1.4.2 Retinoid signaling in Hematopoiesis .....................................................................18
1.4.3 Functions of PML and PML-RARA ......................................................................34
1.4.4 Functions of NPM and NPM-RARA .....................................................................37
1.4.5 Functions of NuMA and NuMA-RARA ...............................................................45
1.5 Secondary genetic events in APL ......................................................................................47
1.5.1 Candidate gene alterations as approaches to elucidate secondary genetic events in APL .........................................................................................................47
1.5.2 Whole genome approaches to determining secondary events ...............................50
viii
1.5.3 Use of high throughput assays to understand leukemia pathogenesis ...................52
1.5.4 ChIP-seq and ChIP-on-chip technology: Global analysis of DNA binding profiles ...................................................................................................................56
2.4.1 APL blasts exhibit deregulated expression of NF-κB signaling genes..................75
ix
2.4.2 Primary APL cells and cell lines U937-NPM-RARΑ and NB4 over-express NF-κB/p65. ............................................................................................................84
2.4.3 TNFα inhibits colony formation and cellular survival in U937 control, but not X-RARΑ+ cells. .....................................................................................................87
2.4.4 Enhanced NF-κB/p65 activation upon TNFα signaling in X-RARA cells. ..........90
2.4.5 TNFα induces a distinct NF-κB/p65 target gene expression profile in X-RARΑ+ cells compared to controls. ......................................................................93
2.4.6 Treatment with ATRA restores wild type TNF α cell viability response in APL. .......................................................................................................................93
2.4.7 Pharmacological inhibition of p65 restores TNFα sensitivity in NPM-RARΑ+ cells. .......................................................................................................................99
3 Comparative analysis of downstream genetic targets of the variant Acute Promyelocytic Leukemia fusion proteins NPM-RARA and NuMA-RARA ..................................................111
3.4.1 Transcriptional targets of NPM-RARA and NuMA-RARA are involved in diverse cellular functions. ....................................................................................118
x
3.4.2 Promoter sequences of X-RARA gene targets show over-representation of binding motifs for transcription factors including RELA and FOX. ...................130
3.4.3 A subset of downstream gene targets of NPM-RARA contains regulatory motifs that are directly bound by PML-RARA and PLZF-RARA. .....................136
3.4.4 Potential direct transcriptional targets of NPM-RARA are functionally involved in diverse cellular processes..................................................................137
3.4.5 Potential direct transcriptional targets of NPM-RARA contain known RARE binding sites as well as PU.1 binding sites. .........................................................137
3.4.6 Comparison of NPM- and NuMA-RARA expression profiles with PML-RARA. .................................................................................................................150
3.4.7 NPM-RARA and NuMA-RARA induced gene signatures overlap with AML LSC. .....................................................................................................................150
3.4.9 Gene expression based signature identifies compounds that confer expression changes that are correlated with ATRA treatment in APL cells. .........................170
4.4.1 NPM is aberrantly expressed in NPM-RARA and PML-RARA cells. ...............185
4.4.2 NPM is post-translationally stabilized in NPM-RARA+ cells. ...........................198
4.4.3 NPM-RARA+ cells have a cell growth phenotype consistent with disrupted nucleolar function. ...............................................................................................203
4.4.4 Assessment of pre-rRNA and 18S rRNA expression in APL patient samples. ...203
4.4.5 NPM and X-RARA interact with RXRA in COS X-RARAV5 cells. ...................211
5.2 Downstream genetic targets of the acute promyelocytic leukemia fusion proteins NPM-RARA and NuMA-RARA. ....................................................................................223
VAD Vitamin A Deficient VDR Vitamin D Receptor VDRE Vitamin D Response Element WHO World Health Organization WNT Wingless-type MMTV integration site family X-RARA X-Retinoic Acid Receptor Alpha
xvi
List of Tables
Chapter 1: Introduction
Table 1.1: WHO/FAB classification of acute leukemias
Table 1.2: Evidence for the role of RXR in myelopoeisis
Table 1.3: RAR and RXR deficient models in hematopoiesis
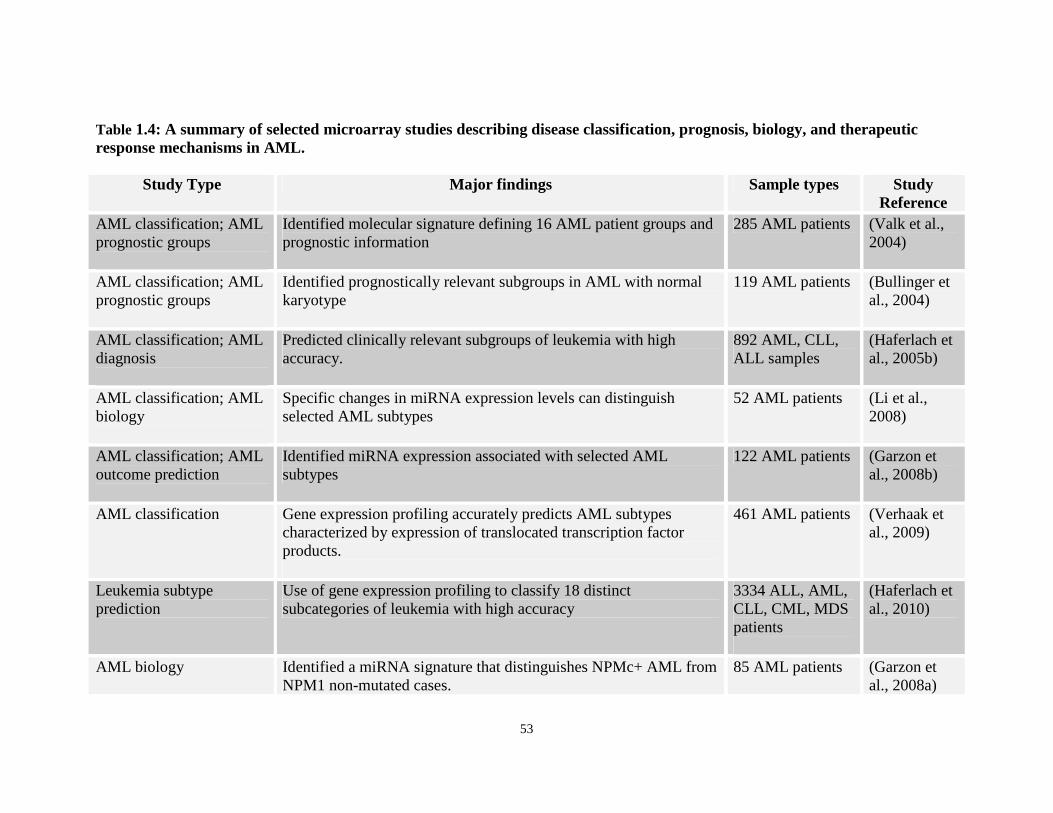
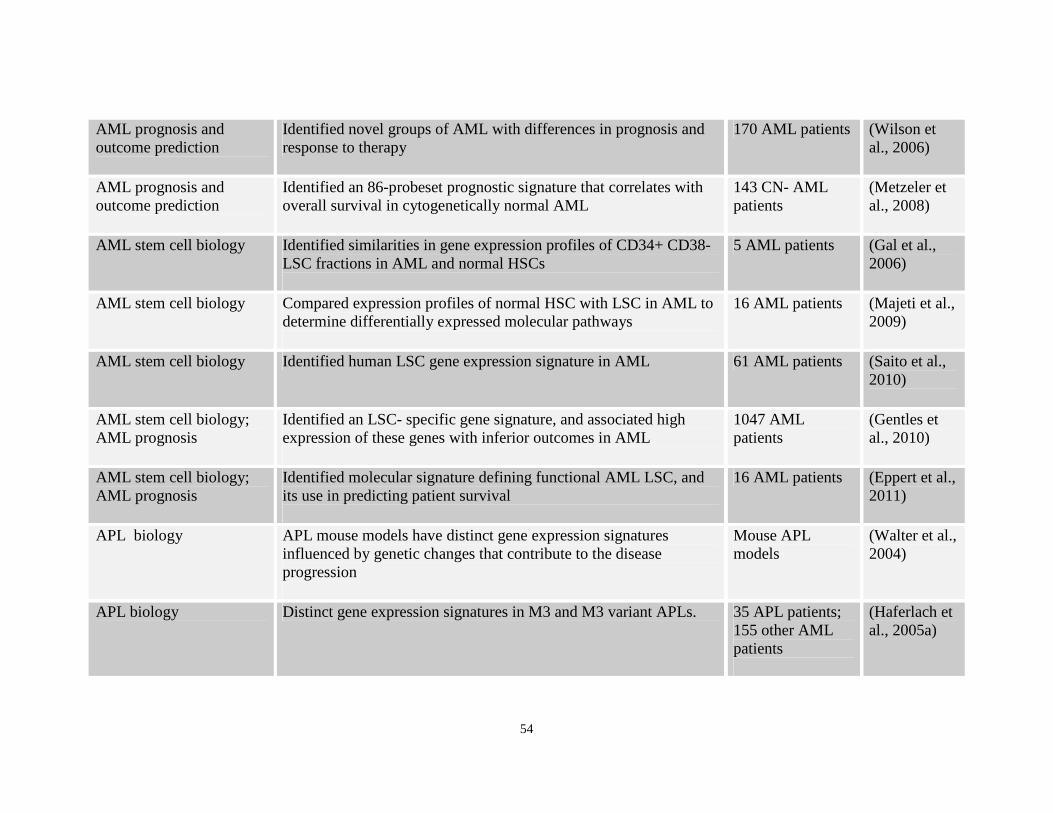
Table 1.4: Array studies demonstrating use of gene expression studies in AML
Chapter 2: Deregulated NF-κκκκB signaling in APL
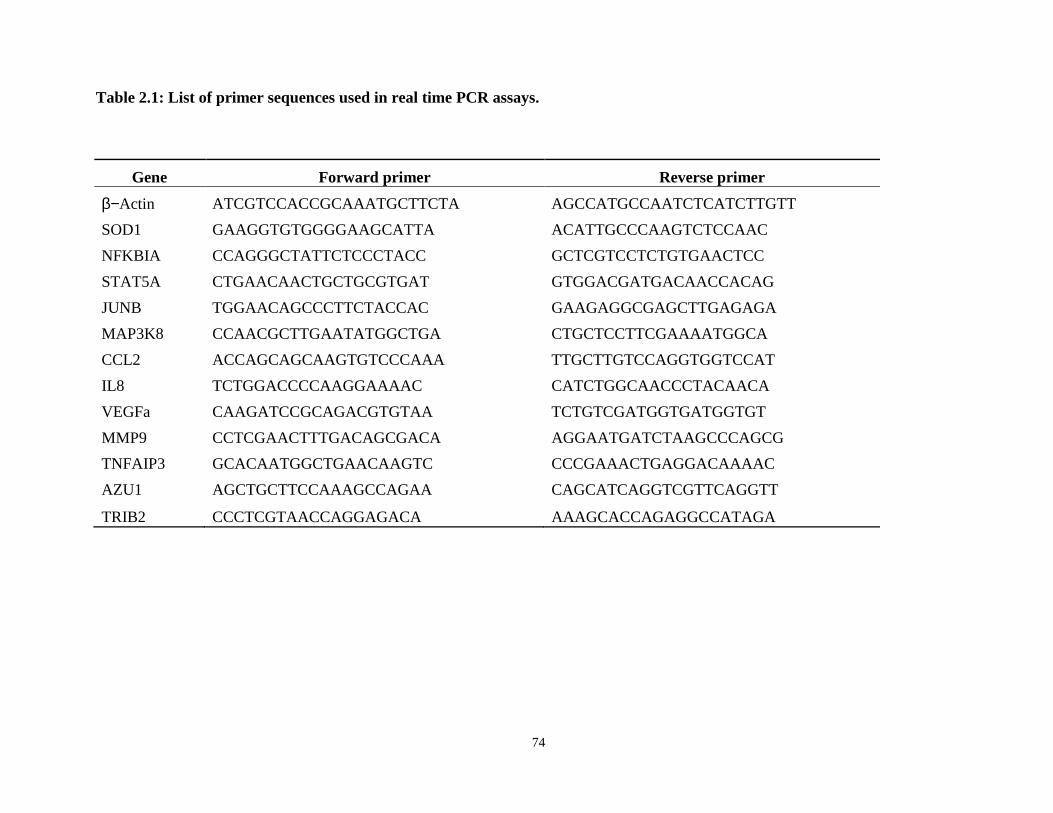
Table 2.1: Genes and primer sequences used in real time PCR assays.
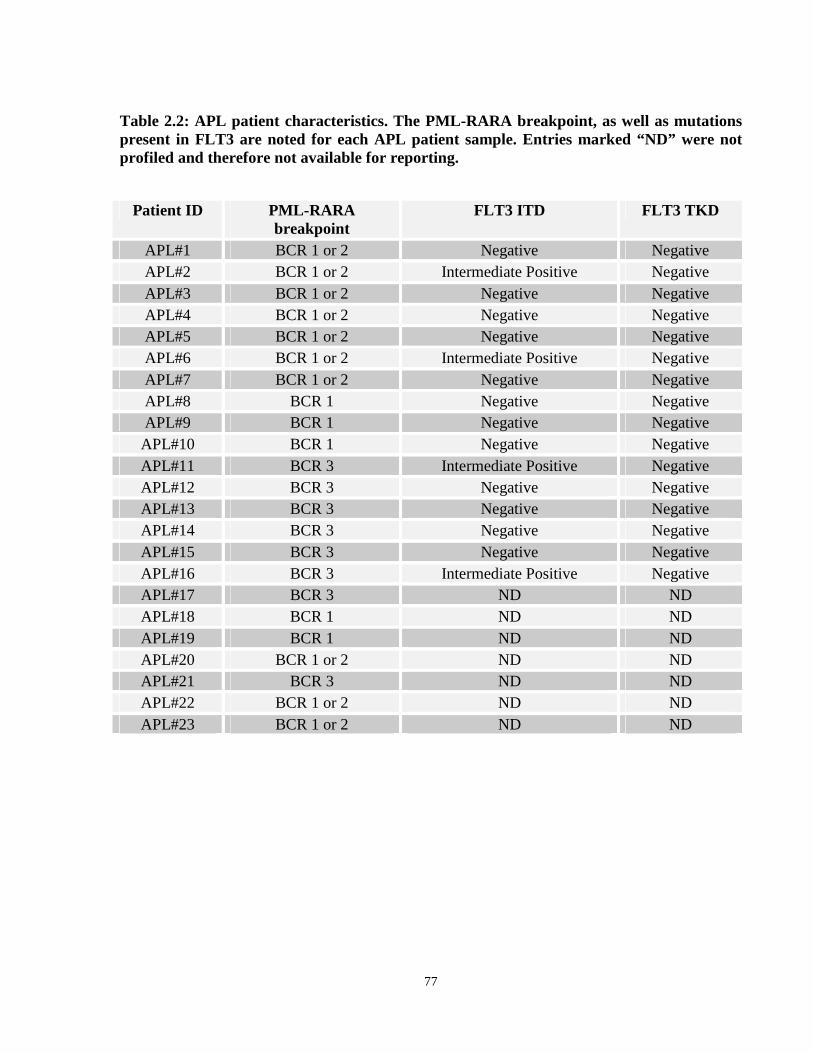
Table 2.2: APL patient characteristics
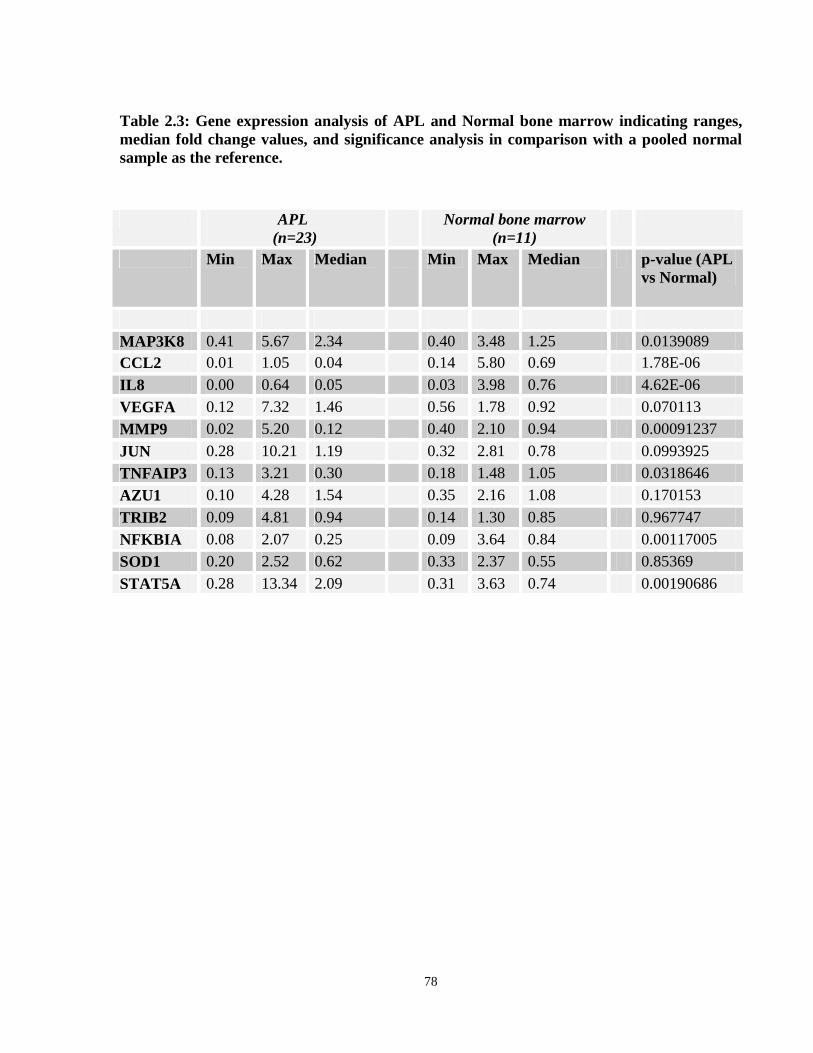
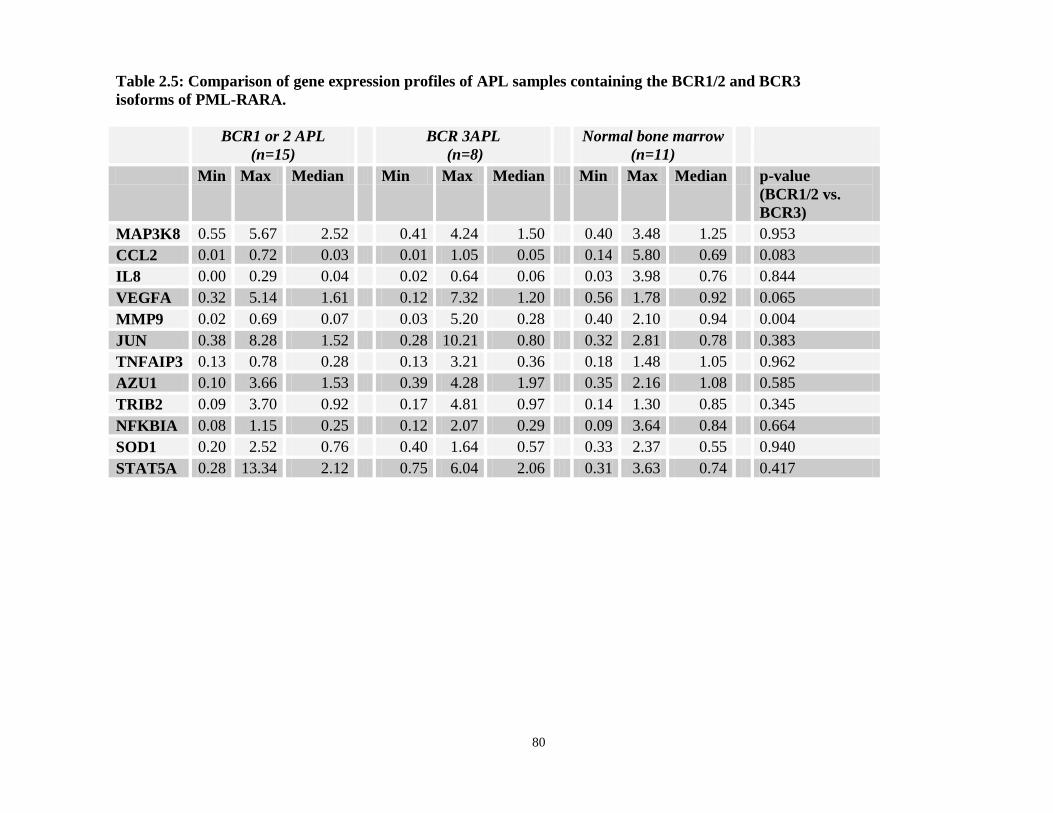
Table 2.3: Gene expression comparisons between APL patients containing the bcr1/2 and bcr3 isoforms of PML-RARA.
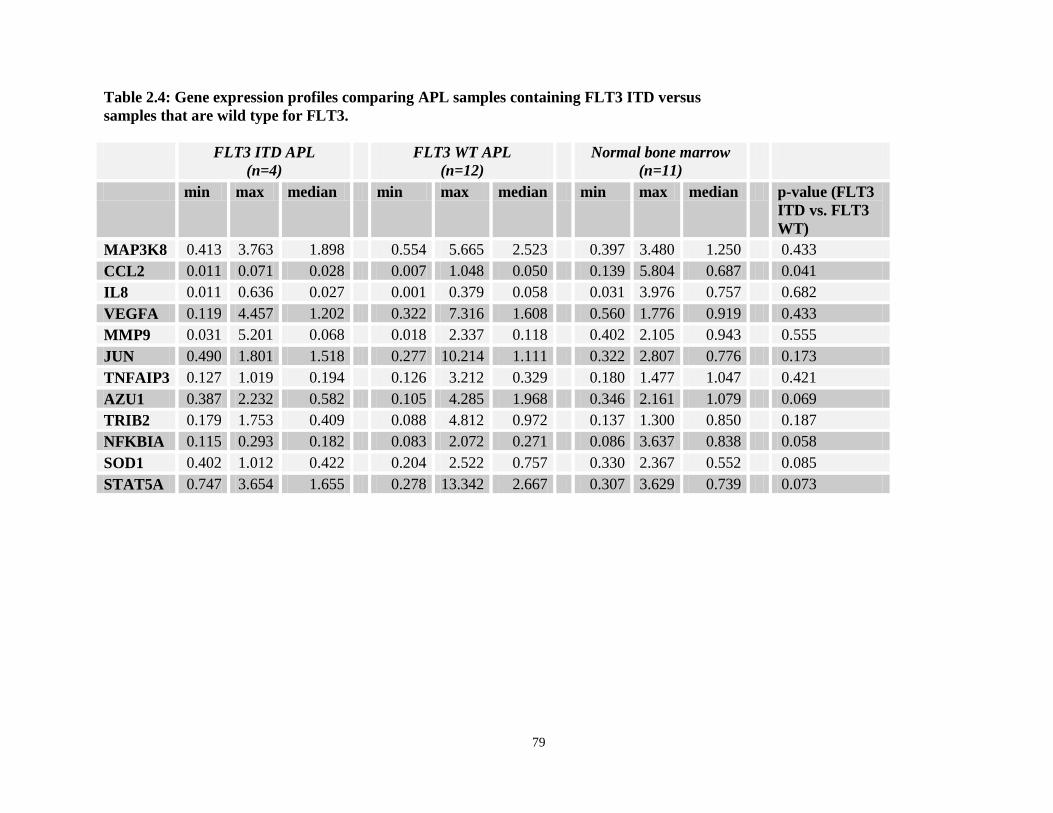
Table 2.4: Gene expression comparisons between APL patients containing FLT3-ITD versus FLT3 wild type patients.
Chapter 3: Gene expression profiling of NPM-RARA and NuMA-RARA variant APL
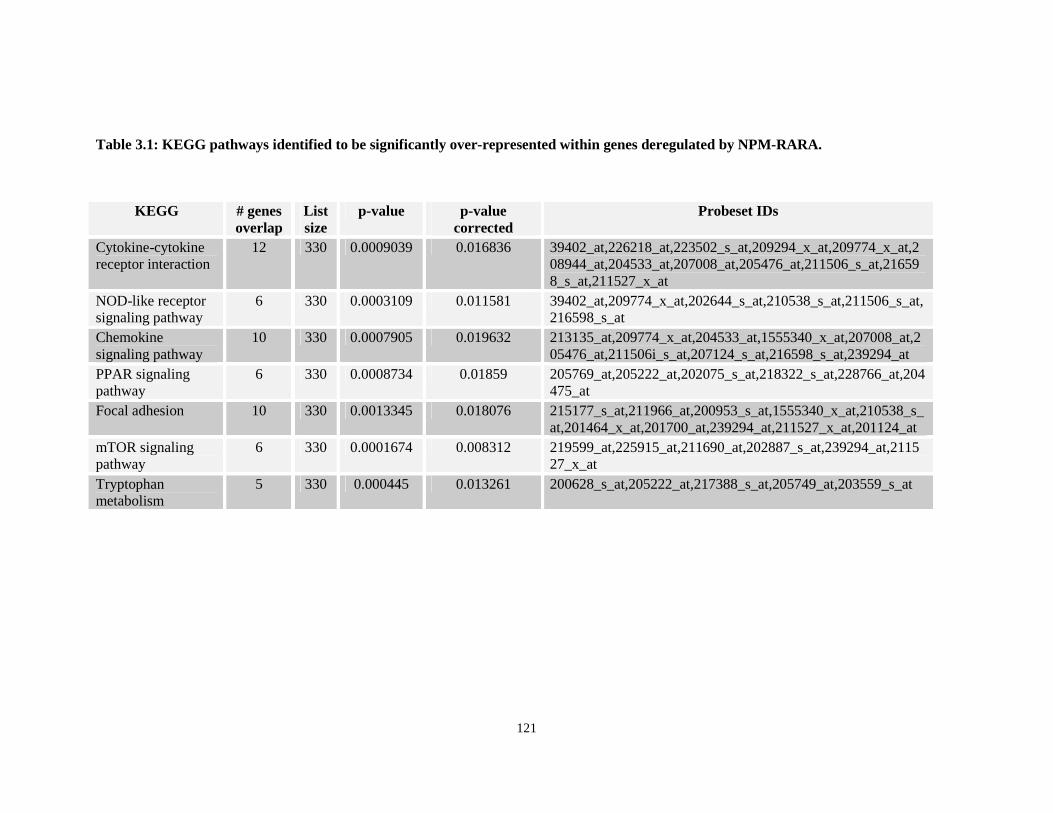
Table 3.1: KEGG pathways identified to be significantly over-represented within genes deregulated by NPM-RARA.
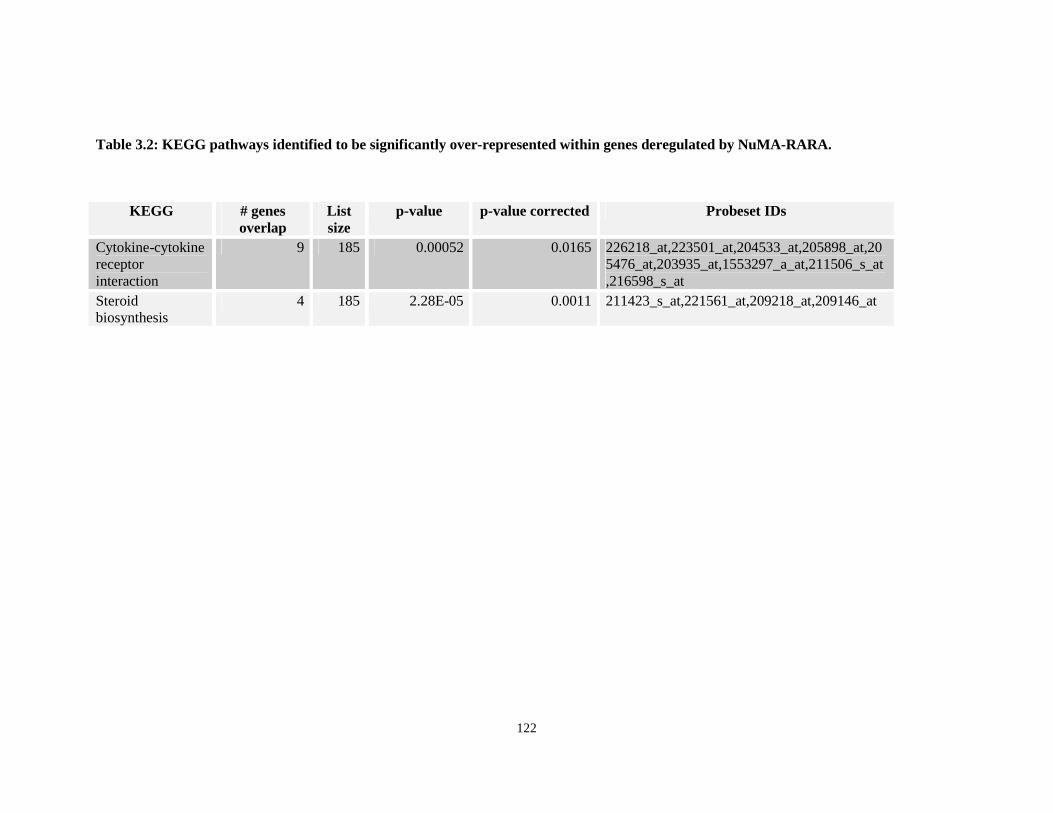
Table 3.2: KEGG pathways identified to be significantly over-represented within genes deregulated by NuMA-RARA.
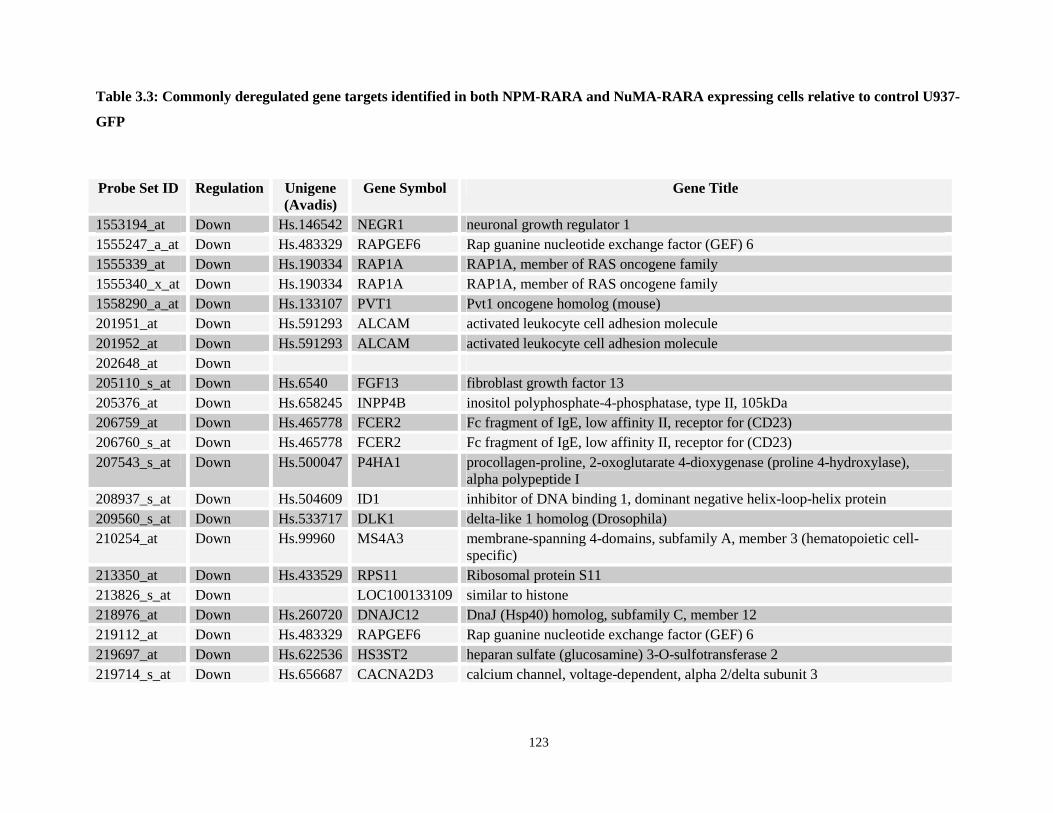
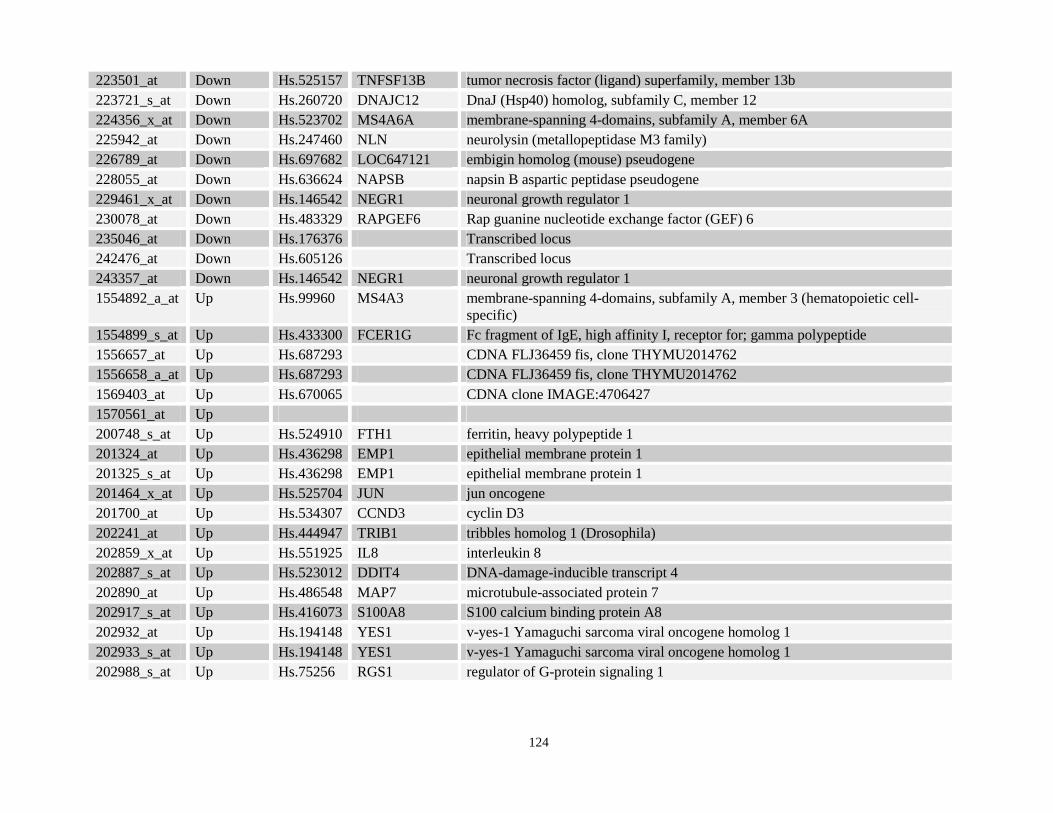
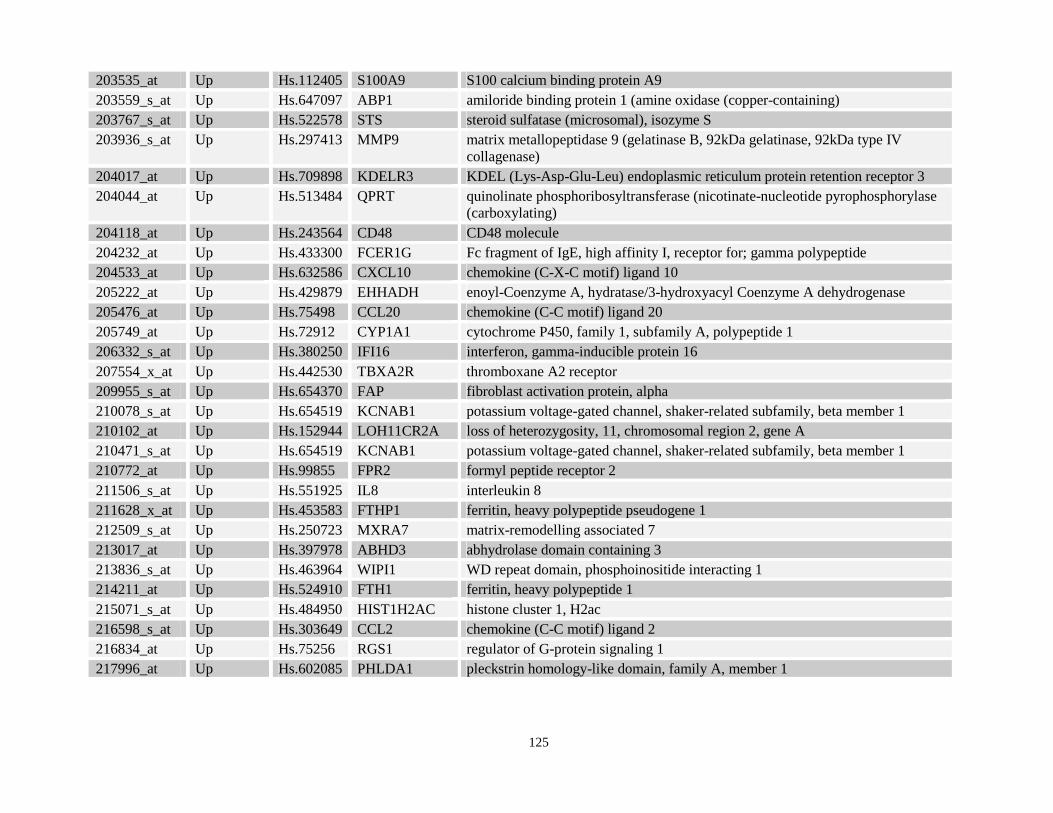
Table 3.3: Commonly deregulated gene targets identified in both NPM-RARA and NuMA-RARA expressing cells relative to control U937-GFP
Table 3.4: KEGG pathways identified to be significantly over-represented within genes commonly deregulated by NPM-RARA and NuMA-RARA.
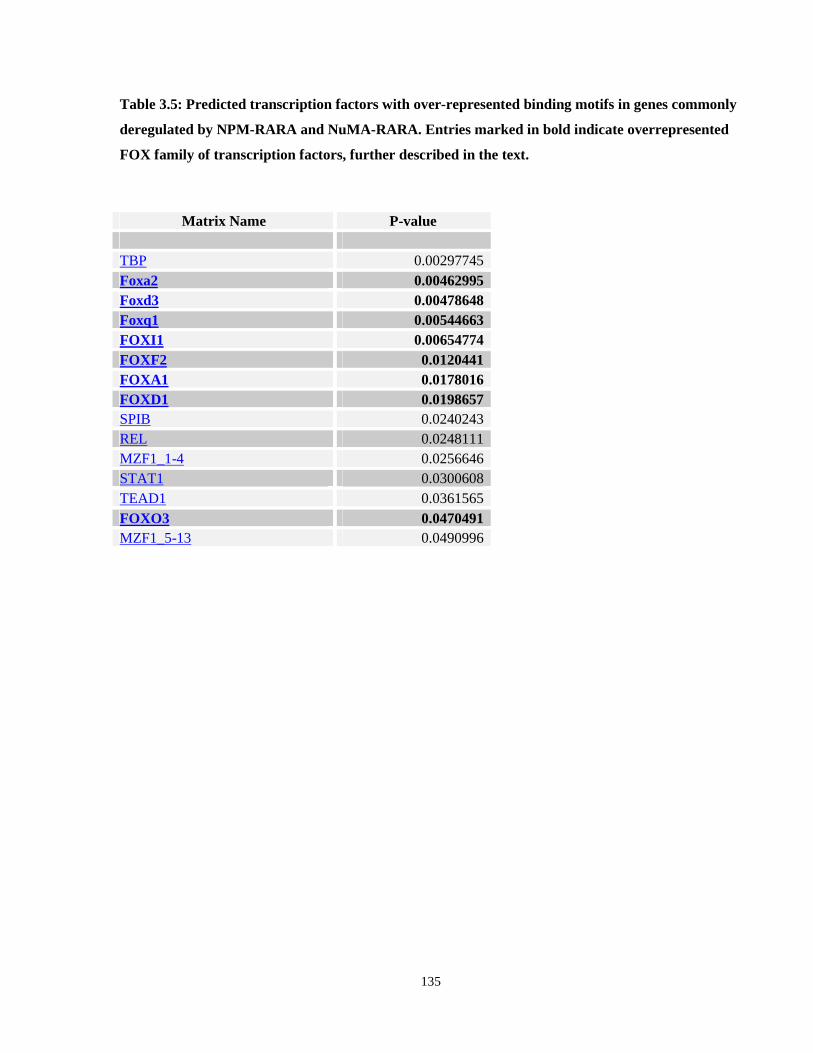
Table 3.5: Predicted transcription factors with over-represented binding motifs in genes commonly deregulated by NPM-RARA and NuMA-RARA.
Table 3.6: List of NPM-RARA deregulated genes that are also direct targets of PML-RARA identified from the Wang et al., and Martens et al. published datasets. Entries marked with an
xvii
asterisk are PML-RARA binding targets, which were reported in both of these independent published datasets.
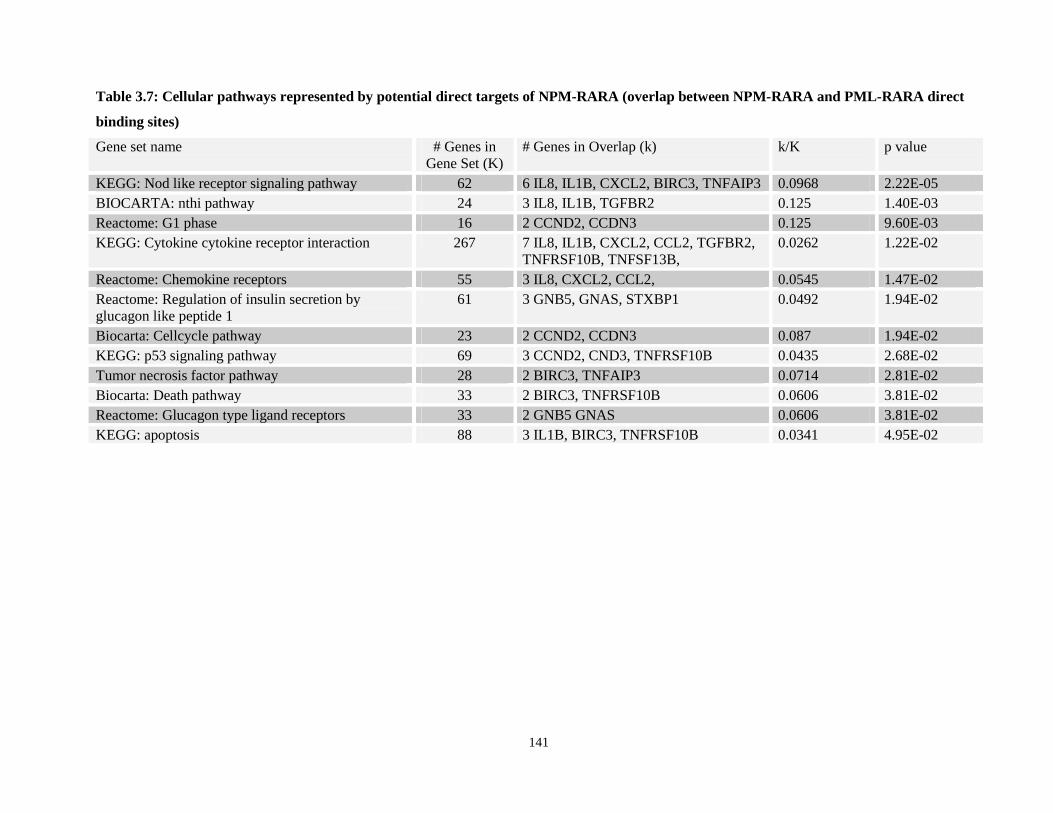
Table 3.7: Cellular pathways represented by potential direct targets of NPM-RARA (overlap between NPM-RARA and PML-RARA direct binding sites)
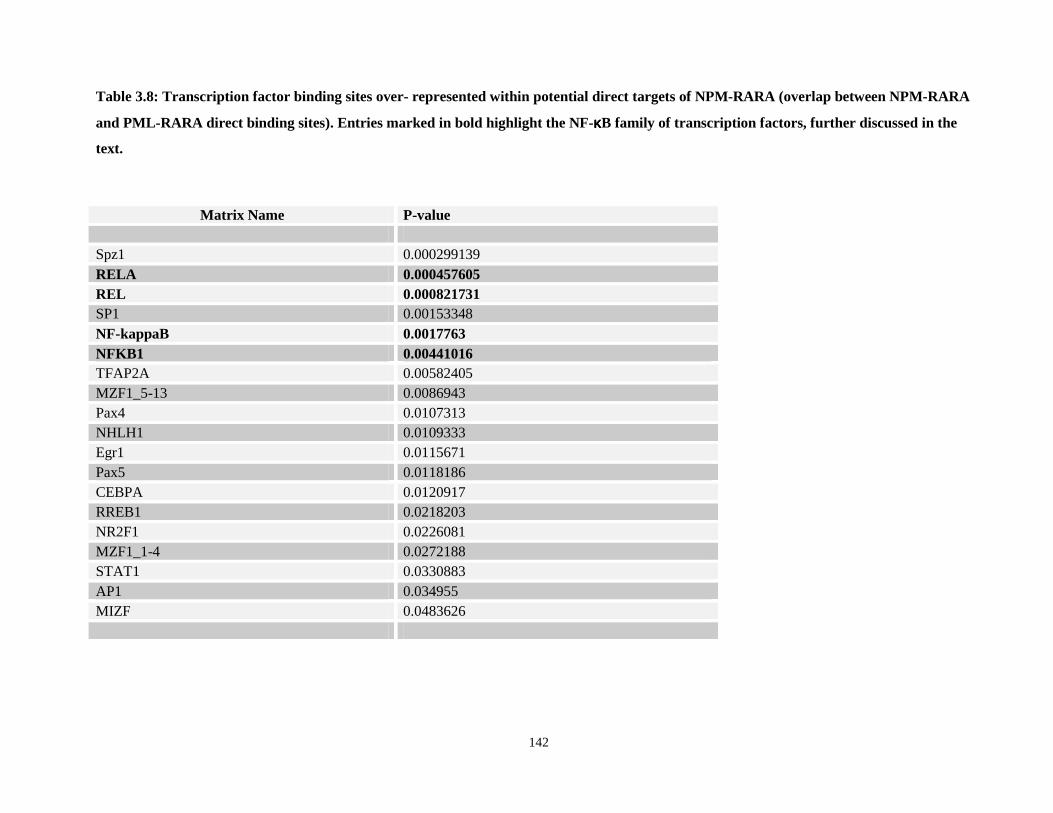
Table 3.8: Transcription factor binding sites over- represented within potential direct targets of NPM-RARA (overlap between NPM-RARA and PML-RARA direct binding sites)
Table 3.9: List of NPM-RARA deregulated genes that are also direct targets of PLZF-RARA identified from the Rice et al., and Spicuglia et al. published datasets.
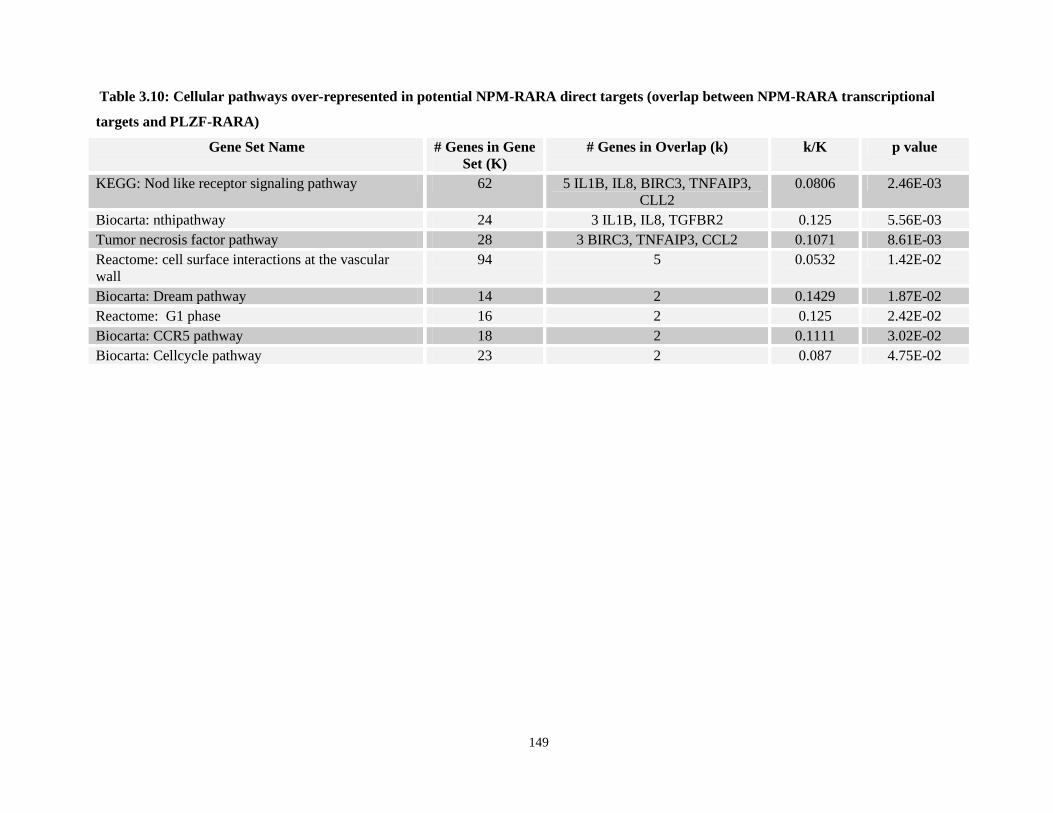
Table 3.10: Cellular pathways over-represented in potential NPM-RARA direct targets (overlap between NPM-RARA transcriptional targets and PLZF-RARA)
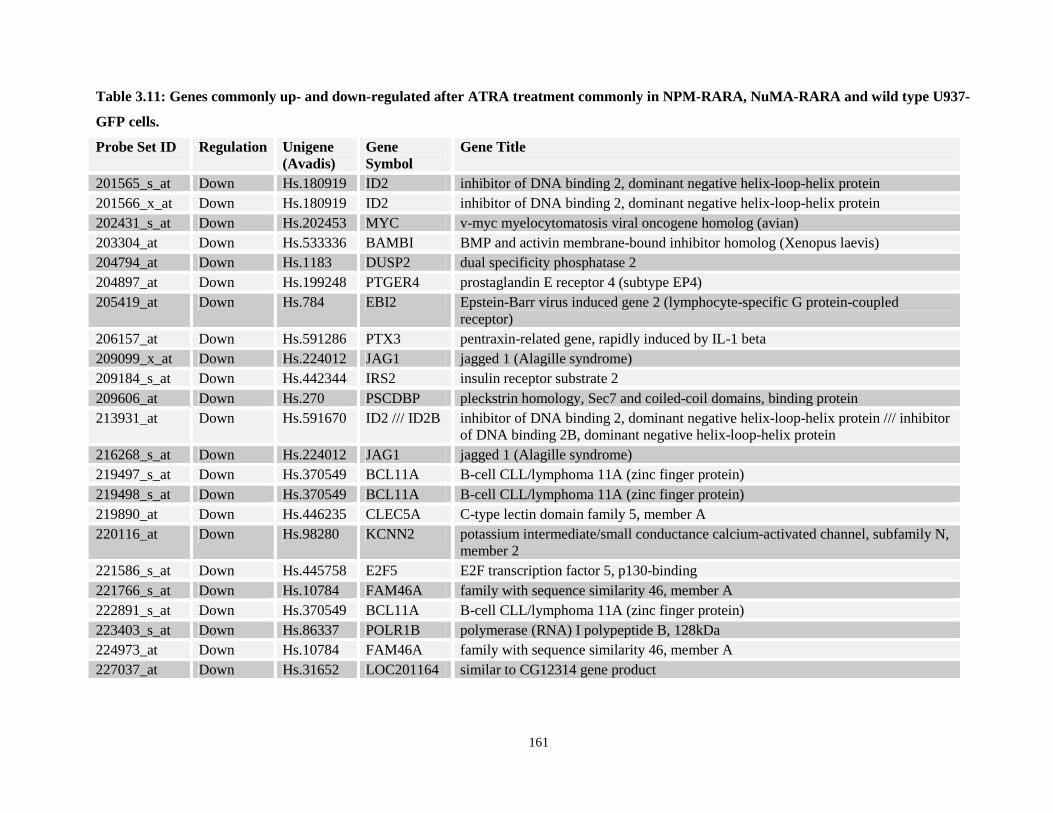
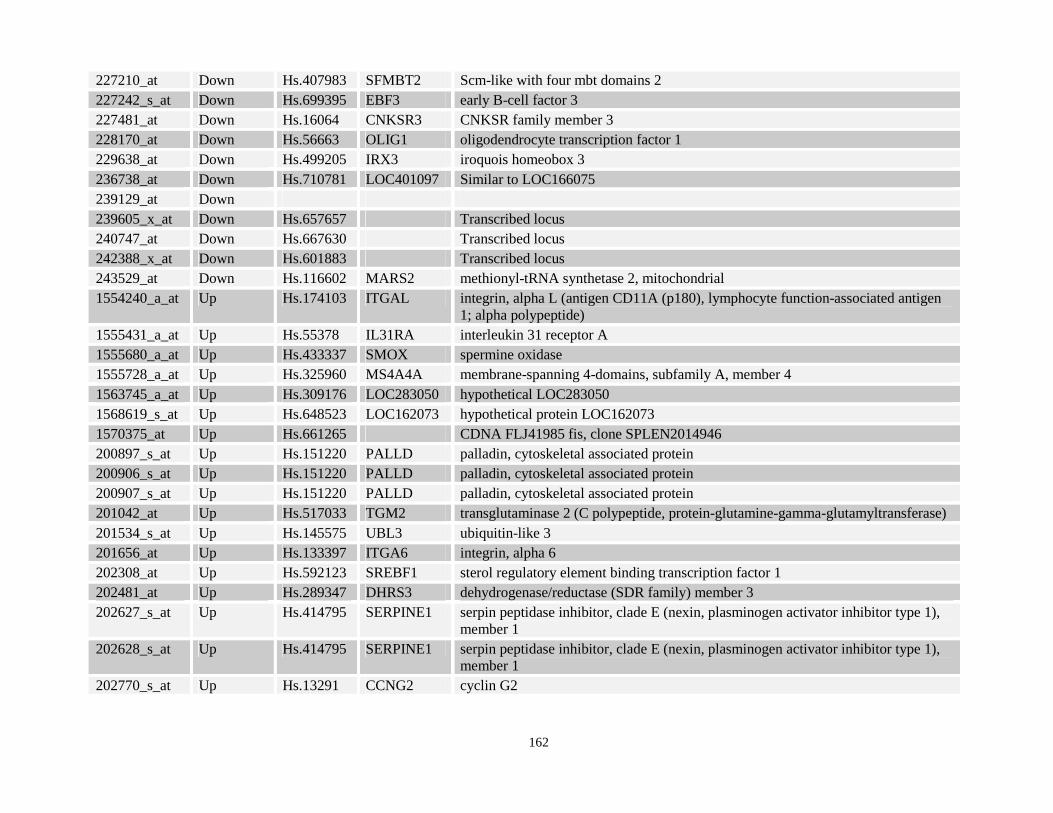
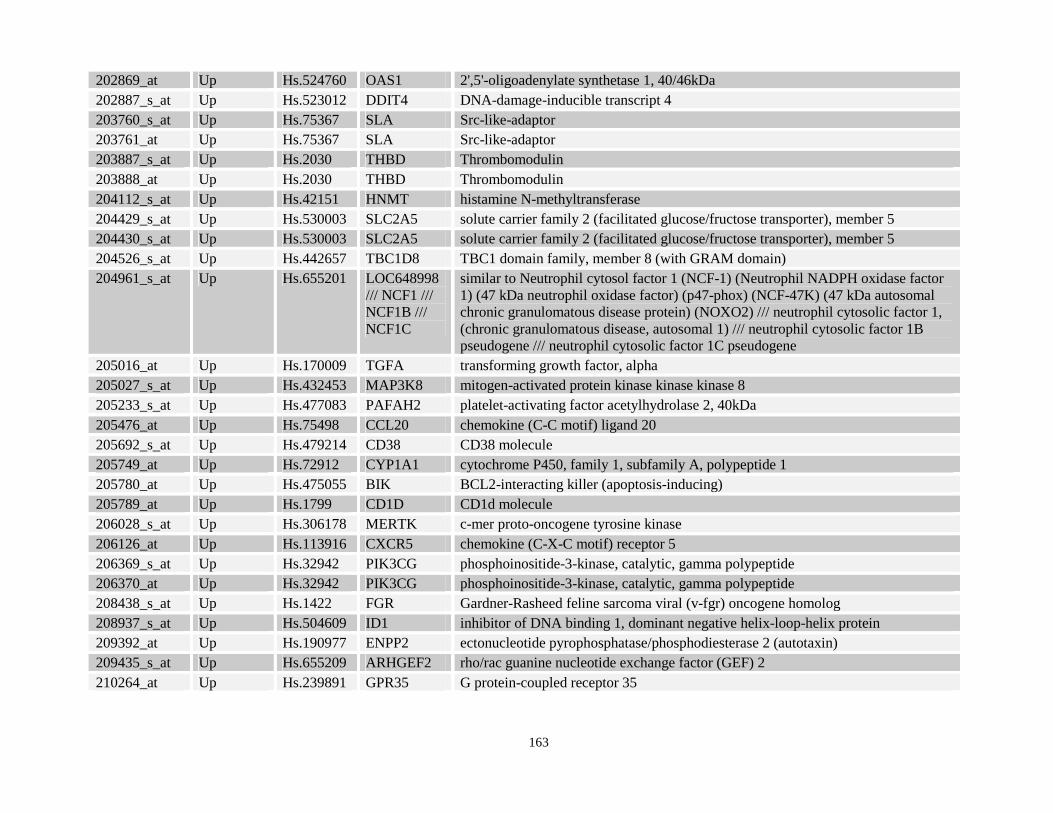
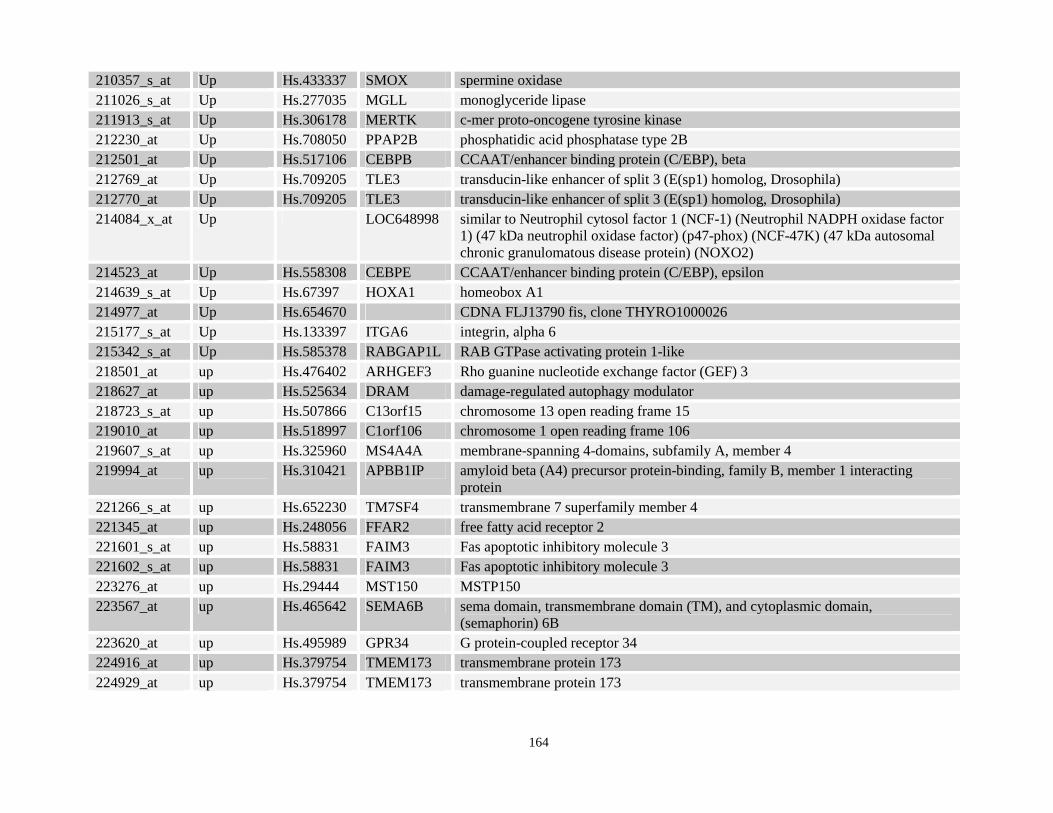
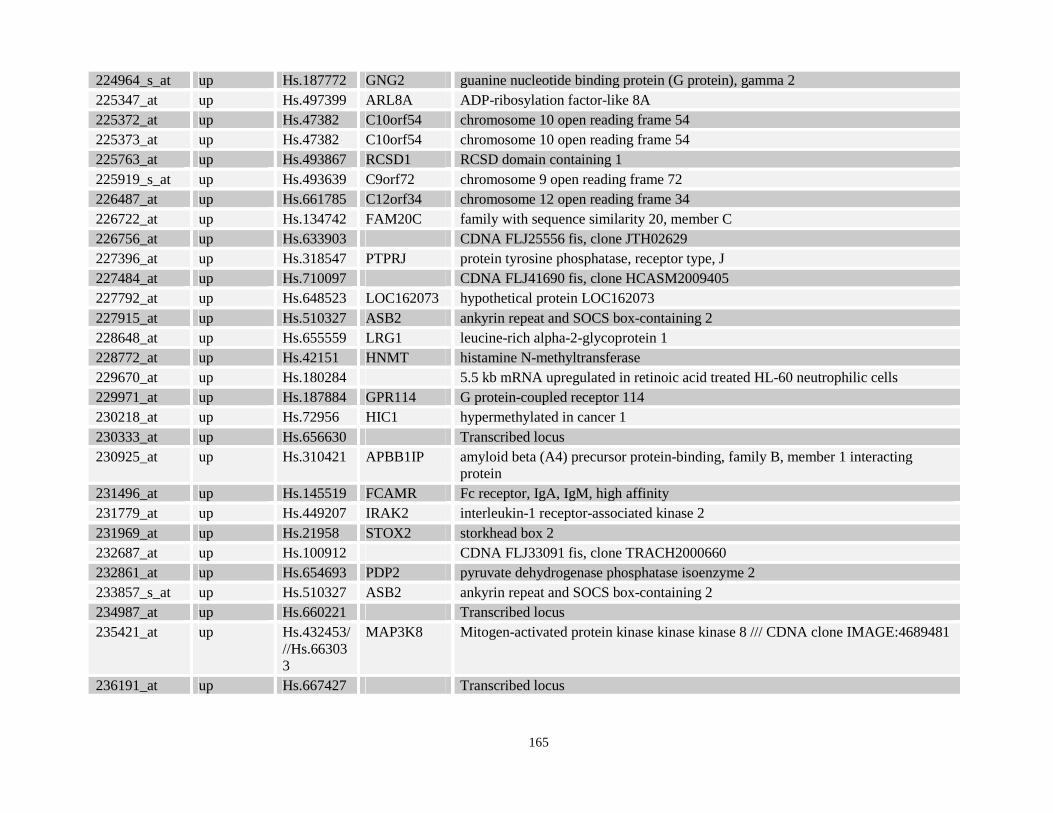
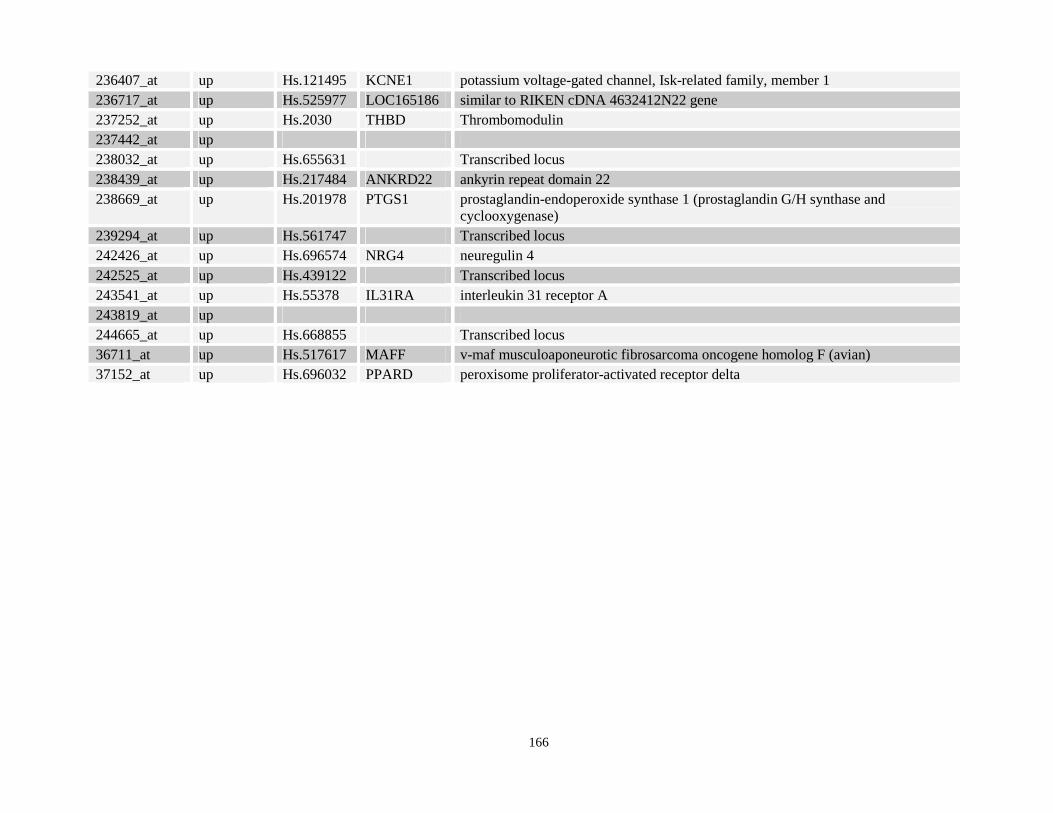
Table 3.11: Genes commonly up- and down-regulated after ATRA treatment commonly in NPM-RARA, NuMA-RARA and wild type U937-GFP cells.
