Implication of different effector mechanisms by cord bloodederived and peripheral bloodederived cytokine-induced killer cells to kill precursor B acute lymphoblastic leukemia cell lines LUDOVIC DURRIEU 1,2 , WILLIAM LEMIEUX 1,2 , MAME MASSAR DIENG 1 , FRANÇOIS FONTAINE 1 , MICHEL DUVAL 1,2,3 , FRANÇOISE LE DEIST 1,2,3 & ELIE HADDAD 1,2,3 1 CHU Sainte-Justine Research Center, Center of Cancerology Charles-Bruneau, Montreal, Quebec, Canada, and 2 Department of Microbiology and Immunology and 3 Department of Pediatrics, University of Montreal, Montreal, Quebec, Canada Abstract Background aims. Cytokine-induced killer (CIK) cells ex vivoeexpanded from cord blood (CB) or peripheral blood (PB) have been shown to be cytotoxic against autologous and allogeneic tumor cells. We have previously shown that CD56 þ CIK cells (CD3 þ CD56 þ and CD3 CD56 þ ) are capable of killing precursor B-cell acute lymphoblastic leukemia (B-ALL) cell lines. However, the lytic pathways used by CD56 þ PB and CB-CIK cells to kill B-ALL cell lines have not been studied. Methods. CB and PB-CIK cells were differentiated. CD56 þ CB- and PB-CIK cells were compared for expression of different phenotypic markers and for the lytic pathways used to kill B-ALL cell lines. Results. We found that cytotoxic granule proteins were expressed at higher levels in CD56 þ PB-CIK than in CD56 þ CB-CIK cells. However, CD56 þ CB-CIK cells expressed more tumor necrosis factorerelated apoptosis-inducing ligand (TRAIL) compared with CD56 þ PB-CIK cells. We observed that CD56 þ CB-CIK cells used both the NKG2D and TRAIL cytotoxic pathways and were more effective at killing REH cells than CD56 þ PB-CIK cells that used only the NKG2D pathway. In contrast, CD56 þ PB-CIK cells used both NKG2D and TRAIL pathways to kill NALM6 cells, whereas CD56 þ CB-CIK cells used only the NKG2D pathway. Conclusions. Our results suggest that both the source of CIK and the type of B-ALL cell line have an impact on the intensity of the cytolytic activity and on the pathway used. These findings may have clinical implications with respect to optimizing therapeutic efficacy, which may be dependent on the source of the CIK cells and on the target tumor cells. Key Words: cord blood, cytokine-induced killer cells, peripheral blood, precursor B acute lymphoblastic leukemia, NKG2D, TRAIL Introduction In vitroegenerated cytokine-induced killer (CIK) cells, first described by Schmidt-Wolf et al. (1,2), are a mix of three subpopulations: CD3 þ CD56 e , CD3 - CD56 þ and CD3 þ CD56 þ . They can be amplified from multiple cell sources, including pe- ripheral blood (PB) (1) and cord blood (CB) (3). The protocol involves culturing mononuclear cells in the presence of recombinant human interferon-g (rhIFNg), followed by the addition of an anti-CD3 monoclonal antibody (OKT3) and recombinant human interleukin 2 (rhIL2), which leads to a greatly increased expansion in the number of CD3 þ CD56 þ cells over a 3-week period. Lu and Negrin (4) have shown that this subpopulation is derived from CD3 þ CD56 e T cells that acquire the CD56 marker. There are a number of in vitro and in vivo studies that describe the ability of CIK cells to kill a variety of tumor cells (5e12). Furthermore, we and others have shown that the cytotoxic activity against tumor cells is associated with the CD56 þ cell subpopula- tion (that comprise mainly CD3 þ CD56 þ and some CD3 e CD56 þ cells) (13,14). CIK cells have potent major histocompatibility complexeunrestricted cytotoxic activity against tu- mor cells, which they carry out through several possible pathways. As in other lymphocyte functional subsets, adhesion through lymphocyte functione associated antigen 1 on CIK cells and intracellular adhesion molecule 1 on target cells is necessary for CIK cells to kill tumor cells (2). In addition, the NKG2D receptor (15) has been identified as playing a major role in the activation of CIK cell cytotoxic activity. NKG2D is a glycoprotein expressed on the Correspondence: Elie Haddad, MD, PhD, Service d’immunologie et rhumatologie pediatriques, CHU Sainte-Justine, 3175 Cote Sainte-Catherine, Montreal, Quebec H3T 1C5, Canada. E-mail: [email protected]Cytotherapy, 2014; 0: 1e12 (Received 6 June 2013; accepted 26 December 2013) ISSN 1465-3249 Copyright Ó 2014, International Society for Cellular Therapy. Published by Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.jcyt.2013.12.010
Transcript
Cytotherapy, 2014; 0: 1e12
Implication of different effector mechanisms by cord bloodederived andperipheral bloodederived cytokine-induced killer cells to kill precursorB acute lymphoblastic leukemia cell lines
LUDOVIC DURRIEU1,2, WILLIAM LEMIEUX1,2, MAME MASSAR DIENG1,FRANÇOIS FONTAINE1, MICHEL DUVAL1,2,3, FRANÇOISE LE DEIST1,2,3 &ELIE HADDAD1,2,3
1CHU Sainte-Justine Research Center, Center of Cancerology Charles-Bruneau, Montreal, Quebec, Canada, and2Department of Microbiology and Immunology and 3Department of Pediatrics, University of Montreal, Montreal,Quebec, Canada
AbstractBackground aims.Cytokine-induced killer (CIK) cells ex vivoeexpanded from cord blood (CB) or peripheral blood (PB) havebeen shown to be cytotoxic against autologous and allogeneic tumor cells. We have previously shown that CD56þ CIK cells(CD3þCD56þ and CD3�CD56þ) are capable of killing precursor B-cell acute lymphoblastic leukemia (B-ALL) cell lines.However, the lytic pathways used by CD56þ PB and CB-CIK cells to kill B-ALL cell lines have not been studied. Methods.CB and PB-CIK cells were differentiated. CD56þ CB- and PB-CIK cells were compared for expression of differentphenotypic markers and for the lytic pathways used to kill B-ALL cell lines. Results.We found that cytotoxic granule proteinswere expressed at higher levels in CD56þ PB-CIK than in CD56þ CB-CIK cells. However, CD56þ CB-CIK cells expressedmore tumor necrosis factorerelated apoptosis-inducing ligand (TRAIL) compared with CD56þ PB-CIK cells. We observedthat CD56þ CB-CIK cells used both the NKG2D and TRAIL cytotoxic pathways and were more effective at killing REHcells than CD56þ PB-CIK cells that used only the NKG2D pathway. In contrast, CD56þ PB-CIK cells used both NKG2Dand TRAIL pathways to kill NALM6 cells, whereas CD56þ CB-CIK cells used only the NKG2D pathway. Conclusions. Ourresults suggest that both the source of CIK and the type of B-ALL cell line have an impact on the intensity of the cytolyticactivity and on the pathway used. These findings may have clinical implications with respect to optimizing therapeuticefficacy, which may be dependent on the source of the CIK cells and on the target tumor cells.
