INFECTION AND IMMUNITY, Feb. 1981, p. 660-667 Vol. 31, No. 2 0019-9567/81/020660-08S02.00/0 Immunological Reactivity of Herpes Simplex Virus 1 and 2 Polypeptides Electrophoretically Separated and Transferred to Diazobenzyloxymethyl Paper B. NORRILD,' B. PEDERSEN,' AND B. ROIZMAN2* Institute of Medical Microbiology, University of Medical Microbiology, DK-2100 Copenhagen, Denmark,1 and Marjorie B. Kovler Viral Oncology Laboratories, University of Chicago, Chicago, Illinois 606372 In this paper we report that viral polypeptides from herpes simplex virus 1 (HSV-1) and 2 (HSV-2)-infected cells electrophoretically separated in sodium dodecyl sulfate-polyacrylamide-agarose gels and transferred to diazobenzyloxy- methyl paper can react with rabbit hyperimmune sera, both polyvalent and prepared against specific antigens. The polyvalent hyperimmune sera against HSV-1 reacted with 17 HSV-1 polypeptide bands and 8 HSV-2 polypeptide bands. Concordantly, polyvalent sera against HSV-2 reacted with at least 16 HSV-2 polypeptide bands and 8 HSV-1 polypeptide bands. The antisera prepared against the specific antigens reacted with a smaller number of polypeptide bands. Preim- mune sera and immune sera did not react with electrophoretically separated polypeptides from infected and uninfected cells, respectively. The immune local- ization of separated antigens test provides a powerful technique for identification of immunogenic viral polypeptides, especially those which are normally insoluble and therefore unavailable for immunological reactivity in immune precipitation tests. Herpes simplex viruses 1 and 2 (human her- pesvirases 1 and 2; HSV-1, HSV-2) are genet- ically and immunologically related. Their ge- nomes share 50% of the nucleotide sequences, with good matching of base pairs (7, 11-13, 21), and numerous studies have shown that cells infected with either one of the viruses contain antigens which react with antisera produced against both the homologous and heterologous virus (3-6, 8, 9, 23-25, 29). However, detailed analysis of the virus-specific infected cell poly- peptides (ICPs) has been restricted to only a subset of the approximately 50 polypeptides specified by each virus in the course of produc- tive infection. Predominant among these are the glycoproteins gA, gB, gC, gD, and gE, which are readily solubilized in nonionic detergents, and a small number of other polypeptides which are also soluble and therefore available for immune precipitation (2-5, 17, 18, 26-28). With respect to the glycoproteins, the results obtained to date show that HSV-1 gC reacts with antisera to HSV-1 only, whereas all other glycoproteins of HSV-1 are precipitable by antisera to both HSV- 1 and HSV-2 (26-28). Similar analyses have not been extended to other virus-specific ICPs, mainly because a large fraction of the ICPs are insoluble under conditions appropriate for im- munological reactivity and therefore their im- munological specificity cannot be readily meas- ured. In this paper we show that a large fraction of the ICPs transferred from sodium dodecyl sul- fate (SDS)-polyacrylamide-agarose gels to diaz- obenzyloxymethyl (DBM) paper according to the procedure of Renart et al. (20) retain their immunological reactivity. This technique, which we shall designate as immunological localization of separated antigens, can be used to study the immunological specificity of HSV-1- and HSV- 2-specific ICPs. (A preliminary account of this study was pre- sented at the International Conference on Hu- man Herpesviruses, Atlanta, Ga., March 1980.) MATERIALS AND METHODS Virus strains. HSV-1 strain F [HSV-1(F)] and HSV-2 strain G [HSV-2 (G)] were propagated in HEp- 2 cells grown in Eagle minimal essential medium sup- plemented with 1 % fetal calf serum (22). The mutant of HSV-1, HSV-1(HFEM)tsB5, was obtained from A. Buchan, Department of Microbiology, University of Birmingham, Birmingham, England (14). Infection of cells and labeling with radioiso- topes. Vero cells were propagated as monolayers in 25-cm2 tissue culture flasks in Eagle minimal essential medium with 10% fetal calf serum. The cells were infected with HSV-1(F), HSV-2(G), or HSV- l(HFEM)tsB5 in 0.5 ml at a multiplicity of infection of 10 plaque-forming units per cell. After adsorption at 34°C for 1 h, the inoculum was replaced with 5 ml of Eagle minimal essential medium supplemented with 1% fetal calf serum. 660 on September 17, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript
INFECTION AND IMMUNITY, Feb. 1981, p. 660-667 Vol. 31, No. 20019-9567/81/020660-08S02.00/0
Immunological Reactivity of Herpes Simplex Virus 1 and 2Polypeptides Electrophoretically Separated and Transferred
to Diazobenzyloxymethyl PaperB. NORRILD,' B. PEDERSEN,' AND B. ROIZMAN2*
Institute ofMedical Microbiology, University ofMedical Microbiology, DK-2100 Copenhagen, Denmark,1and Marjorie B. Kovler Viral Oncology Laboratories, University of Chicago, Chicago, Illinois 606372
In this paper we report that viral polypeptides from herpes simplex virus 1(HSV-1) and 2 (HSV-2)-infected cells electrophoretically separated in sodiumdodecyl sulfate-polyacrylamide-agarose gels and transferred to diazobenzyloxy-methyl paper can react with rabbit hyperimmune sera, both polyvalent andprepared against specific antigens. The polyvalent hyperimmune sera againstHSV-1 reacted with 17 HSV-1 polypeptide bands and 8 HSV-2 polypeptide bands.Concordantly, polyvalent sera against HSV-2 reacted with at least 16 HSV-2polypeptide bands and 8 HSV-1 polypeptide bands. The antisera prepared againstthe specific antigens reacted with a smaller number of polypeptide bands. Preim-mune sera and immune sera did not react with electrophoretically separatedpolypeptides from infected and uninfected cells, respectively. The immune local-ization of separated antigens test provides a powerful technique for identificationof immunogenic viral polypeptides, especially those which are normally insolubleand therefore unavailable for immunological reactivity in immune precipitationtests.