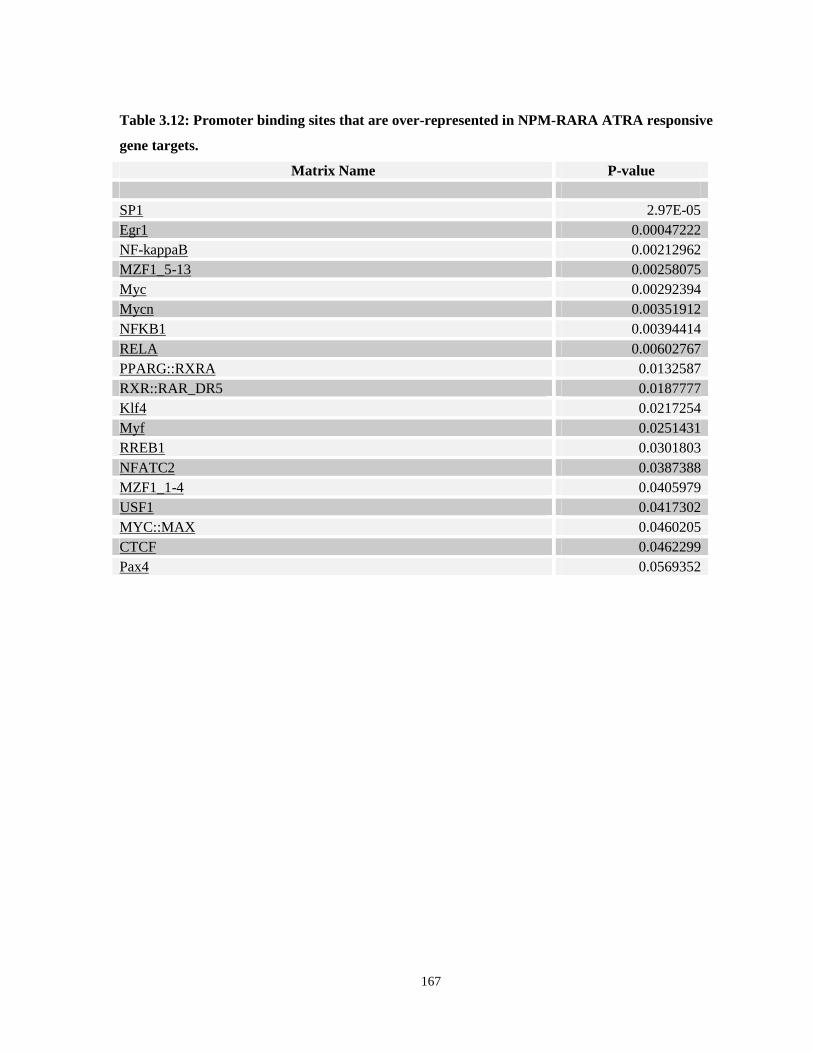
Table 3.12: Promoter binding sites that are over-represented in NPM-RARA ATRA responsive gene targets.
Table 3.13: Promoter binding sites that are over-represented in ATRA responsive gene sets in NuMA-RARA
Table 3.14: Promoter binding sites that are over-represented in ATRA responsive gene targets of wild type U937-GFP control cells.
Table 3.15: Compounds identified from gene expression matching analysis using ATRA responsive gene targets in NPM-RARA.
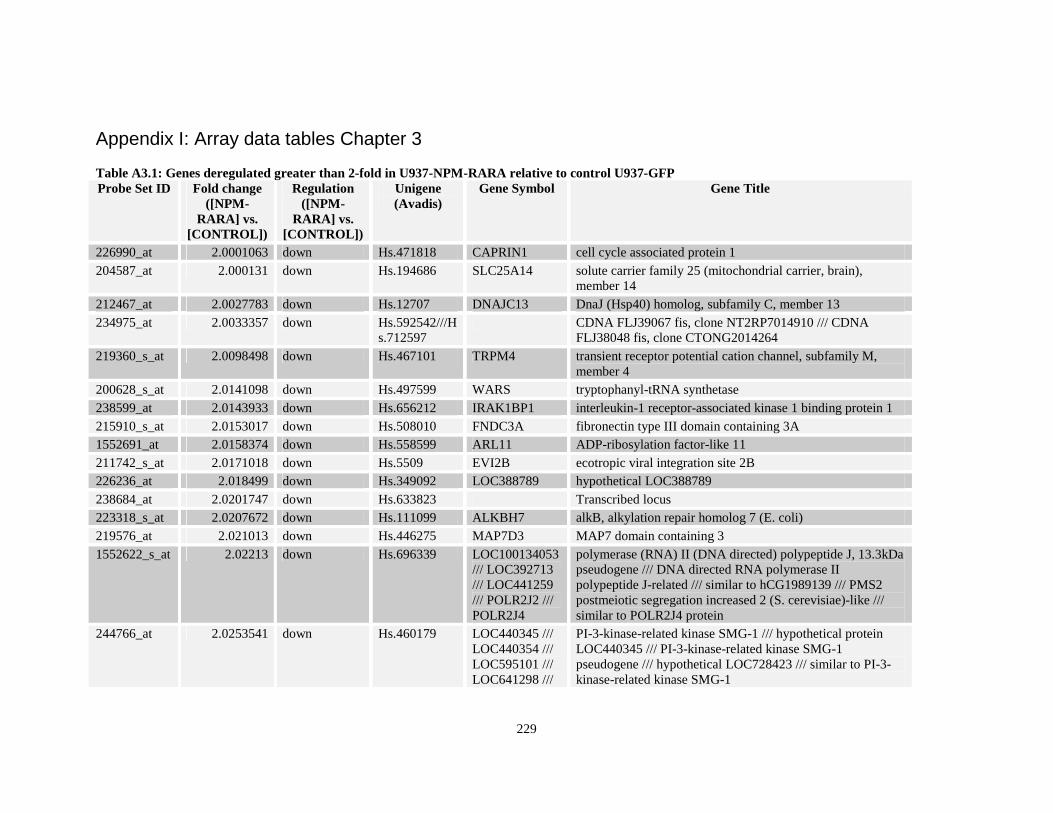
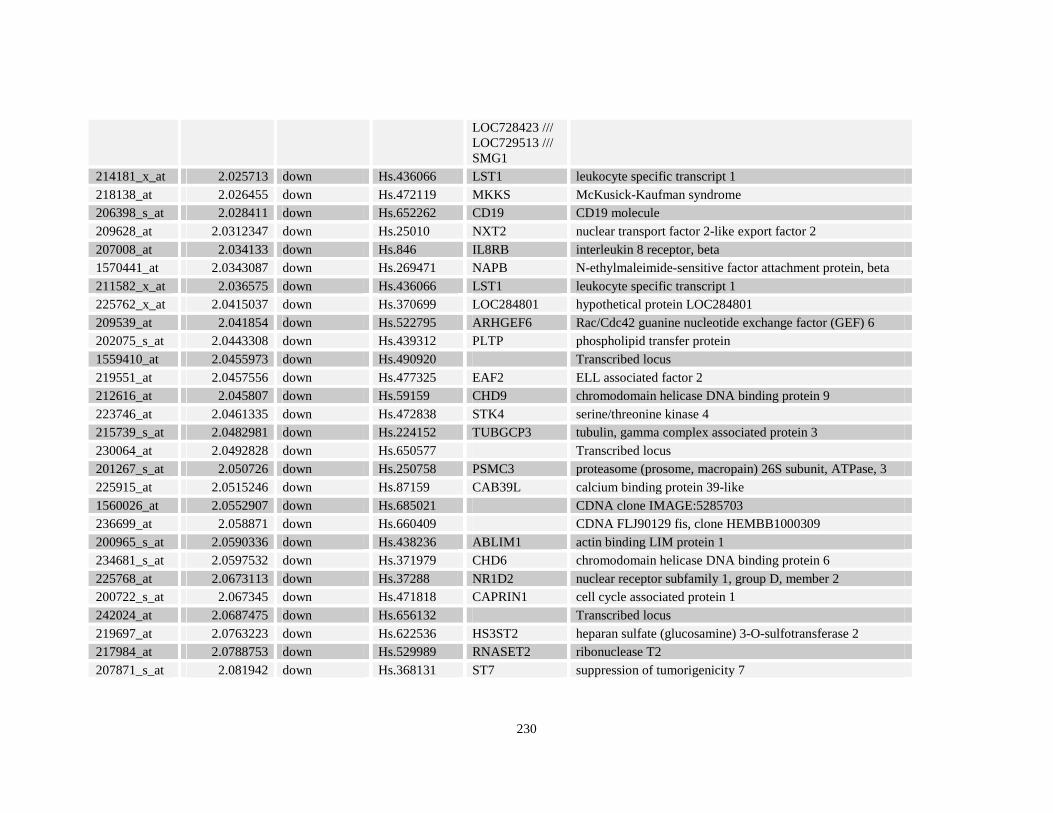
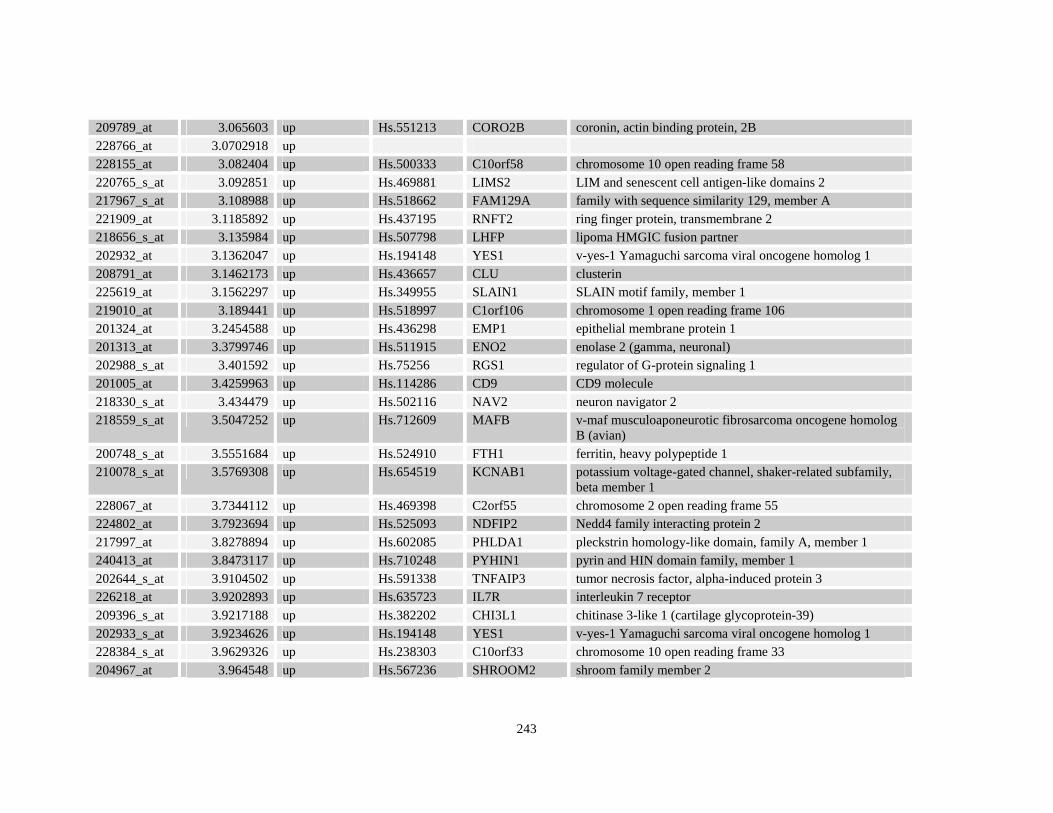
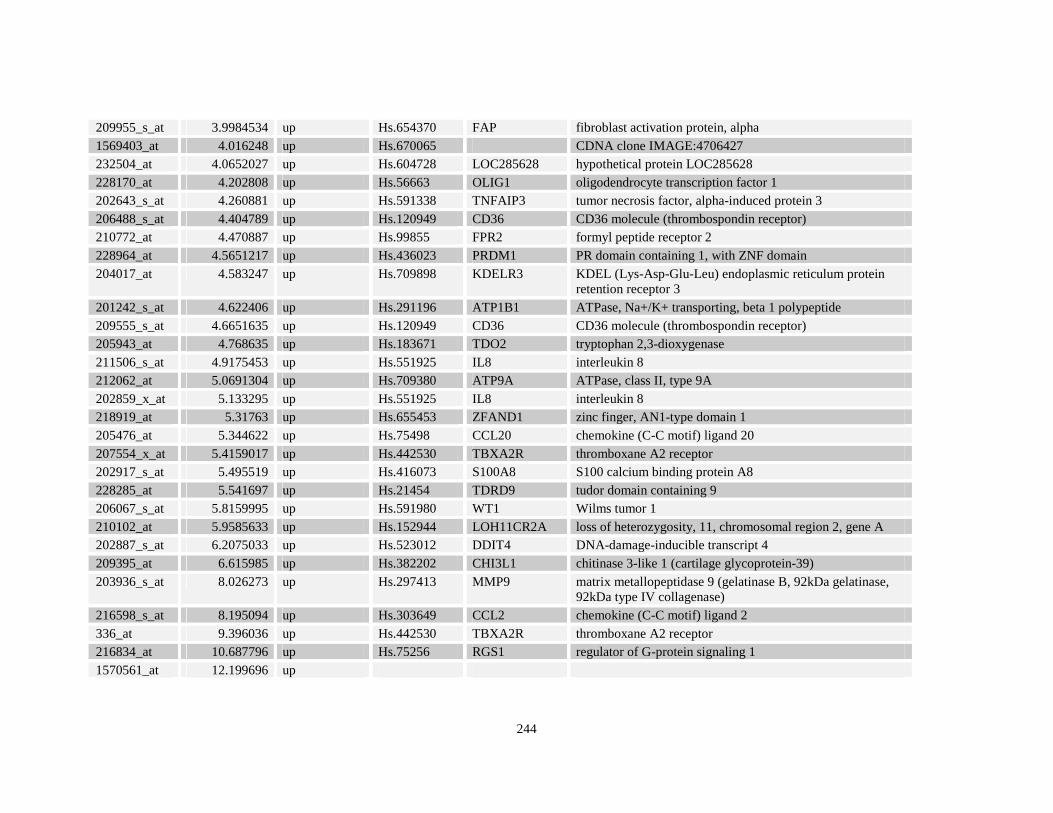
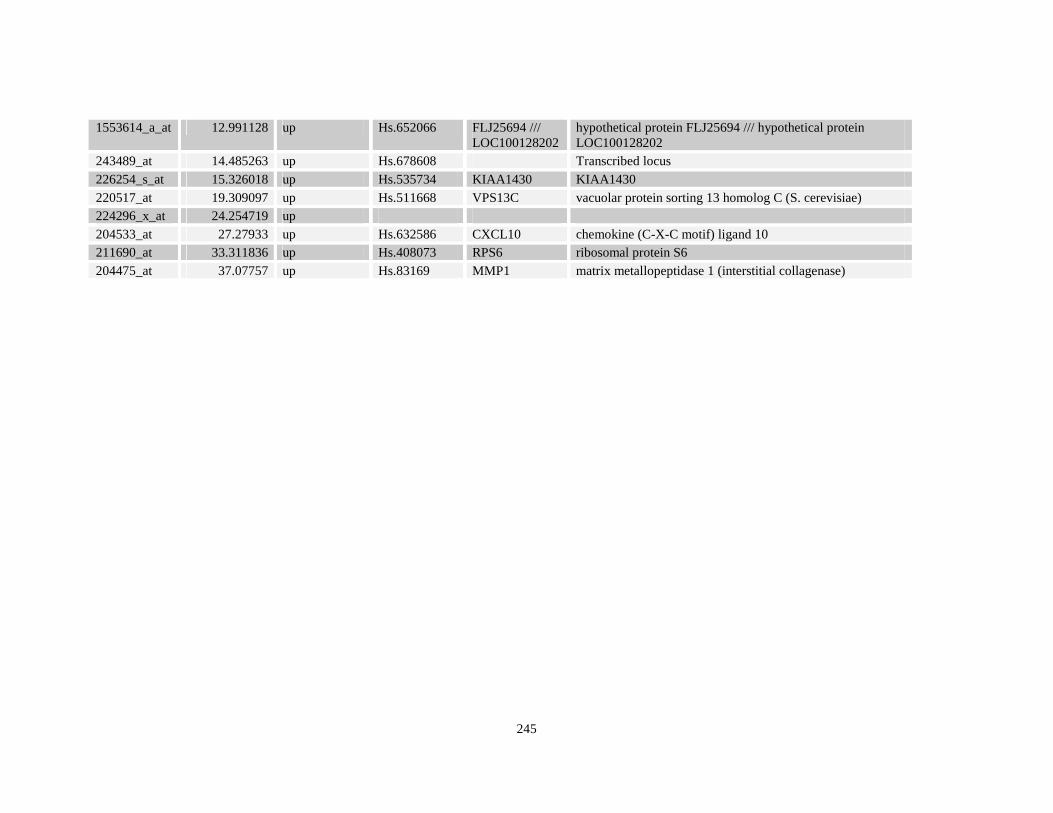
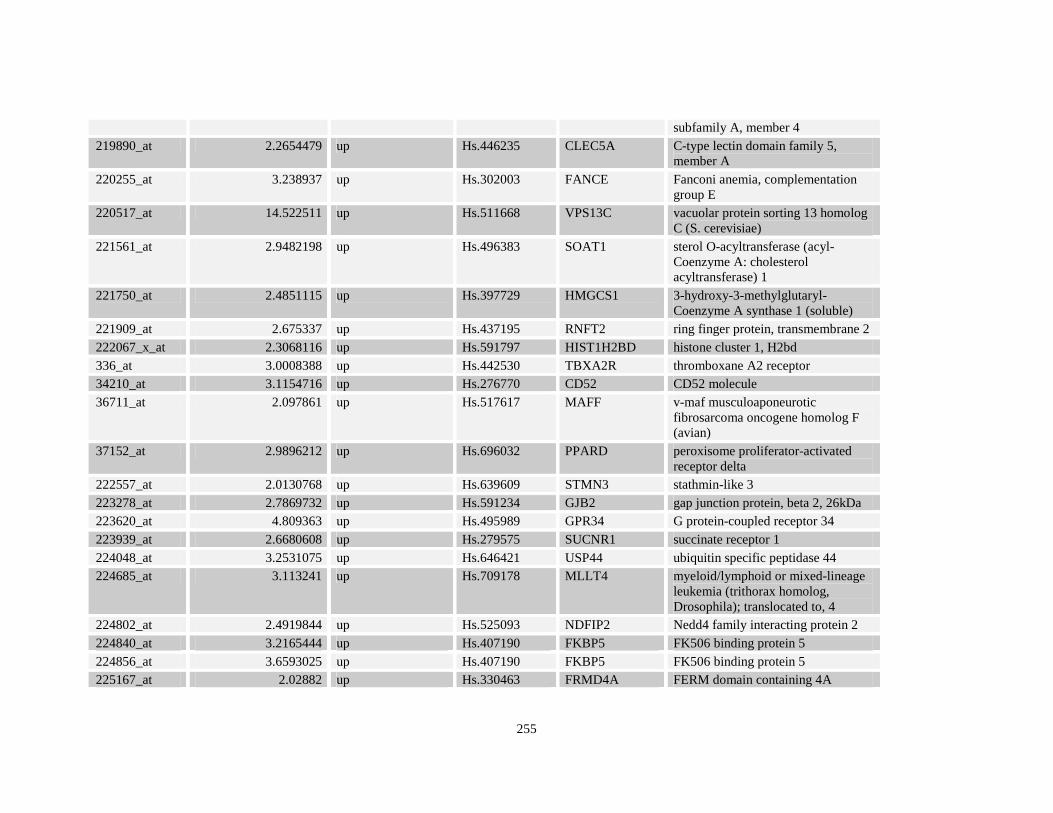
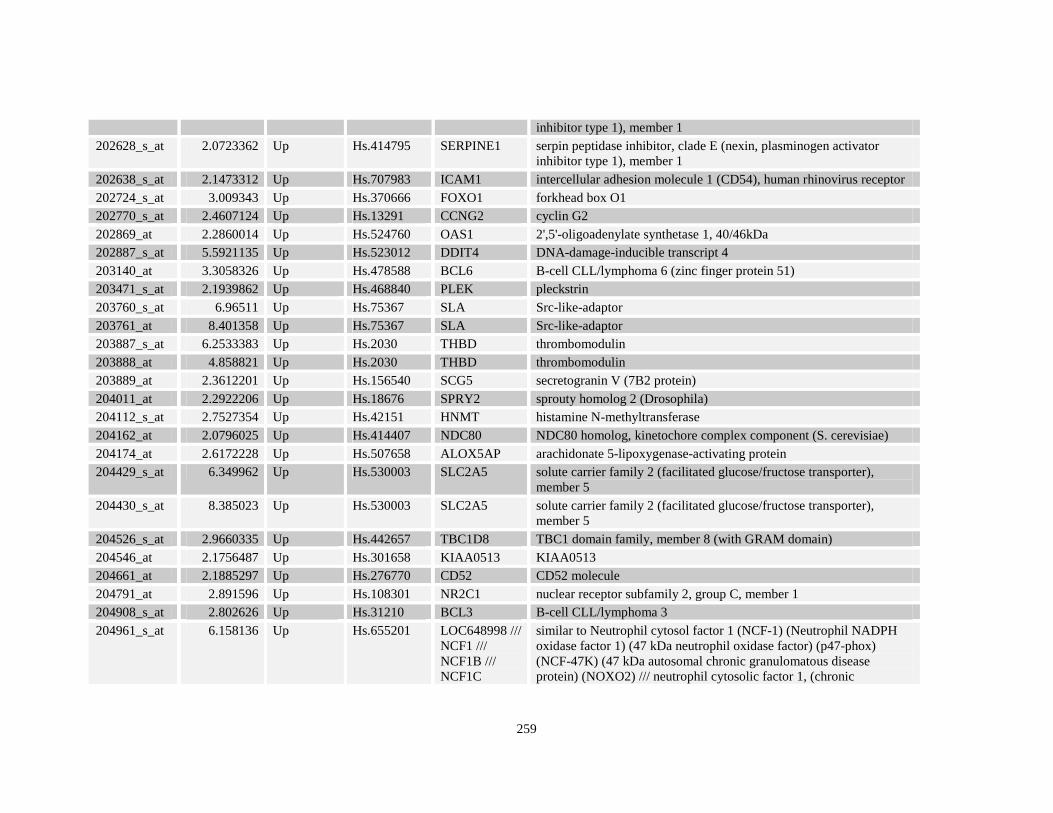
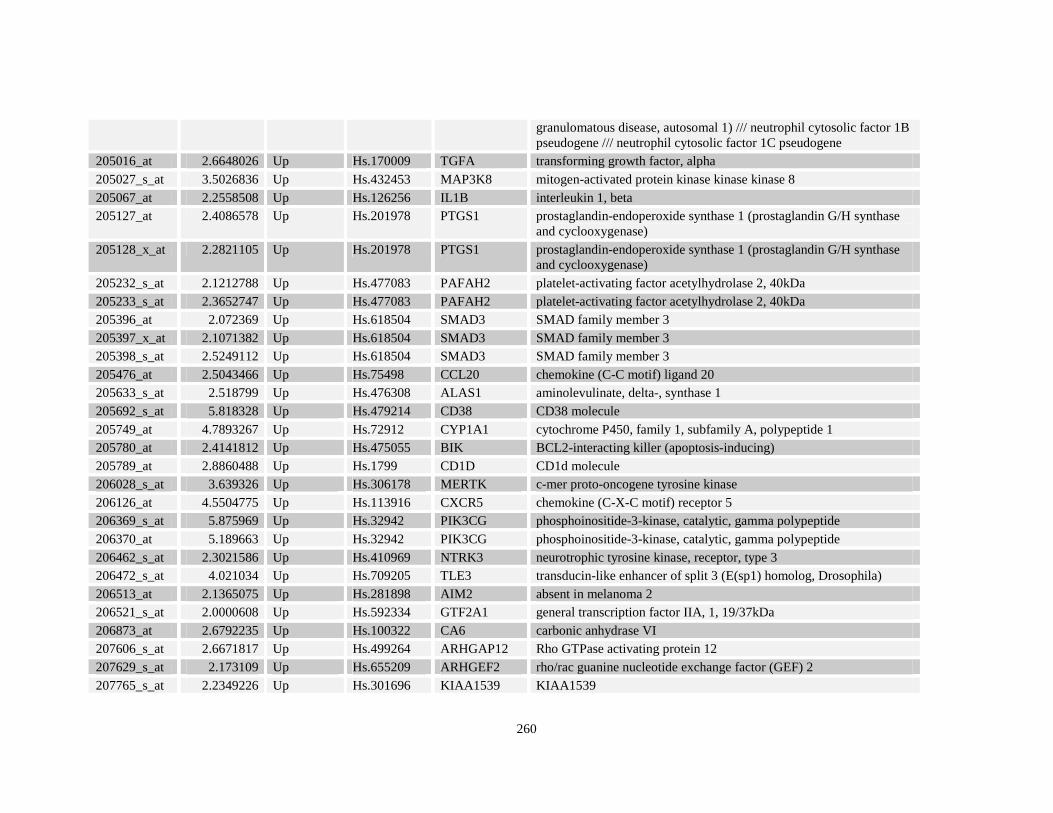
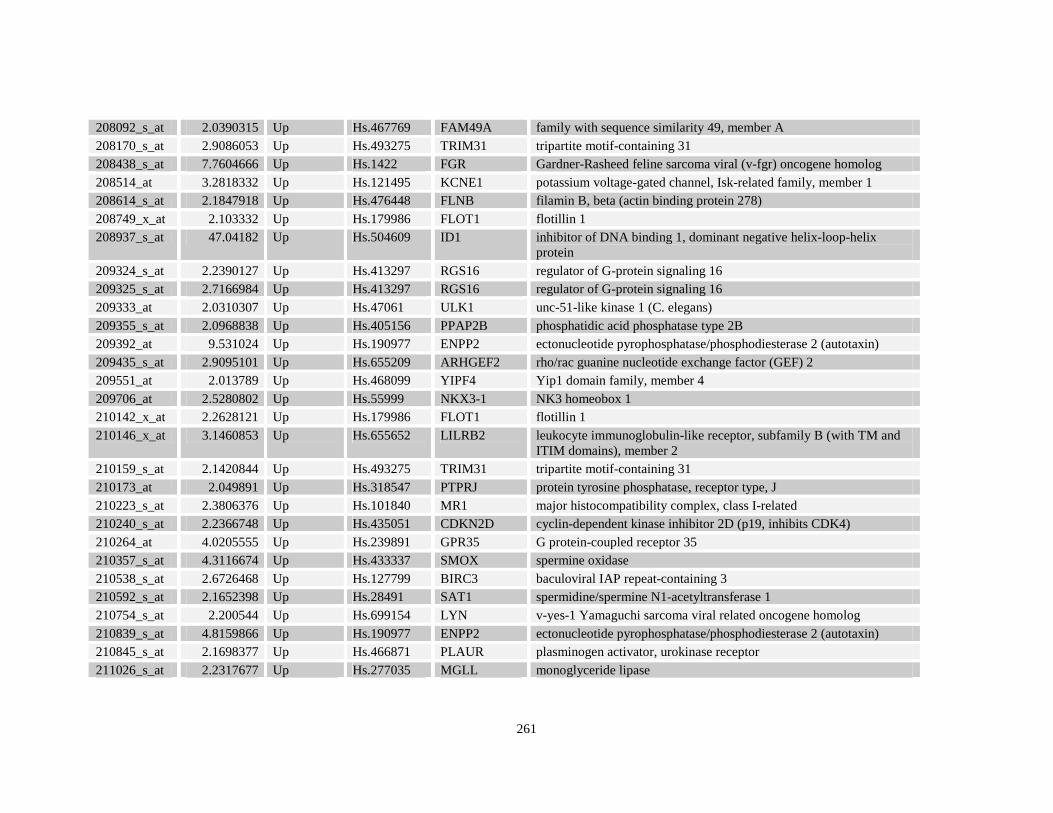
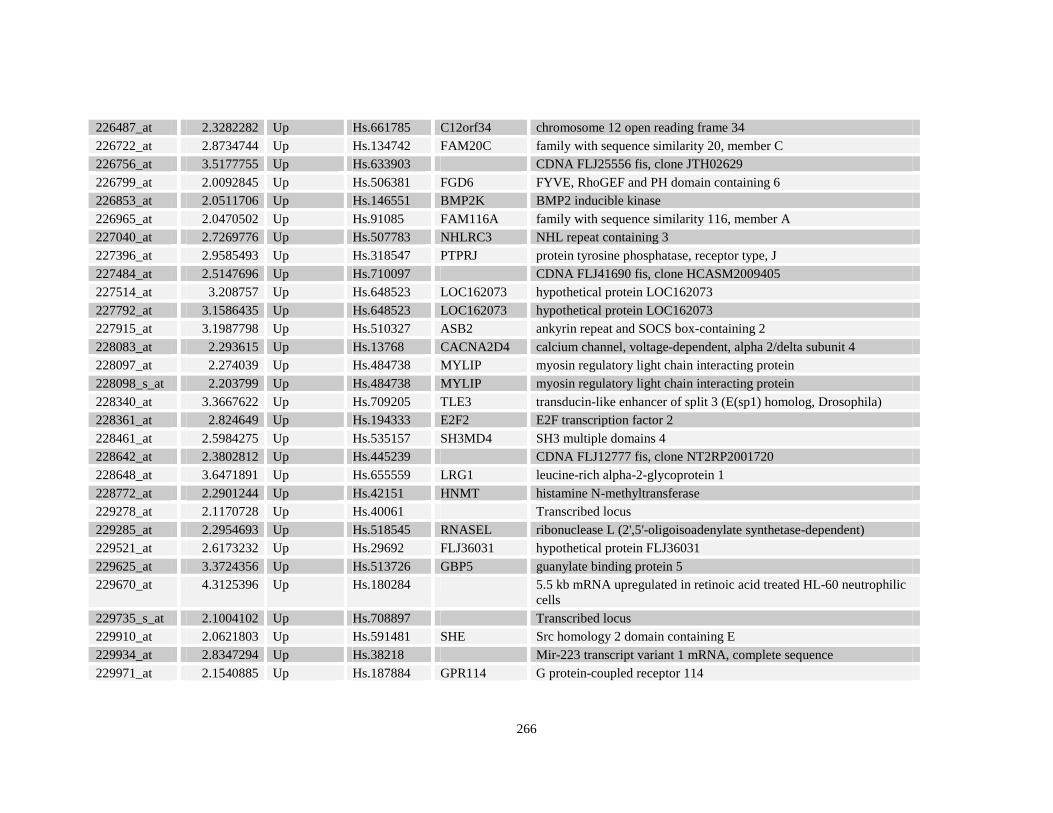
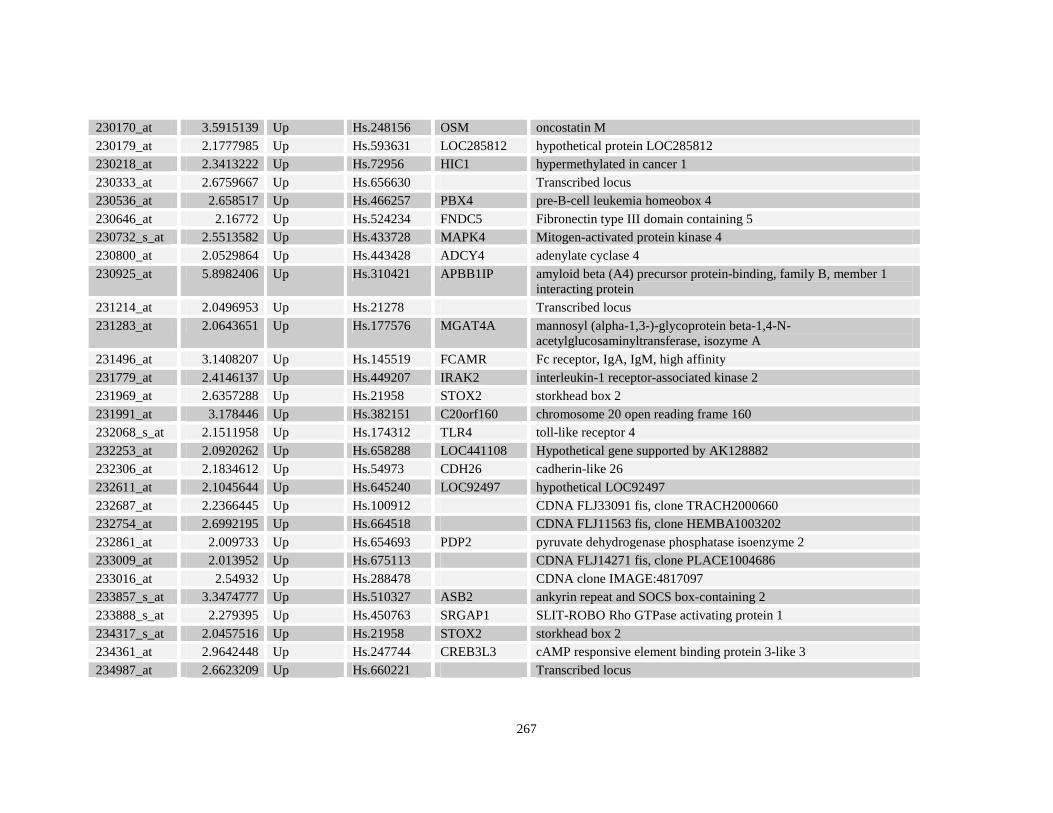
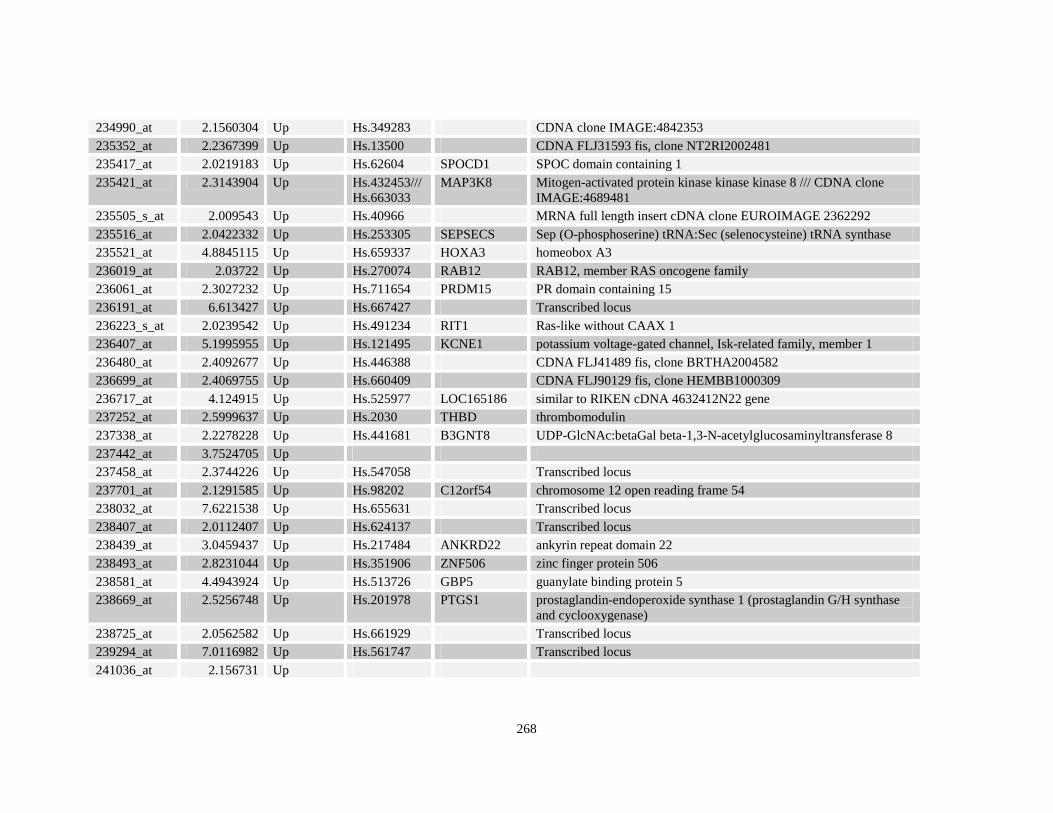
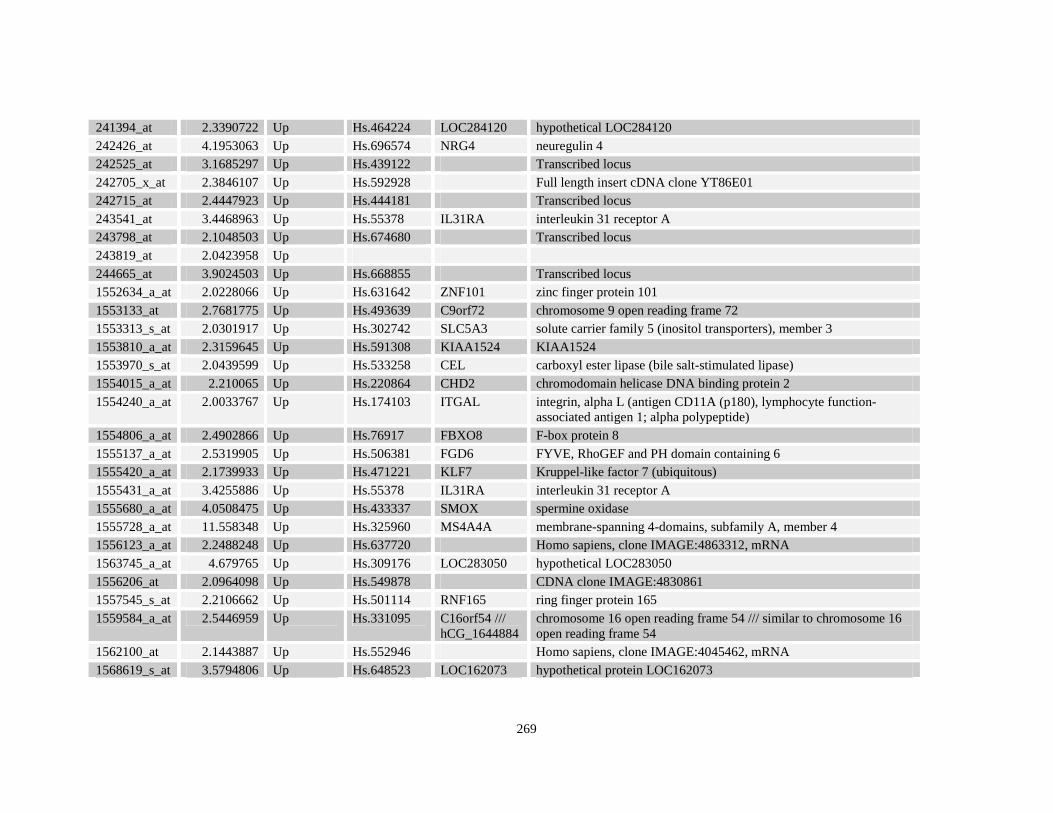
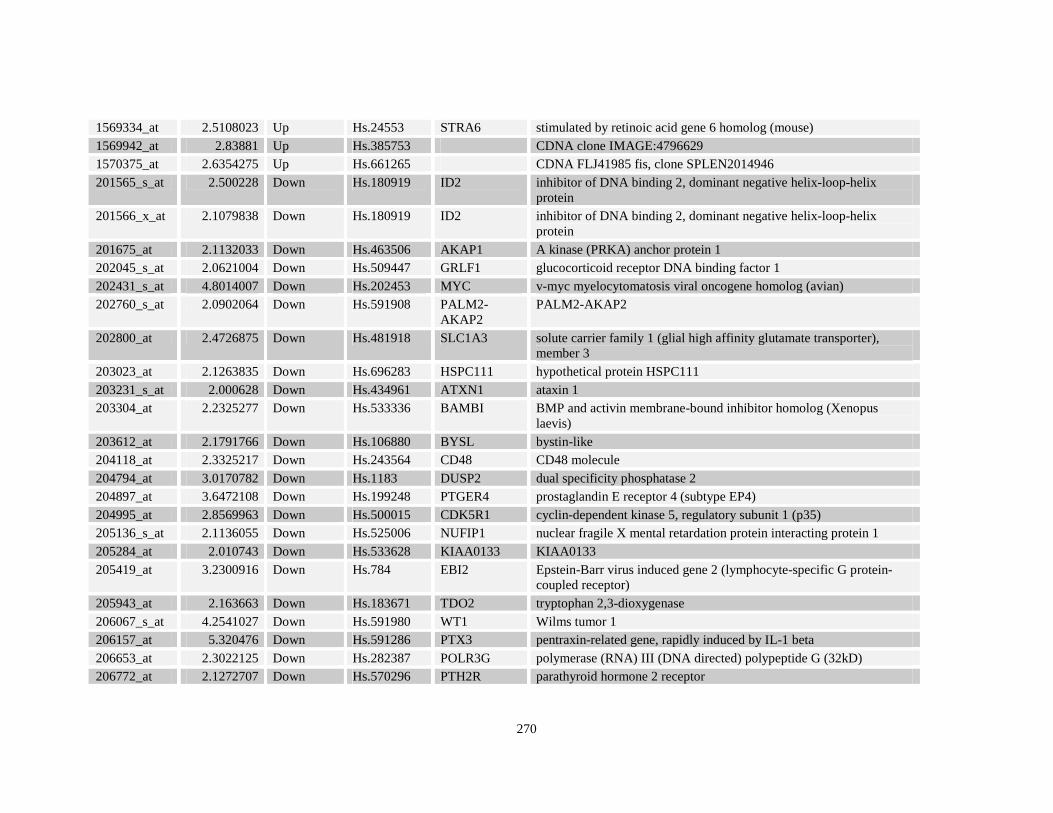
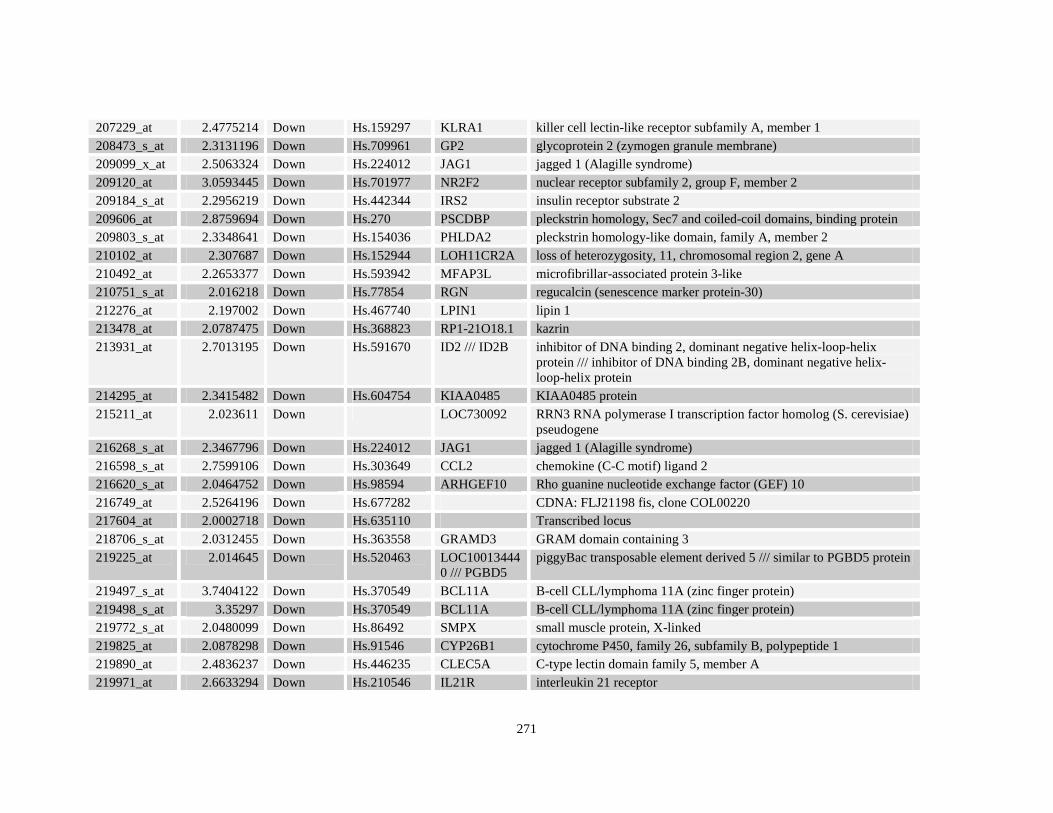
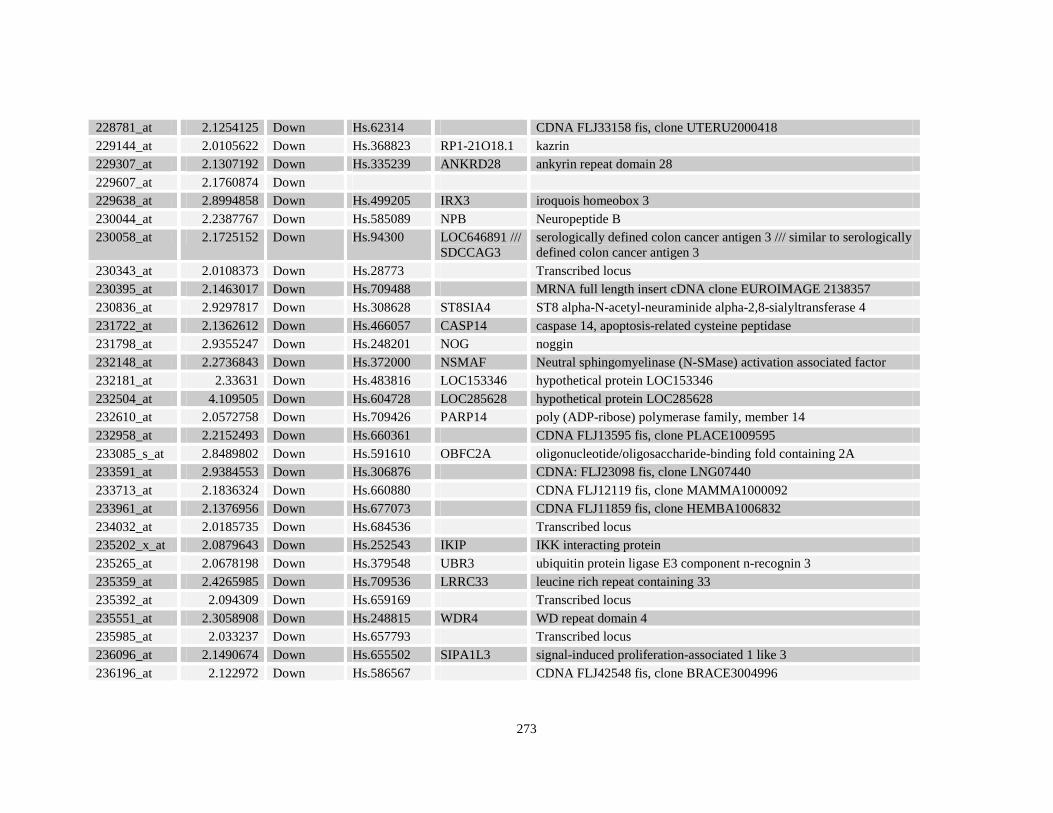
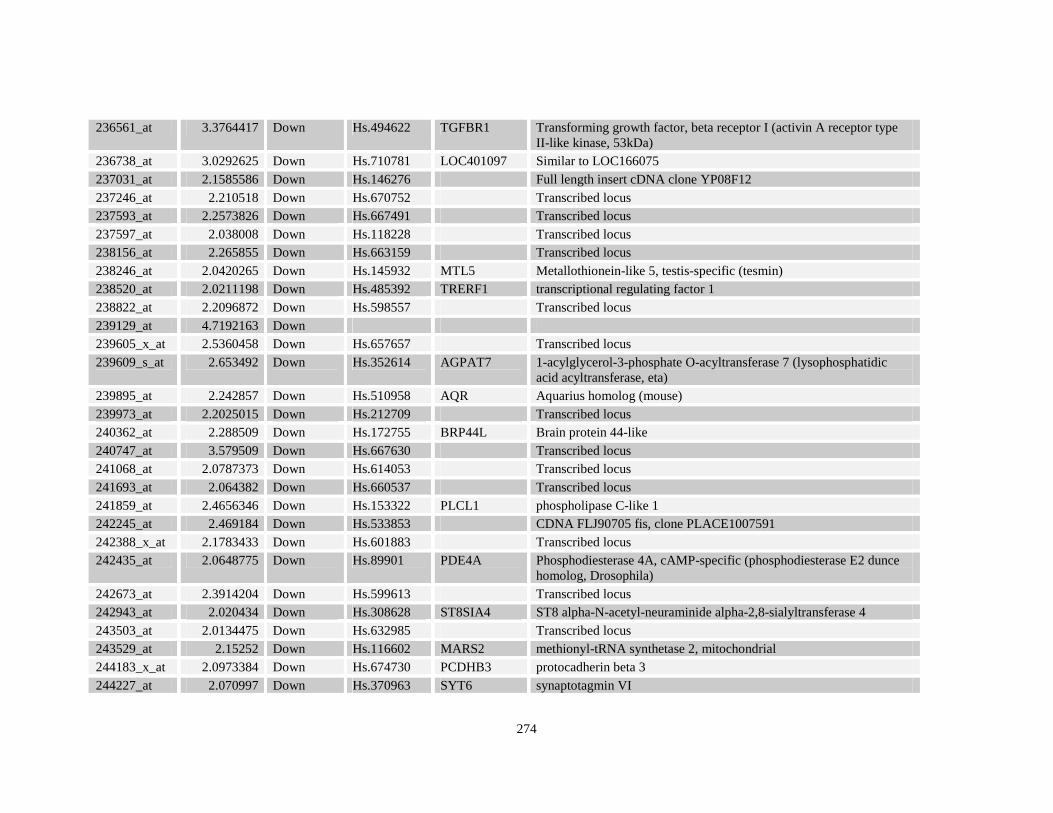
Table A3.1: Genes deregulated greater than 2-fold in U937-NPM-RARA relative to control U937-GFP
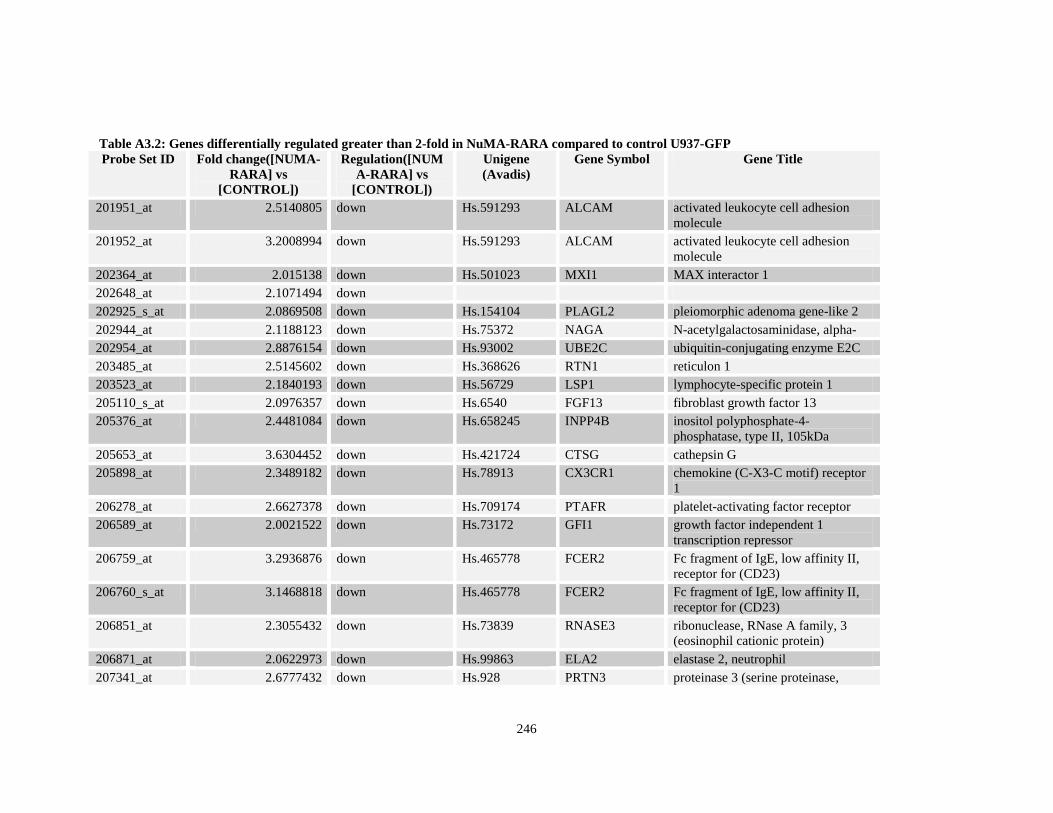
Table A3.2: Genes differentially regulated greater than 2-fold in NuMA-RARA compared to control U937-GFP
Table A3.3: Retinoid responsive gene targets of NPM-RARA
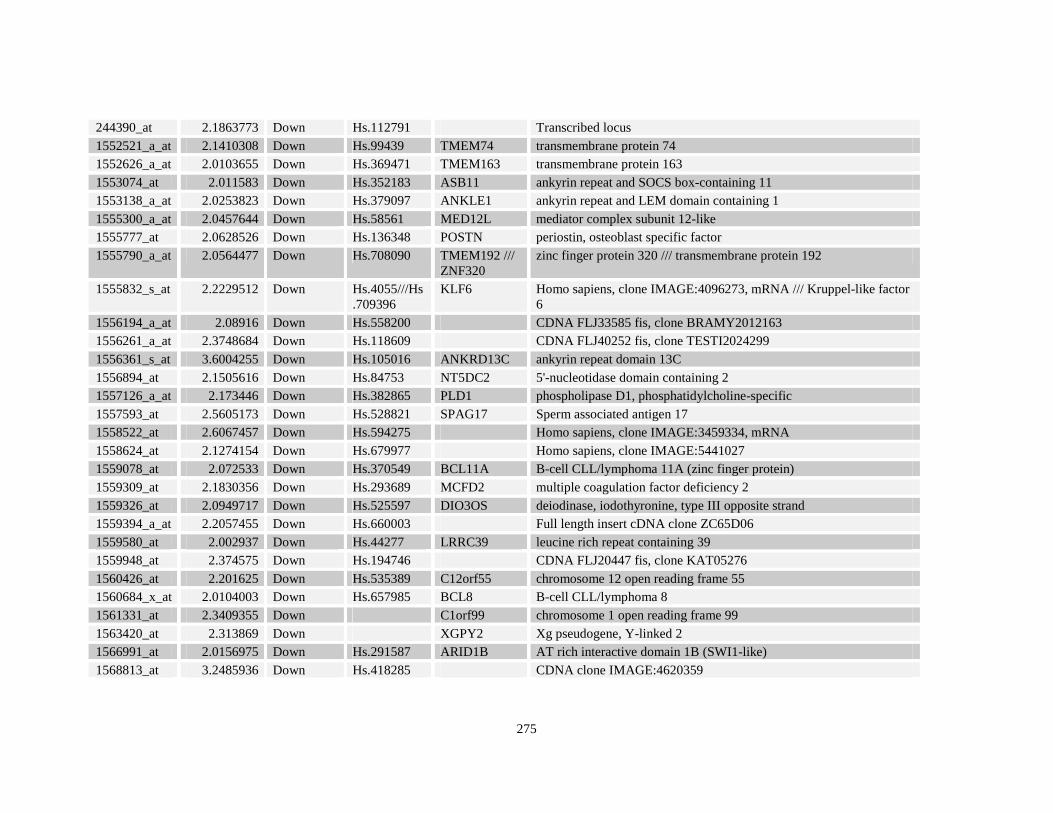
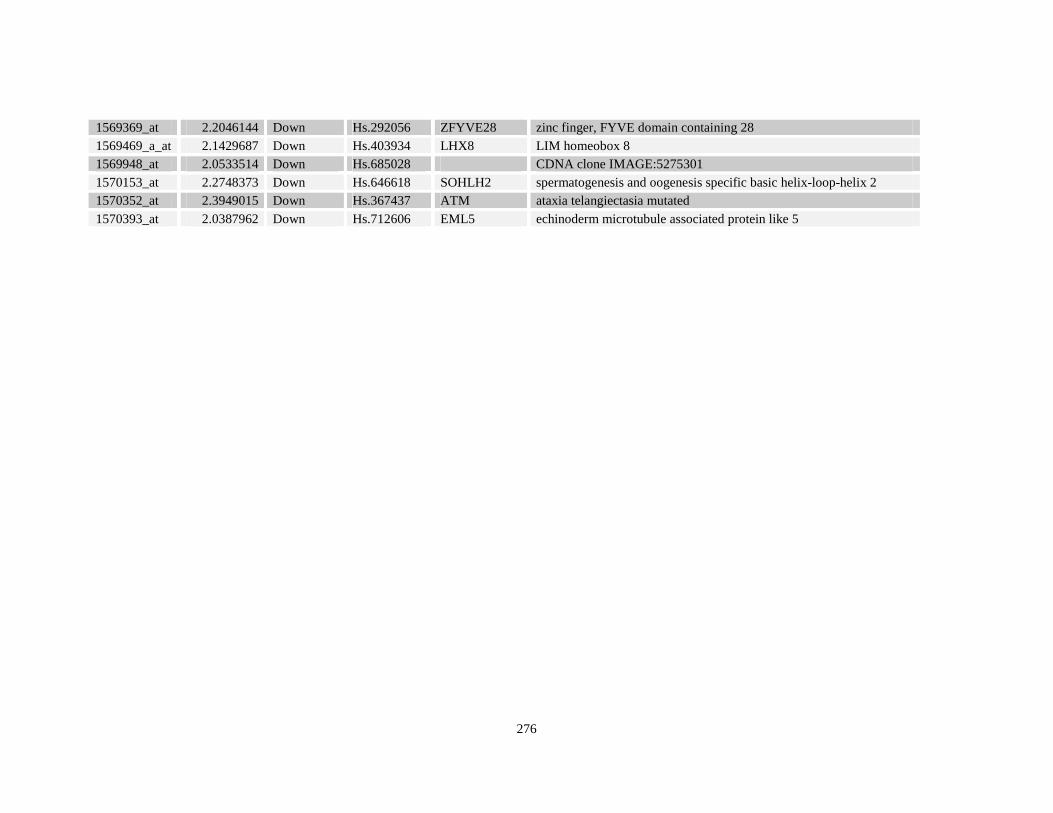
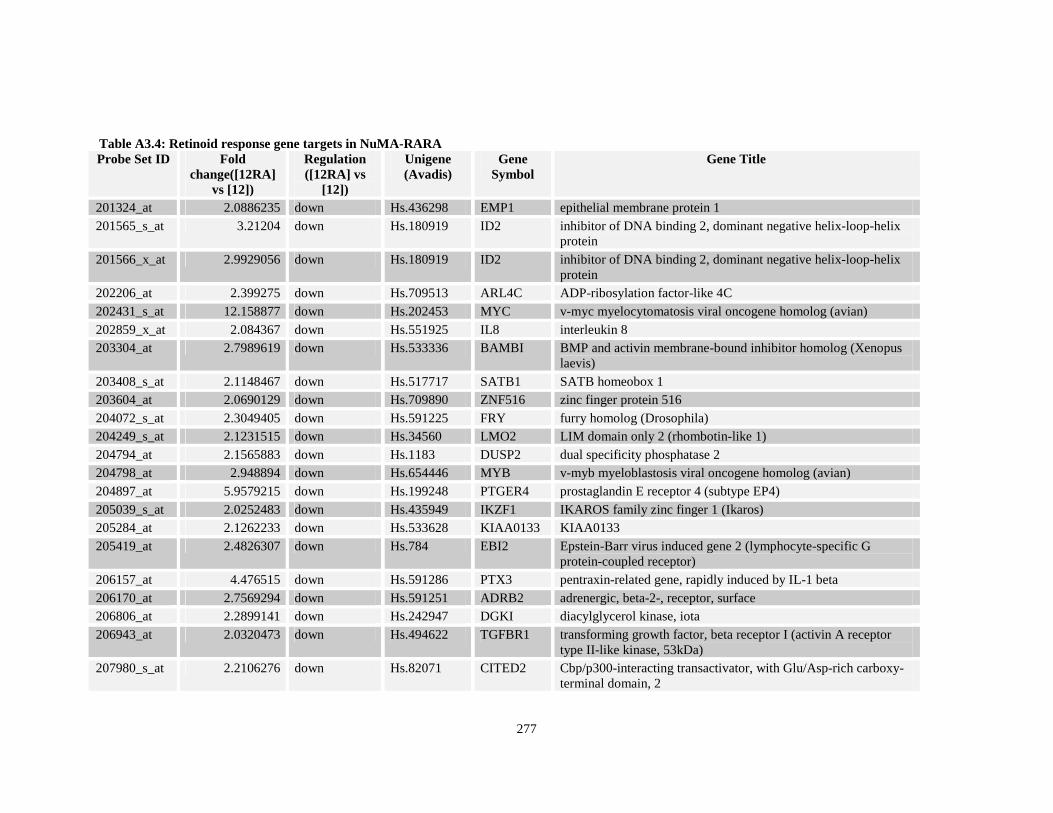
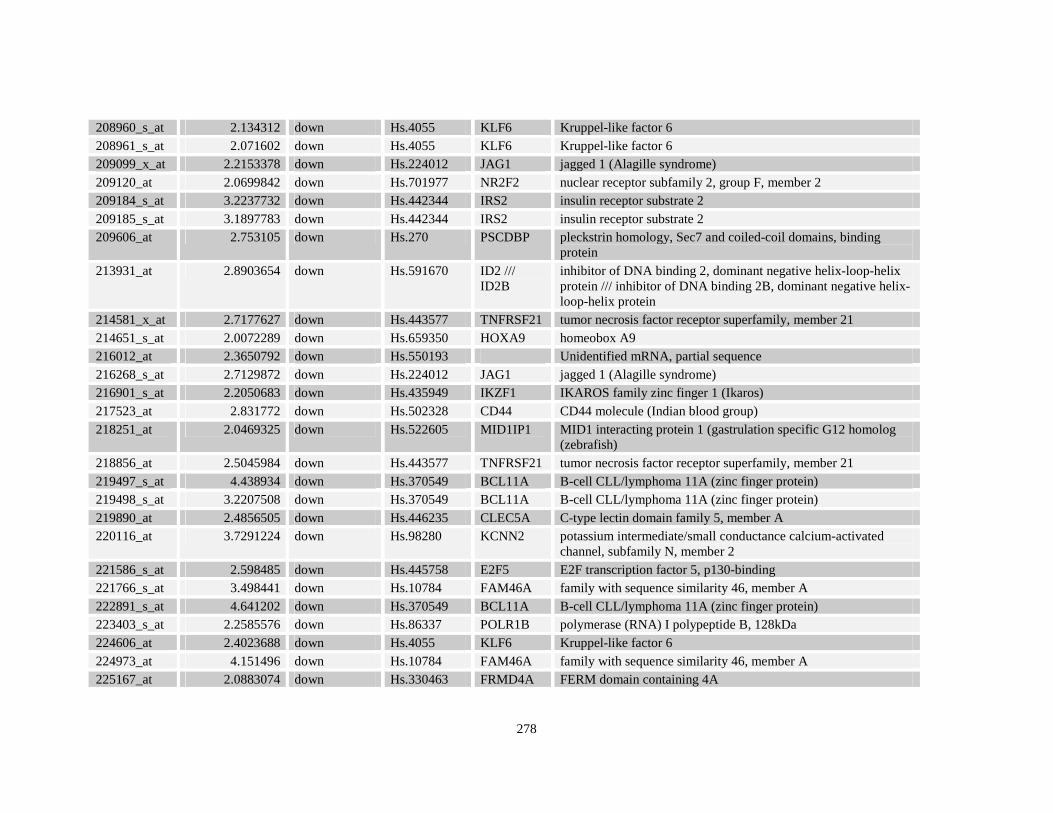
Table A3.4: Retinoid response gene targets in NuMA-RARA
Table A3.5: Retinoid responsive gene targets in control GFP
xviii
Chapter 4: Nucleophosmin is universally deregulated In Acute Promyelocytic Leukemia
Table 4.1: Genes and primer sequences used in real time PCR assays.
