Received for publication November 26, 1990Accepted February 18, 1991
Implication of Gibberellins in Head Smut(Sporisorium reilianum) of Sorghum bicolor'
Anne-Marie Matheussen2, Page W. Morgan*, and Richard A. FrederiksenDepartment of Soil and Crop Sciences (A.M.M., P.W.M.) and Department of Plant Pathology and
Microbiology (R.A.F.), Texas A&M University, College Station, Texas 77843-2474
ABSTRACT
The head smut fungus, Sporisorium reilianum ([Kuhn] Landonand Fullerton), was shown to reduce plant height in infectedSorghum bicolor ([L.] Moench) plants. The major reductions oc-curred in the internodes nearest the panicle and were moresevere in naturally infected than in inoculated plants. Less af-fected plants developed reproductively sterile panicles, andeventually smutted panicles developed phyllodied growths whichprogressed into leafy shoots. Extracts of smutted, sterile, andhealthy (control) panicles of field-grown plants exhibited gibber-ellin (GA)-like activity in the dwarf rice bioassay. When extractswere purified and assayed with deuterium-labeled GA standardsby gas chromatography-mass spectrometry-selected ion moni-toring (GC-MS-SIM), GA1, GA3, GA19, GA20, and GA53 were de-tected based on coelution with the standards, identical Kovatsretention index values, and matching ion masses and relativeabundances for three major ions. In addition, based on publishedKovats retention index values, ion masses, and relative abun-dance values, GA4, GA7, GA8, GA14, GA29, and GA44 were tenta-tively identified. Quantitative analysis revealed that panicles ofhealthy control plants contained from 60 to 100% higher totalconcentrations of GAs than panicles of smutted plants. Thesecomparisons were most striking for the early 13-hydroxylationpathway precursors GA53, GA44, and GA19 but not for GA20. Ex-tracts of S. reilianum sporidia and culture medium exhibited GA-like bioactivity, and GA1 and GA3 were detected based on GC-MS-SIM assay with 2H-labeled internal standards. Quantitativeanalysis of these GAs showed increasing concentrations from 4to 7 to 10 days of culture and a decline at 20 days. This is thefirst GC-MS-SIM detection of GAs in a non-Ascomycete fungus,and the disease symptoms and quantitative data suggested thatfungal infection may interfere with biosynthesis of GAs by thehost plant.
Throughout the world, sorghum (Sorghuim bicolor [L.]Moench) is attacked by the head smut disease organism,Sporisoriium reilianuim (Kuhn [Langdon and Fullerton]) (10,29). The pathogen primarily colonizes the plant's reproductivestructures and there develops its own sorus filled withteliospores.
' Contribution of the Texas Agricultural Experiment Station, paperNo. 25963 in the Technical Article Series. Supported by TexasAdvanced Research Program grant 2878 and the Texas AgriculturalExperiment Station.
Present address: Provinciaal Instituut voor Hygiene. Kronen-burgstraat 45. 2000 Antwerpen. Belgium.
Some aspects of head smut in sorghum suggest that planthormones are involved in the disease. Inflorescences of in-fected plants that do not develop sori are sterile reproductively(8, 9, 29). Inflorescences with sori only on the lower paniclebranches are always sterile in the upper branches. Less com-monly, panicles exhibit leafy or phyllodied development, andheavily infected panicles will eventually differentiate leafyshoots (5, 8, 9, 22, 29). These leafy shoots can be removedand cultured as separate, although infected, plants (5). It haslong been noted that head smut causes a substantial reductionin plant height (8) and that effect was documented recently(18). We have also observed that head smut alters plant colorto a more bluish green.
Because plant height and sexual development both arealtered in maize mutants with blocks in their GA' biosyntheticpathway (13, 21), the symptoms of sorghum head smut sug-gested to us that GAs may be involved in the expression ofdisease symptoms. Based on the GAs detected in vegetativeplants (2, 25), sorghum apparently contains the same biosyn-thetic pathway as maize (21).We have examined the effect of head smut on the height of
both naturally infected and inoculated sorghum plants andon the GA content of panicles. In addition, we have examinedthe possibility that S. reilianuim produces GAs in culture.
MATERIALS AND METHODS
Plant Studies
Sorghum (Sorghum bicolor [L.] Moench) inbred and hybridlines and varieties were grown on the Texas A&M UniversityFarm in Burleson County. Planting was on April 3, 1989, ata density to produce 65,000 plants/ha. Normal agronomicpractices were followed, including the application of 60 kg/ha of N, cultivation, and treatment with herbicides.