In vitroegenerated cytokine-induced killer (CIK)cells, first described by Schmidt-Wolf et al. (1,2),are a mix of three subpopulations: CD3þCD56e,CD3-CD56þ and CD3þCD56þ. They can beamplified from multiple cell sources, including pe-ripheral blood (PB) (1) and cord blood (CB) (3).The protocol involves culturing mononuclear cells inthe presence of recombinant human interferon-g(rhIFNg), followed by the addition of an anti-CD3monoclonal antibody (OKT3) and recombinanthuman interleukin 2 (rhIL2), which leads to a greatlyincreased expansion in the number of CD3þCD56þ
cells over a 3-week period. Lu and Negrin (4) haveshown that this subpopulation is derived fromCD3þCD56e T cells that acquire the CD56 marker.There are a number of in vitro and in vivo studies that
Correspondence: Elie Haddad, MD, PhD, Service d’immunologie et rhumatologiQuebec H3T 1C5, Canada. E-mail: [email protected]
(Received 6 June 2013; accepted 26 December 2013)
ISSN 1465-3249 Copyright � 2014, International Society for Cellular Therapy. Phttp://dx.doi.org/10.1016/j.jcyt.2013.12.010
describe the ability of CIK cells to kill a variety oftumor cells (5e12). Furthermore, we and othershave shown that the cytotoxic activity against tumorcells is associated with the CD56þ cell subpopula-tion (that comprise mainly CD3þCD56þ and someCD3eCD56þ cells) (13,14).
CIK cells have potent major histocompatibilitycomplexeunrestricted cytotoxic activity against tu-mor cells, which they carry out through severalpossible pathways. As in other lymphocyte functionalsubsets, adhesion through lymphocyte functioneassociated antigen 1 on CIK cells and intracellularadhesion molecule 1 on target cells is necessary forCIK cells to kill tumor cells (2). In addition, theNKG2D receptor (15) has been identified as playinga major role in the activation of CIK cell cytotoxicactivity. NKG2D is a glycoprotein expressed on the
e pediatriques, CHU Sainte-Justine, 3175 Cote Sainte-Catherine, Montreal,
membrane of NK and T cells. The stimulation ofthese cells through the NKG2D pathway inducescytotoxicity on the target cells and can induce theproduction of cytokines (16). The implication ofNKG2D has been shown to play a role in the tumorsurveillance (17). This cell membrane receptor,which is present on CIK cells (14,18), recognizesligands expressed on tumor cells, mainly major his-tocompatibility complex class I chain-related gene A(MICA) and MICB (14,15). Proof that the NKG2Dreceptor plays an important role in CIK cellemedi-ated cytotoxic activity has been established throughthe use of NKG2D receptor blocking antibodiesthat inhibit CIK cells from killing several cell linesincluding myeloma (15) and B lymphoma (19). Inaddition, it has been shown that CIK cells can useother receptors to kill tumor cells. Cell signalingthrough one of these, natural killer p30 (NKp30), hasbeen shown to lead to CIK cell activation, resultingin tumor cell lysis (19). It has also been shown thatCIK cells can provoke lysis through the tumor ne-crosis factorerelated apoptosis-inducing ligand(TRAIL)/TRAIL-receptors pathway (20). TRAIL isa member of the tumor necrosis factor superfamilythat can provoke the apoptosis through activation ofits receptors (DR4 and DR5) on target cells. TheTRAIL pathway has also been shown to act in thetumor surveillance (21).
Although, as noted above, CIK cells can kill avariety of tumor cells, precursor B-cell acute lym-phoblastic leukemia (B-ALL) cell lines have provento be less sensitive to lysis mediated by CIK cells(22,23). However, we have recently shown thatwhereas bulk cordbloodCIK (CB-CIK) cells could killB-ALL cell lines, albeit weakly (13), the purifiedCD56þ cell fraction (CD56þ CB-CIK) could effec-tively lyse B-ALL cell lines. Furthermore, this activityresided mainly in the CD3þCD56þ CB-CIK subpop-ulation, whereas the CD3þCD56e CB-CIK cell frac-tion had no cytotoxic activity against these cell lines.
Various research groups working with humanCIK cells have derived them from either PB or CB(3,14,15). They have shown that CIK cells expressvarious markers including CD3, CD56, CD8,NKG2D and others markers (3,4,18). Nevertheless,no one has compared the expression of multiplemarkers on both CB-CIK and PB-CIK cells. Inaddition, whereas the CIK cell cytotoxic pathwaysused to kill various tumors has been reported(15,19,20), no study has specifically described thecytotoxic pathways used by PB- or CB-CIK cells tokill B-ALL cell lines. Therefore, we have investi-gated the phenotype of CD56þ CB-CIK and PB-CIK cells and the cytotoxic pathways involved inCD56þ CB-CIK and CD56þ PB-CIK cell-mediatedkilling of B-ALL cell lines.
Methods
Cells
Two cell lines were used as positive controls in cyt-otoxicity assays, K562, a myelogenous leukemia cellline (ATCC, Manassas, VA, USA), and U266, ahuman cell line multiple myeloma cell line (ATCC).Human B-cell precursor leukemia cell lines REH,NALM6 and KOPN-8 were purchased from DSMZ(Braunschweig, Germany) and 697 from ATCC. Allcell lines were maintained in Roswell Park MemorialInstitute (RPMI)-1640 medium supplemented with2 mmol/L L-glutamine (Life Technologies, Bur-lington, ON, Canada) and 10% fetal bovine serum(FBS) (Life Technologies).
Generation of CB-CIK and PB-CIK cells
CB units were obtained from the CHU Sainte-Justine Research CB bank, with approval from theethics committee. PB samples were collected fromconsenting healthy donors. CB mononuclear cells(CBMCs) and PB mononuclear cells (PBMCs) wereisolated with the use of Ficoll-Paque PLUS (GEHealthcare Bio-Science AB, Uppsala, Sweden).CBMCs and PBMCs were generated as previouslydescribed (13); they were cultured in X-VIVO 15medium plus FBS (10%) with 1000 IU/mL ofrhIFNg (Feldan Bio, Qu�ebec City, QC, Canada)added on day 1 and then supplemented with 50 ng/mLof OKT3 monoclonal antibody (eBioscience, SanDiego, CA, USA) and 300 IU/mL of rhIL2 (Novartis,Basel, Switzerland) on day 2. Over the culture period,cells were counted, stained, replated in flasks andstimulated with rhIL2 (300 IU/mL) every 3e4 days,and fresh medium was added as needed to maintain acell density of 0.8e1 � 106 cells/mL for a total of21e28 days.
Extracellular staining of CB-CIK and PB-CIK cells
CB-CIK and PB-CIK cells were stained with the useof anti-human CD3 (clone UCHT1) and anti-human CD56 (clone B159) antibodies. CD56þ cellswere characterized with the following antibodies:anti-human NKp30 (clone p30e15), anti-humanNKp44 (clone p44e8.1), anti-human NKp46 (clone9E2/NKp46), anti-human TRAIL (clone RIK-2), andanti-human NKG2D (clone 1D11). All antibodieswere purchased from BD Biosciences (Mississauga,ON, Canada) except for anti-human TRAIL, whichwas purchased from BioLegend (San Diego, CA,USA). Samples were acquired on a fluorescence-activated cell sorter (FACS) Fortessa flow cytometerand analyzed with the use of FACSDiva software
(BD Biosciences) or FlowJo (Tree Star Inc, Ashland,OR, USA).
