1 11'" ,_,~ ... "_~_._ .. c ... ~~~ .... fl'l'-
Machnicka, B. 1972. TricHinella
lymphocytic serum on mouse
Parfis1tologyc/ 31: i. 72-177.
spir~lis: influence of anti
infections. Experimental
\
100
Madsen, H. 1976. Life cycle of !. spiralis. Acta Parasito1og1ca Polonica~, 14: 143-158'.
Mansfield, J.M. and J.H. Wallace. 1974. Suppression of ceU
mediated immuni ty in experimental African trypanosomiasis.
Infection and Immu~ity, ~: 335~339.
Marbrook, J. 1967. Primary immune respo,nse in cultures of
spleen cells. Lancet ii: 1279-1281. \
Matossian, R.M., 1. Sa1ti and E. Stephan. 1977. Variation in J
serwn immunoglobulin, levels in acute ,trichinosis. Journal
of Helminthology, 51: 1-4 • . -Matov, k. and ~ Kamburov. 1968. Experiments for the produftion
qf cross immuni ty between Trichinella spiralis and Ascaris
suum. Izvestiia na Tsentralnata ~helmintologichna Laborator1iia (Sofia), 12: 167-173. '
Mauss, E.A. 1940a. Transmission of immunfty to Trichinella , '
spiralis, from, infected. animaIs to their offsprinq • . ' . American Journal bf Hygiene, ~2(Section Dl: 75-79.'
o
Mauss, E .A. 1940b. The in vitro effect 0(' immune -seiuin upon
Trichinel1a spiralis larvae. American Journal of Hyqlene,
32 (Section D): 80-83. ," ..
Mauss, E.A. 1941. The serum fraction with which anti-trichinE!11a . . (Trichine1la spiralis) antibody i8 associat'ed. American "
Journal of Hygiene, U(Section' D): -Tl-S0.
MCCoy, O.R. 1931. Immunity of rats to reinfection with
Trichinel1a spiralis. American Journal of Hygiene, 14:
484-494. '\
McCoy, O. R. 1935. Artificial immunization of rats agalnst
Trichine1la spiralis. ,American Journal of Hyqien~, ~: . •
200-213.
\
--' \
l"
!: "
Page 125
(j
)
o.
- .=+ :
',,1 ..: ),
101 1
~
McCoy, O. R.... 1940. Rapid 1055 of Trichine11a larvae fed to
imrnun~ rats and - i ts béaring on the' mechanisrn of irnmulli ty.
Arnerican Journal of Hygiene, E: 105-116.
MeCoy, O. R. and F. F. Bond. 1941. Attempted passive transfer to
rats and mice of, immuni ty to Trichinella spiralis. Journal
,of Paràsitology, 27 (Supplement}:' 24;;'25. -------- J
Meerovitch, E. and S.J. Aekerman. 1974. Trypanosomiasis in
rats with trichinosis. Transactions of the Royal Society
of Tropical Medicine and Hygiene, §: 411.
Micklem, H.S,. and J.F. Loutit~ 1966. Tissue Grafting and
Radiation. Academie Press, New York. '\ 1}
Miller, J.F.A.P. 1961. Inununological function of the thymus.
Lancet, ii: 748-749.
Mills, C.K. and N.H. Kent. - 1965. Excretions and secretions ~
of Trichinella spira1is and their role in immuni ty.
Experimental Parasitology, 16: 300-310.
J.F.A.'P. Miller. 1968. Cenl to cell inter"::
immune re,sponse. II .~he source of hemo1ysin-
irradiated mice given bone marrow and ..
\
)
ic duct lymphocytes. Journal of Exp.:;rimental
Medicine, 821-837. c::;;;;-....
\ Mochizuki, H., T. Tomimura and T. Oka. . 1954. Cerebrospinal
~ nematodiasis as a provoking factor in Japanese B encepha
litis: an experirnenta.1 approach. Journ~1 bf Infectious
Oiseases, ~ ~ 260:"266. \ . MOlinari,r J.A. and R. Cypess. :f975. lnununo1@gica1 sequelae of
Trichine11a spiralis infection in mice: effect of viability ,. ,
and route of BCG administration on nematode induced
immunopotentiation. In'fection and Immunity, .li: 919-:921. l .
Molinari, J.A., R. Cypess and J.L. Ebersole. 1974.~ Effect of
Trichinel1a spiralis infectioR on the ce ll-mediated
immunÈl response to BCG. International Archives of A11ergy 1
• . '
,.'
