IAWA Bulletin n.s., Vol. 12 (4),1991: 389-417 WOOD ANATOMY OF THE BIGNONIACEAE, WI'IH A COMPARISON OF TREES AND LlANAS by Peter GBS80n 1 and David R. Dobbins 2 Summary The secondary xylem anatomy of trees and lianas was compared in the family Big- noniaceae. General descriptions of the family and the six woody tribes are provided. Lianas belong to the tribes Bignonieae, Tecomeae and Schlegelieae, and most have ve.ssels of two distinct diameters, many vessels per unit area, large intervascular pits, septate fibres, large heterocellular rays often of two distinct sizes, scanty paratracheal and vasicentric axial parenchyma and anomalous growth. Conver- sely, trees, which belong to the tribes Coleeae, Crescentieae, Oroxyleae and Tecomeae gen- erally have narrower vessels in one diameter class, fewer vessels per unit area, smaller intervascular pits, non-septate fibres, small homocellular rays, scanty paratracheal, ali- form or confluent parenchyma, and none ex- hibits anomalous growth. The majority of both trees and Hanas possess growth rings, are diffuse-porous, have non-solitary vessels which lack helical thickenings, and few have apotracheal parenchyma or storied structure. All species have alternate intervascular pitting and simple perforation plates. Key words: Lianas, trees, wood, anomalous structure, xylem, Bignoniaceae. Introduction This paper has two purposes: to present descriptions of the secondary xylem anatomy of the Bignoniaceae, and to compare the wood anatomy of trees and Hanas within the family. Metcalfe and Chalk (1950) have surveyed the wood anatomy of this family, but there is no comprehensive publication, although many genera and species are covered in papers on particular geographical regions (Record & Hess 1943; Jain & Singh 1980; see also Greg- ory 1980 and in preparation). Den Outer and Veenendaal (1983) have compared Bignonia- ceae wood anatomy with that of Uncarina (Pedaliaceae) and Santos has recently describ- ed the New World Tecomeae for an M.S. thesis (Santos 1990) and Flora Neotropica (Santos in press). The family Bignoniaceae has a wide dis- tribution from about 400N to 35°S, encom- passing North and South America, Africa south of the Sahara, Asia, Indonesia, New Guinea and eastern Australia. It is mainly tropical, with most species in northern South America, and consists of lianas, trees and shrubs with very few herbs. Estimates of the number of genera and species vary: 650 species in 120 genera (Willis 1973) or 800 species in 110 genera (Takhtajan 1987), al- though Gentry (1973) considers that the num- ber of genera is too high. We have examined specimens from 35 tree and 27 liana genera from six of the eight tribes recognized by Gentry (1980), Cronquist (1981), and Takhtajan (1987). Two tribes, both neotropical and monogeneric, Tourret- tieae and Eccremocarpeae, are not covered in this paper since they are herbaceous vines (Gentry 1980). Much of the world distribu- tion of the family is also covered. An overall comparison of trees and lianas has been made, and anatomical descriptions of the six tribes are presented. The tribes Coleeae, Crescen- tieae and Oroxyleae consist entirely of trees and shrubs, whereas all members of the tribes Bignonieae and Schlegelieae are lianas. The Tecomeae are mainly trees and shrubs, but Campsis and Tecomanthe are climbers. The trees come from a wide range of habitats 1) Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3DS, U.K. 2) Biology Department, Millersville University, Millersville, Pennsylvania 17.5.51, U.S.A. Downloaded from Brill.com01/11/2022 05:37:50PM via free access
Transcript
IAWA Bulletin n.s., Vol. 12 (4),1991: 389-417
WOOD ANATOMY OF THE BIGNONIACEAE, WI'IH A COMPARISON OF TREES AND LlANAS
by
Peter GBS80n 1 and David R. Dobbins2
Summary The secondary xylem anatomy of trees
and lianas was compared in the family Bignoniaceae. General descriptions of the family and the six woody tribes are provided. Lianas belong to the tribes Bignonieae, Tecomeae and Schlegelieae, and most have ve.ssels of two distinct diameters, many vessels per unit area, large intervascular pits, septate fibres, large heterocellular rays often of two distinct sizes, scanty paratracheal and vasicentric axial parenchyma and anomalous growth. Conversely, trees, which belong to the tribes Coleeae, Crescentieae, Oroxyleae and Tecomeae generally have narrower vessels in one diameter class, fewer vessels per unit area, smaller intervascular pits, non-septate fibres, small homocellular rays, scanty paratracheal, aliform or confluent parenchyma, and none exhibits anomalous growth. The majority of both trees and Hanas possess growth rings, are diffuse-porous, have non-solitary vessels which lack helical thickenings, and few have apotracheal parenchyma or storied structure. All species have alternate intervascular pitting and simple perforation plates. Key words: Lianas, trees, wood, anomalous
structure, xylem, Bignoniaceae.
Introduction
This paper has two purposes: to present descriptions of the secondary xylem anatomy of the Bignoniaceae, and to compare the wood anatomy of trees and Hanas within the family. Metcalfe and Chalk (1950) have surveyed the wood anatomy of this family, but there is no comprehensive publication, although many genera and species are covered in papers on particular geographical regions (Record &
Hess 1943; Jain & Singh 1980; see also Gregory 1980 and in preparation). Den Outer and Veenendaal (1983) have compared Bignoniaceae wood anatomy with that of Uncarina (Pedaliaceae) and Santos has recently described the New World Tecomeae for an M.S. thesis (Santos 1990) and Flora Neotropica (Santos in press).
The family Bignoniaceae has a wide distribution from about 400N to 35°S, encompassing North and South America, Africa south of the Sahara, Asia, Indonesia, New Guinea and eastern Australia. It is mainly tropical, with most species in northern South America, and consists of lianas, trees and shrubs with very few herbs. Estimates of the number of genera and species vary: 650 species in 120 genera (Willis 1973) or 800 species in 110 genera (Takhtajan 1987), although Gentry (1973) considers that the number of genera is too high.
