144
Introduction to Biomechanics Part II

I n t r o d u c t i o n t o
B i o m e c h a n i c s
P a r t I I


E d ito rJohn D . Enderle, University o f Connecticut
In tro d u c tio n to B iom edical E ng ineering : B iom echanics and B ioelectricity - P a rt IDouglas A. Christensen 2009
In tro d u c tio n to B iom edical E ng ineering : B iom echanics and B ioelectricity - P a rt IIDouglas A. Christensen 2009
Basic Feedback C ontro ls in B iom edicineCharles S. Lessard 2008
U nderstand ing A tria l F ib rilla tion : T h e S ignal P rocessing C o n trib u tio n , P a rt ILuca Mainardi, Leif Sornmo, and Sergio Cerutti 2008
U nderstand ing A tria l F ib rilla tion : T h e S ignal P rocessing C o n trib u tio n , P a rt IILuca Mainardi, Leif Sornmo, and Sergio Cerutti 2008
In tro d u c to ry M edical Im agingA. A. Bharath 2008
L u n g Sounds: A n Advanced S ignal P rocessing PerspectiveLeontios T. Hadiileontiadis 2008
A n O u tlin e o f In fo rm atio n G eneticsGerard Battail 2008

iv
N eural In terfacing: Forg ing th e H u m an -M ach in e C onnectionThomas D. Coates, Jr.2008
Q u an tita tiv e N europhysiologyJoseph V. Tranquillo 2008
T rem or: From P athogenesis to T rea tm en tGiuliana Grimaldi and Mario Manto 2008
In tro d u c tio n to C o n tinuum B iom echanicsKyriacos A. Athanasiou and Roman M. Natoli 2008
T h e E ffects o f H ypergravity and M icrogravity on B iom edical E xperim entsThais Russomano, Gustavo Dalmarco, and Felipe Prehn Falcao 2008
A B iosystem s A pproach to In d u stria l P a tien t M o n ito rin g and D iagnostic D evicesGail Baura 2008
M ultim odal Im ag ing in N eurology: Special Focus on M R I A pplications and M E GHans-Peter Muller and Jan Kassubek 2007
E stim atio n o f C ortical C onnectiv ity in H um ans: A dvanced S ignal Processing T echniquesLaura Astolfi and Fabio Babiloni 2007
B rain -M achine In terface E ng ineeringJustin C. Sanchez and Jose C. Principe 2007
In tro d u c tio n to S tatistics fo r B iom edical E ngineersKristina M. Ropella 2007
C apstone D esign Courses: P roducing Industry -R eady B iom edical E ngineersJay R. Goldberg 2007
B ioN anotechnologyElisabeth S. Papazoglou and Aravind Parthasarathy 2007

V
B io instrum en ta tionJohn D. Enderle 2006
F undam entals o f R esp irato ry Sounds and A nalysisZahra Moussavi 2006
A dvanced P ro b ab ility T h eo ry for B iom edical E ngineersJohn D. Enderle, David C. Farden, and Daniel J. Krause 2006
In term ed ia te P ro b ab ility T h eo ry fo r B iom edical E ngineersJohn D. Enderle, David C. Farden, and Daniel J. Krause 2006
Basic P ro b ab ility T h eo ry for B iom edical E ngineersJohn D. Enderle, David C. Farden, and Daniel J. Krause 2006
Sensory O rg an R eplacem ent and R epairGerald E. Miller 2006
A rtificial O rgansGerald E. Miller 2006
Signal P rocessing o f R andom Physiological SignalsCharles S. Lessard 2006
Im age and Signal P rocessing for N etw orked E -H ea lth A pplicationsIlias G. Maglogiannis, Kostas Karpouzis, and Manolis Wallace 2006

Copyright © 2009 by Morgan & Claypool
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means—electronic, mechanical, photocopy, recording, or any other except for brief quotations in printed reviews, without the prior permission of the publisher.
Introduction to Biomedical Engineering: Biomechanics and Bioelectricity - Part II
Douglas A. Christensen
www.morganclaypool.com
ISBN: 9781598298444 paperback ISBN: 9781598298468 ebook
DOI 10.22 00/S000183ED1V01Y200903BME029
A Publication in the Morgan & Claypool Publishers series SYNTHESIS LECTURES ON BIOMEDICAL ENGINEERING
Lecture #29Series Editor: John D. Enderle, University of Connecticut
Series ISSNSynthesis Lectures on Biomedical Engineering Print 1932-0328 Electronic 1932-0336

I n t r o d u c t i o n t o
B i o m e c h a n i c s
P a r t I I
D oug las A . C h ris ten senUniversity of Utah
SYNTHESIS LECTURES ON BIOMEDICAL ENGINEERING #29
M O R G A N C L A Y P O O L P U B L I S H E R S

ABSTRACTIntended as an introduction to the field of biomedical engineering, this book covers the topics of biomechanics (Part I) and bioelectricity (Part II). Each chapter emphasizes a fundamental principle or law, such as Darcy’s Law, Poiseuille’s Law, Hooke’s Law, Starling’s Law, levers and work in the area of fluid, solid, and cardiovascular biomechanics. In addition, electrical laws and analysis tools are introduced, including Ohm ’s Law, Kirchhoff’s Laws, Coulomb’s Law, capacitors and the fluid/electrical analogy. Culminating the electrical portion are chapters covering Nernst and membrane potentials and Fourier transforms. Examples are solved throughout the book and problems with answers are given at the end of each chapter. A semester-long Major Project that models the human systemic cardiovascular system, utilizing both aM atlab numerical simulation and an electrical analog circuit, ties many of the book’s concepts together.
KEYWORDSbiomedical engineering, biomechanics, cardiovascular, bioelectricity, modeling, Matlab