Intracellular staining of CB-CIK and PB-CIK cells
To carry out the intracellular staining of CD56þ CBand CD56þ PB-CIK, cells were first stained with anti-human CD3 and anti-human CD56 for 30 min at4�C; they were then washed with phosphate-bufferedsaline (1� PBS; Life Technologies) supplementedwith 2% FBS and 0.02% azide (Sigma-Aldrich, SaintLouis, MO, USA). Subsequently, cells were washedwith 1� PBS supplemented with 2% FBS, 0.02%azide and 0.05% saponin (Sigma-Aldrich). Cellswere then permeabilized with the use of FACSpermeabilizing solution 2 (1� PERM2; BD Bio-sciences) for 10 min at room temperature, washedwith 1� PBS supplemented with 2% FBS, 0.02%azide and 0.05% saponin and were then stained withanti-human perforin (clone dG9), anti-human gran-ulysin (clone DH2) and anti-human granzyme B(clone GB11) monoclonal antibodies for 40 min at4�C. Finally, cells were washed with 1� PBS sup-plemented with 2% FBS, 0.02% azide and 0.05%saponin, followed by 1� PBS supplemented with 2%FBS and 0.02% azide. Samples were analyzed on anFACS Fortessa flow cytometer. Anti-human perforinwas purchased from BD Biosciences; anti-humangranulysin and anti-human granzyme B were pur-chased from BioLegend.
Staining of human B-cell precursor leukemia cell lines
Human B-cell precursor leukemia cell lines werestained with anti-human MICA/B (clone 6D4), anti-CD95 (clone DX2), anti-human TRAIL-R1 (DR4,clone HS101) or anti-human TRAIL-R2 (DR5,clone HS201) monoclonal antibodies for 30 min at4�C. Cells were washed and were then stained withan anti-mouse immunoglobulin (Ig)G1 antibodycoupled to fluorescein isothiocyanate (Life Tech-nologies) for cells stained with anti-TRAIL-R1(DR4) or anti-TRAIL-R2 (DR5) antibodies. Sam-ples were analyzed on an FACS Fortessa flow cy-tometer. Anti-human TRAIL-R1 and TRAIL-R2antibodies were purchased from Enzo Life Sciences(Plymouth, PA, USA).
51Chromium-release cytotoxicity assay
CD56þ CB-CIK and CD56þ PB-CIK cells werepurified through positive selection with CD56 mag-netic microbeads (Miltenyi Biotec, Auburn, CA,USA) and used in all cytotoxicity assays. The cyto-toxicity assays were conducted as previously described(13); 104 target cells (K562, U266, NALM6 and
REH) were labeled with 51chromium (51Cr) (PerkinElmer, Waltham, MA, USA) and plated with effectorCD56þCB-CIK or CD56þPB-CIK cells at effector-to-target cell ratios of 1.25:1, 5:1, 20:1 and 40:1 andincubated for 4 h at 37�C in a 5% CO2 atmosphere.At the end of the assay, 100 mL of supernatant wasrecuperated and radioactivity was counted with theuse of a gamma counter (Packard Bioscience; modelE5002, Waltham, MA, USA).
The percentage of specific 51Cr release wascalculated by means of the following equation: %specific 51Cr release ¼ [(test release)e(spontaneousrelease)/(maximal release)e(spontaneous release)] �100%. Maximal release was obtained after theaddition of 1 mol/L hydrochloric acid.
Inhibition of the NKG2D pathway
To measure inhibition of the NKG2D pathway, acytotoxic assay was performed in the presence20 mg/mL of an anti-human NKG2D blocking anti-body (clone M585; Amgen, WA, USA). CD56þ
CB-CIK or CD56þ PB-CIK cells were incubatedwith the anti-human NKG2D blocking antibody oranti-human IgG1 antibody for 30 min at roomtemperature before the addition of target cells.Incubation then continued for an additional 4 h at37�C, in a 5% CO2 atmosphere. Effector-to-targetcell ratio used was 40:1.
Inhibition of the TRAIL pathway
To measure inhibition of the TRAIL pathway, thecytotoxic assay was performed in the presence of20 mg/mL of both the anti-human TRAIL-R1 (DR4)and TRAIL-R2 (DR5) blocking antibodies. Targetcells were incubated with these two blocking anti-bodies or an anti-human IgG1 antibody for 30 min atroom temperature before the addition of effectorcells. Incubation then continued for an additional 4 hat 37�C in a 5% CO2 atmosphere. Effector-to-targetcell ratio used was 40:1.
CD95-mediated apoptosis assays
B-ALL cell lines were plated in RPMI-1640 me-dium supplemented with 10% FBS and 2 mmol/LL-glutamine at a density of 2 � 105 cells/well in 96-well, flat-bottomed plates. Cells were then incubatedwith an anti-human CD95 antibody (clone APO-1e3,100 or 250 ng/mL; Enzo Life Sciences, Burlington,ON, Canada) for 1 h at 37�C in a 5% CO2 atmo-sphere. At the end of the incubation, an affiniPurerabbit anti-mouse IgG (10 mg/mL, Jackson Immu-noResearch, West Grove, PA, USA) was added for anadditional 23 h at 37�C in a 5% CO2 atmosphere.
B-ALL cell lines were plated in RPMI-1640 me-dium supplemented with 10% FBS and 2 mmol/LL-glutamine at a density of 2 � 105 cells/well in96-well, flat-bottomed plates. Cells were incubatedwith a TRAIL recombinant protein (SuperKillerTRAIL, 50 ng/mL, Enzo Life Sciences) for 24 h at37�C in a 5% CO2 atmosphere.
Assessment of apoptosis
To assess apoptosis, cells were harvested and stainedwith fluorescein isothiocyanateelabeled annexin (BDBioscience) and 7AAD (BD Bioscience) for 15 min atroom temperature. Cells were then resuspended in300 mL of annexin buffer (1�, BD Bioscience) andsubsequently analyzed by means of FACS within 1 h.
Calculation of percentage of specific inhibition of immunepathways
The percentage of specific inhibition was calculatedas follows: % specific inhibition ¼ [(% lysis of labeledcells without the addition of antibody or receptor)e(% lysis of labeled cells with the addition of antibodyor receptor)]/(% lysis of labeled cells without theaddition of antibody or receptor) � 100.
Calculation of percentage of specific apoptosis
The percentage of specific apoptosis was calculated asfollows: % specific apoptosis ¼ [(% of experimen-tal apoptosis)e(% spontaneous apoptosis)]/(100espontaneous apoptosis) � 100.
Statistics
Results were analyzed with the use of GraphPadPrism 5.0 (GraphPad Software Inc, La Jolla, CA,USA). For statistical analysis, the Mann-Whitneytest (one- or two-tailed) and the analysis of variancetest in combination with the Bonferroni post-testwere used according to the experiment. The level ofsignificance was set at P � 0.05.
Results
Differential expression of phenotypic markers on CD56þ
CB-CIK compared with CD56þ PB-CIK cells
In 2006, Introna et al. (3) described for the first timethat CIK cells could be amplified from CBMCs. Theyconfirmed that the increase in the CD3þCD56þ
subpopulation derived from CBMCs was consistentwith what had been reported for PBMCs. Similarly, in
our study, CIK cells were expanded >1000-fold fromboth CBMCs and PBMCs over a 21-day cultureperiod, and we observed a >30% increase in theCD3þCD56þ CB-CIK and PB-CIK cell fractions bythe end of the culture period (Supplementary Figure 1).Thus, CIK cells in general and the CD3þCD56þ CIKsubpopulations in particular could be amplified fromeither CBMCs or PBMCs with the use of the sameprotocol.