Page 126
.~' 1 'l.
, "
"
, , -, . j
.' .; ~ .... ;: ~ . --t , ..;- ~ 1 ~ ~ I}. 10,
~ __ !.!.- • ~i_
.,
47: 483-487.
"', t~ , ;, " .' ,
, .. ""j' " ,
102
Mo ta , 1., ~:H. Sàdun, R.M. Bradshaw and R.W. Gore. 1969. The
immunologiç response of mice infected with Trichinel1a
spir~lis. Biolo9ica~.a~d physicochemical distinct~on of .
• :'two' homocytotropie antIbddies. I~unology,!§.: 7l-81.,J, 1" ", ... MUller,· R. 1975. Wôrms and Ofsease. A Manua1 of MedicaL
Helmintho109l' ~illiàm Heinemann Medical Books Ltd., 1) ~,
LOndon: ... .f{
Munoz, j.J: and R.L. Cole" 1977. Effeét of Trichine1la spira1is " "'"
infection on-passive cutaneous anaphylaxis in m~ce: ~nfec-.. Z.r: ::r'" ,
tion and Immunity, 15: 84-90. ~ -" - \
, 1
Offut t; E. P. 1941.' 'l:.he effects 'of immune s'erum upon the larvae
of Trîchinella spiralis-!n'~~o. Journal of Parasi~ology, , 27 (Supplement): 24. ,}
Ogi1vie, B.M., and ~.J.M. W±fson. 1976. Evasion of the immune
response by parasitès. British Medical Bulletin, 32:' 177-
~81.
Oliver-Gonzalez, J.
on the larvae . ,
of Infectious
. 1940. The in vitro action of· ilt\lPU(le serum
• and adults of Trichinel1a sp~r~lis. Journal
Diseases, ~:. 292:"300. • :'. i ~ 4. !
Oliver-Gonzalez, J. 1941.0 The dual antibddy basis of' acqulred • é" 1
immunity in.tr.i,~hinosis,,.;' Journal of ~n~ectio.ù~ Disea,~es, 69: 254-270 ..
.~ .. 1
Osmond, D.G. and-G.J.:'!. Nossa'l. v '" ; _
1974.' Differentiation of '1ym_ .~ ,
phocYtes in mouse bone marrow. II. Kinetics of ~aturation and renewal of' antig1'Obulin~brnd~ng ,ce11s studied by, i double labelling. Cellular Immunology, 13: 1,32-145. 1
~. ./ Ottescn, lLA., 'l'.Je. Smith imù C:U'. \Kirkpa~rick. 191.5.' Inttrunc
, ~
response to Trichinella spiralis ~n the" rat. 1. Deve op-
ment of cellular, and humoral responses dur'ing ehronic . ) ). . infection. International,' Arcl)ives of Allergy and Appliêd
, p' , ~
Immunology~ !!: 396-410.
, ,
..
. , ' ..
\,
Page 127
t
,'. '" "'. , .... _'" .• __ ...... --........... ..... ~..."..~-"'1"".--___ ... ""'...,. .....
/
Perrudet-Badoux, A., R.A. Binaght" and G. Bi02Zi. 1975. Trichi.nella infestation in mice genetically selected for high . and low antibody production. Immuno1ogy, 29: 387-390.
Perrudet-Badoux, A., R.A. Binaghi and Y. Boussac-Aron. 1976.
Production of different classes of immunoglobulins in rats infect~d with Trichinella ~pira1is. lrnmunochemistry, 13: 443-445.
Perry, R.H. 1974 Transfer of immunity'to Trichinella spiralis • i
from mother to offspring. Journal of Parasi tology', 60: 460-465.
)
103
Persson, U ..... 1977. Lipopolysaccharide-induced suppression of the primary immune response to a thymus-dependent an tigen.
i
Journal of Irnmunology, 118: 789-796.
Piotrowski, R. 1975. Transfer of delayed-type hypersensitivity by cells (spleen, mesenteric 1ymph node) in Trichinella spiralis infection in micè. 21: 623-632.
Wiadomosci"Parazytologiczne, \
Rappaport, 1. and H.S. Wells. 1951. Studiés in trichinosis. 1. Immunity to reinfection in mi ce fol1owing a single light infection. Journal of lnfectious Diseases, ~: 248-253.
Reed, S.G., C.L. Larson and C.A. Speer. 1977. Suppression oi cell-mediated irnrnunity in experimental Chagas' disease. Zeitschrift für Parasitenkunde, 52: 11-17. .'
