Contents lists available at SciVerse ScienceDirect
Experimental Gerontology
j ourna l homepage: www.e lsev ie r .com/ locate /expgero
Improvement of oxidative stress and immunity by melatonin: An age dependentstudy in golden hamster
Dipanshu Kumar Vishwas a, Arun Mukherjee a, Chandana Haldar a,⁎, Debabrata Dash b, Manasa K. Nayak b
a Department of Zoology, Banaras Hindu University, Varanasi-221005, Indiab Department of Biochemistry, Institute of Medical Science, Banaras Hindu University, Varanasi-221005, India
Reactive oxygen species (ROS) have been proposed to play an important role in balancing the pro- and anti-oxidant homeostasis during aging. Melatonin has been suggested as an effective free radical scavenger thatmight have a role during the process of aging. We observed, that melatonin administration (25 μg/100 gbody weight for 30 days) significantly augments the activity of anti-oxidative enzymes like superoxidedismutase (SOD), catalase and glutathione peroxidase (GPx) in the plasma, spleen and bone marrow (BM)of young (6 weeks), adult (30 weeks) and old aged (2.5 years) male golden hamster, Mesocricetus auratus.A sharp decline in generation of ROS was observed in peripheral blood mononuclear cells (PBMC) andsplenocytes upon melatonin administration in different age group of hamsters. Reduction in the level ofthiobarbituric acid-reactive substances (TBARS) and total nitrite and nitrate concentration as metabolitesand indicators of nitric oxide (NO) in plasma, spleen and BM were observed along with night time(22:00 h) melatonin concentration in different age group of hamsters after administration of melatoninand compared to the control group (treated with 0.9% saline). General immune parameters like proliferationof splenocytes, PBMC and colony forming ability of GM-CFU were observed following melatonin treatment indifferent age group, although it was low only in aged hamsters compared to the young and adult. Our dataindicates that the age related increase of oxidative load and simultaneously augments the general immunityin aged hamsters.
Aging is a complex process affecting a wide variety of physiologicalfunctions, including the development and maintenance of the periph-eral immune system (Miller, 1991). The oxidative theory of aging byHarman (1956) states that reactive oxygen species (ROS) and nitrogenspecies (RNS) cause random damage to cells, which includes impairedphysiological functions and increased incidence of diseases as the ageprogresses leading to mortality. ROS and RNS both induce oxidativestress during several physiological conditions like inflammation, andare responsible for increased oxidative load with advancing age(Drew and Leeuwenburgh, 2002). The nitrosative stress due to interac-tion between nitrosants and oxidants may produce products that aremore toxic than either reactant alone causing cellular injury, predomi-nate oxidative damage (Eu et al., 2000; Stamler and Hausladen, 1998).At the cellular level, nitric oxide (NO) has been widely implicated innitrosative stress which is linked to inhibition of cell growth and
+91 542 2307149x209, +91
).
rights reserved.
apoptosis (Marshall et al., 2000). The detection of nitrotyrosine(NO2-Tyr) formation in various tissue following inflammation, andduring the process of aging is recognized as peroxynitrite-triggeredmechanism of nitrosative injury (Beal, 2002; Bian and Murad, 2001;Drew and Leeuwenburgh, 2002; Levrand et al., 2005). Oxidative stressmay also result in wide varieties of diseases and neurodegenerative dis-orders (Pacher and Szabo, 2008).
Decline in the production of several hormones including melato-nin has been reported (Reiter et al., 1981) to play a key regulatoryrole in aging and senescence (Liu and Wang, 2002; Pierpaoli andRegelson, 1992) by preventing the T lymphocyte mediated immuneresponses (Miller, 1995). Melatonin is known for immunomodulation(Ahmad and Haldar, 2010) and reflects a general immunoenhancingproperty in many animal species including human (Nelson, 2004).Numbers of studies suggest that pinealectomy induces variouspathological changes resembling senescence and could be reversedby the administration of pineal extract or melatonin. The mechanismof action of melatonin in immune-enhancement needs to beelucidated.
Though, the direct free radical scavenging capacity (Tan et al.,1993) and indirect antioxidant stimulatory property of melatoninhave been reported in number of studies for improving metabolic
169D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
function (Reiter et al., 2000; Rodriguez et al., 2004). To date, the agerelated protective effect of melatonin in immune function and oxida-tive stress in terms of ROS/RNS generation is lacking. We thereforeexplored the relationship between the age dependent melatonin se-cretion and modulation of immune responses along with oxidativestress in different age group of hamster under ambient condition.We also explored the protective effect of exogenous melatoninagainst age induced oxidative injury by accessing ROS generation,lymphocyte proliferation and colony forming ability of granulocytemacrophages (GM-CFU) in different age group of hamsters.
Fig. 1. Flow cytometric analysis with age and melatonin administration showing ROS gehistograms showing DCFH-detectable ROS level (MFI) in (c) PBMC and (d) splenocytes. V(OH) hamsters M. auratus; apb0.01 vehicle vs melatonin treated hamster.
2. Methods
2.1. Animals
Golden hamsters (outbred strain) were procured from Central DrugResearch Institute (CDRI), Lucknow, India and colony was developedand maintained in animal house facility. Hamsters were kept in aroom having temperature 25±2 °C with alternative light/dark cycle(12 h light, 12 h dark; i.e. lights on 07:00–19:00 h) in order tomaintainthe photoperiod. Animals were maintained in polypropylene cages
neration in (a) PBMC and (b) splenocytes. Results are expressed as representativealues are expressed as MFI±SD, N=7. **pb0.01 young (YH) vs adult (AH) and old
(475×350×200 mm) in a group of seven, supplemented with waterand commercial rodent pellet ad libitum. All experiments wereconducted in accordance with Institutional guidelines following theframework of revised Animal (Specific Procedure) Act of 2002, Govern-ment of India for animal welfare.
2.2. Experiment
Male golden hamster were randomly selected according to theirage and divided into three groups each containing fourteen animals.Group I consists of young hamsters and served as young-aged group(YH; weighing 60±10 g, 6 weak old), group II accounts for adulthamsters (AH; weighing 110±10 g, 30 weak old) and group III in-clude old hamsters (OH; weighing 150±10 g, 2.5 year old). Eachgroup of hamsters was divided into two set, 7 animals in each set,
Fig. 2. Effect of age and melatonin (25 μg/100 g body weight daily) treatment on DCF fluoadult (AH) and old (OH) hamsters. Scale bar represents 50 μm.
treated with melatonin or vehicle control. Melatonin was dissolvedin 10 μl of ethanol followed by dilution with normal saline (0.9%NaCl) in order to make desired concentration for subcutaneous injec-tion at a dose of 25 μg/100 g body weight/hamster/day during theevening hours (between 18:00 and 18:30 h~1 h prior to sunset) for30 days. The control hamsters received ethanolic saline as vehicle.