We have examined specimens from 35 tree and 27 liana genera from six of the eight tribes recognized by Gentry (1980), Cronquist (1981), and Takhtajan (1987). Two tribes, both neotropical and monogeneric, Tourrettieae and Eccremocarpeae, are not covered in this paper since they are herbaceous vines (Gentry 1980). Much of the world distribution of the family is also covered. An overall comparison of trees and lianas has been made, and anatomical descriptions of the six tribes are presented. The tribes Coleeae, Crescentieae and Oroxyleae consist entirely of trees and shrubs, whereas all members of the tribes Bignonieae and Schlegelieae are lianas. The Tecomeae are mainly trees and shrubs, but Campsis and Tecomanthe are climbers. The trees come from a wide range of habitats
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
390
and climates, e.g. temperate (Catalpa), warm temperate and subtropical (Chilo psis), monsoon forest (Millingtonia), secondary tropical forest (Oroxyiurn), arid tropics (Catophrac-tes), dry tropical forest (Tabebuia, Tecoma, Cybistax, Godmania), seasonal savanna (Ki-gelia), mangrove (Dolichandrone spathacea), and moist tropical forest (many genera). The lianas tend to be mostly from moist tropical forest, although some do grow in drier conditions, e. g. Cydista and Macfadyena.
Lianas are relatively rapidly growing climbers which produce very long stems and have limited vascular cambial activity (Schenck 1893; Dobbins 1971; Dobbins & Fisher 1986) whereas trees and shrubs grow more slowly and have greater cambial activity resulting in larger amounts of xylem and phloem. Because of their climbing habit, lianas are subject to different stresses than trees and shrubs. Lianas form an extensive canopy, and the distance water and solutes have to be transported is often greater than for trees (Ewers & Fisher 1989). Moreover, the control of development and subsequent anatomical features in Hanas are demonstrably different from those of trees and shrubs (Schenck 1893), and this paper documents some of these anatomical differences in the Bignoniaceae.
Materials and Methods Apart from some Hanas which were col
lected fresh from the Fairchild Tropical Garden, Florida, U.S.A. (FTG), all the material examined was from the following institutions: Jodrell Laboratory, Kew (KJw); museum collections, Kew (Kw); Institute of Systematic Botany, University of Utrecht, the Netherlands (Uw); Rijksherbarium, Leiden, the Netherlands (Lw); Universidad de Los Andes, Merida, Venezuela (MERw); Forest Products Laboratory, Madison, Wisconsin, U.S.A. (MADw and SJRw). The slides labelled FHOw are in the Jodrell Laboratory collection ( KJw), but originated from Oxford Forestry Institute. The abbreviations for wood collections follow those in Stern (1988). A complete list of the specimens examined is given in the Appendix (pages 415-417).
Microscopical observations of sectioned material were interpreted using the IAWA List of Features for Hardwood Identification
IAWA Bulletin n.s., Vol. 12 (4),1991
(lAWA Committee 1989) as a guide, with emphasis on those characters most pertinent to the comparison of trees with lianas in this family. These are: growth ring definition; vessel distribution, number, diameter, element length and intervascular pit size; fibre type (i.e. septate or non-septate) and wall thickness; axial parenchyma distribution; ray size and composition; storeying of tissues; presence or absence of anomalous secondary thickening.
In order to compare the frequency of these features in trees and lianas, Tables 1 and 2 (pages 398-407) were prepared. In the ten tree genera in which we examined three or more species, some features were of variable occurrence within the genus. For these genera, an index is given to denote the proportion of the genus with the feature (i.e., if three species out of five have a feature, the index is 0.6). The lianas are treated separately in Table 2. The indices give an indication of the variability of a feature within a given genus, and are used to calculate the overall percentage frequencies for the comparison of trees and Hanas, which are on a genus basis.
Information on whether the trees are deciduous or evergreen was rarely available on herbarium sheets or in the floras consulted (see the Appendix), except for Van Steenis (1977). Where available this information is given in Table 1. It is assumed that most of the Hanas are evergreen.
At least 25 measurements were made of each quantitative feature per sample, and these were bulked to obtain the means for species represented by more than one sample. For species with vessels of two distinct diameters, only the wider vessels were measured. Statistical analyses of mean vessel diameter (Fig. 22), mean vessel element length (Fig. 23) and intervascular pit border diameter (Fig. 24) were made using Kolmogorov-Smirnov Chi-square tests. These data, plus number of vessels per square mm were examined using an ANOV A. The relationship between mean vessel diameter in J.LM and vessel number per square mm is shown in Figure 25. The statistical results are given with the captions for Figures 22-25. Quantitative data are incomplete because some samples provided too few measurements for
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 391
inclusion in the tables and graphs; however, a sufficient number of samples was measured to compare trees and lianas in Figures 22-25 (see pages 411 & 412).
De8criptions The descriptions are in tribal order for
trees: Co1eeae, Crescentieae, Oroxyleae and Tecomeae, followed by lianas: Tecom<;:ae, Bignonieae and Schlegelieae. The Tecomeae trees and lianas are described separately, one after the other for easy reference. These tribal descriptions provide more detail and qualify some of the information given in Tables 1 (trees) and 2 (lianas). Since the anatomy of
some species and genera is not uniform, i.e. some genera have more than one feature state, the total percentage of genera with a feature may exceed 100% (e.g. in trees 48% have distinct. 55 % indistinct and 17 % absent growth rings which totals 120%. The same applies to axial parenchyma, where particular distribution patterns like scanty paratracheal, confluent and aliform often occur together and are not mutually exclusive). The final description outlines the features found in the family and refers to the tribes to which certain characters are restricted.
(text continued on page 397)
Legends of Figures 1-13 (trees) and 14-21 (lianas and climbers):
Figs. 1-4. Trees. - 1: Catalpa bignonioides TS. Ring-porous (scale as no.3). - 2: Jacaranda copaia TS. Diffuse-porous, winged aliform axial parenchyma (scale as no.3). - 3: Kigelia afri-cana TS. Diffuse-porous, lozenge aliform and eonfluent parenchyma (scale line = 500 111Il). - 4: Crescentia cujete TS, diffuse-porous, initial, confluent and aliform parenchyma (scale as no.3).
Figs. 5-9 Trees. - 5: Catalpa longissima TS. Diffuse-porous, confluent parenchyma (scale line = 500 J.Lm). - 6: Paratecoma peroba, the only tree with a tendency towards vessels of two distinct sizes (scale as no.5). -7: Oroxylum indicum TS, showing a vessel with a foraminate perforation plate and confluent parenchyma (scale line = 500 J.Lm). - 8: Markhamia sessilis TS, showing a vessel with a foraminate perforation plate (scale line = 100 111Il). - 9: Catalpa longis-sima lLS. Short vessel elements with large intervessel pits, axial parenchyma strands of two or more cells, septate fibres and short rays (scale line = 100 111Il).