Rivera-Ortiz, C. I. and R. Nussenzweig. 1976. Trichinella
spiralis: anaphylactic antibody formation and susceptibility in strains of inbred mice. Experimental Parasitology, 39: 7-17.
Romanovitch, M. 1912. Recherches sur la trichinose. Annales d'Institut Pasteur, ~: 351-370.
'- . Roth, H. 1939. Studies on the course of trichina reinfection
with Trichine11a spira1is. III. Immunity of guinea piqs. to rcinfection with Trichine1la spiralis. American Journal of nygiene, ~: 35"64.
"
Page 128
o
~'1, ,. ~;,,,". ~ , M
,,', , ; .. ,,( , '1"'1.$-
•• ~ .... _"" __ ~--..._~';' f"'''-~ ~~'f>"1Ulr~"'"
1 -
. Roth, H., 1941. The in vitro action of trichina larvae in
immune serum. A new precipitin test in trichinosis . . Acta Pathologièa et Microbio1ogica Scandinavica, 18:
160-167.
Ruitenberg, E.J. and M.J.M. Duyzings. 1972. Immunohistological ,
study of the inununological respbnse Of the rat to infection
with Trichinella spiralis. Journal of Comparative
Patho1ogy, 82: 401-407. ~
Ruitenberg, E.J. and A. Elgersma. 1976. Absence of intestinal ~
mast cell response in congenitally athymie nude mice
during Trichinella spiralis infection. Nature, 264:
258-260.
Ruitenberg, E.J. and P.A. Steerenberg. 1974. Intestinal phase
of Trichinella spiralis in congenita11y athymie (nude)
mice. Journal of parasitology, ~: 1056-1057.
Ruitenberg, E.J., J.S. Teppema and P.A. Steerenberg. 1974. The
intestinal phase of Trichinella spiralis infection. In:
Parasitic Zoonoses: Clinica1 ~ Experimental Studies.
E.J.L. Soulsby, editor. Academic PreEls, New York. 319-
326.
Ryser, J.E. and P. Vassalli. 1~74. Mouse bone marrow lympho
cytes and ,their _differentiation. Journal of Immunology,
113: 719-728.
Sadun, E.H., I. Mota and· R.W. Gore. 1968. Demonstration of
homocytotropic reagin-1ike antibodles in mice and , ,.\
rabbits infected with 1:. spiralis. Journal of Parasito-
10gy, 54: 814-821 • • , 1
Schwartz,' B. 1917. Serum therapy for trièhinosis. JOurnal of
, the'American Medical Association, 69: 884-886 •
L
.. , Semrad, J.E. and M.J. Coora. 1951. The riae and fall of
immunity to !. spiralis in the ~bino rat and its effecj:a
on growth and reprodu~tion of th parasite. Trana.ctions
of the' Illinois Academy of Scie ce, !!: 253-258.
104·
, ,
Page 129
~~""~" .. _'. -
o·
•
. . , . ' , . ........ - - ----_ ..... -- " _.,,_ ~ ............ _ ... '.- .. ,,.,. __ ~""\ ................ 'o!l'i~ ... '~ ... 11,,,,,. ~ ~
*
,
105
, ' . , '
Sengers, R.C •. A~, C.R. Jerusalem and W.H. Doesburg. 1971. Murine
malaria. IV. Disturbeà immuno1ogica1 responsi veness during • l' -'-
Plasmodium berghei infection. Experimental Parasitology,
lQ.:' 41-53.
Shikhobalova, N.P. 1952. Experimental investigation of irnrnunity
against Trich~e1~a. II. Immuni"ty acquired as a' result "
of disease. Trudy Gelmi'ntologi~heskaia Laboratoriia
Akademii Nauk SSSR, ~: 60-71.
$himp, R.G., R.B. Cranda11 ana C.A. Cranda11.' 1975. He,iigmo
somoides ,polygyrus (Nematospiroides dubius): suppre§lsion
of antibody response ta orally -administered sheep
erythrocytes in infected mice. Experimental Parasi tology, 1
38: 257-269.
Shope, R.E. f1943. The lungworm as a reservoir and intermediate
host for swine influenza virus. ·III. Factors influencing . 1
transmission of the virus and the provGcatipn of influenza. L
Journal of Experimental Medicine, 77: 11-126. - . Sinclair, I.J. 1970. The relationship,between circulating
l ,
antibodies and irnrnunity to helminthic infections. Advances
in Parasi ta109Y, 8: 97-138.