Herpes simplex viruses 1 and 2 (human her-pesvirases 1 and 2; HSV-1, HSV-2) are genet-ically and immunologically related. Their ge-nomes share 50% of the nucleotide sequences,with good matching of base pairs (7, 11-13, 21),and numerous studies have shown that cellsinfected with either one of the viruses containantigens which react with antisera producedagainst both the homologous and heterologousvirus (3-6, 8, 9, 23-25, 29). However, detailedanalysis of the virus-specific infected cell poly-peptides (ICPs) has been restricted to only asubset of the approximately 50 polypeptidesspecified by each virus in the course of produc-tive infection. Predominant among these are theglycoproteins gA, gB, gC, gD, and gE, which are
readily solubilized in nonionic detergents, and a
small number of other polypeptides which are
also soluble and therefore available for immuneprecipitation (2-5, 17, 18, 26-28). With respectto the glycoproteins, the results obtained to dateshow that HSV-1 gC reacts with antisera toHSV-1 only, whereas all other glycoproteins ofHSV-1 are precipitable by antisera to both HSV-1 and HSV-2 (26-28). Similar analyses have notbeen extended to other virus-specific ICPs,mainly because a large fraction of the ICPs are
insoluble under conditions appropriate for im-munological reactivity and therefore their im-munological specificity cannot be readily meas-
ured.
In this paper we show that a large fraction ofthe ICPs transferred from sodium dodecyl sul-fate (SDS)-polyacrylamide-agarose gels to diaz-obenzyloxymethyl (DBM) paper according tothe procedure of Renart et al. (20) retain theirimmunological reactivity. This technique, whichwe shall designate as immunological localizationof separated antigens, can be used to study theimmunological specificity of HSV-1- and HSV-2-specific ICPs.
(A preliminary account of this study was pre-sented at the International Conference on Hu-man Herpesviruses, Atlanta, Ga., March 1980.)
MATERIALS AND METHODSVirus strains. HSV-1 strain F [HSV-1(F)] and
HSV-2 strain G [HSV-2 (G)] were propagated in HEp-2 cells grown in Eagle minimal essential medium sup-plemented with 1 % fetal calf serum (22). The mutantof HSV-1, HSV-1(HFEM)tsB5, was obtained from A.Buchan, Department of Microbiology, University ofBirmingham, Birmingham, England (14).
Infection of cells and labeling with radioiso-topes. Vero cells were propagated as monolayers in25-cm2 tissue culture flasks in Eagle minimal essentialmedium with 10% fetal calf serum. The cells wereinfected with HSV-1(F), HSV-2(G), or HSV-l(HFEM)tsB5 in 0.5 ml at a multiplicity of infection of10 plaque-forming units per cell. After adsorption at34°C for 1 h, the inoculum was replaced with 5 ml ofEagle minimal essential medium supplemented with1% fetal calf serum.
For preparation of labeled viral glycoprotein, in-fected cells were incubated at 340C for 4 h, the mediumwas then aspirated, and 2.5 ml of fresh minimal essen-tial medium, supplemented with 1 % fetal calf serumand D-["4C]glucosamine (2 #Ci/ml; Amersham, Eng-land; specific activity, 60.8 mCi/mmol), was added.HSV-1(HFEM)tsBs-infected cells were incubatedfrom the end of the adsorption period at either 34 or390C as indicated in the text.
Labeling of viral proteins with L-[`S]methionine(New England Nuclear, Dreieich, West Germany; spe-cific activity, 756.86 Ci/mmol) was done in a mediumcontaining 1/10 of the amount of methionine con-tained in Eagle minimal essential medium. The label-ing of viral proteins was from 4 to 18 h postinfectionin all experiments.
Unlabeled viral proteins were prepared by extrac-tion of cells 18 h postinfection as described (15).