2.3. Sample collection
The animals were sacrificed by decapitation following deep etheranesthesia during night time between (20:00 h and 22:00 h, IST).The trunk blood was collected in heparinized tubes and centrifugedimmediately. Plasma was kept at −20 °C for hormonal and biochem-ical estimations. Bone marrow (BM) cells were collected in culturemedium and processed for setting of GM-CFU culture. Splenocytes
rescence intensity for ROS generation in (a) PBMC and (b) splenocytes of young (YH),
171D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
and PBMC were processed for proliferation assay which is expressedin terms of % stimulation ratio (%SR) of lymphocyte proliferationcompared to control. A portion of the cell preparation was also keptfor FACS analysis and biochemical estimations for ROS/RNS genera-tion. Tissue homogenate was prepared in 0.01 M phosphate buffer(pH=7.4) from bone marrow and spleen for determination of nitriteand nitrate concentration.
2.4. Total nitrite and nitrate concentration for NO estimation
Total nitrite and nitrate concentration, an indication of NO synthe-sis, was measured in plasma, spleen and BM following the method de-scribed by Sastry et al. (2002) and Di Fiore et al. (2008). Briefly, plasma,5% tissue homogenate (5% w/v, spleen and BM) or KNO3 (100 μL ofeach) was added to 400 μL of carbonate buffer followed by the additionof small amount (0.15 g) of activated copper–cadmium alloy fillingsand incubated at room temperature with intermittent shaking. The re-action was stopped by the addition of 100 μL of 0.35 M NaOH followed
by the addition of 120 mM ZnSO4 solution. The solution was mixedthoroughly and allowed to stand for 10 min followed by centrifugationat 800 rpm for 10 min. 100 μl supernatant was added in 96 well platein presence of Griess reagent (50 μl of 1% sulphanilamide in 2.5%orthophosphoric acid+50 μL of 0.1% N-naphthyl ethylenediamine indistilled water) in quadruplicate. After 10 min, the absorbance wasmeasured at 545 nm with ELISA reader (ELx-800, Biotek Instruments,VT, USA). A standard graph was plotted against different concentra-tions (0, 20, 40, 60, 80 and 100 μM) of KNO3.
2.5. Measurement of ROS levels
ROS generationwasmeasured byflowcytometry usingDCFH-DAdyeas described previously (Degli Esposti andMcLennan, 1998). Splenocytesand PBMCwere isolated in PBS supplemented with 50 mM glucose, andincubated with 10 μM DCFH-DA at 37 °C for 1 h. Fluorescence genera-tion due to the hydrolysis of DCFH-DA to dichlorodihydrofluorescein(DCFH) by non-specific cellular esterases and the subsequent oxidation
of DCFH by peroxides was measured in terms of mean fluorescence in-tensity (MFI) and analyzed using ‘Cell Quest 3.3’ software.
2.6. Determination of reactive oxygen species (ROS)
To determine the DCFH-detectable ROS, the fluorescent probeDCFH-DA was used. DCFH-DA is cleaved by intracellular esterases toproduce an impermeable, non-fluorescent active form (DCFH),which further reacts with ROS to form a fluorescent product DCF(Jakubowski and Bartosz, 2000; Rastogi, 2010). The cells werewashed with PBS and then stained with DCFH-DA (10 μM) for30 min at 37 °C. The cells were then washed with PBS and fluores-cence images were obtained by a fluorescence microscopy (LeitzVario Orthomat 2, Wetzlar GMBH, Germany) with digital camera(Leica EC3 with software) at an excitation setting of 488 nm.
2.7. Lipid peroxidation (LPO) assay
A 10% homogenates of spleen tissue in 20 mM Tris Hydrochloride(HCl) buffer (pH 7.4) was centrifuged and supernatant was subjectedto thiobarbituric acid (TBA) assay by mixing it with 8.1% SDS, 20%
Fig. 3. Total nitrite and nitrate concentration in (a) plasma and homogenates of(b) spleen and (c) bone marrow of young (YH), adult (AH) and old (OH) hamsters.Values are expressed as mean±SD, N=7. **pb0.01YH vs AH and OH; apb0.01 vehiclevs melatonin treatment.
acetic acid, 0.8% TBA and boiling for 1 h at 95 °C. The reaction mixturewas cooled and shaken with n-butanol and pyridine reagent (15:1)and centrifuged (Ohkawa et al., 1978). The absorbance of reaction mix-turewasmeasured at 534 nm. LPOwas expressed as thiobarbituric acidreactive substances (TBARS) in nmol/g tissue weight having 1, 1, 3,3-tetraethoxy propane (TEP) as standard. The standard curve was cali-brated using 10 nM concentration of TEP.
2.8. Estimations of antioxidant activity
The spleen and BM tissue was homogenized in PBS and the proteinlevels were estimated using BSA as the standard (Bradford, 1976).The supernatant of 10% homogenate was centrifuged and processedfor total Cu, Zn-Superoxide dismutase (SOD) and Mn-SOD activity.The reaction was stopped by adding freshly prepared Griess reagentand the OD was taken at 543 nm. SOD activity expressed as specificactivity (units/mg protein) (Das et al., 1999).
Fig. 4. Age related changes and effect of melatonin administration on concentration ofthiobarbituric acid-reactive substances (TBARS) in (a) plasma and homogenates of(b) spleen and (c) bone marrow of young (YH), adult (AH) and old (OH) hamsters.Values are expressed as mean±SD, N=7. **pb0.01 and *pb0.05 YH vs AH and OH;apb0.01 vehicle vs melatonin treatment.
173D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
The indirect catalase activity was measured using hydrogenperoxide (H2O2) as a substrate and was expressed as mM/ml/mg ofprotein (Sinha, 1972). The supernatant of 10% homogenates in PBSwas processed for the assay in a reaction mixture comprising0.8 mM H2O2 and potassium dichromate in glacial acetic acid heatedin a water bath for 10 min and then OD was taken at 570 nm and thedecrease in H2O2 content was calculated. The standard curve was cal-ibrated with varying concentrations of 0.2 mM H2O2 in PBS.
Glutathione peroxidase (GPx) activity was assayed as described byMantha et al. (1993). The reactionmixture (1 ml) contained 50 μl sam-ple, 398 μl of 50 mM phosphate buffer (pH 7.0), 2 μl of 1 mM EDTA,10 μl of 1 mM sodium azide, 500 μl of 0.5 mM NADPH, 40 μl of0.2 mM GSH and 1 U glutathione reductase. The reaction mixturewas allowed to equilibrate for 1 min at room temperature. After this,the reaction was initiated by addition of 100 mMH2O2. The absorbancemeasured kinetically at 340 nm for 3 min. The GPx activity wasexpressed as nmol of oxidized NADPH oxidized to NADP+ per minper mg of protein using an extinction coefficient (6.22 mM−1 cm−1)for NADPH.
Fig. 5. Aging and effect of melatonin administration on catalase (CAT) enzyme activityin (a) plasma and homogenates of (b) spleen and (c) bone marrow of young (YH),adult (AH) and old (OH) hamsters. Values are expressed as mean±SD, N=7.**pb0.01 YH vs AH and OH; apb0.01 vehicle vs melatonin treatment.
