Page 1
Subscriber access provided by Penn State | University Libraries
Journal of Proteome Research is published by the American Chemical Society. 1155Sixteenth Street N.W., Washington, DC 20036Published by American Chemical Society. Copyright © American Chemical Society.However, no copyright claim is made to original U.S. Government works, or worksproduced by employees of any Commonwealth realm Crown government in the courseof their duties.
Article
In Pursuit of Protein Targets: Proteomiccharacterization of Bacterial Spore Outer Layers.
Wishwas Abhyankar, Abeer H. Hossain, André Djajasaputra, Patima Permpoonpattana, AlexanderS. Ter Beek, Henk L. Dekker, Simon Cutting, Stanley Brul, Leo J de Koning, and Chris G de Koster
J. Proteome Res., Just Accepted Manuscript • DOI: 10.1021/pr4005629 • Publication Date (Web): 02 Sep 2013
Downloaded from http://pubs.acs.org on September 9, 2013
Just Accepted
“Just Accepted” manuscripts have been peer-reviewed and accepted for publication. They are postedonline prior to technical editing, formatting for publication and author proofing. The American ChemicalSociety provides “Just Accepted” as a free service to the research community to expedite thedissemination of scientific material as soon as possible after acceptance. “Just Accepted” manuscriptsappear in full in PDF format accompanied by an HTML abstract. “Just Accepted” manuscripts have beenfully peer reviewed, but should not be considered the official version of record. They are accessible to allreaders and citable by the Digital Object Identifier (DOI®). “Just Accepted” is an optional service offeredto authors. Therefore, the “Just Accepted” Web site may not include all articles that will be publishedin the journal. After a manuscript is technically edited and formatted, it will be removed from the “JustAccepted” Web site and published as an ASAP article. Note that technical editing may introduce minorchanges to the manuscript text and/or graphics which could affect content, and all legal disclaimersand ethical guidelines that apply to the journal pertain. ACS cannot be held responsible for errorsor consequences arising from the use of information contained in these “Just Accepted” manuscripts.
Page 2
1
In Pursuit of Protein Targets: Proteomic characterization of
Bacterial Spore Outer Layers.
Wishwas Abhyankar†,×, Abeer H. Hossain†,×, André Djajasaputra†,×, Patima Permpoonpattana‡,¤,
Alexander Ter Beek×, Henk L. Dekker†, Simon M. Cutting‡, Stanley Brul×, Leo J. de Koning†,
Chris G. de Koster*,†
†Mass Spectrometry of Bio-macromolecules and ×Molecular Biology & Microbial Food safety
Swammerdam Institute for Life Sciences, University of Amsterdam, The Netherlands;
‡School of Biological Sciences, Royal Holloway, University of London, UK.
¤ Current address : Faculty of Science and Industrial Technology, Prince of Songkla University, Surat
Thani Campus, Surat Thani 84000, Thailand.
Abstract
Bacillus cereus, responsible for food poisoning and Clostridium difficile, causative agent
of Clostridium difficile-associated diarrhoea (CDAD) are both spore forming pathogens involved
in food spoilage, food intoxication and other infections in humans and animals. The
proteinaceous coat and the exosporium layers from spores are important for their resistance and
pathogenicity characteristics. The exosporium additionally provides an ability to adhere to
surfaces eventually leading to spore survival in food. Thus studying these layers and identifying
suitable protein targets for rapid detection and removal of spores is of utmost importance. In this
study, we identified 100 proteins from B. cereus spore coat, exosporium and 54 proteins from the
C. difficile coat insoluble protein fraction. In an attempt to define a universal set of spore outer
layer proteins we identified 11 superfamily domains common to the identified proteins from two
Bacilli and a Clostridium species. The evaluated orthologue relationships of identified proteins
Page 1 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 3
2
across different spore formers resulted in a set of 13 coat proteins conserved across the spore
formers and 12 exosporium proteins conserved in the B. cereus group which could be tested for
quick and easy detection or targeted in strategies aimed at removal of spores from surfaces.
Keywords: B. cereus ATCC 14579, C. difficile 630, spore coat, exosporium, insoluble protein
fraction
Introduction
Studies with Bacillus subtilis have provided good insights in the sporulation and
germination processes of spores. However, studies of more pathogenic species like aerobic or
facultatively anaerobic Bacillus cereus and strictly anaerobic Clostridium difficile are needed to
gain more insights in the spore structure and germination characteristics of these spores. The B.
cereus group (also known as B. cereus sensu lato), is comprised of several closely related
species: Bacillus mycoides, Bacillus pseudomycoides, Bacillus weihenstephanensis, Bacillus
anthracis, Bacillus thuringiensis and Bacillus cereus1. B. cereus is mainly known as a food-
borne pathogen causing two types of gastrointestinal diseases: emesis and diarrhea2. B. cereus
related emesis is caused by the cereulide toxin, which once produced remains stable upon
enzyme, heat or acid treatment. Diarrhea, on the other hand, is caused by enterotoxins
haemolysin BL (Hbl), non-haemolytic enterotoxin (Nhe) and cytotoxin K (CytK)2, 3 that are
produced by the vegetative cells. B. cereus strains have also been reported in few cases to be
associated with non-gastrointestinal diseases including: respiratory tract infections,
endophthalmitis, central nervous system infections, gas gangrene-like infections, cutaneous
infections, endocarditis, osteomyelitis and urinary tract infections4. Spore forming Clostridium
spp. (e.g. Clostridium botulinum, Clostridium tetani), like Bacilli, are also known for the
Page 2 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 4
3
infections and diseases caused by them. Pathogenic C. difficile can colonize the intestinal tracts
of humans and other mammals5, 6. Routine treatments containing use of antibiotics such as
Metronidazole, Vancomycine have led to emergence of resistant strains7, 8 and thus prolonged
treatment with such antibiotics can result in C. difficile overgrowth and can lead to diseases
ranging from diarrhea to life-threatening pseudomembranous colitis, especially in
immunocompromised people9, 10. C. difficile is reported to be a continuously evolving species11,
12 and in recent times the organism has emerged as one of the major causes of nosocomial
diarrhea (Clostridium difficile-associated diarrhea; CDAD). CDAD is mainly caused by the
secretion of two cytotoxic, enterotoxic and proinflammatory toxins known as toxin A (TcdA)
and toxin B (TcdB)13. CDAD is particularly problematic to treat and to avert because of the
robust endospores that can persist and be easily transferred, person-to-person, in a hospital
environment and thus the morbidity and mortality rates have been increasing in recent years.
Endospores, are metabolically dormant multi-layered cellular forms which upon
germination, lose their protective external layers and resume vegetative growth14. In B. subtilis
the outermost thick concentric proteinaceous layers - the inner and outer spore coat15 - aid in
resistance against variety of environmental assaults. Additionally in spores of many Bacillus and
Clostridium species surrounding the coats an external loosely-fitting, hydrophobic, glycosylated,
balloon-like layer - the exosporium - is present, which assists in adherence to the surfaces16.
Though the cortex layer from spores might have evolved from the cell wall homologous to those
of vegetative cells, the proteinaceous coat is quite unique. The immense resistance of the coat to
attack by microorganisms and by free enzymes is unmatched in the bacterial domain. The wide
variety of germination signals amongst the closely related species suggests that germination
receptor assembly is a relatively later event in the evolution of the spore structure as the ancestral
Page 3 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 5
4
Bacilli and Clostridia moved to different niches after their divergence about 2.3 billion years
ago17. Albeit the evolutionary distance, the ability to form spores and the overall sporulation
process are conserved in these organisms with significant differences still existing both in the
regulation of spore formation and in spore structure18. Thus it is imperative to study the spore
coat and exosporium layers from the spores of these two organisms in order to obtain detailed
knowledge about the spore structure as well as to design quick and simple techniques for
detection and thereby facilitating eradication of spores form the environment.
Previous extensive research has led to the identification of up to 70 different proteins
from the spore coat layers in B. subtilis. The exosporium is a relatively less studied layer,
possibly due to the difficulties in obtaining its large quantities, and is reported to comprise of at
least 25 proteins as studied from B. cereus and B. anthracis spores 15, 19-21. Mostly these studies
have focused on the soluble fraction of proteins. Our “gel-free” method22 allowed us for the first
time to focus on the insoluble protein fraction from the spore coat, which makes up to 30% of the
proteins15, and is characterized by extensive inter-protein cross-linking and thus is difficult to
analyze using conventional PAGE gels. We have showed that our gel-free method is
comprehensive in isolating and identifying proteins from the insoluble protein fraction of B.
subtilis spore coats with identification of 19 new proteins. Here, we have extended our method to
B. cereus and C. difficile to identify protein targets for early and rapid detection of spores and
have characterized for the first time the insoluble proteome of the coat and exosporium of B.
cereus ATCC 14579 spores and of the coat layers of C. difficile 630 spores. Recently, McKenney
et al.23 mentioned that the coat morphogenetic proteins may be the targets for evolutionary
adaptation for spore formers. Therefore to obtain a universal set of spore coat and exosporium
Page 4 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 6
5
proteins we also assess different spore formers for the conservation of the spore surface proteins
that were identified from B. subtilis22, B. cereus and C. difficile in our studies.
Materials and Methods
Media and bacterial strains used The strains used in this study were B. cereus ATCC 14579
and C. difficile 630 (tcdA+ tcdB
+). For B. cereus ATCC 14579, growth and sporulation were
carried out in Tryptic Soy Broth (TSB) medium as well as a chemically defined growth and
sporulation (CDGS) medium, as described previously24. The CDGS medium consisted of the
following components (final concentrations): 10 mM D-glucose, 20 mM L-glutamic acid, 6 mM
leucine, 2.6 mM L-valine, 1.4 mM L-threonine, 0.47 mM L-methionine, 0.32 mM L-histidine, 5
mM D/L-lactic acid, 1 mM acetic acid, 50µM FeCl3, 2.5 µM CuCl2, 12.5 mM ZnCl2, 66 µM
MnSO4, 1 mM MgCl2, 5 mM (NH4)2SO4, 2.5 µM Na2MoO4, 2.5 µM CoCl2, 1 mM Ca(NO3)2,
buffered to pH 7.2 using 100 mM potassium phosphate buffer. C. difficile 630 was routinely
grown in vegetative stage on Brain-Heart Infusion Supplemented (BHIS) agar plates and in
Tryptone Glucose Yeast extract (TGY)-vegetative medium (3% TSB, 2% glucose, 1% yeast
extract, 0.1% L-cysteine). Pre-culturing was done in SMC broth (90 g peptone, 5 g proteose
peptone, 1 g (NH4)2SO4, 1.5 g Tris) containing 0.1% L-cysteine, while sporulation was initiated
on SMC agar plates25.
Conditions for growth and sporulation For B. cereus ATCC 14579 and C. difficile 630
respectively four and three independent biological spore crops were made. A B. cereus culture
was grown overnight in 50 mL TSB medium (pH 7.5) at 30⁰C in a shaker-incubator at 200 rpm.
Cells were harvested by centrifugation at 10000 rpm for 30 min at 4⁰C, resuspended in 500 mL
CDGS medium, and incubated at 30⁰C for 96 h at 200 rpm. After 4 days, the spores were
Page 5 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 7
6
harvested by centrifugation. To lyse the remaining vegetative cells, the spore pellet was treated
twice with 0.1% (final concentration) Tween-20 and subsequently washed five times with sterile
Milli-Q water. From each B. cereus spore crop, three sample fractions were prepared. The first
fraction contained spores with intact exosporium and coat (Fraction 1), the second fraction
contained spores with exosporium removed (spore coat sample; Fraction 2) and the third fraction
contained the concentrated exosporium fragments (Fraction 3).
For C. difficile, a colony from a BHIS agar plate was inoculated into 10 ml TGY medium
and incubated at 37°C overnight. This TGY culture was then subcultured into SMC broth and
incubated until the cells achieved the logarithmic phase, and then plated onto SMC agar plates.