SDS-polyacrylamide-agarose gel electropho-resia. HSV-infected cell extracts were separated in9.25% (wt/vol) acrylamide gels cross-linked with 0.24%(wt/vol) N,N-diallyltartamide (Bio-Rad Labora-tories, Richmond, Calif.) and containing in addition1% (wt/vol) agarose (Litex, Copenhagen, Denmark) asdescribed by Renart et al. (20). The stacking gel was3% (wt/vol) acrylamide-0.08% (wt/vol) N,N-diallyltar-tardiamide. The buffer conditions and the proceduresfor staining, drying, and autoradiography were as pre-viously described (15). Before electrophoresis, the cellswere disrupted with 2% (wt/vol) SDS and 5% (vol/vol) 2-mercaptoethanol. The ICPs were numberedaccording to the nomenclature of Morse et al. (15),and the glycoproteins were numbered according to thenomenclature adapted at the 1979 Herpesvirus Work-shop held at Cold Spring Harbor, N.Y.
Transfer of HSV proteins to DBM paper. TheDBM paper was freshly prepared for each experimentfrom aminobenzyloxymethyl paper as described byAlwine et al. (1). The SDS-polyacrylamide agarose gelwas processed for the transfer of proteins largely asdescribed by Renart et al. (20). Briefly, the gel wastreated twice with 100 ml of 2% periodic acid for 30min at room temperature, followed by a 15- to 30-mintreatment with 0.5 M sodium phosphate buffer (pH7.5). Finally the gel was washed twice with 0.05 Msodium phosphate buffer (pH 7.5). Extraction of theproteins from the gel and immobilization by transferto the DBM paper were done according to the methodsdescribed by Southern for transfer of deoxyribonucleicacid (25). The transfer was done for 20 h at roomtemperature; after transfer the paper was soaked at370C for 2 h in a buffer containing 0.1 M tris-(hydroxymethyl)aminomethane-hydrochloride (pH9.0)-10% (vol/vol) ethanolamine-0.25% (wt/vol) gela-tin (19).
Antisera. Rabbit sera were used throughout thestudy. Sera 1 and 2 were hyperimmune to HSV-1 andHSV-2, respectively. The animals were injected witheither HSV-1- or HSV-2-infected rabbit cornea cellsgrown in rabbit serum. The sera neutralized bothHSV-1 and HSV-2 viruses, but neutralized the homol-ogous virus more strongly (28). These sera precipitatedviral proteins in crossed immunoelectrophoresis. andlysed both HSV-1- and HSV-2-infected cells in im-mune cytolytic tests (16). Activity against uninfected
cells was not demonstrable by any of the tests. Serum0 was obtained from nonimmunized animals.Serum 11 was a hyperimmune serum reactive with
glycoproteins gA and gB; it was produced by immu-nizing rabbits with immunoprecipitates of antigenfraction 11 (Ag-il) of HSV-1 cut out of agarose gels(29). The serum neutralized both HSV-1 and HSV-2,and in crossed immunoelectrophoretic tests it reactedwith both HSV-1 and HSV-2.Serum 8 was a hyperimmune serum reactive with
glycoprotein gD; it was produced by immunizing rab-bits with immunoprecipitates of Ag-8 of HSV-1 cutout of agarose gels (29). The antiserum neutralizedHSV-1 and HSV-2 and caused lysis of both HSV-1-and HSV-2-infected cells in immune cytolytic tests. Inimmune precipitation tests only gD and its precursorsof both HSV-1 and HSV-2 were precipitated by thisserum (16).Serum 6 was a hyperimmune serum reactive with
glycoprotein gC of HSV-1; it was produced by theimmunization of rabbits with Ag-6 of HSV-1. Thisserum neutralized HSV-1 but not HSV-2. The speci-ficity of the serum for HSV-1 was also demonstrablein immunoprecipitation and in immunocytolytic tests(16, 28).Serum 4 was obtained from a rabbit immunized
with antigen Ag-4 of HSV-2; it neutralized both HSV-1 and HSV-2, but the neutralization ofthe homologousHSV-2 was stronger. In crossed immunoelectropho-retic tests the serum reacted only with Ag-4 of HSV-2 (29). In antibody-dependent cell-mediated cytolysisthe serum mediated the lysis of both HSV-1- andHSV-2-infected cells. After extensive absorption withHSV-1-infected cells in monolayer culture, the serummediated the lysis of HSV-2-infected cells only. Thisabsorbed preparation was used in the present study.None of these rabbit sera reacted with uninfected
host cell antigens in any of the control tests done inparallel with tests designed to measure the reactivityof the sera with infected cell preparations.