Figs. 10-13 Trees. - 10: Dolichandrone spathacea TLS. Vessel elements with small intervessel pits, non-septate fibres, short uniseriate rays (scale line = 100 111Il). - 11: Tabebuia impetiginosa TLS. Tissues storied, vessel elements with large intervessel pits, short biseriate rays (scale as no. 10). - 12: Tabebuia caraiba RLS, showing rays tending towards heterocellular (scale line = 500 111Il). - 13: Deplanchea bancana RLS, showing heterocellular rays with one row of marginal square and upright cells (scale line = 100 111Il).
Figs. 14-17 Lianas. - 14: Macfadyena unguis-cati TS, showing the included phloem arrangement typical oflianas in the tribe Bignonieae (scale line = 1 mm). -15: Cydista aequinoctialis TS. Close up of staircase type phloem (like no.14) with very wide phloem sieve cells (scale line = 500 J.Lm). - 16: Pyrostegia venusta TS. Vessels of two distinct sizes (scale as no.15). - 17. Campsis radicans TS. A climber, not usually considered a liana. Ring-porous, without phloem wedges (scale as no.15).
Figs. 18-21 Lianas. -18: Schlegelia parasitica TS. Diffuse-porous, with sparse narrow vessels, and no phloem wedges (scale line = 500 111Il). -19: Phryganocydia corymbosa lLS. Very tall rays, septate fibres (scale as no.18). - 20: Arrabidaea corallina lLS. Short wide and narrow rays of two distinct sizes, septate fibres (scale as no.18). - 21: Schlegelia albiflora RLS. Heterocellular rays with more than one row of square and upright cells (scale as no.18).
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
392 IAWA Bulletin n.s., Vol. 12 (4),1991
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 393
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
394 IAWA Bulletin n.s., Vol. 12 (4),1991
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus Hanas in Bignoniaceae 395
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
396 IAWA Bulletin n.s., Vol. 12 (4),1991
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 397
Trees by Tribe: See Table 1.
Tribe Coleeae: Kigelia, Ophiocolea, Rhodo-colea. See Table 1.
Growth rings indistinct or absent. Diffuseporous.
Vessels solitary, in pairs and a few small clusters and radial multiples up to 6. .
Fibres non-septate, fibre walls thin to thick in Ophiocolea, thick in Kigelia and Rhodo-colea.
Axial parenchyma scanty paratracheal and diffuse in aggregates in Ophiocolea and Rho-docolea, some confluent in Rhodocolea; in Kigelia lozenge aliform to confluent (Fig. 3), tangential band width 5 cells or more, sometimes so wide that the ground tissue in some areas is entirely of axial parenchyma with no fibres.
Rays uniseriate, up to 20 cells high in Rhodocolea and 40 cells high in Ophiocolea, 2-4 cells wide and up to 15 cells high in Ki-gelia. Rays homocellular, but a few square and upright cells within rays of Ophiocolea, many unchambered ray cells containing one or more small rhomboidal crystals in Ophio-coleaonly.
Tribe Crescentieae: Amphitecna, Crescentia, Enallagma, Parmentiera. See Table 1.
Growth rings distinct in Parmentiera, otherwise indistinct or absent. Diffuse-porous.
Vessels solitary and in pairs in Amphitec-na and Crescentia, also in a few small clusters in Enallagma and Parmentiera.
Fibres non-septate. Axial parenchyma aliform and confluent
in all genera (Fig. 4), tangential bands 2-7 cells wide in Enallagma, initial in Crescentia and Parmentiera, also scanty paratracheal in Parmentiera. One to several small rhomboidal crystals in some unchambered axial parenchyma cells in Amphitecna and Crescentia cujete.
Rays uniseriate and up to 20 cells high in Amphitecna, uniseriate and biseriate and up to 15 cells high in other 3 genera; homocellular except in Parmentiera, which has some rays with one row of square marginal cells. One to several small crystals of various shapes in some unchambered ray cells in P.
cereifera, and one to several small rhomboidal crystals in some unchambered ray cells of C. alata (MADw 42263). Axial parenchyma and rays usually storied in Amphitecna and Crescentia, axial parenchyma irregularly storied in Enallagma.
Tribe Oroxyleae: Millingtonia, Oroxy1wn. See Table 1.
Growth rings indistinct to distinct. Diffuseporous.
Vessels solitary, in pairs, radial multiples and a few small clusters up to 5. Perforation plates simple, except for occasional foraminate plates in both genera (Fig. 7).
Fibres non-septate. Axial parenchyma confluent and initial,
also scanty paratracheal in Millingtonia and vasicentric and lozenge aliform in Oroxylum (Fig. 7).
Rays 2-3 cells wide and up to 15-30 cells high in Millingtonia, 1-4 cells wide and up to 25 cells high in Oroxylum, rays homocellular. One to several small crystals of various shapes in some unchambered ray cells in Millingtonia (KJw 3645), small rhomboidal crystals in some unchambered ray cells in Oroxylum.
Tribe Tecomeae: 26 genera. See Table 1.
Growth rings distinct in 18, indistinct in 15 and absent in 9 genera. Ring-porous in Catalpa bignonioides (Fig. 1) , C. bungei, C .. ovata and C. speciosa, Markhamia acumina-ta, M. sessilis and M. stipulata; semi-ring~
porous in Catophractes, Chilopsis and Mark-hamia zanzibarica; all other species diffuseporous (Figs. 5-7).
Vessels: Slight tendency towards vessels of two distinct sizes in some parts of Para-tecoma cross sections. Vessels solitary, in pairs, radial multiples and a few clusters up to 7 in most species, occasionally more, i.e. 8-17 in Dolichandrone, to 11 in Jacaranda, to 20 in Paratecoma. Radial multiples most common in Tabebuia chrysantha, T. chrysea, T. rosea and T. stenocalyx. Latewood vessels in clusters and tangential arrangement in Catalpa ovata and C. speciosa and Chilo psis. Perforation plates simple, except for occasio-
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
398 IAWA Bulletin n.s., Vol. 12 (4),1991
Table 1. Trees. Secondary xylem characters (98 species in 35 genera). Key: + = present Index: 1; all species in the genus have this feature; 0.1 to 0.9 is the proportion of species in the
genus with this feature.
~ E E.
~ £ £ ~ 01)
] " ~ 1) ~ u = li '" £ <: ~ '::1 = '" ~ '" " " ; '::1 :a '" 01) N
~ S eo .S:! B
.~ '" :I .;; .S 'S £ S .S ~ S " "0 l '" 0 = 0; £' '" '" '" :I ~ ~ S os. 9 £ B 01) 01) eo '"
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
404 IAWA Bulletin n.s., Vol. 12 (4),1991
Table 2. Lianas. Secondary xylem characters (39 species in 27 genera). Key: + = presenL Index: I, all species in the genus have this feature; 0.1 to 0.9 is the proportion of species in the
genus with this feature.