Fig. 6. Aging and effect of melatonin administration on SOD enzyme activity in(a) plasma and homogenates of (b) spleen and (c) bone marrow of young (YH),adult (AH) and old (OH) hamsters. Values are expressed as mean±SD, N=7.**pb0.01 YH vs AH and OH; apb0.01 vehicle vs melatonin treatment.
2.9. Cell cycle analysis
Splenocytes and PBMC were isolated at a concentration of 1×106
cells/ml, washed and centrifuged. The cell pellet was resuspended in50 μl cold PBS and fixed in 2 ml of 70% ice-cold ethanol, againcentrifuged and treated with 0.1% Triton X-100 for 5 min. Ribonucle-ase (100 μg/ml) was added and the cells were incubated at 37 °C for30 min. After centrifugation cells were resuspended in 1 ml of PBScontaining 50 μg/ml propidium iodide (PI) and incubated for30 min at 4 °C (Srivastava et al., 2012). The samples were analyzedby flow-cytometer.
2.10. Morphological analysis of apoptotic cells
Apoptotic cells were microscopically analyzed following AcridineOrange–Ethidium Bromide (AO–EB) double staining. AO–EB dye ofvolume 0.01 ml (1×) was admixed gently with 0.2 ml of the dilutedsample (1×106 cells/ml in PBS). A drop of this mixture was placedunderneath coverslip on a clean slide and cells were observed imme-diately under fluorescence microscope (Leitz MPV3, Wetzlar, Hesse,Germany) at 440–520 nm. A minimum of 200 cells was counted inevery sample to observed cell death (Sharma and Haldar, 2009).
Fig. 7. Aging and effect of melatonin administration on glutathione peroxidase (GPx)activity in (a) plasma and homogenates of (b) spleen and (c) bone marrow of young(YH), adult (AH) and old (OH) hamsters. Values are expressed as mean±SD, N=7.**pb0.01 YH vs AH and OH; apb0.01 vehicle vs melatonin treatment.
2.11. Measurement of cell proliferation by MTT assay
Cell-mediated immune function was assessed by measuringsplenocytes proliferation in response to the T-cell specific mito-gen, Concanavalin-A (Con-A), using a colorimetric assay basedon the reduction of tetrazolium salt Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) (Ahmad et al., 2012; Mosmann,1983). In short spleen of hamsters was removed in sterile conditionand a single-cell suspension was prepared with culture medium(RPMI-1640 supplemented with penicillin, streptomycin, L-glutamine,2-mercaptoethanol and heat-inactivated fetal calf serum. Viability wasdetermined with a hemocytometer and trypan blue exclusion method.Viable cells (which exceeded 95%) were adjusted to 1×107 cells/ml.100 μL aliquots of each cell suspensionwere added to thewells of sterileflat-bottom 96-well culture plates. Concanavalin-A (Sigma-Aldrich,St. Louis, USA) was added to the culturemedium at the concentrationof 5 μg/ml. Plates were incubated at 37 °C with 5% CO2 for 69 hprior to addition of 10 μL of MTT (SRL, Bombay, India; 5 mg/mlin phosphate-buffered saline) per well. Acidified propanol (0.04 mol/L
HCL in isopropanol) was added and the optical density (OD) was deter-mined with a microplate reader (ELx-800, Biotek Instruments, WinooskiVT, USA) equipped with a 570 nmwavelength filter. Mean OD valuesfor each set of duplicates were used in subsequent statistical analy-ses. Response was calculated as percent stimulation index (%SR)representing the ratio of absorbance of mitogen stimulated cultures tocontrol cultures.
2.12. Isolation and cell proliferation of PBMC
A modified method of Bienzle et al. (2003) was used for isolationof PBMC. PBMC were isolated from the heparinized blood collectedfrom heart in a 15 ml tube. Blood was diluted in 1:1 ratio with PBSand under layered with Ficoll hicep™ LSM (Himedia Lab, India) asdescribed by Boyum (1968). White ring of mononuclear cells collect-ed, washed with PBS, centrifuged and suspended in RPMI 1640supplemented with 10% fetal calf serum and 100 U of penicillin andstreptomycin. The cell concentration was adjusted to 1×107 viablecells/ml and plated in 96 well plate in duplicate having concentrationof (2×106cells/well) for the measurement of cell proliferation byMTT assay and OD values at 570 nm were taken and the %SR wascalculated.
2.13. Bone marrow isolation and GM-CFU culture
Modified method of Haldar et al. (1992) was used for the isolationof bone marrow mononuclear cells (BM-MNC). In brief femur bonewas dissected out and bone marrow was collected by flushing itwith PBS, washed with sterile medium IMDM (GIBCO). 1.0×107
mononuclear cells (isolated from the BM with the Ficoll Hicep™ LSMpreparation and counted in Neubauer's chamber) per 35/10 mm dishwere cultured in an assay medium of methylcellulose (Sigma-AldrichChemicals, St. Louis, USA) with fetal calf serum (heat inactivated,Sigma-Aldrich, St. Louis, Missouri, USA) and IMDM 2× medium(GIBCO). The culture plates were incubated for total 7 days at 37 °Cwith 5% CO2 and in a HERA cell CO2 incubator (Kendro LaboratoryProducts (India) Pvt. Ltd., New Delhi, India) after stimulating the col-onies with 250 U of recombinant murine colony stimulating factor(rGM-CSF; Peprotech, USA). The developed colonies (containing 50or more cells) were counted using an inverted microscope at ×20magnification.
2.14. ELISA for plasma melatonin
The assay was performed according to the manufacturer's protocolprovided with the kit of Uscn Life Science Inc. USA.
2.15. Statistical analysis
Statistical analysis of the data was performed with two-wayANOVA comparing effect of melatonin treatment with age group byStatistical Package for the Social Sciences (SPSS). Further, one-wayANOVA followed by Student Newman–Keuls multiple range tests toobserve the age dependent variation while t-test was performed forcontrol vs melatonin in all age group. The differences were consid-ered significant when pb0.05 (Bruning and Knitz, 1977). Correlationanalysis was also performed with different parameters in relationwith age and melatonin treatment. Microsoft Excel program wasused for statistical analyses and documentation.
3. Results
3.1. DCFH-detectable ROS generation
The DCFH-detectable ROS levels were determined in terms of MFIin PBMC (Fig. 1a) and splenocytes (Fig. 1b) of hamsters. A two-way
175D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
ANOVA was conducted that examined the effect of melatonin treat-ment with age on ROS. There was significant interaction betweenthe effects of age and melatonin treatment in DCFH-detectedROS level in PBMC F(2, 12)=583.362, p b0.01; and splenocytes(F(2, 12)=469.638, pb0.01. Further, old (pb0.01) and adult(pb0.05) hamsters showed significant increase in ROS productionwhen compared with young hamsters and showed a strong positivecorrelation (PBMC r=0.992, pb0.01; and splenocytes r=0.719,pb0.01) with age. Administration of melatonin decreased significantly(pb0.01) DCFH-detectable ROS production by PBMC and splenocytesof different aged hamsters whichwas presented in histogram followingFACS data analysis (Fig. 1c and d). ROS generation was also observed byfluorescence microscopy in different age group after administration ofmelatonin that presented a decrease in ROS level (Fig. 2a and b) andhad negative correlation (PBMC r=−0.979, p>0.05) and no correlation(splenocytes r=0.366, pb0.01) with circulatory melatonin level.
