Identification of heavy metal regulated genes from the root associated ascomycete Cadophora finlandica using a genomic microarray Markus GORFER*, Helene PERSAK, Harald BERGER 1 , Sabine BRYNDA 2 , Dragana BANDIAN, Joseph STRAUSS Department of Applied Genetics and Cell Biology, Fungal Genomics Unit, Austrian Research Centers and BOKU-University Vienna, Muthgasse 18, 1190 Vienna, Austria article info Article history: Received 6 July 2009 Received in revised form 4 September 2009 Accepted 15 September 2009 Available online 19 September 2009 Corresponding Editor: Geoffrey Michael Gadd Keywords: Cadmium Cadophora finlandica Genomic microarray Heavy metal Lead Zinc abstract The ascomycete Cadophora finlandica, which can form mycorrhizas with ectomycorrhizal and ericoid hosts, is commonly found in heavy metal polluted soils. To understand the se- lective advantage of this organism at contaminated sites heavy metal regulated genes from C. finlandica were investigated. For gene identification a strategy based on a genomic micro- array was chosen, which allows a rapid, genome-wide screening in genetically poorly char- acterized organisms. In a preliminary screen eleven plasmids covering eight distinct genomic regions and encoding a total of ten Cd-regulated genes were identified. Northern analyses with RNA from C. finlandica grown in the presence of either Cd, Pb or Zn revealed different transcription patterns in response to the heavy metals present in the growth me- dium. The Cd-regulated genes are predicted to encode several extracellular proteins with unknown functions, transporters, a centaurin-type regulator of intracellular membrane trafficking, a GNAT-family acetyltransferase and a B-type cyclin. ª 2009 The British Mycological Society. Published by Elsevier Ltd. All rights reserved. Introduction Soils are frequently contaminated by heavy metals (HMs), mostly due to industrial activities (Anonymus 2005). Indepen- dent of their origindeither from naturally metalliferous rock or from anthropogenic pollutiondHMs in soil pose a challenge to all kinds of soil organisms including bacteria, fungi, plants and animals. Most studies on how fungi deal with HMs were carried out with model ascomycetes like Saccharomyces cerevisiae (Fauchon et al. 2002), Candida albicans (Riggle & Kumamoto 2000) and Schizosaccharomyces pombe (Bae & Chen 2004; Chen et al. 2003; Coblenz & Wolf 1994; Ortiz et al. 1992) or with ecto- mycorrhizal (ECM) basidiomycetes (Bellion et al. 2006). Only few studies addressed HM