! ! ! t ...
£ Jj
.., - g
~ c ~ u .= C ., c ., 1;l ., .., .., .= :a .., ., bI)
N S S bI) :a ., :s .0; c .~ .5 i ·2 S .~
.., 0 .,
~ <l ., ., ., ~
:s ~ S "" .s, bI) bI) bI) ., e
~ 8- i i c c c :s 8. '- <l ·c ·c ·c [ bI) 0
., ., ., c b ~ .., .,
£ £ £ ·c ., Ol i > ..,
.~ ., <l c
~ t ~ ~ ~ 00 6 ., .>l to £ ~ ~ ~ ·8
., <l
., .., :a .., ..,
S S .5 ., > .c > Bignonieae Adenocalymna alliaceum + + 85 55 232 4.7
Fibres thin-walled in Tabebuia cassinoides and T. rosea; thick-walled in Catophractes, Cotema, Ekmanianthe, Phyllarthron, Tecoma-ria and Zeyheria; very thick-walled in Tabe-
'" = @-., u
13 <a I .5 .~ ~ e
'" S S e i i) .... '" ~ .E '" 0 ., N = '" .;;; § ;; .,
~ g ~ e 0
-5 ~ ~ ~ e '<t .~ ~ ., :9 .... g ..c: A ~ 0 ..c: :9 '" '" '" '" >. ~ e- e- >. >. .8 e e e ....
buia barbata, T. capitata, T. cf. catinga, T. chrysantha, T. guayacan and T. serrati/olia, Tecoma pedicellata, and T. spectabilis. Fibres entirely non-septate in 15 and in some spe-
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
408
cies of another 5 genera. Most if not all fibres septate in Catalpa ovata, C. speciosa, C. den-ticulata, C. longissima (Fig. 9) and C. punc-tata, Delostoma, Fernandoa, Spathodea gi-gantea, Tecoma castanifolia, T. garrocha, T. stans and Tecomaria. At least some fibres septate in Markhamia acuminata and M. sessilis, Pajanelia, Radermacheragigantea andR. sini-ca, Romeroa, Spathodea campanulata and S. serratula, Stereospermumfimbriatum, Teco-ma grandis, T. serrati folia and T. undulata.
Axial parenchyma paratracheal in all species. Some vasicentric in Catalpa ovata, Ek-manianthe, Pajanelia, Paratecoma, Phyllar-thron, Radermachera, Spathodea serratuia, Stereospermum neuranthum and S. xylo-carpum, Tabebuia angustata and T. roseo-alba. Diffuse in aggregates only in Ekman-ianthe. Scanty paratracheal in 22 genera (not in Deplanchea, Godmania, Newbouldia and Pajanelia). Alifonn in 14 genera (Fig. 2). Con-fluent in 19 genera. Unilateral paratracheal in Jacaranda copaia, J. cuspidifolia, J. glabra, J. mimosifolia and J. obtusifolia. Initial in 16/26 genera. Tangential bands less than 3 cells wide in Deplanchea and Dolichandrone, more than 3 cells wide in Dolichandrone atrovirens, D. heterophylla and D. spathacea, Godmania and Spathodea campanulata.
Rays mainly 1-3 cells wide. Uniseriate in Dolichandrone spathacea (Fig. 10), Jacaranda caucana, J. coeruIea, J. hesperia, J. mimosi-folia and J. obtusifolia, Phyllanhron, Spath-odea serratula, Tabebuia caraiba and T. cas-sinoides; 1-2 cells wide in some species in 13 genera. Biseriate in Radermachera gigan-tea, Stereospermum fimbriatum, Tabebuia capitata and T. impetiginosa (Fig. 11), Teco-ma castanifolia, T. conspicua, T. serratifolia and T. spectabilis. Rays up to 10 cells high in Cotema, Phyllanhron, Tabebuia angustata, T. barbata, T. chrysantha, T. guayacan, T. haemantha and T. serratifolia, Tecoma con-spicua, T. pedicel/ata, T. spectabilis and Zey-heria. Rays up to 20 cells high in some species of 16 genera, up to 21-50 cells high in some species of 12 genera and up to 70 cells high in Markhamia sessilis. Rays mainly homocellular in 22 genera, but heterocellular, with one or more rows of square or upright marginal cells (Fig. 13), or square and upright cells within the body of the rays in 20
IAWA Bulletin n.s., Vol. 12 (4),1991
genera (Fig. 12). Rays of two distinct sizes in Romeroa, almost so in Markhamia sessilis, Spathodea campanulata and S. gigantea.
Crystals of various shapes, several per cell in non-chambered axial parenchyma in Pajanelia, and in ray cells in 8 genera, a single crystal per ray cell in Markhamia zanzibarica. Storied in Cotema, Godmania, Tabebuia angustata, T. barbata, T. capitata, T. cf. catinga, T. chrysantha, T. guayacan, T. haemantha, T. impetiginosa (Fig. 11), T. pallida, T. rosea, T. roseo-alba and T. ser-ratifolia, Tecoma conspicua, T. grandis, T. leucoxylon, T. pedicellata and T. spectabilis and Zeyheria. Some tendency towards storeying of axial parenchyma in Catalpa, especially C. ovata, and of some parenchyma in Tecoma aurea, and irregularly storied in rays of T abebuia caraiba and Tecoma serrati folia.
Lianas by Tribe: See Table 2.
Tribe Tecomeae: Campsis, Tecomanthe. See Table 2.
Growth rings indistinct in Tecomanthe, distinct in Campsis. Ring-porous in C. radi-cans (Fig. 17), diffuse-porous in C. chinen-sis and Tecomanthe.
Vessels of two distinct sizes in C. chinen-sis. Vessels solitary, in pairs, radial multiples and a few clusters up to 6, but longer radial multiples are abundant in smaller vessels in C. chinensis and latewood vessels in C. radi-cans. Helical thickenings in narrow vessels in Campsis.
Fibres septate in Campsis, some fibres septate in Tecomanthe, thin- to thick-walled.
Axial parenchyma scanty paratracheal. Rays 1-4 cells wide and up to 70 cells
high. Rays heterocellular with several rows of square and upright cells at ray margins in Campsis and mixed within the body of the rays in Tecomanthe. Several tiny needle-like crystals present in some ray cells in Teco-manthe. Axial parenchyma and narrow vessels storied in Campsis.