Table 4.2: APL patient characteristics.
xix
List of Figures
Chapter 1: Introduction
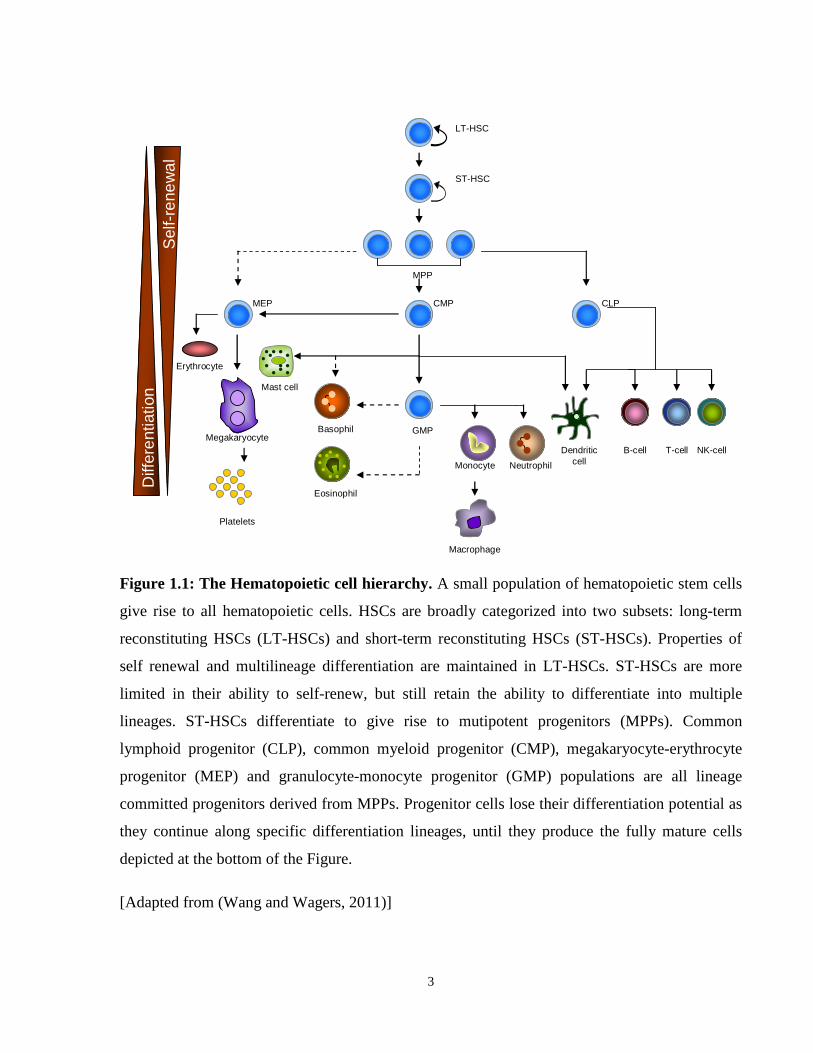
Figure 1.1: Hematopoietic cell hierarchy.
Figure 1.2: Asymmetric versus symmetric cell division in hematopoietic stem cells.
Figure 1.3: BMI-p53 and the PI3K signaling networks regulating HSC self-renewal.
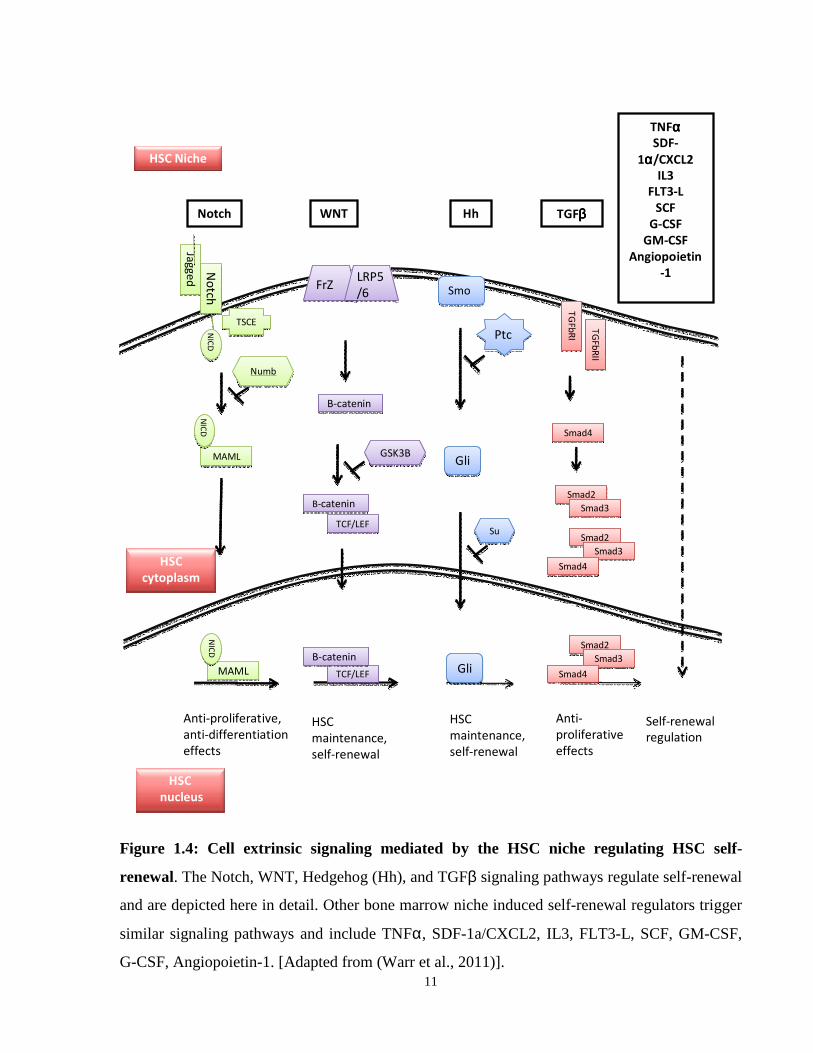
Figure 1.4: Cell extrinsic signaling mediated by the HSC niche regulating HSC self-renewal.
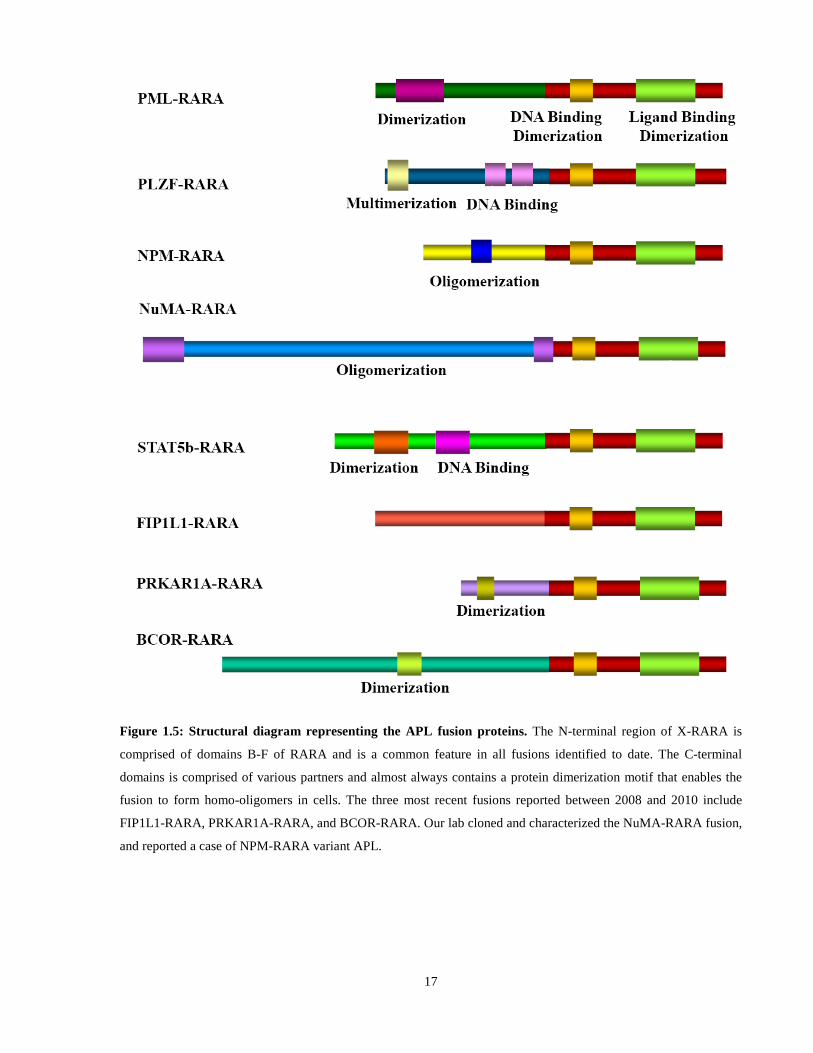
Figure 1.5: General structure of Acute Promyelocytic Leukemia associated fusion proteins.
Figure 1.6: Structural features of Retinoic Acid Receptors (RAR).
Figure 1.7: Structural features of Retinoid X Receptors (RXR).
Figure 1.8: RAR signaling in response to retinoic acid.
Figure 1.9: X-RARA signaling in response to retinoic acid.
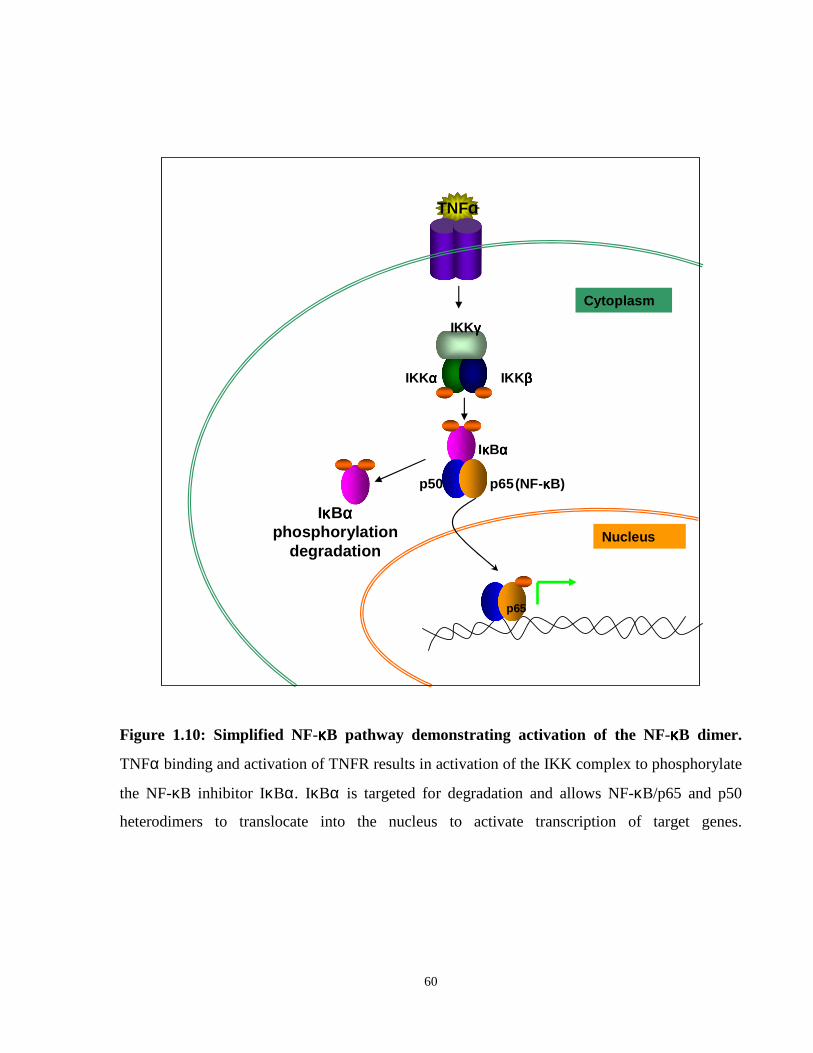
Figure 1.10: NF-κB signaling pathway.
Chapter 2: Deregulated NF-κκκκB signaling in APL
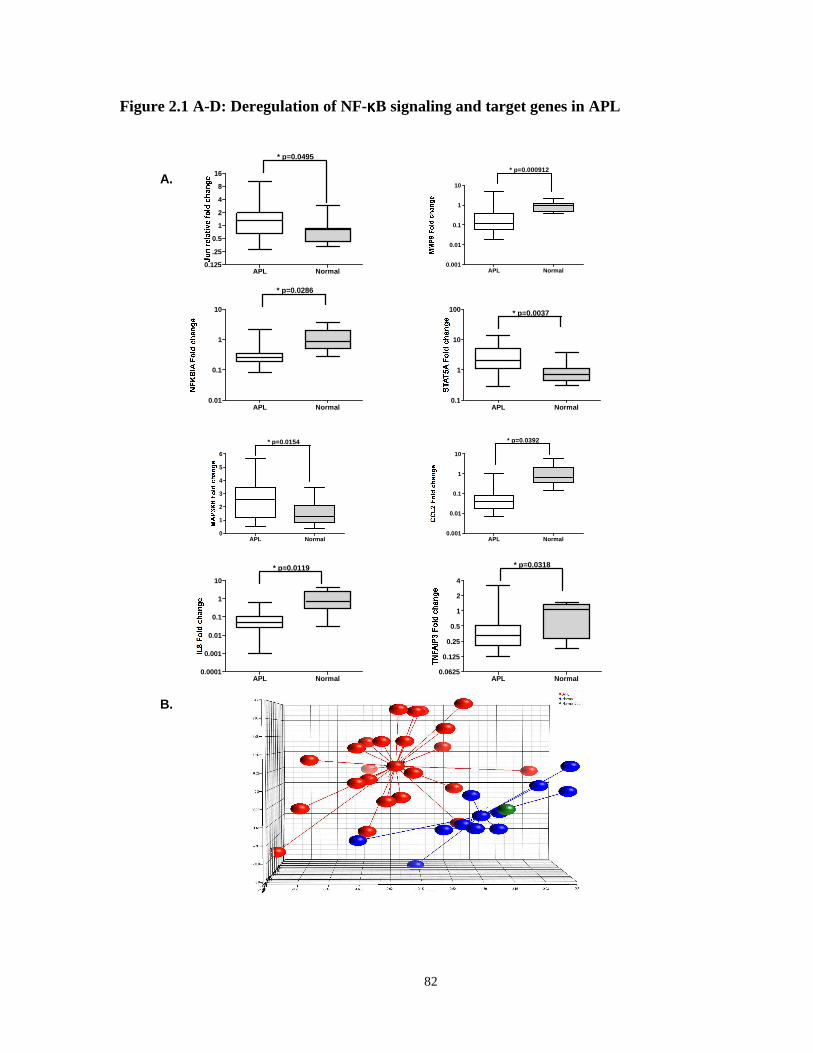
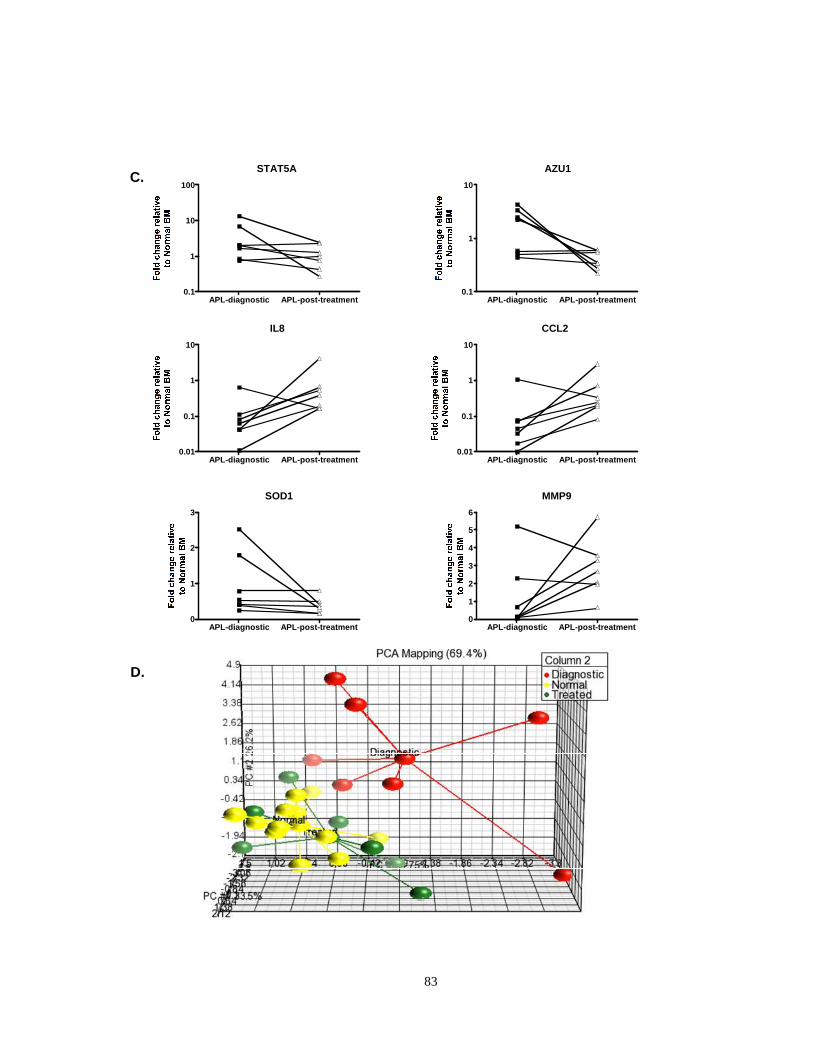
Figure 2.1: Deregulation of NF-κB signaling and target genes in APL.
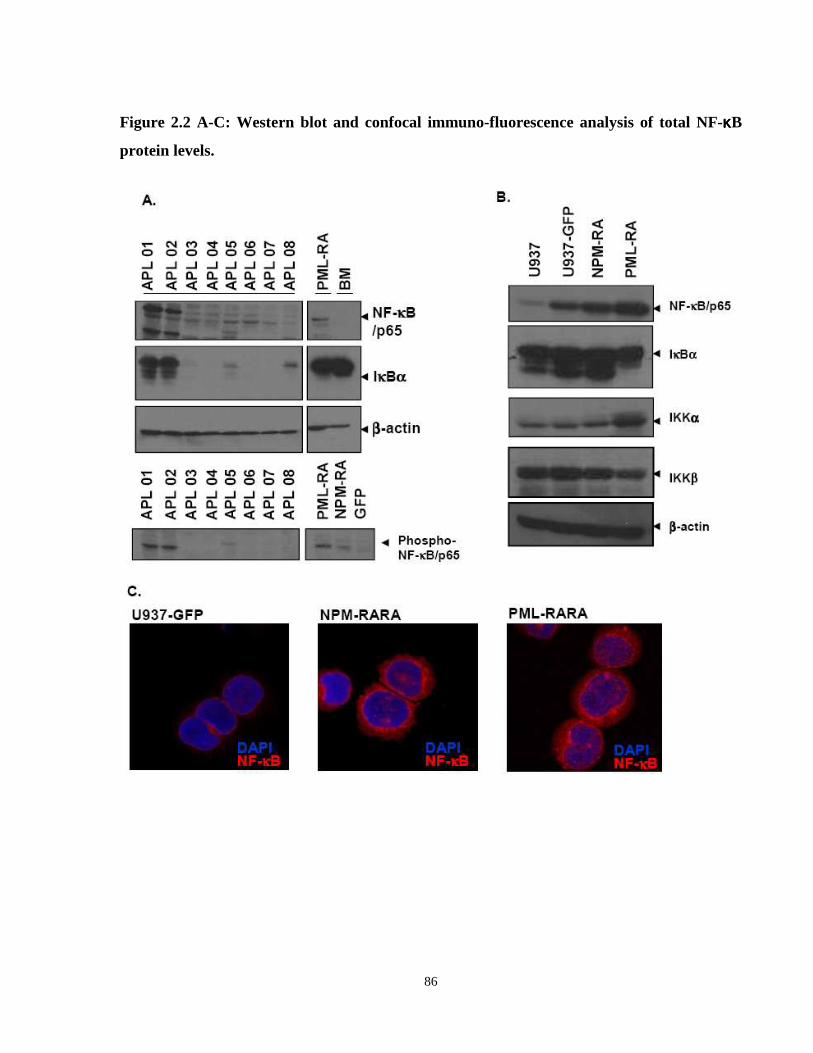
Figure 2.2: Western blot and confocal immunofluorescence analysis of total NF-κB protein levels.
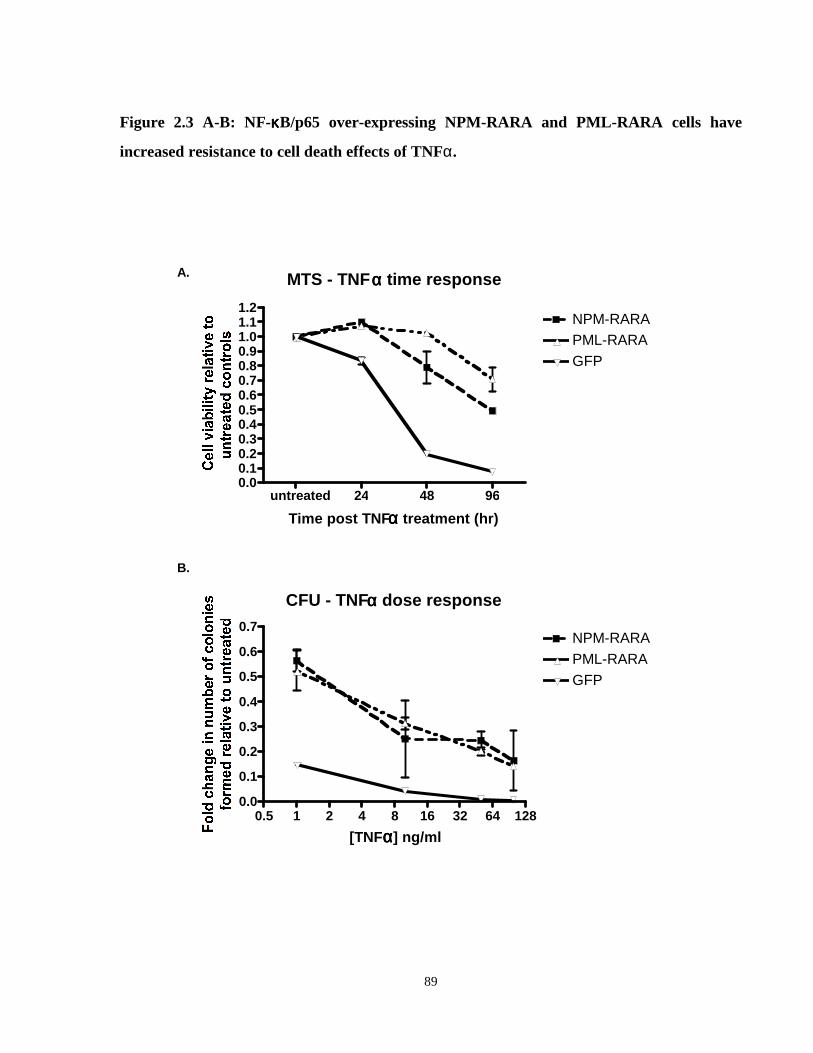
Figure 2.3: NF-κB/p65 over-expressing NPM-RARA and PML-RARA cells have increased resistance to cell death effects of TNFα.
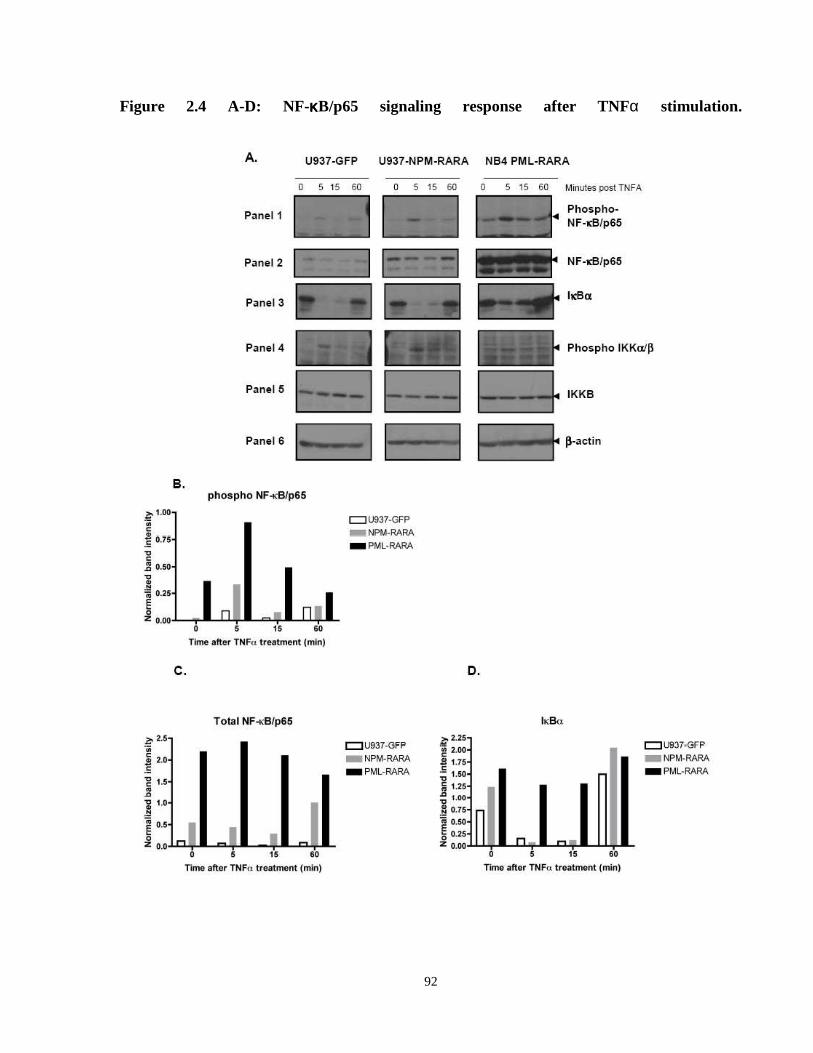
Figure 2.4: NF-κB/p65 signaling response after TNFα stimulation.
Figure 2.5: Expression profiles of the NF-κB/p65 gene interaction network and target genes after TNFα stimulation.
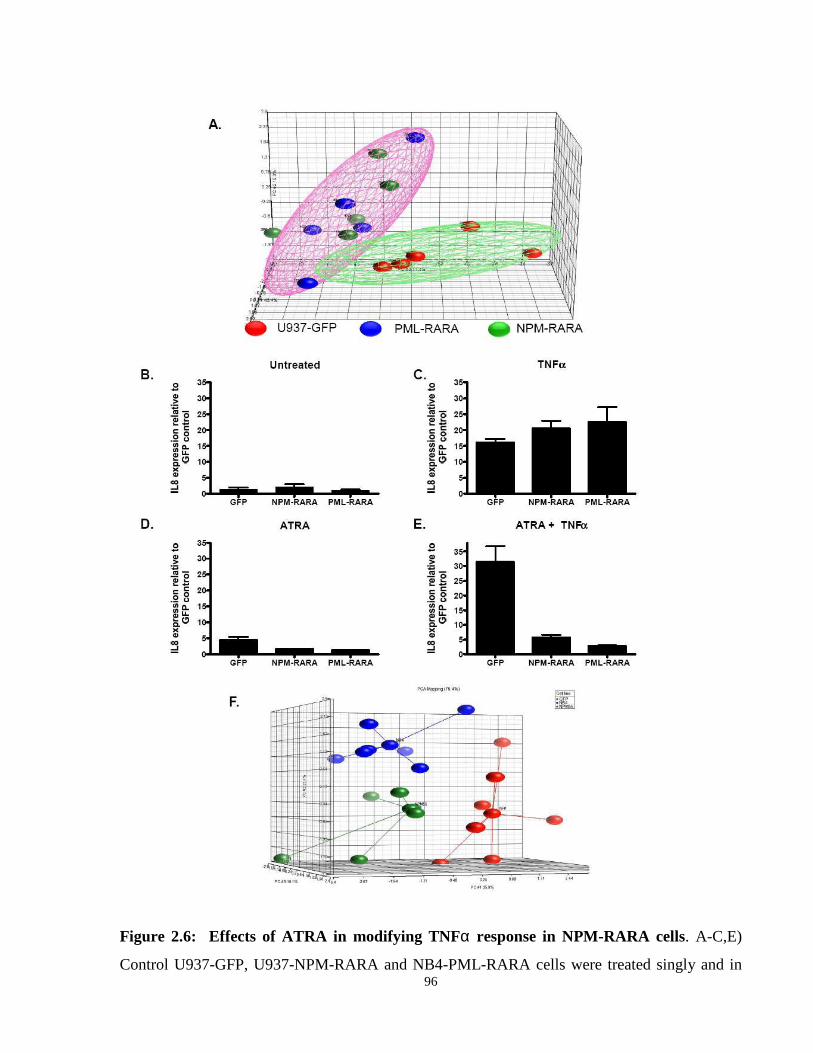
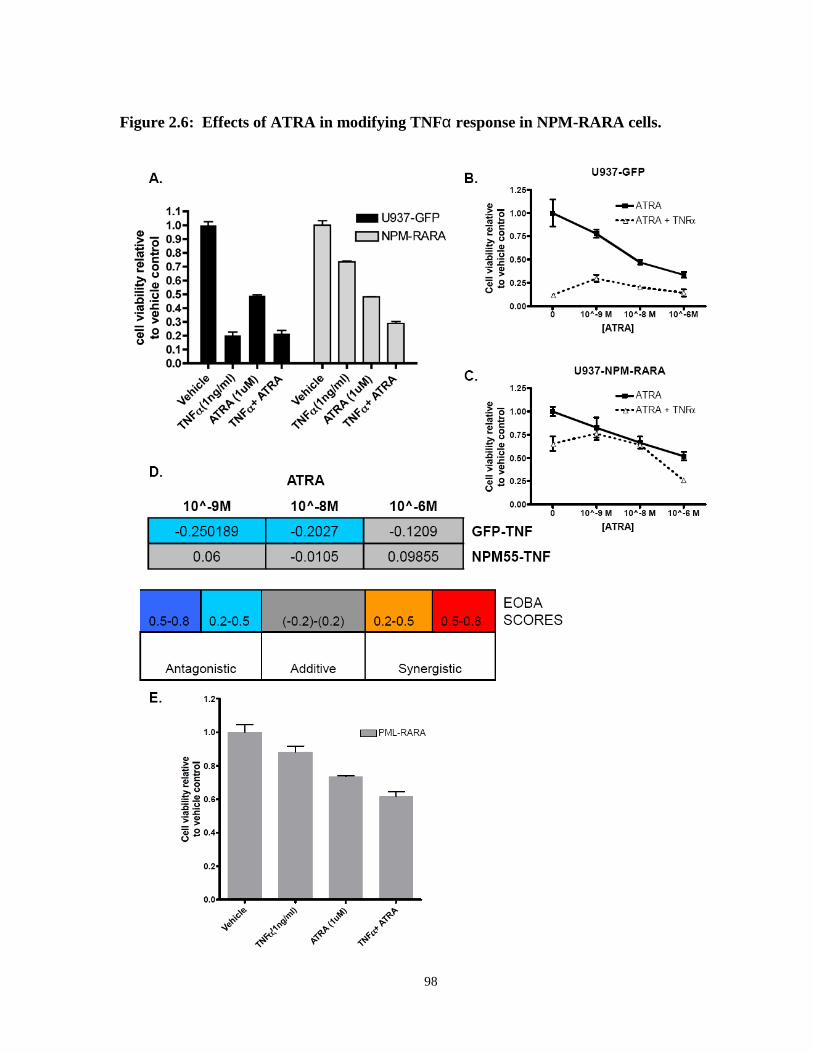
Figure 2.6: Effects of ATRA in modifying TNFα response in NPM-RARA cells.
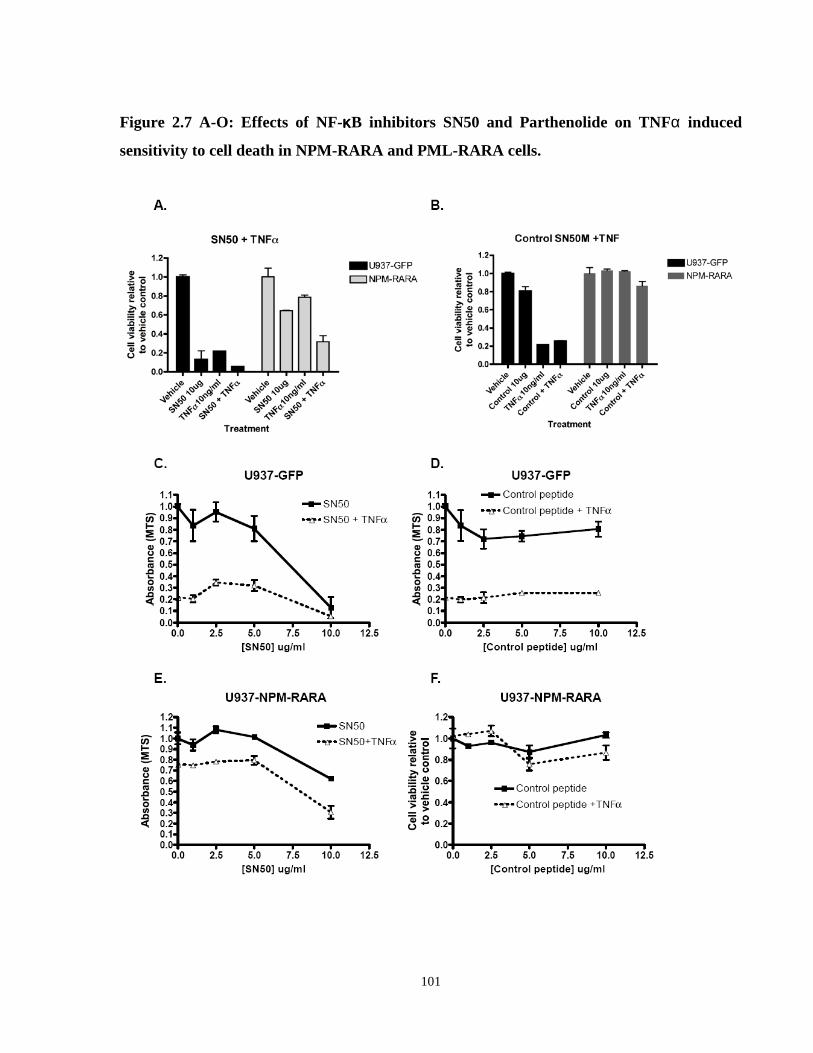
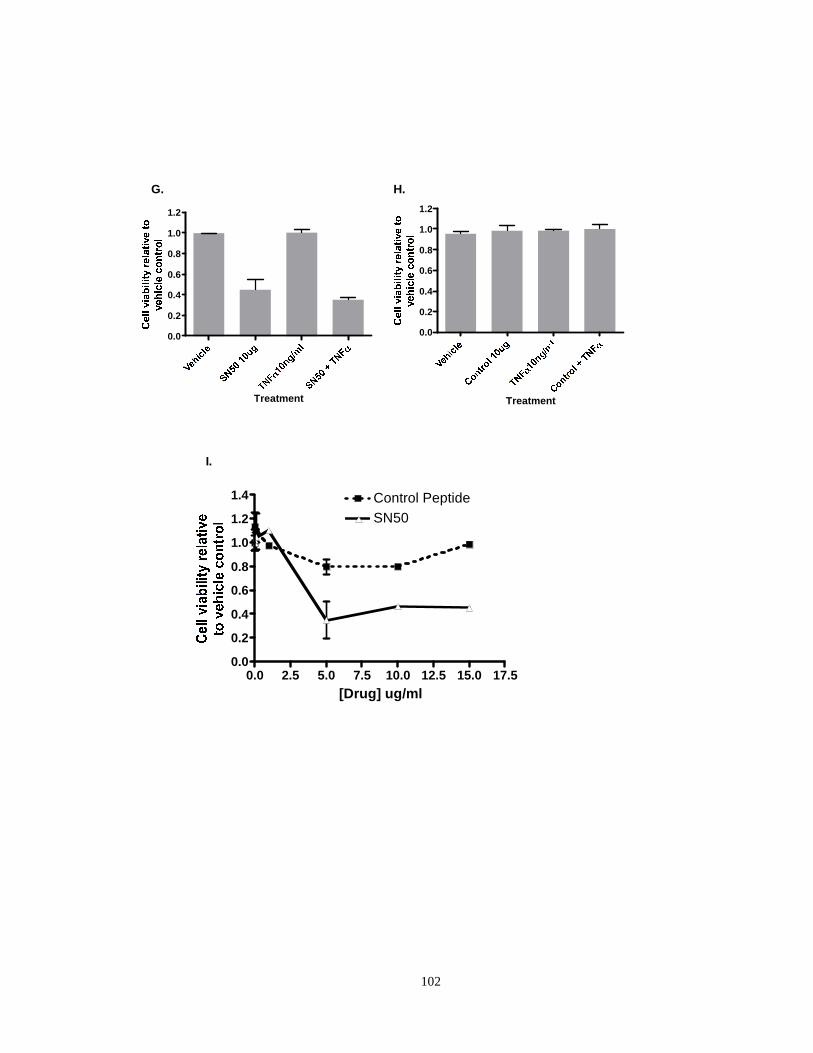
Figure 2.7: Effects of NF-κB inhibitors (SN50 and parthenolide) on TNFα induced sensitivity to cell death in NPM-RARA and PML-RARA cells.
xx
Chapter 3: Gene expression profiling in NPM-RARA and NuMA-RARA variant APL
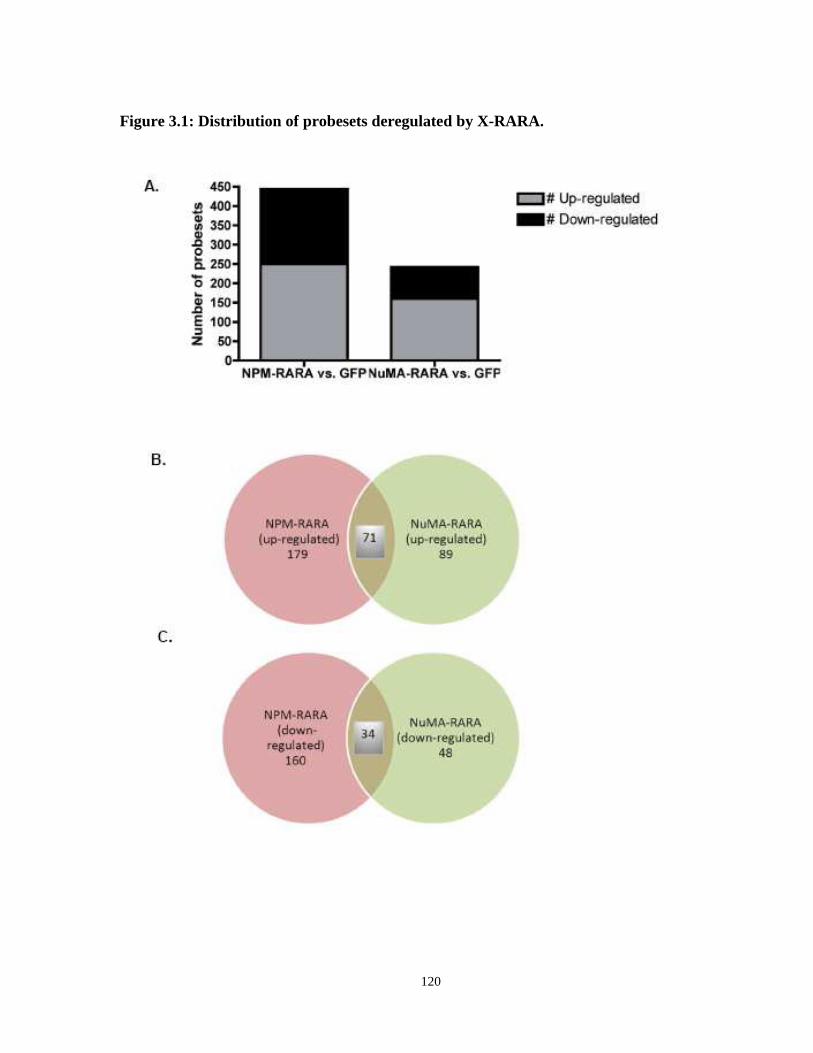
Figure 3.1: Distribution of probesets deregulated by X-RARA.
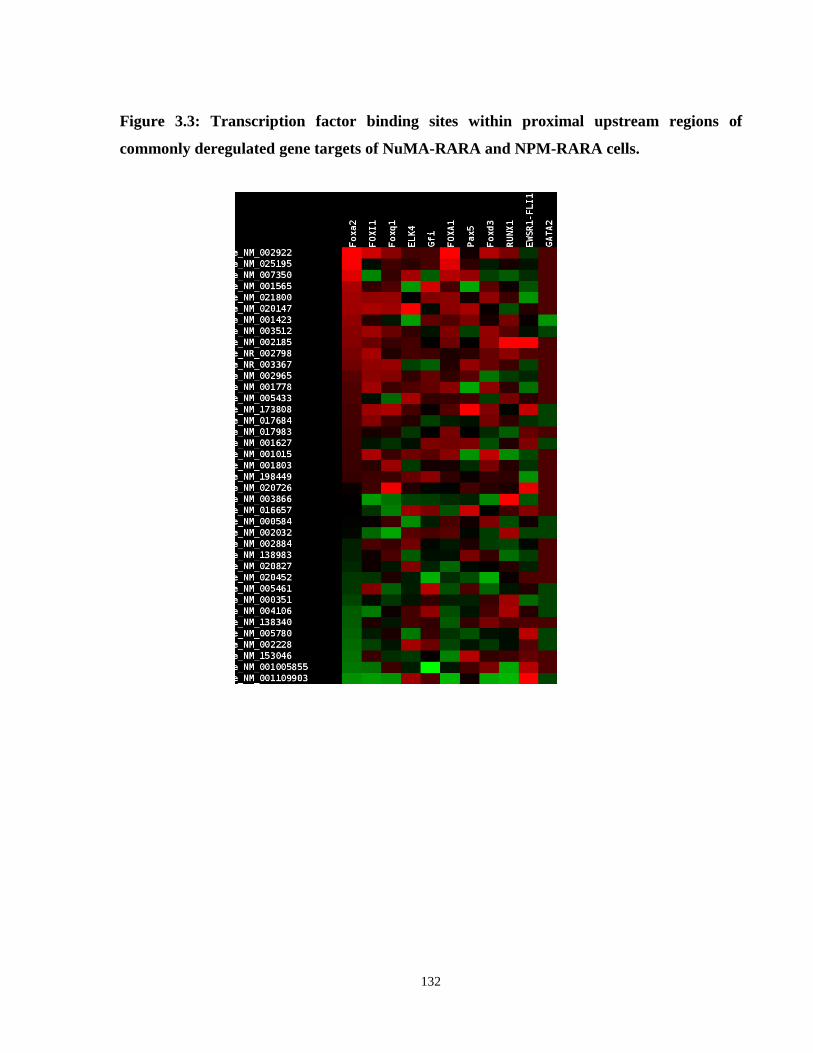
Figure 3.3: Transcription factor binding sites within proximal upstream regions of commonly deregulated gene targets of NuMA-RARA and NPM-RARA cells.
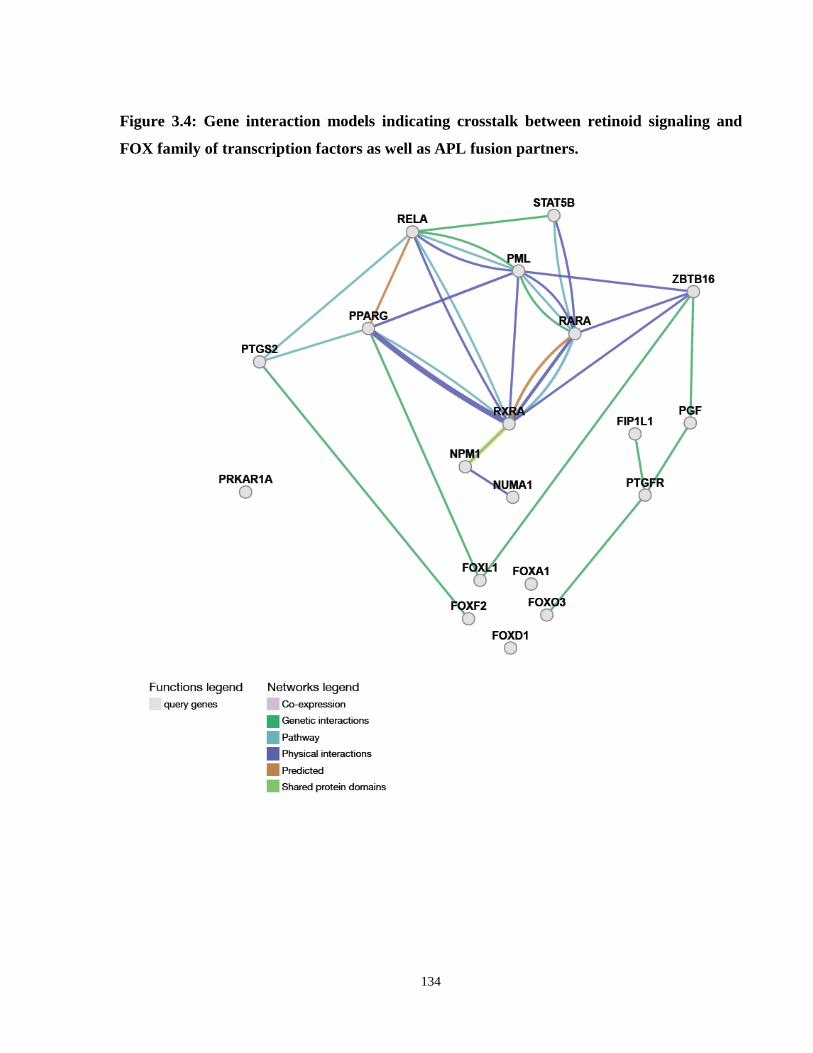
Figure 3.4: Gene interaction models indicating crosstalk between retinoid signaling and FOX family of transcription factors as well as APL fusion partners.
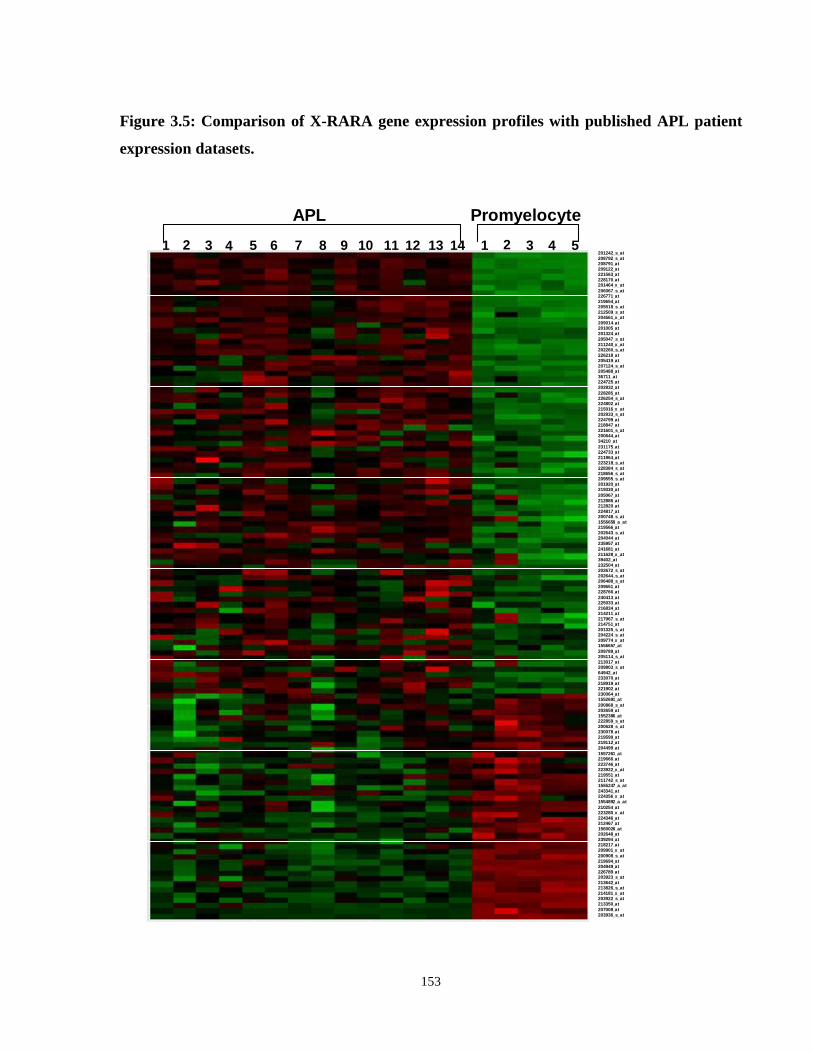
Figure 3.5: Comparison of X-RARA gene expression profiles with published APL patient gene expression datasets.
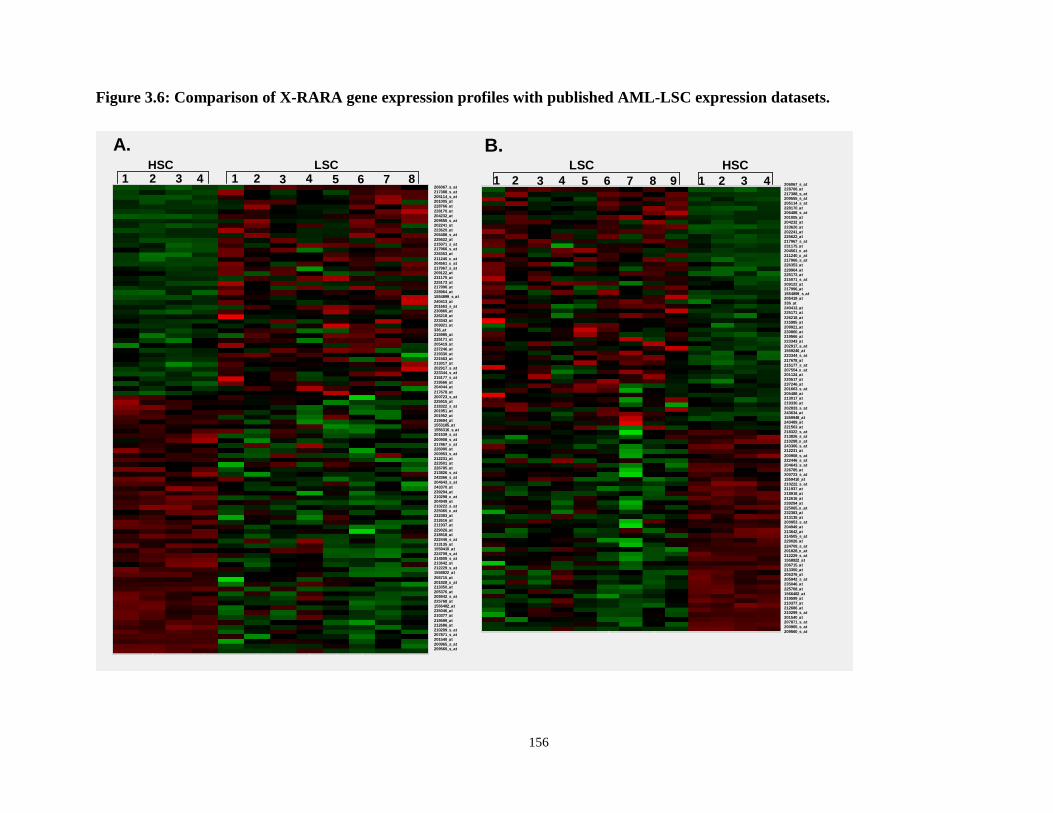
Figure 3.6: Comparison of X-RARA gene expression profiles with published AML-LSC expression datasets.
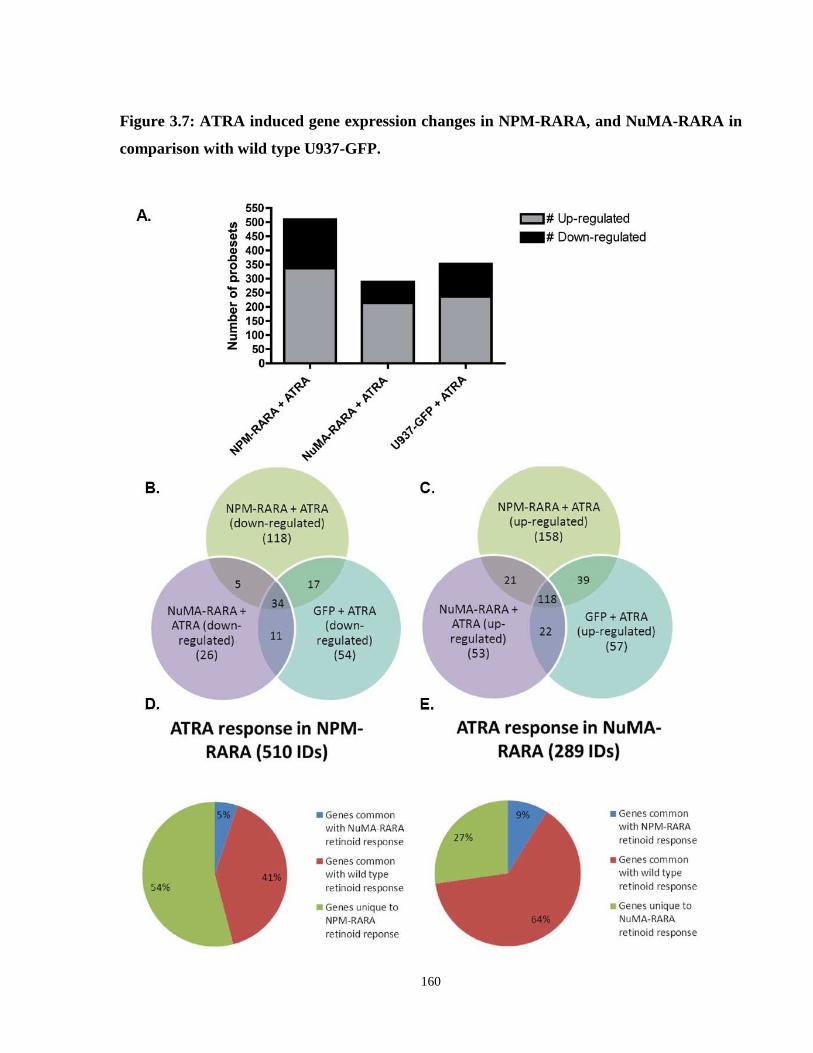
Figure 3.7: ATRA induced gene expression changes in NPM-RARA, and NuMA-RARA in comparison with wild type U937-GFP.
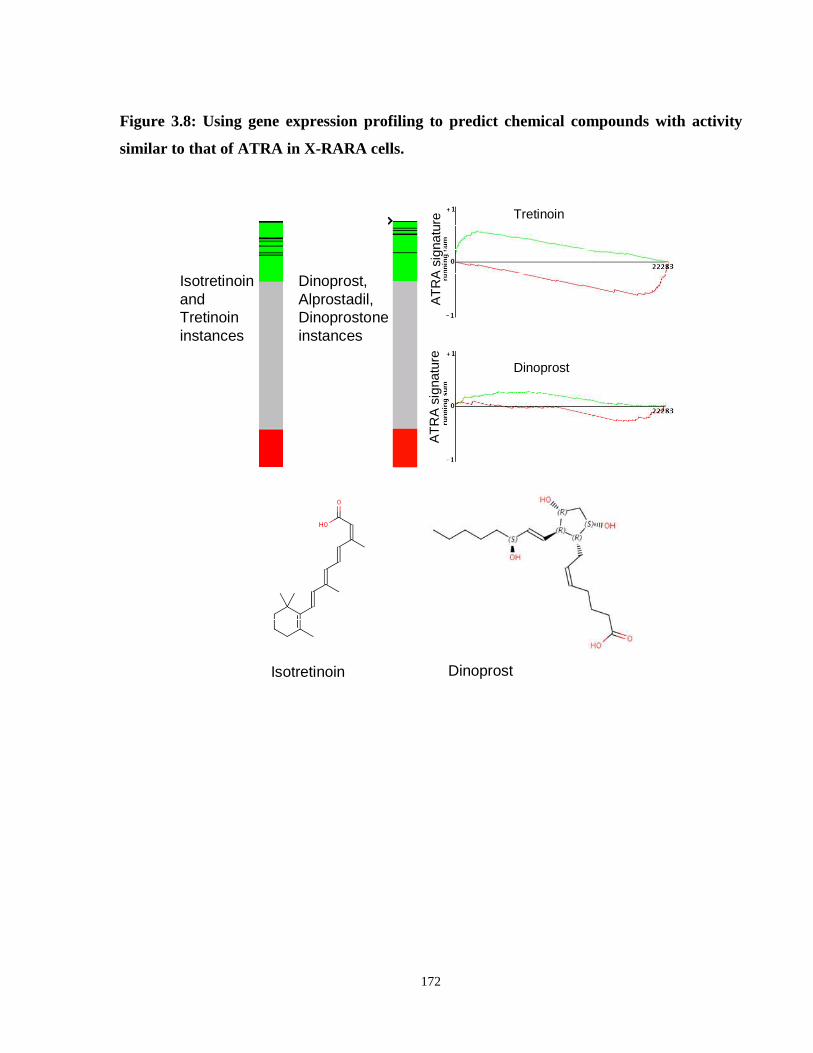
Figure 3.8: Using gene expression profiling to predict chemical compounds with activity similar to that of ATRA in X-RARA cells.
Chapter 4: Deregulated NPM in APL
Figure 4.1: NPM expression in X-RARA+ cells.