The plates were further incubated for 7 days at 37°C. After 7 days, the spores were harvested by
washing twice in water and followed by suspension in phosphate-buffered saline (PBS)
containing 125 mM Tris, 200 mM EDTA, 0.3 mg/ml proteinase K (Fermentas), and 1% sarcosyl
with further incubation and gentle shaking at 37°C for 2 h. Finally, spores were centrifuged; the
pellets were resuspended in water, and washed a further 10 times25. Only the coat fraction was
analyzed from C. difficile in this study.
Removal of exosporium To separate the exosporium layer from B. cereus spores, spores (1010
spores / mL) were resuspended in 50 mM Tris-HCl, 0.5 mM Na-EDTA (pH 7.5) and passaged 4
times through a French pressure cell (Thermo Fisher Scientific, MA, USA) at 20000 psi. Spores
were separated from exosporium fragments by centrifugation at 10000 rpm for 15 min at 4⁰C.
The supernatant (containing loose exosporium fragments) was centrifuged one more time, at
10000 rpm for 15 min at 4oC and pressed through a 0.22 µm PES filter (VWR International, PA,
USA) to remove any remaining spores and stored at 4oC. The pelleted spore fraction was
Page 6 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 8
7
resuspended in sterile Milli-Q water and stored at 4oC until further use. The protocol was adapted
from the work of Todd and co-workers26.
Isolation of spore coat The spore fraction of B. cereus ATCC 14579 (obtained after French
press treatment) and C. difficile 630 was centrifuged and the spore pellets were resuspended in
10 mM Tris-HCl (pH 7.5). The spores were disintegrated by bead-beating with 0.1 mm Zirconia-
Silica beads (BioSpec Products, USA) using a Precellys 24 homogenizer (Bertin Technologies,
Aix en Provence, France). Nine rounds of bead-beating were performed, with each round
consisting of a 40 s cycle at 6000 rpm and a 1 min interval between each round for cooling. After
every three rounds of beating, 10 min cooling periods (on ice) were allowed to prevent protein
degradation by overheating. The disintegrated spore pellets were then washed 5-6 times with 1
M NaCl to remove non-covalently bound proteins and intracellular contaminants. Spore coat
protein extraction was done by thermal heating for 10 min using a water bath, starting at ambient
temperature and reaching a final temperature of 80⁰C in SDS extraction buffer consisting of (in
final concentrations): 50 mM Tris-HCl (pH 7.8), 100 mM Na-EDTA, 150 mM NaCl, 0.2% SDS
and 100 mM β-mercaptoethanol. After protein extraction, the remaining SDS insoluble spore
coat protein pellets were washed four times with sterile Milli-Q water to remove SDS. The
pellets were then frozen in liquid nitrogen and freeze-dried overnight and stored at -80⁰C.
Concentration and analysis of exosporium For B. cereus spores, the suspension (obtained after
French press) containing the exosporium fragments, was concentrated by ultracentrifugation at
40000 rpm for 1 h at 4⁰C. The pellet containing exosporium fragments was then washed in 1 M
NaCl and subsequently twice in TEP buffer, consisting of 50 mM Tris-HCl (pH 7.5), 10 mM Na-
EDTA, 0.2% SDS and 2 mM β-mercaptoethanol. After washing in TEP buffer, the pellet was
washed once in TEP buffer without SDS. Exosporium protein extraction was done as mentioned
Page 7 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 9
8
above for spore coat protein extraction. After protein extraction, the remaining SDS insoluble
exosporium pellet was washed four times with sterile Milli-Q water to remove residual SDS. The
obtained pellet was then frozen in liquid nitrogen, freeze-dried overnight and stored at -80 ⁰C.
Though C. difficile spores have exosporium, its presence in our sample was variable. This is
likely because of its loss during spore harvest. Therefore we did not proceed to isolate
exosporium from C. difficile spores. The method was adapted from the work of Redmond et al27.
Analysis of spore cortex peptidoglycan The muramic acid from the spore cortex peptidoglycan
is an efficient marker to estimate the purity (i.e. the absence of cortex peptidoglycan) of the spore
coat sample. Therefore muramic acid assays were performed where the muramic acid is
estimated based on the absorbance measurements as described previously28, 29. To eliminate any
possible interference in the assay from the exosporial sugars (indicating inefficient exosporium
removal as reported previously by Thompson & co-workers30) deglycosylation of the sample
pellets was done by β-elimination according to the manufacturer’s instructions (GlycoProfile™
IV Chemical Deglycosylation Kit, Sigma-Aldrich Co. LLC). Lysozyme is known to act on the
1,4-β-linkage between N-acetylmuramic acid and N-acetyl-D-glucosamine from the vegetative
bacterial cell wall thus we also subjected the pellets to lysozyme (Sigma-Aldrich Chemie B.V.;
final concentration 250 µg/ml) treatment31 to remove the remaining muramic acid residues, if
any. Intact spores, disintegrated spores before and after NaCl wash, and the final reduced and
alkylated spore extracts were used for the assay.
Preparation for MS analysis Preparation of samples from spore coat & exosporium fraction,
spore coat fraction and exosporium fraction was done separately in the following manner. For
reduction of disulfide bridges, the freeze-dried samples were incubated with 10 mM
dithiothreitol (DTT) in 100 mM NH4HCO3 for 1 h at 55⁰C. The reducing reaction was followed
Page 8 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 10
9
by an alkylation treatment with 55 mM iodacetamide (IAA) in 100 mM NH4HCO3 for 45 min. at
room temperature in the dark. The samples were immediately digested for 18 h at 37⁰C with
trypsin (Trypsin Sequencing Grade Promega, Madison, WI, USA) using 1:60 w/w protease:
protein ratio. The tryptic digests were desalted using Omix µC18 pipette tips (80 µg capacity,
Varian, Palo Alto, CA, USA) according to the manufacturer’s instructions and the peptides were
collected in 25µL 50% acetonitrile (ACN), 0.1 % trifluoroacetic acid (TFA) and stored at -80⁰C.
Before analysis a fraction of eluted peptide material was freeze-dried and concentrated in 10 µL
of 0.1% TFA and peptide concentration was measured at 205 nm32 with a Nanodrop ND1000
spectrophotometer (Isogen Life Sciences, De Meern, The Netherlands).
LC-FT-ICR MS/MS analysis LC-MS/MS data were acquired with an Bruker ApexUltra
Fourier transform ion cyclotron resonance mass spectrometer (Bruker Daltonics, Bremen,
Germany) equipped with a 7 T magnet and a nano-electrospray Apollo II DualSource™ coupled
to an Ultimate 3000 (Dionex, Sunnyvale, CA, USA) HPLC system. Samples containing up to 60
ng of the tryptic peptide mixtures were injected as a 10 µl 0.1% TFA, 3% ACN aqueous solution
and loaded onto a PepMap100 C18 (5-µm particle size, 100-Å pore size, 300-µm inner diameter
x 5 mm length) precolumn. Following injection, the peptides were eluted via an Acclaim
PepMap 100 C18 (3-µm particle size, 100-Å pore size, 75-µm inner diameter x 250 mm length)
analytical column (Thermo Scientific, Etten-Leur, The Netherlands) to the nano-electrospray
source. Gradient profiles of up to 120 minutes were used from 0.1% formic acid / 3% CH3CN /
97% H2O to 0.1% formic acid / 50% CH3CN / 50% H2O at a flow rate of 300 nL/min. Data
dependent Q-selected peptide ions were fragmented in the hexapole collision cell at an Argon
pressure of 6x10-6 mbar (measured at the ion gauge) and the fragment ions were detected in the
ICR cell at a resolution of up to 60000. In the MS/MS duty cycle, 3 different precursor peptide
Page 9 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 11
10
ions were selected from each survey MS. The MS/MS duty cycle time for 1 survey MS and 3
MS/MS acquisitions was about 2 s. Instrument mass calibration was better than 1 ppm over a
m/z range of 250 to 1500.
Raw FT-MS/MS data were processed with the MASCOT DISTILLER program, version
2.4.3.1 (64bits), MDRO 2.4.3.0 (MATRIX science, London, UK), including the Search toolbox
and the Quantification toolbox. Peak-picking for both MS and MS/MS spectra were optimized
for the mass resolution of up to 60000. Peaks were fitted to a simulated isotope distribution with
a correlation threshold of 0.7, with minimum signal to noise of 2. The processed data, combined
from the three independent biological replicates, were searched in a MudPIT approach with the
MASCOT server program 2.3.02 (MATRIX science, London, UK) against a complete B. cereus
ATCC 14579 and C. difficile 630 ORF translation database (Uniprot 2011 update, downloaded
from http://www.uniprot.org/uniprot), respectively. The databases were complemented with their
corresponding decoy data bases for statistical analyses of peptide false discovery rate (FDR).
Trypsin was used as enzyme and 2 missed cleavages were allowed. Carbamidomethylation of
cysteine was used as a fixed modification and oxidation of methionine as a variable
modification. The peptide mass tolerance was set to 30 ppm and the peptide fragment mass
tolerance was set to 0.03 Dalton. If needed the search was repeated with the same parameters but
with semitrypsin as the enzyme. Error-tolerant MASCOT search was also done to identify
possible modification in the peptides. In addition, MS/MS data were matched, allowing semi-
tryptic peptides. MASCOT MudPIT peptide identification score was set to a cut-off of 20. At
this cut-off and based on the number of assigned decoy peptide sequences, a peptide false
discovery rate (FDR) of ~2% for all analyses was obtained. The identified coat and/or
Page 10 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 12
11
exosporium proteins of Bacillus cereus ATCC 14579 and Clostridium difficile 630 are listed in
Table 1 and 2 respectively. The identified cytosolic proteins are listed in Supplementary Table 6.
Tertiary structure prediction and Sequence alignment For tertiary structure prediction Phyre2
tool (www.sbg.bio.ic.ac.uk/phyre2/) was used in the default mode. Sequence alignment was done
using the Clustal Omega tool (http://www.ebi.ac.uk/Tools/msa/clustalo/) with the default
parameters.
Conserved Domain Search The annotation of protein sequences for the identification of
domains is important in the protein sequence analysis. The identification of a conserved domain
may aid in understanding the cellular and molecular function of a protein, as well as the
evolutionary history of proteins. Identification of conserved domains from the identified proteins
was performed using the automatic mode of the Conserved Domain (CD)-search tool from NCBI
database (http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi). The default parameters
were used.
Orthologue identification and PSI-BLAST analysis For orthologue identification, PSI-BLAST
(Position-Specific Iterated Basic Local Alignment Search Tool) option from BLASTP tool
(http://blast.ncbi.nlm.nih.gov/Blast.cgi) version 2.2.27+ was used. The thresholds for orthologue
identification: maximum identity ≥30%, E-value ≤0.005. Only single iteration for PSI-BLAST
was performed. Proteins were said to be conserved if orthologues were found in at least 80% of
the strains selected for comparison.
Results and Discussion
Spore preparation The harvested B. cereus ATCC 14579 spore crop contained >95% purified
spores and for the C. difficile strain 630 the sporulation efficiency was noted to be ~75%. In
Page 11 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 13
12
contrast to the B. cereus spores, the presence of exosporium in harvested C. difficile spores was
inconsistent and if present only remnants of the exosporium were seen under the phase contrast
microscope, as observed previously25.
Spore peptidoglycan analysis Protein isolation and extensive washing of the pellets with salt
reduced the amount of cortex muramic acid below1% compared to the intact spores. However
the final reduced, alkylated pellet unexpectedly showed a slight increase in the amount of
muramic acid. This could be due to the release of sugar moieties post-digestion. Lysozyme
treatment did not further lower the muramic acid content of the pellet material. Deglycosylation
of the reduced and alkylated pellet did significantly reduce the muramic acid content to values
below 0.05% (results not shown). In conclusion, the final pellets used for proteomic analysis
hardly contained any cortex muramic acid contamination.