Cambial variants absent, although fOUT
slight indentations occur in cambium ofTe-comanthe, which could possibly develop into a similar variant to that in Bignonieae, but no wider stems available for examination.
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 409
Tribe Bignonieae: 24 genera. See Table 2.
Growth rings distinct in 9, indistinct in 23 genera.
Vessels solitary, in pairs and often frequent clusters and radial multiples up to 12. Vessels of two distinct sizes (Fig. 16) in all species except Adenocalymna and Anemopaeg-ma robustufn. Vasicentric tracheids sOpletimes present in association with narrow vessel elements.
Fibres septate and thin- to thick-walled, except in Callichlamys and Tanaecium which have non-septate and thin-walled fibres.
Axial parenchyma scanty paratracheal and vasicentric (poorly defined) in all genera, also confluent in Arrabidaea chica, A. corallina and A. mollissima and initial in Arrabidaea pubescens and A. triplinervia, Memora flavi-flora, Neomacjadya, Petastoma broadwayi, Pithecoctenium dolichoides, Tanaecium, Tyn-nanthus and Urbanolophium.
Rays 1-16 cells wide. Rays 1-2 cells wide in Callichlamys, Macrodiscus artd Saritaea, up to 3 cells wide in Arrabidaea pubescens, Neomacfadya, Pleonotoma and Pyrostegia, up to 50 cells high in Arrabidaea chica, A. corallina, Macrodiscus, Neomacfadya and Pleonotoma, up to 50-100 cells high in Ar-rabidaea mollissima and A. pubescens, Cal-lichlamys, Petastoma broadwayi, Pyrostegia, Saritaea, Tanaecium and Tynnanthus micran-thus. Other species with rays up to 101-600 cells high (Fig. 19). Rays of two distinct sizes except in Adenocalymna, Arrabidaea chica, Callichlamys, Macrodiscus, Neomac-fadya, P/eonotoma, Pyrostegia and Saritaea. Rays mainly heterocellular, often mixed or with many rows of square and upright cells. Axial parenchyma more or less storied in Ar-rabidaea triplinervia, Chodanthus, Cydista, Macfadyena (with small rays), Mansoa (with short fibres), Pithecoctenium and Tynnanthus elegans.
One or more crystals of various shapes in unchambered ray cells in Adenocalymna and Tanaecium.
Cambial variant in the form of phloem indentations (wedges) present in all species (Figs. 14, 15). Four indentations in Adeno-calymna, Arrabidaea, Chodanthus, Memora, Neomacfadya, Petastoma, Pithecoctenium,
Pyrostegia, Saritaea, Tanaecium and Tynnan-thus; eight or more indentations in Cydista, Macfadyena (Fig. 14), Mansoa, Paragonia, Phryganocydia and Pseudocalymma; number of indentations could not be determined for some genera because of the samples' incomplete cross sections. Phloem indentations stepped (i. e. staircase-like) in Anemopaegma, Ar-rabidaea pubescens and A. triplinervia, ' Cydis-ta (Fig. 15), Macjadyena, Mansoa, Memora, Mussatia, Paragonia, Phryganocydia, Pithe-coctenium dolichoides, Pleonotoma, Pseudo-calymma, Tynnanthus and Urbanolophium. Sieve cells in indentations much larger than those in normal phloem in Arrabidaea chica, Cydista, Neomacjadya, Petastoma patellife-rum, Pithecoctenium crucigerum, Tanaecium jaroba and Tynnanthus elegans, and slightly so in Saritaea.
Tribe Schlegelieae: Schlegelia. See Table 2.
Growth rings indistinct to distinct Diffuseporous.
Vessels of two distinct sizes in S. albiflora and S. nicaraguensis, but not S. parasitica (Fig. 18). Vessels solitary and in pairs in S. albiflora, also in radial multiples and a few clusters up to 6 in S. nicaraguensis and S. parasitica.
Fibres non-septate, except for some septate fibres in S. parasitica, thin-walled.
Axial parenchyma scanty paratracheal and vasicentric, also initial in S. albiflora and S. nicaraguensis.
Rays 1-2 cells wide and up to 30 cells high in S. parasitica; 1-3 cells wide in S. ni-caraguensis and 1-6 cells wide in S. albiflora, up to 55 cells high in both. Rays heterocellular, with rays composed of mixed procumbent, square and upright cells and several rows of square and upright cells at ray margins (Fig. 21).
No storeying or cambial variants.
Family Description
Trees: 35 genera; 3 genera and 3 species of Coleeae; 4 genera and 7 species of Crescentieae; 2 genera and 2 species of Or-oxyleae, 26 genera and 87 species of Tecomeae.
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
410
Lianas: 27 genera; 24 genera and 33 species of Bignonieae, 1 genus and 3 species of Schlegelieae, 2 genera and 3 species of Tecomeae.
Growth rings distinct, indistinct or absent. Most species diffuse-porous, but some species of Tecomeae ring-porous or semi-ringporous.
Vessels solitary, in pairs, clusters and radial multiples of varying number. Vessel clusters commoner in lianas and ring-porous trees. Vessels of two distinct sizes almost always present in lianas but virtually absent in trees. Intervessel and vessel-ray pitting alternate and of the same size, perforation plates exclusively simple, except for very occasional foraminate plates in some species of Oroxyleae and Tecomeae. Helical thickenings rare, only found in narrow vessels in ring, semi-ring-porous or diffuse-porous (with vessels of two distinct sizes) Tecomeae trees and lianas. Vasicentric tracheids present in some Bignonieae lianas.
Fibres either non-septate (in most trees) or septate (in most lianas), occasionally both types present. Simple or very minutely bordered pits usually present in the radial fibre walls. Fibre walls usually thin to thick, occasionally thin or very thick.
Axial parenchyma mainly scanty paratracheal, vasicentric, aliform, confluent and initial in trees, scanty paratracheal and vasicentric (poorly defined) in lianas.
Rays 1-5 cells wide and up to 50 cells high in trees, 1-16 cells wide and usually over 20 and up to 600 cells high in lianas. Rays mainly homocellular in trees, heterocellular in lianas. Rays of two distinct sizes common in lianas but virtually absent in trees. Storied or irregularly storied tissues equally uncommon in trees and lianas, but characteristic of certain genera of Crescentieae and Tecomeae trees.
Crystals uncommon, usually small and of varying shape, several per non-chambered ray or less commonly non-chambered axial parenchyma cell. Prismatic crystals even rarer.