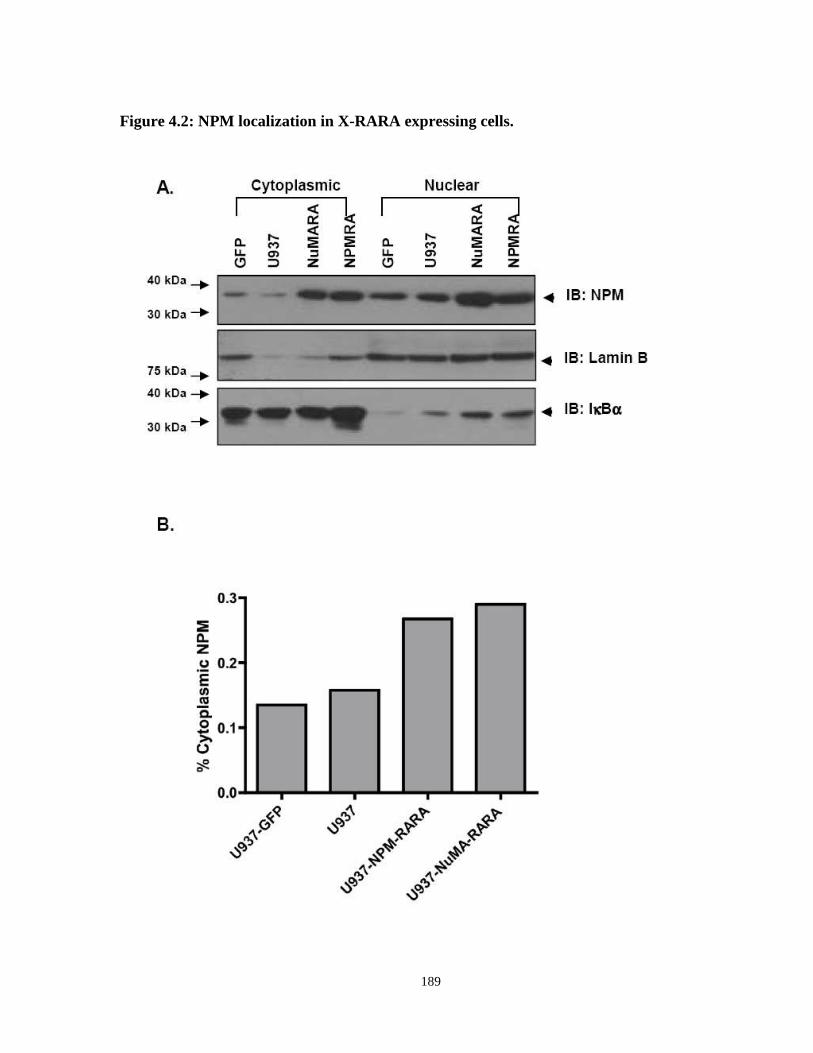
Figure 4.2: NPM localization in X-RARA expressing cells.
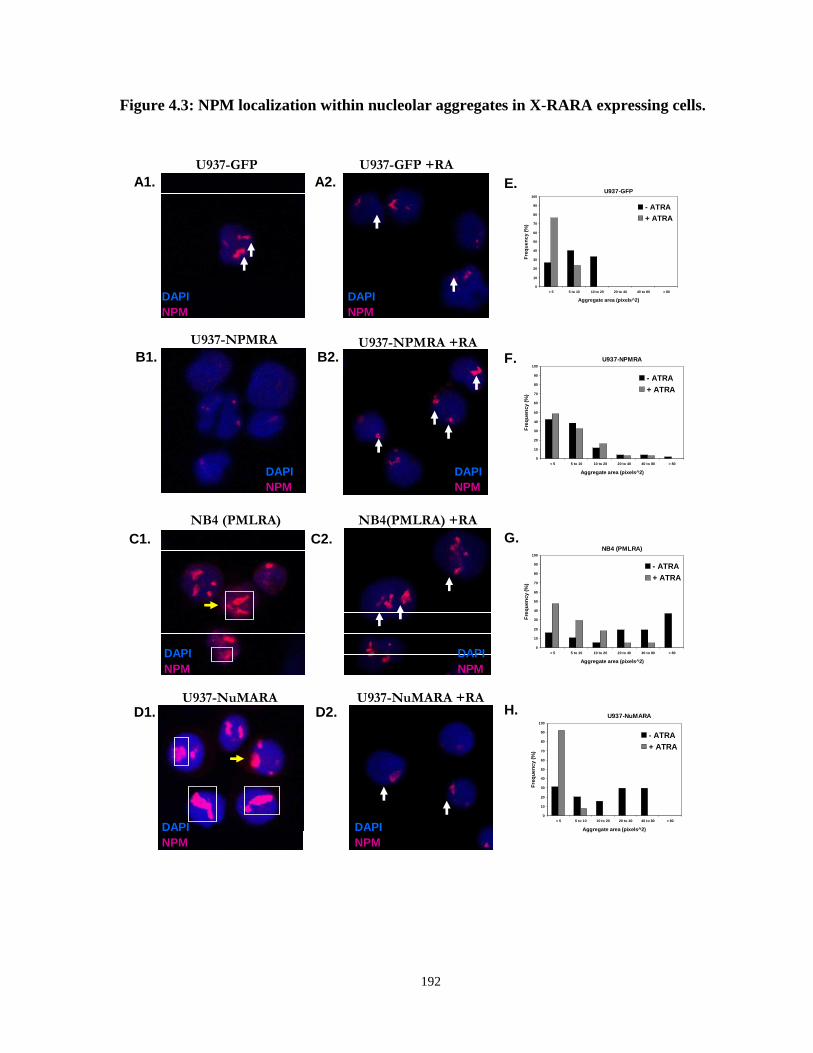
Figure 4.3: NPM localization within nucleolar aggregates in X-RARA expressing cells.
Figure 4.4: NPM half-life in NB4-PML-RARA cells.
Figure 4.5: Localization of NPM in NPMc+ OCI-AML3 cells.
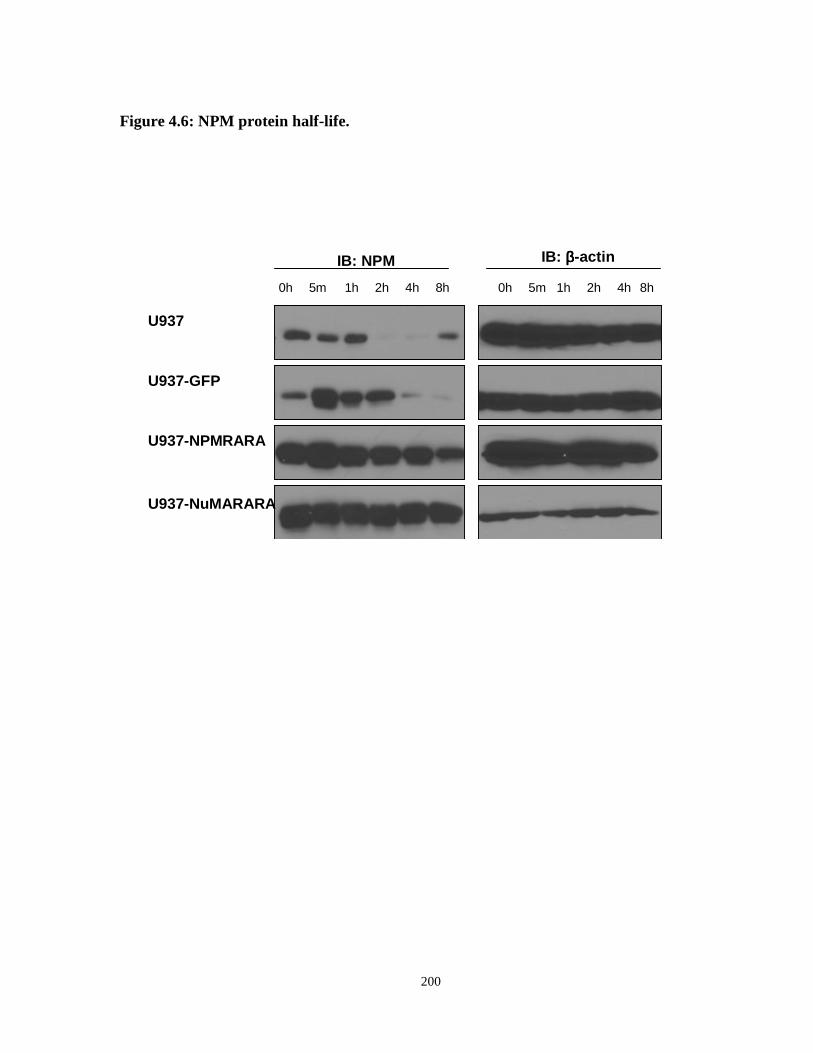
Figure 4.6: NPM protein half-life.
Figure 4.7: Localization of Npm in immature neutrophils from wild-type and transgenic mice.
Figure 4.8: Nucleolar organizing regions in X-RARA+ cells.
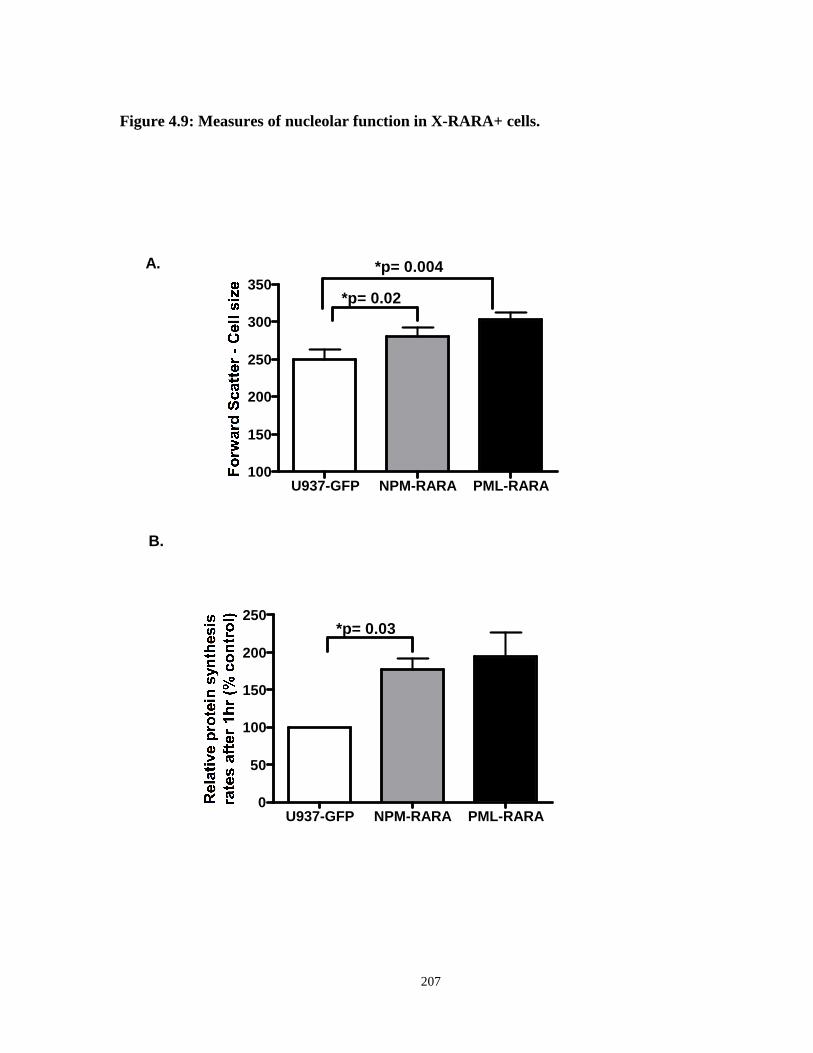
Figure 4.9: Measures of nucleolar function in X-RARA+ cells.
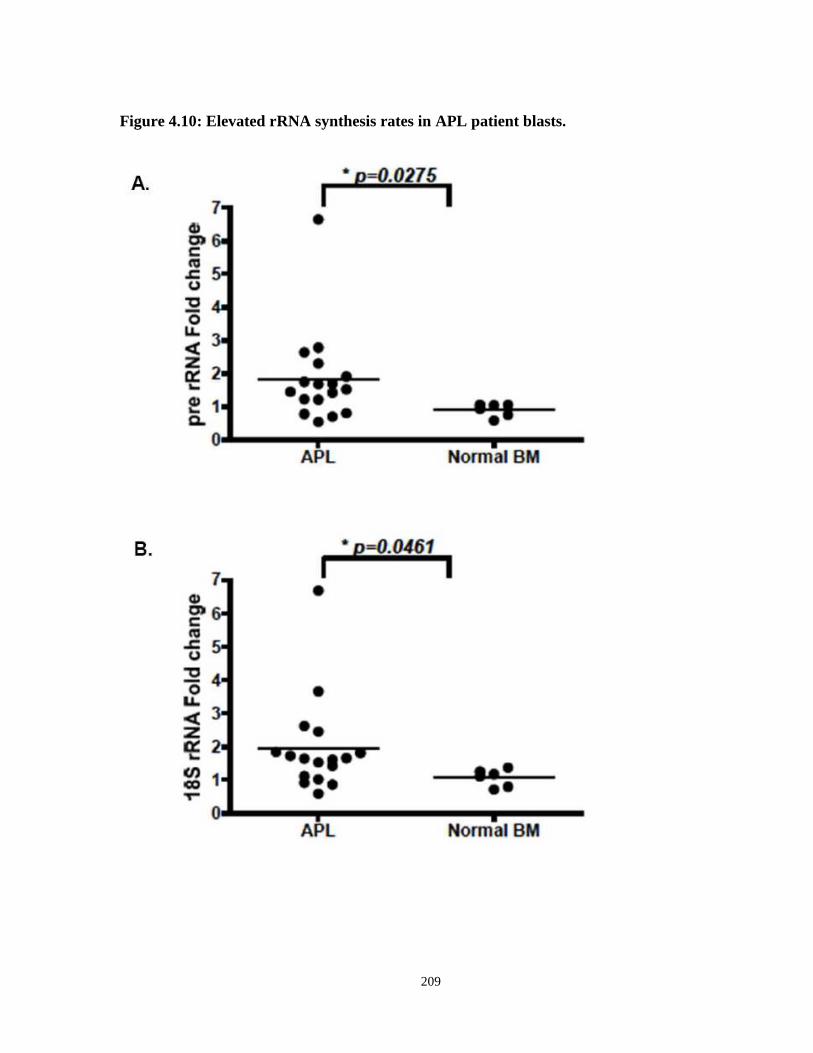
Figure 4.10: Elevated rRNA synthesis rates in APL patient blasts.
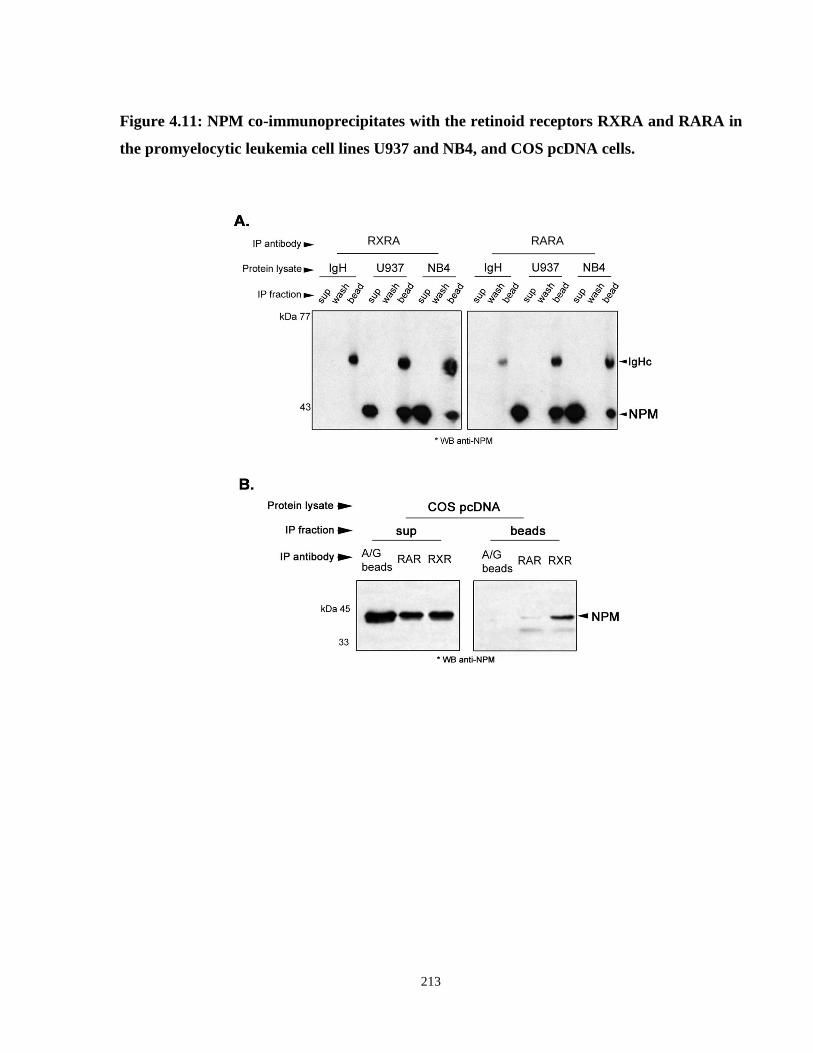
Figure 4.11: NPM co-immunoprecipitates with the retinoid receptors RXRA and RARA in the promyelocytic leukemia cell lines U937 and NB4, and COS pcDNA cells.
xxi
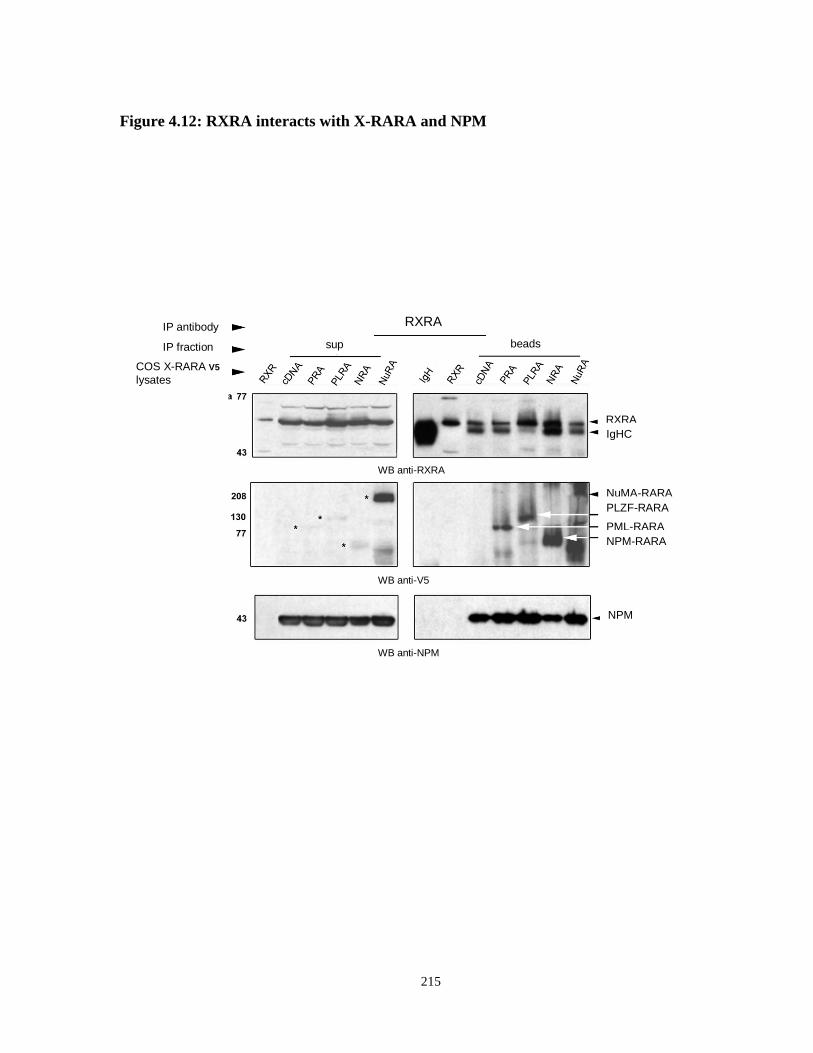
Figure 4.12: NPM and X-RARA co-immunoprecipitate with RXRA in COS X-RARA-V5 cells.
xxii
List of Appendices
Appendix I: Array data tables (Chapter 3)
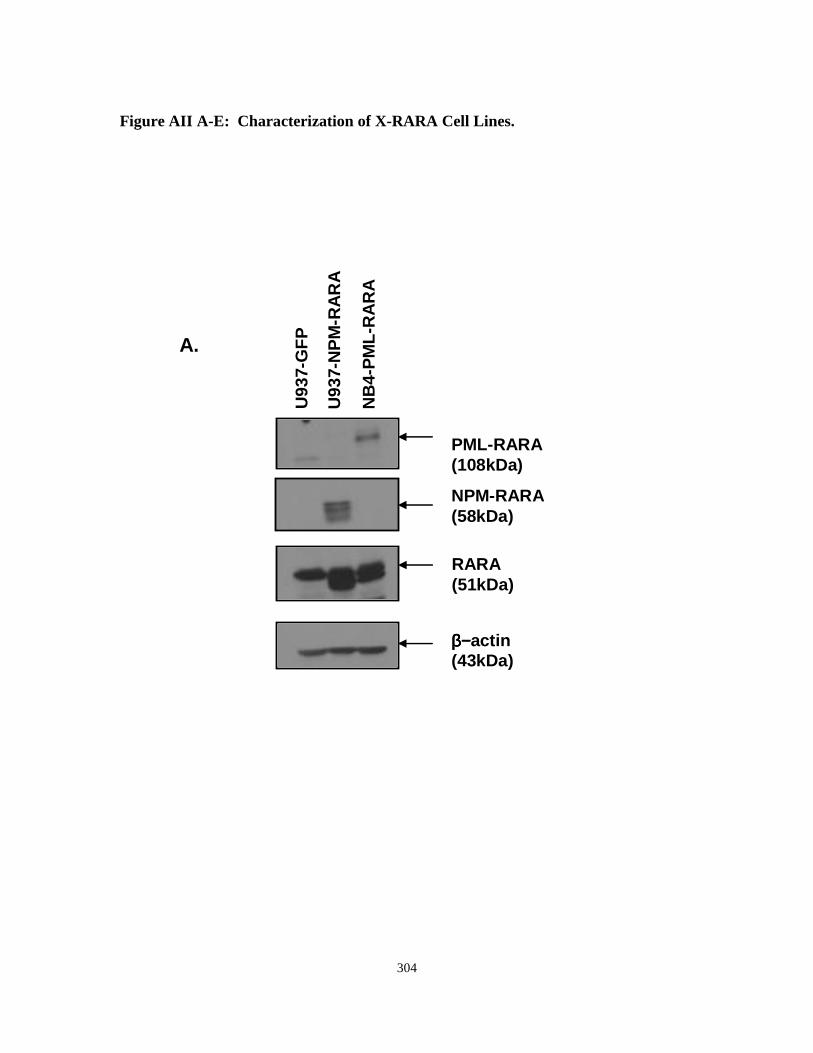
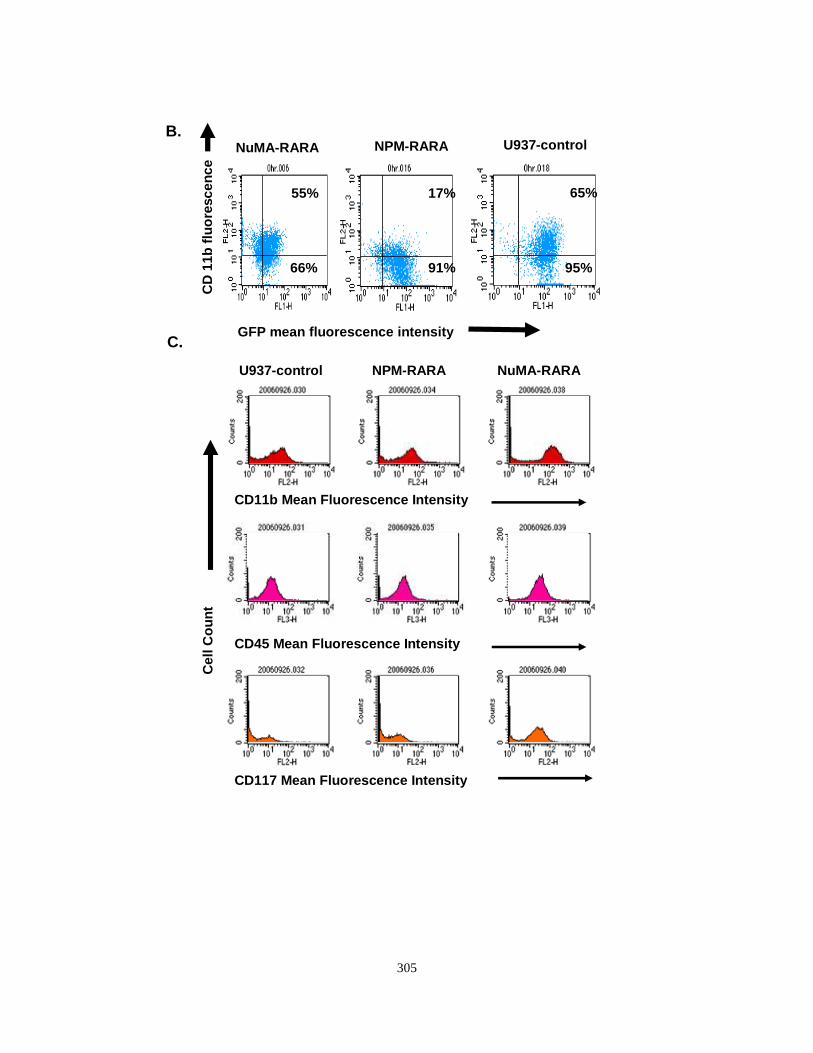
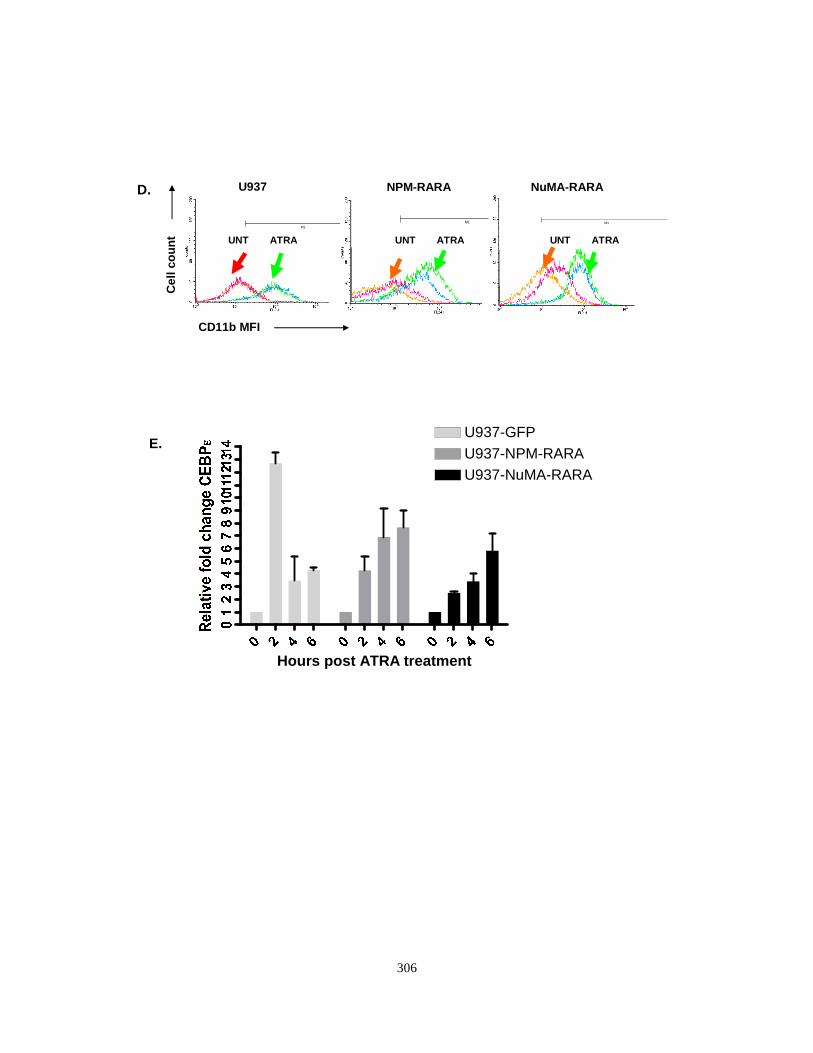
Appendix II: Characterization of U937 cell lines expressing NuMA-RARA and NPM-RARA
1
Chapter 1
Introduction
Material contained in section 1.4 was adapted from the following publication:
Thomas, M., Sukhai, M., and Kamel-Reid, S. (2008). Many paths to one disease: the role of the
variant fusion proteins NPM-RARA and NuMA-RARA in acute promyelocytic leukemia
biology. Cell Science Reviews 4.
Material contained in sections 1.2 and 1.4 were adapted from the following publication:
Thomas, M., Sukhai, M., Kamel-Reid, S. (2012). An Emerging Role for Retinoid X Receptor A
in Malignant Hematopoiesis. Leukemia Research 36(9): 1075-81.
Material contained in sections 1.2, and 1.5 were adapted from the following publication:
Goswami, R. S*, Sukhai, M. A.*, Thomas, M.*, Reis, P. P., and Kamel-Reid, S. (2009).
Applications of microarray technology to Acute Myelogenous Leukemia. Cancer Inform 7, 13-
28. (* Equal contribution).
2
1.1 Normal Hematopoiesis
1.1.1 Model of hematopoietic hierarchy
Hematopoiesis, or blood cell formation, is the process whereby hematopoietic stem cells (HSCs)
give rise to lineage-committed progenitors and end-stage mature cells. HSCs are present at the
apex of the hematopoietic differentiation hierarchy, and have properties of multipotency and
self-renewal. Multipotency is the ability to differentiate into multiple types of cells of the blood
system, while self-renewal is the ability to give rise to identical daughter cells with the same
multipotent properties as the parent cell. Under steady-state conditions, HSCs are maintained in a
quiescent state and their numbers are tightly regulated (Jude et al., 2008). HSC’s are functionally
classified as long term repopulating (LT-HSC) or short term repopulating (ST-HSC) according to
their ability to establish life long, or transient hematopoiesis (Chumsri et al., 2007). When
stimulated to differentiate, HSCs give rise to multipotent progenitors (MPP), which then
differentiate into lineage committed progenitors, and ultimately produce all the differentiated
cells comprising the blood system (Figure 1.1) (Jude et al., 2008). HSCs initially lose the
property of self-renewal upon differentiation to progenitors, and then lose multipotency as they
become progressively committed to a particular lineage. Hematopoietic progenitors are
developmentally more restricted in their lineage commitment, and give rise to developmentally
mature blood cells, which lose the ability to proliferate and self-renew. Models of hematopoiesis
have classified ten HSC-derived blood cell lineages comprising of myeloid/erythroid cells
Figure 1.2: Asymmetric versus symmetric cell division. Quiescent HSCs remain adhered to
the stem cell niche through cell adhesion molecules. Once activated, HSCs can undergo
asymmetric cell division where they can generate a daughter cell that completely resembles the
parent HSC through self-renewal, and a progenitor cell that is more differentiated. Following cell
division and differentiation, HSCs lose properties of self renewal and multipotency.
[Adapted from (Suda et al., 2005)]
9
1.3.1 Signaling networks involved in maintaining self-renewal
Regulation of HSC self-renewal is critical to ensure the maintenance of adequate numbers of
HSCs. Maintaining HSC numbers through self-renewal requires the activation of proliferation
pathways, and a suppression of signals leading to apoptosis, differentiation and cell death. (Akala
2006). Cellular networks involved in positively regulating self-renewal in HSCs include the cell
cycle regulation (Bmi-p53) network, and the PI3K signaling pathway (reviewed in (Warr et al.,
2011) (Figure 1.3). In addition, HSC self-renewal also involves cell extrinsic signaling and
critical signals provided by the bone marrow niche including WNT, TGFβ, Hedgehog, and
Notch. The “bone marrow niche” is a term used to describe the micro-environment surrounding
the HSC, as well as the physical interaction between HSCs and the specialized cells that support
them. Niche cells provide membrane-bound and secreted factors, cytokines, chemokines,
adhesion molecules, and their receptors which are essential for survival, quiescence, and
differentiation (Schofield, 1978; Wilson et al., 2007). Other critical extrinsic signals that have
been identified to date include SDF-1α/CXCL12, Angiopoietin-1, IL-3, FLT3-Ligand, SCF, G-
CSF, and GM-CSF (Trumpp et al., 2010). Additional factors that are expressed in the BM niche,
and are essential for HSC self-renewal include TGF-β, Wnt, Hedgehog, and Notch pathway
components (Figure 1.4).
10
BMI-1BMI-1
p16p16
RBRB
p19p19
P53P53
Cell cycle
arrest •Apoptosis
•DNA repair
•Senesence
PI3KPI3K
AKTAKT
FOXOFOXOTSC1
/2
TSC1
/2
mTORC
1
mTORC
1
•Cell cycle
•Apoptosis
•Protection
from O2
stress
•Cell growth
•Translation
•Cell cycle
progression
PMLPML
PTENPTEN
HSC
Growth Factors
Oxygen levels
Nutrients
DNA damage
Figure 1.3: BMI-p53 and the PI3K signaling networks. Both cellular networks affect HSC
self-renewal by regulating cellular survival, cell cycle, DNA repair, growth and senescence.
Molecules coloured in green are positive modulators of self-renewal in HSCs; those coloured in
red are inhibitors of self-renewal and promote quiescence or apoptosis.
[Adapted from (Warr et al., 2011)]
11
Notch WNT Hh TGFββββ
Jag
ge
dJa
gg
ed
No
tchN
otch
MAMLMAML
MAMLMAML
FrZFrZLRP5
/6
LRP5
/6
B-cateninB-catenin
TCF/LEFTCF/LEF
SmoSmo
PtcPtc
GliGli
TG
Fb
RI
TG
Fb
RI
TG
FbR
IIT
GFb
RII
Smad4Smad4
Smad2Smad2
Smad3Smad3
Smad2Smad2
Smad3Smad3
Smad4Smad4
TSCETSCE
NIC
DN
ICD
NIC
D
B-cateninB-catenin
TCF/LEFTCF/LEF
B-cateninB-catenin
GliGli
Smad2Smad2
Smad3Smad3
Smad4Smad4
SuSu
GSK3BGSK3B
NumbNumb
HSC
cytoplasm
HSC Niche
Anti-
proliferative
effects
HSC
maintenance,
self-renewal
HSC
maintenance,
self-renewal
Anti-proliferative,
anti-differentiation
effects
HSC
nucleus
TNFααααSDF-
1αααα/CXCL2
IL3
FLT3-L
SCF
G-CSF
GM-CSF
Angiopoietin
-1
Self-renewal
regulation
Figure 1.4: Cell extrinsic signaling mediated by the HSC niche regulating HSC self-
renewal. The Notch, WNT, Hedgehog (Hh), and TGFβ signaling pathways regulate self-renewal
and are depicted here in detail. Other bone marrow niche induced self-renewal regulators trigger
similar signaling pathways and include TNFα, SDF-1a/CXCL2, IL3, FLT3-L, SCF, GM-CSF,
G-CSF, Angiopoietin-1. [Adapted from (Warr et al., 2011)].
12
1.3.1.1 Negative regulators of self-renewal
Cell intrinsic negative regulators of HSC maintenance and self-renewal include the cyclin
dependent kinase inhibitor, p21CIP1/WAF1 (Cheng et al., 2000), GFI1 (Hock et al., 2004), and LNK
(Buza-Vidas et al., 2006). In general, these factors work to regulate the size of the HSC
compartment by restricting entry of stem cells into the cell cycle (Cheng et al., 2000), and
restricting their proliferation (Hock et al., 2004), or negatively regulating cytokine-induced HSC
expansion (Buza-Vidas et al., 2006). Cell extrinsic factors such as transforming growth factor
(TGF)-β (Larsson et al., 2005), interferon (IFN)-γ (Yang et al., 2005), interleukin (IL)-3
(Yonemura et al., 1996) and tumor necrosis factor (TNF)-α (Bryder et al., 2001) negatively
affect HSC growth in vitro. Multiple cytokines involved in promoting growth have been
implicated in regulation of the HSC pool; however, the potential roles of these negative
regulators are not very clear. Such negative regulators of self-renewal can work by accelerating
differentiation of stem and progenitor cells (Dybedal et al., 2001), and abrogating self-renewal
and expansion of stem cells (Yonemura et al., 1996).
1.3.1.2 TNF signaling as a negative regulator of hematopoiesis
Our results in Chapter 2 describe deregulation in TNFα mediated cellular survival in acute
promyelocytic leukemia. TNFα is a pleiotropic cytokine that exerts inhibitory and stimulatory
effects on a variety of cell types. TNF receptor (TNFR) and Fas ligand-mediated signaling have
been thought to suppress hematopoiesis both in vitro and in vivo. In a recent study, TNFα was
shown to be a potent endogenous suppressor of normal HSC activity in vivo in a mouse model
(Pronk et al., 2011). TNF signals through 2 receptors – TNFR-p55, and TNFR-p75 (Aggarwal,
2003). While the p55 form contains an intracellular death domain, and is therefore associated
with apoptotic signals, the p75 form is implicated in the control of proliferation (Aggarwal,
2003). TNF receptors are activated by 2 isoforms of TNF, TNFα and TNFβ. Knock out studies
have demonstrated that mice lacking TNFRSF 1 and 2 have normal counts of steady state HSCs.
However, both receptors were important in restricting HSC expansion, as HSCs from double
knockout donor mice had a significantly increased competitive advantage in reconstituting the
hematopoietic system in the bone marrow of recipient mice. Clonal growth of bone marrow with
wild type TNFRs was inhibited in the presence of TNFα. TNFα mediated growth suppression
was not rescued by over-expression of BCL2. This indicated that suppression of growth by
13
TNFα does not involve induction of cell death via inhibition of BCL2, and suggests that
induction of apoptosis has little role in TNFα induced growth inhibition. Pronk et al. (2011) also
showed that cell cycle activation enhanced TNFα induced suppression of HSC activity. TNFα
treatment of adult wild-type steady state mice resulted in decreased bone marrow cellularity and
reduced HSC activity as assessed in a bone marrow transplant assay. These effects of TNFα on
bone marrow cellularity and HSC activity were enhanced in mice treated with 5-fluorouracil, a
compound that is known to deplete actively cycling cells, and as a result induce active
proliferation of dormant HSCs. These studies indicate that TNFα primarily exerts negative
regulatory effects on actively cycling but not quiescent HSCs.
TNFα was previously reported to also exert negative effects on the committed progenitor cell
population. (Bryder et al., 2001) used purified, long term repopulating bone marrow stem cells
from mice and demonstrated that TNFα treatment severely compromised the short- and long-
term muti-lineage reconstitution of the hematopoietic system. In addition to previous work that
shows TNFα has effects on more differentiated clonogenic progenitor cells (Jacobsen et al.,
1994), this work demonstrated that TNFα mediated suppression can also target HSCs.
Interestingly, they also demonstrated that the negative effects on self-renewal were limited to
HSCs that were actively cycling. Quiescent HSCs have low TNFR expression and do not
respond to TNFα suppression (Bryder et al., 2001).
These studies suggest that TNFα plays an important regulatory role in normal hematopoiesis by
affecting both long term and short term repopulating progenitor and stem cells. TNFα may also
play an important role in malignant hematopoiesis. Chapter 2 of this thesis describes how while
TNFα exerts negative effects on growth and survival of wild type cells, leukemic cells
expressing oncogenic fusion proteins have evolved mechanisms to evade this normal negative
regulation. Mutations and deregulation of these genes involved in positive or negative
modulation of stem cell function and maintaining properties of self-renewal can lead to
deregulated growth of normal and malignant cells and contribute to disease development.
14
1.3.2 Summary
Taken together, normal hematopoiesis is critical for the maintenance of adequate levels of blood
cell elements required for survival. This regulation of HSC proliferation and self-renewal, as
well as the tight control of hematopoietic cell lineages, is a complex process involving bone
marrow microenvironment signals, as well as cell intrinsic regulators. Deregulation of any
regulatory component within this complex web can result in aberrant hematopoiesis and in some
cases, malignant transformation or other bone marrow disorders. Disruption in the normal
homeostatic balance between mature differentiated cells and HSCs/progenitors can occur due to
deregulated proliferation of progenitors or other mature cells, or the acquisition of self-renewal
properties by cells that normally do not self-renew (Passegue and Weisman, 2005).
15
1.4 Acute Promyelocytic Leukemia (APL)
Acute promyelocytic leukemia (APL; AML FAB M3) accounts for ~10% of all AML cases
worldwide. It is characterized by accumulation of abnormal hematopoietic cells with
promyelocytic features in the bone marrow, as well as balanced chromosomal translocations
involving the retinoic acid receptor alpha (RARA) locus on chromosome 17q21 (Beck, 1991;
Grignani et al., 1994; Kalantry et al., 1997; Warrell et al., 1993) Normal hematopoiesis is
inhibited, leading to pancytopenia, and diminished immunity. APL was first described by a
Swedish author, Hillestad (Hillestad, 1957), in a report of three patients characterized by a fatal
condition with white blood cells dominated by promyelocytes and a severe bleeding tendency. In
1985, all-trans retinoic acid (ATRA) was introduced as a treatment option in APL, and since then
has been extremely successful in treating APL (Wang and Chen, 2008). ATRA-based treatment
regimens combining ATRA and chemotherapy brought rates of complete remission up to 90%-
95%, and 6-year disease free survival up to 86% (+/- 10%) in low-risk patients (Asou et al.,
2007). Since the early 1990s the introduction of arsenic trioxide (ATO) further improved clinical
outcome of refractory and relapsed patients, and also newly diagnosed cases of APL (Powell et
al., 2010).
1.4.1 APL molecular pathology
More than 99% of APL cases involve a translocation t(15;17)(q22;q21) between the
promyelocytic leukemia (PML) and RARA genes (Alcalay et al., 1991; Goddard et al., 1991;
Pandolfi et al., 1991). Other “variant” partner genes collectively called “X”, have been
characterized: PLZF (Chen et al., 1993a); NPM (Hummel et al., 1999; Redner et al., 1996);
NuMA (Wells et al., 1997; Wells et al., 1996); STAT5b (Arnould et al., 1999); PRKAR1A
(Catalano et al., 2007); FIP1L1 (Kondo et al., 2008), (Menezes et al., 2011); BCOR (Yamamoto
et al., 2010). These translocations result in the creation of a functional chimeric protein (X-
RARA), whose N-terminus is derived from the partner gene, and retains the X protein’s
oligomerization domain; and whose C-terminus is derived from RARA (Melnick and Licht,
1999) (Figure 1.5). X-RARA, therefore, can be viewed as aberrant RARA transcription factors,
containing heterologous oligomerization and protein-protein interaction domains fused to an N-
terminal truncated RARA.
16
While the vast majority of the APL literature has focused on understanding the mechanism of
action of PML-RARA, and the clinically resistant variant PLZF-RARA, functional studies using
STAT5b-RARA, NPM-RARA, NuMA-RARA, and the other recently identified fusion proteins
are lacking, specifically on the role of the “X” partner protein in disease pathogenesis. While the
clinical manifestation of the disease induced by all the RA responsive APL fusion proteins are
virtually identical, the biology of the fusions are quite distinct, as a result of distinct fusion
partner proteins. In studying and understanding these variants in relation to PML-RARA, we are
in a better position to identify those minimally required molecular events and cellular pathways
that give rise to the common disease.
17
Figure 1.5: Structural diagram representing the APL fusion proteins. The N-terminal region of X-RARA is
comprised of domains B-F of RARA and is a common feature in all fusions identified to date. The C-terminal
domains is comprised of various partners and almost always contains a protein dimerization motif that enables the
fusion to form homo-oligomers in cells. The three most recent fusions reported between 2008 and 2010 include
FIP1L1-RARA, PRKAR1A-RARA, and BCOR-RARA. Our lab cloned and characterized the NuMA-RARA fusion,
and reported a case of NPM-RARA variant APL.
18
1.4.1.1 The leukemia initiating cell in APL
Analysis and purification of leukemia initiating cells in APL has been challenging, as a result of
the poor transplantibility of human APL cells as xenografts (Bonnet and Dick, 1997; Kogan,
2009). Studies have suggested that the APL LIC may be present within the committed myeloid
progenitor population, rather than a stem cell compartment. The true nature of the APL LIC,
whether a distinct cell population or cells at a particular stage of the cell cycle, or those localized
to particular bone marrow microenvironments, is still an open question. (Nasr et al., 2008)
studied the APL LIC at a functional level, to report that the combination of ATRA and ATO
resulted in clearance of the LIC. The mechanism of clearance, whether mediated by a loss of
self-renewal or induction of apoptosis is still unclear, and is one question that remains difficult to
address until such time as when the LIC cell population can be fully characterized. This work
also indicates that retinoids can mediate self-renewal of normal HSCs in addition to their well
known functions for promoting myeloid differentiation. The newly uncovered role for retinoid
signaling can potentially explain how deregulation of this signaling pathway by RARA fusion
proteins further contributes to the APL phenotype.
1.4.2 Retinoid signaling in Hematopoiesis
All of the APL associated fusion proteins involve the retinoic acid receptor (RAR), implying that
RAR mediated signaling and its effects on the cell is an important step in the pathogenesis of the
disease. Two classes of nuclear receptors, retinoid (RAR) and rexinoid (RXR), mediate retinoid
signaling in cells. Both RAR and RXRs are encoded by three genes, giving rise to closely related
isoforms α, β, and γ (Mangelsdorf and Evans, 1995). Like other nuclear receptors, RARs consist
of six evolutionarily conserved domains A-F, including the DNA binding domain that mediates
binding to retinoic acid response elements (RARE) (Chambon, 1996). The A/B (AF1) domain is
a ligand-independent, promoter context-dependent transactivation domain (Nagpal et al., 1993).
The C domain contains the DNA binding domain, and the E domain contains the all trans
retinoic acid (ATRA) binding site, the ligand-dependent transactivation domain (AF2), the
retinoid X receptor alpha (RXRA) dimerization interface and the nuclear co-repressor and co-
activator binding sites (Nagpal et al., 1993) (Figure 1.6). RAREs are located in the promoter
elements of retinoid target genes and consist of direct repeats (DRs) (A/G)G(G/T)TCA,
19
separated by 2 or 5 nucleotides (Naar et al., 1991; Umesono et al., 1991). RAR/RXR
heterodimerization is mediated by the DNA binding and ligand binding domains of RARs and is
required for binding to RAREs (Zechel et al., 1994) (Figure 1.6).
20
Retinoid X receptors (RXRs) were identified as co-regulators, required for the efficient binding
of RARs to their response elements (Hallenbeck et al., 1992; Leid et al., 1992; Yu et al., 1991).
Structurally RXRs are very similar to RARs and retain most of the domains found in RAR,
except for the F domain (Figure 1.7). RXRs can affect multiple biological pathways because of
their unique ability to heterodimerize with different nuclear receptors, including the thyroid
hormone receptor (TR), peroxisome proliferator-activated receptor (PPAR), and vitamin D
receptor (VDR) (Berrodin et al., 1992; Bugge et al., 1992; Kliewer et al., 1992). The three RXR
isoforms (α, β, and γ) have a high degree of homology, suggesting that they have common target
sequences, and respond to common ligands (Mangelsdorf et al., 1992) The isoforms differ in
their expression patterns: RXR alpha and RXR beta can be found in almost every tissue type,
while RXR gamma is restricted to muscle and brain cells (Mangelsdorf et al., 1992).
Retinoid signaling plays an important role in the development and differentiation of a number of
tissues in the body, including the eye, the heart and circulatory systems, the central nervous
system, the urogenital and respiratory systems, the musculoskeletal system and hematopoiesis
(Mark et al., 1999); (Ross et al., 2000). Retinoid signaling has been implicated in hematopoiesis,
specifically terminal neutrophil differentiation, through a number of different lines of
investigation (Table 1.2; Table 1.3). Additional means of retinoid-mediated control of
hematopoiesis include control of retinoic acid metabolism, and non-ligand-mediated crosstalk
among hematopoietic signaling pathways. Numerous studies involving vitamin-A deficient
animal systems and in vitro culture models clearly establish a role for retinoids in controlling
signaling pathways involved in hematopoiesis (Oren et al., 2003). Active retinoid signaling starts
with cellular enzymes and RA binding proteins, which mediate the processing of dietary retinol
(vitamin A) to retinoic acid (Oren et al., 2003). The different rates of retinol metabolism in
hematopoietic cells constitute a means of regulating cellular levels of retinoic acid. This ensures
that RARA transcriptional activity is differentially regulated in hematopoietic cells in spite of
their exposure to uniform physiological concentrations of retinoids (1-10 nM) in the serum
(Collins, 2002).
21
Table 1.2: Evidence for the role of RXR in myelopoeisis
Evidence
Study
In HL-60 cells, activation of RXR signaling was shown to be required for the induction of apoptosis, and a similar link was reported in the NB4 cell line.
Mehta et al., 1996; Shiohara et al., 1999;
Nagy et al., 1995; Benoit et al., 2001
Evidence of crosstalk between the RXR and protein kinase A (PKA) signaling pathways, as activation of both RXR and PKA, using specific ligands and agonists, was found to induce cell maturation in NB4 cells
Benoit et al., 1999
Conditional knockout of RXRA in the hematopoietic compartment. Strikingly, these mice did not exhibit an abnormal hematopoietic phenotype in vivo, suggesting that RXRA is dispensable for normal adult hematopoiesis.
Ricote et al., 2006
Down-regulation of RXRA is essential for terminal neutrophil differentiation, as RXRA is highly expressed in granulocyte/monocytes progenitors, and in terminally differentiated monocytes, but not in mature granulocytes. Furthermore, the ectopic over-expression of RXRA in GM progenitors resulted in a block of neutrophil differentiation, while the expression of a dominant negative form of RXRA, lacking amino acids 1-197, did not impair neutrophil differentiation.
Taschner et al., 2007
22
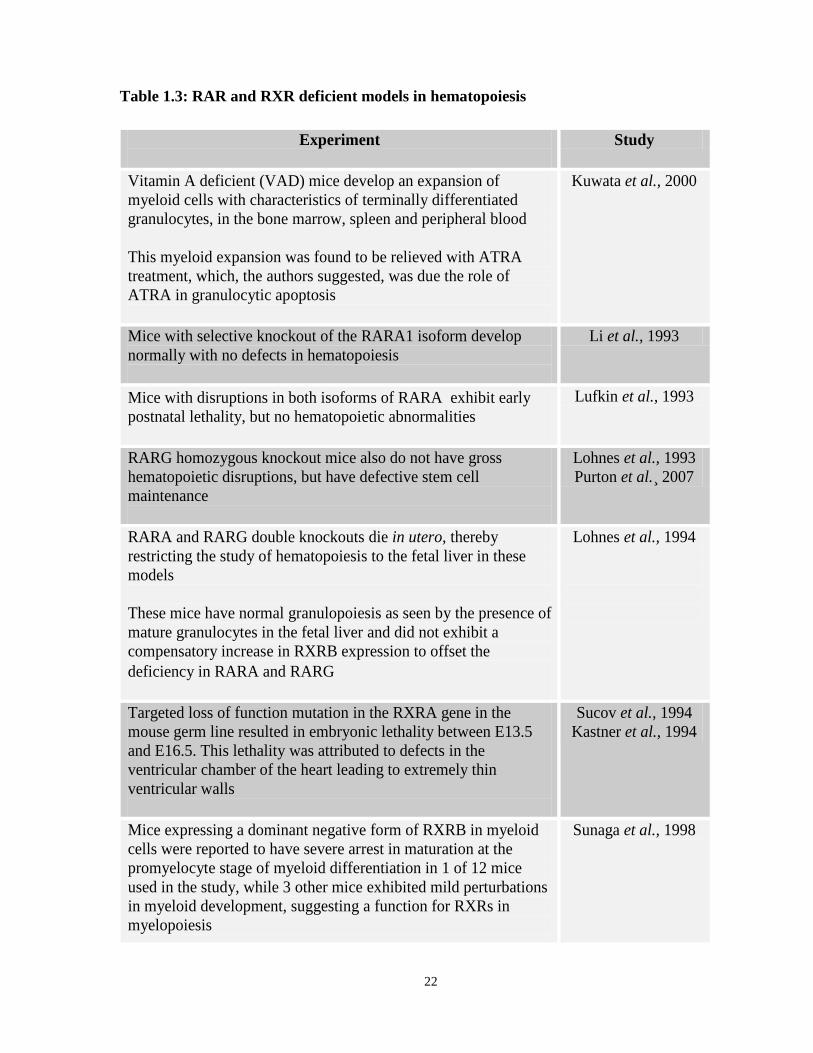
Table 1.3: RAR and RXR deficient models in hematopoiesis
Experiment
Study
Vitamin A deficient (VAD) mice develop an expansion of myeloid cells with characteristics of terminally differentiated granulocytes, in the bone marrow, spleen and peripheral blood This myeloid expansion was found to be relieved with ATRA treatment, which, the authors suggested, was due the role of ATRA in granulocytic apoptosis
Kuwata et al., 2000
Mice with selective knockout of the RARA1 isoform develop normally with no defects in hematopoiesis
Li et al., 1993
Mice with disruptions in both isoforms of RARA exhibit early postnatal lethality, but no hematopoietic abnormalities
Lufkin et al., 1993
RARG homozygous knockout mice also do not have gross hematopoietic disruptions, but have defective stem cell maintenance
Lohnes et al., 1993 Purton et al.¸ 2007
RARA and RARG double knockouts die in utero, thereby restricting the study of hematopoiesis to the fetal liver in these models These mice have normal granulopoiesis as seen by the presence of mature granulocytes in the fetal liver and did not exhibit a compensatory increase in RXRB expression to offset the deficiency in RARA and RARG
Lohnes et al., 1994
Targeted loss of function mutation in the RXRA gene in the mouse germ line resulted in embryonic lethality between E13.5 and E16.5. This lethality was attributed to defects in the ventricular chamber of the heart leading to extremely thin ventricular walls
Sucov et al., 1994 Kastner et al., 1994
Mice expressing a dominant negative form of RXRB in myeloid cells were reported to have severe arrest in maturation at the promyelocyte stage of myeloid differentiation in 1 of 12 mice used in the study, while 3 other mice exhibited mild perturbations in myeloid development, suggesting a function for RXRs in myelopoiesis
Sunaga et al., 1998
23
AF-1 Transcriptional Activation
AF-1 Transcriptional Activation
DNA Binding
Hinge Region
Ligand Binding
AF-2 AD AF2 Activation Domain
Figure 1.7: Structural features of Retinoid X Receptor (RXR). RXR structural organization is
similar to that of other nuclear hormone receptors, but lacks C-terminal F domain. AF-1,
In the absence of the RA ligand, the transcriptional activity of the RAR/RXR heterodimer is
inhibited by the binding of co-repressors, including nuclear receptor co-repressor (N-CoR),
silencing mediator for retinoid and thyroid receptors (SMRT) and histone deacetylase (HDAC)-
containing Sin3A complexes (Hu and Lazar, 2000). Binding of all-trans retinoic acid (ATRA) to
RARA creates a conformational transition in the RARA ligand binding domain (Renaud et al.,
1995). This disrupts the co-repressor interaction and promotes the sequential recruitment of co-
activator complexes, which remodel chromatin to facilitate binding of the transcriptional
machinery on the promoter. Co-activators like p300/CBP, locally modify chromatin structure
through their histone acetyl transferase (HAT) activity, which acetylates lysine residues on the
N-terminal tails of histones and weakens their interaction with DNA, allowing for the activation
of gene transcription (Wade et al., 1997).
Non-ligand mediated activation of RARs has also been recorded during different stages of
myelopoiesis (Collins, 2002). These include interactions of RARs with transcription factors like
the STAT family members and their synergism with other receptors such as the protein kinase A
(PKA) receptor (Collins, 2002). A number of hematopoietic cytokines including IL-3, GM-CSF,
and IL-1, have been reported to enhance transcriptional activity of RA receptors (Nakamaki et
al., 1994). IL-3 and GM-CSF mediate their cellular effects by activation of the JAK/STAT
pathway. Activated cytokine receptors phosphorylate STATs by their association with JAKs.
STATs then translocate to the nucleus, where they act as transcription factors (Darnell, 1997).
There exists significant functional crosstalk between RARs and STAT receptors, as seen by the
direct role of STAT5 (Si and Collins, 2002) in mediating IL-3 induced enhancement of RAR
activity (Johnson et al., 2002). Furthermore, a number of overlapping STAT/RAR binding sites
have been reported in the RAREs of different genes and illustrate the role of non-ligand
mediated activation of RAR transcriptional activities (Si and Collins, 2002).
25
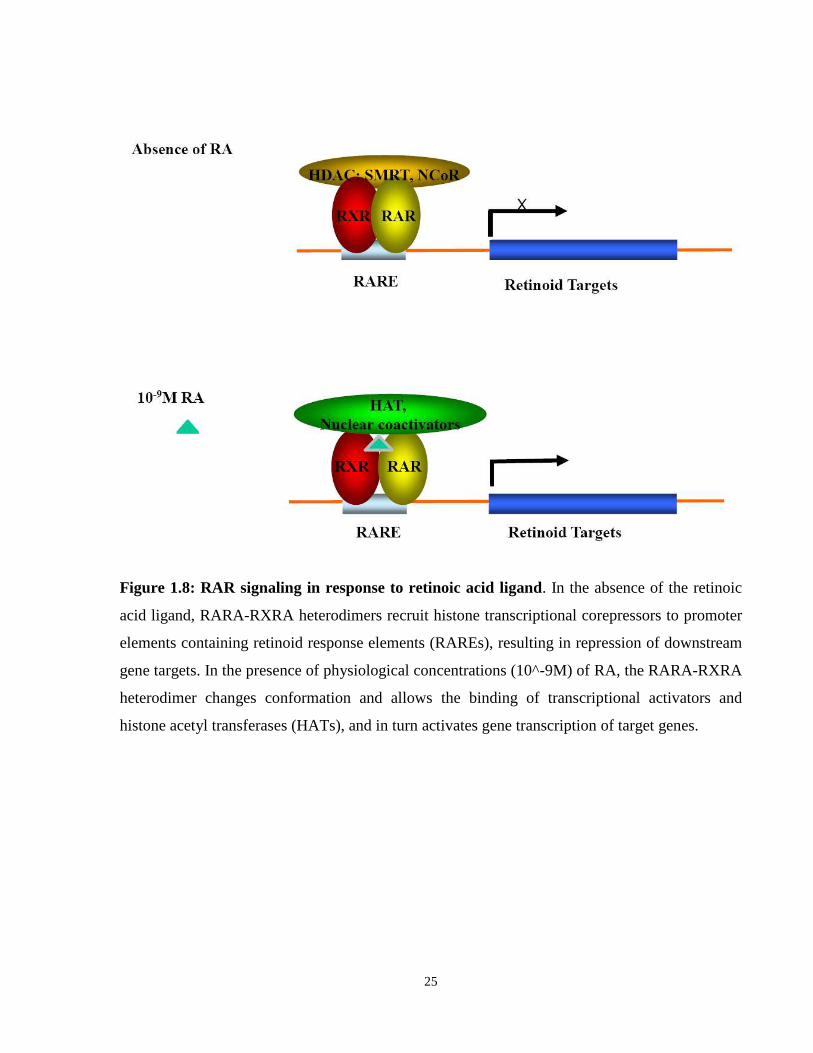
Figure 1.8: RAR signaling in response to retinoic acid ligand. In the absence of the retinoic
acid ligand, RARA-RXRA heterodimers recruit histone transcriptional corepressors to promoter
elements containing retinoid response elements (RAREs), resulting in repression of downstream
gene targets. In the presence of physiological concentrations (10^-9M) of RA, the RARA-RXRA
heterodimer changes conformation and allows the binding of transcriptional activators and
histone acetyl transferases (HATs), and in turn activates gene transcription of target genes.
26
1.4.2.1 The Role of RXRA in Hematopoiesis
While the role of RARA in hematopoiesis is well characterized, the role of RXRs in myeloid cell
differentiation is not well understood beyond their function as obligatory heterodimerization
partners for RARs (Szanto et al., 2004). This is partially because of the complexity of the
system; different blood cell lineages express different nuclear receptors that heterodimerize with
RXRA, thus making the phenotype of RXRA deficient hematopoiesis difficult to analyze. The
other major problem is one of construction of the experimental system: The RXRA -/- mouse is
embryonic lethal at ED17.5 due to myocardial defects (Sucov et al., 1994) This is attributed to
the critical roles RXRA plays throughout the rest of the organism, though it renders it impossible
to study the role of RXRA in adult hematopoiesis. Several studies have identified a role for
RXRA in hematopoiesis (Benoit et al., 1999; Benoit et al., 2001a; Benoit et al., 2001b; Mehta et
al., 1996; Ricote et al., 2006; Shiohara et al., 1999; Taschner et al., 2007). Some studies, in
particular, are intriguing: Conditional knockout of RXRA in the hematopoietic compartment
(Ricote et al., 2006) did not lead to an abnormal hematopoietic phenotype in vivo, suggesting that
RXRA is not critical for normal adult hematopoiesis. However, down-regulation of RXRA is
essential for terminal neutrophil differentiation, as RXRA is highly expressed in
granulocyte/monocytes progenitors, and in terminally differentiated monocytes, but not in
mature granulocytes (Taschner et al., 2007). Furthermore, the ectopic over-expression of RXRA
in GM progenitors resulted in blocked neutrophil differentiation, while the expression of a
dominant negative form of RXRA did not impair neutrophil differentiation.
A number of studies have characterized the phenotype of RXRA mutant mice to elucidate the
role of rexinoid signaling during development. These studies have reported the use of both
complete knockouts as well as tissue specific, temporally regulated RXRA disruption in the
mice. Tissue specific functional knockout of RXRA was created to study RXRA effects
specifically in the ventricular chamber of the heart (Chen et al., 1998), in epidermal and hair
follicle keratinocytes (Li et al., 2001; Li et al., 2000) in thymocytes and T-lymphocytes
(Stephensen et al., 2007) and in hepatocytes (Imai et al., 2001). The mouse models used in these
studies all employ the cre-loxP strategy to generate tissue specific knockouts. We used this
system to selectively knock out RXRA in the hematopoietic compartment, using mice with exon
4 of RXRA flanked with loxP sites, also expressing cre under the Flk-1 promoter (Sukhai et al.,
2008). (Kastner et al., 1994) reported the use of RXRA null mice with a disruption in exon 4. A
27
similar report of a complete RXRA knockout using mice that lacked part of exon 3 (encoding
part of the DNA-binding domain) was also reported by (Sucov et al., 1994). Others have looked
at the role of other domains of the protein including the AF-1 (Mascrez et al., 2001), and AF-2
(Mascrez et al., 1998) using transgenic mice lacking these domains of RXRA. Compound
knockout mice exhibit a wider array of abnormalities, and are often embryonic lethal.