We then analyzed the expression of differentmarkers, including natural cytotoxicity receptors(NCRs), cytotoxic granule proteins (perforin, gran-ulysin and granzyme B), NKG2D and TRAIL,which can all contribute to the activation of cytotoxicactivity of effector against tumor cells on CD56þ
CB-CIK and CD56þ PB-CIK cells (Figure 1). Toanalyze the expression of different markers, CD56þ
cells were gated on total viable lymphocytes that wereselected by means of CD3, CD56 and 7AADmarkers (Supplementary Figure 2). Expression ofNKp30 was more pronounced on CD56þ CB-CIKcells than on CD56þ PB-CIK cells (Figure 1A).Expression of NKp44 appeared to be stronger onCD56þ CB-CIK cells than on CD56þ PB-CIK cells,but it was not significant, whereas NKp46 was veryweakly expressed on both CD56þ CB and PB-CIKcells.
With regard to cytotoxic granule proteins, weobserved that CD56þ CB-CIK subpopulationsexpressed granzyme B and granulysin, albeit to amuch lesser degree than in CD56þ PB-CIK cells(Figure 1B). Perforin expression appeared to bereduced in CD56þ CB-CIK compared with CD56þ
PB-CIK cells, but it was not statistically significant(Figure 1B).
TRAIL expression was consistently higher despitevariabilities between the donors on CD56þ CB-CIKsubpopulations compared with CD56þ PB-CIKcells (Figure 1C,E). Finally, NKG2D expression wassimilarly detected on the CD56þ CB and PB-CIKcells (Figure 1D).
Comparison of the cytotoxic activity of CD56þ CB-CIKand CD56þ PB-CIK against several B-ALL cell lines
We observed that CD56þ CB-CIK cells lysed K562and U266 control cell lines as effectively as didCD56þ PB-CIK cells (Figure 2). Whereas CD56þ
PB-CIK and CB-CIK cells showed similar cytotoxicactivity against the NALM6 and 697 cell lines(although with a weak efficacy against 697), weobserved that CD56þ CB-CIK cells were statisticallymore efficient than CD56þ PB-CIK cells at lysingREH cells; in contrast to this, CD56þ PB-CIK cellswere statistically more efficient than CD56þ CB-CIK cells at lysing KOPN-8 cells (Figure 2).
Figure 1. Phenotype of CD56þ CB-CIK compared with CD56þ PB-CIK cells. On day 21, CIK cells were stained with CD3 and CD56 andsubsequently analyzed for the expression of several markers: (A) NKp30, NKp44 and NKp46 (n ¼ 6 independent CB- and PB-CIK cellcultures); (B) perforin, granzyme B and granulysin (n ¼ 5 independent CB- and PB-CIK cell cultures), (C) TRAIL (n ¼ 5 independent CB-and PB-CIK cell cultures) and (D) NKG2D (n ¼ 5 independent CB- and PB-CIK cell cultures) on each subpopulation: CD56þ CB-CIKcells in white bars; CD56þ PB-CIK cells in black bars. Results are represented as the mean � standard error of the mean. *P � 0.05 and**P � 0.01, Mann-Whitney test (two-tailed). (E) Representative example of an overlay of TRAIL expression with the use of flow cytometry.Filled gray area: isotype control; filled line: CD56þ CB-CIK cells; dashed line: CD56þ PB-CIK cells.
CD56+ CB and PB-CIK differently kill B-ALL 5
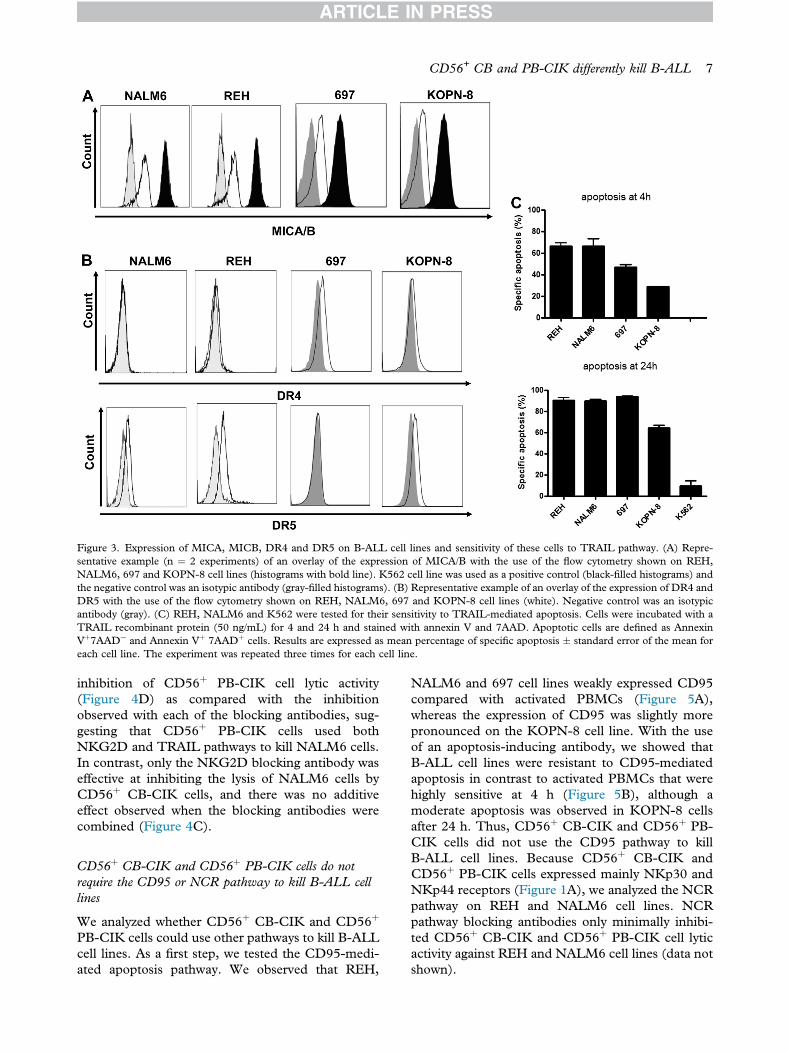
Expression of MICA, MICB, DR4 and DR5 onB-ALL cell lines and sensitivity of these cells to TRAILpathway
To determine if the CD56þ CB-CIK and PB-CIKcells could kill B-ALL cell lines through the NKG2Dand TRAIL pathways, we analyzed the expression ofNKG2D receptor ligands and the expression of DR4and DR5 receptors on these target cells. The fourB-ALL cell lines expressed MICA and/or MICB(Figure 3A) at an intermediate level compared withthe positive control K562 cells. In addition, NALM6and REH cell lines did not express DR4, whereasboth expressed low levels of DR5 (Figure 3B), withslightly higher levels detected on REH cells. TheKOPN-8 cell line weakly expressed both DR4 andDR5, with slightly higher levels for DR5, whereasthe 697 cell line did not express DR5 but expressedlow levels of DR4 (Figure 3B). We then tested if theexpression of DR4 and/or DR5 on B-ALL cell lineswas sufficient to sensitize them to TRAIL-mediatedapoptosis. We treated the four B-ALL cell lines aswell as the K562 cell line, which has been shown tobe resistant to TRAIL-mediated apoptosis (24,25),with a TRAIL recombinant protein for 24 h. Indeed,
the K562 cell line showed resistance to TRAIL-mediated apoptosis at 24 h, as expected. Becausethe cytotoxic activity of CD56þ CIK cells againstB-ALL cell lines was measured in a 4-h test, weassessed the sensitivity of B-ALL cell lines toTRAIL-mediated apoptosis at 4 h as well. Althoughthe apoptosis at 4 h was less pronounced than at24 h, all 4 B-ALL cell lines at this time point weresensitive to TRAIL-mediated apoptosis, with 697and KOPN-8 cell lines showing a partial sensitivity(Figure 3C).