3.2. Nitrite and nitrate ion estimation
Age-dependent increase in nitrite/nitrate ions (as metabolite and in-dicator of NO) in plasma (Fig. 3a), spleen (Fig. 3b) and BM (Fig. 3c) ofhamsters was noted. A two-way ANOVA presented significantinteraction between the effects of age and melatonin treatment innitrite/nitrate ions concentration in plasma (F(2, 12)=14.959, pb0.01),spleen (F(2, 12)=26.692, pb0.01), and BM (F(2, 12)=0.038, p>0.05).Further, adult and old hamsters showed significant (pb0.01) increasein NO concentration in plasma and spleen while only BM of old ham-sters showed significant (pb0.05) increase in NO concentration when
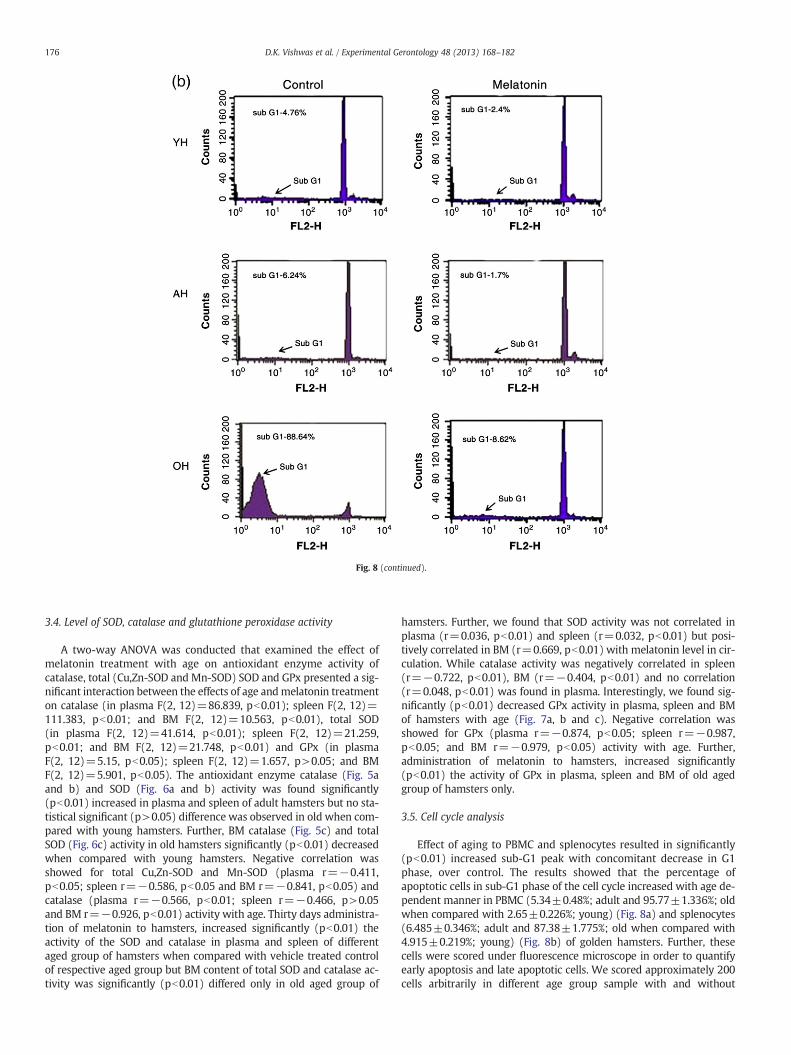
Fig. 8. Effect of age and melatonin administration on cell cycle progression in (a) PBMC andsured by flow-cytometry analysis. Control cells of young and adult hamsters had no sub-G1 phamsters had some cells in sub-G1phase. Propidium Iodide (PI) fluorescence intensity was
compared to the young hamsters (Fig. 3a, b and c) and had positive cor-relation (plasma r=0.844, p>0.05; spleen r=0.758, p>0.05) withage. Administration of melatonin decreased significantly (pb0.01) NOconcentration in plasma and spleen of adult and old hamsters and hadnegative correlation (plasma r=−0.887, p>0.05; spleen r=−0.751,p>0.05; and BM r=−0.833, pb0.05) with circulatory melatoninlevel.
3.3. Level of lipid peroxidation
The levels of plasma TBARS (as product of LPO) increased in adultand old hamsters. Two-way ANOVA suggested a significant inter-action between the effects of age and melatonin treatmentwith TBARS level in plasma (F(2, 12)=145.049, pb0.01), spleen(F(2, 12)=9.321, pb0.01), and BM (F(2, 12)=4.715, pb0.05). Fur-ther, plasma and spleen TBARS levels were found to be significantly(pb0.01) higher in adult and old hamsters than the young ones(Fig. 4a and b). Significantly (pb0.05) high BM TBARS level was alsonoted in adult and old (pb0.01) hamsters when compared withyoung one (Fig. 4c) and showed a positive correlation (plasma r=0.957, pb0.05; spleen r=0.857, p>0.05 and BM r=0.746, pb0.01)with age. Melatonin significantly (pb0.01) decreased the level ofLPO in the plasma, spleen and BM of adult and old hamsters whencompared with vehicle treated control of respective age group ofhamsters and found no correlation in plasma (r=0.373, pb0.01),and negative correlation in spleen (r=−0.661, pb0.01) and BM(r=−0.612, pb0.01) with circulatory melatonin level.
(b) splenocytes of young (YH), adult (AH) and old (OH) hamsters M. auratus as mea-eak, whereas old hamsters all cells were in sub-G1phase. Further, melatonin treated oldmeasured as an indicator of cellular DNA content.