1.4.2.2 Retinoid signaling in APL and X-RARA effects on transcription
For a number of years, the prevailing hypothesis was that forced homo-dimerization of RARA
was responsible for the development of APL. PML-RARA is associated with nuclear complexes
that are much greater in apparent molecular weight than PML-RARA alone (Nervi et al., 1992).
Wild-type PML and RXRA were identified as some of the proteins associated with these
complexes (Nervi et al., 1992) It has been hypothesized that APL fusion proteins aberrantly
repress gene transcription and hence de-regulate genes important in myeloid differentiation,
resulting in the observed block in maturation (Melnick and Licht, 1999) (Figure 1.9). In many
cases, the differentiation block can be overcome by treatment with pharmacological doses of
ATRA (>10-7 M) (Melnick and Licht, 1999). The APL fusion proteins have altered DNA binding
properties (Chang et al., 1992) and can bind RAREs as homodimers (Perez et al., 1993) while
wild type RARA does not (Leid et al., 1992). The impaired ability of APL fusion proteins to
activate certain promoters used to be explained by their increased ability to bind corepressors
SMRT and N-CoR, thus requiring pharmacological doses of ATRA for dissociation. PML-
RARA acts as an aberrant transcriptional repressor on RA target genes by having a stronger
association with NCoR/SMRT. The mechanism responsible for the more stable recruitment of
NCoR/SMRT by PML-RARA (as opposed to wild-type RARA) is based on the presenceof a
strong self-association domain (Lin and Evans, 2000; Minucci et al., 2000). Oligomerization of
RARA through the coiled coil domain of PML is responsible for abnormal recruitment of
NCoR/SMRT (Lin and Evans, 2000; Minucci et al., 2000). In contrast with wild-type RARs,
which bind only one NCoR/SMRT molecule, it is thought that PML-RARA oligomers can
associate with multiple NCoR/SMRT complexes simultaneously (Lin and Evans, 2000; Minucci
et al., 2000). This results in increased local concentration of the HDAC complex on target gene
promoters, leading to enhanced transcriptional repression in the presence of physiological
concentrations of RA. Recent evidence also suggests that PML-RARA and PLZF-RARA can
recognize and bind to other splice variants of SMRT and NCoR that are not recognized by wild
type RARA (Mengeling et al., 2011). These data suggest that the acquired ability to interact with
28
alternative splice variants of NCoR and SMRT contributes to the oncogenicity of the APL fusion
proteins. The presence of RXRA in heterotetrameric complexes of APL fusions favored co-
repressor recruitment and binding, in stark contrast to wild type RARA-RXRA heterodimers
(Mengeling et al., 2011). In addition to release of co-repressors and stimulation of genes
responsible for myeloid differentiation, ATRA also acts to degrade the PML-RARA protein and
upregulate wild-type RARs to restore retinoid signaling.
29
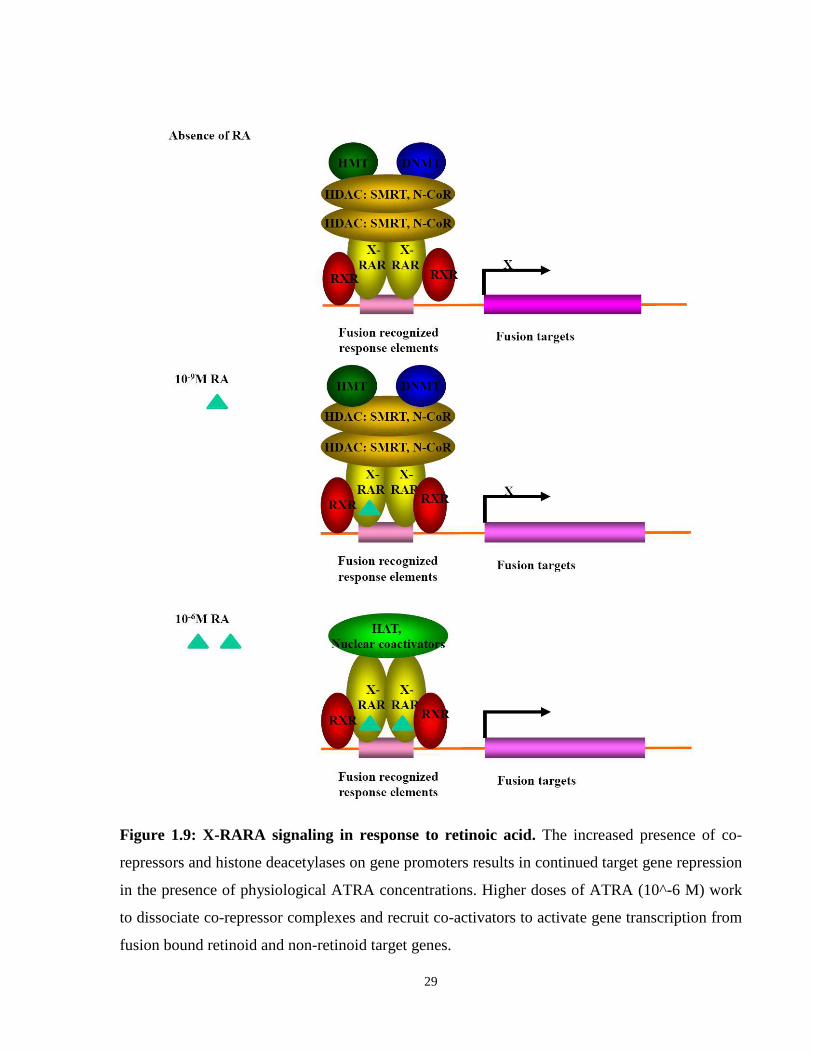
Figure 1.9: X-RARA signaling in response to retinoic acid. The increased presence of co-
repressors and histone deacetylases on gene promoters results in continued target gene repression
in the presence of physiological ATRA concentrations. Higher doses of ATRA (10^-6 M) work
to dissociate co-repressor complexes and recruit co-activators to activate gene transcription from
fusion bound retinoid and non-retinoid target genes.
30
Much like PML-RARA, PLZF-RARA interacts with SMRT and N-CoR, resulting in
transcriptional repression at RAREs. Interestingly, PLZF-RARA may form repressor complexes
resistant to pharmacological concentrations of ATRA (Grignani et al., 1998; Guidez et al., 1998).
PLZF itself has the capacity to bind SMRT through its POZ domain, located in its N-terminus
and retained in the fusion protein (Hong et al., 1997); thus, PLZF-RARA homodimers can
potentially interact with four SMRT complexes (Grignani et al., 1998; Guidez et al., 1998; Hong
et al., 1997), two of which, by interacting with the N-terminal POZ domain of PLZF, cannot be
removed on ATRA treatment. Both PML-RARA and PLZF-RARA have an affinity for RA
comparable to that of wild-type RARA (Benedetti et al., 1997; Dong et al., 1996) and can bind to
RAREs as homodimers or multimeric complexes containing RXRA (Dong et al., 1996; Perez et
al., 1993). Strikingly, a cell line model expressing PLZF-RARA (the tetracycline-repressible
U937-PLZF-RARA line (Rice et al., 2009), is at least partially sensitive to ATRA treatment, an
observation that runs counter to the above model.
More recent evidence has uncovered additional details of transcriptional repression mediated by
X-RARA, which broaden the potential impact that the fusion has in the cell. PML-RARA binds a
wider range of DNA response elements in the genome than the retinoic acid response element
(RARE) (Kamashev et al., 2004). A number of these elements are required by other nuclear
hormone receptors in order to bind DNA. While RARA, as a heterodimer with RXRA, will only
bind DR2 and DR5 elements, PML-RARA has been shown to bind a range of direct repeat (DR),
everted repeat (ER) and inverted repeat (IR) sequences in vitro (Jansen et al., 1995; Kamashev et
al., 2004; Perez et al., 1993). Our group demonstrated this relaxed DNA binding specificity for
all X-RARA, specifically for the DR1 PPRE, as well as for NPM-RARA with DR3 and DR4
elements (Kamel-Reid et al., 2003); Hamadanizadeh SA, Kamel-Reid S, unpublished).
PML-RARA requires RXRA in its transcriptional complex (Kamashev et al., 2004), suggesting
that the fusion does not merely sequester RXRA away from its sites of action within the cell, but
instead forms a functional complex with it in order to have a more direct effect at the gene
expression level. PLZF-RARA/RXRA heterodimers bind to RAREs with higher affinity than
PLZF-RARA homodimers in vitro (Dong et al., 1996; Licht et al., 1996). Furthermore, PLZF-
RARA/RXRA heterodimers have the capacity to bind to non-consensus RAREs, thus suggesting
that these heterodimers contribute to APL pathogenesis through the regulation of novel gene
expression (Hauksdottir and Privalsky, 2001; So et al., 2000). STAT5b-RARA also requires
31
RXRA for strong association with HDAC complexes and transcriptional repression (Zeisig et al.,
2007). Our work further demonstrated that functional loss of RXRA resulted in an amelioration
of the NuMA-RARA-mediated leukemic phenotype in vivo (Sukhai et al., 2008). Similar results
were published for PML-RARA as well (Zhu et al., 2007).
NPM-RARA and NuMA-RARA can form heterodimeric complexes with RXRA; these
complexes are capable of binding to and repressing transcription from the RARE (Kamel-Reid et
al., 2003); Hamadanizadeh SA, Kamel-Reid S, unpublished). Like PML- and PLZF-RARA,
NPM-RARA expression enhances primitive marrow progenitor cell proliferation, while ATRA
treatment of NPM-RARA-expressing cells induces differentiation and inhibits cell growth (Du et
al., 1999).
1.4.2.3 Rexinoid signaling in APL pathogenesis
Recent studies have shed light on an increasingly important role for RXRA in APL pathogenesis.
X-RARA has the capability of forming homodimers, as reported for PML-RARA (Jansen et al.,
1995; Perez et al., 1993), PLZF-RARA (Grignani et al., 1998; Guidez et al., 1998) NPM-RARA
(Lin and Evans, 2000) and NuMA-RARA [(Dong et al., 2003); and our own work, unpublished].
In a previous study (Hummel et al., 2002), our lab showed that X-RARA can form heterodimers
with RXRA, as well as the wild-type X protein. Until very recently, this interaction was not
thought to be important in leukemogenesis. Recent reports, however, when taken together, allow
us to view the role of RXRA in APL, and more broadly, AML, from a novel perspective.
All APL fusions retain the RARA/RXRA dimerization interface found in wild-type RARA. A
physical interaction between X-RARA and RXRA is therefore expected, and indeed, observed,
for all fusions studied (Dong et al., 2004; Perez et al., 1993; Sukhai et al., 2008; Yamamoto et
al., 2010). For example, PML-RARA is associated with nuclear complexes that are much greater
in apparent molecular weight than PML-RARA alone (Nervi et al., 1992). Wild-type PML and
RXRA were identified as some of the proteins associated with these complexes (Perez et al.,
1993). Very recent analysis of genome wide binding sites of PML-RARA confirms the presence
of RXRA in 99% of PML-RARA binding sites (Martens et al., 2010). Furthermore, PLZF-
RARA can also bind to RAREs as homodimers or multimeric complexes containing RXRA
(Dong et al., 1997). Our previous studies demonstrated physical interaction between NPM- and
NuMA-RARA, and RXRA (Hummel et al., 2002). The latter finding was independently
corroborated in separate studies (Dong et al., 2003; Dong et al., 2004). This physical interaction
32
between X-RARA and RXRA was preserved after ATRA treatment, as one can follow the
mobilization of the X-RARA/RXRA complex within the cell after treatment with
pharmacological concentrations of RA (Dong et al., 2004). Studies have strongly implicated
RXRA as having a critical functional role in PML-RARA mediated transcriptional repression.
(Zeisig et al., 2007), and (Zhu et al., 2007) show that disrupting the binding between RXRA and
X-RARA by introducing mutations in RXR binding sites of PML-RARA, impairs the
development of leukemia in transgenic mice, while still being able to cause transformation in
vitro. Silencing RXRA using shRNA mediated knock-down completely abrogates the in vitro
transforming potential of the PML-RARA. These studies indicate that RXR plays an important
role in mediating PML-RARA mediated transformation.
1.4.2.4 Therapeutic potential of rexinoids
The basis of differentiation therapy is to force cells along the normal differentiation pathway
through the restoration/reactivation of signal transduction pathways that are otherwise
suppressed during tumour development. Guiding cells through the differentiation lineage will
eventually result in post-differentiation induced cell death. The APL fusion protein complex
consists of higher order hetero-oligomers with RXRA as described above. In addition to the
classically targeted RARA moiety, studies by two groups (Zeisig et al., 2007; Zhu et al., 2007)
have implicated RXRA as a potential therapeutic target. In APL blasts, RA can trigger a death
signaling cascade through the activation of IFN regulated factor-1, which is recruited to the
promoter elements of TRAIL, which along with Death Receptor 5 (DR5) and DR4, selectively
targets and kills tumour cells (Clarke et al., 2004). As a result of their lower toxicity profile
compared to retinoids, rexinoids are preferred as drug candidates (Altucci et al., 2007). A
characteristic feature of RXRs is the inability of RXR agonists to transactivate RAR-RXR
signaling when used as a single agent. This process referred to as “RXR subordination” only
allows RXR agonists to enhance the retinoid response initiated by an RAR agonist, and not to be
able to transactivate signaling from RAR-RXR heterodimers as a single agent (de Lera et al.,
2007). Altucci et al., demonstrated that corepressor complexes can be dissociated from APL
heterodimers when PKA is activated, thereby allowing rexinoids to induce transactivation of
gene expression through coactivator recruitment (Altucci et al., 2005). Benoit and colleagues
also showed that raising intracellular cAMP levels allows RXR ligands to induce differentiation
in APL cells that have developed RA resistance (Benoit et al., 1999). Despite RXR
subordination, rexinoids can act through different mechanisms to activate RXR signaling. In the
33
presence of increased cAMP levels, rexinoid signaling can induce differentiation and post
maturation cell death (Benoit et al., 1999). This is the case even in ATRA resistant AML cells,
suggesting that this mechanism of rexinoid activation is acting independently of retinoid
signaling. RXR agonists can however activate other signaling pathways, as RXR is known to
heterodimerize with other proteins including VDR and PPARγ (de Lera et al., 2007). Rexinoids
induce cell death in AML cells under reduced serum and growth factor conditions (Benoit et al.,
2001a). This signaling was shown to be mediated through rexinoid activation of permissive
RXR-PPARγ heterodimers (Indra et al., 2007; Shankaranarayanan et al., 2009). These studies
indicate that targeting rexinoid mediated pathways can result in the activation of one of two
cellular pathways: cellular differentiation, or cell death.
1.4.2.5 Targeting the oncogenic PML-RARA in APL therapy
Induction of cellular differentiation was classically thought to be the basis by which ATRA
therapy worked effectively in patients presenting with APL, as RA induces rapid differentiation
of primary blasts into terminally differentiated granulocytes (Breitman et al., 1981). Recent
observations have called for a re-evaluation of the importance of differentiation in mediating
treatment efficacy in APL. Arsenic trioxide (ATO) is a potent anti-leukemia therapy with
profound effects on APL cell clearance, even when administered as a single agent (Mathews et
al.; Mathews et al., 2006; Zhu et al., 2002). It does little to affect gene expression of PML-
RARA target genes (Shao et al., 1998; Wiese et al., 2001), but is very effective in the treatment
of relapsed APL cases. The oncogenic fusion PML-RARA exerts self-renewal and growth
properties on the leukemic cell, in addition to inducing a block in differentiation. In patients, RA
treatment alone is not effective in inducing complete remission unless combined with
chemotherapy (Warrell et al., 1993). Even though RA induces complete differentiation in most
cases of APL, only a small percentage of patients respond durably with RA when used as a
single agent (Hu et al., 1999). This suggests that the differentiation process alone does not induce
stable depletion of APL cells. In vivo, much higher concentrations of ATRA are required for
clearing APL cells, compared to smaller amounts used for inducing transcriptional activation
(Ablain and de The, 2011). Some patients only respond to liposomal ATRA formulations that
considerably increase RA levels in the blood (Tsimberidou et al., 2006). Also studies in mouse
models have shown that mutations in PML-RARA’s phosphorylation site S873, which modulates
its degradation, impaired disease remission, while differentiation induction by RA treatment
remained unaffected (Nasr et al., 2008) In the case of the RA resistant fusion PLZF-RARA, the
34
resistance was thought to result from a stronger repression of target genes by the fusion
(Grignani et al., 1998; He et al., 1998; Lin et al., 1998). Recent evidence indicates that PLZF-
RARA cells can fully differentiate upon RA treatment [(Nasr et al., 2008; Rice et al., 2009) and
our own unpublished observations]. These data suggest that the block in differentiation alone
may not be driving leukemogenesis and that only reversing this inhibition may be insufficient for
APL therapy.
In addition to blocked differentiation, PML-RARA was also shown to boost the self-renewal
properties of APL blasts (Welch et al., 2011). Although phenotypically they resemble committed
progenitors, mouse APL leukemic blasts have increased self-renewal, thereby indicating that
PML-RARA confers self-renewal properties to more differentiated progenitor cells. LICs have to
be targeted for tumour eradication, as their persistence after conventional therapy strongly
contributes to disease recurrence. APL is described as an oncogene-derived disease, where one
can expect that degradation of the oncogene would be sufficient to eradicate the disease. Both
retinoic acid and arsenic trioxide degrade PML-RARA. They are synergistic in their effects in
mouse models, as would be expected from the fact that they target the oncoprotein through two
different mechanisms. Loss of the fusion protein may also elicit spontaneous differentiation
(Ablain and de The, 2011). These observations indicate that differentiation block is not the result
of irreversible chromatin modifications induced by the APL fusions, and can be relieved by
removing the presence of the fusion.
1.4.3 Functions of PML and PML-RARA
The t(15;17) reciprocal balanced chromosomal translocation, characteristic of the vast majority
of APL cases, produces PML-RARA and RARA-PML fusion proteins. PML-RARA is the main
oncogenic APL fusion with the ability to transform hematopoietic progenitors (de The and Chen,
2010; Piazza et al., 2001). While PML-RARA mediates transcriptional repression by suppressing
retinoid signaling as discussed previously and blocks differentiation, it is also known to disrupt
PML nuclear bodies (PML-NBs) through PML-RARA interaction with wild type PML (de The
and Chen, 2010; Piazza et al., 2001). The PML moiety in PML-RARA is thought to function by
promoting the formation of fusion multimers which contribute to transformation (Minucci et al.,
2000). However, it is also known that the loss of PML expression increases the penetrance and
latency of leukemia in APL mouse models, indicating that PML mediated functional pathways
play a role in disease pathogenesis. PML expression is also lost in some solid tumours including
35
prostate adenocarcinomas, colon adenocarcinomas, breast carcinomas and lung carcinomas
(Gurrieri et al., 2004), as well as in other hematological malignancies, including 83% of diffuse
large cell lymphomas (DLCL) and 77% of follicular lymphomas (Gurrieri et al., 2004).
We will review the contribution of wild type PML, as well as PML-RARA, to cellular
transformation through deregulating pathways involved in cellular survival, apoptosis, and self-
renewal.
1.4.3.1 PML functions in cellular growth and apoptosis.
PML interacts with a wide range of protein targets, however the physiological role of many of
these interactions have not be firmly established in vivo. In the following sections, a brief
overview of PML’s biological role that implicates it in oncogenesis will be surveyed. PML’s role
as a tumour suppressor works through multiple mechanisms including the control of key factors
involved in modulating the cell death response, regulation of protein synthesis, maintenance of
genome stability, and modulating cell cycle regulation.
PML has been thought to be involved in regulation of growth suppressive signals. Studies have
implicated PML in maintaining cellular senescence and programmed cell death. PML was shown
to mediate apoptosis induced by FAS ligand (FASL) and TNFα (Guardiola-Serrano et al., 2010).
Lymphocytes lacking PML have decreased cell death in response to FasL treatment (Wang et al.,
1998). PML is also known to potentiate cell death through interferon alpha, by inducing
production of TRAIL in cancer cells (Ashkenazi, 2008; Falschlehner et al., 2009; Gurrieri et al.,
2004; Schneider-Jakob et al., 2010) .
PML’s interaction with pro-apoptotic transcription factors also plays a role in regulating cell
death. PML regulates p53 tumour suppressor degradation (through inhibition of Mdm2, which is
the E3 ubiquitin ligase for p53) (Bernardi et al., 2004; Kurki et al., 2003). PML also promotes
p53 post translational modifications including p53 acetylation (Pearson et al., 2000) and
phosphorylation (Hofmann and Will, 2003). PML regulates the PI3K pathway at multiple levels
(Ito et al., 2009) (Song et al., 2008) (Trotman et al., 2007), as well as the TGFβ pathway (Lin et
al., 2004), both of which have pro-neoplastic effects.
Studies have implicated the regulation of transcription as a key step in tumourigenesis (Ruggero
and Pandolfi, 2003). PML can interact with the eukaryotic initiation factor (eIF4E), and inhibits
36
its function in mRNA export. This affects expression of cell cycle regulators targets such as
cyclin D1, and further results in decreased proliferative capacity (Cohen et al., 2001).
1.4.3.2 PML-RARA and self-renewal
X-RARA fusion proteins block terminal differentiation and increase the self-renewal capability
of X-RARA expressing cells (Puccetti and Ruthardt, 2004). The activation of Wnt signaling is
one mechanism by which X-RARA may increase self-renewal (Muller-Tidow et al., 2004).
Gamma and beta-catenin are increased transcriptionally and this allows for Wnt signaling
activation by X-RARA. (Steinert et al., 2011) show that derivatives of the drug Sundilac down-
regulated key components of the Wnt signaling network in APL cells through down-regulation
of beta-catenin and gamma-catenin. Sundilac is known to be used as a type of non-steroidal anti-
inflammatory agent, which inhibits Wnt signaling in tumour models (Boon et al., 2004).
(Welch et al., 2011) developed a mouse model expressing PML-RARA under the control of the
endogenous mouse PML promoter, specifically in the myeloid compartment. They utilized this
model to investigate the effects of PML-RARA on hematopoiesis after the acquisition of the
oncogenic fusion protein as a somatic event in these mice. They reported that PML-RARA
increased hematopoietic self-renewal as they observed that bone marrow CFU’s from these mice
could be replated for 6 or more weeks. Also of note was the progressive accumulation of cells
expressing PML-RARA in the bone marrow, without evidence of myeloproliferation. This
suggested that PML-RARA alters self-renewal in hematopoietic precursors without altering the
bone marrow cellular feedback mechanisms that regulate the size of the myeloid compartment.
Their work also showed that PML-RARA cannot reprogram late myeloid cells to self-renew, but
instead that deregulation of self-renewal is induced in early hematopoietic cells. Taken together,
(Welch et al., 2011) demonstrated that PML-RARA affects multipotent progenitor populations,
rather than the more committed myeloid progenitors, which were thought to be the cell of origin
according to the established paradigm (Bonnet and Dick, 1997; Guibal et al., 2009; Turhan et al.,
1995; Wojiski et al., 2009). It has been suggested that the maturation defect in APL is a
cooperating event during development, or that mutations in genes specifically expressed in the
myeloid compartment contributed to the differentiation block. This is in contrast to other work
from (Wojiski et al., 2009) which used an older model of PML-RARA expressed under the
mouse cathepsin G promoter, where PML-RARA was able to confer self-renewal properties to
committed progenitors or leukemic promyelocytes. The discrepancies in the two studies most
37
likely stem from differences inherent in the two mouse models. These studies support a role for
PML-RARA in promoting self-renewal and show this to be an important step in the pathogenesis
of APL.
1.4.3.3 PML-RARA and apoptosis
(Tao et al., 2011) showed that the wild type PML binds to Fas, which is a potent death receptor.
Disabling mutations in Fas have been reported in a minority of cancers. Deregulated expression
of mediators of Fas death signaling has also been described in various cancers including lung and
colon (Pitti et al., 1998). Defective Fas signaling has been implicated in resistance to therapy, as
intact signals are required for effective function of many genotoxic therapies including radiation
(Muller et al., 1998). PML-RARA was found to suppress Fas-mediated signaling and apoptosis.
Both PML and PML-RARA were identified to directly interact with Fas in APL cell lines and
primary blasts (Tao et al., 2011). PML-RARA after binding to Fas interferes with Fas mediated
apoptosis by recruiting cFLIP and forming an inhibitory complex. cFLIP recruitment blocks the
initiation of Fas mediated apoptotic signaling by inhibiting the binding and activation of pro-
caspases to the complex. PML-RARA also is known to block both p53- dependent and p53
independent pathways of cell death (Wang et al., 1998) and is thought to be mediated through
PML-RARA effects on transcriptional repression (Guo et al., 2000). We investigated PML-
RARA and the variant fusions ability to interfere with the TNFα and NF-κB mediated survival
signaling in an effort to understand the role of the fusions in promoting cellular survival (Chapter
2).
1.4.4 Functions of NPM and NPM-RARA
1.4.4.1 NPM: Structure and expression
Nucleophosmin (NPM, B23, NO38, numatrin) is an abundant nucleolar phosphoprotein, ~37
kDa in size present in the granular region of the nucleolus (Kang et al., 1975). Two isoforms of
NPM have been reported – a longer 294-amino-acid variant, prevalent in all tissues and localized
to the nucleolus (Chan et al., 1989); and a shorter 259-amino-acid isoform, differing at the C-
terminus, present in both nucleoplasm and cytoplasm (Colombo et al., 2006; Dalenc et al., 2002).
Several functional domains have been identified in NPM: An N-terminal hydrophobic segment
(involved in oligomerization and chaperone activities); and two acidic sections (essential for
histone binding). The central portion between the acidic segments and the C-terminal region are
38
involved in nucleic-acid binding and ribonuclease activity (Hingorani et al., 2000). Two
tryptophan residues (288 and 290) are necessary for NPM nucleolar localization (Nishimura et
al., 2002). The NPMc cytoplasmic mutant, found in ~40% of normal karyotype AML, carries a
frameshift mutation in exon 12 that disrupts this nucleolar localization signal (Falini et al.,
2005).
NPM also contains nuclear localization (Hingorani et al., 2000) (NLS; Hingorani 2000) and
nuclear export signals (NES; (Wang et al., 2005). The wide range of protein interacting domains
within NPM enables it to bind a number of partners in different compartments within the cell,
including transcription factors (IRF1, NF-κB), nucleolar proteins (nucleolin, fibrillarin), histones
(H3, H4), proteins associated with proliferation (DNA polymerase-α), and mitosis (NuMA,
NEK2A). NPM undergoes dynamic changes over the course of the cell cycle. NPM expression
peaks at S or G2 phases of the cell cycle and is at minimal levels in G0 (Feuerstein et al., 1988a;
Feuerstein et al., 1988b; Feuerstein et al., 1988c). This might be related to the fact that NPM
specifically stimulates the activity of DNA polymerase α (Takemura et al., 1994) or may simply
be a reflection of the metabolic demand of the cell. During G2 and M phase, NPM is heavily
phosphorylated by CDC2 (Peter et al., 1990). The fact that NPM is intimately involved in events
taking place at the G2/M regulatory point also underlines the strong relation between NPM and
cellular proliferation.
NPM is thought to have roles in cellular transformation, growth and proliferation. NPM is
commonly over-expressed in a variety of tumours including gastric, colon, ovarian and prostate
carcinomas (Bernard et al., 2003; Nozawa et al., 1996; Skaar et al., 1998; Subong et al., 1999;
Tsui et al., 2004), and is involved in chromosomal translocations, or deleted in various tumours
and hematological malignancies including APL (Redner et al., 1996), Anaplastic Large Cell
Lymphoma (ALCL) (Morris et al., 1994) Myelodysplastic syndrome (MDS) (Yoneda-Kato et
al., 1996) and AML (Mendes-da-Silva et al., 2000; Olney et al., 2002). Close to 40% of acute
myeloid leukemia patients also harbour mutations in NPM, making it one of the most frequently
mutated genes in AML (Falini et al., 2005).
1.4.4.2 NPM in hematological malignancies
Unlike the other RARA partner genes, NPM is also fused to genes other than RARA in
hematologic malignancies, as in the t(2;5)(p23;q35) translocation found in Ki-1+ anaplastic large
39
cell lymphoma (Morris et al., 1994). Here, NPM is linked to ALK, a gene encoding a membrane
spanning tyrosine kinase (Morris et al., 1994; Nakamura et al., 1997) which is normally not
expressed in lymphoid tissue. (Downing et al., 1995; Morris et al., 1994; Nakamura et al., 1997).
As a result, the ubiquitously expressed NPM drives the expression of an aberrant fusion tyrosine
kinase (Bischof et al., 1997; Shiota et al., 1994). Moreover, the NPM portion of NPM-ALK has
been shown to be essential for its role in oncogenesis (Bischof et al., 1997). Cases of
myelodysplastic syndrome and a subset of AML have also been shown to be associated with a
t(3;5)(q25;q35) translocation that fuses NPM to myelodysplasia/myeloid leukemia factor (MLF-
1) (Yoneda-Kato et al., 1996), although little is known about this fusion protein.
NPM mutations have also been found to be associated with ~40% of AMLs with normal
karyotype. These mutations lie within exon 12 of NPM, thus introducing a nuclear export signal
and rendering the protein cytoplasmic (NPMc). Mutational status of NPM is now an important
diagnostic and prognostic marker in AML with a normal karyotype, along with FLT3 status.
NPM mutations in AML occur in an age-dependent fashion and are stable over disease evolution
(Chou et al., 2006). NPM mutation status can therefore be used as a marker of disease remission.
The copy number of mutant NPM predicted relapse in AML It has emerged as an important
prognostic indicator in NPM mutant AML. Patients achieving greatest reduction in mutant NPM
copy number were associated with better outcome (Gorello et al., 2006). NPMc mutations are
infrequent in chronic myeloid disorders, but were found in 3 CMML patients progressing to
AML (Caudill et al., 2006). Furthermore, haploinsufficiency of NPM (e.g., through
chromosomal rearrangement) may play a role in some AML and in MDS (Berger et al., 2006).
NPMc+ AML also bears a distinct gene expression signature, separate from other AMLs,
characterized by upregulation of genes involved in regulation of the stem cell compartment
(Alcalay et al., 2005).
1.4.4.3 Oncogenic roles of NPM
NPM over-expression results in increased growth and proliferation. Normal NPM expression
levels are tightly correlated with proliferation. Highly proliferative cells show increased
expression of NPM compared to quiescent cells (Dergunova et al., 2002). NPM’s role in
proliferation is through various mechanisms including the following: NPM is a target gene of the
proto-oncogene Myc (Boon et al., 2001; Zeller et al., 2001); NPM is correlated with stimulation
of DNA polymerase activity (Takemura et al., 1999); also, cells deficient in NPM1 exhibit
40
impaired proliferation and induction of apoptosis when the nuclear-cytoplasmic shuttling of
NPM is inhibited (Brady et al., 2004; Grisendi and Pandolfi, 2005)
NPM expression is increased in proliferating and malignant cells (Borer et al., 1989), (Chan et
al., 1989) including leukemic blasts. This may influence protein synthesis through NPM’s roles
in ribosome biogenesis, although this is not well-defined within the literature. Our data indicates
that increased NPM protein levels are associated with increased ribosomal RNA levels in APL.
Interestingly, under hypoxic conditions, HIF1α is an activator of NPM expression (Li et al.,
2004) . This may be due to an increased requirement for ribosomal precursors (Derenzini et al.,
1995; Kondo et al., 1997). Down-regulation of NPM delayed entry into mitosis (Jiang and Yung,
1999) suggesting that NPM may have an active role in growth control. In support of this concept,
NPM binds to the tumor suppressor IRF-1, inhibiting its anti-proliferative effects (Kondo et al.,
1997). Finally, NPM is implicated in initiating the DNA damage response, as it binds to
damaged chromatin during initiation of this process (Lee et al., 2005). Interestingly, Npm-/- mice
are embryonic lethal (ED11.5), due to defects in hematopoiesis and organ development. Npm-/-
cells have significant mitotic and centrosomal defects, chromosome copy number alterations, and
cannot properly respond to genotoxic signals. Npm+/- mice are viable at birth, but develop a
myelodysplastic syndrome (Grisendi et al., 2005). Thus, there may be a strong requirement for
NPM in proper cellular function, and in hematopoietic development. Over-expression of NPM
enhances the proliferative potential of hematopoietic stem cells (HSC), and promotes self-
renewal of long-term repopulating HSCs. Over-expression of NPM also promotes survival after
DNA damage, oxidative stress and hematopoietic injury (Li et al., 2006a).
1.4.4.4 NPM as a tumour suppressor
Translocations involving NPM1 or mutations in NPM are commonly found in leukemias, and
lead to the loss of one functional copy of NPM. Phenotyping of mice with NPM knockouts has
indicated that NPM is required for genome stability. NPM interaction with ARF is essential for
its contribution to growth suppressive effects. NPM associates with Arf in the nucleolus. ARF
inhibits MDM2, resulting in p53 activation and therefore suppression of cell proliferation.
Therefore loss of NPM can also promote tumourigenesis.
Increased expression of NPM leads to accumulation of ARF by preventing its turnover and
increasing its stability. Arf is responsible for triggering cell cycle arrest and apoptotic programs
41
in response to oncogenic stimuli. ARF also negatively regulates ribosomal RNA processing.
ARF inhibits MDM2, which is a negative regulator of p53, by relocalizing it to the nucleolus
(Weber et al., 1999). ARF mutants that are unable to associate with NPM have decreased protein
stability and function (Kuo et al., 2004). Loss of NPM function can promote tumourigenesis
through destabilizing and inactivating ARF, and as a result deregulating cell proliferation
through p53-dependent and independent mechanisms (Sherr, 2001). It has been proposed that
when NPM is over-expressed, it sequesters ARF in the nucleolus (Korgaonkar et al., 2005). It
remains unknown if ARF bound to NPM can interact with MDM2. If this is possible then it has
been suggested that NPM’s ability to stabilize ARF would be tumour suppressive.
NPM and ARF regulation has also been studied in NPMc+ AML. All NPM1 alleles that are
mutated encode proteins with an added nuclear export signal motif (Nakagawa et al., 2005), and
lose at least one of the 2 tryptophan residues at positions 288 and 290 – these are thought to be
important for nucleolar localization of NPM (Falini et al., 2005). NPMc+ also can interact with
ARF and results in its cytoplasmic relocalization. This reduces SUMOylation of MDM2, and
reduces ARF dependent p53 activation. Also, wild type NPM is also delocalized to the
cytoplasm, which affects its ability to inhibit ARF turnover. Cells with NPMc+ mutation may be
characterized by inactivation of the ARF tumour suppressor pathway. Analysing its effects on
the cells and function of mutant NPM, it is clear that this gene belongs to the class of proteins
with both tumour suppressive and oncogenic functions.
1.4.4.5 NPM in ribosome biogenesis
Ribosome biogenesis is an integral part of cellular growth. Approximately half of the cellular
energy reserve is directed towards ribosome biogenesis (Moss, 2004). Since the early 1960s the
nucleolus was recognized as the site for ribosomal DNA transcription and ribosome biogenesis
(Birnstiel et al., 1963), (Ritossa and Spiegelman, 1965). The nucleolus is comprised of three
distinct regions based on ultrastructural morphology: fibrillar centers, dense fibrillar
compartment, and granular zone. Transcription of rDNA occurs in the regions between the
fibrillar centers and dense fibrillar components, while the transcribed rRNA is processed in the
dense fibrillar component. The granular regions further process the rRNA and assemble them
into specific ribosomal subunits (Hernandez-Verdun and Roussel, 2003). NPM has been
implicated in the processing and assembly of ribosomes. The specific localization of the long
isoform of NPM (B23.1) to the granular region of the nucleolus correlates with its function in
42
transcribing ribosomal DNA (Murano et al., 2008), and processing pre-rRNA (Savkur and
Olson, 1998). Decreased NPM expression has been shown to inhibit processing of pre-rRNA.
Increased NPM expression can disrupt ribosomal rRNA synthesis by leading to sustained
ribosome activation and increased protein synthesis, both of which are found in cancer cells
(Roussel and Hernandez-Verdun, 1994). Cancer cells have been reported to have enlarged
nucleoli, which lends support to the fact that sustained ribosome activity supports aberrant cell
growth.
The ARF tumour suppressor is another protein that localizes to the nucleolus, where it integrates
growth signals to suppress cellular growth and proliferation. ARF also binds and affects the
function of NPM (Bertwistle et al., 2004; Brady et al., 2004; Itahana et al., 2003) . Arf knock out
mice have significant changes in nucleolar morphology, and displayed increased ribosomal
biogenesis (Apicelli et al., 2008). Interestingly, this increase in ribosome biogenesis that was
associated with the loss of Arf was found to be reversible upon removal of NPM. This indicates
that increased amounts of free cellular NPM (not tethered to Arf) can promote ribosome
biogenesis. This study demonstrates that the NPM-ARF interaction is functionally important in
controlling ribosome biogenesis and protein synthesis rates within cells.
1.4.4.6 NPM in survival and apoptosis.
Over-expression of NPM promotes cell survival through inhibition of pro-apoptotic pathways –
IRF1 DNA binding is inhibited; NPM binds and inhibits function of eukaryotic initiation factor 2
kinase PKR; DNA fragmentation activity of caspase activated DNAse (CAD) in neural cells is
inhibited; and, p53 activation is inhibited by interaction in response to apoptotic stimuli. All of
this works to suppress the pro-apoptotic machinery within the cells.
Over-expression of NPM in tumor cells is also associated with increased cell growth and
proliferation, and inhibition of differentiation and apoptosis (Grisendi et al., 2005). Knocking
down NPM expression induces cell death (Itahana et al., 2003; Ye, 2005). NPM is up-regulated
in normal cells upon exposure to different types of apoptotic stimuli, leading to a survival
response (Grisendi et al., 2005). Under conditions of cellular stress, NPM translocates from the
nucleolus to the cytoplasm and/or nucleoplasm. Stimuli capable of inducing this response
include cytotoxic drugs and intercalating agents such as daunorubicin (Milligan 2006), inhibitors
of RNA polymerase I and II, and UV irradiation (Bor et al., 1992; Chan, 1992; Chan et al., 1987;
Kurki et al., 2004; Yung et al., 1990). NPM translocation may impair apoptosis by inhibiting
43
caspases and nucleases or by regulating tumor suppressors such as ARF and p53 (Ye, 2005),
(Korgaonkar et al., 2005). NPM mediates the anti-apoptotic activity of nerve growth factor
(NGF) by acting as a receptor for nuclear phosphatidylinositol (3, 4, 5)-triphosphate (PIP3) in
NGF-treated PC12 cells. The nuclear NPM-PIP3 complex inhibits the DNA fragmentation
activity of caspase-activated DNAse (CAD) (Ahn et al., 2005; Grisendi et al., 2006), and
contributes to the survival activity of NGF. Overall, deregulated NPM may promote
tumourigenesis (Ahn et al., 2005), partially by its anti-apoptotic functions.
NPM functions have also been linked to cell proliferation in response to mitogenic stimuli and
cell cycle progression. Indirectly, NPM can function as anti-apoptotic protein – malignant cells
with NPM over-expression can have suppressed activity of IRF1, which is a transcription factor
and tumour suppressor, involved in DNA damage induced apoptosis (Kondo et al., 1997). NPM
was also found to suppress apoptosis through the inhibition of the double stranded RNA
dependant protein kinase (PKR), which is a factor that induces apoptosis in response to various
stimuli (Pang et al., 2003). Hematopoietic cells with NPM over-expression can exhibit defective
PKR activity as over-expressed NPM directly interacts with it, which can result in abnormal
proliferation and survival.
44
1.4.4.7 NPM-RARA in APL
The t(5;17) chromosomal translocation results in the formation of the NPM-RARA
fusion protein. Phenotypically, NPM-RARA expressing blasts are very similar to the
more commonly occurring PML-RARA expressing APL. These leukemic blasts are also
sensitive to differentiation induced by ATRA. One case of NPM-RARA associated APL
which showed evidence of disease relapse also responded favourably multiple times with
ATO based therapy (Chen et al., 2010).
Alternative splicing results in the creation of two isoforms of the NPM-RARA fusion
protein: NPMs-RARA containing 110 amino acids of NPM; and NPMl-RARA
containing an additional 43 amino acids (Nicci et al., 2005; Redner et al., 2000). Redner
et al. first described this fusion in APL (Redner et al., 1996); our group has since then
reported another case of this fusion, followed by other studies. Subsequent case reports
describing NPM-RARA expressing APL, including our report only describe the presence
of the shorter NPM-RARA transcript (Grimwade et al., 2000; Hummel et al., 1999; Xu et
al., 2001).(Redner et al., 2000) demonstrate that both forms of NPM-RARA interact with
RAREs as homodimers or heterodimers with RXRA. Similar to PML-RARA, NPM-
RARA also binds corepressors and coactivators that respond to retinoic acid. Studies
from our lab have shown that NPM-RARA, in addition to repressing wild type retinoid
signaling can also mediate its effects in APL by interfering with other signaling
pathways. One such pathway is the peroxisome proliferator-activated receptor gamma
(PPARγ) signaling pathway. The ability of NPM-RARA to alter PPARγ signaling was
tested using the PPARγ specific agonist Troglitazone. While control cells exhibited no
death response to Troglitazone, the presence of NPM-RARA sensitized the pro-
monocytic U937 line and resulted in inhibition of cell growth and induction of apoptosis
(Kamel-Reid et al., 2003). Furthermore, NPM-RARA was also shown to bind the PPRE
as a heterodimer with RXRA, and thereby block normal PPARγ signaling induced by
Troglitazone.
45
1.4.5 Functions of NuMA and NuMA-RARA
1.4.5.1 NuMA structure, and roles in the cell
NuMA is a large (~240 kDa), nuclear phosphoprotein first observed in a human/hamster
somatic cell hybrid where it strongly associated with the mitotic spindle (Lydersen and
Pettijohn, 1980). Using the changes in the subcellular localization patterns of NuMA
during various stages of the cell cycle, several groups have identified its diverse role
within cells. In interphase cells NuMA is localized in diffuse and speckled nuclear
patterns (Kallajoki et al., 1991; Compton et al., 1992) and is associated with the nuclear
matrix (Zeng et al., 1994). During metaphase, NuMA accumulates in a crescent-shaped
pattern that parallels that of the spindle poles (Tang et al., 1994) and then is released in
late telophase for transportation to the nuclei of daughter cells (Compton et al., 1992; Hsu
and Yeh, 1996; Kallajoki et al., 1991; Wiese et al., 2001). Drosophila Mud is an
orthologue of NuMA and Lin-5 (Siller et al., 2006), and coordinates spindle orientation
and promotes asymmetric cell division. Mud is required for neuroblasts to align with the
polarity axis during development; loss of Mud leads to symmetric cell division, mis-
specification of cell fate, and tumor-like proliferation of cells (Bowman et al., 2006). It is
reasonable to hypothesize that NuMA may likewise play a role in asymmetric cell
division during development and differentiation in the mouse and human. Such a role
offers intriguing possibilities in terms of the ability of the NuMA-RARA fusion protein
to block differentiation during neutrophil development, and to foster self-renewal in a
leukemia initiating cell. Interestingly, NuMA is differentially overexpressed in leukemic
cells and is significantly associated with complex karyotypic abnormalities (Ota et al.,
2003). Overexpression of NuMA in mouse myeloid cell cultures leads to aneuploidy, cell
cycle arrest and apoptosis (Ota et al., 2003).
1.4.5.2 NuMA-RARA in APL
Like most patients that have PML-RARA positive APL, the 6 month old infant in whom
NuMA-RARA was first identified also responded fully to treatment with ATRA (Wells et
al., 1997) . NuMA-RARA is therefore another ATRA sensitive variant of APL. Since the
identification and characterization of the t(11;17)(q13;q21) NuMA-RARA fusion, our
46
laboratory has described a transgenic mouse model (hCG-NuMA-RARA) of APL. Mice
carrying the NuMA-RARA transgene developed a myeloproliferative disease like
myeloid leukemia with promyelocytic features, with a phenotype similar to the original
patient (Sukhai et al., 2004). The disease development and latency is inversely correlated
with NuMA-RARA fusion copy number in mice (Sukhai et al., 2011).
X-RARA have the capability of forming homodimers, as reported for PML-
RARA (Jansen et al., 1995); (Perez et al., 1993), PLZF-RARA (Grignani et al., 1998);
(Guidez et al., 1998), NPM-RARA (Lin and Evans, 2000), and NuMA-RARA (Dong et
al., 2003); and our own work, unpublished), X-RARA also have the ability to form
heterodimers with RXRA, and the wild-type X protein (Hummel et al., 2002). In order to
further examine the role of RXRA in NuMA-RARA pathogenesis, we created a
functional knockout of RXRA within the transgenic NuMA-RARA mouse model. The
resulting mice expressed the oncogenic fusion protein and a functionally inert version of
RXRA that lacked the DNA binding domain. Upon characterizing this model, we
demonstrated that functional loss of RXRA resulted in an amelioration of the NuMA-
RARA-mediated leukemic phenotype in vivo (Sukhai et al., 2008). This also suggested
that the RXR molecule is an important part of the NuMA-RARA complex on the DNA
and is required for the development of leukemia.
In addition to impairing retinoid/rexinoid signaling in APL, other work has also described
NuMA-RARA and its effects in other cellular signaling networks. The effect of NuMA-
RARA on the transcriptional activity of the oncogene STAT3 has been elucidated (Dong
and Tweardy, 2002). Similar to PML-RARA, PLZF-RARA and STAT5b-RARA,
NuMA-RARA has the ability to enhance the transcriptional activity of STAT3. These
results are the first indication that APL fusion proteins may disrupt additional signaling
pathways within the cell that are not directly related to RAR or RXR signaling.
47
1.5 Secondary genetic events in APL
Several transgenic mouse models of APL have been developed over the years, and
extensive work in this area has been done to identify the in vivo role of various fusion
proteins in APL biology. Transgenic mice expressing NPM-RARA (Cheng et al., 1999),
(Rego et al., 2006), PML-RARA (Grisolano et al., 1997), (Welch et al., 2011), PLZF-
RARA (He et al., 1998), (Cheng et al., 1999), and NuMA-RARA (Sukhai et al., 2011;
Sukhai et al., 2008; Sukhai et al., 2004) have been generated to express the specific
fusion protein in the myeloid/promyelocytic compartment of mouse bone marrow.
Various improvements to the APL model have been made, the most recent one being the
introduction of PML-RARA knocked-in to the mouse PML promoter in order to more
closely resemble the background of the human APL cell, and mimic the acquisition of the
fusion as a somatic event leading to the transformation of the cells. A common feature of
all models thus far is the long latency to leukemia development, thus strongly suggesting
the presence of secondary genetic events that cooperate with the fusion to give rise to
leukemia. Analysis of human APL patient blasts, as well as leukemic mice to identify
additional mutations and gene/pathway deregulation has been traditionally used in order
to identify relevant secondary events. More recent efforts have also focused on high
throughput gene expression arrays and whole genome sequencing to determine
deregulated or mutated genes and pathways that can contribute to disease development.
The following sections describe these candidate gene and whole genome based
approaches and their findings.
1.5.1 Candidate gene alterations as approaches to elucidate secondary genetic events in APL
Analysis of transgenic and knock-in models of APL expressing PML-RARA show
leukemia development only after a long latency, indicating that while PML-RARA is
necessary, it is not sufficient to induce a leukemia (Pollock et al., 2001), (Grisolano et al.,
1997; Westervelt et al., 2003). Candidate gene studies have targeted genes and pathways
for analysis as potential cooperating events, based either on their appearance in APL
patient cells (e.g. FLT3, K-ras), or their characteristic as biologically relevant pathways
48
in transformation and leukemogenesis e.g. BCL2 and PIM2 (apoptosis), and DNMT3A1
(epigenetic modifiers).
Mutations that lead to the constitutive activation of K-Ras or FLT3 shorten the latency
and increase the penetrance of leukemia development in PML-RARA (Chan et al., 2006;
Sohal et al., 2003). These mutations are also found in APL patients (Callens et al., 2005;
Kiyoi et al., 1997), (Longo et al., 1993). Kelly et al. (2002) showed that introduction of
FLT3 ITD in bone marrow from PML-RARA mice resulted in a short latency disease like
APL that resembles that which the single transgenic PML-RARA mice develop after a
long latency. FLT3 is mutated in about 40% of human APL, and FLT3 ITD induces
myeloproliferative disease in a mouse model (Kelly et al., 2002). While not sufficient to
induce AML in mouse models, Kelly and colleagues demonstrated that FLT3-ITD
cooperates with PML-RARA to induce an APL-like disease. Chan et al. 2006 report the
cooperation between oncogenic K-Ras and PML-RARA in APL development in mice.
Oncogenic K-Ras mutations are observed in 4-10% of APL patients; in PML-RARA
transgenic mice, mutant K-Ras was thought to trigger proliferative and cell survival
signals that cooperate with the fusion in causing an APL-like phenotype with increased
penetrance and decreased latency.
In addition to these activating mutations, alterations in epigenetic modifications may also
result in enhancing PML-RARA induced leukemia. DNA methylation, histone
methylation, and histone acetylation have recently been associated with APL target genes
(Martens et al., 2010). Further, a recent study by Subramanyam et al., (2011) looked at
the contribution of epigenetic modifiers such as DNMT3, which also physically interact
with PML-RARA as part of a complex within the nucleus. PML-RARA recruits DNA
methyltransferases (DNMTs) to target promoters and this causes methylation of CpG
islands and repression of transcription in APL cells (Di Croce et al., 2002).
(Subramanyam et al., 2011) tested the ability of Dnmt3a1 to cooperate with the PML-
RARA fusion to induce APL. They found that over-expression of Dnmt3a1 shortened
survival of transgenic PML-RARA mice. Bone marrow transplant assays demonstrated
that mice recipient of bone marrow over-expressing PML-RARA and Dnmt3a1
developed leukemia with shorter latencies than PML-RARA donor mice. This indicated
49
that over-expression of Dnmt3a cooperates with PML-RARA in enhancing the
development of leukemia.
PIM2 belongs to a family of serine threonine kinases that are expressed in the
hematopoietic system (White, 2003). PIM2 functions to phosphorylate BAD and reverse
its apoptotic effects, and thereby has anti-apoptotic effects on the cell (Yan et al., 2003),
(Fox et al., 2003). PIM2 is a transcriptional target of FLT3-ITD, which is an alteration
that occurs in 30% of APL patients and induces transformation both in vitro and in vivo.
STAT5 activates transcription of PIM2 after signaling through FLT3-ITD (Mizuki et al.,
2003). While Pim2 overexpression alone does not result in hematological malignancies, it
was found to cooperate with other partially transforming FLT3 mutants to promote
transformation (Agrawal et al., 2008). PIM2 kinase is upregulated in AML blasts and
primary APL cells, in comparison to normal CD34+ cells. PML-RARA expressing bone
marrow cells infected with PIM2 retrovirus and transplanted into lethally irradiated
recipients, resulted in an APL like disease in ~64% of mice, while none of the control
transplanted mice showed evidence of disease. Cells from these primary recipients caused
a leukemia when harvested and introduced into secondary and tertiary transplant mice.
(Agrawal-Singh et al., 2011) also note that expansion of Pim2 PML-RARA cells did not
occur until a short period prior to the development of leukemia, suggesting that Pim2
over-expression does not induce a rapid polyclonal expansion of leukemic cells.
It is also known that the fusion confers increased resistance to apoptosis in myeloid cell
lines (Fu et al., 1995; Grignani et al., 1993), and enhances survival in primary myeloid
cells in response to pro-apoptotic stimuli (Wang et al., 1998). These observations led
groups to investigate candidate proteins in pathways controlling cellular apoptosis, as
drivers of leukemia and cooperating events in APL. Accordingly, BCL2, which functions
to inhibit caspases and thereby negatively regulate apoptosis was investigated in the
context of promoting leukemogenesis in the PML-RARA mouse model background.
Kogan and colleagues (2001) observed that simultaneous expression of Bcl2 and PML-
RARA lead to a marked accumulation of myeloid cells in mice, and accelerated the
development of acute leukemia in the PML-RARA mouse model.
50
1.5.2 Whole genome approaches to determining secondary events
Candidate gene based approaches to determine secondary genetic events are limited in
their comprehensiveness, and their ability to use an unbiased approach to identify
biologically relevant, and functionally important drivers of APL pathogenesis. Genetic
changes that accompany progression of leukemia can be characterized directly using
cytogenetics, and or DNA sequencing (Ding et al., 2012; Ley et al., 2010), or indirectly
by studying the gene expression profiles of leukemic cells (Golub et al., 1999). This
approach has been useful in distinguishing between different types of leukemia, and
developing markers of specific mutations, however it is more challenging to identify
mutations that contribute to disease initiation and progression, and leukemic cells are
usually detected post transformation (Walter et al., 2004). Mouse models offer a more
effective system whereby a single genetic event (e.g. chromosomal translocation) can be
engineered into cells, and followed to detect disease progression and the associated
changes. (Wartman et al., 2011) used a whole genome sequencing based approach to
investigate functionally relevant and recurrent somatic gene mutations that occur in the
mCtsg-PML-RARA knock in mouse model of APL. Their analysis identified a recurrent
point mutation in Jak1 (Jak1 V657F) and a deletion in histone demethylase (Kdm6a).
They had further shown that the Jak1 variant can cooperate with PML-RARA to induce a
rapidly fatal APL like disease in the mouse. Importantly, this mutation in Jak1 is identical
to the recently described mutation in the JAK1 pseudokinase domain (V658F) in human
APL and adult ALL (Flex et al., 2008), (Jeong et al., 2008), (Mullighan and Downing,
2009).
Whole genome transcript expression analysis is more commonly used to identify global
changes in gene expression that are associated with the APL fusion proteins. Cellular
pathways that are identified to be over-represented within gene sets deregulated by the
APL fusions, are thought to be involved in disease pathogenesis, and have the potential to
drive leukemia development. Array analysis of gene targets of PML-RARA and PLZF-
RARA was performed to compare the set of downstream deregulated targets of these two
fusions on gene expression (Park et al., 2003). In this analysis, deregulated expression of
a number of genes identified to be involved in tumor necrosis factor (TNF) α signaling
51
was observed. Another group reported the effect of ATRA on gene expression in ATRA-
sensitive and –resistant NB4 cells also involved induction of TNF-response genes in
response to retinoic acid treatment (Witcher et al., 2003; Witcher et al., 2004). A
combined analysis of PML-RARA, PLZF-RARA and AML1-ETO, also in U937 cells
indicated that pathways involved in stem cell maintenance and in the regulation of DNA
repair were commonly altered by these fusions (Casorelli et al., 2006). These
observations suggested that AML fusions induced a state in leukemic blasts that
promoted their self-renewal capabilities.
The WNT/β-Catenin signaling pathway was another commonly deregulated network of
genes identified from array analysis of the three AML fusions (Muller-Tidow et al.,
2004). Wnt signaling is involved in cellular proliferation, cell-to-cell signaling, and is
implicated in the self-renewal of stem cells, suggesting that AML fusions may induce
abnormal cellular proliferation in part through deregulated WNT signaling. AML-
associated translocation products (including PML-RARA) increase the expression of γ-
Catenin by activating its promoter region. Increased expression of γ-Catenin results in
increased replating efficiency of HSC (Zheng et al., 2004).
Array analysis was performed in the mCG-PML-RARA transgenic line after treatment
with external irradiation, in order to measure the accumulation of genetic changes after
DNA damage (Walter et al., 2004). Further analysis demonstrated a network of
deregulated myeloid transcription, postulated to contribute to leukemogenesis in mice
These studies demonstrated that individual PML-RARA+ mice had variable gene
expression profiles, suggesting that no single, unifying cooperating downstream gene
expression change may be required for leukemogenesis in these mice.
It still remains to be seen what genetic changes drive leukemogenesis after acquisition of
the chromosomal translocation. While studies to date are limited in their ability to detect
these driver changes, emerging sequencing technologies and improved mouse models can
be used to address these issues and better understand the pathogenesis of APL.
52
1.5.3 Use of high throughput assays to understand leukemia pathogenesis
Whole genome based analysis allows for comprehensive interrogation of the mechanisms
of disease development and disease biology. We have used whole genome transcriptome
profiling to characterize the effects of the variant fusion proteins on cellular gene
expression and transcriptional regulation, described in Chapter 3. Gene expression array
technology has been put to a number of uses in order to more fully elucidate AML
biology. Broadly, array analysis has been applied to the classification of AML subtypes;
diagnosis and prognosis of AML; development and understanding of AML therapies; and
elucidating the mechanisms of AML pathogenesis. Selected cases demonstrating the use
of array technology in each of these areas of AML biology are described in Table 1.4
below.
A number of factors determine the ability to classify samples, and discover novel
subtypes within a heterogeneous disease using gene expression profiling studies. Both the
size of the study group, as well as demographic variations influence expression results. In
addition, technical factors such as the use of different array platforms between studies,
sample preparation and array hybridization techniques play an integral part to the final
expression output. Further differences in bioinformatics approaches and filtering methods
between studies factor into the outcomes of the study. In most cases, these factors do not
drastically affect disease subtypes with a distinctive expression signature, but will likely
affect classification of sample sets containing more subtle expression changes.
In addition to subtype identification, gene expression profiling studies have also been
frequently used to determine prognostic signatures in AML using overall survival and
other outcome measures as the end point. Additionally, gene expression profiling studies
have proved useful in understanding the downstream genetic targets of oncogenes, or
identifying novel changes that are causative for disease development.
53
Table 1.4: A summary of selected microarray studies describing disease classification, prognosis, biology, and therapeutic response mechanisms in AML.
Study Type Major findings Sample types Study Reference
AML classification; AML prognostic groups
Identified molecular signature defining 16 AML patient groups and prognostic information
285 AML patients (Valk et al., 2004)
AML classification; AML prognostic groups
Identified prognostically relevant subgroups in AML with normal karyotype
119 AML patients (Bullinger et al., 2004)
AML classification; AML diagnosis
Predicted clinically relevant subgroups of leukemia with high accuracy.
892 AML, CLL, ALL samples
(Haferlach et al., 2005b)
AML classification; AML biology
Specific changes in miRNA expression levels can distinguish selected AML subtypes
52 AML patients (Li et al., 2008)
AML classification; AML outcome prediction
Identified miRNA expression associated with selected AML subtypes
122 AML patients (Garzon et al., 2008b)
AML classification Gene expression profiling accurately predicts AML subtypes characterized by expression of translocated transcription factor products.
461 AML patients (Verhaak et al., 2009)
Leukemia subtype prediction
Use of gene expression profiling to classify 18 distinct subcategories of leukemia with high accuracy
3334 ALL, AML, CLL, CML, MDS patients
(Haferlach et al., 2010)
AML biology Identified a miRNA signature that distinguishes NPMc+ AML from NPM1 non-mutated cases.
85 AML patients (Garzon et al., 2008a)
54
AML prognosis and outcome prediction
Identified novel groups of AML with differences in prognosis and response to therapy
170 AML patients (Wilson et al., 2006)
AML prognosis and outcome prediction
Identified an 86-probeset prognostic signature that correlates with overall survival in cytogenetically normal AML
143 CN- AML patients
(Metzeler et al., 2008)
AML stem cell biology Identified similarities in gene expression profiles of CD34+ CD38- LSC fractions in AML and normal HSCs
5 AML patients (Gal et al., 2006)
AML stem cell biology Compared expression profiles of normal HSC with LSC in AML to determine differentially expressed molecular pathways
16 AML patients (Majeti et al., 2009)
AML stem cell biology Identified human LSC gene expression signature in AML 61 AML patients (Saito et al., 2010)
AML stem cell biology; AML prognosis
Identified an LSC- specific gene signature, and associated high expression of these genes with inferior outcomes in AML
1047 AML patients
(Gentles et al., 2010)
AML stem cell biology; AML prognosis
Identified molecular signature defining functional AML LSC, and its use in predicting patient survival
16 AML patients (Eppert et al., 2011)
APL biology APL mouse models have distinct gene expression signatures influenced by genetic changes that contribute to the disease progression
Mouse APL models
(Walter et al., 2004)
APL biology Distinct gene expression signatures in M3 and M3 variant APLs. 35 APL patients; 155 other AML patients
(Haferlach et al., 2005a)
55
APL biology Identified deregulated genes and pathways contributing to disease progression in APL mouse models
APL mouse models
(Yuan et al., 2007)
APL sub-classification; APL biology
Identified differentially expressed genes in APL samples with presence of Flt3 ITDs
18 APL patients (Marasca et al., 2006)
APL expression signature Identified gene expression signatures specific to APL and distinct from other AML subtypes
15 APL patients; 62 other AML patients.
(Payton et al., 2009)
Therapeutic sensitivity prediction
Identified gene expression changes correlated with response to chemotherapy, and predictive of chemosensitivity
76 AML patients (Okutsu et al., 2002)
Therapeutic mechanisms gene expression profiles of cells after 12-72 hrs of ATRA induced differentiation
NB4 cell line (Yang et al., 2003)
Therapeutic mechanisms Genes modulated by valproic acid in AML OCI/AML-2 cell line
(Trus et al., 2005)
Therapeutic mechanisms Gene expression signature determining resistance or sensitivity to differentiation agents in in vitro cell lines
through gene expression array analysis of the U937-NPM-RARA cell line (c.f. Chapter 3). We
determined that 8/11 genes, including STAT5A, MAP3K8, NFKBIA, CCL2, IL8, MMP9, JUNB
and TNFAIP3 were significantly differentially regulated (p<0.05, by one-way ANOVA) in
diagnostic APL samples compared to normal bone marrow, and had expression profiles that
could distinguish between APL and normal bone marrow in Principle Components Analysis
(Figure 2.1A-B).
Fms-like tyrosine kinase receptor 3 (FLT3) alterations including internal tandem duplications
(ITDs) are found in approximately 30% of APL patients and are associated with more severe
clinical features (Callens et al., 2005). In order to evaluate whether FLT3 mutation states are
associated with greater NF-κB transcriptional deregulation, we undertook a separate gene
expression analysis on mutated and non-mutated APL samples. FLT3 mutation analysis
conducted on a subset of our APL samples (17/23) indicated the presence of FLT3 internal
tandem duplications (ITDs) in 4/16 (25%) samples tested. However, FLT3 mutation status did
not correlate with significant changes in gene expression profiles (Table 2.4).
Furthermore, we also tested whether NF-κB network gene expression levels correlated with
PML-RARA breakpoints. APL patients expressing either the BCR1/2, or the BCR3 specific form
of PML-RARA were analyzed, but revealed no statistically significant correlations with
expression levels of the NF-κB gene set (Table 2.5).
Having observed deregulated expression in patient samples at diagnosis, we next asked whether
expression profiles revereted to normal in the same patients after treatment. Comparing 7 pairs of
diagnostic and post-treatment samples, indicated that expression levels of STAT5A, AZU1,
CCL2, SOD1, MMP9, and IL-8 significantly changed in diagnostic compared to treated samples.
76
In many cases, a clear reversion of gene expression patterns to normal levels was noted in the
post-treatment samples, demonstrating that these genes were responsive to ATRA therapy
(Figure 2.1C-D).