Conserved Domain Search Amongst all the identified proteins from both species, 148 different
superfamily domains including 22 distinct Domains of Unknown Function (DUFs) were
identified by a domain searching tool from NCBI (see Supplementary Table S1). The hydrolases
and peptidases identified from B. cereus ATCC 14579 and C. difficile 630 in this study might be
important for spore germination and spore structure. Amongst the proteins identified from C.
difficile, a preference towards proteins with catalase-domains, ferritin-like domains and metal-
binding domains that might have a role in resistance towards oxidative stress (see below), was
observed. There were 11 superfamily domains that emerged as common to the identified proteins
from B. subtilis 16822, B. cereus ATCC 14579 and C. difficile 630 (Figure 1).
Protein identification and Orthologue evaluation i.e. PSI-BLAST analysis Orthologues are
genes (in different species) evolved from a common ancestral gene by speciation. This suggests
that orthologues retain the same function in the course of evolution. Evolutionary pressure
Page 12 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 14
13
however is not equal on all residues of a protein. For instance, buried residues at an active site or
at a binding site are generally more conserved than residues in loops. Also evolutionary pressure
may force addition or deletion of gene segments from the genomes leading to different gene and
protein sizes. Orthologue identifications has been done by various different approaches amongst
which a Best-Reciprocal BLAST Hit (BBH) or Bi-directional Best BLAST hit (BDBH) approach
has been considered the best33. However, this approach has its own flaws in the sense that only
one-to-one orthologous pairs are found, for duplicated genes or paralogues only a single hit is
found. The PSI-BLAST search tool helps in identification of regions of importance (not variable)
and gives them more weight in subsequent comparisons. The PSI-BLAST tool may also detect
subtle relationships between proteins that are distant structural or functional homologues. Such
relationships are often not detected by a simple BLAST search.
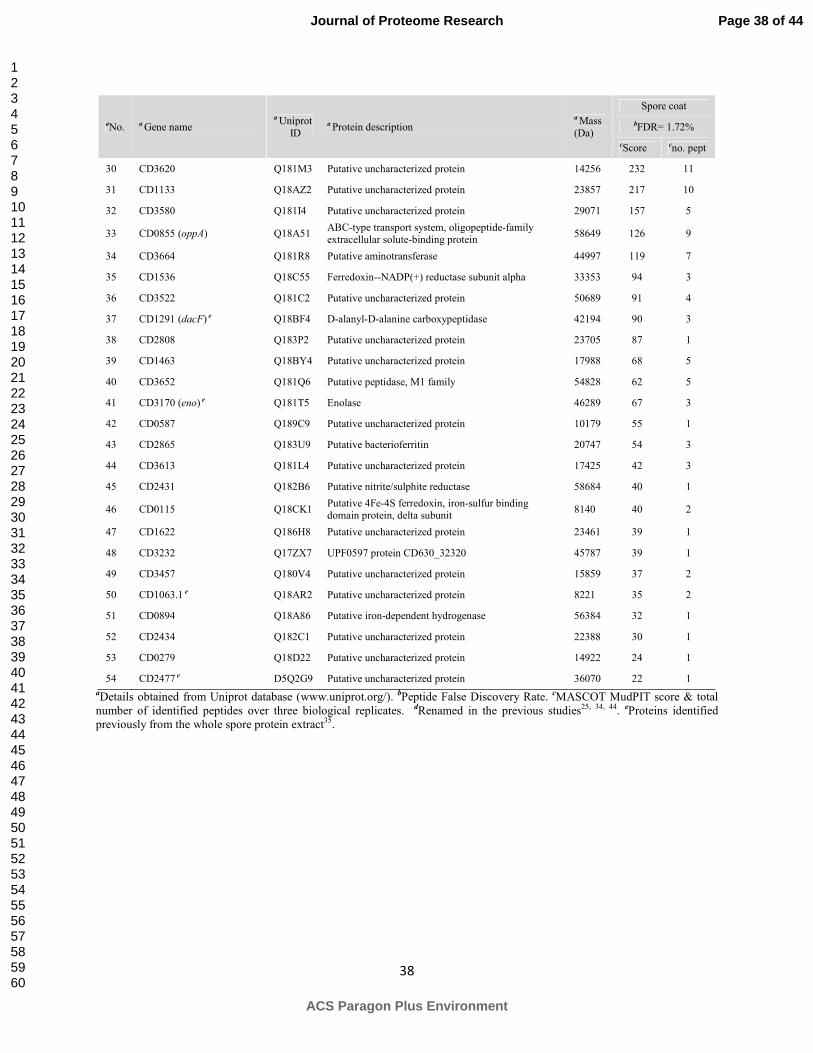
Our current study focused for the first time on the insoluble fraction and led to the
identification of total 100 proteins from the spore coat and exosporium layers of B. cereus ATCC
14579 and 54 proteins from the spore coat layers of C. difficile 630 (Table 1 & 2). These proteins
were then used for orthologue evaluation using PSI-BLAST search (see Supplementary Tables 2,
3, 4 and 5). As mentioned above, due to the possibility of various sizes of proteins, we assigned
the sequence identity threshold at 30% and an E-value of 0.005 was the default value chosen.
The experimentally identified proteins with conserved orthologues in two Bacilli and a
Clostridium species analyzed by us are listed in Table 3. As seen proteins CotJC, DacF, SpoIVA
and YisY were identified experimentally from all the three organisms. For further comparative
analysis, we selected five Bacillus species (11 different strains) and four Clostridium species
(total 13 different strains) available on the Genolist database (http://genolist.pasteur.fr/GenoList).
Proteins SafA (BC_4420), CotE (BC_3770), YhcN (BC_4419), YusW (BC_0212), CotD
Page 13 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 15
14
(BC_1560), YxeE (BC_3534) and others (Table 1) are among the proteins identified only from
coat + exosporium (fraction 1) and coat (fraction 2) confirming their localization in the spore
coat while proteins InA (BC_1284), CalY (BC_1279) and BC_1591 were among those identified
only from fraction 1 & exosporium (fraction 3). As seen there was a considerable overlap
between fractions 2 & 3 id est between spore coat and exosporium fractions implying two
possibilities - either these proteins may be spread across both the layers or the removal of the
exosporium layer was inefficient. Five proteins - BC_0944, SodF (BC_1468), YppG (BC_1559),
BC_2237 and BclB (BC_2382) - were identified only by a semi-trypsin enzyme search.
In case of C. difficile 630, CdeC34 (CD1067), CD1581, CotA (CD1613), CotB (CD1511),
CotJB1/CotCA (CD0597), CotJC1/CotCB (CD0598), CotJC2/CotD (CD2401), CotE (CD1433),
CotF (CD0596), CotJB2 (CD2400), Rbr (CD2845), SipL (CD3567), were among the high
scoring proteins with > 10 peptides identified per protein. In total, from the identified C. difficile
proteins 24 proteins have been identified previously35 from the whole spore protein extracts. We
did not study a separate exosporium fraction for C. difficile 630 as the stability of the exosporial
layer may depend on the spore preparation method as well as their storage and in our sample the
presence of exosporium was inconsistent possibly due to its loss during spore harvest. (see
Materials & Methods).
We divided the identified proteins from both species into six different categories based
on their possible function as indicated by their descriptions in the database - (1) proteins
involved in spore coat morphogenesis and other known spore coat proteins; (2) spore coat
proteins with a possible role in spore resistance; (3) exosporium proteins likely to be involved in
attachment to surfaces; (4) exosporium proteins possibly involved in pathogenicity; (5) spore
Page 14 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 16
15
coat proteins possibly involved in spore germination and (6) other putative spore coat and/or
exosporium proteins.
1. Proteins involved in spore coat morphogenesis and other known spore coat proteins. In
the previous studies in B. subtilis different morphogenetic proteins important for the
morphogenesis of the spore coat were assigned. These proteins include, starting from the cortex
and moving towards the outer coat, SpoVM, SpoIVA, SpoVID, SafA and CotE. For the
biogenesis of the inner coat SpoIVA and SafA are important while CotE plays a pivotal role in
outer coat development15, 23. In our current study we identified CotE (BC_3770), SafA
(BC_4420) and SpoIVA (BC_1509) from B. cereus ATCC 14579 and SpoIVA (CD2629) as well
as SipL (CD3567, recently identified36 morphogenetic protein) from C. difficile 630. The CotE
orthologue of B. subtilis was not found by protein sequence comparison in the C. difficile 630
proteome. However CD1433 renamed as CotE25, was identified. Orthologues of other
morphogenetic proteins i.e. SpoVID, SpoVM were not identified but for these two proteins it is
important to keep in mind that they are mostly identified in the soluble fraction of spores from B.
subtilis37, 38. In B. subtilis coat proteins CotC, CotG and CotU are known to be important for the
coat structure wherein cross-linking among these proteins plays a major role39-42. Genes for
CotC, CotG, CotS, CotT and CotW are not present in the B. cereus genome15. Likewise,
orthologues of SpoVID, SpoVM, CotC, CotG, CotT, CotU and CotW are not observed in the C.
difficile genome indicating a possible difference in the spore structure. Spore coat G (BC_2030)
identified in our study should not be confused with CotG from B. subtilis as it is 89% identical to
the highly phosphorylated exosporium protein ExsB from B. anthracis. The phosphorylated
region of BC_2030 is 96% identical to the region identified from ExsB from B. anthracis43.
Identical to B. anthracis ExsB43, we identified Threonine residues at positions 76, 79, 82, 105,
Page 15 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 17
16
114, 117, 129 from BC_2030 as phosphorylated in our error-tolerant MASCOT search.
Identified protein CD2399 is a paralogue of CD0596 (renamed as CotF44) also identified in the
current study. Both these proteins contain a CotJA superfamily domain of spore proteins and
thus could function as the CotJA orthologues. Two copies of each, spore coat B (BC_0389 and
BC_0390), spore coat Y (BC_1218) and BC_1222) and spore coat X (BC_2872 and BC_2874),
are present in the genome of B. cereus ATCC 14579, and these were all identified in this study.
Considering the Bacillus species in which the orthologues of these proteins are found and based
on the fact that B. subtilis has an orthologue of one of each duo (as predicted from the GenoList
database), it is possible that these proteins are localized either in the spore coat, the exosporium
or in the basal layer of the exosporium. For instance, protein spore coat Y (BC_1218) is in fact a
close orthologue of exosporium morphogenetic protein BAS1141 or ExsY from B. anthracis45.
Another structural protein identified in our study was spore coat protein D (BC_1560), which is
an orthologue of CotD - an inner coat protein in B. subtilis identified prominently from the
soluble fraction. This indicates a possible structural difference amongst B subtilis and B. cereus
spore coats.
From B. cereus, we also identified protein BC_4419, an orthologue of YhcN from B.
subtilis (< 30% sequence identity and thus left out of the orthologue data (Supplementary Table
2)) that contains a Spore_YhcN_YlaJ motif. This motif is reported to be found in lipoproteins
present in spores and not in vegetative cells. In B. subtilis 168 spores YhcN is suggested to be
located in the inner membrane or integument of the spore46. Although the expression of YhcN in
B. subtilis is controlled by the transcription factor σG (expressed late in the forespore during
sporulation47) due to the signal sequence carried by the protein it may be identified in the
insoluble coat fraction. The collagen adhesion protein (BC_5056) contains a TQxA domain and
Page 16 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 18
17
is conserved amongst the B. cereus group. TQxA domain containing proteins have been reported
to be associated with LPXTG-containing sortase target domains. Additionally, LPXTG proteins
are located at the cell wall peptidoglycan48. It is plausible that BC_5056 is in the spore coat or in
the integument region connecting cortex and the coat. Another interesting protein is the L-alanyl-
D-glutamate peptidase BC_2677, an orthologue of CwlK which is thought to be involved in cell
wall peptidoglycan hydrolysis in B. subtilis49. This protein has a VanY domain that is present in
proteins involved in cell wall biosynthesis. Additionally, BC_2677 also has SH3 superfamily
domain. Proteins belonging to the SH3 family are involved in the formation of multi-protein
complex assemblies50. This is an essential feature in spore coat build-up and this protein could be
an interesting candidate for studying structural built-up in B. cereus spores. Superoxide
dismutase (SodA) also plays an important role in spore coat assembly in B. subtilis by mediating
protein cross-linking. We did not identify the enzyme in B. cereus ATCC 14579. By a
semitrypsin enzyme search with MASCOT we identified SodF (BC_1468) from the spore coat
fraction. The superoxide dismutase (CD1631) from the genome of C. difficile was not identified
in our study but, to our surprise, PSI-BLAST with one iteration did not predict orthologue for
SodA either. In fact CD1631 is more identical to SodF identified from exosporium fractions
previously (Supplementary Table 5).