3.4. Level of SOD, catalase and glutathione peroxidase activity
A two-way ANOVA was conducted that examined the effect ofmelatonin treatment with age on antioxidant enzyme activity ofcatalase, total (Cu,Zn-SOD and Mn-SOD) SOD and GPx presented a sig-nificant interaction between the effects of age andmelatonin treatmenton catalase (in plasma F(2, 12)=86.839, pb0.01); spleen F(2, 12)=111.383, pb0.01; and BM F(2, 12)=10.563, pb0.01), total SOD(in plasma F(2, 12)=41.614, pb0.01); spleen F(2, 12)=21.259,pb0.01; and BM F(2, 12)=21.748, pb0.01) and GPx (in plasmaF(2, 12)=5.15, pb0.05); spleen F(2, 12)=1.657, p>0.05; and BMF(2, 12)=5.901, pb0.05). The antioxidant enzyme catalase (Fig. 5aand b) and SOD (Fig. 6a and b) activity was found significantly(pb0.01) increased in plasma and spleen of adult hamsters but no sta-tistical significant (p>0.05) difference was observed in old when com-pared with young hamsters. Further, BM catalase (Fig. 5c) and totalSOD (Fig. 6c) activity in old hamsters significantly (pb0.01) decreasedwhen compared with young hamsters. Negative correlation wasshowed for total Cu,Zn-SOD and Mn-SOD (plasma r=−0.411,pb0.05; spleen r=−0.586, pb0.05 and BM r=−0.841, pb0.05) andcatalase (plasma r=−0.566, pb0.01; spleen r=−0.466, p>0.05and BM r=−0.926, pb0.01) activity with age. Thirty days administra-tion of melatonin to hamsters, increased significantly (pb0.01) theactivity of the SOD and catalase in plasma and spleen of differentaged group of hamsters when compared with vehicle treated controlof respective aged group but BM content of total SOD and catalase ac-tivity was significantly (pb0.01) differed only in old aged group of
hamsters. Further, we found that SOD activity was not correlated inplasma (r=0.036, pb0.01) and spleen (r=0.032, pb0.01) but posi-tively correlated in BM (r=0.669, pb0.01) with melatonin level in cir-culation. While catalase activity was negatively correlated in spleen(r=−0.722, pb0.01), BM (r=−0.404, pb0.01) and no correlation(r=0.048, pb0.01) was found in plasma. Interestingly, we found sig-nificantly (pb0.01) decreased GPx activity in plasma, spleen and BMof hamsters with age (Fig. 7a, b and c). Negative correlation wasshowed for GPx (plasma r=−0.874, pb0.05; spleen r=−0.987,pb0.05; and BM r=−0.979, pb0.05) activity with age. Further,administration of melatonin to hamsters, increased significantly(pb0.01) the activity of GPx in plasma, spleen and BM of old agedgroup of hamsters only.
3.5. Cell cycle analysis
Effect of aging to PBMC and splenocytes resulted in significantly(pb0.01) increased sub-G1 peak with concomitant decrease in G1phase, over control. The results showed that the percentage ofapoptotic cells in sub-G1 phase of the cell cycle increased with age de-pendent manner in PBMC (5.34±0.48%; adult and 95.77±1.336%; oldwhen compared with 2.65±0.226%; young) (Fig. 8a) and splenocytes(6.485±0.346%; adult and 87.38±1.775%; old when compared with4.915±0.219%; young) (Fig. 8b) of golden hamsters. Further, thesecells were scored under fluorescence microscope in order to quantifyearly apoptosis and late apoptotic cells. We scored approximately 200cells arbitrarily in different age group sample with and without
177D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
melatonin treatment. Acridine orange (AO) stained both apoptotic andviable cells emitting green fluorescence when bound to double strand-ed DNA and red fluorescence when bound to single stranded RNA. Vi-able cells fluoresced uniform bright green nuclei with organizedstructure while early apoptotic cells showed intact membranes andchromatin condensation as bright green patches or fragments andlate apoptotic cells showed orange to red nuclei with condensed orfragmented chromatin. Ethidium bromide (EB) is taken up only bydead cells and emitted red fluorescence when intercalated with DNA.Necrotic cells showed a uniformly orange to red nuclei with organizedstructure. We found almost all the cells in young and adult hamsterwith or without treatment of melatonin emitted green fluorescencewith intact nuclear structure. In old aged hamster a large number ofstructure known as early as well as late apoptotic cells having yellow-ish orange color indicating fragmented DNA. In addition, the late stagesof apoptosis such as the presence of reddish orange color due to thebinding of AO to denatured DNA were observed. However treatmentof melatonin to old hamster reduced above mentioned staining
Fig. 9. Morphological changes observed in (a) PBMC and (b) splenocytes stained with Acrimagnification. Arrow (→) indicates control, plus (+) indicates early apoptotic cells, asteris
expression and more green cells (normal) were observed when com-pared with control old hamster (Fig. 9 a and b).
3.6. Lymphocyte proliferation index of PBMC and Splenocytes
There was no significant interaction between the effects of age andmelatonin treatment on %SR of PBMC F(2,12)=2.341, p>0.05 whilesignificant interaction was observed in %SR of splenocytes F(2, 12)=4.415, pb0.05. Old vehicle treated control hamsters showed a signifi-cant (pb0.01) decrease in %SR of PBMC (Fig. 10a) and splenocytes(Fig. 10b) when compared with young hamsters and showed negativecorrelation (PBMC r=−0.863, pb0.01; and splenocytes r=−0.769,pb0.01) with age. Melatonin treatment significantly (pb0.01) en-hanced the %SR of PBMC and splenocytes in all age group when com-pared with vehicle treated controls of respective age groups ofhamsters and found strong positive correlation (PBMC r=0.932,pb0.01; and splenocytes r=0.857, pb0.01) with circulatory melatoninlevel.
dine Orange–Ethidium Bromide (AO–EB) dye under fluorescence microscope at 920×k (*) indicates late apoptotic cells and $ indicates necrotic/dead cells.
Following two-way ANOVA, there was significant interaction be-tween the effects of age and melatonin treatment on GM-CFU countF(2, 12)=4.587, pb0.05. Significant (pb0.01) decrease in numberof colonies of CFU-GM was noted in old hamster when comparedwith adult and young hamsters. Melatonin treatment significantly en-hanced the GM-CFU colonies in young (pb0.01), adult and old(pb0.05) hamster when compared with vehicle treated control of re-spective age group of hamsters (Fig. 11a and b). Further, the prolifer-ation of GM-CFU was negatively correlated (r=−0.917, pb0.05)with age and showed strongly positive correlation (r=0.966,pb0.01) with melatonin level in circulation.
3.8. Peripheral level of melatonin
There was significant interaction observed following two wayANOVA between the effects of age and melatonin treatment on plas-ma melatonin level F(2, 12)=6.706, pb0.05. Plasma melatonin
(night time; 22:00 h) level was significantly (pb0.01) low in oldage but no significant difference was noted in adult when comparedwith young hamster. Further, melatonin treatment significantly(pb0.01) increased the level of night time plasma melatonin whencompared with vehicle treated control of respective age group ofhamsters (Fig. 12).
4. Discussion
Aging might be a result of the failure or slow down of various pro-tective mechanisms that counteract the damages induced by ROS/RNS radicals. We therefore, explored the ROS/RNS generation in dif-ferent age group of hamsters following the treatment with melatonin,in order to note its effects as a counteracting compound for aging.Aging also has been associated with changes in cellular, humoraland innate immunity in many mammals (Ahmad et al., 2012; Raiand Haldar, 2003; Srinivasan et al., 2005) but its relationship with ox-idative stress has not been explored. We evaluated oxidative stress bymeasuring the levels of ROS/RNS in peripheral blood and lymphoid
Fig. 10. Effect of age and melatonin administration on blastogenic immune response (%SR) in (a) PBMC and (b) splenocytes of young (YH), adult (AH) and old (OH) hamsters.Values are expressed as mean±SD, N=7. **pb0.01 YH vs OH; apb0.01 and bpb0.05 vehicle vs melatonin treatment.