77
Table 2.2: APL patient characteristics. The PML-RARA breakpoint, as well as mutations present in FLT3 are noted for each APL patient sample. Entries marked “ND” were not profiled and therefore not available for reporting.
Table 2.3: Gene expression analysis of APL and Normal bone marrow indicating ranges, median fold change values, and significance analysis in comparison with a pooled normal sample as the reference.
APL (n=23)
Normal bone marrow (n=11)
Min Max Median Min Max Median p-value (APL vs Normal)
Table 3.3: Commonly deregulated gene targets identified in both NPM-RARA and NuMA-RARA expressing cells relative to control U937-
GFP
Probe Set ID Regulation Unigene (Avadis)
Gene Symbol Gene Title
1553194_at Down Hs.146542 NEGR1 neuronal growth regulator 1 1555247_a_at Down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange factor (GEF) 6 1555339_at Down Hs.190334 RAP1A RAP1A, member of RAS oncogene family 1555340_x_at Down Hs.190334 RAP1A RAP1A, member of RAS oncogene family 1558290_a_at Down Hs.133107 PVT1 Pvt1 oncogene homolog (mouse) 201951_at Down Hs.591293 ALCAM activated leukocyte cell adhesion molecule 201952_at Down Hs.591293 ALCAM activated leukocyte cell adhesion molecule 202648_at Down 205110_s_at Down Hs.6540 FGF13 fibroblast growth factor 13 205376_at Down Hs.658245 INPP4B inositol polyphosphate-4-phosphatase, type II, 105kDa 206759_at Down Hs.465778 FCER2 Fc fragment of IgE, low affinity II, receptor for (CD23) 206760_s_at Down Hs.465778 FCER2 Fc fragment of IgE, low affinity II, receptor for (CD23) 207543_s_at Down Hs.500047 P4HA1 procollagen-proline, 2-oxoglutarate 4-dioxygenase (proline 4-hydroxylase),
alpha polypeptide I 208937_s_at Down Hs.504609 ID1 inhibitor of DNA binding 1, dominant negative helix-loop-helix protein 209560_s_at Down Hs.533717 DLK1 delta-like 1 homolog (Drosophila) 210254_at Down Hs.99960 MS4A3 membrane-spanning 4-domains, subfamily A, member 3 (hematopoietic cell-
specific) 213350_at Down Hs.433529 RPS11 Ribosomal protein S11 213826_s_at Down LOC100133109 similar to histone 218976_at Down Hs.260720 DNAJC12 DnaJ (Hsp40) homolog, subfamily C, member 12 219112_at Down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange factor (GEF) 6 219697_at Down Hs.622536 HS3ST2 heparan sulfate (glucosamine) 3-O-sulfotransferase 2 219714_s_at Down Hs.656687 CACNA2D3 calcium channel, voltage-dependent, alpha 2/delta subunit 3
124
223501_at Down Hs.525157 TNFSF13B tumor necrosis factor (ligand) superfamily, member 13b 223721_s_at Down Hs.260720 DNAJC12 DnaJ (Hsp40) homolog, subfamily C, member 12 224356_x_at Down Hs.523702 MS4A6A membrane-spanning 4-domains, subfamily A, member 6A 225942_at Down Hs.247460 NLN neurolysin (metallopeptidase M3 family) 226789_at Down Hs.697682 LOC647121 embigin homolog (mouse) pseudogene 228055_at Down Hs.636624 NAPSB napsin B aspartic peptidase pseudogene 229461_x_at Down Hs.146542 NEGR1 neuronal growth regulator 1 230078_at Down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange factor (GEF) 6 235046_at Down Hs.176376 Transcribed locus 242476_at Down Hs.605126 Transcribed locus 243357_at Down Hs.146542 NEGR1 neuronal growth regulator 1 1554892_a_at Up Hs.99960 MS4A3 membrane-spanning 4-domains, subfamily A, member 3 (hematopoietic cell-
specific) 1554899_s_at Up Hs.433300 FCER1G Fc fragment of IgE, high affinity I, receptor for; gamma polypeptide 1556657_at Up Hs.687293 CDNA FLJ36459 fis, clone THYMU2014762 1556658_a_at Up Hs.687293 CDNA FLJ36459 fis, clone THYMU2014762 1569403_at Up Hs.670065 CDNA clone IMAGE:4706427 1570561_at Up 200748_s_at Up Hs.524910 FTH1 ferritin, heavy polypeptide 1 201324_at Up Hs.436298 EMP1 epithelial membrane protein 1 201325_s_at Up Hs.436298 EMP1 epithelial membrane protein 1 201464_x_at Up Hs.525704 JUN jun oncogene 201700_at Up Hs.534307 CCND3 cyclin D3 202241_at Up Hs.444947 TRIB1 tribbles homolog 1 (Drosophila) 202859_x_at Up Hs.551925 IL8 interleukin 8 202887_s_at Up Hs.523012 DDIT4 DNA-damage-inducible transcript 4 202890_at Up Hs.486548 MAP7 microtubule-associated protein 7 202917_s_at Up Hs.416073 S100A8 S100 calcium binding protein A8 202932_at Up Hs.194148 YES1 v-yes-1 Yamaguchi sarcoma viral oncogene homolog 1 202933_s_at Up Hs.194148 YES1 v-yes-1 Yamaguchi sarcoma viral oncogene homolog 1 202988_s_at Up Hs.75256 RGS1 regulator of G-protein signaling 1
125
203535_at Up Hs.112405 S100A9 S100 calcium binding protein A9 203559_s_at Up Hs.647097 ABP1 amiloride binding protein 1 (amine oxidase (copper-containing) 203767_s_at Up Hs.522578 STS steroid sulfatase (microsomal), isozyme S 203936_s_at Up Hs.297413 MMP9 matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa type IV
collagenase) 204017_at Up Hs.709898 KDELR3 KDEL (Lys-Asp-Glu-Leu) endoplasmic reticulum protein retention receptor 3 204044_at Up Hs.513484 QPRT quinolinate phosphoribosyltransferase (nicotinate-nucleotide pyrophosphorylase
(carboxylating) 204118_at Up Hs.243564 CD48 CD48 molecule 204232_at Up Hs.433300 FCER1G Fc fragment of IgE, high affinity I, receptor for; gamma polypeptide 204533_at Up Hs.632586 CXCL10 chemokine (C-X-C motif) ligand 10 205222_at Up Hs.429879 EHHADH enoyl-Coenzyme A, hydratase/3-hydroxyacyl Coenzyme A dehydrogenase 205476_at Up Hs.75498 CCL20 chemokine (C-C motif) ligand 20 205749_at Up Hs.72912 CYP1A1 cytochrome P450, family 1, subfamily A, polypeptide 1 206332_s_at Up Hs.380250 IFI16 interferon, gamma-inducible protein 16 207554_x_at Up Hs.442530 TBXA2R thromboxane A2 receptor 209955_s_at Up Hs.654370 FAP fibroblast activation protein, alpha 210078_s_at Up Hs.654519 KCNAB1 potassium voltage-gated channel, shaker-related subfamily, beta member 1 210102_at Up Hs.152944 LOH11CR2A loss of heterozygosity, 11, chromosomal region 2, gene A 210471_s_at Up Hs.654519 KCNAB1 potassium voltage-gated channel, shaker-related subfamily, beta member 1 210772_at Up Hs.99855 FPR2 formyl peptide receptor 2 211506_s_at Up Hs.551925 IL8 interleukin 8 211628_x_at Up Hs.453583 FTHP1 ferritin, heavy polypeptide pseudogene 1 212509_s_at Up Hs.250723 MXRA7 matrix-remodelling associated 7 213017_at Up Hs.397978 ABHD3 abhydrolase domain containing 3 213836_s_at Up Hs.463964 WIPI1 WD repeat domain, phosphoinositide interacting 1 214211_at Up Hs.524910 FTH1 ferritin, heavy polypeptide 1 215071_s_at Up Hs.484950 HIST1H2AC histone cluster 1, H2ac 216598_s_at Up Hs.303649 CCL2 chemokine (C-C motif) ligand 2 216834_at Up Hs.75256 RGS1 regulator of G-protein signaling 1 217996_at Up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1
126
217997_at Up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 218330_s_at Up Hs.502116 NAV2 neuron navigator 2 218559_s_at Up Hs.712609 MAFB v-maf musculoaponeurotic fibrosarcoma oncogene homolog B (avian) 218656_s_at Up Hs.507798 LHFP lipoma HMGIC fusion partner 218919_at Up Hs.655453 ZFAND1 zinc finger, AN1-type domain 1 219434_at Up Hs.283022 TREM1 triggering receptor expressed on myeloid cells 1 219596_at Up Hs.591123 THAP10 THAP domain containing 10 220517_at Up Hs.511668 VPS13C vacuolar protein sorting 13 homolog C (S. cerevisiae) 221909_at Up Hs.437195 RNFT2 ring finger protein, transmembrane 2 223620_at Up Hs.495989 GPR34 G protein-coupled receptor 34 224048_at Up Hs.646421 USP44 ubiquitin specific peptidase 44 224802_at Up Hs.525093 NDFIP2 Nedd4 family interacting protein 2 226218_at Up Hs.635723 IL7R interleukin 7 receptor 226254_s_at Up Hs.535734 KIAA1430 KIAA1430 226771_at Up Hs.435700 ATP8B2 ATPase, class I, type 8B, member 2 227682_at Up Hs.595314 Transcribed locus 228155_at Up Hs.500333 C10orf58 chromosome 10 open reading frame 58 228170_at Up Hs.56663 OLIG1 oligodendrocyte transcription factor 1 228285_at Up Hs.21454 TDRD9 tudor domain containing 9 228384_s_at Up Hs.238303 C10orf33 chromosome 10 open reading frame 33 228964_at Up Hs.436023 PRDM1 PR domain containing 1, with ZNF domain 336_at Up Hs.442530 TBXA2R thromboxane A2 receptor 34210_at Up Hs.276770 CD52 CD52 molecule 36711_at Up Hs.517617 MAFF v-maf musculoaponeurotic fibrosarcoma oncogene homolog F (avian)
127
Table 3.4: KEGG pathways identified to be significantly over-represented within genes commonly deregulated by NPM-RARA and NuMA-
TNFAIP3* Tumor necrosis factor, alpha-induced protein 3 hydrolase activity; double-stranded DNA binding Yes Yes S100P Protein S100-P calcium ion binding; calmodulin binding No Yes IFIH1 Interferon-induced helicase C domain-containing
ALKBH7 Alkylated DNA repair protein alkB homolog 7 ANKRD13C Ankyrin repeat domain-containing protein 13C ARHGAP18 Rho GTPase-activating protein 18 protein binding;small GTPase
CST3 Cystatin-C protein binding;cysteine-type endopeptidase inhibitor activity
CUL4A Cullin-4A CYBASC3 Cytochrome b ascorbate-dependent protein 3 oxidoreductase activity DDIT4 DNA-damage-inducible transcript 4 protein DMXL2 DmX-like protein 2 DNAJC13 DnaJ homolog subfamily C member 13 DUSP10 Dual specificity protein phosphatase 10 phosphoprotein phosphatase
PYHIN1 Pyrin and HIN domain-containing protein 1 transcription factor activity RAP1A Ras-related protein Rap-1A GTPase activity;protein binding Heterotrimeric G-protein signaling
pathway-Gq alpha and Go alpha mediated pathway->Ras associated protein 1
RAPGEF6 Rap guanine nucleotide exchange factor 6 protein binding;small GTPase
To determine early ATRA responsive transcripts in NuMA- and NPM-RARA cells, cells were
treated with 1 µM ATRA for 4 hours and RNA was extracted and analyzed. These data
determined 510 (174 down, 336 up) early ATRA-responsive transcripts in U937-NPM-RARA
cells (Figure 3.7; see Appendix I Table A3.3). A total of 289 (75 down, 214 up) ATRA-
regulated transcripts were recorded for U937-NuMA-RARA cells (see Appendix I Table A3.4).
U937-GFP cells were used to determine the wild type ATRA response, and exhibited 352 ATRA
responsive targets (116 down, 236 up) (see Appendix I Table A3.5). The ATRA response in
NuMA-RARA, NPM-RARA, and the GFP system included 152 commonly regulated targets;
these potentially represent early wild-type retinoid responsive signaling genes that are also
affected by the fusions (Table 3.11; Figure 3.7). Interestingly, 41% of ATRA responsive targets
in NPM-RARA, and 27% of ATRA responsive probesets in NuMA-RARA were unique to these
specific fusions, and not regulated by ATRA in wild type cells; these may represent novel
retinoid responsive targets affected by the variant APL fusion proteins.
Our goal in this analysis was to compare the retinoid response at the level of transcriptional
control, and understand how retinoid induced signaling and transcriptional control is affected in
cells expressing NPM-RARA and NuMA-RARA. To this end, we identified that, of the 22
transcription factor binding motifs significantly over-represented in the wild type ATRA
response dataset (in GFP control cells), the majority (13/22 = 59%) of these were shared by
NPM-RARA cells (Table 3.12; Table 3.14). This set included both RAR:RXR and NF-κB
binding sites. Interestingly, the binding motifs identified to be unique to NPM-RARA alone
includes dimers containing PPARγ:RXRA. This is in agreement with previous studies from our
lab identifying the deregulation of PPARγ signaling in NPM-RARA expressing cells.
Additionally, ras responsive element binding protein 1 (RREB1), nuclear factor of activated T-
cells, cytoplasmic, calcineurin-dependent 2 (NFATC2), CCCTC-binding factor (CTCF) and
Paired box 4 (PAX4) binding sites were significantly enriched in the fusion target list, implying
that the global transcriptional disruption induced by the fusion is in part mediated through
deregulated activity in these other transcriptional control networks. NuMA-RARA ATRA
response targets had enriched binding sites that overlapped with those of control cells (Table
3.13, Table 3.14). This overlap suggests that early ATRA targets in NuMA-RARA cells
158
classical retinoid signaling genes, which are also involved in the ATRA response profile of
control wild type cells.
We next compared the ATRA responsive gene targets in NPM-RARA with that of genes
deregulated in APL patient blasts. This analysis identified that 151 ATRA responsive gene
targets in NPM-RARA were also deregulated in APL patient samples. Furthermore, these probe
sets showed opposite regulation patterns in APL patients versus the ATRA responsive case, as
expected. This indicates that the ATRA responsive gene sets identified in the NPM-RARA cell
line are physiologically relevant, as they are also direct/indirect targets of the PML-RARA fusion
protein in primary APL patient blasts.
Having identified early ATRA responsive gene targets of the fusion, we next asked whether any
of these targets overlapped with PML-RARA and PLZF-RARA direct binding sites that were
also identified to be deregulated by the NPM-RARA fusion. A total of 10 such genes including
BIRC3, C1orf106, CCL2, CD48, IL1B, MAFF, OLIG1, TREM1, TRIB1, and ID1 were
identified as gene targets that are deregulated by NPM-RARA, directly bound by PML-RARA,
and transcriptionally responsive to treatment with ATRA. These genes which are modulated
early after retinoic acid treatment and also deregulated by the presence of NPM-RARA represent
a critical subset of fusion direct targets.
159
Figure 3.7: ATRA induced gene expression changes in NPM-RARA, and NuMA-RARA in
comparison with wild type U937-GFP. A) Probesets differentially regulated at least 2-fold
after 4-hrs of ATRA treatment in NuMA-RARA, NPM-RARA, and control U937-GFP are
quantified and represented according to their regulation pattern. B) Venn diagrams representing
genes up-regulated; and C) down-regulated after ATRA treatment in NuMA-RARA, NPM-
RARA, and control GFP are compared to indicate degree of overlap between the two fusion
retinoid responses and the wild type response. D) The NPM-RARA retinoid response; and E)
NuMA-RARA retinoid response were compared to highlight the presence of wild type retinoid
response genes in both NPM-RARA and NuMA-RARA ATRA response targets. In addition, the
presence of novel retinoid targets present uniquely within the NPM-RARA and NuMA-RARA
datasets are also noted.
160
Figure 3.7: ATRA induced gene expression changes in NPM-RARA, and NuMA-RARA in
comparison with wild type U937-GFP.
161
Table 3.11: Genes commonly up- and down-regulated after ATRA treatment commonly in NPM-RARA, NuMA-RARA and wild type U937-
GFP cells.
Probe Set ID Regulation Unigene (Avadis)
Gene Symbol
Gene Title
201565_s_at Down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix protein 201566_x_at Down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix protein 202431_s_at Down Hs.202453 MYC v-myc myelocytomatosis viral oncogene homolog (avian) 203304_at Down Hs.533336 BAMBI BMP and activin membrane-bound inhibitor homolog (Xenopus laevis) 204794_at Down Hs.1183 DUSP2 dual specificity phosphatase 2 204897_at Down Hs.199248 PTGER4 prostaglandin E receptor 4 (subtype EP4) 205419_at Down Hs.784 EBI2 Epstein-Barr virus induced gene 2 (lymphocyte-specific G protein-coupled
receptor) 206157_at Down Hs.591286 PTX3 pentraxin-related gene, rapidly induced by IL-1 beta 209099_x_at Down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 209184_s_at Down Hs.442344 IRS2 insulin receptor substrate 2 209606_at Down Hs.270 PSCDBP pleckstrin homology, Sec7 and coiled-coil domains, binding protein 213931_at Down Hs.591670 ID2 /// ID2B inhibitor of DNA binding 2, dominant negative helix-loop-helix protein /// inhibitor
of DNA binding 2B, dominant negative helix-loop-helix protein 216268_s_at Down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 219497_s_at Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219498_s_at Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219890_at Down Hs.446235 CLEC5A C-type lectin domain family 5, member A 220116_at Down Hs.98280 KCNN2 potassium intermediate/small conductance calcium-activated channel, subfamily N,
member 2 221586_s_at Down Hs.445758 E2F5 E2F transcription factor 5, p130-binding 221766_s_at Down Hs.10784 FAM46A family with sequence similarity 46, member A 222891_s_at Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 223403_s_at Down Hs.86337 POLR1B polymerase (RNA) I polypeptide B, 128kDa 224973_at Down Hs.10784 FAM46A family with sequence similarity 46, member A 227037_at Down Hs.31652 LOC201164 similar to CG12314 gene product
162
227210_at Down Hs.407983 SFMBT2 Scm-like with four mbt domains 2 227242_s_at Down Hs.699395 EBF3 early B-cell factor 3 227481_at Down Hs.16064 CNKSR3 CNKSR family member 3 228170_at Down Hs.56663 OLIG1 oligodendrocyte transcription factor 1 229638_at Down Hs.499205 IRX3 iroquois homeobox 3 236738_at Down Hs.710781 LOC401097 Similar to LOC166075 239129_at Down 239605_x_at Down Hs.657657 Transcribed locus 240747_at Down Hs.667630 Transcribed locus 242388_x_at Down Hs.601883 Transcribed locus 243529_at Down Hs.116602 MARS2 methionyl-tRNA synthetase 2, mitochondrial 1554240_a_at Up Hs.174103 ITGAL integrin, alpha L (antigen CD11A (p180), lymphocyte function-associated antigen
1; alpha polypeptide) 1555431_a_at Up Hs.55378 IL31RA interleukin 31 receptor A 1555680_a_at Up Hs.433337 SMOX spermine oxidase 1555728_a_at Up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 1563745_a_at Up Hs.309176 LOC283050 hypothetical LOC283050 1568619_s_at Up Hs.648523 LOC162073 hypothetical protein LOC162073 1570375_at Up Hs.661265 CDNA FLJ41985 fis, clone SPLEN2014946 200897_s_at Up Hs.151220 PALLD palladin, cytoskeletal associated protein 200906_s_at Up Hs.151220 PALLD palladin, cytoskeletal associated protein 200907_s_at Up Hs.151220 PALLD palladin, cytoskeletal associated protein 201042_at Up Hs.517033 TGM2 transglutaminase 2 (C polypeptide, protein-glutamine-gamma-glutamyltransferase) 201534_s_at Up Hs.145575 UBL3 ubiquitin-like 3 201656_at Up Hs.133397 ITGA6 integrin, alpha 6 202308_at Up Hs.592123 SREBF1 sterol regulatory element binding transcription factor 1 202481_at Up Hs.289347 DHRS3 dehydrogenase/reductase (SDR family) member 3 202627_s_at Up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1),
member 1 202628_s_at Up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1),
member 1 202770_s_at Up Hs.13291 CCNG2 cyclin G2
163
202869_at Up Hs.524760 OAS1 2',5'-oligoadenylate synthetase 1, 40/46kDa 202887_s_at Up Hs.523012 DDIT4 DNA-damage-inducible transcript 4 203760_s_at Up Hs.75367 SLA Src-like-adaptor 203761_at Up Hs.75367 SLA Src-like-adaptor 203887_s_at Up Hs.2030 THBD Thrombomodulin 203888_at Up Hs.2030 THBD Thrombomodulin 204112_s_at Up Hs.42151 HNMT histamine N-methyltransferase 204429_s_at Up Hs.530003 SLC2A5 solute carrier family 2 (facilitated glucose/fructose transporter), member 5 204430_s_at Up Hs.530003 SLC2A5 solute carrier family 2 (facilitated glucose/fructose transporter), member 5 204526_s_at Up Hs.442657 TBC1D8 TBC1 domain family, member 8 (with GRAM domain) 204961_s_at Up Hs.655201 LOC648998
214523_at Up Hs.558308 CEBPE CCAAT/enhancer binding protein (C/EBP), epsilon 214639_s_at Up Hs.67397 HOXA1 homeobox A1 214977_at Up Hs.654670 CDNA FLJ13790 fis, clone THYRO1000026 215177_s_at Up Hs.133397 ITGA6 integrin, alpha 6 215342_s_at Up Hs.585378 RABGAP1L RAB GTPase activating protein 1-like 218501_at up Hs.476402 ARHGEF3 Rho guanine nucleotide exchange factor (GEF) 3 218627_at up Hs.525634 DRAM damage-regulated autophagy modulator 218723_s_at up Hs.507866 C13orf15 chromosome 13 open reading frame 15 219010_at up Hs.518997 C1orf106 chromosome 1 open reading frame 106 219607_s_at up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 219994_at up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B, member 1 interacting
protein 221266_s_at up Hs.652230 TM7SF4 transmembrane 7 superfamily member 4 221345_at up Hs.248056 FFAR2 free fatty acid receptor 2 221601_s_at up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 221602_s_at up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 223276_at up Hs.29444 MST150 MSTP150 223567_at up Hs.465642 SEMA6B sema domain, transmembrane domain (TM), and cytoplasmic domain,
(semaphorin) 6B 223620_at up Hs.495989 GPR34 G protein-coupled receptor 34 224916_at up Hs.379754 TMEM173 transmembrane protein 173 224929_at up Hs.379754 TMEM173 transmembrane protein 173
165
224964_s_at up Hs.187772 GNG2 guanine nucleotide binding protein (G protein), gamma 2 225347_at up Hs.497399 ARL8A ADP-ribosylation factor-like 8A 225372_at up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225373_at up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225763_at up Hs.493867 RCSD1 RCSD domain containing 1 225919_s_at up Hs.493639 C9orf72 chromosome 9 open reading frame 72 226487_at up Hs.661785 C12orf34 chromosome 12 open reading frame 34 226722_at up Hs.134742 FAM20C family with sequence similarity 20, member C 226756_at up Hs.633903 CDNA FLJ25556 fis, clone JTH02629 227396_at up Hs.318547 PTPRJ protein tyrosine phosphatase, receptor type, J 227484_at up Hs.710097 CDNA FLJ41690 fis, clone HCASM2009405 227792_at up Hs.648523 LOC162073 hypothetical protein LOC162073 227915_at up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 228648_at up Hs.655559 LRG1 leucine-rich alpha-2-glycoprotein 1 228772_at up Hs.42151 HNMT histamine N-methyltransferase 229670_at up Hs.180284 5.5 kb mRNA upregulated in retinoic acid treated HL-60 neutrophilic cells 229971_at up Hs.187884 GPR114 G protein-coupled receptor 114 230218_at up Hs.72956 HIC1 hypermethylated in cancer 1 230333_at up Hs.656630 Transcribed locus 230925_at up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B, member 1 interacting
protein 231496_at up Hs.145519 FCAMR Fc receptor, IgA, IgM, high affinity 231779_at up Hs.449207 IRAK2 interleukin-1 receptor-associated kinase 2 231969_at up Hs.21958 STOX2 storkhead box 2 232687_at up Hs.100912 CDNA FLJ33091 fis, clone TRACH2000660 232861_at up Hs.654693 PDP2 pyruvate dehydrogenase phosphatase isoenzyme 2 233857_s_at up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 234987_at up Hs.660221 Transcribed locus 235421_at up Hs.432453/
236407_at up Hs.121495 KCNE1 potassium voltage-gated channel, Isk-related family, member 1 236717_at up Hs.525977 LOC165186 similar to RIKEN cDNA 4632412N22 gene 237252_at up Hs.2030 THBD Thrombomodulin 237442_at up 238032_at up Hs.655631 Transcribed locus 238439_at up Hs.217484 ANKRD22 ankyrin repeat domain 22 238669_at up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H synthase and
cyclooxygenase) 239294_at up Hs.561747 Transcribed locus 242426_at up Hs.696574 NRG4 neuregulin 4 242525_at up Hs.439122 Transcribed locus 243541_at up Hs.55378 IL31RA interleukin 31 receptor A 243819_at up 244665_at up Hs.668855 Transcribed locus 36711_at up Hs.517617 MAFF v-maf musculoaponeurotic fibrosarcoma oncogene homolog F (avian) 37152_at up Hs.696032 PPARD peroxisome proliferator-activated receptor delta
167
Table 3.12: Promoter binding sites that are over-represented in NPM-RARA ATRA responsive
polymerase (RNA) II (DNA directed) polypeptide J, 13.3kDa pseudogene /// DNA directed RNA polymerase II polypeptide J-related /// similar to hCG1989139 /// PMS2 postmeiotic segregation increased 2 (S. cerevisiae)-like /// similar to POLR2J4 protein
PI-3-kinase-related kinase SMG-1 /// hypothetical protein LOC440345 /// PI-3-kinase-related kinase SMG-1 pseudogene /// hypothetical LOC728423 /// similar to PI-3-kinase-related kinase SMG-1
230
LOC728423 /// LOC729513 /// SMG1
214181_x_at 2.025713 down Hs.436066 LST1 leukocyte specific transcript 1 218138_at 2.026455 down Hs.472119 MKKS McKusick-Kaufman syndrome 206398_s_at 2.028411 down Hs.652262 CD19 CD19 molecule 209628_at 2.0312347 down Hs.25010 NXT2 nuclear transport factor 2-like export factor 2 207008_at 2.034133 down Hs.846 IL8RB interleukin 8 receptor, beta 1570441_at 2.0343087 down Hs.269471 NAPB N-ethylmaleimide-sensitive factor attachment protein, beta 211582_x_at 2.036575 down Hs.436066 LST1 leukocyte specific transcript 1 225762_x_at 2.0415037 down Hs.370699 LOC284801 hypothetical protein LOC284801 209539_at 2.041854 down Hs.522795 ARHGEF6 Rac/Cdc42 guanine nucleotide exchange factor (GEF) 6 202075_s_at 2.0443308 down Hs.439312 PLTP phospholipid transfer protein 1559410_at 2.0455973 down Hs.490920 Transcribed locus 219551_at 2.0457556 down Hs.477325 EAF2 ELL associated factor 2 212616_at 2.045807 down Hs.59159 CHD9 chromodomain helicase DNA binding protein 9 223746_at 2.0461335 down Hs.472838 STK4 serine/threonine kinase 4 215739_s_at 2.0482981 down Hs.224152 TUBGCP3 tubulin, gamma complex associated protein 3 230064_at 2.0492828 down Hs.650577 Transcribed locus 201267_s_at 2.050726 down Hs.250758 PSMC3 proteasome (prosome, macropain) 26S subunit, ATPase, 3 225915_at 2.0515246 down Hs.87159 CAB39L calcium binding protein 39-like 1560026_at 2.0552907 down Hs.685021 CDNA clone IMAGE:5285703 236699_at 2.058871 down Hs.660409 CDNA FLJ90129 fis, clone HEMBB1000309 200965_s_at 2.0590336 down Hs.438236 ABLIM1 actin binding LIM protein 1 234681_s_at 2.0597532 down Hs.371979 CHD6 chromodomain helicase DNA binding protein 6 225768_at 2.0673113 down Hs.37288 NR1D2 nuclear receptor subfamily 1, group D, member 2 200722_s_at 2.067345 down Hs.471818 CAPRIN1 cell cycle associated protein 1 242024_at 2.0687475 down Hs.656132 Transcribed locus 219697_at 2.0763223 down Hs.622536 HS3ST2 heparan sulfate (glucosamine) 3-O-sulfotransferase 2 217984_at 2.0788753 down Hs.529989 RNASET2 ribonuclease T2 207871_s_at 2.081942 down Hs.368131 ST7 suppression of tumorigenicity 7
231
225065_x_at 2.083929 down Hs.368934 C17orf45 chromosome 17 open reading frame 45 231939_s_at 2.094253 down Hs.258272 BDP1 B double prime 1, subunit of RNA polymerase III
transcription initiation factor IIIB 225305_at 2.094777 down Hs.578109 SLC25A29 solute carrier family 25, member 29 222765_x_at 2.0996466 down Hs.369284 ESF1 ESF1, nucleolar pre-rRNA processing protein, homolog (S.
cerevisiae) 211581_x_at 2.1002464 down Hs.436066 LST1 leukocyte specific transcript 1 209901_x_at 2.1039147 down Hs.76364 AIF1 allograft inflammatory factor 1 226436_at 2.107545 down Hs.522895 RASSF4 Ras association (RalGDS/AF-6) domain family member 4 243341_at 2.111661 down 204205_at 2.112657 down Hs.660143 APOBEC3F ///
APOBEC3G apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3G /// apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3F
244779_at 2.1131718 down Hs.587465 CDNA FLJ34038 fis, clone FCBBF2005645 210222_s_at 2.1211119 down Hs.368626 RTN1 reticulon 1 200868_s_at 2.1215606 down Hs.144949 ZNF313 zinc finger protein 313 242476_at 2.1248302 down Hs.605126 Transcribed locus 243366_s_at 2.1272874 down Hs.72981 Transcribed locus 202659_at 2.1324925 down Hs.9661 PSMB10 proteasome (prosome, macropain) subunit, beta type, 10 244718_at 2.1389098 down Hs.471067 LOC339483 Hypothetical LOC339483 202854_at 2.1476676 down Hs.412707 HPRT1 hypoxanthine phosphoribosyltransferase 1 (Lesch-Nyhan
syndrome) 219599_at 2.1498694 down Hs.648394 EIF4B eukaryotic translation initiation factor 4B 210943_s_at 2.150597 down Hs.532411 LYST lysosomal trafficking regulator 1566482_at 2.160243 down Hs.684006 Transcribed locus 222859_s_at 2.1645083 down Hs.436271 DAPP1 dual adaptor of phosphotyrosine and 3-phosphoinositides 1558822_at 2.1657143 down Hs.684614 Full length insert cDNA clone YP59C02 225943_at 2.1674364 down Hs.247460 NLN neurolysin (metallopeptidase M3 family) 240772_at 2.1703398 down Hs.687668 Transcribed locus 227485_at 2.1742713 down Hs.496829 DDX26B DEAD/H (Asp-Glu-Ala-Asp/His) box polypeptide 26B 200723_s_at 2.174671 down Hs.471818 CAPRIN1 cell cycle associated protein 1 204643_s_at 2.1786573 down Hs.171458 ENOX2 ecto-NOX disulfide-thiol exchanger 2
232
1562194_at 2.183296 down Hs.684712 Full length insert cDNA clone YW28D08 221514_at 2.1872556 down Hs.458598 UTP14A UTP14, U3 small nucleolar ribonucleoprotein, homolog A
(yeast) 218322_s_at 2.1907299 down Hs.11638 ACSL5 acyl-CoA synthetase long-chain family member 5 223922_x_at 2.1933236 down Hs.523702 MS4A6A membrane-spanning 4-domains, subfamily A, member 6A 201539_s_at 2.1956432 down Hs.435369 FHL1 four and a half LIM domains 1 1554892_a_at 2.207399 down Hs.99960 MS4A3 membrane-spanning 4-domains, subfamily A, member 3
(hematopoietic cell-specific) 211937_at 2.2092397 down Hs.648394 EIF4B eukaryotic translation initiation factor 4B 226289_at 2.2250307 down Hs.471818 CAPRIN1 cell cycle associated protein 1 220252_x_at 2.22847 down Hs.665009 CXorf21 chromosome X open reading frame 21 202201_at 2.2379 down Hs.515785 BLVRB biliverdin reductase B (flavin reductase (NADPH) 222003_s_at 2.2533574 down Hs.591002 DOCK6 dedicator of cytokinesis 6 209009_at 2.2579336 down Hs.432491 ESD esterase D/formylglutathione hydrolase 239017_at 2.2613833 down Hs.351043 Transcribed locus 243370_at 2.2642157 down Hs.471818 CAPRIN1 cell cycle associated protein 1 207543_s_at 2.2717898 down Hs.500047 P4HA1 procollagen-proline, 2-oxoglutarate 4-dioxygenase (proline
4-hydroxylase), alpha polypeptide I 1561939_at 2.2795866 down Hs.503721 DYNC2H1 dynein, cytoplasmic 2, heavy chain 1 229498_at 2.2831652 down Hs.291319 Transcribed locus 1557261_at 2.2937984 down Hs.655308 LOC652637 ///
WHDC1L1 /// WHDC1L2
WAS protein homology region 2 domain containing 1-like 1 /// WAS protein homology region 2 domain containing 1-like 2 /// similar to junction-mediating and regulatory protein
243514_at 2.2962644 down Hs.661045 Transcribed locus 209294_x_at 2.3005726 down Hs.521456 TNFRSF10B tumor necrosis factor receptor superfamily, member 10b 1556316_s_at 2.3006198 down Hs.707281 LOC284889 hypothetical protein LOC284889 1553185_at 2.3046198 down Hs.657750 RASEF RAS and EF-hand domain containing 224789_at 2.3079128 down Hs.709474 WDR40A WD repeat domain 40A 229367_s_at 2.3168159 down Hs.647105 GIMAP6 GTPase, IMAP family member 6 223328_at 2.3228679 down Hs.287412 ARMC10 armadillo repeat containing 10 217867_x_at 2.3252523 down Hs.529408 BACE2 beta-site APP-cleaving enzyme 2 1564972_x_at 2.3369355 down Hs.631789 SETDB2 SET domain, bifurcated 2
233
232466_at 2.3381941 down Hs.339735 CUL4A Cullin 4A 230435_at 2.3404386 down Hs.710370 LOC375190 hypothetical LOC375190 204499_at 2.3487406 down Hs.494321 AGTPBP1 ATP/GTP binding protein 1 239294_at 2.3601356 down Hs.561747 Transcribed locus 234303_s_at 2.362843 down Hs.152009 GPR85 G protein-coupled receptor 85 228523_at 2.3698564 down Hs.591918 NANOS1 nanos homolog 1 (Drosophila) 223502_s_at 2.3810837 down Hs.525157 TNFSF13B tumor necrosis factor (ligand) superfamily, member 13b 226789_at 2.388231 down Hs.697682 LOC647121 embigin homolog (mouse) pseudogene 226785_at 2.3931837 down Hs.88252 ATP11C ATPase, class VI, type 11C 243357_at 2.398746 down Hs.146542 NEGR1 neuronal growth regulator 1 226999_at 2.399876 down Hs.632430 RNPC3 RNA-binding region (RNP1, RRM) containing 3 220331_at 2.399885 down Hs.25121 CYP46A1 cytochrome P450, family 46, subfamily A, polypeptide 1 202304_at 2.405207 down Hs.508010 FNDC3A fibronectin type III domain containing 3A 209199_s_at 2.4231865 down Hs.699175 MEF2C myocyte enhancer factor 2C 210254_at 2.424963 down Hs.99960 MS4A3 membrane-spanning 4-domains, subfamily A, member 3
(hematopoietic cell-specific) 212686_at 2.4263275 down Hs.435479 PPM1H protein phosphatase 1H (PP2C domain containing) 201360_at 2.4286895 down Hs.304682 CST3 cystatin C 200953_s_at 2.4785903 down Hs.376071 CCND2 cyclin D2 226835_s_at 2.5095668 down Hs.356766 C20orf199 chromosome 20 open reading frame 199 203630_s_at 2.5241778 down Hs.239631 COG5 component of oligomeric golgi complex 5 212229_s_at 2.5375736 down Hs.159699 FBXO21 F-box protein 21 212231_at 2.5438178 down Hs.159699 FBXO21 F-box protein 21 213642_at 2.5552883 down 210298_x_at 2.5561078 down Hs.435369 FHL1 four and a half LIM domains 1 224346_at 2.5577912 down 204949_at 2.5660028 down Hs.654563 ICAM3 intercellular adhesion molecule 3 235520_at 2.5661733 down Hs.308418 ZNF280C zinc finger protein 280C 218918_at 2.56725 down Hs.197043 MAN1C1 mannosidase, alpha, class 1C, member 1 229937_x_at 2.567973 down Hs.667388 LILRB1 Leukocyte immunoglobulin-like receptor, subfamily B (with
TM and ITIM domains), member 1
234
229026_at 2.5713942 down Hs.653643 Transcribed locus 228055_at 2.576628 down Hs.636624 NAPSB napsin B aspartic peptidase pseudogene 206403_at 2.6169062 down Hs.378901 ZNF536 zinc finger protein 536 223501_at 2.6326554 down Hs.525157 TNFSF13B tumor necrosis factor (ligand) superfamily, member 13b 226814_at 2.654285 down Hs.656071 ADAMTS9 ADAM metallopeptidase with thrombospondin type 1 motif,
9 200952_s_at 2.6592095 down Hs.376071 CCND2 cyclin D2 205376_at 2.6637907 down Hs.658245 INPP4B inositol polyphosphate-4-phosphatase, type II, 105kDa 202648_at 2.7345772 down 219463_at 2.7355642 down Hs.22920 C20orf103 chromosome 20 open reading frame 103 200908_s_at 2.7451262 down Hs.437594 RPLP2 ribosomal protein, large, P2 1558290_a_at 2.7996411 down Hs.133107 PVT1 Pvt1 oncogene homolog (mouse) 218217_at 2.8231165 down Hs.514950 SCPEP1 serine carboxypeptidase 1 217948_at 2.8297436 down Hs.460924 FAM127B family with sequence similarity 127, member B 206759_at 2.8359363 down Hs.465778 FCER2 Fc fragment of IgE, low affinity II, receptor for (CD23) 224407_s_at 2.8525004 down Hs.444247 RP6-213H19.1 serine/threonine protein kinase MST4 223280_x_at 2.8559701 down Hs.523702 MS4A6A membrane-spanning 4-domains, subfamily A, member 6A 237899_at 2.8577225 down Hs.631532 LOC729994 hypothetical LOC729994 219694_at 2.866797 down Hs.591751 FAM105A family with sequence similarity 105, member A 214505_s_at 2.874888 down Hs.435369 FHL1 four and a half LIM domains 1 210299_s_at 2.8758771 down Hs.435369 FHL1 four and a half LIM domains 1 204057_at 2.8920434 down Hs.137427 IRF8 interferon regulatory factor 8 205942_s_at 2.9905415 down Hs.706754 ACSM3 acyl-CoA synthetase medium-chain family member 3 225942_at 3.023462 down Hs.247460 NLN neurolysin (metallopeptidase M3 family) 1552612_at 3.0316563 down Hs.508829 CDC42SE2 CDC42 small effector 2 209560_s_at 3.037277 down Hs.533717 DLK1 delta-like 1 homolog (Drosophila) 218476_at 3.050557 down Hs.522449 POMT1 protein-O-mannosyltransferase 1 227976_at 3.074569 down Hs.42239 LOC644538 hypothetical protein LOC644538 201828_x_at 3.1145337 down Hs.522789 FAM127A family with sequence similarity 127, member A 218499_at 3.146391 down Hs.444247 RP6-213H19.1 serine/threonine protein kinase MST4 201601_x_at 3.2108214 down Hs.458414 IFITM1 interferon induced transmembrane protein 1 (9-27)
235
232383_at 3.2426195 down Hs.125962 TFEC transcription factor EC 214022_s_at 3.248919 down Hs.458414 IFITM1 interferon induced transmembrane protein 1 (9-27) 210377_at 3.2570925 down Hs.706754 ACSM3 acyl-CoA synthetase medium-chain family member 3 219666_at 3.3481128 down Hs.523702 MS4A6A membrane-spanning 4-domains, subfamily A, member 6A 201952_at 3.3782434 down Hs.591293 ALCAM activated leukocyte cell adhesion molecule 206760_s_at 3.428722 down Hs.465778 FCER2 Fc fragment of IgE, low affinity II, receptor for (CD23) 208937_s_at 3.5412586 down Hs.504609 ID1 inhibitor of DNA binding 1, dominant negative helix-loop-
helix protein 213826_s_at 3.5833752 down LOC100133109 similar to histone 1553194_at 3.6176825 down Hs.146542 NEGR1 neuronal growth regulator 1 223721_s_at 3.6589942 down Hs.260720 DNAJC12 DnaJ (Hsp40) homolog, subfamily C, member 12 224356_x_at 3.6768126 down Hs.523702 MS4A6A membrane-spanning 4-domains, subfamily A, member 6A 201951_at 3.6924899 down Hs.591293 ALCAM activated leukocyte cell adhesion molecule 213135_at 3.7599144 down Hs.517228 TIAM1 T-cell lymphoma invasion and metastasis 1 229461_x_at 3.8539445 down Hs.146542 NEGR1 neuronal growth regulator 1 205081_at 3.9021432 down Hs.70327 CRIP1 cysteine-rich protein 1 (intestinal) 222446_s_at 4.0326624 down Hs.529408 BACE2 beta-site APP-cleaving enzyme 2 235046_at 4.413527 down Hs.176376 Transcribed locus 213350_at 4.5241256 down Hs.433529 RPS11 Ribosomal protein S11 205110_s_at 4.5955734 down Hs.6540 FGF13 fibroblast growth factor 13 218976_at 4.647823 down Hs.260720 DNAJC12 DnaJ (Hsp40) homolog, subfamily C, member 12 206715_at 4.648782 down Hs.125962 TFEC transcription factor EC 215047_at 5.0228643 down Hs.323858 TRIM58 tripartite motif-containing 58 228617_at 5.0417824 down Hs.441975 XAF1 XIAP associated factor 1 239340_at 5.53087 down Hs.444181 Transcribed locus 1552386_at 5.5491457 down Hs.547697 C5orf29 chromosome 5 open reading frame 29 201540_at 5.8108406 down Hs.435369 FHL1 four and a half LIM domains 1 219112_at 6.622824 down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange factor (GEF) 6 203923_s_at 6.8786955 down Hs.292356 CYBB cytochrome b-245, beta polypeptide (chronic granulomatous
224709_s_at 7.61829 down Hs.508829 CDC42SE2 CDC42 small effector 2 1552613_s_at 7.6911745 down Hs.508829 CDC42SE2 CDC42 small effector 2 242715_at 10.302398 down Hs.444181 Transcribed locus 1555247_a_at 12.315356 down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange factor (GEF) 6 219714_s_at 14.661497 down Hs.656687 CACNA2D3 calcium channel, voltage-dependent, alpha 2/delta subunit 3 230078_at 34.80333 down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange factor (GEF) 6 1555339_at 47.755352 down Hs.190334 RAP1A RAP1A, member of RAS oncogene family 1555340_x_at 66.98855 down Hs.190334 RAP1A RAP1A, member of RAS oncogene family 241808_at 2.0012462 up Hs.658042 CDNA FLJ36977 fis, clone BRACE2006344 217678_at 2.006411 up Hs.390594 SLC7A11 solute carrier family 7, (cationic amino acid transporter, y+
system) member 11 207124_s_at 2.0091777 up Hs.155090 GNB5 guanine nucleotide binding protein (G protein), beta 5 224655_at 2.0091856 up Hs.493362 AK3 adenylate kinase 3 226771_at 2.010245 up Hs.435700 ATP8B2 ATPase, class I, type 8B, member 2 224733_at 2.0119143 up Hs.298198 CMTM3 CKLF-like MARVEL transmembrane domain containing 3 201920_at 2.0148942 up Hs.187946 SLC20A1 solute carrier family 20 (phosphate transporter), member 1 227425_at 2.0177138 up Hs.186810 REPS2 RALBP1 associated Eps domain containing 2 210471_s_at 2.0186863 up Hs.654519 KCNAB1 potassium voltage-gated channel, shaker-related subfamily,
beta member 1 206332_s_at 2.0190833 up Hs.380250 IFI16 interferon, gamma-inducible protein 16 241681_at 2.020528 up Hs.656858 Transcribed locus 202073_at 2.0241623 up Hs.332706 OPTN optineurin 212509_s_at 2.0265996 up Hs.250723 MXRA7 matrix-remodelling associated 7 217388_s_at 2.0278747 up Hs.470126 KYNU kynureninase (L-kynurenine hydrolase) 1557038_s_at 2.0323508 up Hs.225914 Clone IMAGE:110862, mRNA sequence 204561_x_at 2.0337727 up Hs.655423 APOC2 ///
APOC4 apolipoprotein C-II /// apolipoprotein C-IV
232422_at 2.0343502 up Hs.350868 RP11-151A6.2 hypothetical protein BC004360 231912_s_at 2.0359485 up Hs.632303 DKFZP434B03
35 DKFZP434B0335 protein
202672_s_at 2.0361893 up Hs.460 ATF3 activating transcription factor 3 225173_at 2.040095 up Hs.486458 ARHGAP18 Rho GTPase activating protein 18
237
219654_at 2.0449255 up Hs.114062 PTPLA protein tyrosine phosphatase-like (proline instead of catalytic arginine), member A
207229_at 2.0467234 up Hs.159297 KLRA1 killer cell lectin-like receptor subfamily A, member 1 218376_s_at 2.0478377 up Hs.33476 MICAL1 microtubule associated monoxygenase, calponin and LIM
domain containing 1 230511_at 2.0480478 up Hs.200250 CREM cAMP responsive element modulator 1559240_at 2.0485454 up 220387_s_at 2.049341 up Hs.142245 HHLA3 HERV-H LTR-associating 3 222347_at 2.0498004 up Hs.652926 LOC644450 hypothetical protein LOC644450 201663_s_at 2.0590868 up Hs.58992 SMC4 structural maintenance of chromosomes 4 221841_s_at 2.05947 up Hs.376206 KLF4 Kruppel-like factor 4 (gut) 219157_at 2.059561 up Hs.388668 KLHL2 kelch-like 2, Mayven (Drosophila) 201201_at 2.0599608 up Hs.695 CSTB cystatin B (stefin B) 209921_at 2.0627186 up Hs.390594 SLC7A11 solute carrier family 7, (cationic amino acid transporter, y+
system) member 11 224048_at 2.063752 up Hs.646421 USP44 ubiquitin specific peptidase 44 208944_at 2.0670264 up Hs.82028 TGFBR2 transforming growth factor, beta receptor II (70/80kDa) 204351_at 2.068727 up Hs.2962 S100P S100 calcium binding protein P 208792_s_at 2.0700374 up Hs.436657 CLU clusterin 202833_s_at 2.0704043 up Hs.525557 SERPINA1 serpin peptidase inhibitor, clade A (alpha-1 antiproteinase,
antitrypsin), member 1 225622_at 2.0795598 up Hs.266175 PAG1 phosphoprotein associated with glycosphingolipid
microdomains 1 221190_s_at 2.081298 up Hs.529006 C18orf8 chromosome 18 open reading frame 8 218701_at 2.0813134 up Hs.118554 LACTB2 lactamase, beta 2 228652_at 2.0828218 up Hs.109540 ZNF776 zinc finger protein 776 220194_at 2.0833938 up Hs.590923 NSUN7 NOL1/NOP2/Sun domain family, member 7 219330_at 2.084181 up Hs.515130 VANGL1 vang-like 1 (van gogh, Drosophila) 231842_at 2.084993 up Hs.533953 KIAA1462 KIAA1462 211240_x_at 2.0872247 up Hs.166011 CTNND1 catenin (cadherin-associated protein), delta 1 228370_at 2.0919597 up Hs.564847 SNRPN Small nuclear ribonucleoprotein polypeptide N 210512_s_at 2.0931418 up Hs.73793 VEGFA vascular endothelial growth factor A
238
232664_at 2.0939298 up Hs.289062 FLJ12334 hypothetical gene supported by AK022396; AK097927 215177_s_at 2.0963833 up Hs.133397 ITGA6 integrin, alpha 6 219566_at 2.0967424 up Hs.466383 PLEKHF1 pleckstrin homology domain containing, family F (with
FYVE domain) member 1 209774_x_at 2.0992193 up Hs.590921 CXCL2 chemokine (C-X-C motif) ligand 2 209859_at 2.0999236 up Hs.654750 TRIM9 tripartite motif-containing 9 225389_at 2.1008906 up Hs.7367 BTBD6 BTB (POZ) domain containing 6 205067_at 2.10146 up Hs.126256 IL1B interleukin 1, beta 201124_at 2.1058223 up Hs.536663 ITGB5 integrin, beta 5 209122_at 2.1069572 up Hs.3416 ADFP adipose differentiation-related protein 231164_at 2.1108537 up Hs.376281 ABCA17P ATP-binding cassette, sub-family A (ABC1), member 17
(pseudogene) 1559948_at 2.112178 up Hs.194746 CDNA FLJ20447 fis, clone KAT05276 221398_at 2.1144664 up Hs.533755 TAS2R8 taste receptor, type 2, member 8 209803_s_at 2.122951 up Hs.154036 PHLDA2 pleckstrin homology-like domain, family A, member 2 209014_at 2.1318061 up Hs.5258 MAGED1 melanoma antigen family D, 1 202260_s_at 2.1331933 up Hs.288229 STXBP1 syntaxin binding protein 1 231779_at 2.1337702 up Hs.449207 IRAK2 interleukin-1 receptor-associated kinase 2 218706_s_at 2.13423 up Hs.363558 GRAMD3 GRAM domain containing 3 201700_at 2.1414373 up Hs.534307 CCND3 cyclin D3 207329_at 2.145752 up Hs.161839 MMP8 matrix metallopeptidase 8 (neutrophil collagenase) 227290_at 2.1460938 up Hs.60257 CDNA FLJ13598 fis, clone PLACE1009921 212886_at 2.1466742 up Hs.655336 CCDC69 coiled-coil domain containing 69 1560739_a_at 2.1565313 up Hs.663724 CDNA clone IMAGE:5300703 226909_at 2.1572163 up Hs.455089 ZNF518B zinc finger protein 518B 219489_s_at 2.1590781 up Hs.527989 NXN nucleoredoxin 224832_at 2.1613 up Hs.536535 DUSP16 dual specificity phosphatase 16 205769_at 2.162935 up Hs.11729 SLC27A2 solute carrier family 27 (fatty acid transporter), member 2 203559_s_at 2.1633184 up Hs.647097 ABP1 amiloride binding protein 1 (amine oxidase (copper-
containing) 238727_at 2.1733625 up Hs.635193 CDNA clone IMAGE:5311842 226353_at 2.187416 up Hs.401537 SPPL2A signal peptide peptidase-like 2A
239
212820_at 2.188495 up Hs.511386 DMXL2 Dmx-like 2 230569_at 2.1918945 up Hs.535734 KIAA1430 KIAA1430 221601_s_at 2.192935 up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 1565698_at 2.193966 up Hs.656960 HECTD2 HECT domain containing 2 207111_at 2.1982448 up Hs.2375 EMR1 egf-like module containing, mucin-like, hormone receptor-
like 1 225436_at 2.2078629 up Hs.459072 FAM108C1 family with sequence similarity 108, member C1 217552_x_at 2.208279 up Hs.334019 CR1 complement component (3b/4b) receptor 1 (Knops blood
group) 225685_at 2.2088935 up Hs.696089 CDNA FLJ31353 fis, clone MESAN2000264 243634_at 2.2166958 up Hs.205952 CDNA FLJ26764 fis, clone PRS02668 225078_at 2.228193 up Hs.531561 EMP2 epithelial membrane protein 2 211966_at 2.242716 up Hs.508716 COL4A2 collagen, type IV, alpha 2 215985_at 2.244619 up Hs.653168 C6orf12 HLA complex group 8 225079_at 2.256898 up Hs.531561 EMP2 epithelial membrane protein 2 237246_at 2.2613916 up Hs.670752 Transcribed locus 211964_at 2.2618506 up Hs.508716 COL4A2 collagen, type IV, alpha 2 1569469_a_at 2.2747583 up Hs.403934 LHX8 LIM homeobox 8 202241_at 2.2756593 up Hs.444947 TRIB1 tribbles homolog 1 (Drosophila) 209288_s_at 2.2815948 up Hs.369574 CDC42EP3 CDC42 effector protein (Rho GTPase binding) 3 215016_x_at 2.2828763 up Hs.631992 DST dystonin 223688_s_at 2.286348 up Hs.699597 LY6K lymphocyte antigen 6 complex, locus K 221902_at 2.289728 up Hs.531581 GPR153 G protein-coupled receptor 153 224725_at 2.2921298 up Hs.140903 MIB1 mindbomb homolog 1 (Drosophila) 223343_at 2.2989476 up Hs.530735 MS4A7 membrane-spanning 4-domains, subfamily A, member 7 230493_at 2.3104346 up Hs.433791 SHISA2 shisa homolog 2 (Xenopus laevis) 205222_at 2.3131294 up Hs.429879 EHHADH enoyl-Coenzyme A, hydratase/3-hydroxyacyl Coenzyme A
dehydrogenase 211590_x_at 2.3155444 up Hs.442530 TBXA2R thromboxane A2 receptor 211527_x_at 2.3224564 up Hs.73793 VEGFA vascular endothelial growth factor A 238390_at 2.3403041 up Hs.584762 Transcribed locus 224358_s_at 2.340462 up Hs.530735 MS4A7 membrane-spanning 4-domains, subfamily A, member 7
240
228731_at 2.3407497 up Hs.24321 CDNA clone IMAGE:5273964 205749_at 2.3446243 up Hs.72912 CYP1A1 cytochrome P450, family 1, subfamily A, polypeptide 1 224817_at 2.3510356 up Hs.709448 SH3PXD2A SH3 and PX domains 2A 204232_at 2.3589032 up Hs.433300 FCER1G Fc fragment of IgE, high affinity I, receptor for; gamma
polypeptide 201034_at 2.3661902 up Hs.501012 ADD3 adducin 3 (gamma) 231579_s_at 2.376706 up Hs.633514 TIMP2 TIMP metallopeptidase inhibitor 2 214751_at 2.3812814 up Hs.467223 ZNF468 zinc finger protein 468 1552727_s_at 2.3860312 up Hs.513200 ADAMTS17 ADAM metallopeptidase with thrombospondin type 1 motif,
pyrophosphorylase (carboxylating) 204224_s_at 2.4215453 up Hs.86724 GCH1 GTP cyclohydrolase 1 (dopa-responsive dystonia) 205047_s_at 2.4257452 up Hs.489207 ASNS asparagine synthetase 213836_s_at 2.4561102 up Hs.463964 WIPI1 WD repeat domain, phosphoinositide interacting 1 201325_s_at 2.4683442 up Hs.436298 EMP1 epithelial membrane protein 1 210773_s_at 2.4940493 up Hs.99855 FPR2 formyl peptide receptor 2 200733_s_at 2.4943774 up Hs.227777 PTP4A1 protein tyrosine phosphatase type IVA, member 1 230860_at 2.504897 up Hs.282800 Transcribed locus 209683_at 2.5064533 up Hs.467769 FAM49A family with sequence similarity 49, member A
241
244390_at 2.5098689 up Hs.112791 Transcribed locus 205419_at 2.513219 up Hs.784 EBI2 Epstein-Barr virus induced gene 2 (lymphocyte-specific G
protein-coupled receptor) 200644_at 2.5132656 up Hs.75061 MARCKSL1 MARCKS-like 1 205488_at 2.5173492 up Hs.90708 GZMA granzyme A (granzyme 1, cytotoxic T-lymphocyte-
associated serine esterase 3) 214157_at 2.5319889 up Hs.125898 GNAS GNAS complex locus 1557104_at 2.5332897 up Hs.464896 ZNF397OS Zinc finger protein 397 opposite strand 201753_s_at 2.5353084 up Hs.501012 ADD3 adducin 3 (gamma) 1554899_s_at 2.5357373 up Hs.433300 FCER1G Fc fragment of IgE, high affinity I, receptor for; gamma
polypeptide 231175_at 2.5378351 up Hs.582993 C6orf65 chromosome 6 open reading frame 65 217966_s_at 2.5471883 up Hs.518662 FAM129A family with sequence similarity 129, member A 201631_s_at 2.5666416 up Hs.591785 IER3 immediate early response 3 34210_at 2.5698426 up Hs.276770 CD52 CD52 molecule 242271_at 2.5714705 up Hs.164073 SLC26A9 solute carrier family 26, member 9 219609_at 2.5732338 up Hs.497600 WDR25 WD repeat domain 25 215071_s_at 2.5748265 up Hs.484950 HIST1H2AC histone cluster 1, H2ac 218810_at 2.576255 up Hs.656294 ZC3H12A zinc finger CCCH-type containing 12A 213017_at 2.5767097 up Hs.397978 ABHD3 abhydrolase domain containing 3 227682_at 2.5885766 up Hs.595314 Transcribed locus 225033_at 2.5925033 up Hs.374257 LOC286167 hypothetical LOC286167 209651_at 2.6035943 up Hs.513530 TGFB1I1 transforming growth factor beta 1 induced transcript 1 218847_at 2.6044652 up Hs.35354 IGF2BP2 insulin-like growth factor 2 mRNA binding protein 2 205768_s_at 2.6062658 up Hs.11729 SLC27A2 solute carrier family 27 (fatty acid transporter), member 2 209120_at 2.612503 up Hs.701977 NR2F2 nuclear receptor subfamily 2, group F, member 2 1552578_a_at 2.619047 up Hs.671900 MYO3B myosin IIIB 224735_at 2.6243396 up Hs.22546 CYBASC3 cytochrome b, ascorbate dependent 3 204118_at 2.630754 up Hs.243564 CD48 CD48 molecule 1556658_a_at 2.632578 up Hs.687293 CDNA FLJ36459 fis, clone THYMU2014762 235957_at 2.6423335 up Hs.594436 Transcribed locus 224799_at 2.6737282 up Hs.525093 NDFIP2 Nedd4 family interacting protein 2
242
39402_at 2.6869698 up Hs.126256 IL1B interleukin 1, beta 219209_at 2.6904526 up Hs.163173 IFIH1 interferon induced with helicase C domain 1 203767_s_at 2.6911764 up Hs.522578 STS steroid sulfatase (microsomal), isozyme S 64942_at 2.7049022 up Hs.531581 GPR153 G protein-coupled receptor 153 202890_at 2.705124 up Hs.486548 MAP7 microtubule-associated protein 7 223344_s_at 2.7198555 up Hs.530735 MS4A7 membrane-spanning 4-domains, subfamily A, member 7 205114_s_at 2.7348754 up Hs.514107 CCL3 ///
201464_x_at 2.7501729 up Hs.525704 JUN jun oncogene 206026_s_at 2.7533123 up Hs.437322 TNFAIP6 tumor necrosis factor, alpha-induced protein 6 210538_s_at 2.7620296 up Hs.127799 BIRC3 baculoviral IAP repeat-containing 3 244313_at 2.762246 up Hs.334019 CR1 complement component (3b/4b) receptor 1 (Knops blood
group) 1555832_s_at 2.807976 up Hs.4055///Hs.