Plants used in an evaluation of resistance to head smut,both F, hybrids and selected inbred lines, were inoculated onMay 1, 1989, and individual plants were marked with smallstakes for identification. Teliospores ofSporisorilum reilianuim([Kuhn] Langdon and Fullerton) obtained from BurlesonCounty, TX, were surface sterilized in 95% ethanol, air dried,and streaked onto acidified potato dextrose agar for germi-nation (27). Colonies of sporidia from individually germi-nated teliospores were maintained separately. These cultureswere incubated at 25°C for 4 d. The products of a single
3Abbreviations: SIM, selected ion monitoring: GA. gibberellin;KRI. Kovats retention index.
teliospore colony, sporidia, were transferred to potato dextrosebroth and increased on a rotary shaker at room temperatureand 100 rpm for 4 d. This was then filtered through fourlayers of cheesecloth. The sporidial products of three individ-ual teliospores were mixed before inoculation. A hypodermicsyringe was used to introduce the mixed sporidia into 4-week-old sorghum seedlings immediately below the apical meristem(7). Ten plants were inoculated in each plot. A natural infec-tion of S. reilianum occurred in the nursery.
Plots of 44 different hybrids, all with ATx 631, a susceptibleline, as the female parent, were sampled to evaluate the effectsof head smut on growth. Control, naturally infected, andinoculated plants, either three or five each per plot, wereharvested between July 11 and July 21 and allowed to air dryat ambient temperatures. Measurements of plant height, in-ternode lengths of each of the upper three internodes, nodenumber, tiller number, and dry weight were made during thefirst 2 weeks in August.A second experiment was conducted in the same sorghum
disease nursery in 1990: planting date March 24, inoculationdate May 5. Typical diseased and healthy control plants wereharvested August 1 and 2 for cv Tx7078 from 14 naturallyinfected and inoculated plots. Data were obtained as for theprevious experiment.
Inflorescences were harvested from inoculated and healthycontrol plants of Tx7078, a smut-susceptible test variety, onJune 16, 1989. Plants exhibiting advanced sori developmenton diseased panicles, typified by appearance of mature (black)teliospores, were harvested from plot No. 1095 yielding threesmutted panicles, three sterile (floral development incomplete,neither pollen nor seeds produced) panicles, and three healthycontrol panicles of uniform size and stage of plant develop-ment. A second plot (No. 1132) with plants exhibiting anearlier stage of smut development (sori not yet opened) washarvested to yield two smutted and two control panicles. Athird plot (No. 1580), in which some smutted plants exhibitedextensive tissue proliferation and leafy growths on panicles,was harvested yielding three smutted, three sterile, and threecontrol panicles (50% anthesis). These panicles were trans-ported to the laboratory on dry ice, frozen (-80°C), lyophi-lized, ground to a fine powder in a blender, and then storedin sealed containers in a -80°C freezer until samples wereextracted for GA determinations. Thus, each plant sampleanalyzed represented a composite of tissue from two or threepanicles. The plot numbers are used to identify the paniclesamples in the tables and figures.
Fungus Studies
To study the possible presence of GAs in S. reilianum, weused teliospores from a field collection which had been char-acterized and routinely used in our laboratory (No. 10-Fh-GS, collection date 6/23/87; OD-SC, 170-6-17, collectionsite Hondo, TX). Sterile techniques were used in all stages ofthis work. The fungus was inoculated into 1-L flasks contain-ing 500 mL of Richard's medium (27) and then put on arotary shaker (100 rpm) at 25 ± 2°C for 4, 7, 10, and 20 d(pH 3.5-4.0). Because the fungus grows in this medium bybudding, purity of the cultures was verified by plating smallamounts of culture liquid on Petri plates containing Nash and
Synder agar (19) for detection of Fusarium species or onnutrient agar (27) for bacteria. After 24 h, the test plates wereobserved for purity of S. reilianum. This procedure wasrepeated a second time before cultures were harvested andprocessed for isolation ofGAs as an assurance that they werepure. To prepare these fungal cultures for extraction of GAs,fungus and culture medium were separated by centrifugationfor 20 min at 10,000g (4 L each for cultures 4 or 7 d old and2 L each for those 10 or 20 d old). The pellet was frozen at-80°C and lyophilized. Because the high sugar content of theculture medium made column chromatography difficult, thesupernatants of each sample were pooled, acidified to pH 2.9with 2 N HCI, and partitioned three times with water-saturatedethylacetate. The acidic ethylacetate phase was dried in vacuoat 35°C and stored in a -80°C freezer. The remaining aqueousphase was discarded.As an additional control, 500 mL of 4-d-old Richard's
medium (27), which contained no fungus, was extracted todetermine whether components of this medium containedGAs.