2. Spore coat proteins with a role in spore resistance. CotA from B. subtilis is known to play a
role in UV-resistance of spores. Interestingly CotA from B. subtilis has orthologues present in C.
botulinum A ATCC 19397, C. botulinum A ATCC 3502 and C. botulinum strain A Hall but not
in any other species and/or strains considered for orthologue evaluation (Supplementary Table
4). On the other hand CD1613 (renamed as CotA) suggested to be involved in assembly of the
outer coat in C. difficile 630 spores44 was identified. The newly identified protein BC_4047 is an
Page 17 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 19
18
orthologue of the outer coat protein Cotα (BAS3957) in B. anthracis Sterne and contains a
DUF3915 domain of unknown function. There are three Cys residues at the amino terminus (first
20 residues) and similar to the case in B. anthracis spores51, these are likely to be cross-linked.
The absence of Cotα is reported to affect visual appearance of the outer spore coat as well as the
chemical resistance and sensitivity properties of the spores towards phenol, chloroform, and
hypochlorite51. In anaerobic organisms defense against oxidative stress is of prime importance.
Since spores are important in the dissemination of the clostridial species it is likely that proteins
with ability to scavenge the oxidative radicals are present in the outermost layers of the spores.
This is also evident from the identified catalases (CotJC1/CotCB (CD0598), CotG (CD167)),
ferritin like proteins, ruberythrins (CD0825, CD1524, CD2845) and oxidoreductases (CD0117,
CD0176, CD1623) from C. difficile spores (Supplementary Table 1). These proteins may play a
dual role - one in structural built-up of spores coat layers by mediating dityrosine cross-linking
among proteins and/or second in resistance against oxidative stress52, 53 or they may have some
other hitherto unknown functions. For instance, in addition to the radical scavenging activity,
the oxidoreductase CD1623 also has a predicted β-lactamase domain and thus might be
important in antibiotic resistance.
3. Exosporium proteins likely to be involved in attachment to surfaces. The exosporium
layer has been less extensively studied than the spore coat. BclA from B. anthracis is one of the
few exosporium proteins that is well characterized and is an important structural component of
the hair-like nap in exosporium54, 55. We could not identify this protein (accession no.
HM071986.116) in our B. cereus exosporium preparations but proteins BC_3345 & BC_2569
(56% and 58% identical to HM071986.1 respectively) and BclC (BC_3712) were identified in
this study. These Bcl-family proteins, in general, form an extended rod-like structure in the
Page 18 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 20
19
regions that possess the (Gly-X-Y)n motifs, which is characteristic for a triple helix, rod-like
structure formation56. A recent study57 showed that the BC_2569 protein, similar to BclA,
contains a potential NTD attachment motif (-INPDLLGPTLPAI-) necessary to dock this protein
in the basal layer of the exosporium. From C. difficile, CD0332 (BclA1) was identified which
indicates that the presence of the exosporium in spores was inconsistent in our study. As opposed
to the predictions of algorithms used by databases such as KEGG (www.genome.jp/kegg/),
orthologues for Bcl family of proteins were not identified to be conserved by the PSI-BLAST
analysis by a single PSI-BLAST iteration from any other clostridial species considered in this
study.
Recently other exosporium proteins BxpA, BxpB, BclB19, 58, 59 have also been identified
in B. anthracis. Protein BxpA (BC_2149) has an amino acid sequence unusually rich in
glutamine and proline60. Interestingly, VrrA protein in B. anthracis is also rich in glutamine and
proline residues which are suggested to be involved in cross-linking of protein subunits61. BxpB
is an exosporium basal layer protein in B. anthracis Sterne58 and its orthologue BC_1221 in B.
cereus ATCC 14579 was identified in the current study. In B. anthracis BxpB has a paralogue
~80% identical, named ExsFB (BAS2303) and the identified protein BC_2374 is an orthologue
of ExsFB. Both BC_1221 & BC_2374 are 75% identical to BxpB (BAS1144) and ExsFB
(BAS2303) as shown by a Clustal Omega alignment tool. This suggests that BC_1221 and
BC_2374 may be localized in the basal layer of the B. cereus ATCC 14579 exosporium in
providing attachment sites for BclA-like proteins. In bxpB mutants of B. anthracis, the function
of BxpB could not be complemented with BAS2303, implying a different function for BAS2303
in B. anthracis spores58. Orthologues for BxpA (BC_2149), BxpB (BC_1221) and ExsFB
(BC_2374) are well conserved in the B. cereus group. Protein BC_2493 is an orthologue of
Page 19 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 21
20
exosporium protein ExsK identified from the exosporium fraction in B. anthracis (BA_2554) in
a study of Redmond et al.27
Identified cell surface proteins BC_2639 and BC_3547 (523.5 kDa and 97.9 kDa,
respectively) contain multiple unknown domains named DUF11 and non-specific domains called
B_ant_repeat domains. These B_ant_repeat domain containing proteins are shown to be encoded
by one, two or three very conserved large genes in the B. cereus group62. These genes are
expressed in the last developmental time waves during sporulation63. Although a function is
unknown it has been reported that these proteins may have a similar function to that of proteins
that form ribbon-like appendage structure on the exosporium of Clostridium taeniosporum
spores62. These proteins denoted P29a and P29b in C. taeniosporum are smaller in size than the
proteins we identified, but the size may be extremely variable between species. These
appendage-like structures may be common in spores of Bacilli and of Clostridia64 and may
facilitate spore dissemination, assist in spore nutrition during formation, or have no function and
result from a deranged metabolism65.
4. Exosporium proteins possibly involved in pathogenicity. Since the exosporium is the
outermost layer of the spores in many species, it is the initial point of contact with the host cells
and tissues. Therefore, in addition to providing unique adherence and hydrophobic properties to
spores, it may express molecules that play a significant role in spore attachment and
pathogenicity. The BclA protein is suggested to be the immunodominant epitope on the surface
of Bacillus anthracis spores and contains a C-terminal TNF-like domain55, 66. Though not
required for pathogenesis67 BclA may mediate internalization of spores by host cells via the
C1q/TNF-like domain. Putative uncharacterized proteins BC_2266 and BC_2267, identified in
our study both possess a domain belonging to the TNF superfamily and has a weak homology
Page 20 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 22
21
with the C1q domain. The C1q domain has been reported to play a role in spore attachment and
in host entry mechanisms where spores are suggested to bind to the gC1qR receptors on the host
cell via some unidentified molecules, in a Ca2+ dependent manner68. Thus, BC_2266 and
BC_2267 could very well be mediators involved in a spore attachment and host entry process.
Tertiary structure modeling with Phyre2 tool for both molecules showed that they have 99-100%
homology with TNF in the region from amino acids 70 to 150. About 79% to 84% of the
sequence could be modeled with over 90% confidence. Further studies are required to
understand the precise location and the function (e.g. using animal models) of these two proteins.
5. Spore coat proteins involved in spore germination. While the spores of Bacillus and
Clostridium species can survive for years in the dormant state, they can be rapidly converted to a
metabolically active state via the process of spore germination and outgrowth. A variety of small
molecules can trigger spore germination preliminary by binding to the germination receptors in
the inner membrane. L-alanine is known to be a potent germinant of B. cereus spores, whereas
D-alanine inhibits germination. Alanine racemase (Alr1; BC_0264) by converting L-alanine to
its stereoisomer thereby preventing germination69 and the inosine-uridine preferring nucleoside
hydrolase (IunH; BC_2889) by converting the germinant inosine to D-ribose could play a role in
preventing germination70. Once germination is triggered, the coat needs to be broken down and
peptidases identified in this study (Supplementary Table 3.) may be involved in germination.
Subsequent spore outgrowth requires that the spore cortex is degraded immediately once
germination is triggered. Cortex lytic enzymes (CLEs) like CwlJ and SleB play an important role
in cortex degradation and are located in the B. subtilis spore coat. In B. cereus ATCC 14579 we
identified SleB (BC_2753) and CwlJ (BC_5390). Identified protein BC_5391 is an orthologue of
GerQ/YwdL that is involved in proper localization of the cortex lytic enzyme CwlJ in the B.
Page 21 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 23
22
subtilis spore coat71. However, in other studies GerQ/YwdL and CwlJ were identified in the
exosporium fraction of B. cereus ATCC 10876 and B. anthracis spores, respectively60, 72.
Another enzyme identified from the insoluble fraction in this study and reported to be involved
in spore cortex degradation73 during spore germination was YaaH orthologue BC_3607. Newly
identified BC_1591 contains a predicted pectate lysase domain, which is related to the family of
glycosyl hydrolase proteins and may therefore be involved in peptidoglycan modification,
degradation or synthesis. In addition there were also other hydrolases identified in our study that
can be involved in spore germination (Supplementary Table 3.). From C. difficile the
predominant cortex lytic enzyme SleC (CD0551) was identified in this study. Of further interest,
the transcriptomic data of Dembek and colleagues74 shows that genes cd0115, cd0116, cd0117,
cd0176, cd0279, cd0587, cd0825, cd0855, cd1133, cd1524, cd1536, cd1622, cd1623, cd3232,
cd3664 are down-regulated while genes cd0213, cd1511, cd1613, cd2845, cd3567 are up-
regulated upon initiation of germination. It is noteworthy that genes cd0115, cd0116, cd0117,
cd0176, cd1536, cd1623 (all encoding oxidoreductases) and cd0825, cd1524, cd2845 (all
encoding ruberythrins) may play a role in energy-dependent germination75 of Clostridium spores.
We identified all these gene products and thus it could be worthwhile to study their role in spore
germination.
6. Other putative spore coat and/or exosporium proteins. Another newly identified protein in
our study could be important target for the B. cereus group of spore formers. It is an orthologue
of BAS5303 in B. anthracis Sterne76 and BA5699 in B. anthracis Ames, identified previously in
the exosporium fraction in the Ames strain60 and also shown to be immunogenic in animal
models76, 77. BC_0996 could therefore also be located in the exosporium of B. cereus ATCC
14579, although its function remains unclear. Protein BC_0987 is much less conserved in the B.
Page 22 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 24
23
cereus group and could very well be a candidate marker for the B. cereus spores. There were
many peptidases and hydrolases identified (Supplementary Table 1 & 2) which we could not
assign to any particular category and were classified as putative coat and or exosporium proteins.
The plasmid encoded BC_p0002 was found to be unique to the B. cereus ATCC 14579 strain.
Notably, this protein might have a distant relation (20% sequence identity, E-value 1.8) with the
envelope glycoprotein GP120 (residues 193 - 372) from Human Immunodeficiency Virus (HIV).
The highly abundant35 and cysteine-rich (~9%) protein CD1067, recently shown to be
important for exosporium assembly34, was identified and could very well be a potential marker
for this species. Another abundant protein CD1581 was identified as well and is uniquely present
in C. difficile 630. Identified protein CD3613 belongs to the Stay-green (SGR) protein family.
The SGR family is conserved in plants and is involved in chlorophyll degradation. Distant SGR
homologs are also present in algae and, unexpectedly, in species of Bacillus and Clostridium, but
not in other bacterial genomes78. However, the exact role of these proteins in bacterial species
remains to be studied. Protein CD3652 from C. difficile 630 has a characteristic GluZincin
protease domain. The GluZincin family (thermolysin-like peptidases or TLPs) includes several
zinc-dependent metallopeptidases and contain His-Glu-X-X-His and Glu-X-X-X-Asp motifs as
part of their active site. Interestingly, the light-chain (LC, 448 a.a from the N-terminus of the
protein) of the potent neurotoxin BoNT/A from C. botulinum also has this domain79. Previous
studies have confirmed that zinc-binding plays an essential role in the catalytic activity of the
light chain of BoNT/A. Thus protease CD3652 is an interesting target to study for its role in
spore-mediated infections.