Identification of HSV proteins immobilized bybinding toDBM paper. The paper strips correspond-ing to individual slots from the SDS-polyacrylamide-agarose gels were incubated in 10 ml of 1:10 dilutionsof the different rabbit hypernmmune sera as detailedin legends to figur. The sera were diluted in a 0.05M tris(hydroxymethyl)aminomethane-hydrochloridebuffer (pH 7.4) containing 0.15 M NaCl, 0.005 Methylenediaminetetraacetate, 0.25% (wt/vol) gelatin,and 0.05% (vol/vol) Triton X-100. The strips wereincubated at 37°C for 5 to 6 h, then washed in 50 mlof the same buffer for 18 h and incubated for 2 h inthe same buffer containing 0.25 yCi of 125I-labeledprotein A at 37°C (Amersham, England; specific activ-ity, 30 mCi/mg of protein). The final wash of the stripswas done for 2 h in the same buffer supplemented with1 M NaCl and 0.4% (vol/vol) Sarkosyl.
Autoradiographic images were developed on KodakXPR-1 film. Exposure was at 40C for 3 to 7 days.
RESULTSTransfer ofHSV proteins to DBM paper.
Figure 1 shows the autoradiographic images ofpolypeptides labeled in HSV-1-infected cells
A B C D from 4 to 21 h postinfection, electrophoreticallyseparated in SDS-polyacrylamide gels, thentransferred to DBM paper as described in Ma-terials and Methods. Comparisons of slots A, B,and C of Fig. 1, representing the autoradi-ographic images of polypeptides separated in apolyacrylamide-agarose gel and of the polypep-tides transferred to DBM paper after electro-phoretic separation, indicate that with few ex-ceptions the polypeptides transferred to DBM
5 paper reflected the molar abundance in the poly-6- _ _ acrylamide agarose gels and that the separation>8 i - of the polypeptides was not significantly im-
lopaired by the transfer. Analyses of the polyacryl-
w10 ^ =. - amide-agarose gels after transfer indicated thats1 l r1 no more than 50% of the polypeptides in each
15 : _ band were transferred to the DBM paper (datanot shown).
1 7 Immunological reactivity of HSV-1 poly-peptides after transfer to DBM paper. Un-labeled HSV-1 proteins were electrophoretically
20 separated in SDS-acrylamide-agarose gels,*;:transferred to the DBM paper, and then reactedIXwith HSV-1 serum (no. 1) as described in Ma-
[2 5 ^ s 8_w terials and Methods. As shown in Fig. 1, slot D,the antiserum reacted with most of the HSVpolypeptides immobilized on DBM paper. The
29 _ autoradiographic image shown in Fig. 1 wasproduced with dilutions of antisera as high as 1:
x! 200. Of particular interest is the observation thatamong the polypeptides strongly reactive with
3 the antiserum to HSV-1 were ICPs 5, 6, 8, 25,and 36, none of which is a surface-glycosylatedpolypeptide. It is noteworthy also that no im-munological reactivity was observed with ICPs10 and 20.
It should be emphasized that the specificity ofthe immune reaction was tested as follows. (i)Polypeptides from mock-infected cells were sep-
44 . arated in SDS-polyacrylamide-agarose gels,transferred to DBM paper, and incubated with
45 :wlFtI HSV-1 antibodies. No binding to host cell poly-peptides was demonstrable (data not shown). (ii)
35 3 5 125 Binding ofpreimmune rabbit serum to the HSV-S - S - 1 polypeptides was not demonstrable (Fig. 2, slot
Iu Me Me Prot-A D).Autoradiographic images ofelectrophoret- Reactivity of HSV-1 and HSV-2 immobi-aratedpolypeptides from cells infected with lized polypeptides with rabbit hyperim-). Slots A, B, and C are controls: (A) cell mune sera. Labeled and unlabeled HSV-1 and)m infected cells labeled with D-['4CJgluco- HSV-2 polypeptides from infected cells were
samine from 4 to 18 h postinfection; (B) cell lysatefrom infected cells labeled with L-[35SJmethioninefrom 4 to 18 h postinfection; (C) as (B) except thatafter electrophoresis in SDS-polyacrylamide-agarosegels thepolypeptides were transferred toDBMpaper.(D) Cell lysate from unlabeled cells harvested 18 hpostinfection, subjected to electrophoresis in SDS-polyacrylamide-agarose gels, transferred to DBMpa-per, and then reacted with serum no. 1, followed by
binding of 125I-labeled protein A. Slots A and Brepresent autoradiograms of the original SDS-poly-acrylamide-agarose gel. The autoradiograms ofslotsC and D represent images from polypeptides boundto DBM paper. The polypeptides were numbered as
described in the text. Note that the pattern in slot Cis a mirror image of the pattern shown in slot B.