gene regulation in soil inhabiting ascomycetes like the ericoid mycorrhizal (ERM) fungus Oidio- dendron maius (Vallino et al. 2005; Vallino et al. 2009). Fungi have developed a complex defence system to coun- teract cell toxicities by HM (for a recent review see Gadd 2007). The first barrier includes excreted substances like * Corresponding author. Tel.: þ43 50550 3627; fax: þ43 810 80 30 30. E-mail address: [email protected]1 Present address: The Wellcome Trust, Centre for Cell Biology, Edinburgh, UK. 2 Present address: Institut fu ¨ r Molekulare Biotechnologie GmbH, Dr.-Bohr-Gasse 3, 1030 Vienna, Austria. journal homepage: www.elsevier.com/locate/mycres mycological research 113 (2009) 1377–1388 0953-7562/$ – see front matter ª 2009 The British Mycological Society. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.mycres.2009.09.005
Transcript
m y c o l o g i c a l r e s e a r c h 1 1 3 ( 2 0 0 9 ) 1 3 7 7 – 1 3 8 8
journa l homepage : www.e l sev i er . com/ loca te /mycres
Identification of heavy metal regulated genes from the rootassociated ascomycete Cadophora finlandica usinga genomic microarray
Markus GORFER*, Helene PERSAK, Harald BERGER1, Sabine BRYNDA2,Dragana BANDIAN, Joseph STRAUSS
Department of Applied Genetics and Cell Biology, Fungal Genomics Unit, Austrian Research Centers
and BOKU-University Vienna, Muthgasse 18, 1190 Vienna, Austria
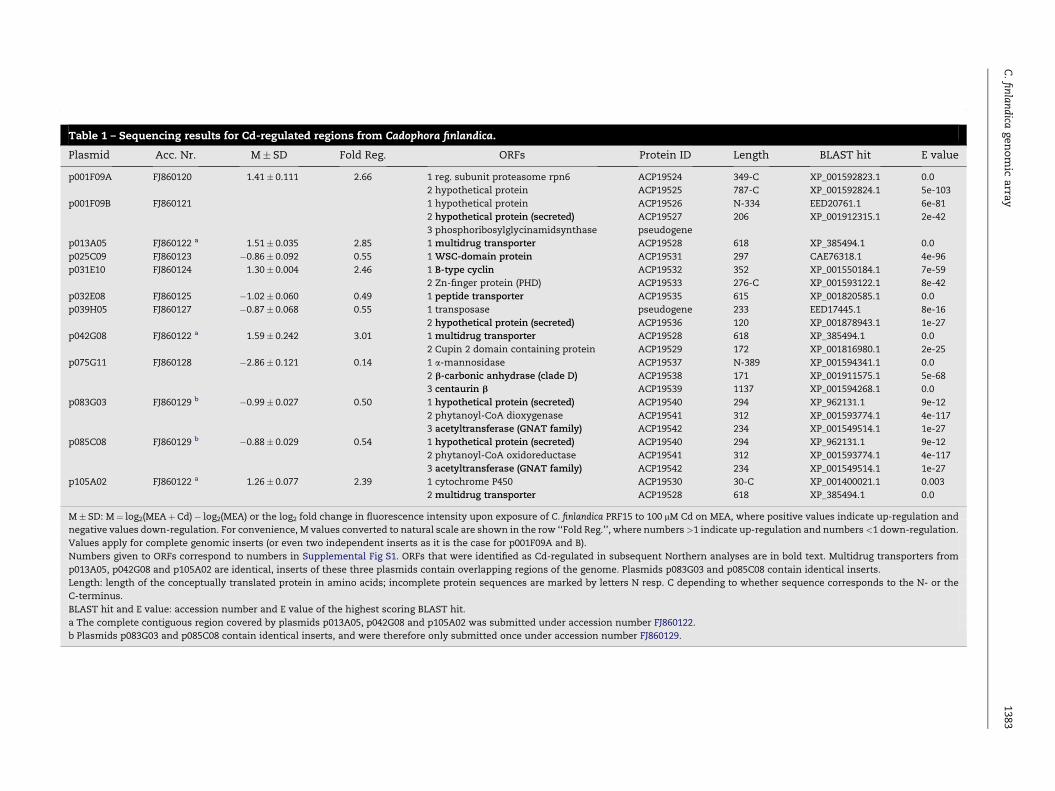
p083G03 FJ860129 b �0.99� 0.027 0.50 1 hypothetical protein (secreted)
2 phytanoyl-CoA dioxygenase
3 acetyltransferase (GNAT family)
ACP19540
ACP19541
ACP19542
294
312
234
XP_962131.1
XP_001593774.1
XP_001549514.1
9e-12
4e-117
1e-27
p085C08 FJ860129 b �0.88� 0.029 0.54 1 hypothetical protein (secreted)
2 phytanoyl-CoA oxidoreductase
3 acetyltransferase (GNAT family)
ACP19540
ACP19541
ACP19542
294
312
234
XP_962131.1
XP_001593774.1
XP_001549514.1
9e-12
4e-117
1e-27
p105A02 FJ860122 a 1.26� 0.077 2.39 1 cytochrome P450
2 multidrug transporter
ACP19530
ACP19528
30-C
618
XP_001400021.1
XP_385494.1
0.003
0.0
M� SD: M¼ log2(MEAþCd)� log2(MEA) or the log2 fold change in fluorescence intensity upon exposure of C. finlandica PRF15 to 100 mM Cd on MEA, where positive values indicate up-regulation and
negative values down-regulation. For convenience, M values converted to natural scale are shown in the row ‘‘Fold Reg.’’, where numbers>1 indicate up-regulation and numbers <1 down-regulation.
Values apply for complete genomic inserts (or even two independent inserts as it is the case for p001F09A and B).