As mentioned above, for orthologue evaluation using PSI-BLAST and to study the
conservation of coat and exosporium proteins we used the identified proteins from B. cereus
Page 23 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 25
24
ATCC 14579 (this study), C. difficile 630 (this study) and B. subtilis 16815, 22 as the initial
dataset. Figure 2 shows the distribution of orthologues respective to the three species mentioned.
As seen, identified B. subtilis coat proteins are very unique to the organism with 25 being only
present in B. subtilis. Although B. cereus and B. subtilis are evolutionary distant, 53 B. subtilis
coat proteins had orthologues present in B. cereus ATCC 14579 with > 30% sequence identity.
From the identified B. subtilis 168 spore coat protein dataset, 9 proteins - CotJB, CotJC, DacF,
SpoIVA, SleB, YabG, YhxC, and YtfJ were found to be conserved in selected Bacilli and
Clostridia (Fig. 2, Supplementary Table 4). Focusing on the identified proteins from B. cereus
ATCC 14579, it was seen that 77 proteins are conserved in the B. cereus group while 44 had
orthologues in B. subtilis 168 and 11 in C. difficile 630. Eleven of the identified proteins
emerged to be conserved in all of the spore forming species considered for comparative analysis.
Amongst the known exosporium proteins, 27 proteins have conserved orthologues
(Supplementary Table 5) in the B. cereus group including the Bcl-family of proteins. Alanine
racemase is conserved throughout the spore formers while in addition to BclA, only SodF has an
orthologue present in C. difficile and few strains of C. botulinum. From the C. difficile 630
identified proteins, 13 are unique to this strain, 12 have orthologues in B. subtilis 168, 13 have
orthologues in B. cereus and 8 are conserved across the spore formers considered for
comparison. Our orthologue predictions for certain B. subtilis and C. difficile proteins were also
supported by previous transcriptomic80 and the phylogenetic studies81 making our orthologue
identification method confident. Figure 3. summarizes the orthologue conservation results
highlighting potential protein targets for easy detection and removal of spores. There are 13 coat
proteins conserved in all spore formers considered in this study. From the exosporium proteins,
11 are conserved in the B. cereus group and only Alr has orthologues in all Clostridia. The exact
Page 24 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 26
25
localization of three proteins BC_0987, BC_0996 both conserved in the B. cereus group and
CD3613 conserved in Clostridia is not known.
Lastly, despite severe and harsh treatments to the sample pellets, we identified a number
of cytosolic proteins including highly abundant ribosomal proteins and elongation factors
(Supplementary Table 8). We think that since in both species the exosporium was not completely
removed, these proteins might have been trapped in the interspace region between the coat and
exosporium layers during sporulation. The fact that Liu et al.60 also identified orthologues of
many of these proteins from the exosporium fraction of B. anthracis indicates that the process of
exosporium attachment to the coat occurs in a highly regulated and precise manner inside the
mother cell cytoplasm. It is suggested that efficient removal of the exosporium is a must to
diminish the contribution of cytosolic proteins. Recently an effort has been made to remove
exosporium from C. difficile 630 spores82 but in Bacillus spp. efficient removal of exosporium
has not been achieved30. For the same reason, it is also difficult to classify the identified proteins
as solely coat or exosporium proteins.
Concluding Remarks
In conclusion, our gel-free method performs comprehensively by identifying coat and
exosporium proteins from different aerobic and anaerobic spore formers. We provide potential
protein targets emerging from this study, for use in food and health sectors for selective removal
of spores from the samples, and these targets need to be studied in detail. Also our study outlines
the general classes of proteins that are necessary for the spore integrity namely - the structural
proteins (eg. SpoIVA, SafA, CotE), proteins involved in resistance mechanisms (eg.
oxidoreductases, catalases, ferritin like proteins) and proteins that facilitate germination (eg.
Page 25 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 27
26
hydrolases, peptidases). Additionally, we provide a wide view of protein conservation in the
structure of spores from different aerobic and anaerobic pathogenic spore formers in an attempt
to derive a universal set of spore surface proteins. We propose that proteins CotJB, CotJC, DacF,
Gpr, Eno, SleB, SpoIVA, RnjA, YabG, YabP, YhxC, YloB, YtfJ make up the universally
conserved set of proteins in bacterial spore formers. Nevertheless, functional annotation of the
genomes, localization studies for the identified putative spore coat and exosporium proteins as
well as quantitative studies are necessary for gaining more detailed insights of the spore
structure.
Associated content
Supplementary Table 1. Conserved superfamily domains from the identified spore coat and
exosporium proteins from Bacillus cereus ATCC 14579 and Clostridium difficile 630.
Supplementary Table 2. Orthologue identification for the identified spore coat and exosporium
proteins from Bacillus cereus ATCC 14579.
Supplementary Table 3. Orthologue identification for the identified spore coat and exosporium
proteins from Clostridium difficile 630.
Supplementary Table 4. Orthologue identification for the previously assigned spore coat
proteins in Bacillus subtilis 168.
Supplementary Table 5. Orthologue identification for the previously assigned exosporium
proteins.
Supplementary Table 6. Cytosolic proteins (possibly from the interspace) identified in this
study.
Page 26 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 28
27
Author information
Corresponding author
*E-mail: [email protected]
Notes
The authors declare no competing financial interest.
Acknowledgements
Rokus van den Dool and Jolanda Verheul are thanked for the experimental help in this study.
W.A. acknowledges the Erasmus Mundus program (EMECW 15) for funding of his PhD project.
A.T.B. is supported by a grant from the Dutch Foundation for Applied Sciences (STW 10431).
References
1. Tourasse, N. J.; Helgason, E.; Okstad, O. A.; Hegna, I. K.; Kolsto, A. B., The Bacillus cereus group: novel aspects of population structure and genome dynamics. Journal of applied
microbiology 2006, 101, (3), 579-93. 2. Stenfors Arnesen, L. P.; Fagerlund, A.; Granum, P. E., From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS microbiology reviews 2008, 32, (4), 579-606. 3. Lund, T.; De Buyser, M. L.; Granum, P. E., A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Molecular microbiology 2000, 38, (2), 254-61. 4. Bottone, E. J., Bacillus cereus, a volatile human pathogen. Clinical microbiology reviews
2010, 23, (2), 382-98. 5. Bartlett, J. G.; Willey, S. H.; Chang, T. W.; Lowe, B., Cephalosporin-associated pseudomembranous colitis due to Clostridium difficile. JAMA : the journal of the American
Medical Association 1979, 242, (24), 2683-5. 6. Songer, J. G.; Anderson, M. A., Clostridium difficile: an important pathogen of food animals. Anaerobe 2006, 12, (1), 1-4. 7. Huang, H.; Weintraub, A.; Fang, H.; Nord, C. E., Antimicrobial resistance in Clostridium
difficile. International Journal of Antimicrobial Agents 2009, 34, (6), 516-522. 8. Razavi, B.; Apisarnthanarak, A.; Mundy, L. M., Clostridium difficile: emergence of hypervirulence and fluoroquinolone resistance. Infection 2007, 35, (5), 300-7. 9. Bartlett, J. G., New drugs for Clostridium difficile infection. Clinical infectious diseases : an
official publication of the Infectious Diseases Society of America 2006, 43, (4), 428-31.
Page 27 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 29
28
10. Brazier, J. S.; Raybould, R.; Patel, B.; Duckworth, G.; Pearson, A.; Charlett, A.; Duerden, B. I., Distribution and antimicrobial susceptibility patterns of Clostridium difficile PCR ribotypes in English hospitals, 2007-08. Euro surveillance : bulletin europeen sur les maladies transmissibles
= European communicable disease bulletin 2008, 13, (41). 11. Cairns, M. D.; Stabler, R. A.; Shetty, N.; Wren, B. W., The continually evolving Clostridium
difficile species. Future microbiology 2012, 7, (8), 945-57. 12. Dawson, L. F.; Valiente, E.; Wren, B. W., Clostridium difficile--a continually evolving and problematic pathogen. Infection, genetics and evolution : journal of molecular epidemiology and
evolutionary genetics in infectious diseases 2009, 9, (6), 1410-7. 13. Carter, G. P.; Rood, J. I.; Lyras, D., The role of toxin A and toxin B in Clostridium difficile-associated disease: Past and present perspectives. Gut microbes 2010, 1, (1), 58-64. 14. Setlow, P., Spore germination. Curr Opin Microbiol 2003, 6, (6), 550-6. 15. Henriques, A. O.; Moran, C. P., Jr., Structure, assembly, and function of the spore surface layers. Annu Rev Microbiol 2007, 61, 555-88. 16. Lequette, Y.; Garénaux, E.; Tauveron, G.; Dumez, S.; Perchat, S.; Slomianny, C.; Lereclus, D.; Guerardel, Y.; Faille, C., Role played by exosporium glycoproteins in the surface properties of Bacillus cereus spores and in their adhesion properties to stainless steel. Applied and
environmental microbiology 2011. 17. Paredes, C. J.; Alsaker, K. V.; Papoutsakis, E. T., A comparative genomic view of clostridial sporulation and physiology. Nature reviews. Microbiology 2005, 3, (12), 969-78. 18. de Hoon, M. J.; Eichenberger, P.; Vitkup, D., Hierarchical evolution of the bacterial sporulation network. Current biology : CB 2010, 20, (17), R735-45. 19. Leski, T. A.; Caswell, C. C.; Pawlowski, M.; Klinke, D. J.; Bujnicki, J. M.; Hart, S. J.; Lukomski, S., Identification and Classification of bcl Genes and Proteins of Bacillus cereus Group Organisms and Their Application in Bacillus anthracis Detection and Fingerprinting. Applied and environmental microbiology 2009, 75, (22), 7163-7172. 20. Thompson, B. M.; Hoelscher, B. C.; Driks, A.; Stewart, G. C., Localization and assembly of the novel exosporium protein BetA of Bacillus anthracis. Journal of bacteriology 2011, 193, (19), 5098-104. 21. Waller, L. N.; Stump, M. J.; Fox, K. F.; Harley, W. M.; Fox, A.; Stewart, G. C.; Shahgholi, M., Identification of a second collagen-like glycoprotein produced by Bacillus anthracis and demonstration of associated spore-specific sugars. Journal of bacteriology 2005, 187, (13), 4592-7. 22. Abhyankar, W.; Beek, A. T.; Dekker, H.; Kort, R.; Brul, S.; de Koster, C. G., Gel-free proteomic identification of the Bacillus subtilis insoluble spore coat protein fraction. Proteomics
2011, 11, (23), 4541-50. 23. McKenney, P. T.; Driks, A.; Eichenberger, P., The Bacillus subtilis endospore: assembly and functions of the multilayered coat. Nature reviews. Microbiology 2013, 11, (1), 33-44. 24. de Vries, Y. P.; Hornstra, L. M.; de Vos, W. M.; Abee, T., Growth and sporulation of Bacillus cereus ATCC 14579 under defined conditions: temporal expression of genes for key sigma factors. Applied and environmental microbiology 2004, 70, (4), 2514-9. 25. Permpoonpattana, P.; Tolls, E. H.; Nadem, R.; Tan, S.; Brisson, A.; Cutting, S. M., Surface layers of Clostridium difficile endospores. Journal of bacteriology 2011, 193, (23), 6461-70. 26. Todd, S. J.; Moir, A. J.; Johnson, M. J.; Moir, A., Genes of Bacillus cereus and Bacillus
anthracis encoding proteins of the exosporium. Journal of bacteriology 2003, 185, (11), 3373-8.