FIG. 2. Autoradiographic images of electrophoretically separated polypeptides from cells infected withHSV-1(F). Slots A, B, and C represent controls as described in Fig. 1. Slots D to Jrepresentpolypeptides fromunlabeled cells harvested at 18 hpostinfection, electrophoretically separated on SDS-polyacrylamide-agarosegels, transferred to DBMpaper, and reacted with sera and then with "LI-labeledprotein A. The sera were as
follows: (D) serum 0; (E) serum 1; (F) serum 2; (G) sera 8 and 11; (H) serum 8; (I) serum 6; and (J) serum 4.
solubilized, electrophoretically separated on
SDS-polyacrylanide-agarose gels, and trans-ferred to DBM paper. The DBM-paper strips
with unlabeled polypeptides were then incu-bated with various preparations of hyperim-mune rabbit sera as specified in the legends to
the figures. The results shown in Fig. 2 and 3were as follows. The hyperimmune rabbit serumto HSV-1 (serum no. 1) reacted with approxi-mately 17 HSV-1 polypeptide bands but onlywith 8 HSV-2 polypeptide bands (Fig. 2, slot E;Fig. 3, slot E). Concordantly, the hyperimmunerabbit serum to HSV-2 proteins (serum no. 2)reacted with 16 polypeptide bands of HSV-2 and8 polypeptide bands of HSV-1 (Fig. 2, slot F;Fig. 3, slot F). Of interest is the observation thatthe HSV-1 CPs 5, 6, 8, 11, 25, 29, 39 or 40, and48 reacted with both HSV-1 and HSV-2 antisera,and therefore these polypeptides contained bothHSV-1 and HSV-2 antigen determinant sites. Incontrast, HSV-1 ICPs 15, 17, 36, and 37 boundantibody to HSV-1 (serum 1) only, and thereforethese polypeptides may contain HSV-1 antigenicdeterminant sites only. The HSV-2 ICPs 11, 18,20, 25, 36, 39, and 44 bound HSV-2 antiserum(serum 2) only, and therefore these polypeptidesmay contain HSV-2 antigenic deterninant sitesonly.The major glycoproteins of HSV-1 and HSV-
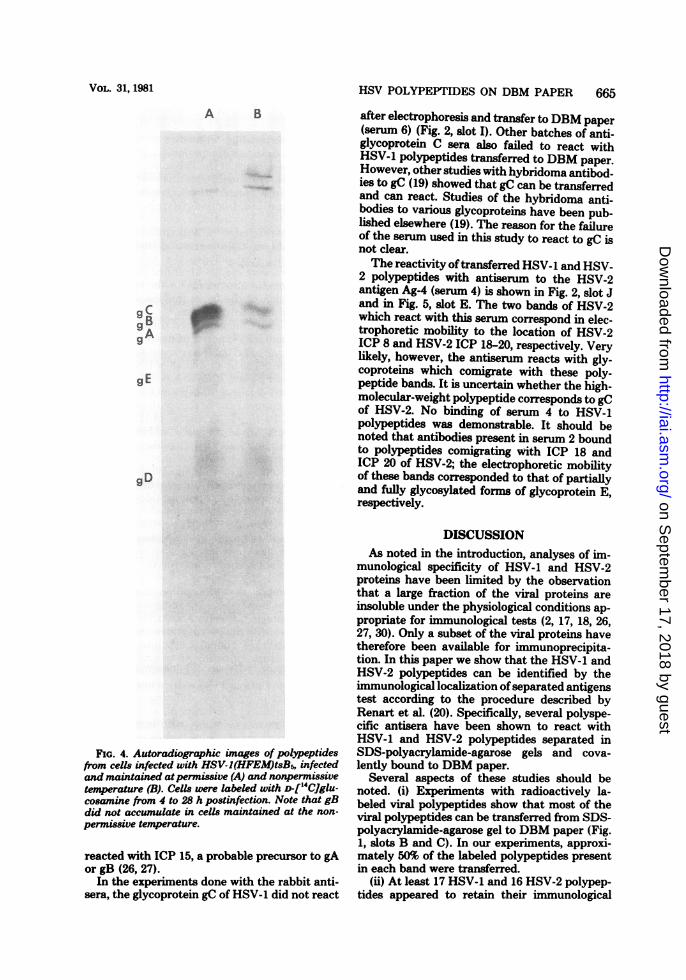
2 were identified among the immobilized pro-teins by the incubation of the paper strips withrabbit hyperimmune sera produced to the indi-vidual glycoproteins gD and gC and the mixtureof gA and gB. The resolution of glycoproteinsgA, gB, and gC in the gel system was analyzedby separation of ["4C]glucosamine-labeled gly-coproteins extracted from cells infected with themutant HSV-1(HFEM)tsB5. This mutant istemperature sensitive for the accumulation ofglycoprotein gB (14). Comparison of the electro-phoretic profile of proteins labeled at the per-missive temperature (Fig. 4, slot A) with that ofproteins labeled at the nonpermissive tempera-ture (Fig. 4, slot B) indicates that gA, gB, andgC were well separated. The results of the im-mune reactions were as follows.Serum no. 8 reacted strongly with glycopro-
tein gD of HSV-1 (Fig. 2, slot H). Three poly-peptide bands were visible, and the polypeptidebinding the antibodies most strongly was thepolypeptide ICP 29, which previously has beenshown to correspond to gD. The serum reactedless strongly with the band containing the poly-peptides with a lower apparent molecular weightthan that of ICP 29, but these polypeptidesmight be precursor molecules to ICP 29. Therelationship to glycoprotein gD of the poly-peptide with a higher molecular weight thanthat of ICP 29 is unknown.