179D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
tissues from different age group of hamsters. Increased level ofnitrite/nitrate ion was recorded in plasma, spleen and BM of hamsterswith advancing age along with a low level of melatonin (Fig. 12). Thisdecrease in level of melatonin might be due to decrease in sensitivityof SCN as reported earlier (Garidou et al., 2002; Pang et al., 1990;Reiter et al., 1980; Selmaoui and Touitou, 1999) and hence loses its ef-ficiency to detoxify RNS (NO\ and ONOO ) in a concentration depen-dent manner (Acuna-Castroviejo et al., 2005; Reiter et al., 2001). Ourresults also suggested that circulatory melatonin level was inverselyproportion to the age dependent increase in ROS generation and theDNA damage as reflected by AO-EB staining, which gets supportfrom report of Lopez et al. (2010) that melatonin is an excellent anti-oxidant and protector counteracts the generation of ROS/RNS and im-proves cell function by maintaining the ATP production and cellsurvival.
We found that melatonin treatment played an important role ininducing the pre-dormant cells to enter in G1 phase of cell cycle,suggesting the role of melatonin in immune cell proliferation. Previ-ously, we showed that the melatonin induction augments prolifera-tion of lymphocytes by increasing the blastogenic response in termsof %SR (Ahmad and Haldar, 2010). In the present study we for thefirst time, presented the datum that melatonin induces cell prolifera-tion at G1 phase onwards and that melatonin treatment maintainedthe normal cell cycle at G1 phase in old hamsters. Aging has been as-sociated with increased oxidative damage, cell dysfunction, DNAdamage and cell death (Shi et al., 2009). Cell death may induce an in-flammatory reaction and induction of inducible nitric oxide synthase
(iNOS) which is responsible for the production of an excess of NO andperoxynitrite leading to irreversible damage to the mitochondrial re-spiratory complexes, ATP depletion, and increased DNA damage andthereby cell death (Agostini et al., 2002; Payne et al., 1995;Wiseman and Halliwell, 1996).
We also observed that ROS generation by PBMC (r=−0.863,pb0.01) and splenocytes (r=−0.769, pb0.01) were directly propor-tional with age of hamsters supporting the hypothesis that reducedlevel of melatonin with age was unable to counteract the elevatedlevels of ROS in aged hamsters. The major effects of increased ROS/RNS noted from bone marrow, spleen, PBMC as well from plasmasuggested a compromised state of immune function in aged hamsters.This negative influence of age factor on immunity was furthersupported by the reduced number of colony of BM progenitor cells(GM-CFU) and splenocytes proliferation rate (%SR) recorded in oldhamsters. Damage of cells and rate of apoptosis in old hamsters getssignificantly reduced following melatonin administration as judgedby AO-EB staining. Thus, we hypothesize that melatonin may improvethe general immunity by up-regulating immune functions and simul-taneously reducing oxidative load and DNA damages, which having apositive correlation with age.
Lipid peroxidation is one of the pathways which also get induced byROS/RNS and can damage the immune cells by causing cellular injurydue to cleavage of DNA, destruction of cell membrane lipids and inhibi-tion of critical enzymes through nitration in electron transport chain(Vincent and Maiese, 1999; Wang et al., 2003; Yamamoto et al.,2002). TBARS is typically produced when the membrane lipoproteins
Fig. 11. Effect of age and melatonin administration on proliferation of granulocyte macrophage colony forming unit (GM-CFU) from bone marrow. (a) Histogram representsmean±SD, N=7. **pb0.01 YH vs OH; apb0.01 and bpb0.05 vehicle vs melatonin treatment. (b) Photograph showing GM-CFU colony after 7 days of bone marrow mononuclearcell (BM-MNC) culture of golden hamster (×20).
and polyunsaturated fatty acids are attacked by ROS, another marker ofoxidative stress (Floyd, 1990), increased in plasma, spleen and BM ofaged hamsters presenting a positive correlation (plasma r=0.957,pb0.05; spleen r=0.857, p>0.05; and BM r=0.746, pb0.01) withage.
Antioxidant enzymes such as total SOD, catalase and GPx togetherplay an important role in the oxidative defense at all the levels(Mates, 2000). Transcriptional and translational up-regulation of theabove mentioned antioxidant enzymes by melatonin has alreadybeen reported (Esparza et al., 2005; Hung et al., 2008; Munoz-Casares et al., 2006). Therefore, we examined the antioxidant enzymeactivities to stress upon the oxidative injury if occurred in immunecells of old hamsters. We observed that SOD and catalase activities
Fig. 12. Effect of age and melatonin administration on plasma melatonin concentrationin golden hamster. Values are expressed as mean±SD, N=7.**pb0.01 YH vs OH;apb0.01 vehicle vs melatonin treatment.
were decreased with increased age, suggesting a negative correlationwith aging process as noted earlier by Andersen et al. (1997) andGlass and Gershon (1981) in rats.
Further, GPx being a key enzyme of antioxidative defense, its ac-tivity in cells of various organs can be stimulated by melatonin(Agapito et al., 2001; Barlow-Walden et al., 1995; Chen and Chuang,1999; Guletkin et al., 2001; Okatani et al., 2000, 2001; Pablos et al.,1995). We observed that melatonin administration to hamsters sig-nificantly elevated the activity of GPx in plasma, spleen and BM ofold aged hamsters. This increase in GPx activity might be due tohigher expression of GPx mRNA level following melatonin treatmentas noted by Jose et al. (2007) in aged rats than in young groups.Therefore, the age related decrease in the anti-oxidant enzyme SOD,catalase and GPx may be considered as a generalized consequenceof deterioration of the physiological functions with age (Fabian etal., 2012), andmelatoninmight have induced the activity of antioxidantenzymes in aged hamsters to alleviate the imbalance of antioxidant/pro-oxidant level.
Thus, low level of melatonin with progressing age of hamstersmay lead to reduction in the sensitivity of the lymphoid organ tothe melatonin. This is further aggravated by the increased ROS/RNSproduction resulting in oxidative stress of immune cells and thereby,compromised the immune function. In summary, the present studysuggests that apart from declining level of melatonin and increasingoxidative stress with age, there also exists a possible role of ROS me-diated toxicity for DNA damage, which may be improved by adminis-tration of melatonin acting as a potent free radical scavenger toprevent age associated immune disorders marked by the enhancedoxidative stress for better survival.
181D.K. Vishwas et al. / Experimental Gerontology 48 (2013) 168–182
Author contributions
All the listed authors were involved in: 1) the conception and de-sign, or analysis and interpretation of data; 2) drafting the article orrevising it critically for important intellectual content, and 3) final ap-proval of the version to be published.
Grants
This work was supported by grants from the Council of Scientificand Industrial Research, New Delhi, government of India (grantscheme number: 37/1462/11/EMR-II) for junior research fellowship(AM). Financial support as senior research fellowship from UniversityGrant Commission, New Delhi, India (DKV) is gratefully acknowledged.
Conflicts of interest
None.
Acknowledgement
The authors acknowledge the gift of instrument by Alexander vonHumboldt Foundation, Bonn, Germany and copper-cadmium alloy byDr. Jagmohan, Barely, India. Various technical helps by Mr. AmitKumar Srivastava is also acknowledged.
References
Acuna-Castroviejo, D., Escames, G., Lopez, L.C., Hitos, A.B., Leon, J., 2005. Melatonin andnitric oxide. Endocrine 27, 159–168.