Cambial variant in the form of phloem indentations restricted to Bignonieae lianas.
IAWA Bulletin n.s., Vol. 12 (4),1991
Discussion This paper outlines some of the overall
wood anatomical differences between trees and lianas, and for the first time provides accounts of the wood anatomy of all the woody tribes in the Bignoniaceae.
The wood anatomical features of Bignoniaceae trees and lianas fall into three groups: 1) features restricted to or more common in trees, 2) features restricted to or more common in lianas, and (3) features equally common in trees and lianas, including those common to the entire family.
1) Features restricted to or more common in trees - Rays exclusively uniseriate (12%), and the occurrence of very infrequent foraminate perforation plates in a scattering of species from Oroxyleae and Tecomeae come into this category (although Handa, 1936, records foraminate plates in Campsis grandi-flora, which would be treated as a liana in this paper). Growth rings are absent in 19% of trees. In lianas they are always present, but are often indistinct. Aliform (50%) and initial parenchyma (49%) were recorded only in trees and confluent parenchyma was found in 70% of trees and only 2% oflianas. Banded parenchyma (up to 3 cells wide in 8% and over 3 cells wide in 12%) was found only in trees, and so were rays up to 20 cells high (72%).
2) Features restricted to or more common in lianas - Most of the lianas have four or eight wedges of phloem resulting from the failure of the cambium to produce secondary xylem while continuing phloem production (Dobbins 1971). This cambial variant was restricted to Bignonieae in our samples, and Gentry (1980) believes it is restricted to this tribe. The only sample of Tecomanthe (Tecomeae) available was very narrow, only 13 mm in diameter including bark, and had four very slight phloem indentations. Wider stems of Tecomanthe would be needed to see whether phloem wedges occur, which would extend their occurrence outside the Bignonieae. Internal phloem has been recorded in Campsis (Tecomeae) by Handa (1936), but this is an entirely different variant from the phloem wedges in Bignonieae.
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 411
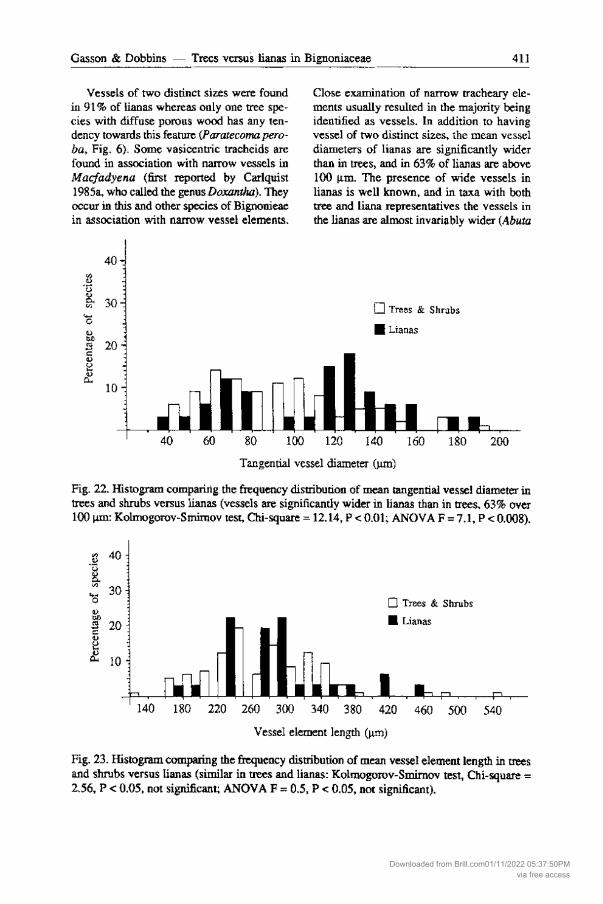
Vessels of two distinct sizes were found in 91 % of lianas whereas only one tree species with diffuse porous wood has any tendency towards this feature (Paratecoma pero-ba, Fig. 6). Some vasicentric tracheids are found in association with narrow vessels in Macfadyena (first reported by Carlquist 1985a, who called the genus Doxantha). They occur in this and other species of Bignonieae in association with narrow vessel elements.
40 r/J G)
'<3 G) c. 30 Vl
..... 0 G) OJ)
20 .:s i3 8 &
10
40 60 80 100
Close examination of narrow tracheary elements usually resulted in the majority being identified as vessels. In addition to having vessel of two distinct sizes, the mean vessel diameters of lianas are significantly wider than in trees, and in 63% of lianas are above 100 11m. The presence of wide vessels in lianas is well known, and in taxa with both tree and liana representatives the vessels in the lianas are almost invariably wider (Abuta
o Trees & Shrubs
• Lianas
120 140 160 180 200
Tangential vessel diameter (~)
Fig. 22. Histogram comparing the frequency distribution of mean tangential vessel diameter in trees and shrubs versus lianas (vessels are significantly wider in lianas than in trees, 63% over 100~: Kolmogorov-Smirnov test, Chi-square = 12.14, P < 0.01; ANOVA F = 7.1, P < 0.008).
r/J 40 .~ u
G) C. r/J
30 ..... 0 o Trees & Shrubs G) OJ) • Lianas 00 20 E G)
~ c.. 10
140 180 220 260 300 340 380 420 460 500 540
Vessel element length (~)
Fig. 23. Histogram comparing the frequency distribution of mean vessel element length in trees and shrubs versus lianas (similar in trees and lianas: Kolmogorov-Smirnov test, Chi-square = 2.56, P < 0.05, not significant; ANOV A F = 0.5, P < 0.05, not significant).
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
412
Percentage of species
40
30
20
10
o. j CIl
2345678910 Intervessel pit diameter (J.1ffi)
Fig. 24. Histogram comparing the frequency distribution of mean intervessel pit border diameter in trees and shrubs versus lianas (intervessel pits are significantly larger in lianas than in trees, 50% over 6 j.Lm: Kolmogorov-Smirnov test, Chi-square = 10.18, P < 0.01).
grandijlora, Menispermaceae, Mennega 1982; Zanthoxylum, Rutaceae, Caesalpinia and Bauhinia, Leguminosae, Bamber 1984; Carlquist 1985b; Ewers et al. 1990). Lianas also have significantly more vessels per unit area than trees (P < 0.0001). The relationship between vessel density and mean vessel diameter in trees and lianas is shown in Fig. 25. The intervascular pits of lianas are also significantly larger than in trees. Half of the liana species have pit borders exceeding 6 11m.