Involvement of both NKG2D and TRAIL pathways byCD56þ CB-CIK and CD56þ PB-CIK cells is dependenton the B-ALL cell line
We studied in detail the mechanism of cytotoxic ac-tivity of CD56þ CIK cells against REH and NALM6cell lines. We tested whether an NKG2D blockingantibody could inhibit the CD56þ CB-CIK andPB-CIK cells from lysing B-ALL cell lines. As a firststep, we confirmed, as previously shown by Verneriset al. (15), that the NKG2D blocking antibody couldinhibit the lytic activity of CD56þ CB-CIK and
Figure 2. Cytotoxic activity of CD56þ CB-CIK compared with CD56þ PB-CIK cells. On day 21, CB- and PB-CIK cells were purified withthe use of CD56 magnetic microbeads to recuperate the CD56þ fraction. The cytotoxic activity of CD56þ CB-CIK (dotted line) and CD56þ
PB-CIK (filled line) were tested against K562 (n ¼ 6 independent CB- and PB-CIK cell cultures), U266 (n ¼ 4 independent CB- and PB-CIK cell cultures), REH (n ¼ 6 independent CB- and PB-CIK cell cultures), NALM6 (n ¼ 4 independent CB- and PB-CIK cell cultures),697 (n ¼ 2 independent CB- and PB-CIK cell cultures) and KOPN-8 (n ¼ 2 independent CB- and PB-CIK cell cultures) cell lines atdifferent effector/target ratios. Results are represented as the mean � standard error of the mean. *P � 0.05 and **P � 0.01, analysis ofvariance test with a Bonferroni post-test. Percent specific lysis was calculated as follows: % specific lysis ¼ [(test release)e(spontaneousrelease)]/[(maximal release)e(spontaneous release)] � 100%.
6 L. Durrieu et al.
CD56þ PB-CIK cells against the U266 cell line byan average of 60% (data not shown). The NKG2Dblocking antibody was not as effective at inhibitingthe lytic activity of both CD56þ CB-CIK andCD56þ PB-CIK cells against the REH cell line(mean of 32.6 � 8% and mean of 40 � 4%,respectively) (Figure 4A,B). This result might sug-gest that CD56þ CB- and PB-CIK cells could useadditional cytotoxic pathways to kill REH cells. Forthe NALM6 cell line, the inhibition by the NKG2Dblocking antibody was similar to that observedfor the U266 cell line (mean of 65 � 8% for CD56þ
CB-CIK cells and mean of 61 � 15% for CD56þ
PB-CIK cells) (Figure 4C,D), with no differenceobserved for either CIK cell source.
We next analyzed if we could inhibit the lyticactivity of CD56þ CB-CIK and CD56þ PB-CIKcells against REH and NALM6 cell lines by use ofblocking antibodies to both DR4 and DR5. First, weconfirmed that these blocking antibodies couldcompletely inhibit TRAIL-mediated apoptosis of theREH cell line (data not shown). We observed thatthese antibodies inhibited the lysis of REH cells byCD56þ CB-CIK cells by 57 � 8% (Figure 4A). Insharp contrast, CD56þ PB-CIK cell lytic activity on
REH cells was not significantly inhibited by the DR4and DR5 blocking antibodies compared with theisotype control (Figure 4B). Finally, DR4 and DR5blocking antibodies did not inhibit the lytic activity ofCD56þ CB-CIK cells against the NALM6 cell linecompared with the isotype control (Figure 4C) butinhibited the lytic activity of CD56þ PB-CIK cells by38.5 � 2% (Figure 4D).
In the next set of experiments, we compared theability of NKG2D, DR4 and DR5 blocking anti-bodies, tested in combination, to inhibit the cyto-toxic activity of CD56þ CIK cells against B-ALLcell lines by NKG2D, DR4 and DR5 blocking an-tibodies. As shown in Figure 4A, CD56þ CB-CIKcells used both NKG2D and TRAIL pathways tokill REH cells, and there was an additive inhibitoryeffect when the blocking antibodies were addedtogether. On the other hand, only the NKG2Dblocking antibody was effective at inhibiting CD56þ
PB-CIK cells from lysing REH cells, and the com-bination of blocking antibodies was not better thanthe NKG2D blocking antibody alone (Figure 4B).For the NALM6 cell line, the results mirrored thosefor the REH cell line. Indeed, the combined block-ing antibodies resulted in a significantly stronger
Figure 3. Expression of MICA, MICB, DR4 and DR5 on B-ALL cell lines and sensitivity of these cells to TRAIL pathway. (A) Repre-sentative example (n ¼ 2 experiments) of an overlay of the expression of MICA/B with the use of the flow cytometry shown on REH,NALM6, 697 and KOPN-8 cell lines (histograms with bold line). K562 cell line was used as a positive control (black-filled histograms) andthe negative control was an isotypic antibody (gray-filled histograms). (B) Representative example of an overlay of the expression of DR4 andDR5 with the use of the flow cytometry shown on REH, NALM6, 697 and KOPN-8 cell lines (white). Negative control was an isotypicantibody (gray). (C) REH, NALM6 and K562 were tested for their sensitivity to TRAIL-mediated apoptosis. Cells were incubated with aTRAIL recombinant protein (50 ng/mL) for 4 and 24 h and stained with annexin V and 7AAD. Apoptotic cells are defined as AnnexinVþ7AADe and Annexin Vþ 7AADþ cells. Results are expressed as mean percentage of specific apoptosis � standard error of the mean foreach cell line. The experiment was repeated three times for each cell line.
CD56+ CB and PB-CIK differently kill B-ALL 7
inhibition of CD56þ PB-CIK cell lytic activity(Figure 4D) as compared with the inhibitionobserved with each of the blocking antibodies, sug-gesting that CD56þ PB-CIK cells used bothNKG2D and TRAIL pathways to kill NALM6 cells.In contrast, only the NKG2D blocking antibody waseffective at inhibiting the lysis of NALM6 cells byCD56þ CB-CIK cells, and there was no additiveeffect observed when the blocking antibodies werecombined (Figure 4C).