Incubation of the electrophoretically sepa-rated polypeptides transferred to DMB-paperstrips with a mixture of sera 11 and 8 resulted inthe visualization of glycoproteins gA and gB inaddition to glycoprotein gD (ICP 29), as shownin Fig. 2, slot G. The reaction with gA wasstronger than that with gB. In addition, the sera
Original Gel Transferred
A B C D
A.*.
6.
gBm9A
39 ..
44 .....
45
14C- 35S 35 _ 1251kIu Me Me Protei n A
FIG. 3. Autoradiographic images ofelectrophoret-ically separatedpolypeptides from cells infected withHSV-2(G). Slots A, B, and C represent controls as
described in Fig. 1. Slots D to F represent polypep-tides from unlabeled cells harvested at 18 h postin-fection, electrophoretically separated on SDS-poly-acrylamide-agarose gels, transferred to DBMpaper,and reacted with sera and then with '251-labeledprotein A. The sera were as follows: (D) serum 0; (E)serum 1; (F) serum 2.
FIG. 4. Autoradiographic images of polyeptidesfrom cells infected with HSV-I(HFEM)tsB5, infectedand maintained atpermissive (A) and nonpermissivetemperature (B). Cells were labeled with D-114C]glu-cosamine from 4 to 28 h postinfection. Note that gBdid not accumulate in cells maintained at the non-
permissive temperature.
reacted with ICP 15, a probable precursor to gAor gB (26, 27).
In the experiments done with the rabbit anti-
sera, the glycoprotein gC of HSV-1 did not react
after electrophoresis and transfer to DBM paper(serum 6) (Fig. 2, slot I). Other batches of anti-glycoprotein C sera also failed to react withHSV-1 polypeptides transferred to DBM paper.However, other studies with hybridoma antibod-ies to gC (19) showed that gC can be transferredand can react. Studies of the hybridoma anti-bodies to various glycoproteins have been pub-lished elsewhere (19). The reason for the failureof the serum used in this study to react to gC isnot clear.The reactivity oftransferred HSV-1 and HSV-
2 polypeptides with antiserum to the HSV-2antigen Ag-4 (serum 4) is shown in Fig. 2, slot Jand in Fig. 5, slot E. The two bands of HSV-2which react with this serum correspond in elec-trophoretic mobility to the location of HSV-2ICP 8 and HSV-2 ICP 18-20, respectively. Verylikely, however, the antiserum reacts with gly-coproteins which comigrate with these poly-peptide bands. It is uncertain whether the high-molecular-weight polypeptide corresponds to gCof HSV-2. No binding of serum 4 to HSV-1polypeptides was demonstrable. It should benoted that antibodies present in serum 2 boundto polypeptides comigrating with ICP 18 andICP 20 of HSV-2; the electrophoretic mobilityof these bands corresponded to that of partiallyand fully glycosylated forms of glycoprotein E,respectively.
DISCUSSIONAs noted in the introduction, analyses of im-
munological specificity of HSV-1 and HSV-2proteins have been limited by the observationthat a large fraction of the viral proteins areinsoluble under the physiological conditions ap-propriate for immunological tests (2, 17, 18, 26,27, 30). Only a subset of the viral proteins havetherefore been available for immunoprecipita-tion. In this paper we show that the HSV-1 andHSV-2 polypeptides can be identified by theimmunological localization ofseparated antigenstest according to the procedure described byRenart et al. (20). Specifically, several polyspe-cific antisera have been shown to react withHSV-1 and HSV-2 polypeptides separated inSDS-polyacrylamide-agarose gels and cova-lently bound to DBM paper.
Several aspects of these studies should benoted. (i) Experiments with radioactively la-beled viral polypeptides show that most of theviral polypeptides can be transferred from SDS-polyacrylamide-agarose gel to DBM paper (Fig.1, slots B and C). In our experiments, approxi-mately 50% of the labeled polypeptides presentin each band were transferred.
(ii) At least 17 HSV-1 and 16 HSV-2 polypep-tides appeared to retain their immunological
FIG. 5. Autoradiographic images of electrophoret-ically separatedpolypeptides from cells infected withHSV-2(G). Slots A, B, and C represent controls as
described in Fig. 1. Slots D and E represent polypep-tides from unlabeled cells harvested at 18 h postin-fection, electrophoretically separated on SDS-poly-acrylamide-agarose gels, transferred to DBM paper,
and reacted with sera and then with '251-labeledprotein A. The sera were as follows: (D) serum 0; (E)serum 4.
reactivity after electrophoretic separation inSDS-polyacrylamide gels and transfer to DBMpaper (Fig. 2, slot E; Fig. 3, slot E). The failure
INFECT. IMMUN.
of other polypeptides to react with the homolo-gous hyperimmune sera 1 and 2 may be due tolack of corresponding antibodies in the poly-specific sera or to loss of immunological specific-ity as a consequence of the denaturation of thepolypeptides. We should note that none of theantisera in use reacted to electrophoreticallyseparated and DBM-bound polypeptides fromuninfected cells. Furthermore, none of the viralpolypeptides reacted with the preimmune rabbitsera. In addition, the observation that most ifnot all of the viral polypeptides that reactedwith antisera were readily identified by electro-phoretic mobility also argues for the specificityof the reaction of the hyperimmune sera withimmobilized viral polypeptides.