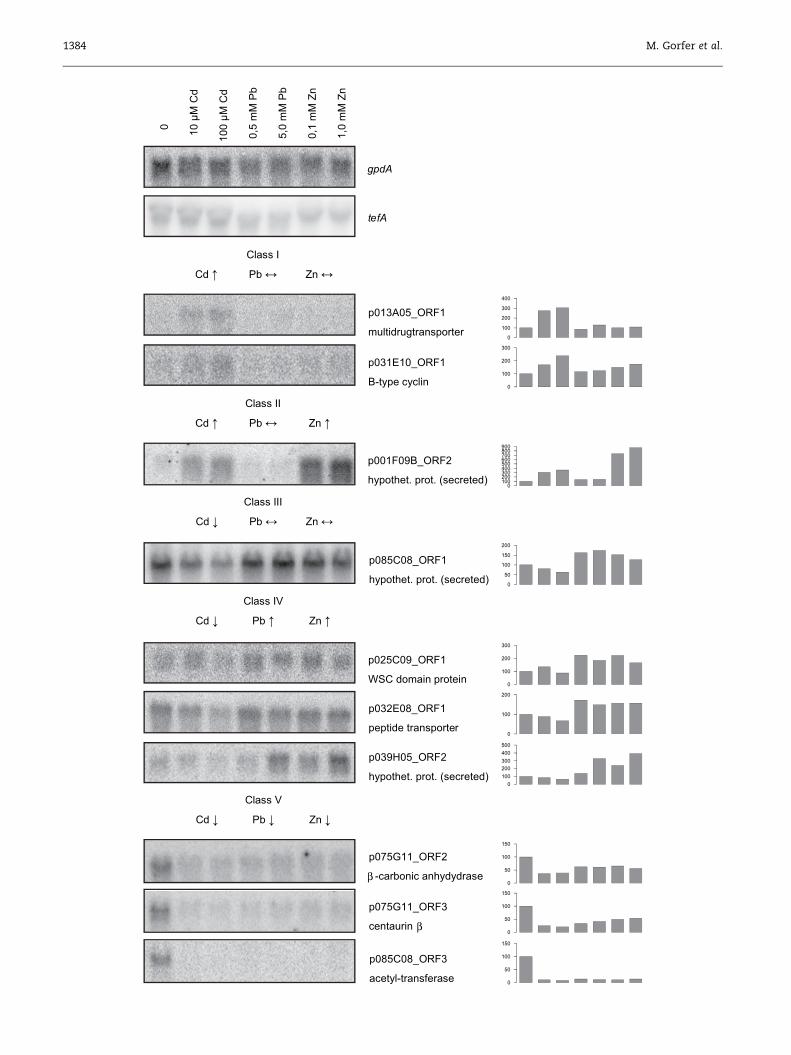
Numbers given to ORFs correspond to numbers in Supplemental Fig S1. ORFs that were identified as Cd-regulated in subsequent Northern analyses are in bold text. Multidrug transporters from
p013A05, p042G08 and p105A02 are identical, inserts of these three plasmids contain overlapping regions of the genome. Plasmids p083G03 and p085C08 contain identical inserts.
Length: length of the conceptually translated protein in amino acids; incomplete protein sequences are marked by letters N resp. C depending to whether sequence corresponds to the N- or the
C-terminus.
BLAST hit and E value: accession number and E value of the highest scoring BLAST hit.
a The complete contiguous region covered by plasmids p013A05, p042G08 and p105A02 was submitted under accession number FJ860122.
b Plasmids p083G03 and p085C08 contain identical inserts, and were therefore only submitted once under accession number FJ860129.
Al-Lahham A, Rohde V, Heim P, Leuchter R, Veeck J,Wunderlich C, Wolf K, Zimmermann M, 1999. Biosynthesis ofphytochelatins in the fission yeast. Phytochelatin synthesis:a second role for the glutathione synthetase gene of Schizo-saccharomyces pombe. Yeast 15: 385–396.
Anonymus, 2005. The European EnvironmentdState and Outlook2005. European Environment Agency, Copenhagen.
Bae W, Chen X, 2004. Proteomic study for the cellular responses toCd2þ in Schizosaccharomyces pombe through amino acid-codedmass tagging and liquid chromatography tandem mass spec-trometry. Molecular & Cellular Proteomics 3: 596–607.