CD56þ CB-CIK and CD56þ PB-CIK cells do notrequire the CD95 or NCR pathway to kill B-ALL celllines
We analyzed whether CD56þ CB-CIK and CD56þ
PB-CIK cells could use other pathways to kill B-ALLcell lines. As a first step, we tested the CD95-medi-ated apoptosis pathway. We observed that REH,
NALM6 and 697 cell lines weakly expressed CD95compared with activated PBMCs (Figure 5A),whereas the expression of CD95 was slightly morepronounced on the KOPN-8 cell line. With the useof an apoptosis-inducing antibody, we showed thatB-ALL cell lines were resistant to CD95-mediatedapoptosis in contrast to activated PBMCs that werehighly sensitive at 4 h (Figure 5B), although amoderate apoptosis was observed in KOPN-8 cellsafter 24 h. Thus, CD56þ CB-CIK and CD56þ PB-CIK cells did not use the CD95 pathway to killB-ALL cell lines. Because CD56þ CB-CIK andCD56þ PB-CIK cells expressed mainly NKp30 andNKp44 receptors (Figure 1A), we analyzed the NCRpathway on REH and NALM6 cell lines. NCRpathway blocking antibodies only minimally inhibi-ted CD56þ CB-CIK and CD56þ PB-CIK cell lyticactivity against REH and NALM6 cell lines (data notshown).
Figure 4. Inhibition of cytotoxic activity of CD56þ CB- and PB-CIK cells by both the NKG2D and TRAIL pathways. Cytotoxic activity ofCD56þ CB- and PB-CIK was tested in the presence of anti-human IgG1 antibody (20 mg/mL), anti-human NKG2D blocking antibody (20mg/mL), anti-human DR4 and DR5 blocking antibodies (20 mg/mL each) or both NKG2D and DR4/DR5 blocking antibodies against (Aand B) REH and (C and D) NALM6 cell lines. Effector-to-target cell ratio was 40:1, and the specific lysis observed without blockingantibody was between 25e30%. Results are expressed as mean percent of inhibition � standard error of the mean. The experiment wasrepeated three times for each cell line with the use of independent CD56þ CB- and PB-CIK cells. *P � 0.05, **P � 0.01; Mann-Whitneytest (one- or two-tailed) was used. Percentage of specific inhibition was calculated as follows: % specific inhibition ¼ [(% lysis of labeled cellswithout the addition of antibody)e(% lysis of labeled cells with the addition of antibody)]/(% lysis of labeled cells without the addition ofantibody) � 100.
8 L. Durrieu et al.
Discussion
Several research groups have studied CIK cells iso-lated from either PBMCs (1,9,15) or CBMCs (3,26)and have demonstrated their cytotoxic activityagainst several tumors. The pathways involved in thecytotoxic activity of CIK cells against tumor cellshave been well studied (15,19,20). In contrast, thelytic mechanisms used by CB and PB CIK cells tokill B-ALL cell lines are still unknown. Furthermore,until now, a comparison of the expression of differentreceptors on PBMC- and CBMC-derived CIK cellstogether with an analysis of the cytotoxic activity ofPB-CIK versus CB-CIK cells in the same study hasnot been done. By performing this study, we have
shown that the expression of some receptors wasdifferent on CD56þ CB-CIK compared with CD56þ
PB-CIK cells. In addition, we have observed for thefirst time that not only do these different CIK cellpreparations kill B-ALL cell lines through activationof different cytotoxic pathways but that the pathwaysinvolved also differ according to the B-ALL cell line.
CIK cells express various markers, with the twomajor ones being CD3 and CD56. Additionalmarkers expressed by CIK cells include NKG2D,CD8, CD4 and NCR, as well as other receptors(2,18). Several of these have been described in theliterature as playing a role in immune cellemediatedcytotoxic activity, including the NCR receptors,
Figure 5. Sensitivity of B-ALL cell lines to CD95-mediated apoptosis. (A) REH, NALM6 and PBMC activated for 8 days were tested forthe expression of CD95 with the use of flow cytometry. The expression of CD95 is represented in white and the isotypic antibody in grayhistograms; representative of three experiments. (B) REH, NALM6, 697, KOPN-8 and PBMCs were tested for sensitivity to CD95-mediated apoptosis. Cells were incubated with an anti-CD95 antibody (Apo1-3, 250 ng/mL) cross-linked with anti-mouse antibody (10 mg/mL) for 4 and 24 h and stained with annexin V and 7AAD. Apoptotic cells are defined as Annexin Vþ 7AADe and Annexin Vþ 7AADþ cells.Results are expressed as mean percentage of specific apoptosis � standard error of the mean for each cell line. The experiment was repeatedthree times for each cell line.
CD56+ CB and PB-CIK differently kill B-ALL 9
recently shown to be involved in NK cellemediatedkilling of the REH B-ALL cell line (27). TheNKG2D receptor has been described as being themain pathway through which CIK cells mediatetumor cell killing (15), whereas activation of theTRAIL signaling pathway has been implicatedin CIK cellemediated cytotoxic activity againstrhabdomyosarcoma cells (20). It has also beenshown that cytotoxic granules were involved in cellkilling when NKG2D and NCR pathways wereactivated (16).
By analyzing NKp30, NKp44, NKp46, TRAILand NKG2D and proteins expressed in cytotoxicgranules, we observed that CD56þ CB-CIK cellsdifferentially expressed several of the receptorscompared with CD56þ PB-CIK cells. For some re-ceptors (NKp30 and TRAIL), the expression wasmore pronounced on CD56þ CB-CIK than onCD56þ PB-CIK cells, whereas for others either theopposite was found (granzyme B and granulysine) orthere was no difference (NKp44, perforin andNKG2D). These differences in receptor expression
could be due to the intrinsic differences observedbetween CB- and PB-derived mononuclear cells andparticularly T cells, which have been shown to havedifferences in their phenotype, such as the relativefrequency of naive and memory T cells, and in theirfunction, such as their capacity to produce cytokinesunder stimulation (28).
Despite these differences in receptor expressionbetween CD56þ CB- and CD56þ PB-CIK cells, weobserved that both could kill three of the six tumorcell lines (K562, U266 and NALM6) with similarefficacy and that both had a weak cytotoxic activityagainst the 697 cell line. Although the cytotoxicactivity of CD56þ CB- and PB-CIK cells againstB-ALL cell lines, particularly NALM6 and 697 celllines, was lower than reported for NK cell andcytotoxic T-lymphocyte lysis (29,30), it was suffi-cient to lyse these B-ALL cells. Importantly, ourresults demonstrated that CD56þ CB-CIK cellswere significantly more effective than were CD56þ
PB-CIK cells to kill REH cell line, and in contrast,less effective to kill the KOPN-8 cell line. This
difference probably is not caused by the fractionationof CIK cells, because (i) in our hands, the cytotoxicactivity of CIK cells was predominantly related to theCD3þCD56þ subset, with a little participation ofthe CD3eCD56þ subset (13); (ii) the proportion ofCD3þCD56þ and CD3-CD56þ cells in the CD56þ
CB- and PB-CIK fractions was similar in both(97�1% of CD3þCD56þ cells and 2�1% ofCD3-CD56þ cells; data not shown). In addition, it isunlikely that any difference in NKp30 expression onCD56þ CB- or CD56þ PB-CIK cells could explainthis result because we did not observe any inhibitionof REH cell lysis with soluble NCR receptors.However, the difference in TRAIL expression mayexplain what we found because CD56þ CB-CIKcells expressed more TRAIL than did CD56þ PB-CIK cells. Also, CD56þ CB-CIK cells were effec-tively inhibited, albeit partially, from lysing REHcells by DR4/DR5 blocking antibodies, whereasCD56þ PB-CIK cells were not inhibited.