(iii) This study demonstrated that several ofthe HSV-1 and HSV-2 polypeptides reacted withboth homotypic and heterotypic antisera (ICPs5, 6, 8, 11, 25, 29, 39 or 40, and 48 of HSV-1 [Fig.2, slots E and F] and ICPs 5, 6, 8, 29, 39 or 40,and 48 of HSV-2 [Fig. 3, slots E and F]), whereasother polypeptides reacted only with homotypicantisera (ICPs 15, 17, 36, and 47 of HSV-1 andICPs 11, 18, 20, 25, 36, 39, and 44 of HSV-2 [Fig.1, slot E, and Fig. 2, slot F]). It is likely that thepolypeptides reactive with both homotypic andheterotypic antisera indeed contain type-specificas well as type-common antigen determinantsites. With respect to the polypeptides reactivewith homotypic sera, additional studies will haveto be done to establish whether heterotypic an-tigenic sites have been lost or whether the anti-sera used in this study for unknown reasons didnot contain antibodies to heterotypic antigenicsites.The immunological localization of separated
antigens test has several advantages comparedto immunoprecipitation tests. First, the test spe-cifically identifies the polypeptides carrying theantigen sites. This is of particular interest in thecase of proteins consisting of several differentpolypeptides. Second, the test identifies both theprecursor and products as well as immunologi-cally related polypeptides differing in electro-phoretic mobility. Furthermore, sensitivity ofthe test compares favorably with immune pre-cipitation tests inasmuch as it does not requirean antibody excess. For example, positive reac-tions were obtained with sera diluted severalhundred-fold. A major disadvantage of the testis that some polypeptides might lose their im-munological reactivity.The immunological localization of separated
antigens test has many obvious applications,especially in the identification of viral antigensto which antibodies are produced in the courseof human and experimental infections withHSV.
Studies done at the University of Copenhagen were sup-
ported by a grant from the NOVO Foundation. The studiesdone at the University of Chicago were supported by grantsfrom the National Cancer Institute (Public Health Servicegrants CA-08494-15 and CA 19264-05) and the American Can-cer Society (MV-2-P).
LITERATURE CITED
1. Alwine, J. C., D. J. Kemp, and G. R. Stark. 1977.Method for detection of specific RNA's in agarose gelsby transfer to diazobenzyloxymethyl paper and hybrid-ization with DNA probes. Proc. Natl. Acad. Sci. U.S.A.74:5350-5354.
2. Baucke, B., and P. G. Spear. 1979. Membrane proteinsspecified by herpes simplex viruses. V. Identification ofan Fc-binding glycoprotein. J. Virol. 32:779-789.
3. Cohen, G. H., M. Katze, C. Hydrean-Stern, and R. J.Eisenberg. 1978. Type-common CP-1 antigen ofherpessimplex virus is associated with a 59,000-molecular-weight envelope glycoprotein. J. Virol. 27:172-181.
4. Courtney, R. J., and K.L Powell. 1975. Immunologicaland biochemical characterization of polypeptides in-duced by herpes simplex virus types 1 and 2. IARC Sci.Publ. 11:63-73.
5. Eisenberg, R. J., C. Hydrean-Stern, and G. H. Cohen.1979. Structural analysis of precursor and product formsof type-common envelope glycoprotein D (CP-1 anti-gen) of herpes simplex virus type 1. J. Virol. 31:608-620.
6. Glorioso, J. C., L A. Wilson, T. W. Fenger, and J. W.Smith. 1978. Complement mediated cytolysis of HSV-1 and HSV-2 infected cells: plasma membrane antigensreactive with type-specific and cross-reactive antibody.J. Gen. Virol. 40:443-454.
7. Goodheart, C. R., G. Plummer, and J. L Waner. 1968.Density differences of DNA of human herpes simplexviruses types 1 and 2. Virology 35:473-475.
8. Honess, R. W., K.-L Powell, D. J. Robinson, C. Sim,and D. H. Watson. 1974. Type specific and type com-
mon antigens in cells infected with herpes simplex virustype 1 and on the surfaces of naked and envelopedparticles of the virus. J. Gen. Virol. 22:159-169.
9. Honess, R. W., and D. H. Watson. 1974. Herpes simplexvirus-specific polypeptides studied by polyacrylamidegel electrophoresis of immune precipitates. J. Gen. Vi-rol. 22:171-185.
10. Honess, R. W., and D. H. Watson. 1977. Review article.Unity and diversity in the herpesviruses. J. Gen. Virol.37:15-37.
11. Kieff, E. D., S. L Bachenheimer, and B. Roi2man.1971. Size, composition and structure of the DNA ofsubtypes 1 and 2 herpes simplex virus. J. Virol. 8:125-132.