Page 28 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 30
29
27. Redmond, C.; Baillie, L. W.; Hibbs, S.; Moir, A. J.; Moir, A., Identification of proteins in the exosporium of Bacillus anthracis. Microbiology 2004, 150, (Pt 2), 355-63. 28. Hadžija, O., A simple method for the quantitative determination of muramic acid. Analytical
biochemistry 1974, 60, (2), 512-7. 29. Taylor, K. C. C., A simple colorimetric assay for muramic acid and lactic acid. Appl Biochem
Biotechnol 1996, 56, (1), 49-58. 30. Thompson, B. M.; Binkley, J. M.; Stewart, G. C., Current physical and SDS extraction methods do not efficiently remove exosporium proteins from Bacillus anthracis spores. J
Microbiol Methods 2011, 85, (2), 143-8. 31. Klobutcher, L. A.; Ragkousi, K.; Setlow, P., The Bacillus subtilis spore coat provides "eat resistance" during phagocytic predation by the protozoan Tetrahymena thermophila. Proceedings
of the National Academy of Sciences of the United States of America 2006, 103, (1), 165-70. 32. Scopes, R. K., Measurement of protein by spectrophotometry at 205 nm. Analytical
biochemistry 1974, 59, (1), 277-82. 33. Abecasis, A. B.; Serrano, M.; Alves, R.; Quintais, L.; Pereira-Leal, J. B.; Henriques, A. O., A genomic signature and the identification of new sporulation genes. Journal of bacteriology 2013. 34. Barra-Carrasco, J.; Olguin-Araneda, V.; Plaza-Garrido, A.; Miranda-Cardenas, C.; Cofre-Araneda, G.; Pizarro-Guajardo, M.; Sarker, M. R.; Paredes-Sabja, D., The Clostridium difficile Exosporium Cysteine (CdeC) Rich Protein is Required for Exosporium Morphogenesis and Coat Assembly. Journal of bacteriology 2013. 35. Lawley, T. D.; Croucher, N. J.; Yu, L.; Clare, S.; Sebaihia, M.; Goulding, D.; Pickard, D. J.; Parkhill, J.; Choudhary, J.; Dougan, G., Proteomic and genomic characterization of highly infectious Clostridium difficile 630 spores. Journal of bacteriology 2009, 191, (17), 5377-86. 36. Putnam, E. E.; Nock, A. M.; Lawley, T. D.; Shen, A., SpoIVA and SipL Are Clostridium
difficile Spore Morphogenetic Proteins. Journal of bacteriology 2013, 195, (6), 1214-25. 37. Cutting, S.; Anderson, M.; Lysenko, E.; Page, A.; Tomoyasu, T.; Tatematsu, K.; Tatsuta, T.; Kroos, L.; Ogura, T., SpoVM, a small protein essential to development in Bacillus subtilis, interacts with the ATP-dependent protease FtsH. Journal of bacteriology 1997, 179, (17), 5534-42. 38. Ozin, A. J.; Henriques, A. O.; Yi, H.; Moran, C. P., Jr., Morphogenetic proteins SpoVID and SafA form a complex during assembly of the Bacillus subtilis spore coat. Journal of bacteriology
2000, 182, (7), 1828-33. 39. Henriques, A. O.; Melsen, L. R.; Moran, C. P., Jr., Involvement of superoxide dismutase in spore coat assembly in Bacillus subtilis. Journal of bacteriology 1998, 180, (9), 2285-91. 40. Isticato, R.; Esposito, G.; Zilhao, R.; Nolasco, S.; Cangiano, G.; De Felice, M.; Henriques, A. O.; Ricca, E., Assembly of multiple CotC forms into the Bacillus subtilis spore coat. Journal of
bacteriology 2004, 186, (4), 1129-35. 41. Isticato, R.; Pelosi, A.; Zilhao, R.; Baccigalupi, L.; Henriques, A. O.; De Felice, M.; Ricca, E., CotC-CotU heterodimerization during assembly of the Bacillus subtilis spore coat. Journal of
bacteriology 2008, 190, (4), 1267-75. 42. Zilhão, R.; Serrano, M.; Isticato, R.; Ricca, E.; Moran, C. P., Jr.; Henriques, A. O., Interactions among CotB, CotG, and CotH during assembly of the Bacillus subtilis spore coat. Journal of bacteriology 2004, 186, (4), 1110-9. 43. McPherson, S. A.; Li, M.; Kearney, J. F.; Turnbough, C. L., Jr., ExsB, an unusually highly phosphorylated protein required for the stable attachment of the exosporium of Bacillus
anthracis. Molecular microbiology 2010, 76, (6), 1527-38.
Page 29 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 31
30
44. Permpoonpattana, P.; Phetcharaburanin, J.; Mikelsone, A.; Dembek, M.; Tan, S.; Brisson, M. C.; La Ragione, R.; Brisson, A. R.; Fairweather, N.; Hong, H. A.; Cutting, S. M., Functional Characterization of Clostridium difficile Spore Coat Proteins. Journal of bacteriology 2013. 45. Boydston, J. A.; Yue, L.; Kearney, J. F.; Turnbough, C. L., Jr., The ExsY protein is required for complete formation of the exosporium of Bacillus anthracis. Journal of bacteriology 2006, 188, (21), 7440-8. 46. Bagyan, I.; Noback, M.; Bron, S.; Paidhungat, M.; Setlow, P., Characterization of yhcN, a new forespore-specific gene of Bacillus subtilis. Gene 1998, 212, (2), 179-88. 47. Errington, J., Regulation of endospore formation in Bacillus subtilis. Nat Rev Microbiol
2003, 1, (2), 117-26. 48. Aucher, W.; Davison, S.; Fouet, A., Characterization of the sortase repertoire in Bacillus
anthracis. PloS one 2011, 6, (11), e27411. 49. Fukushima, T.; Yao, Y.; Kitajima, T.; Yamamoto, H.; Sekiguchi, J., Characterization of new L,D-endopeptidase gene product CwlK (previous YcdD) that hydrolyzes peptidoglycan in Bacillus subtilis. Molecular genetics and genomics : MGG 2007, 278, (4), 371-83. 50. Mirey, G.; Soulard, A.; Orange, C.; Friant, S.; Winsor, B., SH3 domain-containing proteins and the actin cytoskeleton in yeast. Biochemical Society transactions 2005, 33, (Pt 6), 1247-9. 51. Kim, H. S.; Sherman, D.; Johnson, F.; Aronson, A. I., Characterization of a major Bacillus
anthracis spore coat protein and its role in spore inactivation. Journal of bacteriology 2004, 186, (8), 2413-7. 52. Lehmann, Y.; Meile, L.; Teuber, M., Rubrerythrin from Clostridium perfringens: cloning of the gene, purification of the protein, and characterization of its superoxide dismutase function. Journal of bacteriology 1996, 178, (24), 7152-8. 53. Riebe, O.; Fischer, R. J.; Wampler, D. A.; Kurtz, D. M., Jr.; Bahl, H., Pathway for H2O2 and O2 detoxification in Clostridium acetobutylicum. Microbiology 2009, 155, (Pt 1), 16-24. 54. Brahmbhatt, T. N.; Janes, B. K.; Stibitz, E. S.; Darnell, S. C.; Sanz, P.; Rasmussen, S. B.; O'Brien, A. D., Bacillus anthracis exosporium protein BclA affects spore germination, interaction with extracellular matrix proteins, and hydrophobicity. Infection and immunity 2007, 75, (11), 5233-9. 55. Kailas, L.; Terry, C.; Abbott, N.; Taylor, R.; Mullin, N.; Tzokov, S. B.; Todd, S. J.; Wallace, B. A.; Hobbs, J. K.; Moir, A.; Bullough, P. A., Surface architecture of endospores of the Bacillus
cereus/anthracis/thuringiensis family at the subnanometer scale. Proceedings of the National
Academy of Sciences of the United States of America 2011, 108, (38), 16014-9. 56. Beck, K.; Brodsky, B., Supercoiled protein motifs: the collagen triple-helix and the alpha-helical coiled coil. Journal of structural biology 1998, 122, (1-2), 17-29. 57. Tan, L.; Turnbough, C. L., Jr., Sequence motifs and proteolytic cleavage of the collagen-like glycoprotein BclA required for its attachment to the exosporium of Bacillus anthracis. Journal of
bacteriology 2010, 192, (5), 1259-68. 58. Steichen, C. T.; Kearney, J. F.; Turnbough, C. L., Jr., Characterization of the exosporium basal layer protein BxpB of Bacillus anthracis. Journal of bacteriology 2005, 187, (17), 5868-76. 59. Thompson, B. M.; Hoelscher, B. C.; Driks, A.; Stewart, G. C., Assembly of the BclB glycoprotein into the exosporium and evidence for its role in the formation of the exosporium 'cap' structure in Bacillus anthracis. Molecular microbiology 2012, 86, (5), 1073-84.
Page 30 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 32
31
60. Liu, H.; Bergman, N. H.; Thomason, B.; Shallom, S.; Hazen, A.; Crossno, J.; Rasko, D. A.; Ravel, J.; Read, T. D.; Peterson, S. N.; Yates, J., 3rd; Hanna, P. C., Formation and composition of the Bacillus anthracis endospore. Journal of bacteriology 2004, 186, (1), 164-78. 61. Andersen, G. L.; Simchock, J. M.; Wilson, K. H., Identification of a region of genetic variability among Bacillus anthracis strains and related species. Journal of bacteriology 1996, 178, (2), 377-84. 62. Walker, J. R.; Gnanam, A. J.; Blinkova, A. L.; Hermandson, M. J.; Karymov, M. A.; Lyubchenko, Y. L.; Graves, P. R.; Haystead, T. A.; Linse, K. D., Clostridium taeniosporum spore ribbon-like appendage structure, composition and genes. Molecular microbiology 2007, 63, (3), 629-43. 63. Reiter, L.; Tourasse, N. J.; Fouet, A.; Loll, R.; Davison, S.; Okstad, O. A.; Piehler, A. P.; Kolsto, A. B., Evolutionary history and functional characterization of three large genes involved in sporulation in Bacillus cereus group bacteria. Journal of bacteriology 2011, 193, (19), 5420-30. 64. Driks, A., Surface appendages of bacterial spores. Molecular microbiology 2007, 63, (3), 623-5. 65. Rode, L. J.; Crawford, M. A.; Williams, M. G., Clostridium spores with ribbon-like appendages. Journal of bacteriology 1967, 93, (3), 1160-73. 66. Réty, S.; Salamitou, S.; Garcia-Verdugo, I.; Hulmes, D. J.; Le Hegarat, F.; Chaby, R.; Lewit-Bentley, A., The crystal structure of the Bacillus anthracis spore surface protein BclA shows remarkable similarity to mammalian proteins. The Journal of biological chemistry 2005, 280, (52), 43073-8. 67. Bozue, J.; Cote, C. K.; Moody, K. L.; Welkos, S. L., Fully virulent Bacillus anthracis does not require the immunodominant protein BclA for pathogenesis. Infection and immunity 2007, 75, (1), 508-11. 68. Ghebrehiwet, B.; Tantral, L.; Titmus, M. A.; Panessa-Warren, B. J.; Tortora, G. T.; Wong, S. S.; Warren, J. B., The exosporium of B. cereus contains a binding site for gC1qR/p33: implication in spore attachment and/or entry. Advances in experimental medicine and biology
2007, 598, 181-97. 69. Chesnokova, O. N.; McPherson, S. A.; Steichen, C. T.; Turnbough, C. L., Jr., The spore-specific alanine racemase of Bacillus anthracis and its role in suppressing germination during spore development. Journal of bacteriology 2009, 191, (4), 1303-10. 70. Liang, L.; He, X.; Liu, G.; Tan, H., The role of a purine-specific nucleoside hydrolase in spore germination of Bacillus thuringiensis. Microbiology 2008, 154, (Pt 5), 1333-40. 71. Ragkousi, K.; Eichenberger, P.; van Ooij, C.; Setlow, P., Identification of a new gene essential for germination of Bacillus subtilis spores with Ca2+-dipicolinate. Journal of
bacteriology 2003, 185, (7), 2315-29. 72. Terry, C.; Shepherd, A.; Radford, D. S.; Moir, A.; Bullough, P. A., YwdL in Bacillus cereus: its role in germination and exosporium structure. PloS one 2011, 6, (8), e23801. 73. Lambert, E. A.; Popham, D. L., The Bacillus anthracis SleL (YaaH) protein is an N-acetylglucosaminidase involved in spore cortex depolymerization. Journal of bacteriology 2008, 190, (23), 7601-7. 74. Dembek, M.; Stabler, R. A.; Witney, A. A.; Wren, B. W.; Fairweather, N. F., Transcriptional Analysis of Temporal Gene Expression in Germinating Clostridium difficile 630 Endospores. PloS one 2013, 8, (5), e64011.