Our analysis of NKG2D, TRAIL, CD95 andNCR pathways suggest that CB- and PB-CIK cellsuse both the NKG2D and TRAIL pathways to kill B-ALL cells. In the present study, we show for the firsttime that the involvement of these pathways differs,depending on both the source of the immune cellpopulation (ie, CIK cells) and the target cell popu-lation (ie, B-ALL cell lines). In agreement with ourdata, Kuçi et al. (20) showed that CIK cells killedrhabdomyosarcoma cell lines in part through activa-tion of the NKG2D pathway but could also use theTRAIL pathway. However, in our experiments, theinhibition of NKG2D and TRAIL pathways did notcompletly abolish the lytic activity of CD56þ CB-CIK cells against the NALM6 cell line or CD56þ PB-CIK cells against the REH cell line, indicating that atleast another pathway could be involved, althoughother blocking experiments did not permit us to findwhich additional pathway(s) was (were) involved.
With respect to the NKG2D pathway, we cannotascribe any differences in cytotoxic activity to theexpression of MICA/MICB on B-ALL cell lines,which was similar for these two ligands on REH andNALM6 cells and slightly less on 697 and KOPN-8cells. It is possible that other NKG2D ligands, suchas ULBP1, ULBP2 and ULBP3 (31), are differen-tially expressed, which may explain why inhibition ofNKG2D-mediated lysis is more pronounced onsome cells compared with others. In addition, sometumor cells can secrete soluble NKG2D ligandsinto the microenvironment (32), which in essencebinds to NKG2D on effector cells, preventing theirengagement with ligands on tumor cells (33). Thus,REH cells may secrete more soluble NKG2D ligandsthan do NALM6 cells and thereby partially blockCIK cell cytotoxic activity.
The differential expression of TRAIL and itsreceptors (DR4 and DR5) on CD56þ CIK cells andB-ALL cells, respectively, could also play a role inthe observed differences between cell lines with re-gard to the TRAIL pathway. Furthermore, it is likelythat the differential receptor usage/activation by sol-uble and/or membrane-bound TRAIL (34) plays rolein these differences as well. Indeed, a recent studydemonstrated that neuroblastoma cells were resis-tant to soluble TRAIL-mediated apoptosis, whereasthey were sensitive to membrane-bound TRAIL-mediated apoptosis (35). Notably, although REH,NALM6 and KOPN-8 cell lines expressed DR5 tothe same extent, only REH and NALM6 were sen-sitive to soluble TRAIL-mediated apoptosis, whereasKOPN-8 showed only partial sensitivity at 4 and24 h, thus indicating that this mechanism of differ-ential receptor usage/activation can be operational inthese different cell lines. Taken together, our datashow that CD56þ CB-CIK and CD56þ PB-CIKcells use two main pathways to kill B-ALL cell lines,but as to which pathway(s) predominates is depen-dent on both the effector and target cells involved.Several clinical studies have shown that the infusionof CB- or PB-CIK cells was well tolerated and didnot cause severe adverse events (36e39). However,no comparison of the efficacy of CB-CIK versus PB-CIK cells has been addressed. Our data could alsohave clinical consequences because they suggest thatit could be more advantageous to use either CD56þ
CB-CIK cells or CD56þ PB-CIK cells, dependingon the target tumor cells. More generally, our datacould have relevance with regard to the choice ofwhich CIK cells to use in the clinic because it sug-gests that depending on the tumor, CD56þ CB-CIKcells or CD56þ PB-CIK cells may have more thera-peutic efficacy.
Acknowledgments
This project was funded by grants from Fonds derecherche du Qu�ebec-Sant�e (FRQS), the CanadianInstitutes of Health Research MOP-130246 (MDand EH), the Fondation du CHU Sainte-Justine andthe Fondation Charles Bruneau (EH). LD received ascholarship from the Fondation de l’Hôpital Sainte-Justine/Fondation des Étoiles and a doctoral stu-dentship from the Department of Microbiologyand Immunology, University of Montreal. EH is ascholar of the FRQS.
The authors would like to thank the AmgenCompany for the NKG2D blocking antibody.
Disclosure of interests: The authors have nocommercial, proprietary, or financial interest in theproducts or companies described in this article.
1. Schmidt-Wolf IG, Negrin RS, Kiem HP, Blume KG,Weissman IL. Use of a SCID mouse/human lymphoma modelto evaluate cytokine-induced killer cells with potent antitumorcell activity. J Exp Med. 1991;174:139e49.
2. Schmidt-Wolf IG, Lefterova P, Mehta BA, Fernandez LP,Huhn D, Blume KG, et al. Phenotypic characterization andidentification of effector cells involved in tumor cell recogni-tion of cytokine-induced killer cells. Exp Hematol. 1993;21:1673e9.
3. Introna M, Franceschetti M, Ciocca A, Borleri G, Conti E,Golay J, et al. Rapid and massive expansion of cord blood-derived cytokine-induced killer cells: an innovative proposalfor the treatment of leukemia relapse after cord blood trans-plantation. Bone Marrow Transplant. 2006;38:621e7.
4. Lu PH, Negrin RS. A novel population of expanded humanCD3þCD56þ cells derived from T cells with potent in vivoantitumor activity in mice with severe combined immunode-ficiency. J Immunol. 1994;153:1687e96.
5. Verneris MR, Ito M, Baker J, Arshi A, Negrin RS, Shizuru JA.Engineering hematopoietic grafts: purified allogeneic he-matopoietic stem cells plus expanded CD8þ NK-T cells inthe treatment of lymphoma. Biol Blood Marrow Transplant.2001;7:532e42.
6. Edinger M, Cao YA, Verneris MR, Bachmann MH,Contag CH, Negrin RS. Revealing lymphoma growth and theefficacy of immune cell therapies using in vivo biolumines-cence imaging. Blood. 2003;101:640e8.
7. Sweeney TJ, Mailander V, Tucker AA, Olomu AB, Zhang W,Cao Y, et al. Visualizing the kinetics of tumor-cell clearance inliving animals. Proc Natl Acad Sci U S A. 1999;96:12044e9.
8. Wongkajornsilp A, Somchitprasert T, Butraporn R,Wamanuttajinda V, Kasetsinsombat K, Huabprasert S, et al.Human cytokine-induced killer cells specifically infiltratedand retarded the growth of the inoculated human chol-angiocarcinoma cells in SCID mice. Cancer Invest. 2009;27:140e8.
9. Kim HM, Lim J, Park SK, Kang JS, Lee K, Lee CW, et al.Antitumor activity of cytokine-induced killer cells againsthuman lung cancer. Int Immunopharmacol. 2007;7:1802e7.
10. Kim HM, Lim J, Yoon YD, Ahn JM, Kang JS, Lee K, et al.Anti-tumor activity of ex vivo expanded cytokine-inducedkiller cells against human hepatocellular carcinoma. IntImmunopharmacol. 2007;7:1793e801.
11. Kim YJ, Lim J, Kang JS, Kim HM, Lee HK, Ryu HS, et al.Adoptive immunotherapy of human gastric cancer with exvivo expanded T cells. Arch Pharm Res. 2010;33:1789e95.