Extraction and Detection of Endogenous GAs
Extraction, purification, and detection of GAs were doneby methods described by Beall et al. (2). Briefly, approxi-mately 2 to 3 g of lyophilized, finely ground panicle tissuewere used for each extraction. The lyophilized fungal sampleswere weighed, the residue from the culture medium wassolubilized in approximately 10 mL 80% aqueous methanol,and both the fungal and the culture medium residue sampleswere combined. Further extraction and purification were thesame for the plant material and the combined fungal andculture medium samples. Samples were extracted in 80%aqueous methanol to which approximately 50,000 dpm each[l,2,3H]GA1 and [ 1,2-3H]GA4 (respectively, 37.7 and 32.2 Ci/mmol; Amersham) were added. This extract was chromato-graphed on a Davisil C,8 (Alltech) column and then on a SiO2column. A precipitation step previously used at this point (2)was not used in the present work. The free GA fraction wasthen separated by C18 HPLC and the subsequent fractionswere tested with the dwarf rice bioassay ( 17). Active fractionswere grouped, chromatographed on a 10-tim NucleosilN(CH3)2 HPLC column (Alltech), and again bioassayed (17).Only the free GA fraction was analyzed. Previously, theprecursor and conjugated GA fractions were found to containvery small or nondetectable amounts ofGA activity in vege-tative sorghum tissues (2), and they were not analyzed in thisstudy. Addition of deuterated GA internal standards wasdelayed until after this point to ensure that they did notinterfere with the bioassays.
Fractions exhibiting significant bioactivity were preparedfor mass spectrometry by addition of 50 ng of an appropriatedeuterated GA standard and subsequent derivatization byethereal diazomethane and silylation, with bis-(trimethyl-silyl)trifluoro-acetamide in 1% trimethylchlorosilane. Thesamples prepared this way were then analyzed by GC-MS-SIM operated at the conditions described previously (2).
Quantitation was based on the peak area ratios of the mostprominent ion (usually M+) of deuterated to nondeuteratedcompounds after correcting the nondeuterated peak area for
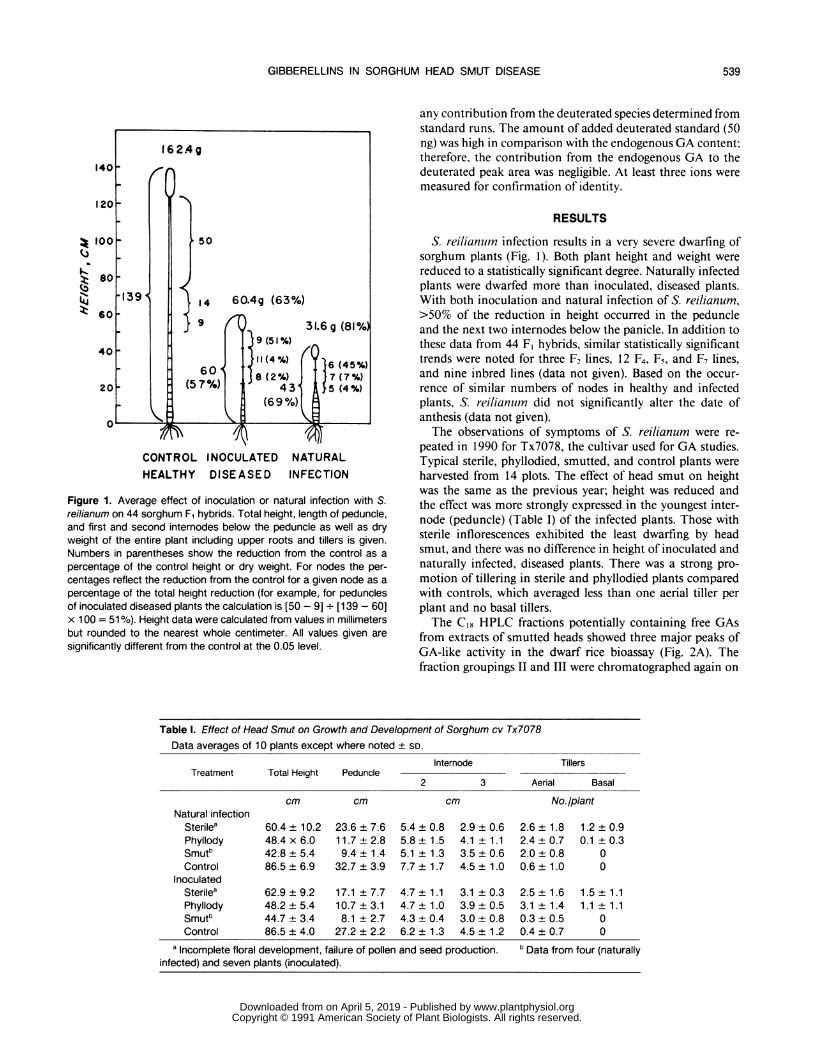
CONTROL INOCULATED NATURALHEALTHY DISEASED INFECTION
Figure 1. Average effect of inoculation or natural infection with S.reilianum on 44 sorghum F1 hybrids. Total height, length of peduncle,and first and second internodes below the peduncle as well as dryweight of the entire plant including upper roots and tillers is given.Numbers in parentheses show the reduction from the control as a
percentage of the control height or dry weight. For nodes the per-centages reflect the reduction from the control for a given node as a
percentage of the total height reduction (for example, for pedunclesof inoculated diseased plants the calculation is [50 - 9] - [139 - 60]x 100 = 51%). Height data were calculated from values in millimetersbut rounded to the nearest whole centimeter. All values given aresignificantly different from the control at the 0.05 level.