709396 KLF6 Homo sapiens, clone IMAGE:4096273, mRNA /// Kruppel-
like factor 6 231603_at 2.8095737 up Hs.112761 RNASE11 ribonuclease, RNase A family, 11 (non-active) 225171_at 2.8266404 up Hs.486458 ARHGAP18 Rho GTPase activating protein 18 1556657_at 2.839919 up Hs.687293 CDNA FLJ36459 fis, clone THYMU2014762 223620_at 2.8479464 up Hs.495989 GPR34 G protein-coupled receptor 34 231688_at 2.8591058 up Hs.557039 Transcribed locus 219596_at 2.8679712 up Hs.591123 THAP10 THAP domain containing 10 203535_at 2.912584 up Hs.112405 S100A9 S100 calcium binding protein A9 211628_x_at 2.9205534 up Hs.453583 FTHP1 ferritin, heavy polypeptide pseudogene 1 1556361_s_at 3.006104 up Hs.105016 ANKRD13C ankyrin repeat domain 13C 233070_at 3.0078309 up Hs.157035 ZNF197 zinc finger protein 197 219434_at 3.0421839 up Hs.283022 TREM1 triggering receptor expressed on myeloid cells 1 217996_at 3.0431302 up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 214211_at 3.0585701 up Hs.524910 FTH1 ferritin, heavy polypeptide 1
243
209789_at 3.065603 up Hs.551213 CORO2B coronin, actin binding protein, 2B 228766_at 3.0702918 up 228155_at 3.082404 up Hs.500333 C10orf58 chromosome 10 open reading frame 58 220765_s_at 3.092851 up Hs.469881 LIMS2 LIM and senescent cell antigen-like domains 2 217967_s_at 3.108988 up Hs.518662 FAM129A family with sequence similarity 129, member A 221909_at 3.1185892 up Hs.437195 RNFT2 ring finger protein, transmembrane 2 218656_s_at 3.135984 up Hs.507798 LHFP lipoma HMGIC fusion partner 202932_at 3.1362047 up Hs.194148 YES1 v-yes-1 Yamaguchi sarcoma viral oncogene homolog 1 208791_at 3.1462173 up Hs.436657 CLU clusterin 225619_at 3.1562297 up Hs.349955 SLAIN1 SLAIN motif family, member 1 219010_at 3.189441 up Hs.518997 C1orf106 chromosome 1 open reading frame 106 201324_at 3.2454588 up Hs.436298 EMP1 epithelial membrane protein 1 201313_at 3.3799746 up Hs.511915 ENO2 enolase 2 (gamma, neuronal) 202988_s_at 3.401592 up Hs.75256 RGS1 regulator of G-protein signaling 1 201005_at 3.4259963 up Hs.114286 CD9 CD9 molecule 218330_s_at 3.434479 up Hs.502116 NAV2 neuron navigator 2 218559_s_at 3.5047252 up Hs.712609 MAFB v-maf musculoaponeurotic fibrosarcoma oncogene homolog
B (avian) 200748_s_at 3.5551684 up Hs.524910 FTH1 ferritin, heavy polypeptide 1 210078_s_at 3.5769308 up Hs.654519 KCNAB1 potassium voltage-gated channel, shaker-related subfamily,
beta member 1 228067_at 3.7344112 up Hs.469398 C2orf55 chromosome 2 open reading frame 55 224802_at 3.7923694 up Hs.525093 NDFIP2 Nedd4 family interacting protein 2 217997_at 3.8278894 up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 240413_at 3.8473117 up Hs.710248 PYHIN1 pyrin and HIN domain family, member 1 202644_s_at 3.9104502 up Hs.591338 TNFAIP3 tumor necrosis factor, alpha-induced protein 3 226218_at 3.9202893 up Hs.635723 IL7R interleukin 7 receptor 209396_s_at 3.9217188 up Hs.382202 CHI3L1 chitinase 3-like 1 (cartilage glycoprotein-39) 202933_s_at 3.9234626 up Hs.194148 YES1 v-yes-1 Yamaguchi sarcoma viral oncogene homolog 1 228384_s_at 3.9629326 up Hs.238303 C10orf33 chromosome 10 open reading frame 33 204967_at 3.964548 up Hs.567236 SHROOM2 shroom family member 2
244
209955_s_at 3.9984534 up Hs.654370 FAP fibroblast activation protein, alpha 1569403_at 4.016248 up Hs.670065 CDNA clone IMAGE:4706427 232504_at 4.0652027 up Hs.604728 LOC285628 hypothetical protein LOC285628 228170_at 4.202808 up Hs.56663 OLIG1 oligodendrocyte transcription factor 1 202643_s_at 4.260881 up Hs.591338 TNFAIP3 tumor necrosis factor, alpha-induced protein 3 206488_s_at 4.404789 up Hs.120949 CD36 CD36 molecule (thrombospondin receptor) 210772_at 4.470887 up Hs.99855 FPR2 formyl peptide receptor 2 228964_at 4.5651217 up Hs.436023 PRDM1 PR domain containing 1, with ZNF domain 204017_at 4.583247 up Hs.709898 KDELR3 KDEL (Lys-Asp-Glu-Leu) endoplasmic reticulum protein
retention receptor 3 201242_s_at 4.622406 up Hs.291196 ATP1B1 ATPase, Na+/K+ transporting, beta 1 polypeptide 209555_s_at 4.6651635 up Hs.120949 CD36 CD36 molecule (thrombospondin receptor) 205943_at 4.768635 up Hs.183671 TDO2 tryptophan 2,3-dioxygenase 211506_s_at 4.9175453 up Hs.551925 IL8 interleukin 8 212062_at 5.0691304 up Hs.709380 ATP9A ATPase, class II, type 9A 202859_x_at 5.133295 up Hs.551925 IL8 interleukin 8 218919_at 5.31763 up Hs.655453 ZFAND1 zinc finger, AN1-type domain 1 205476_at 5.344622 up Hs.75498 CCL20 chemokine (C-C motif) ligand 20 207554_x_at 5.4159017 up Hs.442530 TBXA2R thromboxane A2 receptor 202917_s_at 5.495519 up Hs.416073 S100A8 S100 calcium binding protein A8 228285_at 5.541697 up Hs.21454 TDRD9 tudor domain containing 9 206067_s_at 5.8159995 up Hs.591980 WT1 Wilms tumor 1 210102_at 5.9585633 up Hs.152944 LOH11CR2A loss of heterozygosity, 11, chromosomal region 2, gene A 202887_s_at 6.2075033 up Hs.523012 DDIT4 DNA-damage-inducible transcript 4 209395_at 6.615985 up Hs.382202 CHI3L1 chitinase 3-like 1 (cartilage glycoprotein-39) 203936_s_at 8.026273 up Hs.297413 MMP9 matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase,
92kDa type IV collagenase) 216598_s_at 8.195094 up Hs.303649 CCL2 chemokine (C-C motif) ligand 2 336_at 9.396036 up Hs.442530 TBXA2R thromboxane A2 receptor 216834_at 10.687796 up Hs.75256 RGS1 regulator of G-protein signaling 1 1570561_at 12.199696 up
245
1553614_a_at 12.991128 up Hs.652066 FLJ25694 /// LOC100128202
hypothetical protein FLJ25694 /// hypothetical protein LOC100128202
243489_at 14.485263 up Hs.678608 Transcribed locus 226254_s_at 15.326018 up Hs.535734 KIAA1430 KIAA1430 220517_at 19.309097 up Hs.511668 VPS13C vacuolar protein sorting 13 homolog C (S. cerevisiae) 224296_x_at 24.254719 up 204533_at 27.27933 up Hs.632586 CXCL10 chemokine (C-X-C motif) ligand 10 211690_at 33.311836 up Hs.408073 RPS6 ribosomal protein S6 204475_at 37.07757 up Hs.83169 MMP1 matrix metallopeptidase 1 (interstitial collagenase)
246
Table A3.2: Genes differentially regulated greater than 2-fold in NuMA-RARA compared to control U937-GFP Probe Set ID Fold change([NUMA-
RARA] vs [CONTROL])
Regulation([NUMA-RARA] vs
[CONTROL])
Unigene (Avadis)
Gene Symbol Gene Title
201951_at 2.5140805 down Hs.591293 ALCAM activated leukocyte cell adhesion molecule
201952_at 3.2008994 down Hs.591293 ALCAM activated leukocyte cell adhesion molecule
202364_at 2.015138 down Hs.501023 MXI1 MAX interactor 1 202648_at 2.1071494 down 202925_s_at 2.0869508 down Hs.154104 PLAGL2 pleiomorphic adenoma gene-like 2 202944_at 2.1188123 down Hs.75372 NAGA N-acetylgalactosaminidase, alpha- 202954_at 2.8876154 down Hs.93002 UBE2C ubiquitin-conjugating enzyme E2C 203485_at 2.5145602 down Hs.368626 RTN1 reticulon 1 203523_at 2.1840193 down Hs.56729 LSP1 lymphocyte-specific protein 1 205110_s_at 2.0976357 down Hs.6540 FGF13 fibroblast growth factor 13 205376_at 2.4481084 down Hs.658245 INPP4B inositol polyphosphate-4-
phosphatase, type II, 105kDa 205653_at 3.6304452 down Hs.421724 CTSG cathepsin G 205898_at 2.3489182 down Hs.78913 CX3CR1 chemokine (C-X3-C motif) receptor
1 206278_at 2.6627378 down Hs.709174 PTAFR platelet-activating factor receptor 206589_at 2.0021522 down Hs.73172 GFI1 growth factor independent 1
transcription repressor 206759_at 3.2936876 down Hs.465778 FCER2 Fc fragment of IgE, low affinity II,
receptor for (CD23) 206760_s_at 3.1468818 down Hs.465778 FCER2 Fc fragment of IgE, low affinity II,
receptor for (CD23) 206851_at 2.3055432 down Hs.73839 RNASE3 ribonuclease, RNase A family, 3
(eosinophil cationic protein) 206871_at 2.0622973 down Hs.99863 ELA2 elastase 2, neutrophil 207341_at 2.6777432 down Hs.928 PRTN3 proteinase 3 (serine proteinase,
247
neutrophil, Wegener granulomatosis autoantigen)
207543_s_at 2.1209998 down Hs.500047 P4HA1 procollagen-proline, 2-oxoglutarate 4-dioxygenase (proline 4-hydroxylase), alpha polypeptide I
207808_s_at 2.029618 down Hs.64016 PROS1 protein S (alpha) 207983_s_at 2.0453823 down Hs.496710 STAG2 stromal antigen 2 208937_s_at 2.5915275 down Hs.504609 ID1 inhibitor of DNA binding 1,
dominant negative helix-loop-helix protein
209560_s_at 4.8076034 down Hs.533717 DLK1 delta-like 1 homolog (Drosophila) 210254_at 2.5120962 down Hs.99960 MS4A3 membrane-spanning 4-domains,
subfamily A, member 3 (hematopoietic cell-specific)
211341_at 3.387051 down Hs.654522 LOC100131317 /// POU4F1
POU class 4 homeobox 1 /// similar to hCG1781072
212840_at 2.330931 down Hs.518524 UBXD7 UBX domain containing 7 213293_s_at 2.3205166 down Hs.501778 TRIM22 tripartite motif-containing 22 213350_at 4.1874185 down Hs.433529 RPS11 Ribosomal protein S11 213703_at 2.067904 down Hs.651352 LOC150759 hypothetical protein LOC150759 213826_s_at 2.7210894 down LOC100133109 similar to histone 214575_s_at 2.2711442 down Hs.72885 AZU1 azurocidin 1 (cationic antimicrobial
protein 37) 217100_s_at 2.0867736 down Hs.518524 UBXD7 UBX domain containing 7 217941_s_at 2.0043929 down Hs.591774 ERBB2IP erbb2 interacting protein 218858_at 2.3477619 down Hs.112981 DEPDC6 DEP domain containing 6 218976_at 4.531457 down Hs.260720 DNAJC12 DnaJ (Hsp40) homolog, subfamily
C, member 12 219014_at 2.838491 down Hs.546392 PLAC8 placenta-specific 8 219112_at 2.2908504 down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange
factor (GEF) 6 219295_s_at 2.2198925 down Hs.8944 PCOLCE2 procollagen C-endopeptidase
enhancer 2
248
219410_at 2.0081465 down Hs.658956 TMEM45A transmembrane protein 45A 219697_at 3.588678 down Hs.622536 HS3ST2 heparan sulfate (glucosamine) 3-O-
sulfotransferase 2 219714_s_at 8.406451 down Hs.656687 CACNA2D3 calcium channel, voltage-dependent,
alpha 2/delta subunit 3 221168_at 2.7659276 down Hs.287386 PRDM13 PR domain containing 13 222453_at 2.6440423 down Hs.221941 CYBRD1 cytochrome b reductase 1 223501_at 2.0222945 down Hs.525157 TNFSF13B tumor necrosis factor (ligand)
superfamily, member 13b 223721_s_at 3.2100446 down Hs.260720 DNAJC12 DnaJ (Hsp40) homolog, subfamily
C, member 12 224345_x_at 2.150762 down Hs.584881 C3orf28 chromosome 3 open reading frame
28 224356_x_at 2.171828 down Hs.523702 MS4A6A membrane-spanning 4-domains,
subfamily A, member 6A 224518_s_at 2.5020921 down Hs.655107 ZNF559 zinc finger protein 559 224604_at 2.0568666 down Hs.173705 LOC401152 HCV F-transactivated protein 1 225159_s_at 2.4450445 down 225942_at 2.0562873 down Hs.247460 NLN neurolysin (metallopeptidase M3
family) 225944_at 2.2761114 down Hs.247460 NLN neurolysin (metallopeptidase M3
family) 226352_at 2.0012338 down Hs.482605 JMY junction-mediating and regulatory
protein 226558_at 2.1029706 down LOC653071 similar to CG32820-PA, isoform A 226789_at 2.1677015 down Hs.697682 LOC647121 embigin homolog (mouse)
pseudogene 227198_at 2.2629147 down Hs.444414 AFF3 AF4/FMR2 family, member 3 227663_at 2.3365154 down Hs.664334 CDNA FLJ40901 fis, clone
UTERU2003704 227949_at 2.3743393 down Hs.473218 PHACTR3 phosphatase and actin regulator 3 228055_at 2.2473907 down Hs.636624 NAPSB napsin B aspartic peptidase
pseudogene
249
228623_at 2.0270693 down Hs.374460 Transcribed locus 229461_x_at 2.7272398 down Hs.146542 NEGR1 neuronal growth regulator 1 230078_at 2.5947235 down Hs.483329 RAPGEF6 Rap guanine nucleotide exchange
factor (GEF) 6 230550_at 2.012824 down Hs.523702 MS4A6A membrane-spanning 4-domains,
subfamily A, member 6A 232912_at 2.1449473 down Hs.657472 GPR180 G protein-coupled receptor 180 235046_at 3.150063 down Hs.176376 Transcribed locus 235816_s_at 2.0076482 down Hs.658997 RGL4 ral guanine nucleotide dissociation
stimulator-like 4 236766_at 2.255865 down Hs.624661 Transcribed locus 238902_at 2.02885 down Hs.655574 Transcribed locus 239798_at 2.2176015 down Hs.660359 Transcribed locus 242476_at 2.3789568 down Hs.605126 Transcribed locus 243357_at 2.069857 down Hs.146542 NEGR1 neuronal growth regulator 1 1553194_at 2.3751698 down Hs.146542 NEGR1 neuronal growth regulator 1 1553297_a_at 2.1469352 down Hs.524517 CSF3R colony stimulating factor 3 receptor
(granulocyte) 1554892_a_at 2.212003 down Hs.99960 MS4A3 membrane-spanning 4-domains,
subfamily A, member 3 (hematopoietic cell-specific)
1555339_at 8.438069 down Hs.190334 RAP1A RAP1A, member of RAS oncogene family
1555340_x_at 8.837916 down Hs.190334 RAP1A RAP1A, member of RAS oncogene family
1557370_s_at 2.1244938 down Hs.591221 MYCBP2 MYC binding protein 2 1558290_a_at 2.590345 down Hs.133107 PVT1 Pvt1 oncogene homolog (mouse) 1559391_s_at 2.006825 down Hs.667269 Partial mRNA; ID EE2-8E 200675_at 2.1861107 up Hs.54457 CD81 CD81 molecule 200748_s_at 2.5502985 up Hs.524910 FTH1 ferritin, heavy polypeptide 1 201324_at 4.738356 up Hs.436298 EMP1 epithelial membrane protein 1
250
201325_s_at 2.5761929 up Hs.436298 EMP1 epithelial membrane protein 1 201422_at 2.2819552 up Hs.371344 IFI30 /// PIK3R2 phosphoinositide-3-kinase,
regulatory subunit 2 (beta) /// interferon, gamma-inducible protein 30
201464_x_at 2.4171145 up Hs.525704 JUN jun oncogene 201626_at 2.2931118 up Hs.520819 INSIG1 insulin induced gene 1 201700_at 2.0883832 up Hs.534307 CCND3 cyclin D3 202068_s_at 2.5631108 up Hs.213289 LDLR low density lipoprotein receptor
(familial hypercholesterolemia) 202146_at 2.058797 up Hs.7879 IFRD1 interferon-related developmental
regulator 1 202199_s_at 2.8917031 up Hs.443861 SRPK1 SFRS protein kinase 1 202200_s_at 3.0715544 up Hs.443861 SRPK1 SFRS protein kinase 1 202207_at 2.618532 up Hs.709513 ARL4C ADP-ribosylation factor-like 4C 202241_at 2.4987798 up Hs.444947 TRIB1 tribbles homolog 1 (Drosophila) 202388_at 2.0187805 up Hs.78944 RGS2 regulator of G-protein signaling 2,
24kDa 202440_s_at 3.1627939 up Hs.117715 ST5 suppression of tumorigenicity 5 202627_s_at 2.237239 up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E
(nexin, plasminogen activator inhibitor type 1), member 1
202708_s_at 2.4584754 up Hs.2178 HIST2H2BE histone cluster 2, H2be 202859_x_at 3.3710713 up Hs.551925 IL8 interleukin 8 202887_s_at 3.1278207 up Hs.523012 DDIT4 DNA-damage-inducible transcript 4 202890_at 2.0465117 up Hs.486548 MAP7 microtubule-associated protein 7 202917_s_at 8.607895 up Hs.416073 S100A8 S100 calcium binding protein A8 202932_at 13.4396715 up Hs.194148 YES1 v-yes-1 Yamaguchi sarcoma viral
oncogene homolog 1 202988_s_at 4.1364365 up Hs.75256 RGS1 regulator of G-protein signaling 1 203535_at 4.0339875 up Hs.112405 S100A9 S100 calcium binding protein A9
251
203559_s_at 7.14856 up Hs.647097 ABP1 amiloride binding protein 1 (amine oxidase (copper-containing)
203767_s_at 3.3810694 up Hs.522578 STS steroid sulfatase (microsomal), isozyme S
203935_at 2.1324503 up Hs.470316 ACVR1 activin A receptor, type I 203936_s_at 2.877271 up Hs.297413 MMP9 matrix metallopeptidase 9 (gelatinase
B, 92kDa gelatinase, 92kDa type IV collagenase)
203980_at 2.8758497 up Hs.391561 FABP4 fatty acid binding protein 4, adipocyte
204017_at 2.1915872 up Hs.709898 KDELR3 KDEL (Lys-Asp-Glu-Leu) endoplasmic reticulum protein retention receptor 3
204044_at 2.0071492 up Hs.513484 QPRT quinolinate phosphoribosyltransferase (nicotinate-nucleotide pyrophosphorylase (carboxylating)
204118_at 2.8159168 up Hs.243564 CD48 CD48 molecule 204232_at 2.576512 up Hs.433300 FCER1G Fc fragment of IgE, high affinity I,
receptor for; gamma polypeptide 204533_at 3.6701992 up Hs.632586 CXCL10 chemokine (C-X-C motif) ligand 10 204560_at 3.5530586 up Hs.407190 FKBP5 FK506 binding protein 5 204588_s_at 2.453063 up Hs.513147 SLC7A7 solute carrier family 7 (cationic
amino acid transporter, y+ system), member 7
205222_at 2.1246877 up Hs.429879 EHHADH enoyl-Coenzyme A, hydratase/3-hydroxyacyl Coenzyme A dehydrogenase
205476_at 2.1502717 up Hs.75498 CCL20 chemokine (C-C motif) ligand 20 205541_s_at 2.5884073 up Hs.59523 GSPT2 G1 to S phase transition 2 205749_at 2.0112467 up Hs.72912 CYP1A1 cytochrome P450, family 1,
subfamily A, polypeptide 1 206110_at 3.0578635 up Hs.591778 HIST1H3H histone cluster 1, H3h 206332_s_at 2.1315906 up Hs.380250 IFI16 interferon, gamma-inducible protein
252
16 206584_at 4.9695334 up Hs.660766 LY96 lymphocyte antigen 96 207121_s_at 2.0590255 up Hs.411847 MAPK6 mitogen-activated protein kinase 6 207554_x_at 2.1629765 up Hs.442530 TBXA2R thromboxane A2 receptor 208527_x_at 2.1614985 up Hs.553506 HIST1H2BC ///
218330_s_at 2.0383615 up Hs.502116 NAV2 neuron navigator 2 218559_s_at 4.045437 up Hs.712609 MAFB v-maf musculoaponeurotic
fibrosarcoma oncogene homolog B (avian)
218656_s_at 2.4044502 up Hs.507798 LHFP lipoma HMGIC fusion partner 218793_s_at 2.3413656 up Hs.109655 SCML1 sex comb on midleg-like 1
(Drosophila) 218919_at 5.4285846 up Hs.655453 ZFAND1 zinc finger, AN1-type domain 1 219434_at 2.1268172 up Hs.283022 TREM1 triggering receptor expressed on
myeloid cells 1 219596_at 2.5684 up Hs.591123 THAP10 THAP domain containing 10 219607_s_at 8.440766 up Hs.325960 MS4A4A membrane-spanning 4-domains,
255
subfamily A, member 4 219890_at 2.2654479 up Hs.446235 CLEC5A C-type lectin domain family 5,
member A 220255_at 3.238937 up Hs.302003 FANCE Fanconi anemia, complementation
group E 220517_at 14.522511 up Hs.511668 VPS13C vacuolar protein sorting 13 homolog
C (S. cerevisiae) 221561_at 2.9482198 up Hs.496383 SOAT1 sterol O-acyltransferase (acyl-
Coenzyme A: cholesterol acyltransferase) 1
221750_at 2.4851115 up Hs.397729 HMGCS1 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 1 (soluble)
221909_at 2.675337 up Hs.437195 RNFT2 ring finger protein, transmembrane 2 222067_x_at 2.3068116 up Hs.591797 HIST1H2BD histone cluster 1, H2bd 336_at 3.0008388 up Hs.442530 TBXA2R thromboxane A2 receptor 34210_at 3.1154716 up Hs.276770 CD52 CD52 molecule 36711_at 2.097861 up Hs.517617 MAFF v-maf musculoaponeurotic
fibrosarcoma oncogene homolog F (avian)
37152_at 2.9896212 up Hs.696032 PPARD peroxisome proliferator-activated receptor delta
222557_at 2.0130768 up Hs.639609 STMN3 stathmin-like 3 223278_at 2.7869732 up Hs.591234 GJB2 gap junction protein, beta 2, 26kDa 223620_at 4.809363 up Hs.495989 GPR34 G protein-coupled receptor 34 223939_at 2.6680608 up Hs.279575 SUCNR1 succinate receptor 1 224048_at 3.2531075 up Hs.646421 USP44 ubiquitin specific peptidase 44 224685_at 3.113241 up Hs.709178 MLLT4 myeloid/lymphoid or mixed-lineage
leukemia (trithorax homolog, Drosophila); translocated to, 4
224802_at 2.4919844 up Hs.525093 NDFIP2 Nedd4 family interacting protein 2 224840_at 3.2165444 up Hs.407190 FKBP5 FK506 binding protein 5 224856_at 3.6593025 up Hs.407190 FKBP5 FK506 binding protein 5 225167_at 2.02882 up Hs.330463 FRMD4A FERM domain containing 4A
256
225524_at 2.2465055 up Hs.162963 ANTXR2 anthrax toxin receptor 2 225842_at 2.138169 up Hs.602085 PHLDA1 pleckstrin homology-like domain,
family A, member 1 226218_at 2.4769416 up Hs.635723 IL7R interleukin 7 receptor 226254_s_at 14.906071 up Hs.535734 KIAA1430 KIAA1430 226545_at 2.3461847 up Hs.399891 CD109 CD109 molecule 226771_at 2.0420542 up Hs.435700 ATP8B2 ATPase, class I, type 8B, member 2 227492_at 2.5360022 up Hs.654872 Clone HLS_IMAGE_1881469
mRNA sequence 227682_at 2.3687131 up Hs.595314 Transcribed locus 228155_at 2.0698576 up Hs.500333 C10orf58 chromosome 10 open reading frame
58 228170_at 2.4644752 up Hs.56663 OLIG1 oligodendrocyte transcription factor
1 228285_at 2.9728603 up Hs.21454 TDRD9 tudor domain containing 9 228384_s_at 2.3169546 up Hs.238303 C10orf33 chromosome 10 open reading frame
33 228478_at 2.1306157 up Hs.167619 Full length insert cDNA YH99G08 228964_at 5.8129516 up Hs.436023 PRDM1 PR domain containing 1, with ZNF
domain 229390_at 9.867984 up Hs.381220 FAM26F family with sequence similarity 26,
member F 229391_s_at 2.0939236 up Hs.381220 FAM26F family with sequence similarity 26,
member F 229625_at 2.5347552 up Hs.513726 GBP5 guanylate binding protein 5 230333_at 2.0256205 up Hs.656630 Transcribed locus 231406_at 2.2547612 up Hs.658598 MRNA; cDNA DKFZp667G1412
(from clone DKFZp667G1412) 231982_at 2.531045 up Hs.130714 LOC284422 similar to HSPC323 232035_at 2.2233171 up Hs.278483 HIST1H4A ///
205016_at 2.6648026 Up Hs.170009 TGFA transforming growth factor, alpha 205027_s_at 3.5026836 Up Hs.432453 MAP3K8 mitogen-activated protein kinase kinase kinase 8 205067_at 2.2558508 Up Hs.126256 IL1B interleukin 1, beta 205127_at 2.4086578 Up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H synthase
and cyclooxygenase) 205128_x_at 2.2821105 Up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H synthase
and cyclooxygenase) 205232_s_at 2.1212788 Up Hs.477083 PAFAH2 platelet-activating factor acetylhydrolase 2, 40kDa 205233_s_at 2.3652747 Up Hs.477083 PAFAH2 platelet-activating factor acetylhydrolase 2, 40kDa 205396_at 2.072369 Up Hs.618504 SMAD3 SMAD family member 3 205397_x_at 2.1071382 Up Hs.618504 SMAD3 SMAD family member 3 205398_s_at 2.5249112 Up Hs.618504 SMAD3 SMAD family member 3 205476_at 2.5043466 Up Hs.75498 CCL20 chemokine (C-C motif) ligand 20 205633_s_at 2.518799 Up Hs.476308 ALAS1 aminolevulinate, delta-, synthase 1 205692_s_at 5.818328 Up Hs.479214 CD38 CD38 molecule 205749_at 4.7893267 Up Hs.72912 CYP1A1 cytochrome P450, family 1, subfamily A, polypeptide 1 205780_at 2.4141812 Up Hs.475055 BIK BCL2-interacting killer (apoptosis-inducing) 205789_at 2.8860488 Up Hs.1799 CD1D CD1d molecule 206028_s_at 3.639326 Up Hs.306178 MERTK c-mer proto-oncogene tyrosine kinase 206126_at 4.5504775 Up Hs.113916 CXCR5 chemokine (C-X-C motif) receptor 5 206369_s_at 5.875969 Up Hs.32942 PIK3CG phosphoinositide-3-kinase, catalytic, gamma polypeptide 206370_at 5.189663 Up Hs.32942 PIK3CG phosphoinositide-3-kinase, catalytic, gamma polypeptide 206462_s_at 2.3021586 Up Hs.410969 NTRK3 neurotrophic tyrosine kinase, receptor, type 3 206472_s_at 4.021034 Up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog, Drosophila) 206513_at 2.1365075 Up Hs.281898 AIM2 absent in melanoma 2 206521_s_at 2.0000608 Up Hs.592334 GTF2A1 general transcription factor IIA, 1, 19/37kDa 206873_at 2.6792235 Up Hs.100322 CA6 carbonic anhydrase VI 207606_s_at 2.6671817 Up Hs.499264 ARHGAP12 Rho GTPase activating protein 12 207629_s_at 2.173109 Up Hs.655209 ARHGEF2 rho/rac guanine nucleotide exchange factor (GEF) 2 207765_s_at 2.2349226 Up Hs.301696 KIAA1539 KIAA1539
261
208092_s_at 2.0390315 Up Hs.467769 FAM49A family with sequence similarity 49, member A 208170_s_at 2.9086053 Up Hs.493275 TRIM31 tripartite motif-containing 31 208438_s_at 7.7604666 Up Hs.1422 FGR Gardner-Rasheed feline sarcoma viral (v-fgr) oncogene homolog 208514_at 3.2818332 Up Hs.121495 KCNE1 potassium voltage-gated channel, Isk-related family, member 1 208614_s_at 2.1847918 Up Hs.476448 FLNB filamin B, beta (actin binding protein 278) 208749_x_at 2.103332 Up Hs.179986 FLOT1 flotillin 1 208937_s_at 47.04182 Up Hs.504609 ID1 inhibitor of DNA binding 1, dominant negative helix-loop-helix
protein 209324_s_at 2.2390127 Up Hs.413297 RGS16 regulator of G-protein signaling 16 209325_s_at 2.7166984 Up Hs.413297 RGS16 regulator of G-protein signaling 16 209333_at 2.0310307 Up Hs.47061 ULK1 unc-51-like kinase 1 (C. elegans) 209355_s_at 2.0968838 Up Hs.405156 PPAP2B phosphatidic acid phosphatase type 2B 209392_at 9.531024 Up Hs.190977 ENPP2 ectonucleotide pyrophosphatase/phosphodiesterase 2 (autotaxin) 209435_s_at 2.9095101 Up Hs.655209 ARHGEF2 rho/rac guanine nucleotide exchange factor (GEF) 2 209551_at 2.013789 Up Hs.468099 YIPF4 Yip1 domain family, member 4 209706_at 2.5280802 Up Hs.55999 NKX3-1 NK3 homeobox 1 210142_x_at 2.2628121 Up Hs.179986 FLOT1 flotillin 1 210146_x_at 3.1460853 Up Hs.655652 LILRB2 leukocyte immunoglobulin-like receptor, subfamily B (with TM and
ITIM domains), member 2 210159_s_at 2.1420844 Up Hs.493275 TRIM31 tripartite motif-containing 31 210173_at 2.049891 Up Hs.318547 PTPRJ protein tyrosine phosphatase, receptor type, J 210223_s_at 2.3806376 Up Hs.101840 MR1 major histocompatibility complex, class I-related 210240_s_at 2.2366748 Up Hs.435051 CDKN2D cyclin-dependent kinase inhibitor 2D (p19, inhibits CDK4) 210264_at 4.0205555 Up Hs.239891 GPR35 G protein-coupled receptor 35 210357_s_at 4.3116674 Up Hs.433337 SMOX spermine oxidase 210538_s_at 2.6726468 Up Hs.127799 BIRC3 baculoviral IAP repeat-containing 3 210592_s_at 2.1652398 Up Hs.28491 SAT1 spermidine/spermine N1-acetyltransferase 1 210754_s_at 2.200544 Up Hs.699154 LYN v-yes-1 Yamaguchi sarcoma viral related oncogene homolog 210839_s_at 4.8159866 Up Hs.190977 ENPP2 ectonucleotide pyrophosphatase/phosphodiesterase 2 (autotaxin) 210845_s_at 2.1698377 Up Hs.466871 PLAUR plasminogen activator, urokinase receptor 211026_s_at 2.2317677 Up Hs.277035 MGLL monoglyceride lipase
262
211559_s_at 2.4680865 Up Hs.13291 CCNG2 cyclin G2 211732_x_at 2.0705788 Up Hs.42151 HNMT histamine N-methyltransferase 211913_s_at 3.152666 Up Hs.306178 MERTK c-mer proto-oncogene tyrosine kinase 211964_at 2.5366511 Up Hs.508716 COL4A2 collagen, type IV, alpha 2 211966_at 2.676469 Up Hs.508716 COL4A2 collagen, type IV, alpha 2 212230_at 2.0011234 Up Hs.708050 PPAP2B phosphatidic acid phosphatase type 2B 212419_at 2.4394522 Up Hs.523080 C10orf56 chromosome 10 open reading frame 56 212423_at 2.4178545 Up Hs.523080 C10orf56 chromosome 10 open reading frame 56 212501_at 3.8561795 Up Hs.517106 CEBPB CCAAT/enhancer binding protein (C/EBP), beta 212558_at 2.2629871 Up Hs.436944 SPRY1 sprouty homolog 1, antagonist of FGF signaling (Drosophila) 212769_at 3.297391 Up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog, Drosophila) 212770_at 3.15906 Up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog, Drosophila) 212944_at 3.1856778 Up Hs.302742 SLC5A3 solute carrier family 5 (inositol transporters), member 3 213164_at 2.2114139 Up Hs.302742 SLC5A3 solute carrier family 5 (inositol transporters), member 3 213167_s_at 2.1843853 Up Hs.302742 SLC5A3 solute carrier family 5 (inositol transporters), member 3 213385_at 2.1894703 Up Hs.654611 CHN2 chimerin (chimaerin) 2 213475_s_at 2.1780634 Up Hs.174103 ITGAL integrin, alpha L (antigen CD11A (p180), lymphocyte function-
associated antigen 1; alpha polypeptide) 213810_s_at 2.3184736 Up Hs.485915 C6orf166 Chromosome 6 open reading frame 166 213839_at 2.1172595 Up Hs.593760 KIAA0500 KIAA0500 protein 213926_s_at 2.2072005 Up Hs.591619 HRB HIV-1 Rev binding protein 214084_x_at 5.2366743 Up LOC648998 similar to Neutrophil cytosol factor 1 (NCF-1) (Neutrophil NADPH
214101_s_at 2.1747372 Up Hs.708179 Transcribed locus 214196_s_at 2.0795708 Up Hs.523454 TPP1 tripeptidyl peptidase I 214523_at 3.5253074 Up Hs.558308 CEBPE CCAAT/enhancer binding protein (C/EBP), epsilon 214599_at 2.1587114 Up Hs.516439 IVL involucrin 214637_at 2.5485187 Up Hs.248156 OSM oncostatin M 214639_s_at 3.3741236 Up Hs.67397 HOXA1 homeobox A1 214724_at 5.7931156 Up Hs.655626 DIXDC1 DIX domain containing 1
263
214753_at 2.0603442 Up Hs.507680 N4BP2L2 NEDD4 binding protein 2-like 2 214977_at 2.8935628 Up Hs.654670 CDNA FLJ13790 fis, clone THYRO1000026 215177_s_at 2.9345682 Up Hs.133397 ITGA6 integrin, alpha 6 215268_at 2.1503754 Up Hs.658760 KIAA0754 hypothetical LOC643314 215342_s_at 2.0452404 Up Hs.585378 RABGAP1L RAB GTPase activating protein 1-like 215444_s_at 2.4159238 Up Hs.493275 TRIM31 tripartite motif-containing 31 215627_at 2.0265882 Up Hs.301715 CDNA FLJ13453 fis, clone PLACE1003205 215813_s_at 2.1931105 Up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H synthase
and cyclooxygenase) 216587_s_at 2.2156923 Up Hs.302634 FZD8 frizzled homolog 8 (Drosophila) 217104_at 2.0132565 Up Hs.459049 MTHFS ///
ST20 5,10-methenyltetrahydrofolate synthetase (5-formyltetrahydrofolate cyclo-ligase) /// suppressor of tumorigenicity 20
217486_s_at 2.0104165 Up Hs.4014 ZDHHC17 zinc finger, DHHC-type containing 17 217591_at 2.0515206 Up Hs.677805 Transcribed locus 217897_at 2.0845242 Up Hs.635508 FXYD6 FXYD domain containing ion transport regulator 6 217996_at 2.4030678 Up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 217997_at 2.4917169 Up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 218284_at 2.8421624 Up Hs.618504 SMAD3 SMAD family member 3 218501_at 5.894343 Up Hs.476402 ARHGEF3 Rho guanine nucleotide exchange factor (GEF) 3 218510_x_at 2.72195 Up Hs.481704 FAM134B family with sequence similarity 134, member B 218532_s_at 3.3436737 Up Hs.481704 FAM134B family with sequence similarity 134, member B 218627_at 3.6088104 Up Hs.525634 DRAM damage-regulated autophagy modulator 218676_s_at 2.2792597 Up Hs.285218 PCTP phosphatidylcholine transfer protein 218723_s_at 2.0949621 Up Hs.507866 C13orf15 chromosome 13 open reading frame 15 219010_at 3.37471 Up Hs.518997 C1orf106 chromosome 1 open reading frame 106 219317_at 2.2760723 Up Hs.438533 POLI polymerase (DNA directed) iota 219344_at 3.5494576 Up Hs.438419 SLC29A3 solute carrier family 29 (nucleoside transporters), member 3 219434_at 2.0415852 Up Hs.283022 TREM1 triggering receptor expressed on myeloid cells 1 219607_s_at 13.902484 Up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 219797_at 2.1064587 Up Hs.177576 MGAT4A mannosyl (alpha-1,3-)-glycoprotein beta-1,4-N-
acetylglucosaminyltransferase, isozyme A 219844_at 2.1877847 Up Hs.159066 C10orf118 chromosome 10 open reading frame 118
264
219901_at 2.3943903 Up Hs.506381 FGD6 FYVE, RhoGEF and PH domain containing 6 219994_at 4.150032 Up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B, member 1
interacting protein 220507_s_at 2.2128952 Up Hs.474388 UPB1 ureidopropionase, beta 220682_s_at 2.1781907 Up 220802_at 2.0693676 Up Hs.304081 KCNH4 potassium voltage-gated channel, subfamily H (eag-related), member
4 221142_s_at 2.0912616 Up Hs.281680 PECR peroxisomal trans-2-enoyl-CoA reductase 221249_s_at 3.0015697 Up Hs.514308 FAM117A family with sequence similarity 117, member A 221266_s_at 2.1035686 Up Hs.652230 TM7SF4 transmembrane 7 superfamily member 4 221276_s_at 2.1622188 Up Hs.712631 SYNC1 syncoilin, intermediate filament 1 221345_at 3.9440105 Up Hs.248056 FFAR2 free fatty acid receptor 2 221485_at 2.4908564 Up Hs.370487 B4GALT5 UDP-Gal:betaGlcNAc beta 1,4- galactosyltransferase, polypeptide 5 221561_at 2.2119563 Up Hs.496383 SOAT1 sterol O-acyltransferase (acyl-Coenzyme A: cholesterol
acyltransferase) 1 221601_s_at 3.0838854 Up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 221602_s_at 2.5654364 Up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 221986_s_at 3.6803215 Up Hs.407709 KLHL24 kelch-like 24 (Drosophila) 34210_at 2.0480247 Up Hs.276770 CD52 CD52 molecule 36711_at 4.527568 Up Hs.517617 MAFF v-maf musculoaponeurotic fibrosarcoma oncogene homolog F (avian) 37117_at 2.015902 Up Hs.102336 ARHGAP8 ///
37152_at 2.103844 Up Hs.696032 PPARD peroxisome proliferator-activated receptor delta 37170_at 2.0495327 Up Hs.146551 BMP2K BMP2 inducible kinase 222844_s_at 2.030162 Up Hs.461954 SRR serine racemase 222877_at 2.2359216 Up Hs.660596 CDNA: FLJ21027 fis, clone CAE07110 223044_at 2.2080986 Up Hs.643005 SLC40A1 solute carrier family 40 (iron-regulated transporter), member 1 223276_at 3.9754796 Up Hs.29444 MST150 MSTP150 223567_at 2.9676416 Up Hs.465642 SEMA6B sema domain, transmembrane domain (TM), and cytoplasmic domain,
(semaphorin) 6B 223620_at 2.2369268 Up Hs.495989 GPR34 G protein-coupled receptor 34
265
223634_at 2.1102068 Up Hs.474711 RASD2 RASD family, member 2 223672_at 2.7002747 Up Hs.132121 SGIP1 SH3-domain GRB2-like (endophilin) interacting protein 1 223879_s_at 2.1745186 Up Hs.148778 OXR1 oxidation resistance 1 224035_s_at 2.0562057 Up Hs.708077 BCL2L13 BCL2-like 13 (apoptosis facilitator) 224043_s_at 2.271343 Up Hs.474388 UPB1 ureidopropionase, beta 224341_x_at 2.04793 Up Hs.174312 TLR4 toll-like receptor 4 224357_s_at 2.7210784 Up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 224534_at 2.121217 Up Hs.229335 KREMEN1 kringle containing transmembrane protein 1 224916_at 3.251452 Up Hs.379754 TMEM173 transmembrane protein 173 224917_at 2.088817 Up MIRN21 microRNA 21 224929_at 3.2078555 Up Hs.379754 TMEM173 transmembrane protein 173 224964_s_at 4.348477 Up Hs.187772 GNG2 guanine nucleotide binding protein (G protein), gamma 2 225102_at 2.2773845 Up Hs.277035 MGLL monoglyceride lipase 225347_at 3.602254 Up Hs.497399 ARL8A ADP-ribosylation factor-like 8A 225372_at 3.7230558 Up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225373_at 2.3732738 Up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225763_at 3.5019486 Up Hs.493867 RCSD1 RCSD domain containing 1 225803_at 2.6384509 Up Hs.403933 FBXO32 F-box protein 32 225817_at 2.0570107 Up Hs.148989 CGNL1 cingulin-like 1 225842_at 2.6239111 Up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 225899_x_at 2.000697 Up Hs.465593 LOC643670 ///
LOC728105 /// tcag7.907
hypothetical LOC402483 /// similar to hCG1739109
225919_s_at 3.3289592 Up Hs.493639 C9orf72 chromosome 9 open reading frame 72 226039_at 2.476476 Up Hs.177576 MGAT4A mannosyl (alpha-1,3-)-glycoprotein beta-1,4-N-
acetylglucosaminyltransferase, isozyme A 226099_at 2.2057993 Up Hs.592742 ELL2 elongation factor, RNA polymerase II, 2 226119_at 2.006614 Up Hs.308480 PCMTD1 protein-L-isoaspartate (D-aspartate) O-methyltransferase domain
containing 1 226146_at 2.0748007 Up Hs.655606 CDNA clone IMAGE:5294560 226184_at 3.1302419 Up Hs.654630 FMNL2 formin-like 2 226344_at 2.0736291 Up Hs.496512 ZMAT1 zinc finger, matrin type 1
266
226487_at 2.3282282 Up Hs.661785 C12orf34 chromosome 12 open reading frame 34 226722_at 2.8734744 Up Hs.134742 FAM20C family with sequence similarity 20, member C 226756_at 3.5177755 Up Hs.633903 CDNA FLJ25556 fis, clone JTH02629 226799_at 2.0092845 Up Hs.506381 FGD6 FYVE, RhoGEF and PH domain containing 6 226853_at 2.0511706 Up Hs.146551 BMP2K BMP2 inducible kinase 226965_at 2.0470502 Up Hs.91085 FAM116A family with sequence similarity 116, member A 227040_at 2.7269776 Up Hs.507783 NHLRC3 NHL repeat containing 3 227396_at 2.9585493 Up Hs.318547 PTPRJ protein tyrosine phosphatase, receptor type, J 227484_at 2.5147696 Up Hs.710097 CDNA FLJ41690 fis, clone HCASM2009405 227514_at 3.208757 Up Hs.648523 LOC162073 hypothetical protein LOC162073 227792_at 3.1586435 Up Hs.648523 LOC162073 hypothetical protein LOC162073 227915_at 3.1987798 Up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 228083_at 2.293615 Up Hs.13768 CACNA2D4 calcium channel, voltage-dependent, alpha 2/delta subunit 4 228097_at 2.274039 Up Hs.484738 MYLIP myosin regulatory light chain interacting protein 228098_s_at 2.203799 Up Hs.484738 MYLIP myosin regulatory light chain interacting protein 228340_at 3.3667622 Up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog, Drosophila) 228361_at 2.824649 Up Hs.194333 E2F2 E2F transcription factor 2 228461_at 2.5984275 Up Hs.535157 SH3MD4 SH3 multiple domains 4 228642_at 2.3802812 Up Hs.445239 CDNA FLJ12777 fis, clone NT2RP2001720 228648_at 3.6471891 Up Hs.655559 LRG1 leucine-rich alpha-2-glycoprotein 1 228772_at 2.2901244 Up Hs.42151 HNMT histamine N-methyltransferase 229278_at 2.1170728 Up Hs.40061 Transcribed locus 229285_at 2.2954693 Up Hs.518545 RNASEL ribonuclease L (2',5'-oligoisoadenylate synthetase-dependent) 229521_at 2.6173232 Up Hs.29692 FLJ36031 hypothetical protein FLJ36031 229625_at 3.3724356 Up Hs.513726 GBP5 guanylate binding protein 5 229670_at 4.3125396 Up Hs.180284 5.5 kb mRNA upregulated in retinoic acid treated HL-60 neutrophilic
cells 229735_s_at 2.1004102 Up Hs.708897 Transcribed locus 229910_at 2.0621803 Up Hs.591481 SHE Src homology 2 domain containing E 229934_at 2.8347294 Up Hs.38218 Mir-223 transcript variant 1 mRNA, complete sequence 229971_at 2.1540885 Up Hs.187884 GPR114 G protein-coupled receptor 114
267
230170_at 3.5915139 Up Hs.248156 OSM oncostatin M 230179_at 2.1777985 Up Hs.593631 LOC285812 hypothetical protein LOC285812 230218_at 2.3413222 Up Hs.72956 HIC1 hypermethylated in cancer 1 230333_at 2.6759667 Up Hs.656630 Transcribed locus 230536_at 2.658517 Up Hs.466257 PBX4 pre-B-cell leukemia homeobox 4 230646_at 2.16772 Up Hs.524234 FNDC5 Fibronectin type III domain containing 5 230732_s_at 2.5513582 Up Hs.433728 MAPK4 Mitogen-activated protein kinase 4 230800_at 2.0529864 Up Hs.443428 ADCY4 adenylate cyclase 4 230925_at 5.8982406 Up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B, member 1
interacting protein 231214_at 2.0496953 Up Hs.21278 Transcribed locus 231283_at 2.0643651 Up Hs.177576 MGAT4A mannosyl (alpha-1,3-)-glycoprotein beta-1,4-N-
acetylglucosaminyltransferase, isozyme A 231496_at 3.1408207 Up Hs.145519 FCAMR Fc receptor, IgA, IgM, high affinity 231779_at 2.4146137 Up Hs.449207 IRAK2 interleukin-1 receptor-associated kinase 2 231969_at 2.6357288 Up Hs.21958 STOX2 storkhead box 2 231991_at 3.178446 Up Hs.382151 C20orf160 chromosome 20 open reading frame 160 232068_s_at 2.1511958 Up Hs.174312 TLR4 toll-like receptor 4 232253_at 2.0920262 Up Hs.658288 LOC441108 Hypothetical gene supported by AK128882 232306_at 2.1834612 Up Hs.54973 CDH26 cadherin-like 26 232611_at 2.1045644 Up Hs.645240 LOC92497 hypothetical LOC92497 232687_at 2.2366445 Up Hs.100912 CDNA FLJ33091 fis, clone TRACH2000660 232754_at 2.6992195 Up Hs.664518 CDNA FLJ11563 fis, clone HEMBA1003202 232861_at 2.009733 Up Hs.654693 PDP2 pyruvate dehydrogenase phosphatase isoenzyme 2 233009_at 2.013952 Up Hs.675113 CDNA FLJ14271 fis, clone PLACE1004686 233016_at 2.54932 Up Hs.288478 CDNA clone IMAGE:4817097 233857_s_at 3.3474777 Up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 233888_s_at 2.279395 Up Hs.450763 SRGAP1 SLIT-ROBO Rho GTPase activating protein 1 234317_s_at 2.0457516 Up Hs.21958 STOX2 storkhead box 2 234361_at 2.9642448 Up Hs.247744 CREB3L3 cAMP responsive element binding protein 3-like 3 234987_at 2.6623209 Up Hs.660221 Transcribed locus
268
234990_at 2.1560304 Up Hs.349283 CDNA clone IMAGE:4842353 235352_at 2.2367399 Up Hs.13500 CDNA FLJ31593 fis, clone NT2RI2002481 235417_at 2.0219183 Up Hs.62604 SPOCD1 SPOC domain containing 1 235421_at 2.3143904 Up Hs.432453///
IMAGE:4689481 235505_s_at 2.009543 Up Hs.40966 MRNA full length insert cDNA clone EUROIMAGE 2362292 235516_at 2.0422332 Up Hs.253305 SEPSECS Sep (O-phosphoserine) tRNA:Sec (selenocysteine) tRNA synthase 235521_at 4.8845115 Up Hs.659337 HOXA3 homeobox A3 236019_at 2.03722 Up Hs.270074 RAB12 RAB12, member RAS oncogene family 236061_at 2.3027232 Up Hs.711654 PRDM15 PR domain containing 15 236191_at 6.613427 Up Hs.667427 Transcribed locus 236223_s_at 2.0239542 Up Hs.491234 RIT1 Ras-like without CAAX 1 236407_at 5.1995955 Up Hs.121495 KCNE1 potassium voltage-gated channel, Isk-related family, member 1 236480_at 2.4092677 Up Hs.446388 CDNA FLJ41489 fis, clone BRTHA2004582 236699_at 2.4069755 Up Hs.660409 CDNA FLJ90129 fis, clone HEMBB1000309 236717_at 4.124915 Up Hs.525977 LOC165186 similar to RIKEN cDNA 4632412N22 gene 237252_at 2.5999637 Up Hs.2030 THBD thrombomodulin 237338_at 2.2278228 Up Hs.441681 B3GNT8 UDP-GlcNAc:betaGal beta-1,3-N-acetylglucosaminyltransferase 8 237442_at 3.7524705 Up 237458_at 2.3744226 Up Hs.547058 Transcribed locus 237701_at 2.1291585 Up Hs.98202 C12orf54 chromosome 12 open reading frame 54 238032_at 7.6221538 Up Hs.655631 Transcribed locus 238407_at 2.0112407 Up Hs.624137 Transcribed locus 238439_at 3.0459437 Up Hs.217484 ANKRD22 ankyrin repeat domain 22 238493_at 2.8231044 Up Hs.351906 ZNF506 zinc finger protein 506 238581_at 4.4943924 Up Hs.513726 GBP5 guanylate binding protein 5 238669_at 2.5256748 Up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H synthase
and cyclooxygenase) 238725_at 2.0562582 Up Hs.661929 Transcribed locus 239294_at 7.0116982 Up Hs.561747 Transcribed locus 241036_at 2.156731 Up
269
241394_at 2.3390722 Up Hs.464224 LOC284120 hypothetical LOC284120 242426_at 4.1953063 Up Hs.696574 NRG4 neuregulin 4 242525_at 3.1685297 Up Hs.439122 Transcribed locus 242705_x_at 2.3846107 Up Hs.592928 Full length insert cDNA clone YT86E01 242715_at 2.4447923 Up Hs.444181 Transcribed locus 243541_at 3.4468963 Up Hs.55378 IL31RA interleukin 31 receptor A 243798_at 2.1048503 Up Hs.674680 Transcribed locus 243819_at 2.0423958 Up 244665_at 3.9024503 Up Hs.668855 Transcribed locus 1552634_a_at 2.0228066 Up Hs.631642 ZNF101 zinc finger protein 101 1553133_at 2.7681775 Up Hs.493639 C9orf72 chromosome 9 open reading frame 72 1553313_s_at 2.0301917 Up Hs.302742 SLC5A3 solute carrier family 5 (inositol transporters), member 3 1553810_a_at 2.3159645 Up Hs.591308 KIAA1524 KIAA1524 1553970_s_at 2.0439599 Up Hs.533258 CEL carboxyl ester lipase (bile salt-stimulated lipase) 1554015_a_at 2.210065 Up Hs.220864 CHD2 chromodomain helicase DNA binding protein 2 1554240_a_at 2.0033767 Up Hs.174103 ITGAL integrin, alpha L (antigen CD11A (p180), lymphocyte function-
associated antigen 1; alpha polypeptide) 1554806_a_at 2.4902866 Up Hs.76917 FBXO8 F-box protein 8 1555137_a_at 2.5319905 Up Hs.506381 FGD6 FYVE, RhoGEF and PH domain containing 6 1555420_a_at 2.1739933 Up Hs.471221 KLF7 Kruppel-like factor 7 (ubiquitous) 1555431_a_at 3.4255886 Up Hs.55378 IL31RA interleukin 31 receptor A 1555680_a_at 4.0508475 Up Hs.433337 SMOX spermine oxidase 1555728_a_at 11.558348 Up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 1556123_a_at 2.2488248 Up Hs.637720 Homo sapiens, clone IMAGE:4863312, mRNA 1563745_a_at 4.679765 Up Hs.309176 LOC283050 hypothetical LOC283050 1556206_at 2.0964098 Up Hs.549878 CDNA clone IMAGE:4830861 1557545_s_at 2.2106662 Up Hs.501114 RNF165 ring finger protein 165 1559584_a_at 2.5446959 Up Hs.331095 C16orf54 ///
hCG_1644884 chromosome 16 open reading frame 54 /// similar to chromosome 16 open reading frame 54
1562100_at 2.1443887 Up Hs.552946 Homo sapiens, clone IMAGE:4045462, mRNA 1568619_s_at 3.5794806 Up Hs.648523 LOC162073 hypothetical protein LOC162073
270
1569334_at 2.5108023 Up Hs.24553 STRA6 stimulated by retinoic acid gene 6 homolog (mouse) 1569942_at 2.83881 Up Hs.385753 CDNA clone IMAGE:4796629 1570375_at 2.6354275 Up Hs.661265 CDNA FLJ41985 fis, clone SPLEN2014946 201565_s_at 2.500228 Down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix
protein 201566_x_at 2.1079838 Down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix
protein 201675_at 2.1132033 Down Hs.463506 AKAP1 A kinase (PRKA) anchor protein 1 202045_s_at 2.0621004 Down Hs.509447 GRLF1 glucocorticoid receptor DNA binding factor 1 202431_s_at 4.8014007 Down Hs.202453 MYC v-myc myelocytomatosis viral oncogene homolog (avian) 202760_s_at 2.0902064 Down Hs.591908 PALM2-
AKAP2 PALM2-AKAP2
202800_at 2.4726875 Down Hs.481918 SLC1A3 solute carrier family 1 (glial high affinity glutamate transporter), member 3
203023_at 2.1263835 Down Hs.696283 HSPC111 hypothetical protein HSPC111 203231_s_at 2.000628 Down Hs.434961 ATXN1 ataxin 1 203304_at 2.2325277 Down Hs.533336 BAMBI BMP and activin membrane-bound inhibitor homolog (Xenopus
laevis) 203612_at 2.1791766 Down Hs.106880 BYSL bystin-like 204118_at 2.3325217 Down Hs.243564 CD48 CD48 molecule 204794_at 3.0170782 Down Hs.1183 DUSP2 dual specificity phosphatase 2 204897_at 3.6472108 Down Hs.199248 PTGER4 prostaglandin E receptor 4 (subtype EP4) 204995_at 2.8569963 Down Hs.500015 CDK5R1 cyclin-dependent kinase 5, regulatory subunit 1 (p35) 205136_s_at 2.1136055 Down Hs.525006 NUFIP1 nuclear fragile X mental retardation protein interacting protein 1 205284_at 2.010743 Down Hs.533628 KIAA0133 KIAA0133 205419_at 3.2300916 Down Hs.784 EBI2 Epstein-Barr virus induced gene 2 (lymphocyte-specific G protein-
coupled receptor) 205943_at 2.163663 Down Hs.183671 TDO2 tryptophan 2,3-dioxygenase 206067_s_at 4.2541027 Down Hs.591980 WT1 Wilms tumor 1 206157_at 5.320476 Down Hs.591286 PTX3 pentraxin-related gene, rapidly induced by IL-1 beta 206653_at 2.3022125 Down Hs.282387 POLR3G polymerase (RNA) III (DNA directed) polypeptide G (32kD) 206772_at 2.1272707 Down Hs.570296 PTH2R parathyroid hormone 2 receptor
271
207229_at 2.4775214 Down Hs.159297 KLRA1 killer cell lectin-like receptor subfamily A, member 1 208473_s_at 2.3131196 Down Hs.709961 GP2 glycoprotein 2 (zymogen granule membrane) 209099_x_at 2.5063324 Down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 209120_at 3.0593445 Down Hs.701977 NR2F2 nuclear receptor subfamily 2, group F, member 2 209184_s_at 2.2956219 Down Hs.442344 IRS2 insulin receptor substrate 2 209606_at 2.8759694 Down Hs.270 PSCDBP pleckstrin homology, Sec7 and coiled-coil domains, binding protein 209803_s_at 2.3348641 Down Hs.154036 PHLDA2 pleckstrin homology-like domain, family A, member 2 210102_at 2.307687 Down Hs.152944 LOH11CR2A loss of heterozygosity, 11, chromosomal region 2, gene A 210492_at 2.2653377 Down Hs.593942 MFAP3L microfibrillar-associated protein 3-like 210751_s_at 2.016218 Down Hs.77854 RGN regucalcin (senescence marker protein-30) 212276_at 2.197002 Down Hs.467740 LPIN1 lipin 1 213478_at 2.0787475 Down Hs.368823 RP1-21O18.1 kazrin 213931_at 2.7013195 Down Hs.591670 ID2 /// ID2B inhibitor of DNA binding 2, dominant negative helix-loop-helix
protein /// inhibitor of DNA binding 2B, dominant negative helix-loop-helix protein
214295_at 2.3415482 Down Hs.604754 KIAA0485 KIAA0485 protein 215211_at 2.023611 Down LOC730092 RRN3 RNA polymerase I transcription factor homolog (S. cerevisiae)
pseudogene 216268_s_at 2.3467796 Down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 216598_s_at 2.7599106 Down Hs.303649 CCL2 chemokine (C-C motif) ligand 2 216620_s_at 2.0464752 Down Hs.98594 ARHGEF10 Rho guanine nucleotide exchange factor (GEF) 10 216749_at 2.5264196 Down Hs.677282 CDNA: FLJ21198 fis, clone COL00220 217604_at 2.0002718 Down Hs.635110 Transcribed locus 218706_s_at 2.0312455 Down Hs.363558 GRAMD3 GRAM domain containing 3 219225_at 2.014645 Down Hs.520463 LOC10013444
0 /// PGBD5 piggyBac transposable element derived 5 /// similar to PGBD5 protein
219497_s_at 3.7404122 Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219498_s_at 3.35297 Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219772_s_at 2.0480099 Down Hs.86492 SMPX small muscle protein, X-linked 219825_at 2.0878298 Down Hs.91546 CYP26B1 cytochrome P450, family 26, subfamily B, polypeptide 1 219890_at 2.4836237 Down Hs.446235 CLEC5A C-type lectin domain family 5, member A 219971_at 2.6633294 Down Hs.210546 IL21R interleukin 21 receptor
272
220116_at 4.063247 Down Hs.98280 KCNN2 potassium intermediate/small conductance calcium-activated channel, subfamily N, member 2
220502_s_at 2.0347304 Down Hs.489849 SLC13A1 solute carrier family 13 (sodium/sulfate symporters), member 1 220503_at 2.495743 Down Hs.489849 SLC13A1 solute carrier family 13 (sodium/sulfate symporters), member 1 221018_s_at 2.0035052 Down Hs.333132 TDRD1 tudor domain containing 1 221298_s_at 2.0786602 Down Hs.266223 SLC22A8 solute carrier family 22 (organic anion transporter), member 8 221586_s_at 2.6404374 Down Hs.445758 E2F5 E2F transcription factor 5, p130-binding 221766_s_at 2.619421 Down Hs.10784 FAM46A family with sequence similarity 46, member A 222664_at 2.3220863 Down Hs.221873 KCTD15 potassium channel tetramerisation domain containing 15 222891_s_at 3.474895 Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 223036_at 2.0134509 Down Hs.471452 FARSB phenylalanyl-tRNA synthetase, beta subunit 223403_s_at 2.3991299 Down Hs.86337 POLR1B polymerase (RNA) I polypeptide B, 128kDa 223688_s_at 2.3365293 Down Hs.699597 LY6K lymphocyte antigen 6 complex, locus K 224793_s_at 2.0926368 Down Hs.494622 TGFBR1 transforming growth factor, beta receptor I (activin A receptor type II-
like kinase, 53kDa) 224973_at 3.8480284 Down Hs.10784 FAM46A family with sequence similarity 46, member A 224979_s_at 2.0989978 Down Hs.464243 USP36 ubiquitin specific peptidase 36 225032_at 2.0437646 Down Hs.159430 FNDC3B fibronectin type III domain containing 3B 225962_at 2.5650084 Down Hs.427284 ZNRF1 zinc and ring finger 1 226479_at 2.0452113 Down Hs.534040 KBTBD6 kelch repeat and BTB (POZ) domain containing 6 226811_at 2.9312084 Down Hs.356216 FAM46C family with sequence similarity 46, member C 227037_at 2.4785297 Down Hs.31652 LOC201164 similar to CG12314 gene product 227210_at 2.2683113 Down Hs.407983 SFMBT2 Scm-like with four mbt domains 2 227242_s_at 3.7268121 Down Hs.699395 EBF3 early B-cell factor 3 227243_s_at 2.149735 Down Hs.699395 EBF3 early B-cell factor 3 227265_at 2.0887623 Down Hs.520989 FGL2 fibrinogen-like 2 227410_at 2.3463452 Down Hs.708232 FAM43A family with sequence similarity 43, member A 227481_at 2.1525605 Down Hs.16064 CNKSR3 CNKSR family member 3 227802_at 2.1459513 Down Hs.633432 Transcribed locus 228170_at 3.4621675 Down Hs.56663 OLIG1 oligodendrocyte transcription factor 1 228645_at 2.1142652 Down SNHG9 small nucleolar RNA host gene (non-protein coding) 9
273
228781_at 2.1254125 Down Hs.62314 CDNA FLJ33158 fis, clone UTERU2000418 229144_at 2.0105622 Down Hs.368823 RP1-21O18.1 kazrin 229307_at 2.1307192 Down Hs.335239 ANKRD28 ankyrin repeat domain 28 229607_at 2.1760874 Down 229638_at 2.8994858 Down Hs.499205 IRX3 iroquois homeobox 3 230044_at 2.2387767 Down Hs.585089 NPB Neuropeptide B 230058_at 2.1725152 Down Hs.94300 LOC646891 ///
SDCCAG3 serologically defined colon cancer antigen 3 /// similar to serologically defined colon cancer antigen 3
230343_at 2.0108373 Down Hs.28773 Transcribed locus 230395_at 2.1463017 Down Hs.709488 MRNA full length insert cDNA clone EUROIMAGE 2138357 230836_at 2.9297817 Down Hs.308628 ST8SIA4 ST8 alpha-N-acetyl-neuraminide alpha-2,8-sialyltransferase 4 231722_at 2.1362612 Down Hs.466057 CASP14 caspase 14, apoptosis-related cysteine peptidase 231798_at 2.9355247 Down Hs.248201 NOG noggin 232148_at 2.2736843 Down Hs.372000 NSMAF Neutral sphingomyelinase (N-SMase) activation associated factor 232181_at 2.33631 Down Hs.483816 LOC153346 hypothetical protein LOC153346 232504_at 4.109505 Down Hs.604728 LOC285628 hypothetical protein LOC285628 232610_at 2.0572758 Down Hs.709426 PARP14 poly (ADP-ribose) polymerase family, member 14 232958_at 2.2152493 Down Hs.660361 CDNA FLJ13595 fis, clone PLACE1009595 233085_s_at 2.8489802 Down Hs.591610 OBFC2A oligonucleotide/oligosaccharide-binding fold containing 2A 233591_at 2.9384553 Down Hs.306876 CDNA: FLJ23098 fis, clone LNG07440 233713_at 2.1836324 Down Hs.660880 CDNA FLJ12119 fis, clone MAMMA1000092 233961_at 2.1376956 Down Hs.677073 CDNA FLJ11859 fis, clone HEMBA1006832 234032_at 2.0185735 Down Hs.684536 Transcribed locus 235202_x_at 2.0879643 Down Hs.252543 IKIP IKK interacting protein 235265_at 2.0678198 Down Hs.379548 UBR3 ubiquitin protein ligase E3 component n-recognin 3 235359_at 2.4265985 Down Hs.709536 LRRC33 leucine rich repeat containing 33 235392_at 2.094309 Down Hs.659169 Transcribed locus 235551_at 2.3058908 Down Hs.248815 WDR4 WD repeat domain 4 235985_at 2.033237 Down Hs.657793 Transcribed locus 236096_at 2.1490674 Down Hs.655502 SIPA1L3 signal-induced proliferation-associated 1 like 3 236196_at 2.122972 Down Hs.586567 CDNA FLJ42548 fis, clone BRACE3004996
274
236561_at 3.3764417 Down Hs.494622 TGFBR1 Transforming growth factor, beta receptor I (activin A receptor type II-like kinase, 53kDa)
236738_at 3.0292625 Down Hs.710781 LOC401097 Similar to LOC166075 237031_at 2.1585586 Down Hs.146276 Full length insert cDNA clone YP08F12 237246_at 2.210518 Down Hs.670752 Transcribed locus 237593_at 2.2573826 Down Hs.667491 Transcribed locus 237597_at 2.038008 Down Hs.118228 Transcribed locus 238156_at 2.265855 Down Hs.663159 Transcribed locus 238246_at 2.0420265 Down Hs.145932 MTL5 Metallothionein-like 5, testis-specific (tesmin) 238520_at 2.0211198 Down Hs.485392 TRERF1 transcriptional regulating factor 1 238822_at 2.2096872 Down Hs.598557 Transcribed locus 239129_at 4.7192163 Down 239605_x_at 2.5360458 Down Hs.657657 Transcribed locus 239609_s_at 2.653492 Down Hs.352614 AGPAT7 1-acylglycerol-3-phosphate O-acyltransferase 7 (lysophosphatidic
acid acyltransferase, eta) 239895_at 2.242857 Down Hs.510958 AQR Aquarius homolog (mouse) 239973_at 2.2025015 Down Hs.212709 Transcribed locus 240362_at 2.288509 Down Hs.172755 BRP44L Brain protein 44-like 240747_at 3.579509 Down Hs.667630 Transcribed locus 241068_at 2.0787373 Down Hs.614053 Transcribed locus 241693_at 2.064382 Down Hs.660537 Transcribed locus 241859_at 2.4656346 Down Hs.153322 PLCL1 phospholipase C-like 1 242245_at 2.469184 Down Hs.533853 CDNA FLJ90705 fis, clone PLACE1007591 242388_x_at 2.1783433 Down Hs.601883 Transcribed locus 242435_at 2.0648775 Down Hs.89901 PDE4A Phosphodiesterase 4A, cAMP-specific (phosphodiesterase E2 dunce
homolog, Drosophila) 242673_at 2.3914204 Down Hs.599613 Transcribed locus 242943_at 2.020434 Down Hs.308628 ST8SIA4 ST8 alpha-N-acetyl-neuraminide alpha-2,8-sialyltransferase 4 243503_at 2.0134475 Down Hs.632985 Transcribed locus 243529_at 2.15252 Down Hs.116602 MARS2 methionyl-tRNA synthetase 2, mitochondrial 244183_x_at 2.0973384 Down Hs.674730 PCDHB3 protocadherin beta 3 244227_at 2.070997 Down Hs.370963 SYT6 synaptotagmin VI
275
244390_at 2.1863773 Down Hs.112791 Transcribed locus 1552521_a_at 2.1410308 Down Hs.99439 TMEM74 transmembrane protein 74 1552626_a_at 2.0103655 Down Hs.369471 TMEM163 transmembrane protein 163 1553074_at 2.011583 Down Hs.352183 ASB11 ankyrin repeat and SOCS box-containing 11 1553138_a_at 2.0253823 Down Hs.379097 ANKLE1 ankyrin repeat and LEM domain containing 1 1555300_a_at 2.0457644 Down Hs.58561 MED12L mediator complex subunit 12-like 1555777_at 2.0628526 Down Hs.136348 POSTN periostin, osteoblast specific factor 1555790_a_at 2.0564477 Down Hs.708090 TMEM192 ///
ZNF320 zinc finger protein 320 /// transmembrane protein 192
1555832_s_at 2.2229512 Down Hs.4055///Hs.709396
KLF6 Homo sapiens, clone IMAGE:4096273, mRNA /// Kruppel-like factor 6
1556194_a_at 2.08916 Down Hs.558200 CDNA FLJ33585 fis, clone BRAMY2012163 1556261_a_at 2.3748684 Down Hs.118609 CDNA FLJ40252 fis, clone TESTI2024299 1556361_s_at 3.6004255 Down Hs.105016 ANKRD13C ankyrin repeat domain 13C 1556894_at 2.1505616 Down Hs.84753 NT5DC2 5'-nucleotidase domain containing 2 1557126_a_at 2.173446 Down Hs.382865 PLD1 phospholipase D1, phosphatidylcholine-specific 1557593_at 2.5605173 Down Hs.528821 SPAG17 Sperm associated antigen 17 1558522_at 2.6067457 Down Hs.594275 Homo sapiens, clone IMAGE:3459334, mRNA 1558624_at 2.1274154 Down Hs.679977 Homo sapiens, clone IMAGE:5441027 1559078_at 2.072533 Down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 1559309_at 2.1830356 Down Hs.293689 MCFD2 multiple coagulation factor deficiency 2 1559326_at 2.0949717 Down Hs.525597 DIO3OS deiodinase, iodothyronine, type III opposite strand 1559394_a_at 2.2057455 Down Hs.660003 Full length insert cDNA clone ZC65D06 1559580_at 2.002937 Down Hs.44277 LRRC39 leucine rich repeat containing 39 1559948_at 2.374575 Down Hs.194746 CDNA FLJ20447 fis, clone KAT05276 1560426_at 2.201625 Down Hs.535389 C12orf55 chromosome 12 open reading frame 55 1560684_x_at 2.0104003 Down Hs.657985 BCL8 B-cell CLL/lymphoma 8 1561331_at 2.3409355 Down C1orf99 chromosome 1 open reading frame 99 1563420_at 2.313869 Down XGPY2 Xg pseudogene, Y-linked 2 1566991_at 2.0156975 Down Hs.291587 ARID1B AT rich interactive domain 1B (SWI1-like) 1568813_at 3.2485936 Down Hs.418285 CDNA clone IMAGE:4620359
276
1569369_at 2.2046144 Down Hs.292056 ZFYVE28 zinc finger, FYVE domain containing 28 1569469_a_at 2.1429687 Down Hs.403934 LHX8 LIM homeobox 8 1569948_at 2.0533514 Down Hs.685028 CDNA clone IMAGE:5275301 1570153_at 2.2748373 Down Hs.646618 SOHLH2 spermatogenesis and oogenesis specific basic helix-loop-helix 2 1570352_at 2.3949015 Down Hs.367437 ATM ataxia telangiectasia mutated 1570393_at 2.0387962 Down Hs.712606 EML5 echinoderm microtubule associated protein like 5
277
Table A3.4: Retinoid response gene targets in NuMA-RARA Probe Set ID Fold
change([12RA] vs [12])
Regulation ([12RA] vs
[12])
Unigene (Avadis)
Gene Symbol
Gene Title
201324_at 2.0886235 down Hs.436298 EMP1 epithelial membrane protein 1 201565_s_at 3.21204 down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix
protein 201566_x_at 2.9929056 down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix
protein 202206_at 2.399275 down Hs.709513 ARL4C ADP-ribosylation factor-like 4C 202431_s_at 12.158877 down Hs.202453 MYC v-myc myelocytomatosis viral oncogene homolog (avian) 202859_x_at 2.084367 down Hs.551925 IL8 interleukin 8 203304_at 2.7989619 down Hs.533336 BAMBI BMP and activin membrane-bound inhibitor homolog (Xenopus
laevis) 203408_s_at 2.1148467 down Hs.517717 SATB1 SATB homeobox 1 203604_at 2.0690129 down Hs.709890 ZNF516 zinc finger protein 516 204072_s_at 2.3049405 down Hs.591225 FRY furry homolog (Drosophila) 204249_s_at 2.1231515 down Hs.34560 LMO2 LIM domain only 2 (rhombotin-like 1) 204794_at 2.1565883 down Hs.1183 DUSP2 dual specificity phosphatase 2 204798_at 2.948894 down Hs.654446 MYB v-myb myeloblastosis viral oncogene homolog (avian) 204897_at 5.9579215 down Hs.199248 PTGER4 prostaglandin E receptor 4 (subtype EP4) 205039_s_at 2.0252483 down Hs.435949 IKZF1 IKAROS family zinc finger 1 (Ikaros) 205284_at 2.1262233 down Hs.533628 KIAA0133 KIAA0133 205419_at 2.4826307 down Hs.784 EBI2 Epstein-Barr virus induced gene 2 (lymphocyte-specific G
protein-coupled receptor) 206157_at 4.476515 down Hs.591286 PTX3 pentraxin-related gene, rapidly induced by IL-1 beta 206170_at 2.7569294 down Hs.591251 ADRB2 adrenergic, beta-2-, receptor, surface 206806_at 2.2899141 down Hs.242947 DGKI diacylglycerol kinase, iota 206943_at 2.0320473 down Hs.494622 TGFBR1 transforming growth factor, beta receptor I (activin A receptor
type II-like kinase, 53kDa) 207980_s_at 2.2106276 down Hs.82071 CITED2 Cbp/p300-interacting transactivator, with Glu/Asp-rich carboxy-
terminal domain, 2
278
208960_s_at 2.134312 down Hs.4055 KLF6 Kruppel-like factor 6 208961_s_at 2.071602 down Hs.4055 KLF6 Kruppel-like factor 6 209099_x_at 2.2153378 down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 209120_at 2.0699842 down Hs.701977 NR2F2 nuclear receptor subfamily 2, group F, member 2 209184_s_at 3.2237732 down Hs.442344 IRS2 insulin receptor substrate 2 209185_s_at 3.1897783 down Hs.442344 IRS2 insulin receptor substrate 2 209606_at 2.753105 down Hs.270 PSCDBP pleckstrin homology, Sec7 and coiled-coil domains, binding
protein 213931_at 2.8903654 down Hs.591670 ID2 ///
ID2B inhibitor of DNA binding 2, dominant negative helix-loop-helix protein /// inhibitor of DNA binding 2B, dominant negative helix-loop-helix protein
214581_x_at 2.7177627 down Hs.443577 TNFRSF21 tumor necrosis factor receptor superfamily, member 21 214651_s_at 2.0072289 down Hs.659350 HOXA9 homeobox A9 216012_at 2.3650792 down Hs.550193 Unidentified mRNA, partial sequence 216268_s_at 2.7129872 down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 216901_s_at 2.2050683 down Hs.435949 IKZF1 IKAROS family zinc finger 1 (Ikaros) 217523_at 2.831772 down Hs.502328 CD44 CD44 molecule (Indian blood group) 218251_at 2.0469325 down Hs.522605 MID1IP1 MID1 interacting protein 1 (gastrulation specific G12 homolog
(zebrafish) 218856_at 2.5045984 down Hs.443577 TNFRSF21 tumor necrosis factor receptor superfamily, member 21 219497_s_at 4.438934 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219498_s_at 3.2207508 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219890_at 2.4856505 down Hs.446235 CLEC5A C-type lectin domain family 5, member A 220116_at 3.7291224 down Hs.98280 KCNN2 potassium intermediate/small conductance calcium-activated
channel, subfamily N, member 2 221586_s_at 2.598485 down Hs.445758 E2F5 E2F transcription factor 5, p130-binding 221766_s_at 3.498441 down Hs.10784 FAM46A family with sequence similarity 46, member A 222891_s_at 4.641202 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 223403_s_at 2.2585576 down Hs.86337 POLR1B polymerase (RNA) I polypeptide B, 128kDa 224606_at 2.4023688 down Hs.4055 KLF6 Kruppel-like factor 6 224973_at 4.151496 down Hs.10784 FAM46A family with sequence similarity 46, member A 225167_at 2.0883074 down Hs.330463 FRMD4A FERM domain containing 4A
279
225436_at 2.206226 down Hs.459072 FAM108C1 family with sequence similarity 108, member C1 225974_at 2.0538626 down Hs.567759 TMEM64 transmembrane protein 64 227037_at 3.0964594 down Hs.31652 LOC20116
4 similar to CG12314 gene product
227210_at 2.3620021 down Hs.407983 SFMBT2 Scm-like with four mbt domains 2 227242_s_at 2.3002076 down Hs.699395 EBF3 early B-cell factor 3 227346_at 2.2483537 down Hs.488251 IKZF1 IKAROS family zinc finger 1 (Ikaros) 227410_at 2.2359083 down Hs.708232 FAM43A family with sequence similarity 43, member A 227481_at 2.3246589 down Hs.16064 CNKSR3 CNKSR family member 3 228170_at 3.0674152 down Hs.56663 OLIG1 oligodendrocyte transcription factor 1 229638_at 3.6987438 down Hs.499205 IRX3 iroquois homeobox 3 229723_at 2.0052488 down Hs.529984 TAGAP T-cell activation RhoGTPase activating protein 231786_at 2.0587091 down Hs.592172 HOXA13 homeobox A13 232291_at 2.7976894 down Hs.24115 MIRHG1 microRNA host gene (non-protein coding) 1 232504_at 2.5817292 down Hs.604728 LOC28562
8 hypothetical protein LOC285628
236738_at 8.455545 down Hs.710781 LOC401097
Similar to LOC166075
238607_at 2.3092794 down Hs.192237 ZNF342 zinc finger protein 342 239129_at 2.0111887 down 239605_x_at 2.1959515 down Hs.657657 Transcribed locus 240747_at 2.0800576 down Hs.667630 Transcribed locus 241365_at 2.0684748 down Hs.593276 CDNA FLJ42259 fis, clone TKIDN2011289 242388_x_at 2.214354 down Hs.601883 Transcribed locus 243529_at 2.1790874 down Hs.116602 MARS2 methionyl-tRNA synthetase 2, mitochondrial 244267_at 2.5540445 down Hs.674678 Transcribed locus 1554887_at 2.272857 down 1555832_s_at 2.612295 down Hs.4055///Hs.