Page 31 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 33
32
75. Ando, Y.; Tsuzuki, T., Energy-dependent activation of spore-lytic enzyme precursor by germinated spores of Clostridium perfringens. Biochemical and biophysical research
communications 1984, 123, (2), 463-7. 76. Cybulski, R. J., Jr.; Sanz, P.; McDaniel, D.; Darnell, S.; Bull, R. L.; O'Brien, A. D., Recombinant Bacillus anthracis spore proteins enhance protection of mice primed with suboptimal amounts of protective antigen. Vaccine 2008, 26, (38), 4927-39. 77. Walper, S.; Lee, P.; Anderson, G.; Goldman, E., Selection and Characterization of Single Domain Antibodies Specific for Bacillus anthracis Spore Proteins. Antibodies 2013, 2, (1), 152-167. 78. Barry, C. S., The stay-green revolution: Recent progress in deciphering the mechanisms of chlorophyll degradation in higher plants. Plant Science 2009, 176, (3), 325-333. 79. Fu, Z.; Chen, S.; Baldwin, M. R.; Boldt, G. E.; Crawford, A.; Janda, K. D.; Barbieri, J. T.; Kim, J. J., Light chain of botulinum neurotoxin serotype A: structural resolution of a catalytic intermediate. Biochemistry 2006, 45, (29), 8903-11. 80. Galperin, M. Y.; Mekhedov, S. L.; Puigbo, P.; Smirnov, S.; Wolf, Y. I.; Rigden, D. J., Genomic determinants of sporulation in Bacilli and Clostridia: towards the minimal set of sporulation-specific genes. Environmental microbiology 2012, 14, (11), 2870-90. 81. Traag, B. A.; Pugliese, A.; Eisen, J. A.; Losick, R., Gene conservation among endospore-forming bacteria reveals additional sporulation genes in Bacillus subtilis. Journal of bacteriology
2013, 195, (2), 253-60. 82. Escobar-Cortes, K.; Barra-Carrasco, J.; Paredes-Sabja, D., Proteases and sonication specifically remove the exosporium layer of spores of Clostridium difficile strain 630. Journal of
microbiological methods 2013.
Page 32 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 34
33
Figures
Figure 1. Venn diagramme of conserved superfamily domains in the identified spore coat & exosporium proteins from B. subtilis
168, B. cereus ATCC 14579 and C. difficile 630. A total of 148 superfamily domains were assigned to the identified proteins
from the three species indicated. The numbers correspond to the superfamily domains from the identified proteins. The eleven
superfamily domains common to the three organisms are listed in the box.
Figure 2. Orthologue distribution for identified spore coat & exosporium proteins from B. subtilis 168, B. cereus ATCC 14579
and C. difficile 630. Query protein sequences used for orthologue identification comprised of proteins identified from B. subtilis
168 (92 proteins)15, 22, B. cereus ATCC 14579 (100 proteins; this study) and C. difficile 630 (54 proteins; this study). The bars
indicate the number of proteins from the respective query datasets that have orthologues present in the species or groups
indicated below. * Number of proteins exclusive for indicated organisms studied in our work (See Supplementary Tables 2,3 & 4
for details).
Figure 3. Potential target proteins from spore coat and/or exosporium layers emerging from this study. Spore coat proteins
presented are conserved in the spore formers considered for the comparison. a Exosporium proteins conserved in the B. cereus
group (Alr is conserved in Bacilli as well as in Clostridia). b Localization of the proteins is unknown. BC_0987 & BC_0996 are
conserved in the B. cereus group and CD3613 is conserved in Clostridia.
Page 33 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 35
34
Table 1. Identified spore coat and exosporium proteins from Bacillus cereus ATCC 14579.
aNo. a Gene name a Uniprot ID
a Protein description a Mass (Da)
Spore coat & Exosporium
Spore coat Exosporium
bFDR= 1.77% FDR= 1.62% FDR= 1.99% cScore cno.
pept Score no.
pept Score no.
pept
(I) Proteins involved in spore coat morphogenesis and other known spore coat proteins
1 BC_4420 (safA) d Q812R2 SpoVID-dependent spore coat assembly factor SafA
65354 1286 46 2357 70
2 BC_2872 (cotX) Q81CA3 Spore coat protein X 16345 1145 28 1288 32 216 5
3 BC_2030g Q81EF0 Spore coat protein G 24013 1141 36 1309 41 471 30
4 BC_0389 (cotB) Q81IJ7 Spore coat protein B 19768 1109 67 2763 100 524 23
5 BC_3770 (cotE) d Q81A24 Spore coat protein E 20615 971 37 122 5
6 BC_0390 (cotB) Q81IJ6 Spore coat protein B 17339 744 29 1563 39 328 11
7 BC_4419 (yhcN) d Q817V9 Putative uncharacterized protein 24526 672 21 628 21
8 BC_1222 Q81GH8 Spore coat protein Y 17596 486 19 729 21 5668 188
9 BC_5056 d Q815S6 Collagen adhesion protein 34657 383 19 1427 37
10 BC_2874 (cotX) Q81CA1 Spore coat protein X 21850 306 12 293 12 36 1
11 BC_1509 (spoIVA) Q81FR0 Stage IV sporulation protein A 55636 242 11
12 BC_4640 (ytfJ) d Q817B7 Putative uncharacterized protein 13968 237 10 81 3
13 BC_1560 (cotD) d Q81FM0 Spore coat protein D 15393 214 10 565 16
14 BC_0212 (yusW) d Q81IY1 Putative uncharacterized protein 17725 122 3 204 6
15 BC_4075 (dacF) d Q819B1 D-alanyl-D-alanine carboxypeptidase
44032 85 3 150 5
16 BC_1279 (cotN) Q81GC8 Spore coat-associated protein N 21848 79 3
17 BC_1245 Q81GF8 Putative uncharacterized protein 15287 39 2
18 BC_0822 (cotJB) Q81HI6 CotJB protein 10523 45 1
19 BC_0821 (cotJC) Q81HI7 CotJC protein 21694 37 1
20 BC_0823 (cotJA) Q81HI5 CotJA protein 8475 30 1
21 BC_3534 (yxeE) d Q81AM8 IG hypothetical 17193 16298 21 1 33 1
22 BC_2095 (ytfJ) d Q81E93 Putative uncharacterized protein 15087 120 5 32 1
23 BC_0063 (yabP) Q81J89 Putative uncharacterized protein 11627
45 1
24 BC_2677 Q81CR9 L-alanyl-D-glutamate peptidase 31439
41 1
25 BC_0047 (yabG) Q81JA4 Sporulation-specific protease YabG
33219
23 1
26 BC_1559(yppG)f Q81FM1 Spore coat protein 22228
23 1
(II) Spore coat proteins likely to play a role in spore resistance
27 BC_4047 (cotα) Q819D8 Putative uncharacterized protein 14344 519 24 680 28 310 18
28 BC_4639 d Q817B8 Thiol peroxidase 18055 158 8 48 2
29 BC_1391 (yqfX) d Q81G20 Putative uncharacterized protein 13457 187 5 69 1
30 BC_2099 (yqfX) d Q81E89 Putative uncharacterized protein 12239 26 1 67 1
31 BC_4774 (yisY) d Q816P9 Non-heme chloroperoxidase 30166 54 1 62 1
(III) Exosporium proteins likely to be involved in attachment to surfaces
32 BC_1218 (exsY) Q81GI1 Spore coat protein Y 16761 640 26 1447 46 2274 106
33 BC_2493 (exsK) Q81D85 Putative uncharacterized protein 13556 1128 26 1372 34 10874 236
Page 34 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 36
35
aNo. a Gene name a Uniprot ID
a Protein description a Mass (Da)
Spore coat & Exosporium
Spore coat Exosporium
bFDR= 1.77% FDR= 1.62% FDR= 1.99% cScore cno.
pept Score no.
pept Score no.
pept
34 BC_3712 (bclC) Q812Y5 Hypothetical Membrane Spanning Protein
75838 673 37 483 21 820 30
35 BC_1221 (bxpB) Q813V0 Exosporium basal layer protein 17390 425 19 448 21 1775 65
36 BC_3547 Q81AL6 Cell surface protein 97913 199 5 713 16 848 32
37 BC_2374 (exsFB) Q813L4 Hypothetical Membrane Spanning Protein
17496 141 6 88 3 164 5
38 BC_2382 (bclB)f Q81DI4 Putative uncharacterized protein 39119
50 1
39 BC_2639 d Q81CV2 Cell surface protein 521640 80 1 29 1
40 BC_3345 Q81B46 Collagen-like triple helix repeat protein
77508
116 2
41 BC_2569 Q81D14 Collagen triple helix repeat protein 53405 73 3 24 1 85 2
42 BC_2149 (bxpA) d Q81E43 Putative uncharacterized protein 32484 47 1 412 8
(IV) Exosporium proteins possibly involved in pathogenicity
43 BC_1284 (inA) e Q81GC3 Immune inhibitor A 85713 920 41
137 5
44 BC_2267 Q81DT6 Putative uncharacterized protein 21872 274 11 433 14 261 10
45 BC_2266 Q81DT7 Putative uncharacterized protein 20239 41 2 144 4 42 2
46 BC_1281 (calY) e Q81GC6 Cell envelope-bound metalloprotease (Camelysin)
21801 409 18
21 1
(V) Spore coat proteins involved in spore germination
47 BC_3607 (yaaH) d Q81AG3 Spore peptidoglycan hydrolase (N-acetylglucosaminidase)
48167 1185 54 1235 41
48 BC_5391 (gerQ) d Q814N4 Putative uncharacterized protein 16090 587 21 227 8
49 BC_0264 (alr1) Q81IT5 Alanine racemase 1 43789 382 22 678 18 1393 47
50 BC_2889 (iunH) Q81C90 Inosine-uridine preferring nucleoside hydrolase
36552 306 11 580 20 1797 61
51 BC_2207 Q81DY9 Sporulation-specific N-acetylmuramoyl-L-alanine amidase
35924 129 8
52 BC_5390 (cwlJ) d Q814N5 Cell wall hydrolase cwlJ 16467 93 5 45 3
53 BC_4319 (gpr) Q818E2 Germination protease 40450 90 3
54 BC_1591 e Q81FJ1 Putative uncharacterized protein 58965 87 2
5360 170
55 BC_3552 (iunH) Q81AL1 Inosine-uridine preferring nucleoside hydrolase
34507
27 1
56 BC_2752 (ypeB) Q813I5 Spore germination protein 50109 81 2
57 BC_2753 (sleB) d P0A3V0 Spore cortex-lytic enzyme 28297 66 2 32 1
(VI) Other putative spore coat and/or exosporium proteins
58 BC_0987 Q81H38 Putative uncharacterized protein 14633 736 33 1800 39 878 30
59 BC_0996 Q81H29 Putative uncharacterized protein 15501 536 23 725 31 366 11
60 BC_5135 (eno) d Q815K8 Enolase 46400 516 16 95 4
61 BC_p0002 d Q814F0 Putative uncharacterized protein 17989 333 11 97 4
62 BC_2858 Q81CB5 Putative uncharacterized protein 9626 266 12 232 7 52 1
63 BC_2426 Q81DE1 Putative uncharacterized protein 26265 263 16 365 21 139 5
64 BC_2026 Q81EF3 Oligopeptide-binding protein oppA 63075 237 11
65 BC_1613 d Q81FH4 Zn-dependent hydrolase 62004 193 8 187 5
66 BC_4410 (yajC) d Q817W7 Protein translocase subunit YajC 9486 132 6 72 2
Page 35 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 37
36
aNo. a Gene name a Uniprot ID
a Protein description a Mass (Da)
Spore coat & Exosporium
Spore coat Exosporium
bFDR= 1.77% FDR= 1.62% FDR= 1.99% cScore cno.