any contribution from the deuterated species determined fromstandard runs. The amount of added deuterated standard (50ng) was high in comparison with the endogenous GA content;therefore, the contribution from the endogenous GA to thedeuterated peak area was negligible. At least three ions were
measured for confirmation of identity.
RESULTS
S. reilianlum infection results in a very severe dwarfing ofsorghum plants (Fig. 1). Both plant height and weight were
reduced to a statistically significant degree. Naturally infectedplants were dwarfed more than inoculated, diseased plants.With both inoculation and natural infection of S. reilianum,>50% of the reduction in height occurred in the peduncleand the next two internodes below the panicle. In addition tothese data from 44 F, hybrids, similar statistically significanttrends were noted for three F2 lines, 12 F4, F5, and F7 lines,and nine inbred lines (data not given). Based on the occur-
rence of similar numbers of nodes in healthy and infectedplants, S. reilianlum did not significantly alter the date ofanthesis (data not given).The observations of symptoms of S. reilianum were re-
peated in 1990 for Tx7078, the cultivar used for GA studies.Typical sterile, phyllodied, smutted, and control plants were
harvested from 14 plots. The effect of head smut on heightwas the same as the previous year; height was reduced andthe effect was more strongly expressed in the youngest inter-node (peduncle) (Table I) of the infected plants. Those withsterile inflorescences exhibited the least dwarfing by headsmut, and there was no difference in height of inoculated andnaturally infected, diseased plants. There was a strong pro-motion of tillering in sterile and phyllodied plants comparedwith controls, which averaged less than one aerial tiller per
plant and no basal tillers.The C,8 HPLC fractions potentially containing free GAs
from extracts of smutted heads showed three major peaks ofGA-like activity in the dwarf rice bioassay (Fig. 2A). Thefraction groupings II and III were chromatographed again on
Table I. Effect of Head Smut on Growth and Development of Sorghum cv Tx7O78Data averages of 10 plants except where noted ± SD.
t 4 * * 10 It 14 16 1@ 0 StOgt24 IM 2 30 34 M6 3 4C
GA A B GA4 AIto CK2
4~~~~~~~~ 2 MI =4os' 1~~~~~~~~~~~OTO
10 20 30 40 10 20 30 40
HPLC RETENTION TIME (min)
Figure 2. Dwarf rice leaf sheath growth in response to HPLC frac-tions of extracts of panicles of sorghum plants with head smut diseasein an advanced stage of infection (plot No. 1095). A, results ofbioassays of C18 HPLC fractions; B and C, Nucleosil N(CH3)2 HPLCfractions of, respectively, the combined fractions 11 and IlIl from panelA. Elution of authentic GA standards are indicated.
a Nucleosil N(CH3)2 column, and the fractions showing GA-like activity (Fig. 2, B and C) were combined, an appropriatedeuterated standard was added, and the fractions were deriv-atized and subjected to GC-MS-SIM. The detection of endog-enous GAs was based on HPLC retention times, co-chroma-tography on the GC with appropriate deuterated standards,KRI values, and relative abundances of three ions. These dataare shown for smutted panicles in an advanced stage of thedisease (Table II). GA8 was detected in fraction I, GA, andGA3 in fraction 112, GA14 and GA53 in fraction III,, GA7 andGA20 in fraction 1112, and GA19 in fraction I113. The GA-likebioactivity in fractions II, and III4 could not be identified bythe GC-MS-SIM data obtained on those fractions. In sorghumpanicles showing early stages ofsmut disease symptoms, GA4,GA29, and GA44 were also detected (Table III), although theyhad not been detected in panicle tissue exhibiting a moreadvanced stage of symptom expression (Table II). Similardata for the bioassay as well as for the GC-MS-SIM detectionand quantification of endogenous GAs were obtained for theother samples. Authentic standards for GA4, GA7, GA8, GA14,GA29, and GA44 were not available, but we compared pub-lished KRI values, ion masses, and relative abundances (12,26) to the values in Tables II and III, and we made thetentative detections ofGA4, GA7, GA8, GA,4, GA29, and GA44listed in Tables II and III.