Banerjee S, Flores-Rozas H, 2005. Cadmium inhibits mismatchrepair by blocking the ATPase activity of the MSH2–MSH6complex. Nucleic Acids Research 33: 1410–1419.
Bara MT, Lima AL, Ulhoa CJ, 2003. Purification and characteriza-tion of an exo-beta-1,3-glucanase produced by Trichodermaasperellum. FEMS Microbiology Letters 219: 81–85.
Bellion M, Courbot M, Jacob C, Blaudez D, Chalot M, 2006. Extra-cellular and cellular mechanisms sustaining metal tolerance inectomycorrhizal fungi. FEMS Microbiology Letters 254: 173–181.
Bellion M, Courbot M, Jacob C, Guinet F, Blaudez D, Chalot M,2007. Metal induction of a Paxillus involutus metallothioneinand its heterologous expression in Hebeloma cylindrosporum.New Phytologist 174: 151–158.
Bendtsen JD, Nielsen H, von Heijne G, Brunak S, 2004. Improvedprediction of signal peptides: SignalP 3.0. Journal of MolecularBiology 340: 783–795.
Berger H, Basheer A, Bock S, Reyes-Dominguez Y, Dalik T,Altmann F, Strauss J, 2008. Dissecting individual steps of ni-trogen transcription factor cooperation in the Aspergillus ni-dulans nitrate cluster. Molecular Microbiology 69: 1385–1398.
Bhanoori M, Venkateswerlu G, 2000. In vivo chitin-cadmiumcomplexation in cell wall of Neurospora crassa. Biochimica etBiophysica Acta 1523: 21–28.
Birnboim HC, Doly J, 1979. A rapid alkaline extraction procedurefor screening recombinant plasmid DNA. Nucleic Acids Research7: 1513–1523.
Chen D, Toone WM, Mata J, Lyne R, Burns G, Kivinen K, Brazma A,Jones N, Bahler J, 2003. Global transcriptional responses offission yeast to environmental stress. Molecular Biology of theCell 14: 214–229.
Church GM, Gilbert W, 1984. Genomic sequencing. Proceedings ofthe National Academy of Sciences of the United States of America 81:1991–1995.
Coblenz A, Wolf K, 1994. The role of glutathione biosynthesis inheavy metal resistance in the fission yeast Schizosaccharomycespombe. FEMS Microbiology Reviews 14: 303–308.
Ding Y, Bojja RS, Du L, 2004. Fum3p, a 2-ketoglutarate-dependentdioxygenase required for C-5 hydroxylation of fumonisins inFusarium verticillioides. Applied and Environmental Microbiology70: 1931–1934.
Dos Santos Utmazian MN, Schweiger P, Sommer P, Gorfer M,Strauss J, Wenzel WW, 2007. Influence of Cadophora finlandicaand other microbial treatments on cadmium and zinc uptakein willows grown on polluted soil. Plant, Soil and Environment53: 158–166.
Edelmann SE, Staben C, 1994. A statistical analysis of sequencefeatures within genes from Neurospora crassa. ExperimentalMycology 18: 70–81.
Enjalbert B, Smith DA, Cornell MJ, Alam I, Nicholls S, Brown AJP,Quinn J, 2006. Role of the Hog1 stress-activated protein ki-nase in the global transcriptional response to stress in thefungal pathogen Candida albicans. Molecular Biology of the Cell17: 1018–1032.
Fauchon M, Lagniel G, Aude JC, Lombardia L, Soularue P, Petat C,Marguerie G, Sentenac A, Werner M, Labarre J, 2002. Sulfursparing in the yeast proteome in response to sulfur demand.Molecular Cell 9: 713–723.
Fernandes L, Rodrigues-Pousada C, Struhl K, 1997. Yap, a novelfamily of eight bZIP proteins in Saccharomyces cerevisiae withdistinct biological functions. Molecular and Cellular Biology 17:6982–6993.
Fogarty RV, Tobin JM, 1996. Fungal melanins and their interac-tions with metals. Enzyme and Microbial Technology 19: 311–317.