pept Score no.
pept Score no.
pept
67 BC_0337 d Q814A8 Hypothetical Membrane Spanning Protein
14189 126 8 151 9
68 BC_3195 Q813E6 Hypothetical Cytosolic Protein 15025 113 7 74 3 109 5
69 BC_3787 Q81A08 Zinc protease 48972 112 3
70 BC_3786 Q81A09 Zinc protease 49288 112 2
71 BC_3586 Q81AI2 Oligopeptide-binding protein oppA 63759 104 3
72 BC_3133 Q81BM0 Putative hydrolase 42323 98 3
73 BC_1424 Q81FY8 Ferredoxin--nitrite reductase 60770 97 3
74 BC_3986 d Q812V3 Hypothetical Cytosolic Protein 9170 95 3 61 2
75 BC_4480 (tig) d Q812Q9 Trigger factor 47316 88 4 35 1
76 BC_0344 (rocA) d Q81IP0 1-pyrroline-5-carboxylate dehydrogenase
56418 84 4 23 1
77 BC_2375 Q81DJ0 Putative uncharacterized protein 9489 70 2
78 BC_0825 Q81HI3 Putative uncharacterized protein 30284 65 1
79 BC_2745 Q81CL1 Putative uncharacterized protein 43460 62 3
80 BC_1029 d Q81GZ8 IG hypothetical 18063 33466 57 4 25 1
81 BC_3090 d Q81BR0 Putative uncharacterized protein 18565 44 4 48 2
82 BC_5181 d Q815H3 UPF0145 protein BC_5181 11143 42 1 51 1
83 BC_3992 Q819I6 Putative uncharacterized protein 12700 30 1 44 2 22 1
84 BC_2969 Q81C15 Putative uncharacterized protein 12769 30 1
85 BC_2481 Q81D93 Putative uncharacterized protein 12069 28 1
86 BC_1456 Q81FV9 Putative uncharacterized protein 16272 27 2
87 BC_1708 Q81F89 Putative uncharacterized protein 25183 27 1
88 BC_3977 Q819J9 Zn-dependent hydrolase 62055 27 1
89 BC_3515 d Q813A3 Hypothetical Glycosyltransferase 82417 24 1 38 1
90 BC_0395 Q81IJ1 Metal-dependent hydrolase 23560 23 1
91 BC_3784 Q81A11 IG hypothetical 16623 9209
195 5
92 BC_4387 Q817Y7 Putative uncharacterized protein 4832
58 1
93 BC_3582 (yodI) Q81AI6 Putative uncharacterized protein 14601
213 4 25 1
94 BC_1334 Q813U4 Hypothetical Exported Protein 28367
106 1
95 BC_2878 Q81C97 Putative uncharacterized protein 29564
42 2 66 4
96 BC_0263 Q81IT6 Putative uncharacterized protein 38513
42 1
97 BC_1468 (sodF)f Q81FV0 Superoxide dismutase 26798
29 1
98 BC_0944 f Q81H77 Putative uncharacterized protein 61227
28 1
99 BC_2237f Q81DW2 Putative uncharacterized protein 74111
26 1
100 BC_2427 Q81DE0 Putative uncharacterized protein 34256
26 1 aDetails obtained from Uniprot database (www.uniprot.org/). bPeptide False Discovery Rate. cMASCOT MudPIT score & total number of identified peptides over three biological replicates. d Proteins identified only from coat + exosporium & coat fractions. eProteins identified only from coat + exosporium & exosporium fractions. fProteins identified only be semitrypsin search. g
Phosphorylation sites identified by the error-tolerant MASCOT search. None of the other proteins were identified with modifications.
Page 36 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 38
37
Table 2. Identified spore proteins from Clostridium difficile 630.
aNo. a Gene name a Uniprot ID
a Protein description a Mass (Da)
Spore coat
bFDR= 1.72%
cScore cno. pept
(I) Proteins involved in spore coat morphogenesis and other known spore coat proteins
1 CD1067(cdeC)d, e Q18AS2 Putative uncharacterized protein 46845 6392 173
2 CD1581 e Q186D6 Putative uncharacterized protein 20043 2572 103
3 CD1613 (cotA)d, e Q186G8 Putative uncharacterized protein 34834 1680 50
4 CD2401 (cotJC2/cotD) d, e Q181Y5 Spore coat peptide assembly protein CotJC 2 21497 1231 39
5 CD1433 (cotE) d, e Q18BV5 Putative bifunctional protein: peroxiredoxin/chitinase 82019 957 46
6 CD3567 (sipL) e Q181G7 Putative phage cell wall hydrolase 59373 869 44
7 CD0598 (cotJC1/cotCB) d, e Q189E4 Spore-coat protein 21577 823 38
8 CD2400 (cotJB2) e Q181Y6 Spore coat peptide assembly protein CotJB 2 10663 722 16
9 CD0597 (cotJB1/cotCA) d, e Q189E5 Spore coat peptide assembly protein 10756 389 15
10 CD1511 (cotB) d ,e Q18C29 Putative uncharacterized protein 35056 370 16
11 CD0596 (cotF) d, e Q189D6 Putative uncharacterized protein 8844 336 13
12 CD2399 e Q181Y3 Putative uncharacterized protein 8247 169 5
13 CD2598 Q182T7 Putative oligosaccharide deacetylase 29780 70 2
14 CD2629 (spoIVA) e Q182W3 Stage IV sporulation protein A 55539 55 6
15 CD3569 (yabG) Q181G9 Sporulation-specific protease 32137 33 1
16 CD0213 (cotF) Q18CV2 Putative spore coat protein 10927 31 3
(II) Spore coat proteins likely to play a role in spore resistance
17 CD2845 (rbr) e Q183T4 Rubrerythrin 22603 373 12
18 CD2864 (yisY) e Q183V0 Putative hydrolase 30520 243 13
19 CD1524 Q18C45 Putative rubrerythrin 20319 126 9
20 CD1623 Q186H7 Putative oxidoreductase 94987 110 5
21 CD0825 (rbr) e Q18A24 Rubrerythrin 20829 64 3
22 CD0116 Q18CK8 Putative ferredoxin/flavodoxin oxidoreductase,alpha subunit
39290 106 4
23 CD0117 Q18CK7 Putative ferredoxin/flavodoxin oxidoreductase,beta subunit
27031 61 2
24 CD1567 (cotG) d, e Q186C1 Putative manganese catalase 25329 45 3
25 CD0176 Q18CR6 Putative oxidoreductase, NAD/FAD binding subunit 45889 38 2
(III) Exosporium proteins likely to be involved in attachment to surfaces
26 CD0332 (bclA1)e Q18D69 Putative exosporium glycoprotein 68114 85 5
(IV) Exosporium proteins possibly involved in pathogenicity
-- -- -- -- -- -- --
(V) Spore coat proteins involved in spore germination
27 CD0551 (sleC) e Q188Z5 Spore cortex-lytic enzyme pre-pro-form 47426 227 10
28 CD3102 Q184U1 Putative peptidase, M20 family, peptidase V related 53995 29 2
(VI) Other putative spore coat and/or exosporium proteins
29 CD3032 Q184M0 Putative pyridoxal phosphate-dependent transferase 47977 332 18
Page 37 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 39
38
aNo. a Gene name a Uniprot ID
a Protein description a Mass (Da)
Spore coat
bFDR= 1.72%
cScore cno. pept
30 CD3620 Q181M3 Putative uncharacterized protein 14256 232 11
31 CD1133 Q18AZ2 Putative uncharacterized protein 23857 217 10
32 CD3580 Q181I4 Putative uncharacterized protein 29071 157 5
33 CD0855 (oppA) Q18A51 ABC-type transport system, oligopeptide-family extracellular solute-binding protein
58649 126 9
34 CD3664 Q181R8 Putative aminotransferase 44997 119 7
35 CD1536 Q18C55 Ferredoxin--NADP(+) reductase subunit alpha 33353 94 3
36 CD3522 Q181C2 Putative uncharacterized protein 50689 91 4
37 CD1291 (dacF) e Q18BF4 D-alanyl-D-alanine carboxypeptidase 42194 90 3
38 CD2808 Q183P2 Putative uncharacterized protein 23705 87 1
39 CD1463 Q18BY4 Putative uncharacterized protein 17988 68 5
40 CD3652 Q181Q6 Putative peptidase, M1 family 54828 62 5
41 CD3170 (eno) e Q181T5 Enolase 46289 67 3
42 CD0587 Q189C9 Putative uncharacterized protein 10179 55 1
43 CD2865 Q183U9 Putative bacterioferritin 20747 54 3
44 CD3613 Q181L4 Putative uncharacterized protein 17425 42 3
45 CD2431 Q182B6 Putative nitrite/sulphite reductase 58684 40 1
46 CD0115 Q18CK1 Putative 4Fe-4S ferredoxin, iron-sulfur binding domain protein, delta subunit
8140 40 2
47 CD1622 Q186H8 Putative uncharacterized protein 23461 39 1
48 CD3232 Q17ZX7 UPF0597 protein CD630_32320 45787 39 1
49 CD3457 Q180V4 Putative uncharacterized protein 15859 37 2
50 CD1063.1 e Q18AR2 Putative uncharacterized protein 8221 35 2
51 CD0894 Q18A86 Putative iron-dependent hydrogenase 56384 32 1
52 CD2434 Q182C1 Putative uncharacterized protein 22388 30 1
53 CD0279 Q18D22 Putative uncharacterized protein 14922 24 1
54 CD2477 e D5Q2G9 Putative uncharacterized protein 36070 22 1 aDetails obtained from Uniprot database (www.uniprot.org/). bPeptide False Discovery Rate. cMASCOT MudPIT score & total number of identified peptides over three biological replicates. dRenamed in the previous studies25, 34, 44. eProteins identified previously from the whole spore protein extract35.
Page 38 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 40
39
Table 3. Spore coat and/or exosporium proteins identified in our studies from B. subtilis 168, B. cereus ATCC
14579 and C. difficile 630.
Spore coat and/or exosporium proteins identified in our studies and having conserved orthologues (> 30% protein identity) in the
three spore formers. A tick mark (����) indicates experimental identification of the proteins. Proteins CotJC, DacF, SpoIVA and
YisY were identified experimentally from all the three organisms.
Protein Bacillus cereus
ATCC 14579
Clostridium difficile
630
Bacillus subtilis
168
YabG ���� ����
YabP ����
YdbD ���� ����
CotJC ���� ���� ����
CotJB ���� ����
SpoIVA ���� ���� ����
SleB ����
����
RnjA ����
YmfH ����
YmfF ����
DacF ���� ���� ����
Gpr ����
Tig ����
YtfJ ����
����
YtjP
����
YisY ���� ���� ����
Eno ���� ����
CwlJ ����
����
Page 39 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 41
40
Graphic for abstract
Page 40 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 42
Graphical Abstract
43x22mm (300 x 300 DPI)
Page 41 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 43
Fig.1 Venn diagramme of conserved superfamily domains in the identified spore coat & exosporium proteins from B. subtilis 168, B. cereus ATCC 14579 and C. difficile 630. Total 148 superfamily domains were identified in the proteins from three species indicated. The numbers correspond to the number of
superfamily domains identified. The eleven superfamily domains common to the three organisms are listed in the box.
122x42mm (300 x 300 DPI)
Page 42 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 44
Fig.2 Orthologue distribution for identified spore coat & exosporium proteins from B. subtilis 168, B. cereus ATCC 14579 and C. difficile 630. Initial datasets used for orthologue identification include proteins identified
from B. subtilis 16816, 21, B. cereus ATCC 14579 (this study) and C. difficile 630 (this study). The bars
indicate the number of proteins from the respective initial datasets that have orthologues present in the species or groups indicated below. * Number of proteins exclusive for indicated organisms studied in our
works (See Supplementary Tables 2,3 & 4 for details). 18x20mm (600 x 600 DPI)
Page 43 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
Page 45
69x37mm (300 x 300 DPI)
Page 44 of 44
ACS Paragon Plus Environment
Journal of Proteome Research
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960