Quantitative analysis of endogenous GAs was done forsmutted, sterile (reproductive development incomplete, butsori not visible) and control inflorescences from three differentplots (Table IV). GA3, GA19, GA20, and GA53 were detectedin most analyzed samples. Some panicles also contained GA44
Table II. GC-MS-SIM Results of Fraction Groupings (Fig. 2) from Nucleosil N(CH3)2 HPLC of Extractsof Panicles of Sorghum Plants with Head Smut in an Advanced Stage of Disease Symptom Expression(Plot No. 1095)
The relative abundances of the endogenous GAs have been corrected for contributions from addeddeuterated internal standards. Tentative identification of GA8, GA14, and GA44 is based on publishedKRI values, ions, and relative abundances.
Sample KRI Ions (m/z) and Relative Abundances in Parenthesesa
Table Ill. GC-MS-SIM Data from Fraction Groupings (Fig. 2)Resulting from Nucleosil (N(CH3)2 HPLC of Extracts of Panicles ofSorghum with Head Smut in Early Stage of Disease SymptomExpression (Plot No. 1132) and Control Panicles from the Same Plot
Tentative identification of GA4, GA29, and GA44 is based on com-parison of published KRI values, ions, and relative abundances withthe data here.
Sample KRI Ions (m/z) and Relative Abundancesin Parenthesesa
which suggests that the conversion of GA-3 to GA44 and GA19to GA,(1 leading to GA3 is the pathway that is active ininflorescences. In smutted panicles, in the three plots exam-
ined, the total amount of GA53, GA44, and GA,9 was reducedapproximately 90% in comparison with controls (Table IV).In the same comparison, the total amount of GA53, GA44,and GA, in sterile panicles was reduced by approximately 50to 80% compared to the controls (Table IV, plots 1095 and1580). GA14 was detected only in smutted inflorescences. Inthe smutted heads with teliospores (Table IV, plot 1095), theGA3 level was threefold higher than the control. In addition,GA, and GA7 were detected in the same sample. This was
not the case in smutted panicles exhibiting tissue proliferationand leafy growths, because here GA3 was lower in the smuttedthan in the control panicles, and GA1 and GA7 were not
detected in the smutted panicles (Table IV, plot 1580). Insterile panicles, although the concentration of some GAs was
reduced, others were not significantly different from the con-
trols. GA4, GA8, or GA,, were detected in only a few of thesamples.
Because smutted panicles with open sori produced more
GA3 than control panicles and they also contained GA, andGA7 (Table IV), we suspected that the smut pathogen is ableto produce GAs. Culture filtrates of S. reilianuim containedGA-like activity in the dwarf rice bioassay (Fig. 3). Richard'smedium, which was 4 d old and did not contain fungus, didnot contain any detectable GA by bioassay or GC-MS-SIManalysis (data not shown). Therefore, the GA activity occur-ring in the smut culture filtrates was produced by the fungus.The fractions were combined, as indicated in Fig. 3, andprepared for GC-MS-SIM. In the fraction groupings thatcontain GA4 as internal standard, we did not detect any ofthe GAs that had been found in the smutted sorghum panicletissues (Table IV) (GA4, GAI4, GA19, GA20, and GA53). EvenGA7, only detected in smutted tissue showing mature telios-pores (Table IV), was not present in the S. reilianum samplesanalyzed by GC-MS-SIM (Table V). We did detect GA, andGA3 in the fractions co-eluting with authentic GA, and GA3(Table V). Quantification of these GAs (Table VI) indicatedthat the GA, concentration increased as the cultures aged upto 10 d. The GA3 concentrations show the same pattern, butfrom 10 to 20 d in culture the GA3 concentration was reducedby 85%.
DISCUSSION
Reduction of sorghum plant height has long been recog-nized as a symptom of head smut (8) and that symptom was
strongly expressed in these experiments (Fig. 1). The occur-rence of height reduction across a wide range of sorghumvarieties and hybrids indicates that it is fundamental to thedisease process. That the reduction in height was greater fornaturally infected plants than those inoculated (Fig. 1) sug-
Table IV. Endogenous GA Content in Panicle Tissue of Sorghum Plants Measured by GC-MS-SIM with Deuterated Internal Standards for theIndividual GAs Except for GA8 and GA29 which were Measured Against [2H2]GAl; for GA4, GA7, and GA44 [2H2]GA2O Was Used; for GA14, [2H2]GA53
All values have been corrected for contributions from deuterated internal standards and for the recovery of the [3H]GAs used as internalstandards during the extraction and purification.