Fomina M, Hillier S, Charnock JM, Melville K, Alexander IJ,Gadd GM, 2005. Role of oxalic acid overexcretion in transfor-mations of toxic metal minerals by Beauveria caledonica. Appliedand Environmental Microbiology 71: 371–381.
Frommer WB, Hummel S, Rentsch D, 1994. Cloning of an Arabi-dopsis histidine transporting protein related to nitrate andpeptide transporters. FEBS Letters 347: 185–189.
Gadd GM, 2007. Geomycology: biogeochemical transformations ofrocks, minerals, metals and radionuclides by fungi, biowea-thering and bioremediation. Mycological Research 111: 3–49.
Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB,Storz G, Botstein D, Brown PO, 2000. Genomic expressionprograms in the response of yeast cells to environmentalchanges. Molecular Biology of the Cell 11: 4241–4257.
Gebhart D, Bahrami AK, Sil A, 2006. Identification of a copper-inducible promoter for use in ectopic expression in the fungalpathogen Histoplasma capsulatum. Eukaryotic Cell 5: 935–944.
Goller SP, Gorfer M, Kubicek CP, 1998. Trichoderma reesei prs12encodes a stress- and unfolded-protein-response-inducibleregulatory subunit of the fungal 26S proteasome. CurrentGenetics 33: 284–290.
Gonzalez-Chavez MC, Carrillo-Gonzalez R, Wright SF, Nichols KA,2004. The role of glomalin, a protein produced by arbuscularmycorrhizal fungi, in sequestering potentially toxic elements.Environmental Pollution 130: 317–323.
Gorfer M, Klaubauf S, Bandian D, Strauss J, 2007. Cadophora fin-landica and Phialocephala fortinii: Agrobacterium-mediatedtransformation and functional GFP-expression. MycologicalResearch 111: 850–855.
Halliwell B, Gutteridge J, 1999. Free Radicals in Biology and Medicine.Oxford Science Publications, New York.
Harrington TC, McNew DL, 2003. Phylogenetic analysis places thePhialophora-like anamorph genus Cadophora in the Helotiales.Mycotaxon 87: 141–152.
Hirata D, Yano K, Miyakawa T, 1994. Stress-induced transcriptionalactivation mediated by YAP1 and YAP2 genes that encode theJun family of transcriptional activators in Saccharomyces cerevi-siae. Molecular and General Genetics 242: 250–256.
Hwang L, Hocking-Murray D, Bahrami AK, Andersson M, Rine J,Sil A, 2003. Identifying phase-specific genes in the fungalpathogen Histoplasma capsulatum using a genomic shotgunmicroarray. Molecular Biology of the Cell 14: 2314–2326.
Ivey FD, Magee DM, Woitaske MD, Johnston SA, Cox RA, 2003.Identification of a protective antigen of Coccidioides immitis byexpression library immunization. Vaccine 21: 4359–4367.
Jacob C, Courbot M, Brun A, Steinman HM, Jacquot JP, Botton B,Chalot M, 2001. Molecular cloning, characterization and reg-ulation by cadmium of a superoxide dismutase from the ec-tomycorrhizal fungus Paxillus involutus. European Journal ofBiochemistry 268: 3223–3232.
Jacob C, Courbot M, Martin F, Brun A, Chalot M, 2004. Transcrip-tomic responses to cadmium in the ectomycorrhizal fungusPaxillus involutus. FEBS Letters 576: 423–427.
Jekosch K, Kuck U, 1999. Codon bias in the ß-lactam producerAcremonium chrysogenum. Fungal Genetics Newsletter 46: 11–13.
Jungmann J, Reins HA, Schobert C, Jentsch S, 1993. Resistance tocadmium mediated by ubiquitin-dependent proteolysis. Na-ture 361: 369–371.
1388 M. Gorfer et al.
Lemmon MA, Ferguson KM, 2000. Signal-dependent membranetargeting by pleckstrin homology (PH) domains. BiochemicalJournal 350 (Pt 1): 1–18.
Li ZS, Lu YP, Zhen RG, Szczypka M, Thiele DJ, Rea PA, 1997. A newpathway for vacuolar cadmium sequestration in Saccharomy-ces cerevisiae: YCF1-catalyzed transport of bis(glutathionato)-cadmium. Proceedings of the National Academy of Sciences of theUnited States of America 94: 42–47.