Agapito, M.T., Antolin, Y., Del-Brio, M.T., Lopes-Burillo, S., Pablos, M.I., Recio, J.M., 2001.Protective effect of melatonin against adriamycin toxicity in the rat. J. Pineal Res.31, 23–30.
Agostini, M., Di Marco, B., Nocentini, G., Delfino, D.V., 2002. Oxidative stress andapoptosis in immune diseases. Int. J. Immunopathol. Pharmacol. 15 (3), 157–164.
Ahmad, R., Haldar, C., 2010. Melatonin and androgen receptor expression interplaymodulates cell mediated immunity in tropical rodent Funambulus pennanti.Scand. J. Immunol. 71, 420–430.
Ahmad, R., Gupta, S., Haldar, C., 2012. Age dependent expression of melatonin mem-brane receptor (mt1, mt2) and its role in regulation of nitrosative stress in tropicalrodent Funambulus pennanti. Free Radic. Res. 46 (2), 194–203.
Andersen, H.R., Nielsen, Y.B., Nielsen, F., Grandjean, P., 1997. Antiox-dismutidativeenzyme activities in human erythrocytes. Clin. Chem. 43 (4), 562–568.
Barlow-Walden, L.R., Reiter, R.J., Abe, M., Pablos, M., Menendez-Pelaez, A., Chen, L.D.,Poeggeler, B., 1995. Melatonin stimulates brain glutathione peroxidase activity.Neurochem. Int. 26, 497–502.
Beal, M.F., 2002. Oxidatively modified proteins in aging and disease. Free Radic. Biol.Med. 32, 797–803.
Bian, K., Murad, F., 2001. Diversity of endotoxin-induced nitrotyrosine formation inmacrophageendothelium-rich organs. Free Radic. Biol. Med. 31, 421–429.
Bienzle, D., Reggeti, F., Clark, M., Chow, C., 2003. Immunophenotype and functionalproperties of feline dendritic cells derived from blood and bone marrow. Vet.Immunol. Immunopathol. 96, 19–30.
Boyum, A., 1968. Isolation of mononuclear cells and granulocytes from human blood:isolation of mononuclear cells by one centrifugation, and of granulocytes by com-bining centrifugation and sedimentation at 1 g. Scand. J. Clin. Lab. Invest. Suppl. 97,77–89.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.72, 248–254.
Bruning, J.L., Knitz, B.L., 1977. Computational handbook of statistics. (Eds) Scott,Foresman and Company, Illinois, USA.
Chen, S.T., Chuang, J.I., 1999. The antioxidant melatonin reduces cortical neuronal deathafter intrastriatal injection of kainite in the rat. Exp. Brain Res. 124, 241–247.
Das, K., Samanta, L., Chainy, N.B.G., 1999. A modified spectrophotometric assay of su-peroxide dismutase using nitrite formation by superoxide radicals. J. Biochem.Biophysiol. 37, 201–204.
Degli Esposti, M., McLennan, H., 1998. Mitochondria and cells produce reactive oxygenspecies in virtual anaerobiosis: relevance to ceramide-induced apoptosis. FEBSLett. 430 (3), 338–342.
Di Fiore, M.M., Lamanna, C., Assisi, L., Botte, V., 2008. Opposing effects of d-aspartic acidand nitric oxide on tuning of testosterone production in mallard testis during thereproductive cycle. Reprod. Biol. Endocrinol. 6, 28.
Drew, B., Leeuwenburgh, C., 2002. Aging and the role of reactive nitrogen species. Ann.N. Y. Acad. Sci. 959, 66–81.
Esparza, J.L., Gomez, M., Rosa Nogues, M., 2005. Melatonin reduces oxidative stress andincreases gene expression in the cerebral cortex and cerebellum of aluminum-exposed rats. J. Pineal Res. 39, 129–136.
Eu, J.P., Liu, L., Zeng, M., Stamler, J.S., 2000. An apoptotic model for nitrosative stress.Biochemistry 39, 1040–1047.
Fabian, E., Bogner, M., Elmadfa, I., 2012. Age-related modification of antioxidant en-zyme activities in relation to cardiovascular risk factors. Eur. J. Clin. Invest. 42(1), 42–48.
Floyd, R.A., 1990. Role of oxygen free radicals in carcinogenesis and brain ischemia.FASEB J. 4, 2587–2597.
Garidou, M.L., Ribelayga, C., Pevet, P., Simonneaux, V., 2002. Golden hamster and ratdisplay developmental differences in the regulation of pineal arylalkylamineN-acetyltransferase. J. Neuroendocrinol. 14 (11), 861–868.
Glass, G.A., Gershon, D., 1981. Enzymatic changes in rat erythrocytes with increasingcell and donor age: loss superoxide dismutase activity associated with increasesin catalytically defective forms. Biochem. Biophys. Res. Commun. 103, 1245–1249.
Guletkin, F., Delibas, N., Yasar, S., Kilinc, I., 2001. In vivo changes in antioxidant systemsand protective role of melatonin and a combination of vitamin C and vitamin E onoxidative damage in erythrocytes induced by chlorpyrifos-ethyl in rats. Arch.Toxicol. 75, 88–96.
Haldar, C., Haussler, D., Gupta, D., 1992. Effect of pineal gland on circadian rhythmicityof colony forming units for granulocytes and macrophages (CFU-GM) from ratbone marrow culture. J. Pineal Res. 12, 79–83.
Harman, D., 1956. Aging: a theory based on free radical and radiation chemistry.J. Gerontol. 11, 298–300.
Hung, M.W., George, L., Reiter, R.J., Fung, M.L., 2008. Protective effect of melatoninagainst hippocampal injury of rats with intermittent hypoxia. J. Pineal Res. 44,214–221.
Jakubowski, W., Bartosz, G., 2000. 2,7-Dichlorofluorescin oxidation and reactiveoxygen species: what does it measure? Cell Biol. Int. 24 (10), 757–760.
Jose, L.M., Virginia, M., Maria, V., Garcia, M., Paquita, G., Juan, P.B., Javier, G.G., 2007. Mel-atonin prevents oxidative stress and changes in antioxidant enzyme expression andactivity in the liver of aging rats. J. Pineal Res. 42, 222–230.
Levrand, S., Pesse, B., Feihl, F., Waeber, B., Pacher, P., Rolli, J., Schaller, M.D., Liaudet, L.,2005. Peroxynitrite is a potent inhibitor of NF-kappa B activation triggered by in-flammatory stimuli in cardiac and endothelial cell lines. J. Biol. Chem. 280,34878–34887.
Liu, S.J., Wang, Z.J., 2002. Alzheimer-like tau phosphorylation induced by wortmanninin vivo and its attenuation by melatonin. Acta Pharmacol. Sin. 23, 183–187.
Lopez, L.C., Escames, G., Lopez, A., Garcia, J.A., Doerrier, C., Acuña-Castroviejo, D., 2010.Melatonin, neurogenesis, and aging brain. Open Neuroendocrinol. J. 3, 121–133.
Mantha, S.V., Prasad, M., Kalra, J., Prasad, K., 1993. Anti oxidant enzymes inhypercholesteromia and effect of vitamin E in rabbits. Atherosclerosis 101,135–144.