Figure 3. Dwarf rice leaf sheath growth in response to NucleosilN(CH3)2 HPLC fractions of extracts of S. reilianum from cultures 4(A), 7 (B), 10 (C), and 20 d (D) old. Panels 1 and 2 are the result ofbioassays of Nucleosil N(CH3)2 fractions from, respectively, fractions6 through 18 and 20 through 40 combined from C18 HPLC. Elutionof authentic GA standards and the grouping of fractions used for GC-MS-SIM measurements are indicated. Control refers to growth ofseedlings not treated with HPLC fractions of extracts and is shownas the horizontal line at approximately 10 mm.
gests that fungal invasion occurred earlier after germinationin the former or that the naturally occurring pathogen was
more virulent than the isolate used for inoculation. Except invery late flowering varieties, almost all internode elongationoccurs after floral initiation (23). However, the greatest inhi-bition of elongation occurred in the last internode (peduncle),and the reduction in internode length was progressively lessin internodes farther away from the panicle (Fig. 1). Thissame trend was recently noted for a natural head smut infec-tion in a commercial sorghum field (18). In addition, repro-ductively sterile plants showed less height reduction thansmutted plants ( 18). These observations are consistent with a
hypothesis that the panicle is a major source of GAs and thatinfection with S. reilianum reduces the supply ofGAs, therebyinhibiting elongation of the peduncle and adjacent internodesmost severely as the pathogen overruns the panicle and soridevelop. When symptoms are less severe, i.e. sterile heads,the production of GAs by the panicle is less affected and
dwarfing is not so severe. This hypothesis does not excludethe possibility that GAs are also made in nodes or internodesand thereby influence elongation.Tx7O78, the cultivar used in the GA analysis studies, re-
sponded to head smut the same way the larger and morediverse sorghum population did the year before (Table I).Again, sterile plants with no other visible disease symptoms(i.e. no visible sori) were less dwarfed than plants showingvisible sori or phyllody. In this test, smutted plants showed a
clear promotion of tillering in contrast to the controls (TableI). The most severely dwarfed plants (smutted) exhibited lessdisease-induced tillering than the less dwarfed plants (sterileor phyllody developed on a smutted head). This suggests thatthe most severely diseased plants are unable to support tillergrowth, whereas those with less severe symptoms experiencehormone changes that promote tillering, and they maintainsufficient energy to support tiller growth. It seems possiblethat the promotion of tillering (Table I) results from thereduction in GA levels observed in smutted and sterile plants(Table IV). GA3 application to sorghum inhibits tillering ( 14,16), and a genotype found to contain elevated GA levels doesnot tiller (2). Treatment of the GA-overproducing genotypewith the GA synthesis inhibitor tetcyclacis promoted tillering(2). It seems likely that in Tx7O78 head smut reduced GAlevels and thereby promoted tillering.
Based on the detection of GA53, GA44, GA,9, GA2o, GA,,and GA3 in the panicles (Tables II-IV), we conclude that theearly 1 3-hydroxylation pathway (21) occurs in sorghum pan-icles as previously detected in vegetative tissues (2, 25). Invegetative tissues, GA, has usually been the major biologicallyactive GA, and GA3 was found in smaller amounts (2).
Table V. GC-MS-SIM Results of Fraction Groupings (Fig. 3) fromNucleosil N(CH3)2 HPLC of Extracts of S. reilianum Cultures
The relative abundances of the endogenous GAs have been cor-rected for contributions from added deuterated internal standards.
Sample KRI Ions (m/z) and Relative Abundances inParenthesesa
Table VI. Endogenous GA Content in S. reilianum Cultures ofDifferent Ages Measured by GC-MS-SIM with Deuterated InternalStandards for the Individual GAs
All values have been corrected for contributions from deuteratedinternal standards and for the recovery of the [3H]GAs used as internalstandards during the extraction and purification.
Sample GA, GA3
d ng GA/g dry weight4 NDa 1 9.6a7 3.3a 86.9a
10 54.8a 192.1 a20 48.4 29.4
a Measurements done in duplicate from separate extractions; av-erages presented.