All these vessel features suggest that Bignoniaceae lianas are much more efficient at water conduction than trees. Water conductivity is directly proportional to the fourth power of the radius of a vessel or capillary, so in identical conditions one vessel twice the diameter of another will transport sixteen times as much water (Zimmermann 1983; Ewers & Fisher 1989). The greater density of vessels per unit area and larger intervascular pits would further enhance the ability of a liana to transport water, and the presence of
IAWA Bulletin n.s., Vol. 12 (4),1991
both wide and narrow vessels means that even if some wide vessels become blocked by embolism the narrower ones would remain functional. Vessel element length is similar in both groups regardless of vessel diameter and would be expected to have less influence than vessel diameter on water flow.
Septate fibres are much more common in lianas (83%) than trees (20%) and this seems to be linked with a lower frequency of axial parenchyma and the almost total absence of bands of parenchyma in lianas. Spackman
Trees versus Lianas
Mean vessel diameter in J.1ffi 200,---------------------------,
Fig. 25. Scatter diagram showing the relationship between mean tangential vessel diameter (11m) and number of vessels per square mm in trees and lianas. Trees: closed triangles, y = 117.7 - 2.279 x, r = -0.6184 (p <0.0001). Lianas: open squares, y = 158.49 - 1.997x, r = 0.4557 (p < 0.005). There are significantly more vessels per unit area in lianas, usually more than 20 per square mm: ANOV A F = 51.6, P < 0.0001.
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 413
and Swamy (1949) observed that there is little axial parenchyma in species with many septate fibres, but this is not a simple or universal relationship (Chalk 1983). Septate fibres are found in many taxonomic groups and in plants growing in a wide range of temperate and tropical habitats. Baas and Schweingruber (1987) consider that septate fibres are largely restricted to tropical families (using observations and data from Wheeler et aI. 1986; Metcalfe & Chalk 1950). Fahn et aI. (1986) showed that in Israel, septate fibres occur most frequently in climbers. Wolkinger (1970) and Fahn (1982) also found living protoplasts in both non-septate and septate fibres, which are probably longer-lived in the septate fibres allowing sufficient time for septa to develop.
Rays of two distinct sizes were found in 63% of lianas and only 5% of trees. Rays are wider, taller (usually over 20 cells high, and often much taller) and more heterocellular in lianas than in trees. Large heterocellular rays are considered by Carlquist (1962, 1988) to be a juvenile characteristic. All the lianas we examined were from relatively small stems up to about 5 cm in diameter, so there is a strong possibility that this influenced in part the high incidence of heterocellular rays in the lianas. It seems likely that some lianas have large rays because they receive support from other plants, and do not need a high proportion of fibres to support them. Another possibility is that rays may be important in accommodating the physical torque and stress placed on climbers, as shown by Fisher and Ewers (1989).
3) Features equally common in trees and lianas - There are several features in this category, i.e. distinct growth rings (43% v. 32%), diffuse-porous (97% v. 94%), vessels not solitary, simple perforation plates (all species), alternate intervascular pitting and vessel-ray pitting of the same size (all species), similar vessel element length and paratracheal parenchyma. Fibre wall thickness is similar in the two groups, although none of the lianas have very thick-walled fibres whereas 1 % of the trees do.
The frequency of some of these characters has been compared with the same characters in 4,080 woods from many families world
wide (Wheeler et al. 1986). All Bignoniaceae have alternate intervascular pitting and simple perforation plates in the vessels, as do 87% and 89% of taxa worldwide. The presence of distinct growth rings is higher for Bignonia-ceae than taxa worldwide (48% trees, 32% lianas versus 31 % worldwide). Likewise, vessels not solitary (all trees and lianas versus 83% worldwide), paratracheal parenchyma (100% trees and lianas versus 50% worldwide), storied tissues (18% trees, 25% lianas versus 12% worldwide), and rays three or more cells wide (60% trees, 86% lianas versus 39% worldwide) are greater for Bignoniaceae than world woods.
Acknowledgements
D.F. Cutler, P.J. Rudall, E.A. Wheeler and G.D. Wallace suggested numerous improvements to the manuscript. Angela Pearson and Alison Hughes sectioned much of the material. M. Simmonds provided the statistical analyses. R. Brummitt, D. Hunt, M. Wilmot-Dear, S. Bidgood and A. Gentry advised on the many problems associated with the taxonomy and nomenclature of the family.
This work was supported in part by a Faculty Grants Award from Millersville University and a sabbatical leave to David R. Dobbins.
Baas, P. & F.H. Schweingruber. 1987. Ecological trends in the wood anatomy of trees, shrubs and climbers from Europe. IAWA Bull. n.s. 8: 245-274.
Bamber, R.K. 1984. Wood anatomy of some Australian rainforest vines. Proceedings of Pacific Regional Wood Anatomy Conference October 1-7, 1984, Tsukuba, Japan: 58-60.
Carlquist, S. 1962. A theory ofpaedomorphosis in dicotyledonous woods. Phytomorphology 12: 30-45.
Carlquist, S. 1985a. Vasicentric tracheids as a drought survival mechanism in the woody flora of southern California and similar regions; review of vasicentric tracheids. Aliso 11: 37-68.
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
414
Carlquist, S. 1985b. Observations on functional wood histology of vines and lianas: vessel dimorphism, tracheids, vasicentric tracheids, narrow vessels, and parenchyma. Aliso 11: 139-157.
Carlquist, S. 1988. Comparative wood anatomy: Systematic, ecological and evolutionary aspects of dicotyledon wood. Springer Verlag, New York.
Chalk, L. 1983. Wood structure. In: Anatomy of the Dicotyledons, 2nd Ed. Vol. 2, Ox-ford University Press.
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press, New York.
Diniz, M.A. 1988. Family 124: Bignoniaceae. In Flora Zambesiaca. Vol. 8 Part 3: 61-85.
Dobbins, D.R. 1971. Studies on the anomalous cambial activity in Doxantha unguiscati (Bignoniaceae). ll. A case of differential production of secondary tissues. Amer. J. Bot. 58: 697-705.
Dobbins, D.R. & J.B. Fisher. 1986. Wound responses in girdled stems of lianas. Botanical Gazette 147: 278-289.
Ewers, F. & J.B. Fisher. 1989. Variation in vessel length and diameter in stems of six tropical and subtropical lianas. Amer. J. Bot. 76: 1452-1459.