Marshall, H.E., Merchant, K., Stamler, J.S., 2000. Nitrosation and oxidation in the regu-lation of gene expression. FASEB J. 14, 1889–1900.
Mates, J.M., 2000. Effects of antioxidant enzymes in the molecular control of reactiveoxygen species toxicology. Toxicology 153, 83–104.
Miller, R.A., 1991. Aging and immune function. Int. Rev. Cytol. 124, 187–215.Miller, R.A., 1995. Cellular and biochemical changes in the aging mouse immune
system. Nutr. Rev. 53 (Suppl. 2), 8–17.Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: applica-
tion to proliferation and cytotoxicity assays. J. Immunol. Methods 65, 55–63.Munoz-Casares, F.C., Padillo, F.J., Briceno, J., 2006. Melatonin reduces apoptosis and ne-
crosis induced by ischemia/ reperfusion injury of the pancreas. J. Pineal Res. 40,195–203.
Nelson, R.J., 2004. Seasonal immune function and sickness responses. Trends Immunol.25, 187–192.
Ohkawa, H., Ohishi, N., Yagi, K., 1978. Reaction of linoleic acid hydroperoxide withthiobarbuteric acid. J. Lipid Res. 19, 1053–1057.
Okatani, Y., Wakatsuki, A., Kaneda, C., 2000. Melatonin increases activities ofglutathione peroxidase and superoxide dismutase in fetal rat brain. J. Pineal Res.28, 89–96.
Okatani, Y., Wakatsuki, A., Ahinohara, K., 2001. Melatonin stimulates glutathioneperoxidase activity in human chorion. J. Pineal Res. 30, 199–205.
Pablos, M.I., Agapito, M.T., Gutierrez, R., Recio, J.M., Reiter, R.J., Barlow-Walden, L.R.,Acuña-Castroviejo, D., Menendez-Pelaez, A., 1995. Melatonin stimulates the activ-ity of the detoxifying enzyme glutathione peroxidase in several tissues of chickens.J. Pineal Res. 19, 111–115.
Pacher, P., Szabo, C., 2008. Role of the peroxynitrite-poly(ADP-ribose) polymerasepathway in human disease. Am. J. Pathol. 173, 2–13.
Pang, S.F., Tsang, C.W., Hong, G.X., Yip, P.C., Tang, P.L., Brown, G.M., 1990. Fluctuation ofblood melatonin concentrations with age: result of changes in pineal melatoninsecretion, body growth, and aging. J. Pineal Res. 8 (2), 179–192.
Payne, C.M., Bernstein, C., Bernstein, H., 1995. Apoptosis overview emphasizing the role ofoxidative stress, DNA damage and signal-transduction pathways. Leuk. Lymphoma19 (1–2), 43–93.
Pierpaoli, W., Regelson, W., 1992. Pineal control of aging: effect of melatonin and pinealgrafting on aging mice. Proc. Natl. Acad. Sci. 91, 787–791.
Rai, S., Haldar, C., 2003. Pineal control of immune status and hematological changes inblood and bone marrow of male squirrels (Funambulus pennanti) during theirreproductively active phase. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 136,319–328.
Rastogi, R.P., 2010. Detection of reactive oxygen species (ROS) by the oxidant-sensingprobe 2′,7′-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaenavariabilis PCC 7937. Biochem. Biophys. Res. Commun. 397, 603–607.
Reiter, R.J., Craft, C.M., Johnson, J.E., King Jr., T.S., Richardson, B.A., Vaughan, G.M.,Vaughan, M.K., 1981. Age-associated reduction in nocturnal pineal melatoninlevels in female rats. Endocrinology 109, 1295–1297.
Reiter, R.J., Calvo, J.R., Karbownik, M., Qi, W., Tan, D.X., 2000. Melatonin and its relationto immune system and inflammation. Ann. N. Y. Acad. Sci. 917, 376–386.
Reiter, R.J., Tan, D.S., Manchester, L.C., Qi, W., 2001. Biochemical reactivity of melatoninwith reactive oxygen and nitrogen species. Cell Biochem. Biophys. 34, 237–256.
Rodriguez, C., Mayo, J.C., Sainz, R.M., Antolin, I., Herrera, F., Martin, V., Reiter, R.J., 2004. Reg-ulation of antioxidant enzymes: a significant role for melatonin. J. Pineal Res. 36, 1–9.
Sastry, K.V.H., Moudgal, R.P., Mohan, J., Tyagi, J.S., Rao, G.S., 2002. Spectrophotometricdetermination of serum nitrite and nitrate by copper–cadmium alloy. Anal.Biochem. 306, 79–82.
Selmaoui, B., Touitou, Y., 1999. Age-related differences in serum melatonin and pinealNAT activity and in the response of rat pineal to a 50-Hz magnetic field. Life Sci. 64(24), 2291–2297.
Sharma, S., Haldar, C., 2009. Comparative effect of melatonin and vitamin E onphenylhydrazine-induced toxicity in the spleen of Funambulus pennanti. Environ.Toxicol. 24 (1), 1–9.
Shi, Y., Song, Y., Wang, Y., Liang, X., Hu, Y., Guan, X., 2009. p, p-DDE induces apoptosis ofrat sertoli cells via a FasL-dependent pathway. J. Biomed. Biotechnol. 181282, 1–11.
Miller, S.C., 2005. Melatonin, immune function and aging. Immun. Aging 2, 17.Srivastava, A.K., Ali, W., Singh, R., Bhui, K., Tyagi, S., Al-Khedhairy, A.A., Srivastava, P.K.,
Musarrat, J., Shukla, Y., 2012. Mancozeb-induced genotoxicity and apoptosis incultured human lymphocytes. Life Sci. 90 (21–22), 815–824.
Stamler, J.S., Hausladen, A., 1998. Oxidative modifications in nitrosative stress. Nat.Struct. Biol. 5, 247–249.
Vincent, A.M., Maiese, K., 1999. Nitric oxide induction of neuronal endonucleaseactivity in programmed cell death. Exp. Cell Res. 246, 290–300.
Wang, J.Y., Shum, A.Y., Ho, Y.J., 2003. Oxidative neurotoxicity in rat cerebral cortexneurons: synergistic effects of H2O2 and NO on apoptosis involving activationof p38 mitogen-activated protein kinase and caspase-3. J. Neurosci. Res. 72,508–519.
Wiseman, H., Halliwell, B., 1996. Damage to DNA by reactive oxygen and nitrogen spe-cies: role in inflammatory disease and progression to cancer. Biochem. J. 313,17–29.
Yamamoto, T., Maruyama, W., Kato, Y., 2002. Selective nitration of mitochondrial com-plex I by peroxynitrite: involvement in mitochondria dysfunction and cell death ofdopaminergic SH-SY5Y cells. J. Neural. Transm. 109, 1–13.























![Hamster[1] (3)rt](https://static.documents.pub/doc/80x56/5491fa67b4795943628b46a3/hamster1-3rt.jpg)