Lloyd AT, Sharp PM, 1991. Codon usage in Aspergillus nidulans.Molecular and General Genetics 230: 288–294.
Mao Y, Varoglu M, Sherman DH, 1999. Molecular characterizationand analysis of the biosynthetic gene cluster for the antitumorantibiotic mitomycin C from Streptomyces lavendulae NRRL2564. Chemistry & Biology 6: 251–263.
Markham NR, Zuker M, 2005. DINAMelt web server for nucleicacid melting prediction. Nucleic Acids Research 33: W577–581.
Martin F, Aerts A, Ahren D, Brun A, Danchin EG, Duchaussoy F,Gibon J, Kohler A, Lindquist E, Pereda V, Salamov A,Shapiro HJ, Wuyts J, Blaudez D, Buee M, Brokstein P,Canback B, Cohen D, Courty PE, Coutinho PM, Delaruelle C,Detter JC, Deveau A, DiFazio S, Duplessis S, Fraissinet-Tachet L, Lucic E, Frey-Klett P, Fourrey C, Feussner I, Gay G,Grimwood J, Hoegger PJ, Jain P, Kilaru S, Labbe J, Lin YC,Legue V, Le Tacon F, Marmeisse R, Melayah D, Montanini B,Muratet M, Nehls U, Niculita-Hirzel H, Oudot-Le Secq MP,Peter M, Quesneville H, Rajashekar B, Reich M, Rouhier N,Schmutz J, Yin T, Chalot M, Henrissat B, Kues U, Lucas S, Vande Peer Y, Podila GK, Polle A, Pukkila PJ, Richardson PM,Rouze P, Sanders IR, Stajich JE, Tunlid A, Tuskan G,Grigoriev IV, 2008. The genome of Laccaria bicolor provides in-sights into mycorrhizal symbiosis. Nature 452: 88–92.
Martinez-Pastor MT, Marchler G, Schuller C, Marchler-Bauer A,Ruis H, Estruch F, 1996. The Saccharomyces cerevisiae zinc fingerproteins Msn2p and Msn4p are required for transcriptionalinduction through the stress response element (STRE). TheEMBO Journal 15: 2227–2235.
Meharg AA, 2003. The mechanistic basis of interactions betweenmycorrhizal associations and toxic metal cations. MycologicalResearch 107: 1253–1265.
Nie Z, Randazzo PA, 2006. Arf GAPs and membrane traffic. Journalof Cell Science 119: 1203–1211.
Nittler MP, Hocking-Murray D, Foo CK, Sil A, 2005. Identificationof Histoplasma capsulatum transcripts induced in response toreactive nitrogen species. Molecular Biology of the Cell 16:4792–4813.
O’Connell MJ, Osmani AH, Morris NR, Osmani SA, 1992. An extracopy of nimEcyclinB elevates pre-MPF levels and partially sup-presses mutation of nimTcdc25 in Aspergillus nidulans. The EMBOJournal 11: 2139–2149.
Ortiz DF, Kreppel L, Speiser DM, Scheel G, McDonald G, Ow DW,1992. Heavy metal tolerance in the fission yeast requires anATP-binding cassette-type vacuolar membrane transporter.The EMBO Journal 11: 3491–3499.
Peter BJ, Kent HM, Mills IG, Vallis Y, Butler PJ, Evans PR,McMahon HT, 2004. BAR domains as sensors of membranecurvature: the amphiphysin BAR structure. Science 303:495–499.
Plocke DJ, Kagi JH, 1992. Spectral characteristics of cadmium-containing phytochelatin complexes isolated from Schizosac-charomyces pombe. European Journal of Biochemistry 207: 201–205.
R Development Core Team. 2005. R Foundation for StatisticalComputing, Vienna, Austria.
Randazzo PA, Inoue H, Bharti S, 2007. Arf GAPs as regulators ofthe actin cytoskeleton. Biology of the Cell 99: 583–600.
Ren Q, Chen K, Paulsen IT, 2007. TransportDB: a comprehensivedatabase resource for cytoplasmic membrane transport sys-tems and outer membrane channels. Nucleic Acids Research 35:D274–279.