12. Petvises S, Pakakasama S, Wongkajornsilp A, Sirireung S,Panthangkool W, Hongeng S. Ex vivo generation of cytokine-induced killer cells (CD3þ CD56þ) from post-stem celltransplant pediatric patients against autologous-Epstein-Barrvirus-transformed lymphoblastoid cell lines. Pediatr Trans-plant. 2007;11:511e7.
13. Durrieu L, Gregoire-Gauthier J, Dieng MM, Fontaine F, leDeist F, Haddad E. Human interferon-alpha increases thecytotoxic effect of CD56(þ)cord blood-derived cytokine-induced killer cells on human B-acute lymphoblastic leukemiacell lines. Cytotherapy. 2012;14:1245e57.
14. Sangiolo D, Martinuzzi E, Todorovic M, Vitaggio K,Vallario A, Jordaney N, et al. Alloreactivity and anti-tumoractivity segregate within two distinct subsets of cytokine-induced killer (CIK) cells: implications for their infusionacross major HLA barriers. Int Immunol. 2008;20:841e8.
15. Verneris MR, Karami M, Baker J, Jayaswal A, Negrin RS.Role of NKG2D signaling in the cytotoxicity of activated andexpanded CD8þ T cells. Blood. 2004;103:3065e72.
16. Hayakawa Y, Smyth MJ. NKG2D and cytotoxic effectorfunction in tumor immune surveillance. Semin Immunol.2006;18:176e85.
17. Bauer S, Groh V, Wu J, Steinle A, Phillips JH, Lanier LL,et al. Activation of NK cells and T cells by NKG2D, a re-ceptor for stress-inducible MICA. Science. 1999;285:727e9.
18. Franceschetti M, Pievani A, Borleri G, Vago L, Fleischhauer K,Golay J, et al. Cytokine-induced killer cells are terminallydifferentiated activated CD8 cytotoxic T-EMRA lymphocytes.Exp Hematol. 2009;37:616e628. e2.
19. Pievani A, Borleri G, PendeD,Moretta L,Rambaldi A,Golay J,et al. Dual-functional capability of CD3þCD56þ CIK cells, aT-cell subset that acquires NK function and retains TCR-mediated specific cytotoxicity. Blood. 2011;118:3301e10.
20. Kuci S, Rettinger E, Voss B,Weber G, Stais M, Kreyenberg H,et al. Efficient lysis of rhabdomyosarcoma cells by cytokine-induced killer cells: implications for adoptive immunotherapyafter allogeneic stem cell transplantation.Haematologica. 2010;95:1579e86.
21. Falschlehner C, Schaefer U, Walczak H. Following TRAIL’spath in the immune system. Immunology. 2009;127:145e54.
22. Linn YC, Lau LC, Hui KM. Generation of cytokine-inducedkiller cells from leukaemic samples with in vitro cytotoxicityagainst autologous and allogeneic leukaemic blasts. Br JHaematol. 2002;116:78e86.
23. Marin V, Dander E, Biagi E, Introna M, Fazio G, Biondi A,et al. Characterization of in vitro migratory properties of anti-CD19 chimeric receptor-redirected CIK cells for their po-tential use in B-ALL immunotherapy. Exp Hematol. 2006;34:1219e29.
24. Di Pietro R, Secchiero P, Rana R, Gibellini D, Visani G,Bemis K, et al. Ionizing radiation sensitizes erythroleukemiccells but not normal erythroblasts to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)emediated cyto-toxicity by selective up-regulation of TRAIL-R1. Blood. 2001;97:2596e603.
25. Qiu G, Jiang J, Liu XS. Pentamidine sensitizes chronic mye-logenous leukemia K562 cells to TRAIL-induced apoptosis.Leuk Res. 2012;36:1417e21.
26. Li Y, Schmidt-Wolf IG, Wu YF, Huang SL, Wei J, Fang J,et al. Optimized protocols for generation of cord blood-derived cytokine-induced killer/natural killer cells. AnticancerRes. 2010;30:3493e9.
27. Zhang H, Cui Y, Voong N, Sabatino M, Stroncek DF,Morisot S, et al. Activating signals dominate inhibitory signalsin CD137L/IL-15 activated natural killer cells. J Immunother.2011;34:187e95.
28. Lin SJ, Yan DC, Lee YC, Hsiao HS, Lee PT, Liang YW, et al.Umbilical cord blood immunology: relevance to stem celltransplantation. Clin Rev Allergy Immunol. 2012;42:45e57.
29. Dolstra H, Fredrix H, Maas F, Coulie PG, Brasseur F,Mensink E, et al. A human minor histocompatibility antigenspecific for B cell acute lymphoblastic leukemia. J Exp Med.1999;189:301e8.
30. Reid GS, Bharya S, Klingemann HG, Schultz KR. Differen-tial killing of pre-B acute lymphoblastic leukaemia cells byactivated NK cells and the NK-92 ci cell line. Clin ExpImmunol. 2002;129:265e71.
31. Cosman D, Mullberg J, Sutherland CL, Chin W, Armitage R,Fanslow W, et al. ULBPs, novel MHC class I-related mole-cules, bind to CMV glycoprotein UL16 and stimulate NKcytotoxicity through the NKG2D receptor. Immunity. 2001;14:123e33.
32. Salih HR, Antropius H, Gieseke F, Lutz SZ, Kanz L,Rammensee HG, et al. Functional expression and release ofligands for the activating immunoreceptor NKG2D in leuke-mia. Blood. 2003;102:1389e96.
33. Groh V, Wu J, Yee C, Spies T. Tumour-derived soluble MICligands impair expression of NKG2D and T-cell activation.Nature. 2002;419:734e8.
34. Wajant H, Moosmayer D, Wuest T, Bartke T, Gerlach E,Schonherr U, et al. Differential activation of TRAIL-R1 and-2 by soluble and membrane TRAIL allows selective surfaceantigen-directed activation of TRAIL-R2 by a soluble TRAILderivative. Oncogene. 2001;20:4101e6.
35. Sheard MA, Asgharzadeh S, Liu Y, Lin TY, Wu HW, Ji L,et al. Membrane-bound TRAIL supplements natural killercell cytotoxicity against neuroblastoma cells. J Immunother.2013;36:319e29.
36. Niu Q, Wang W, Li Y, Qin S, Wang Y, Wan G, et al. Cordblood-derived cytokine-induced killer cells biotherapy combinedwith second-line chemotherapy in the treatment of advancedsolid malignancies. Int Immunopharmacol. 2011;11:449e56.
37. Introna M, Pievani A, Borleri G, Capelli C, Algarotti A,Mico C, et al. Feasibility and safety of adoptive
immunotherapy with CIK cells after cord blood trans-plantation. Biol Blood Marrow Transplant. 2010;16:1603e7.
38. Laport GG, Sheehan K, Baker J, Armstrong R, Wong RM,Lowsky R, et al. Adoptive immunotherapy with cytokine-induced killer cells for patients with relapsed hematologicmalignancies after allogeneic hematopoietic cell trans-plantation. Biol BloodMarrowTransplant. 2011;17:1679e87.
39. Linn YC, Niam M, Chu S, Choong A, Yong HX, Heng KK,et al. The anti-tumour activity of allogeneic cytokine-inducedkiller cells in patients who relapse after allogeneic transplantfor haematological malignancies. Bone Marrow Transplant.2012;47:957e66.
Supplementary data
Supplementary data related to this article can befound at http://dx.doi.org/10.1016/j.jcyt.2013.12.010