Ewers, F., J.B. Fisher & S.T. Chiu. 1990. A survey of vessel dimensions in stems of tropical lianas and other growth forms. Oecologia 84: 544-552.
Fahn, A. 1982. Plant Anatomy. 3rd Ed. Pergamon Press, Oxford, New York.
Fahn, A., E. Werker & P. Baas. 1986. Wood anatomy and identification of trees and shrubs from Israel and adjacent regions. The Israel Academy of Sciences and Humanities, Jerusalem.
Fisher, J.B. & F.W. Ewers. 1989. Wound healing in stems of lianas after twisting and girdling injuries. Botanical Gazette 150: 251-265.
Gentry, A.H. 1973. Generic delimitations of Central American Bignoniaceae. Brittonia 25: 226-242.
Gentry, A.H. 1977.178. Bignoniaceae. Flora of Ecuador No.7. Berlings, Lund.
IAWA Bulletin n.s., Vol. 12 (4),1991
Gentry, A.H. 1979. Distribution patterns of neotropical Bignoniaceae: some phytogeographical implications. In Tropical Botany (eds. K. Larsen & L.B. Holm-Nielsen): 339-354. Academic Press, London, New York, San Francisco.
Gentry, A.H. 1980. Bignoniaceae. Part 1 (Crescentieae and Tourrettieae). Flora Neotropica Monograph 25.
Gregory, M. 1980. Wood identification: an annotated bibliography. IA WA Bull. n. s. 1: 3-41.
Handa, T. 1936. Abnormal vascular bundle in the stem of Campsis grandiflora K Schum. Japan. J. Bot. 8: 47-58.
IA WA Committee. 1989. IA WA list of microscopic features for hardwood identification with an appendix on non-anatomical information.IAWA Bull. n.s. 10: 219-332.
Jain, D.K. & V. Singh. 1980. Studies in Bignoniaceae. VIT. Wood anatomy. Proceedings of the Indian Academy of Sciences. Plant Science 89: 443-456.
Liben, L. 1977. Bignoniaceae. In: Flora d' Afrique Centrale (Zaire, Rwanda, Burundi), Belgium.
Mennega, A.M.W. 1982. Stem structure of the New World Menispermaceae. J. Arnold Arbor. 63: 145-171.
Metcalfe, C.R. & L. Chalk. 1950. Anatomy of the Dicotyledons. Clarendon Press, Ox-ford.
Outer, R.W. den & W.L.H. van Veenendaal. 1983. Wood anatomy of Un carina leandrii H. Humb. (Pedaliaceae) and its relation to Bignoniaceae. IA WA Bull. n. s. 4: 53-59.
Perrier de La Bathie, H. 1938. Flore de Madagascar. Family 178: Bignoniaceae.
Record, S.J. & R.W. Hess. 1943. Timbers of the New World. Yale University Press, New Haven.
Santos, G. 1990. Systematic wood anatomy of Tecomeae (Bignoniaceae). Unpublished manuscript. Thesis, University of Missouri, St. Louis.
Santos, G. In press. Systematic wood anatomy of Tecomeae. In A.H. Gentry (ed.), Bignoniaceae Part 2 (Tecomeae). Flora Neotropica Monograph.
Downloaded from Brill.com01/11/2022 05:37:50PMvia free access
Gasson & Dobbins - Trees versus lianas in Bignoniaceae 415
Schenck, H. 1893. Beitriige zur Anatomie der Lianen. In: A.F.W. Schimper (ed.): 1-271. Bot. Mitt. aus den Tropen. Heft 5, Teil2. Gustav Fischer, Jena.
Spackman, W. & B.G.L. Swamy. 1949. The nature and occurrence of septate fibres in dicotyledons. Amer. 1. Bot. 36: 804 (abstract).
Sprague, T. 1906. Flora of Tropical Africa. Vol. IV, Sect. 2, Hydrophyllaceae to. Pedaliaceae. XCVI, Bignoniaceae: 512-538.
Steenis, C.G.G.J. van. 1977. Bignoniaceae. In Flora Malesiana I, 8 (2): 114-186. Sijthoff & Noordhoff, The Netherlands.
Stem, W. L. 1988. Index Xylariorum 3. Institutional wood collections of the world. IAWA Bull. n.s. 9: 203-252.
Takhtajan, A. 1987. Systema Magnoliophytorum. Academia Scientiarum U.R.S.S., Leningrad.
Wheeler, E.A., R.G. Pearson, C.A. La Pasha, T. Zack & W. Hatley. 1986. Computer-aided Wood Identification. Reference Manual. North Carolina Agricultural Research Service Bulletin 474.
Willis, J. C. 1973. A dictionary of the flowering plants. Revised by H. K. Airy Shaw. 8th Ed. Cambridge Univ. Press.
Wolkinger, F. 1970. Das Vorkommen lebender Holzfasem in Striiuchem und Bliumen. Phyton (Austria) 14: 55-67.
Zimmermann, M.H. 1983. Xylem structure and the ascent of sap. Springer Verlag, Berlin, Heidelberg, New York, Tokyo.
APPENDIX
The species examined are listed below. The country or geographical region of origin is that from which the specimen came, not necessarily its native habitat. If the exact source of the specimen is not known, but the native region is, this is in parentheses. Synonymy is taken from Diniz (1988), Gentry (1973, 1977, 1979, 1980), Perrier de la B§.thie (1938), Sprague (1906), Liben (1977), Van Steenis (1977), and herbarium specimens at Kew. The tribal affinity for each genus is taken from Gentry (1980).
TREES:
Coleeae: Kigelia africana, Lw (Lewalle 5327, Bu
rundi), KJw Nigeria, Kw 13640, K. africana (labelledK.pinnata) Mozambique, Kw 13643, K. africana (labelled K. pinnata) S Africa, Uw 22020, K. africana (labelled K. aethio-pum) Ethiopia (Eritrea), Kw 13639. - Ophio-coleafloribunda (labelled Coleafloribunda), Madagascar, Kw 13578. - Rhodocolea tel-fairiae (labelled Colea teljairiae), Madagascar, SJR w10766.
cucurbitina), USA (Florida), Kw 13585. -Crescentia alata, Mexico, SJRw 9599; C. cu-jete, Guyana, Kw 13580; C. cujete (labelled C. acwninata), Jamaica, Kw 13579. - Enal-lagma donnell-smithii, Costa Rica, SJRw
38355; E. latifolia, (New World), MADw 16510. - Parmentiera cereifera, (New World), Kw 13675; P. macrophylla, Malaysia, SJRw 38465. N.B.: Enallagma may be considered part of