Riggle PJ, Kumamoto CA, 2000. Role of a Candida albicans P1-typeATPase in resistance to copper and silver ion toxicity. TheJournal of Bacteriology 182: 4899–4905.
Rozen S, Skaletsky HJ, 2000. Primer3 on the WWW for generalusers and for biologist programmers. In: Krawetz S, Misener S(eds), Bioinformatics Methods and Protocols: methods in molecularbiology. Humana Press, Totowa, NJ, pp. 365–386.
Sa-Correia I, dos Santos SC, Teixeira MC, Cabrito TR, Mira NP,2009. Drug:Hþ antiporters in chemical stress response inyeast. Trends in Microbiology 17: 22–31.
Sambrook J, Russell D, 2001. Molecular Cloning, A Laboratory Man-ual, 3rd edn. Cold Spring Harbor Laboratory Press, USA.
Stuart GW, Searle PF, Palmiter RD, 1985. Identification of multiplemetal regulatory elements in mouse metallothionein-I pro-moter by assaying synthetic sequences. Nature 317: 828–831.
Sugiura M, Georgescu MN, Takahashi M, 2007. A nitrite trans-porter associated with nitrite uptake by higher plant chloro-plasts. Plant and Cell Physiology 48: 1022–1035.
Thakur A, Kaur J, Bachhawat AK, 2008. Pgt1, a glutathionetransporter from the fission yeast Schizosaccharomyces pombe.FEMS Yeast Research 8: 916–929.
Vallee BL, Ulmer DD, 1972. Biochemical effects of mercury, cad-mium, and lead. Annual Review of Biochemistry 41: 91–128.
Vallino M, Drogo V, Abba’ S, Perotto S, 2005. Gene expression ofthe ericoid mycorrhizal fungus Oidiodendron maius in thepresence of high zinc concentrations. Mycorrhiza 15: 333–344.
Vallino M, Martino E, Boella F, Murat C, Chiapello M, Perotto S,2009. Cu, Zn superoxide dismutase and zinc stress in themetal-tolerant ericoid mycorrhizal fungus Oidiodendron maiusZn. FEMS Microbiology Letters 293: 48–57.
Vanden Wymelenberg A, Sabat G, Mozuch M, Kersten PJ, Cullen D,Blanchette RA, 2006. Structure, organization, and transcrip-tional regulation of a family of copper radical oxidase genes inthe lignin-degrading basidiomycete Phanerochaete chrysospo-rium. Applied and Environmental Microbiology 72: 4871–4877.
Vashisht AA, Kennedy PJ, Russell P, 2008. Centaurin-like proteinCnt5 contributes to arsenic and cadmium resistance in fissionyeast. FEMS Yeast Research 9: 257–269.
Vetting MW, de Carvalho LP, Yu M, Hegde SS, Magnet S,Roderick SL, Blanchard JS, 2005. Structure and functions of theGNAT superfamily of acetyltransferases. Archives of Biochem-istry and Biophysics 433: 212–226.
Vralstad T, Myhre E, Schumacher T, 2002. Molecular diversity andphylogenetic affinities of symbiotic root-associated ascomy-cetes of the Helotiales in burnt and metal polluted habitats.New Phytologist 155: 131–148.
Weissman Z, Berdicevsky I, Cavari BZ, Kornitzer D, 2000. The highcopper tolerance of Candida albicans is mediated by a P-typeATPase. Proceedings of the National Academy of Sciences of theUnited States of America 97: 3520–3525.
Wenzel U, Thwaites DT, Daniel H, 1995. Stereoselective uptake ofbeta-lactam antibiotics by the intestinal peptide transporter.British Journal of Pharmacology 116: 3021–3027.
Wenzel WW, Jockwer F, 1999. Accumulation of heavy metals inplants grown on mineralised soils of the Austrian Alps. Envi-ronmental Pollution 104: 145–155.
Zhou JJ, Theodoulou FL, Muldin I, Ingemarsson B, Miller AJ, 1998.Cloning and functional characterization of a Brassica napustransporter that is able to transport nitrate and histidine.Journal of Biological Chemistry 273: 12017–12023.
Zimmerman SA, Ferry JG, 2008. The beta and gamma classesof carbonic anhydrase. Current Pharmaceutical Design 14:716–721.