12. Kieff, E. D., B. Hoyer, S. L Bachenheimer, and B.Roizman. 1972. Genetic relatedness of type 1 and type2 herpes simplex viruses. J. Virol. 9:738-745.
13. Ludwig, H. O., N. Biswal, and M. Benyesh-Melnick.1972. Studies of the relatedness of herpesvirusesthrough DNA-DNA hybridization. Virology 49:95-101.
14. Manservigi, R., P. G. Spear, and A. Buchan. 1977. Cellfusion induced by herpes simplex virus is promoted andsuppressed by different viral glycoproteins. Proc. Natl.Acad. Sci. U.S.A. 74:3913-3917.
15. Morse, L. S., L. Pereira, B. Roizman, and P. A. Schaf-
HSV POLYPEPTIDES ON DBM PAPER 667
fer. 197& Anatomy of herpes simplex virus (HSV)DNA. X. Mapping of viral genes by analysis of polypep-tides and functions specified by HSV-1 x HSV-2 recom-binants. J. Virol. 26:389-410.
16. Norrild, B., S. L Shore, and A. J. Nahmias. 1979.Herpes simplex virus glycoproteins: participation of in-dividual herpes simplex type 1 glycoprotein antigens inimmunocytolysis and their correlation with previouslyidentified glycopolypeptides. J. Virol. 32:741-748.
17. Norrild, B., and B. F. Vestergaard. 1977. Polyacryl-amide gel electrophoretic analysis of herpes simplexvirus type 1 immunoprecipitates obtained by quantita-tive immunoelectrophoresis in antibody-containing aga-rose gel. J. Virol. 22:113-117.
18. Norrild, B., and B. F. Vestergaard. 1979. Immunoelec-trophoretic identification and purification of herpessimplex virus antigens released from infected cells intissue culture. Intervirology 11:104-110.
19. Pereira, L, T. Klassen, and J. R. Baringer. 1980.Type-common and type-specific monoclonal antibodyto herpes simplex virus type 1. Infect. Immun. 29:724-732.
20. Renart, J., J. Reiser, and G. R. Stark. 1979. Transferof proteins from gels to diazobenzyloxymethyl-paperand detection with antisera: a method for studyingantibody specificity and antigen structure. Proc. Natl.Acad. Sci. U.S.A. 76:3116-3120.
21. Roizman, B., and D. Furlong. 1974. The replication ofherpesviruses. Compr. Virol. 3:229-403.
22. Roizman, B., and P. G. Spear. 1971. Herpes virus anti-gens on cell membranes detected by centrifugation ofmembrane-antibody complexes. Science 171:298-300.
23. Schneweis, K. E., and A. J. Nahmias. 1971. Antigensof herpes simplex virus type 1 and 2-immunodiffuLsionand inhibition passive hemagglutination studies. Z. Im-munitaetsforsch. Immunbiol. 141:471-487.
24. Shore, S. L, C. M. Black, F. M. Melewicz, P. A. Wood,and A. J. Nahmia8. 1969. Antibody-dependent cell-mediated cytotoxicity to target cells infected with type1 and type 2 herpes simplex virus. J. Immunol. 116:194-201.
25. Southern, E. M. 1975. Detection of specific sequencesamong DNA fragments separated by gel electrophore-sis. J. Mol. Biol. 98:503-517.
26. Spear, P. G. 1975. Glycoproteins specified by herpessimplex virus type 1: their synthesis, processing andantigenic relatedness to HSV-2 glycoproteins. IARCSci. Publ. 11:49-61.
27. Spear, P. G. 1976. Membrane proteins specified by herpessimplex viruses. I. Identification of four glycoproteinprecursors and their products in type 1-infected cells. J.Virol. 17:991-1008.
28. Vestergaard, B. F., and B. Norrild. 1978. Crossed im-munoelectrophoresis of a herpes simplex virus type 1-specific antigen: immunological and biochemical char-acterization. J. Infect. Dis. 138:639-643.
29. Vestergaard, B. F., and B. Norrild. 1979. Crossed im-munoelectrophoretic analysis and viral neutralizing ac-
tivity of five monospecific antisera against five differentherpes simplex virus glycoproteins. IARC Sci. Publ. 24:225-234.
30. Wilcox, K. W., A. Kohn, E. Sklyanskaya, and B.Roizman. 1980. Herpes simplex virus phosphoproteins.I. Phosphate cycles on and off some viral polypeptidesand can alter their affinity for DNA. J. Virol. 33:167-182.











![RESEARCHARTICLE ......communityabortion stigma andlowersocial support were associatedwithmore negative emotions(b=0.45[0.31,0.58]andb=-0.61[-0.93,-0.29], respectively). Conclusions](https://static.documents.pub/doc/80x56/5f525cae3954ef0fc232381c/researcharticle-communityabortion-stigma-andlowersocial-support-were-associatedwithmore.jpg)