Smithers, S.R. 1967. The induction and nature of antibody
response to parasites. In: ~ American Health Organiza
tion Scientific Publication Number 150: 43-49.
" Sou1sby, E.J .L. 1962. Antigen-antibodt reaction~Hin helminth
~ infections. Advances in Immunology, 2: 265-308.
Stankiewicz, M and N.F. Weatherly. 1975. lnununobiology of . Trichine11a spiralis. Wiadomosci Parazytologiczne, ~:
591-599. ?1
Steel, R.G.D. and J.H-. Torri~. 1960. Principles ~ Procedures
2! Statistics. McGraw-Hill Book Company, Inc., New York.
Steele, J.H. and P.V. Arambulo. 1975. Trichinosis', a world
wide problem with extensive sylv.atic reservoirs. Inter-"
,. ,
Page 130
o
r '.
, .
o
,..~~,,-~rf,~~f.:.r~~~,,\' .. ::. .1>... _ _,(10, ...
t~' : ; .. " t
.. -"''''''''----.:---, .. ~~ ...... ''''
106
national Journal of Zoonoses, ~: 55-75. "
. Ste~apski, kJ. and A. Malezews~L 1972. Specificity of migra
tion inhibition test in parasitic oinvasions. II! Studies
on Triohine!l.la spira1is. Bulletin de l'Academie Polonaise
des Soiences, Serie des Soienoes Biologiques, 20: 261-262.
~ '" Strickland, G. T. and P.C. Sayles . 1977. Depressed antibody
. respon!;!e to a thymus-dependent antigen in toxoplasmosis.
Infect.ion and lnununitY'rll: 184-190. .
Svet.-Moldavsky, G.J., G .. S. Shagiyan, 1. Y. Chernyakhovskaya, \
. D.M. Mkheidze, T.A. Litovchenko, N .N. Ozeretskovskaya and
~·~G. Ka'daghidze:. 1910. Inhibition of sk'i'n allograft
rejection in T-rioltinella-infected mioe. Transplantation,
9: 69-70.
Svet-Moldavskl', G.J., ,G. S.Shagiyan, D.M. M~heidze, T.A. Litov
. ohenko ~ N. N. Ozeretskovskaya, Z. G. Kadaghidze and I. Y •
èhernyakhovskaya. 1969. Mouse transplantation ïrnmunity ,
depressed bl' Triohinella spiralis. Lancet,!: 320.
Taliaferroi W.H. 1940. The meohanism of acquired iIMIunity in
infections with. parasitio worms. Physio1ogica1 Rev~ews,
20: 469-492.
'Taliaferro, W.H., L.G. Taliaferro and B.N. Jaroslow. 1964.
Radiation ~ Immune Meohanisms. Academie Press, New York.
Tanner, C.E. 1968. Relationship between infeeting dose, muscle
parasitism and antibody response in experimenta1 trichino-• 1
sis- in rabbits. JoU7na1 of Parasi to1ogy, ll: 98 ... 107.
Tanner, C. E. '1,974.
rabbits after
In te rna tional \
'>~hancement of ~uscle trichi·nosis in ~' ,4
imiilunization with hetei.0lOQOUS serum proteins. Journal of Parasitology, 4: 193-196. , -
Tanner, C.E. and G.M.. Faubert. 1974. Purification 'of a leuco-l ' , , '
aQql utina tinq factor from TE' iohinella spiralis larva~. .
In: Trichine11osis: proceedingB.'·~ ~ Third Intèrnational
Çpnference ~ Trichirielloais. C.W. li •• editor. ~ational 1
•
, "
: ' ~ ...
Page 131
" " " ~
1 ''J,
\
•
'. '
Publishers, ~ew York. 327-334.
~Tanner, C.E.., H.C. Lim and G.M. Faube,rt. 1978. Trichinel:1,a
spiralis: changes eaused in the mouse r S thymie, splenie, • 1
and lymph node cell populations. Experimental Parasito-
logy, 45: 116-127. ...
Terry, R.j,- and S.R. Smithers. 1975 .. Evasion of the immune'
·response by parasites., Symposia Iilf,the Society for
Experime;ntal Biology, ~: 453-465. ~
Trubcheninova, L.P., K.L. Chimishkyan, G.S. Ovumyani V.A.
Babichev'ànd G.Y: Svet-Moldavsky. 1975. The effect of
Trichinella seiralis infection on the developrnent of
Rauscher leukernia. Voprosy Virusologii, l,-: 344-348.
Vernes, A., F. Flocrh, J. Biguet and R. Tailliez. 1975a.