709396 KLF6 Homo sapiens, clone IMAGE:4096273, mRNA /// Kruppel-like
factor 6 1564906_at 2.2597687 down SNHG4 small nucleolar RNA host gene (non-protein coding) 4 1569054_at 2.2992537 down Hs.481918 SLC1A3 solute carrier family 1 (glial high affinity glutamate transporter),
member 3
280
200644_at 2.9435694 up Hs.75061 MARCKSL1
MARCKS-like 1
200897_s_at 6.706352 up Hs.151220 PALLD palladin, cytoskeletal associated protein 200906_s_at 2.92617 up Hs.151220 PALLD palladin, cytoskeletal associated protein 200907_s_at 4.788296 up Hs.151220 PALLD palladin, cytoskeletal associated protein 201042_at 2.017785 up Hs.517033 TGM2 transglutaminase 2 (C polypeptide, protein-glutamine-gamma-
glutamyltransferase) 201471_s_at 2.0305166 up Hs.437277 SQSTM1 sequestosome 1 201534_s_at 2.1077316 up Hs.145575 UBL3 ubiquitin-like 3 201631_s_at 2.4305413 up Hs.591785 IER3 immediate early response 3 201656_at 2.5098853 up Hs.133397 ITGA6 integrin, alpha 6 202308_at 2.0174117 up Hs.592123 SREBF1 sterol regulatory element binding transcription factor 1 202481_at 4.625771 up Hs.289347 DHRS3 dehydrogenase/reductase (SDR family) member 3 202627_s_at 3.8934524 up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E (nexin, plasminogen activator
inhibitor type 1), member 1 202628_s_at 3.1118069 up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E (nexin, plasminogen activator
inhibitor type 1), member 1 202769_at 2.132356 up Hs.13291 CCNG2 cyclin G2 202770_s_at 2.2489765 up Hs.13291 CCNG2 cyclin G2 202869_at 6.2625756 up Hs.524760 OAS1 2',5'-oligoadenylate synthetase 1, 40/46kDa 202887_s_at 4.862352 up Hs.523012 DDIT4 DNA-damage-inducible transcript 4 203020_at 4.299632 up Hs.585378 RABGAP1
L RAB GTPase activating protein 1-like
203490_at 2.0644116 up Hs.271940 ELF4 E74-like factor 4 (ets domain transcription factor) 203567_s_at 2.0241668 up Hs.584851 TRIM38 tripartite motif-containing 38 203610_s_at 2.135064 up Hs.584851 TRIM38 tripartite motif-containing 38 203760_s_at 4.567203 up Hs.75367 SLA Src-like-adaptor 203761_at 4.160341 up Hs.75367 SLA Src-like-adaptor 203821_at 2.9183996 up Hs.799 HBEGF heparin-binding EGF-like growth factor 203887_s_at 9.840143 up Hs.2030 THBD thrombomodulin 203888_at 7.390962 up Hs.2030 THBD thrombomodulin 203936_s_at 2.7114239 up Hs.297413 MMP9 matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa
281
type IV collagenase) 204112_s_at 2.164019 up Hs.42151 HNMT histamine N-methyltransferase 204429_s_at 2.9717975 up Hs.530003 SLC2A5 solute carrier family 2 (facilitated glucose/fructose transporter),
member 5 204430_s_at 3.808644 up Hs.530003 SLC2A5 solute carrier family 2 (facilitated glucose/fructose transporter),
member 5 204526_s_at 2.1254432 up Hs.442657 TBC1D8 TBC1 domain family, member 8 (with GRAM domain) 204546_at 2.5628142 up Hs.301658 KIAA0513 KIAA0513 204908_s_at 3.4835668 up Hs.31210 BCL3 B-cell CLL/lymphoma 3 204961_s_at 2.6331637 up Hs.655201 LOC64899
214523_at 3.213535 up Hs.558308 CEBPE CCAAT/enhancer binding protein (C/EBP), epsilon 214639_s_at 2.2926025 up Hs.67397 HOXA1 homeobox A1 214977_at 2.097475 up Hs.654670 CDNA FLJ13790 fis, clone THYRO1000026 215177_s_at 2.6028233 up Hs.133397 ITGA6 integrin, alpha 6 215342_s_at 4.092742 up Hs.585378 RABGAP1
L RAB GTPase activating protein 1-like
216015_s_at 2.7958415 up Hs.159483 NLRP3 NLR family, pyrin domain containing 3 217104_at 2.3176522 up Hs.459049 MTHFS ///
ST20 5,10-methenyltetrahydrofolate synthetase (5-formyltetrahydrofolate cyclo-ligase) /// suppressor of tumorigenicity 20
218501_at 7.5679164 up Hs.476402 ARHGEF3 Rho guanine nucleotide exchange factor (GEF) 3 218627_at 2.9676416 up Hs.525634 DRAM damage-regulated autophagy modulator 218723_s_at 2.5117044 up Hs.507866 C13orf15 chromosome 13 open reading frame 15 219010_at 2.7844956 up Hs.518997 C1orf106 chromosome 1 open reading frame 106 219607_s_at 3.3354638 up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 219901_at 2.5995123 up Hs.506381 FGD6 FYVE, RhoGEF and PH domain containing 6 219994_at 2.644592 up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B, member 1
interacting protein 220148_at 2.2781162 up Hs.486520 ALDH8A1 aldehyde dehydrogenase 8 family, member A1 220494_s_at 2.3231566 up 221266_s_at 2.5050628 up Hs.652230 TM7SF4 transmembrane 7 superfamily member 4
284
221345_at 3.761886 up Hs.248056 FFAR2 free fatty acid receptor 2 221601_s_at 4.777954 up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 221602_s_at 3.9625475 up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 221748_s_at 2.7049792 up Hs.471381 TNS1 tensin 1 31845_at 2.1242719 up Hs.271940 ELF4 E74-like factor 4 (ets domain transcription factor) 34210_at 2.6051145 up Hs.276770 CD52 CD52 molecule 36711_at 6.3232007 up Hs.517617 MAFF v-maf musculoaponeurotic fibrosarcoma oncogene homolog F
(avian) 37152_at 2.2212722 up Hs.696032 PPARD peroxisome proliferator-activated receptor delta 38037_at 2.565572 up Hs.799 HBEGF heparin-binding EGF-like growth factor 222996_s_at 2.2260995 up Hs.189119 CXXC5 CXXC finger 5 223276_at 3.4076967 up Hs.29444 MST150 MSTP150 223567_at 6.1853256 up Hs.465642 SEMA6B sema domain, transmembrane domain (TM), and cytoplasmic
domain, (semaphorin) 6B 223620_at 2.1318035 up Hs.495989 GPR34 G protein-coupled receptor 34 224048_at 2.102644 up Hs.646421 USP44 ubiquitin specific peptidase 44 224516_s_at 2.4642644 up Hs.189119 CXXC5 CXXC finger 5 224916_at 3.0291984 up Hs.379754 TMEM173 transmembrane protein 173 224929_at 3.1026132 up Hs.379754 TMEM173 transmembrane protein 173 224964_s_at 2.3579462 up Hs.187772 GNG2 guanine nucleotide binding protein (G protein), gamma 2 225347_at 2.9668434 up Hs.497399 ARL8A ADP-ribosylation factor-like 8A 225372_at 2.9746375 up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225373_at 2.2076936 up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225763_at 2.1507406 up Hs.493867 RCSD1 RCSD domain containing 1 225842_at 2.064913 up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 225919_s_at 2.1124992 up Hs.493639 C9orf72 chromosome 9 open reading frame 72 226039_at 2.0417984 up Hs.177576 MGAT4A mannosyl (alpha-1,3-)-glycoprotein beta-1,4-N-
acetylglucosaminyltransferase, isozyme A 226146_at 2.0081341 up Hs.655606 CDNA clone IMAGE:5294560 226215_s_at 2.629909 up Hs.524800 FBXL10 F-box and leucine-rich repeat protein 10 226487_at 2.6733472 up Hs.661785 C12orf34 chromosome 12 open reading frame 34 226673_at 2.4407449 up Hs.306412 SH2D3C SH2 domain containing 3C
285
226722_at 2.0995586 up Hs.134742 FAM20C family with sequence similarity 20, member C 226756_at 2.6342082 up Hs.633903 CDNA FLJ25556 fis, clone JTH02629 226855_at 3.5249252 up Hs.632214 CDNA FLJ40954 fis, clone UTERU2010525 227396_at 2.8179464 up Hs.318547 PTPRJ protein tyrosine phosphatase, receptor type, J 227484_at 3.5488734 up Hs.710097 CDNA FLJ41690 fis, clone HCASM2009405 227701_at 2.0022275 up Hs.159066 C10orf118 chromosome 10 open reading frame 118 227792_at 2.9687207 up Hs.648523 LOC16207
3 hypothetical protein LOC162073
227915_at 3.3251038 up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 228120_at 2.4339266 up Hs.656677 CDNA: FLJ22073 fis, clone HEP11868 228648_at 5.729146 up Hs.655559 LRG1 leucine-rich alpha-2-glycoprotein 1 228762_at 2.5676794 up Hs.159142 LFNG LFNG O-fucosylpeptide 3-beta-N-acetylglucosaminyltransferase 228772_at 2.638866 up Hs.42151 HNMT histamine N-methyltransferase 228854_at 2.283559 up Hs.586747 Transcribed locus 229399_at 2.0305078 up Hs.159066 C10orf118 chromosome 10 open reading frame 118 229625_at 2.8929 up Hs.513726 GBP5 guanylate binding protein 5 229670_at 3.5084085 up Hs.180284 5.5 kb mRNA upregulated in retinoic acid treated HL-60
neutrophilic cells 229795_at 2.0522285 up Hs.48945 Transcribed locus 229971_at 3.044595 up Hs.187884 GPR114 G protein-coupled receptor 114 230218_at 2.6236181 up Hs.72956 HIC1 hypermethylated in cancer 1 230333_at 2.728373 up Hs.656630 Transcribed locus 230713_at 2.02981 up Hs.476164 Transcribed locus 230925_at 3.1246424 up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B, member 1
interacting protein 231496_at 3.575809 up Hs.145519 FCAMR Fc receptor, IgA, IgM, high affinity 231779_at 2.9554198 up Hs.449207 IRAK2 interleukin-1 receptor-associated kinase 2 231969_at 2.2740314 up Hs.21958 STOX2 storkhead box 2 232687_at 2.5561237 up Hs.100912 CDNA FLJ33091 fis, clone TRACH2000660 232754_at 2.0563302 up Hs.664518 CDNA FLJ11563 fis, clone HEMBA1003202 232861_at 4.6474376 up Hs.654693 PDP2 pyruvate dehydrogenase phosphatase isoenzyme 2 232912_at 2.1348078 up Hs.657472 GPR180 G protein-coupled receptor 180
286
233857_s_at 4.5003366 up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 233955_x_at 2.2768729 up Hs.189119 CXXC5 CXXC finger 5 234361_at 2.4593277 up Hs.247744 CREB3L3 cAMP responsive element binding protein 3-like 3 234987_at 4.1412725 up Hs.660221 Transcribed locus 235421_at 2.1626062 up Hs.432453///
IMAGE:4689481 235456_at 2.520447 up Hs.130853 CDNA clone IMAGE:4819084 235505_s_at 3.9337726 up Hs.40966 MRNA full length insert cDNA clone EUROIMAGE 2362292 235521_at 2.126902 up Hs.659337 HOXA3 homeobox A3 235529_x_at 2.320773 up Hs.660221 Transcribed locus 236191_at 5.362256 up Hs.667427 Transcribed locus 236223_s_at 2.2420454 up Hs.491234 RIT1 Ras-like without CAAX 1 236407_at 6.211272 up Hs.121495 KCNE1 potassium voltage-gated channel, Isk-related family, member 1 236619_at 2.107196 up Hs.196073 Transcribed locus 236717_at 5.53467 up Hs.525977 LOC16518
6 similar to RIKEN cDNA 4632412N22 gene
237252_at 5.35475 up Hs.2030 THBD thrombomodulin 237442_at 2.538061 up 238032_at 2.6913702 up Hs.655631 Transcribed locus 238439_at 2.5457928 up Hs.217484 ANKRD22 ankyrin repeat domain 22 238581_at 7.4942203 up Hs.513726 GBP5 guanylate binding protein 5 238669_at 2.9314187 up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H
synthase and cyclooxygenase) 239196_at 2.1484046 up Hs.217484 ANKRD22 ankyrin repeat domain 22 239258_at 2.9715037 up Hs.713163 Transcribed locus 239294_at 4.0445514 up Hs.561747 Transcribed locus 239808_at 2.0879884 up Hs.656954 Transcribed locus 239843_at 2.2132082 up Hs.491234 RIT1 Ras-like without CAAX 1 240173_at 2.106467 up Hs.602127 Transcribed locus 241742_at 2.1086307 up Hs.465812 PRAM1 PML-RARA regulated adaptor molecule 1 242426_at 4.667349 up Hs.696574 NRG4 neuregulin 4 242525_at 2.6995246 up Hs.439122 Transcribed locus
287
242598_at 2.1197417 up Hs.674063 Transcribed locus 242705_x_at 3.301952 up Hs.592928 Full length insert cDNA clone YT86E01 243463_s_at 2.2107568 up Hs.491234 RIT1 Ras-like without CAAX 1 243541_at 3.0937464 up Hs.55378 IL31RA interleukin 31 receptor A 243745_at 2.017381 up Hs.673919 Transcribed locus 243819_at 2.0161119 up 244014_x_at 2.278581 up Hs.620180 Transcribed locus 244434_at 2.0592644 up Hs.567457 Transcribed locus 244665_at 2.61225 up Hs.668855 Transcribed locus 1553043_a_at 2.9935308 up Hs.567706 CD300LF CD300 molecule-like family member f 1554240_a_at 2.4719732 up Hs.174103 ITGAL integrin, alpha L (antigen CD11A (p180), lymphocyte function-
associated antigen 1; alpha polypeptide) 1555137_a_at 2.1388674 up Hs.506381 FGD6 FYVE, RhoGEF and PH domain containing 6 1555431_a_at 2.369251 up Hs.55378 IL31RA interleukin 31 receptor A 1555680_a_at 3.22602 up Hs.433337 SMOX spermine oxidase 1555728_a_at 3.1336443 up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 1555875_at 2.0978556 up Hs.210751 Homo sapiens, clone IMAGE:3604678, mRNA 1563745_a_at 3.7158568 up Hs.309176 LOC28305
0 hypothetical LOC283050
1556698_a_at 2.4326565 up Hs.605082 GPRIN3 GPRIN family member 3 1557051_s_at 2.0846317 up Hs.445239 CDNA FLJ12777 fis, clone NT2RP2001720 1557545_s_at 2.8805594 up Hs.501114 RNF165 ring finger protein 165 1559214_at 2.42021 up Hs.238914 MRNA full length insert cDNA clone EUROIMAGE 839551 1559391_s_at 2.3651621 up Hs.667269 Partial mRNA; ID EE2-8E 1559883_s_at 2.4473035 up Hs.580681 SAMHD1 SAM domain and HD domain 1 1562255_at 2.885095 up Hs.436977 SYTL3 synaptotagmin-like 3 1568619_s_at 2.378154 up Hs.648523 LOC16207
3 hypothetical protein LOC162073
1569129_s_at 2.0053325 up Hs.476944 Transcribed locus 1569157_s_at 2.0833037 up Hs.665717 LOC16299
3 hypothetical protein LOC162993
1570375_at 2.2113552 up Hs.661265 CDNA FLJ41985 fis, clone SPLEN2014946
288
Table A3.5: Retinoid responsive gene targets in control GFP cells
Probe Set ID Fold change
([GRA] vs [G]) Regulation([G
RA] vs [G]) Unigene (Avadis)
Gene Symbol Gene Title
201565_s_at 3.19715 down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix protein
201566_x_at 3.0895824 down Hs.180919 ID2 inhibitor of DNA binding 2, dominant negative helix-loop-helix protein
201626_at 2.2383513 down Hs.520819 INSIG1 insulin induced gene 1 201627_s_at 2.23358 down Hs.520819 INSIG1 insulin induced gene 1 202431_s_at 5.3314357 down Hs.202453 MYC v-myc myelocytomatosis viral oncogene homolog (avian) 202760_s_at 2.2443905 down Hs.591908 PALM2-AKAP2 PALM2-AKAP2 203304_at 2.6940384 down Hs.533336 BAMBI BMP and activin membrane-bound inhibitor homolog
(Xenopus laevis) 203604_at 2.0618768 down Hs.709890 ZNF516 zinc finger protein 516 204072_s_at 2.1826165 down Hs.591225 FRY furry homolog (Drosophila) 204249_s_at 2.0089493 down Hs.34560 LMO2 LIM domain only 2 (rhombotin-like 1) 204352_at 2.1479347 down Hs.523930 TRAF5 TNF receptor-associated factor 5 204794_at 2.2907133 down Hs.1183 DUSP2 dual specificity phosphatase 2 204897_at 3.9316552 down Hs.199248 PTGER4 prostaglandin E receptor 4 (subtype EP4) 204900_x_at 2.0170596 down Hs.591715 SAP30 Sin3A-associated protein, 30kDa 205419_at 4.3923125 down Hs.784 EBI2 Epstein-Barr virus induced gene 2 (lymphocyte-specific G
protein-coupled receptor) 206157_at 3.9286077 down Hs.591286 PTX3 pentraxin-related gene, rapidly induced by IL-1 beta 206170_at 2.8135798 down Hs.591251 ADRB2 adrenergic, beta-2-, receptor, surface 206653_at 2.602806 down Hs.282387 POLR3G polymerase (RNA) III (DNA directed) polypeptide G (32kD) 206943_at 3.033631 down Hs.494622 TGFBR1 transforming growth factor, beta receptor I (activin A receptor
type II-like kinase, 53kDa) 208056_s_at 2.3279235 down Hs.513811 CBFA2T3 core-binding factor, runt domain, alpha subunit 2; translocated
to, 3 209098_s_at 2.0579336 down Hs.224012 JAG1 jagged 1 (Alagille syndrome)
289
209099_x_at 2.6896167 down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 209184_s_at 2.6711507 down Hs.442344 IRS2 insulin receptor substrate 2 209185_s_at 2.7987473 down Hs.442344 IRS2 insulin receptor substrate 2 209433_s_at 2.06024 down Hs.331420 PPAT phosphoribosyl pyrophosphate amidotransferase 209606_at 2.0527284 down Hs.270 PSCDBP pleckstrin homology, Sec7 and coiled-coil domains, binding
protein 209750_at 2.1113086 down Hs.37288 NR1D2 nuclear receptor subfamily 1, group D, member 2 209803_s_at 2.7973564 down Hs.154036 PHLDA2 pleckstrin homology-like domain, family A, member 2 210347_s_at 2.4175909 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 212875_s_at 2.0140817 down Hs.473894 C2CD2 C2 calcium-dependent domain containing 2 213005_s_at 2.1896367 down Hs.306764 KANK1 KN motif and ankyrin repeat domains 1 213478_at 2.0669103 down Hs.368823 RP1-21O18.1 kazrin 213931_at 3.1132224 down Hs.591670 ID2 /// ID2B inhibitor of DNA binding 2, dominant negative helix-loop-
helix protein /// inhibitor of DNA binding 2B, dominant negative helix-loop-helix protein
214321_at 2.3082538 down Hs.235935 NOV nephroblastoma overexpressed gene 215723_s_at 2.1080801 down Hs.382865 PLD1 phospholipase D1, phosphatidylcholine-specific 216012_at 3.1767006 down Hs.550193 Unidentified mRNA, partial sequence 216211_at 2.2012672 down Hs.659130 MRNA; cDNA DKFZp564A023 (from clone
DKFZp564A023) 216252_x_at 2.0069475 down Hs.244139 FAS Fas (TNF receptor superfamily, member 6) 216268_s_at 2.9269807 down Hs.224012 JAG1 jagged 1 (Alagille syndrome) 216598_s_at 2.0723991 down Hs.303649 CCL2 chemokine (C-C motif) ligand 2 216620_s_at 2.023201 down Hs.98594 ARHGEF10 Rho guanine nucleotide exchange factor (GEF) 10 217647_at 2.0704885 down Hs.660660 CDNA FLJ40920 fis, clone UTERU2005905 218331_s_at 2.0102592 down Hs.699500 C10orf18 chromosome 10 open reading frame 18 218618_s_at 2.0625687 down Hs.159430 FNDC3B fibronectin type III domain containing 3B 219334_s_at 2.2212722 down Hs.591610 OBFC2A oligonucleotide/oligosaccharide-binding fold containing 2A 219497_s_at 5.669927 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219498_s_at 4.8316703 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 219697_at 2.1828601 down Hs.622536 HS3ST2 heparan sulfate (glucosamine) 3-O-sulfotransferase 2 219890_at 3.1078908 down Hs.446235 CLEC5A C-type lectin domain family 5, member A
290
220116_at 5.6960754 down Hs.98280 KCNN2 potassium intermediate/small conductance calcium-activated channel, subfamily N, member 2
220459_at 2.20579 down Hs.709346 MCM3APAS minichromosome maintenance complex component 3 associated protein antisense
221586_s_at 3.1761389 down Hs.445758 E2F5 E2F transcription factor 5, p130-binding 221766_s_at 3.5650196 down Hs.10784 FAM46A family with sequence similarity 46, member A 221778_at 2.3323536 down Hs.308710 JHDM1D jumonji C domain containing histone demethylase 1 homolog
D (S. cerevisiae) 221841_s_at 2.3568435 down Hs.376206 KLF4 Kruppel-like factor 4 (gut) 222071_s_at 2.139667 down Hs.709540 SLCO4C1 solute carrier organic anion transporter family, member 4C1 222322_at 2.6270673 down Hs.661379 Transcribed locus 222668_at 2.0507164 down Hs.221873 KCTD15 potassium channel tetramerisation domain containing 15 222692_s_at 2.0523262 down Hs.159430 FNDC3B fibronectin type III domain containing 3B 222872_x_at 2.4523625 down Hs.591610 OBFC2A oligonucleotide/oligosaccharide-binding fold containing 2A 222891_s_at 7.0744944 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 223397_s_at 2.0809543 down Hs.585728 NIP7 nuclear import 7 homolog (S. cerevisiae) 223403_s_at 2.1982484 down Hs.86337 POLR1B polymerase (RNA) I polypeptide B, 128kDa 223503_at 2.1618946 down Hs.369471 TMEM163 transmembrane protein 163 224793_s_at 2.6721416 down Hs.494622 TGFBR1 transforming growth factor, beta receptor I (activin A receptor
type II-like kinase, 53kDa) 224973_at 3.877564 down Hs.10784 FAM46A family with sequence similarity 46, member A 225032_at 2.0762935 down Hs.159430 FNDC3B fibronectin type III domain containing 3B 225974_at 2.6766124 down Hs.567759 TMEM64 transmembrane protein 64 226350_at 2.055385 down Hs.654545 CHML choroideremia-like (Rab escort protein 2) 226360_at 2.0352998 down Hs.655242 ZNRF3 zinc and ring finger 3 227037_at 2.7701855 down Hs.31652 LOC201164 similar to CG12314 gene product 227210_at 2.4669845 down Hs.407983 SFMBT2 Scm-like with four mbt domains 2 227242_s_at 2.012063 down Hs.699395 EBF3 early B-cell factor 3 227354_at 2.0498126 down Hs.266175 PAG1 phosphoprotein associated with glycosphingolipid
microdomains 1 227481_at 2.2259252 down Hs.16064 CNKSR3 CNKSR family member 3 228170_at 2.047523 down Hs.56663 OLIG1 oligodendrocyte transcription factor 1
291
228176_at 2.069586 down Hs.585118 S1PR3 sphingosine-1-phosphate receptor 3 228333_at 2.0105028 down Hs.621487 Full length insert cDNA clone YT94E02 229307_at 2.0694149 down Hs.335239 ANKRD28 ankyrin repeat domain 28 229638_at 3.289962 down Hs.499205 IRX3 iroquois homeobox 3 229686_at 2.112692 down Hs.111377 P2RY8 purinergic receptor P2Y, G-protein coupled, 8 229723_at 2.766404 down Hs.529984 TAGAP T-cell activation RhoGTPase activating protein 231310_at 2.0057406 down Hs.113170 Transcribed locus 231798_at 2.3127465 down Hs.248201 NOG noggin 232291_at 2.6771522 down Hs.24115 MIRHG1 microRNA host gene (non-protein coding) 1 232530_at 2.0205226 down Hs.382865 PLD1 phospholipase D1, phosphatidylcholine-specific 232629_at 2.1365838 down Hs.528665 PROK2 prokineticin 2 232778_at 2.005665 down Hs.640558 CDNA: FLJ22383 fis, clone HRC07564 232958_at 2.397218 down Hs.660361 CDNA FLJ13595 fis, clone PLACE1009595 233085_s_at 2.4954445 down Hs.591610 OBFC2A oligonucleotide/oligosaccharide-binding fold containing 2A 235359_at 2.527995 down Hs.709536 LRRC33 leucine rich repeat containing 33 236561_at 4.1622524 down Hs.494622 TGFBR1 Transforming growth factor, beta receptor I (activin A receptor
type II-like kinase, 53kDa) 236738_at 5.442677 down Hs.710781 LOC401097 Similar to LOC166075 239129_at 3.6913147 down 239296_at 2.335155 down Hs.672093 Transcribed locus 239605_x_at 3.0961626 down Hs.657657 Transcribed locus 239911_at 2.007892 down Hs.194725 ONECUT2 one cut homeobox 2 240008_at 2.01395 down Hs.656290 Transcribed locus 240452_at 2.5822086 down Hs.528780 GSPT1 G1 to S phase transition 1 240747_at 2.4500487 down Hs.667630 Transcribed locus 241716_at 2.1328788 down HSPD1 heat shock 60kDa protein 1 (chaperonin) 242245_at 2.069497 down Hs.533853 CDNA FLJ90705 fis, clone PLACE1007591 242388_x_at 2.634984 down Hs.601883 Transcribed locus 242550_at 2.1002643 down Hs.371001 EIF3B eukaryotic translation initiation factor 3, subunit B 242905_at 2.5441372 down Hs.262858 PNO1 partner of NOB1 homolog (S. cerevisiae) 243088_at 2.0176904 down
292
243529_at 2.085383 down Hs.116602 MARS2 methionyl-tRNA synthetase 2, mitochondrial 244022_at 2.4184115 down 244165_at 2.3861125 down Hs.699500 C10orf18 chromosome 10 open reading frame 18 244267_at 2.1750958 down Hs.674678 Transcribed locus 1552626_a_at 2.23773 down Hs.369471 TMEM163 transmembrane protein 163 1557360_at 2.2121725 down Hs.368084 LRPPRC leucine-rich PPR-motif containing 1558111_at 2.0698903 down Hs.478000 MBNL1 muscleblind-like (Drosophila) 1559078_at 3.2942047 down Hs.370549 BCL11A B-cell CLL/lymphoma 11A (zinc finger protein) 1561017_at 2.305915 down Hs.684677 Full length insert cDNA clone YW28G08 1566482_at 2.3295524 down Hs.684006 Transcribed locus 200632_s_at 2.2249727 up Hs.372914 NDRG1 N-myc downstream regulated gene 1 200897_s_at 13.084492 up Hs.151220 PALLD palladin, cytoskeletal associated protein 200906_s_at 4.925171 up Hs.151220 PALLD palladin, cytoskeletal associated protein 200907_s_at 8.944003 up Hs.151220 PALLD palladin, cytoskeletal associated protein 201042_at 2.6494477 up Hs.517033 TGM2 transglutaminase 2 (C polypeptide, protein-glutamine-gamma-
glutamyltransferase) 201534_s_at 2.166673 up Hs.145575 UBL3 ubiquitin-like 3 201631_s_at 3.5161445 up Hs.591785 IER3 immediate early response 3 201656_at 4.4946804 up Hs.133397 ITGA6 integrin, alpha 6 201925_s_at 2.3021228 up Hs.527653 CD55 CD55 molecule, decay accelerating factor for complement
(Cromer blood group) 201963_at 2.0353825 up Hs.406678 ACSL1 acyl-CoA synthetase long-chain family member 1 202241_at 3.248599 up Hs.444947 TRIB1 tribbles homolog 1 (Drosophila) 202284_s_at 2.624664 up Hs.370771 CDKN1A cyclin-dependent kinase inhibitor 1A (p21, Cip1) 202308_at 2.7079668 up Hs.592123 SREBF1 sterol regulatory element binding transcription factor 1 202481_at 6.214064 up Hs.289347 DHRS3 dehydrogenase/reductase (SDR family) member 3 202531_at 2.2103148 up Hs.436061 IRF1 interferon regulatory factor 1 202625_at 2.0354729 up Hs.699154 LYN v-yes-1 Yamaguchi sarcoma viral related oncogene homolog 202626_s_at 2.0061784 up Hs.699154 LYN v-yes-1 Yamaguchi sarcoma viral related oncogene homolog 202627_s_at 2.331629 up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E (nexin, plasminogen
activator inhibitor type 1), member 1 202628_s_at 2.2300718 up Hs.414795 SERPINE1 serpin peptidase inhibitor, clade E (nexin, plasminogen
293
activator inhibitor type 1), member 1 202769_at 2.736456 up Hs.13291 CCNG2 cyclin G2 202770_s_at 2.415966 up Hs.13291 CCNG2 cyclin G2 202869_at 3.5392282 up Hs.524760 OAS1 2',5'-oligoadenylate synthetase 1, 40/46kDa 202877_s_at 2.0917723 up Hs.97199 CD93 CD93 molecule 202887_s_at 21.188093 up Hs.523012 DDIT4 DNA-damage-inducible transcript 4 203020_at 2.2809913 up Hs.585378 RABGAP1L RAB GTPase activating protein 1-like 203139_at 2.2186103 up Hs.380277 DAPK1 death-associated protein kinase 1 203140_at 3.248891 up Hs.478588 BCL6 B-cell CLL/lymphoma 6 (zinc finger protein 51) 203234_at 2.2725453 up Hs.488240 UPP1 uridine phosphorylase 1 203243_s_at 2.3842292 up Hs.480311 PDLIM5 PDZ and LIM domain 5 203610_s_at 2.0810134 up Hs.584851 TRIM38 tripartite motif-containing 38 203708_at 2.0438766 up Hs.198072 PDE4B phosphodiesterase 4B, cAMP-specific (phosphodiesterase E4
dunce homolog, Drosophila) 203760_s_at 8.513409 up Hs.75367 SLA Src-like-adaptor 203761_at 9.117418 up Hs.75367 SLA Src-like-adaptor 203887_s_at 10.292902 up Hs.2030 THBD thrombomodulin 203888_at 7.23664 up Hs.2030 THBD thrombomodulin 204112_s_at 2.616031 up Hs.42151 HNMT histamine N-methyltransferase 204174_at 2.9158792 up Hs.507658 ALOX5AP arachidonate 5-lipoxygenase-activating protein 204429_s_at 4.854275 up Hs.530003 SLC2A5 solute carrier family 2 (facilitated glucose/fructose
transporter), member 5 204430_s_at 5.3099704 up Hs.530003 SLC2A5 solute carrier family 2 (facilitated glucose/fructose
transporter), member 5 204526_s_at 2.2889469 up Hs.442657 TBC1D8 TBC1 domain family, member 8 (with GRAM domain) 204961_s_at 7.9495873 up Hs.655201 LOC648998 ///
(biliary glycoprotein) 209593_s_at 2.6336179 up Hs.252682 TOR1B torsin family 1, member B (torsin B) 209930_s_at 2.3035018 up Hs.75643 NFE2 nuclear factor (erythroid-derived 2), 45kDa 209955_s_at 2.3653529 up Hs.654370 FAP fibroblast activation protein, alpha 209993_at 2.184716 up Hs.489033 ABCB1 ATP-binding cassette, sub-family B (MDR/TAP), member 1 210002_at 2.236731 up Hs.514746 GATA6 GATA binding protein 6 210004_at 3.3632 up Hs.412484 OLR1 oxidized low density lipoprotein (lectin-like) receptor 1 210146_x_at 2.7182057 up Hs.655652 LILRB2 leukocyte immunoglobulin-like receptor, subfamily B (with
TM and ITIM domains), member 2 210264_at 3.1211681 up Hs.239891 GPR35 G protein-coupled receptor 35 210357_s_at 5.933463 up Hs.433337 SMOX spermine oxidase 210367_s_at 2.0101428 up Hs.146688 PTGES prostaglandin E synthase 211026_s_at 3.044924 up Hs.277035 MGLL monoglyceride lipase 211192_s_at 2.0021026 up Hs.398093 CD84 CD84 molecule 211352_s_at 2.2821076 up Hs.697989 NCOA3 nuclear receptor coactivator 3 211559_s_at 2.1989374 up Hs.13291 CCNG2 cyclin G2 211913_s_at 4.0624857 up Hs.306178 MERTK c-mer proto-oncogene tyrosine kinase 212226_s_at 2.6206923 up Hs.405156 PPAP2B phosphatidic acid phosphatase type 2B 212230_at 2.7832167 up Hs.708050 PPAP2B phosphatidic acid phosphatase type 2B 212423_at 2.0793097 up Hs.523080 C10orf56 chromosome 10 open reading frame 56 212501_at 3.7421806 up Hs.517106 CEBPB CCAAT/enhancer binding protein (C/EBP), beta 212769_at 3.719843 up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog,
296
Drosophila) 212770_at 3.0558388 up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog,
Drosophila) 214084_x_at 8.358969 up LOC648998 similar to Neutrophil cytosol factor 1 (NCF-1) (Neutrophil
214523_at 4.19656 up Hs.558308 CEBPE CCAAT/enhancer binding protein (C/EBP), epsilon 214639_s_at 2.718203 up Hs.67397 HOXA1 homeobox A1 214708_at 2.0389485 up Hs.655236 SNTB1 syntrophin, beta 1 (dystrophin-associated protein A1, 59kDa,
basic component 1) 214977_at 3.1757998 up Hs.654670 CDNA FLJ13790 fis, clone THYRO1000026 215177_s_at 5.242455 up Hs.133397 ITGA6 integrin, alpha 6 215342_s_at 3.401339 up Hs.585378 RABGAP1L RAB GTPase activating protein 1-like 217997_at 2.2053015 up Hs.602085 PHLDA1 pleckstrin homology-like domain, family A, member 1 218284_at 2.2188303 up Hs.618504 SMAD3 SMAD family member 3 218501_at 9.380894 up Hs.476402 ARHGEF3 Rho guanine nucleotide exchange factor (GEF) 3 218559_s_at 3.3078525 up Hs.712609 MAFB v-maf musculoaponeurotic fibrosarcoma oncogene homolog B
(avian) 218627_at 2.7044392 up Hs.525634 DRAM damage-regulated autophagy modulator 218723_s_at 2.6658025 up Hs.507866 C13orf15 chromosome 13 open reading frame 15 219010_at 5.1608343 up Hs.518997 C1orf106 chromosome 1 open reading frame 106 219607_s_at 16.446184 up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 219777_at 2.1516159 up Hs.647105 GIMAP6 GTPase, IMAP family member 6 219994_at 2.9461663 up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B,
member 1 interacting protein 220005_at 7.095176 up Hs.546396 P2RY13 purinergic receptor P2Y, G-protein coupled, 13 220331_at 2.0530155 up Hs.25121 CYP46A1 cytochrome P450, family 46, subfamily A, polypeptide 1 220507_s_at 2.195543 up Hs.474388 UPB1 ureidopropionase, beta 221249_s_at 2.186694 up Hs.514308 FAM117A family with sequence similarity 117, member A 221266_s_at 3.4274688 up Hs.652230 TM7SF4 transmembrane 7 superfamily member 4 221345_at 6.744044 up Hs.248056 FFAR2 free fatty acid receptor 2
221601_s_at 3.567575 up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 221602_s_at 2.992049 up Hs.58831 FAIM3 Fas apoptotic inhibitory molecule 3 31845_at 2.0390604 up Hs.271940 ELF4 E74-like factor 4 (ets domain transcription factor) 36711_at 6.898643 up Hs.517617 MAFF v-maf musculoaponeurotic fibrosarcoma oncogene homolog F
(avian) 37152_at 2.1214638 up Hs.696032 PPARD peroxisome proliferator-activated receptor delta 222496_s_at 2.0052018 up Hs.518727 RBM47 RNA binding motif protein 47 223276_at 4.3521748 up Hs.29444 MST150 MSTP150 223567_at 6.3490977 up Hs.465642 SEMA6B sema domain, transmembrane domain (TM), and cytoplasmic
domain, (semaphorin) 6B 223620_at 3.3700576 up Hs.495989 GPR34 G protein-coupled receptor 34 223634_at 2.6509604 up Hs.474711 RASD2 RASD family, member 2 224048_at 2.2545755 up Hs.646421 USP44 ubiquitin specific peptidase 44 224916_at 2.9386487 up Hs.379754 TMEM173 transmembrane protein 173 224929_at 2.826263 up Hs.379754 TMEM173 transmembrane protein 173 224964_s_at 4.401167 up Hs.187772 GNG2 guanine nucleotide binding protein (G protein), gamma 2 225102_at 3.0217226 up Hs.277035 MGLL monoglyceride lipase 225347_at 4.439735 up Hs.497399 ARL8A ADP-ribosylation factor-like 8A 225372_at 5.093642 up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225373_at 3.0249772 up Hs.47382 C10orf54 chromosome 10 open reading frame 54 225763_at 3.3711896 up Hs.493867 RCSD1 RCSD domain containing 1 225919_s_at 2.1912513 up Hs.493639 C9orf72 chromosome 9 open reading frame 72 226184_at 2.1509264 up Hs.654630 FMNL2 formin-like 2 226487_at 3.614415 up Hs.661785 C12orf34 chromosome 12 open reading frame 34 226722_at 2.898792 up Hs.134742 FAM20C family with sequence similarity 20, member C 226756_at 3.2073371 up Hs.633903 CDNA FLJ25556 fis, clone JTH02629
298
226855_at 2.2713258 up Hs.632214 CDNA FLJ40954 fis, clone UTERU2010525 227290_at 2.108611 up Hs.60257 CDNA FLJ13598 fis, clone PLACE1009921 227396_at 2.0542467 up Hs.318547 PTPRJ protein tyrosine phosphatase, receptor type, J 227484_at 2.6185803 up Hs.710097 CDNA FLJ41690 fis, clone HCASM2009405 227792_at 3.046168 up Hs.648523 LOC162073 hypothetical protein LOC162073 227915_at 3.4160256 up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 228055_at 3.244248 up Hs.636624 NAPSB napsin B aspartic peptidase pseudogene 228056_s_at 2.75491 up Hs.636624 NAPSB napsin B aspartic peptidase pseudogene 228083_at 2.5030475 up Hs.13768 CACNA2D4 calcium channel, voltage-dependent, alpha 2/delta subunit 4 228120_at 3.458878 up Hs.656677 CDNA: FLJ22073 fis, clone HEP11868 228320_x_at 2.1715071 up Hs.369763 CCDC64 coiled-coil domain containing 64 228340_at 3.3133652 up Hs.709205 TLE3 transducin-like enhancer of split 3 (E(sp1) homolog,
Drosophila) 228361_at 2.0575192 up Hs.194333 E2F2 E2F transcription factor 2 228439_at 2.1450005 up Hs.124840 BATF2 basic leucine zipper transcription factor, ATF-like 2 228461_at 2.023712 up Hs.535157 SH3MD4 SH3 multiple domains 4 228479_at 2.0592828 up Hs.445588 Transcribed locus 228642_at 2.1591246 up Hs.445239 CDNA FLJ12777 fis, clone NT2RP2001720 228648_at 5.1974597 up Hs.655559 LRG1 leucine-rich alpha-2-glycoprotein 1 228772_at 2.7277112 up Hs.42151 HNMT histamine N-methyltransferase 228964_at 2.4908414 up Hs.436023 PRDM1 PR domain containing 1, with ZNF domain 229521_at 2.2940986 up Hs.29692 FLJ36031 hypothetical protein FLJ36031 229670_at 5.11233 up Hs.180284 5.5 kb mRNA upregulated in retinoic acid treated HL-60
neutrophilic cells 229934_at 2.499428 up Hs.38218 Mir-223 transcript variant 1 mRNA, complete sequence 229971_at 2.8952053 up Hs.187884 GPR114 G protein-coupled receptor 114 230218_at 6.501 up Hs.72956 HIC1 hypermethylated in cancer 1 230333_at 3.0846827 up Hs.656630 Transcribed locus 230391_at 2.5469894 up Hs.439064 Transcribed locus 230925_at 3.376848 up Hs.310421 APBB1IP amyloid beta (A4) precursor protein-binding, family B,
member 1 interacting protein 230966_at 2.0041862 up IL4I1 interleukin 4 induced 1
299
231214_at 2.4020123 up Hs.21278 Transcribed locus 231496_at 3.0144367 up Hs.145519 FCAMR Fc receptor, IgA, IgM, high affinity 231779_at 3.3791144 up Hs.449207 IRAK2 interleukin-1 receptor-associated kinase 2 231969_at 2.0790067 up Hs.21958 STOX2 storkhead box 2 232687_at 2.5030003 up Hs.100912 CDNA FLJ33091 fis, clone TRACH2000660 232861_at 2.6882606 up Hs.654693 PDP2 pyruvate dehydrogenase phosphatase isoenzyme 2 233857_s_at 3.7193377 up Hs.510327 ASB2 ankyrin repeat and SOCS box-containing 2 234643_x_at 2.1369362 up Hs.612905 CDNA: FLJ21798 fis, clone HEP00573 234987_at 3.4757702 up Hs.660221 Transcribed locus 235352_at 2.266094 up Hs.13500 CDNA FLJ31593 fis, clone NT2RI2002481 235360_at 2.3207605 up LOC100131989 hypothetical LOC100131989 235421_at 2.6853423 up Hs.432453///
Hs.663033 MAP3K8 Mitogen-activated protein kinase kinase kinase 8 /// CDNA
clone IMAGE:4689481 235529_x_at 2.3057177 up Hs.660221 Transcribed locus 235964_x_at 2.5259798 up Hs.660221 Transcribed locus 236191_at 11.936588 up Hs.667427 Transcribed locus 236407_at 10.117586 up Hs.121495 KCNE1 potassium voltage-gated channel, Isk-related family, member 1 236646_at 2.4213948 up Hs.226422 C12orf59 chromosome 12 open reading frame 59 236717_at 3.0867143 up Hs.525977 LOC165186 similar to RIKEN cDNA 4632412N22 gene 237201_at 2.1277235 up Hs.10305 Transcribed locus 237252_at 5.575421 up Hs.2030 THBD thrombomodulin 237442_at 2.4092128 up 237458_at 2.5117145 up Hs.547058 Transcribed locus 238032_at 4.0849333 up Hs.655631 Transcribed locus 238439_at 4.4300766 up Hs.217484 ANKRD22 ankyrin repeat domain 22 238669_at 2.2470448 up Hs.201978 PTGS1 prostaglandin-endoperoxide synthase 1 (prostaglandin G/H
synthase and cyclooxygenase) 239196_at 2.0093157 up Hs.217484 ANKRD22 ankyrin repeat domain 22 239294_at 4.7089114 up Hs.561747 Transcribed locus 239328_at 2.1184776 up Hs.668429 CDNA FLJ35362 fis, clone SKMUS2000330 239448_at 2.04081 up Hs.658524 Transcribed locus
300
240173_at 2.0289366 up Hs.602127 Transcribed locus 240481_at 2.7046072 up Hs.673398 Transcribed locus 240991_at 2.360695 up Hs.660557 Transcribed locus 241929_at 2.690208 up Hs.656268 Transcribed locus 242426_at 2.4238043 up Hs.696574 NRG4 neuregulin 4 242525_at 4.2519603 up Hs.439122 Transcribed locus 243541_at 3.6648195 up Hs.55378 IL31RA interleukin 31 receptor A 243819_at 2.472962 up 244665_at 5.22692 up Hs.668855 Transcribed locus 244764_at 2.9311552 up Hs.648369 HIVEP3 Human immunodeficiency virus type I enhancer binding
protein 3 1552553_a_at 2.6524854 up Hs.574741 NLRC4 NLR family, CARD domain containing 4 1552690_a_at 2.105542 up Hs.13768 CACNA2D4 calcium channel, voltage-dependent, alpha 2/delta subunit 4 1553740_a_at 2.2439284 up Hs.449207 IRAK2 interleukin-1 receptor-associated kinase 2 1554240_a_at 2.2535973 up Hs.174103 ITGAL integrin, alpha L (antigen CD11A (p180), lymphocyte
function-associated antigen 1; alpha polypeptide) 1554783_s_at 2.046973 up Hs.655209 ARHGEF2 rho/rac guanine nucleotide exchange factor (GEF) 2 1555420_a_at 2.1541228 up Hs.471221 KLF7 Kruppel-like factor 7 (ubiquitous) 1555431_a_at 4.66268 up Hs.55378 IL31RA interleukin 31 receptor A 1555680_a_at 4.149489 up Hs.433337 SMOX spermine oxidase 1555728_a_at 11.893831 up Hs.325960 MS4A4A membrane-spanning 4-domains, subfamily A, member 4 1563745_a_at 4.603027 up Hs.309176 LOC283050 hypothetical LOC283050 1556698_a_at 2.9657874 up Hs.605082 GPRIN3 GPRIN family member 3 1557051_s_at 2.2071733 up Hs.445239 CDNA FLJ12777 fis, clone NT2RP2001720 1559391_s_at 2.1286786 up Hs.667269 Partial mRNA; ID EE2-8E 1559584_a_at 2.1855097 up Hs.331095 C16orf54 ///
hCG_1644884 chromosome 16 open reading frame 54 /// similar to chromosome 16 open reading frame 54
1559882_at 2.0875654 up Hs.580681 SAMHD1 SAM domain and HD domain 1 1559883_s_at 2.4523852 up Hs.580681 SAMHD1 SAM domain and HD domain 1 1568619_s_at 3.5348852 up Hs.648523 LOC162073 hypothetical protein LOC162073 1569942_at 5.638706 up Hs.385753 CDNA clone IMAGE:4796629 1570375_at 3.925189 up Hs.661265 CDNA FLJ41985 fis, clone SPLEN2014946
301
Appendix II: Characterization of in vitro cell line models of APL
In order to assess the downstream genetic targets of the variant APL fusions NPM-, and
NuMA-RARα, we utilized the pro-monocytic U937 cell line retrovirally transduced with
NuMA-RARA or NPM-RARA (Kamel-Reid et al., 2003). All U937-X-RARA lines were
tested to confirm mRNA and protein expression of X-RARA. The highest expressing
clones were selected for the experiments outlined in this thesis (Figure AII.1 A). GFP
positivity in cells was assessed by flow cytometry and used as a surrogate marker for
ensuring fusion expression in the majority of cells (Figure AII.1 B). Flow cytometry
profiling of control and fusion-expressing cells showed that the presence of the NPM-
RARA and NuMA-RARA did not have any significant effect on cellular
immunophenotype. Cells were positive for the myeloid marker CD11b, leukocyte marker
CD45, and moderately positive for the stem cell marker CD117 (Figure AII.1 C).
Response to All Trans Retinoic Acid (ATRA).
In order to establish the validity of all lines as APL model systems, we tested their ability
to undergo differentiation in response to pharmacological concentrations (1 µM) of
ATRA. Differentiation was assessed by flow cytometry using the myeloid differentiation
marker CD11b, after treatment with 1 µM ATRA in culture for 0-96 hrs. Like control
U937 cells, NPM-RARA and NuMA-RARA expressing U937 cells responded to ATRA
with induction of CD11b expression (Figure AII.1 D). ATRA response was also assessed
all cell lines at the transcriptional level, using gene expression changes in the ATRA
responsive maker C/EBPε. The earliest transcriptional changes in response to ATRA
were observed 4 hours post-treatment with a significant induction of C/EBPε expression
in all cell lines (Figure AII.1 E). This time point was used for subsequent examination of
ATRA-induced early global gene expression changes in APL cell lines (Chapter 3).
Our work has also made use of cells derived from a transgenic mouse model of APL
expressing the NuMA-RARA fusion under the control of the human cathepsin G
promoter (hCG-NuMA-RARA). This model was characterized in Sukhai et al 2004, and
bears similarities to the diasease phenotype evident in human APL patients. Mice had
302
increased granulopoesis in the bone marrow and accumulation of promyelocytes,
characteristic of impaired neutrophil differentiation.
303
Figure AII.1: Characterization of X-RARA Cell Line s.
A) Western blot indicating expression of NPM-RARA in U937-NPM-RARA cell line
and PML-RARA in the NB4 cell line. Levels of wild type RARA are also indicated
alongside the β-actin control.
B) Expression of the fusion containing construct was confirmed by assessing levels of
GFP positivity by flow cytometry. Cells were harvested and incubated with anti-CD11b,
and analyzed for expression of this myeloid marker along with expression of GFP.
C) U937-control and X-RARA cells were harvested and incubated with fluorescently
conjugated anti-CD11b, CD45, or CD117 antibodies and analyzed by flow cytometry to
confirm immunophenotype profiles after introduction of the fusions.
D) Validation and effects of pharmacological agents in U937-X-RARA cell lines.
Control and X-RARA cells were treated with 1mM ATRA, prior to analysis of CD11b
expression by flow cytometry using fluorescently conjugated CD11b antibodies.
E) Untreated and ATRA treated cells were harvested for RNA analysis at time points 0-
6hrs and expression of ATRA target gene, CEBPε was analyzed by quantitative real time
PCR.
304
Figure AII A-E: Characterization of X-RARA Cell Li nes.
PML-RARA (108kDa)
RARA (51kDa)
NPM-RARA (58kDa)
ββββ−−−−actin(43kDa)
U93
7-G
FP
U93
7-N
PM
-RA
RA
NB
4-P
ML
-RA
RA
A.
305
CD
11b
flu
ore
scen
ce NuMA-RARA
55%
66%
GFP mean fluorescence intensity
NPM-RARA
17%
91%
U937-control
65%
95%
B.
C.
CD11b Mean Fluorescence Intensity
NuMA-RARANPM-RARAU937-control
CD45 Mean Fluorescence Intensity
CD117 Mean Fluorescence Intensity
Cel
l Co
un
t
306
M1
U937
CD11b MFI
Cel
l co
un
tNPM-RARA
M1
UNT RA
NuMA-RARA
M1
UNT RAUNT RA
D.
E.U937-GFPU937-NPM-RARAU937-NuMA-RARA
Hours post ATRA treatment
UNT ATRA UNT ATRA UNT ATRA
307
308
References
Ablain, J., and de The, H. (2011). Revisiting the differentiation paradigm in acute
promyelocytic leukemia. Blood 117, 5795-5802.
Aggarwal, B. B. (2003). Signalling pathways of the TNF superfamily: a double-edged