However, in panicle tissue, whether infected or healthy, GA3was more frequently detected and occurred in greater concen-trations than GA, (Table IV). Perhaps GA3 is a more stableGA than GA, or perhaps it is the GA stored in seeds. Incontrast, the ratio of GA, to GA3 in vegetative maize tissue is50:1 or greater( II).Because diseased plants were reduced in height (Fig. 1) and
exhibited a 50 to almost 100% reduction in GA concentration(Table IV), it seems possible that GAs are involved in theexpression of smut symptoms. This relationship would besimilar to the case with dwarf maize mutants in which theGA biosynthetic pathway is blocked, the concentration ofGAs is reduced, and plants fail to achieve normal shootelongation (1 1, 21).As noted by Takahashi et al. (26), it was once believed that
among fungi only Gibberella fujikuroi (Fusarium monili-forme) produces GAs. In 1979, GA4 was isolated from Spha-celoma manihoticola, a pathogen of cassava (24). Later, GA3was detected at low levels in Neurospora crassa (15). Thissuggested a wide distribution ofGAs in the Ascomycetes (26);however, the sorghum head smut organism, S. reilianuim, isa Basidiomycete (28) and as such is the first nonAscomycetefungus shown by GC-MS-SIM analysis to produce specificGAs. Previously, culture filtrates from a number of Basidi-omycetes were bioassayed with hybrid maize seedlings andno GA-like activity was detected (6). Later, Pegg (20) extractedfruiting bodies (sporophores) of five Basidiomycetes presentin mycorrhizal associations with trees and found GA-likeactivity in the dwarf-5 maize bioassay for all five species.There have also been two identifications of GAs in bacteria(1, 4). Our confidence in the isolation from S. reilianum isbased on several facts: GA activity did not occur in themedium on which the fungus was cultured, the culturesextracted were determined to be free of bacterial or fungal(especially Fusarium spp.) contamination, the bioassay dataand GC-MS-SIM data were consistent, GAs were detected byboth bioassay and GC-MS-SIM in several different lots offungus and culture medium grown at different times, and theGAs specifically detected (GA, and GA3) were compared todeuterium-labeled internal standards which co-eluted andproduced matching fragmentation patterns and KRIs (TableV). Thus, although it is likely that other GAs occur in S.reilianum, is seems clear that this Basidiomycete produces
GA, and GA3 when cultured on Richard's medium. It will beappropriate now to do large-scale extractions of S. reilianilmand obtain full-scan mass spectra of all the GAs present tofurther confirm their occurrence and determine which onesoccur other than GA, and GA3. We assume that GA4, GA7,and GAI4 (Tables II, II, IV), not associated with the early- 13-hydroxylation pathway (21) and not found in sorghum pre-viously (2, 25), are produced by S. reilianum infection. How-ever, two of the control panicle samples contained smallamounts of GA4, and we have no evidence to identify thesource of this GA.One question is why does a fungus that produces GAs in
culture reduce rather than increase plant height. GA3 foundin the fungal culture medium extract does promote shootelongation in sorghum (2). The most striking comparison inthe data is the reduction in early- I 3-hydroxylation precursorswhich occurred in all smutted or sterile panicle samplescompared to their appropriate controls (Table IV). This re-duction suggests that this pathway is blocked or that precur-sors are diverted to other uses by the fungus. The result couldbe a deficiency of bioactive GAs at the time of internodeelongation. As teliospore development progressed, we ob-served an increase in levels of bioactive GA, and GA3 (TableIV, compare plot 1132 with 1095) which might be associated,along with other hormones, with the initiation of tissue pro-liferation and leafy growths that become obvious after sporematuration. Recently, Bhaskaran et al. (3) demonstrated thataddition of GA3 to tissue culture media causes the reversionof floral primordia of sorghum to vegetative shoots. Thisresponse is similar to the phyllody symptoms of sorghumhead smut. It further suggests that smutted plants may firstexhibit symptoms resulting from low GA levels and latersymptoms resulting from an increase in GA levels. More workwill be necessary to clarify the status ofGAs in smutted plants.
ACKNOWLEDGMENTS
We wish to acknowledge with thanks the assistance of AllisonCollins for culture of S. reilianuin and collection of the data in Figure1 and Table II. In addition. Karen Babb assisted in extracting andassaying GAs and Kevin Childs maintained the GC-MS-SIM whilethis work was in progress. Professor B. 0. Phinney made helpfulsuggestions and encouraged the effort to study the fungal metabolitesseparate from those in diseased plant samples. In the early stages ofthis project. T. Chuteemun and, later, John Ambler observed growthregulator activity in bioassays of S. reilianium cultures.