Trichinose expérime.ntale. 1. Cinlitique des ph~nomènes
107
d rhypersensibilité retardée chez la 'sours CBA et le rat
Wistar. International Journal of Parasitology, ~: 63-70.
Vernes, A., R. Tailliez, F. Floc'h and J. Biguet. 1975b.
Trichinose expérimentale. II. Variations quanti ta tives
et qualitatives des globulines sériques chez la souris
CBA et le rat Wistar. International Journal of Parasi
tology, .§..: 71-75.
Wagner, E.D. and P.A. Nembha.rd. 1976. Concurrent infections in
mice wi th Trypanosoma esuiperdum and Trichinella spiralis. . . '
Japanese ,Journal of Parasi tology, 25: 1-,4. , -
Wakelin, .0. 1976. . Host responses. In: Eco1ogiea1 Aspects of
Parasi tology • C. R. Kennedy, edi tor. North-Holland Pub-....
lj"s~ing Company, Amsterdam.
Wakelin, D. and M. Lloyd. 1976. Acceleratcd expulsion of
adult Trichinclla spiralis. in mice given lymphoid cells c •
and serum f+om infected donors. Parasitology, 72: 307-315.
Walls, R.S., R.L. Carter, E. Leuchars and A.J.S. Davies. 197~.
The immunoBathology of trichiniasis in T-cell deficient
~ .'
Page 132
108
mice'. Clinical and Experimental Inununo1ogy, 13: 231-242.
Walton, B.C. and C.A. Gleiser. 1966. Prqvoking effect of
migrating 1arvae on viral encephalitis in mice. In: 1
proccedings oft
the First Interjnational cong~ess of
Parasi tology . Societa l taliana di Parassi t01,ogia.
Weatherly, N.F. 1970. Inçreased survival of Swis's mice given cr
sub1etha1 infections of Trichinella spiralis. Journal of
Parasitology, 56: 748-752.
Wescott, R.B. and A.C. Todd. 1966. Interaction of Nippostron
r gylus braziliensis and influe.nza virus in mice." 1. "Influ
ence of the nematode on the virus. Experimental Parasi
tology, 22: 242-247.
, .
Woodruff, A.fW. 1968. Helminths as vehicles and syne:rgists of
microbia1 infections. Transactions of the Royal Society
of Tropièal Medicine and Hygiene, 62: 446-452. , --
Wright, G.G. and J. Oliver-Gonzalez. 1943. Electrophoretic
studies on antibodies to Trichinella spiralis in the rabbit.
Journal of Infectious Diseases, 72: 242-245.
Yarinsky, A. 1962. The influence of X-irradiation on the
immunity of mice to infection with Trichine11a spira1is.
Journal of the E1isha Mitchell Science Society, 78:29-43. 1 --
Zaiman, H. 1953. Studies on the nature of immunity to Trichi
[fella spira1is in parabiotic rats. 1. A preliminary study
of the inunune response of the "uninfected" rat as deter
mined by observation Qf the intestinal phase. American
Journal of Hygiene, 57: 297-305.
Zaiman, H. and J. Rubel. 1953. Studies on the nature of immunity
ta Trichinella spiralis in parab~otic rats. III. The inunune
response in parabiot,ic rats surgically separated from
their mates five days after one member of the experimental -:
pairs received an immunizing dose of Trichinella spiralis.
American Journal of Hygiene, 51: 311-315.
Page 133
'1
'.
109
Zaiman, H., G. Omi and L. Gapineki. 1953. Studies on the
nature of immunity to Trichinella spira1is in parabiotic . rats" II. A prelirninary study of the immune resp,onse to,
the uninfected rat as detèrmined by observations on mu~cu
lar invasion. American Journal, 0 f Hygiene, 57: 306- 310.
Zaiman, H., J.M. Stoney and ~N.C. Headly". 1955a. Studies on the
nature of immunity to Trichine11a spiralis in para
biotie rats. VIII. The duration of the immune respons~ in
the uninfected parabioÙc rat followitn infection of o~e twin with Trichinella spiralis. American Journal of Hygiene,
61: 15-23.
Zaiman, .H., J.M. Stoney, J. Rubel and N.C. Headly. 1955b.
Studies on the 'nature-of immunity to Trichine11a spiralis
in parabiotie rats. VII. The immune response of the - '1"
uninfeeted twin one month after its mSte received an immu-• , r
nizing dose of irradiated (X-ray) larvae. American.
, Journal' of Hygiene, 6h 5-14.
)