LITERATURE CITED
1. Atzorn R, Crozier A, Wheeler CT, Sanberg (1988) Production ofgibberellins and indole-3-acetic acid by Rhizobhin phaseolinin relation to nodulation of Phaseoluis i'ulgaris roots. Planta175: 532-538
2. Beall FD, Morgan PW, Mander LN, Miller FR, Babb KH (1991)Genetic regulation of development in Sorghuim bicolor. V. ThemaR3 allele results in gibberellin enrichment. Plant Physiol 95:116-125
3. Bhaskaran S, Smith RH, Frederiksen RA (1990) Gibberellin A,reverts floral primordia to vegetative growth in sorghum. PlantSci 71: 113-118
4. Bottini MF, Pearce D, Pharis RP (1989) Identification of gib-berellins A, A,. and Iso-A3 in cultures of .4zospirillutm lipo-tenln. Plant Physiol 90: 45-47
5. Casady AJ (1969) Propagation from proliferated sorghum spi-kelets caused by head smut infection. Crop Sci 9: 381-382
6. Curtis RW (1957) Survey of fungi and actinomycetes for com-pounds possessing gibberellin-like activity. Science 125: 3249
7. Edmunds LK (1963) Use of sporidial hypodermic injection totest for head smut resistance. Plant Dis Rep 47: 909-913
8. Frederiksen RA (1977) Head smuts ofcorn and sorghum. In HDLoden, D Wilkinson, eds, Proceedings of the 32nd AnnualCorn Sorghum Conference. American Seed Trade Association,Washington, pp 89-105
9. Frederiksen RA (1986) Head smut. In RA Frederiksen, ed,Compendium ofSorghum Diseases. The American Phytopath-ological Society, St. Paul, MN, pp 17-20
10. Frowd JA (1980) A world review of sorghum smuts. In RJWilliams, RA Frederiksen, LK Mughogho, eds, Sorghum Dis-eases: A World Review. International Crops Research Institutefor the Semi-Arid Tropics, Patancheru, India, pp 331-348
11. Fujioka S, Yamane H, Spray CR, Gaskin P. MacMillan J,Phinney BO, Takahashi N (1988) Qualitative and quantitativeanalyses of gibberellins in vegetative shoots of normal, dwarf-1, dwarf-2, dwarf-3 and dwarf-5 seedlings of Zea maYs L. PlantPhysiol 88: 1376-1372
12. Hedden P (1987) Gibberellins. In L Rivier, A Crozier, eds,Principles and Practices of Plant Hormone Analysis, Vol 1.Academic Press, London, pp 9-110
13. Irish EE, Nelson T (1989) Sex determination in monoecious anddioecious plants. Plant Cell 1: 737-744
14. Isbell VR, Morgan PW (1982) Manipulation of apical domi-nance in sorghum with growth regulators. Crop Sci 22: 30-35
15. Kawanabe Y, Yamane H, Murayama T, Takahashi N, NakamuraT (1988) Identification of gibberellin A3 in mycelia of Neuro-spora crassa. Agric Biol Chem 49: 2447-2450
16. Morgan PW, Miller FR, Quinby JR (1977) Manipulation ofsorghum growth and development with gibberellic acid. AgronJ 69: 789-793
17. Murakami Y (1968) A new rice seedling test for gibberellins,"microdrop method," and its use for testing extracts of riceand morning glory. Bot Mag (Tokyo) 81: 33-43
18. Naidoo C, Morgan PW, Collins AC, Frederiksen RA (1991)Effect of Sporisorium reilianum on plant height and internodeelongation in Sorghum bicolor in a naturally infected field.Sorghum Newslett 31: In press
19. Nash SM, Snyder WC (1962) Quantitation estimations by platecounts of propagules of the bean root rot Fusarium in fieldsoils. Phytopathology 52: 567-572
20. Pegg GF (1973) Occurrence of giberellin-like growth substancesin Basidiomycete sporophores. Trans Br Mycol Soc 61:277-286
21. Phinney BO, Spray C (1982) Chemical genetics and the gibber-ellin pathway in Zea mays L. In PF Wareing, ed, Plant GrowthSubstances 1982. Academic Press, London, pp 101-110
22. Potter AA (1914) Head smut of sorghum and maize. J Agric Res2: 339-372
23. Quinby JR (1973) The genetic control of flowering and growthin sorghum. In NC Brady, ed, Advances in Agronomy, Vol25. Academic Press, New York, pp 125-162
24. Rademacher W, Graebe JE (1979) Gibberellin A4 produced bySphaceloma manihoticola, the cause of super-elongation dis-ease of cassava (Manihot esculenta). Biochem Biophys ResCommun 91: 35-40
25. Rood SB, Larsen KM, Mander LN, Abe H, Pharis RP (1986)Identification of endogenous gibberellins from sorghum. PlantPhysiol 82: 575-580
26. Takahashi N, Yamaguchi I, Yamane H (1986) Gibberellins. InN Takahashi, ed, Chemistry of Plant Hormones. CRC Press,Boca Raton, FL, pp 57-151