T o L a r a i n e


xi
C o n t e n t s
Synthesis Lectures on Biomedical Engineering.........................................................................ill
C ontents............................................................................................................................................. xi
Preface............................................................................................................................................... xv
g Ohm ’s Law: Current, Voltage and Resistance............................................................................ 1
8. 1 In troduction ......................................................................................................................... 1
8.1.1 Charge 2
8.1.2 Electric Field 3
8.1.3 Current 3
8.1.4 Voltage 4
8. 2 Ohm ’s L a w ........................................................................................................................... 5
8 .2. 1 Fluid Analogies 6
8.3 Sign Conventions for Voltage and C u rren t.....................................................................7
8.3.1 Resistivity of Bulk Materials 8
8.4 Diodes and O ther Non-Ohmic Circuit E lem ents.........................................................9
8.5 Power Loss in Resistors.................................................................................................... 10
8. 6 P roblem s............................................................................................................................. 11
9 Kirchhoff’s Voltage and Current Laws: Circuit A nalysis.......................................................13
9.1 Introduction........................................................................................................................13
9.2 Kirchhoff’s Voltage Law (KVL)...................................................................................... 15
9.3 Kirchhoff’s Current Law (K C L ).................................................................................... 15
9.4 Resistive Circuit Analysis Using the Branch Current M eth o d ................................. 16
9.5 Problem s............................................................................................................................. 21
1 0 Operational Amplifiers..................................................................................................................25

xii CONTENTS
10.1 Introduction........................................................................................................................25
10. 2 Operational Amplifiers...................................................................................................... 26
10.3 Dependent Sources............................................................................................................29
10.4 Some Standard Op Amp C ircuits.................................................................................. 30
10.4.1 Inverting Amplifier 31
10.4.2Noninverting Amplifier 33
10.4.3 Voltage Follower 34
10.5 Problem s..............................................................................................................................36
Coulomb’s Law, Capacitors and the Fluid/Electrical A nalogy............................................. 39
11. 1 Coulomb’s Law ....................................................................................................................39
11. 2 Capacitors............................................................................................................................40
11.3 Flow Into and O ut of Capacitors.................................................................................... 42
11.4 Analogy Between Fluid and Electrical C ircuits...........................................................43
11.4.1 Scaling the Analog Pairs 45
11.5 Problem s..............................................................................................................................46
1 2 Series and Parallel Combinations of Resistors and Capacitors............................................. 49
12.1 Introduction........................................................................................................................49
12.2 Resistors in Series..............................................................................................................49
12.3 Resistors in Parallel............................................................................................................50
12.4 Capacitors in Series............................................................................................................52
12.5 Capacitors in Parallel........................................................................................................54
12.6 Voltage D iv ider..................................................................................................................56
12.7 Current D ivider..................................................................................................................57
12.8 Problem s..............................................................................................................................59
13 Thevenin Equivalent Circuits and First-Order (RC) Time C onstants............................... 61
13.1 Thevenin Equivalent C ircuits.......................................................................................... 61
13.2 Electrical Behavior of Capacitors.................................................................................... 63
13.3 RC Time C onstan ts..........................................................................................................65
13.4 Problem s..............................................................................................................................70

CONTENTS xiii1 4 Nernst Potential: Cell Membrane Equivalent C ircu it.............................................................73
14.1 In troduction........................................................................................................................73
14.2 Cell Membrane S tructure................................................................................................ 73
14.2.1 Mechanisms of Transport 75
14.3 Nernst Potential..................................................................................................................76
14.4 Equivalent Circuit for the M em brane.............................................................................78
14.5 Action Potentials................................................................................................................84
14.6 Problem s..............................................................................................................................88
15 Fourier Transforms: Alternating Currents (AC) and the Frequency D o m ain ....................91 I
15.1 Introduction - S inusoids..................................................................................................91
15.2 Fourier Series for Arbitrary Repeating W aveforms..................................................... 92
15.3 FFT: Calculating the Discrete Fourier Transform .......................................................94
15.4 Problem s........................................................................................................................... 101
Q Major P roject............................................................................................................................... 105
C .l Background (and Preliminaries).................................................................................... 105
C .1. 1 Modeling the Cardiovascular System 105
C.1.2 Overview of Major Project 106
C .l .3 Notebook Requirements 106
C .l .4 Grading and Checkoff Dates 108
C .l .5 O ur Model 110
C .l .6 Approximations 112
C .l .7 Calculation of R and C Values 113
C.2 MATLAB M o d el............................................................................................................115
C.3 Electrical Circuit A nalog................................................................................................ 119
B ibliography....................................................................................................................................... 127


P r e f a c e
N O T E O N O R G A N IZ A T IO N O F T H IS B O O KThe material in this book naturally divides into two parts:
1. Part I: Chapters 1-7 cover fundamental biomechanics laws, including fluid, cardiovascular, and solid topics (1/ 2 semester).
2, Part II: Chapters 8-15 cover bioelectricity concepts, including circuit analysis, cell potentials, and Fourier topics (1/2 semester).
A Major Project accompanies the book to provide laboratory experience. It also can be divided into two parts, each corresponding to the respective two parts of the book. For a full-semester course, both parts of the book are covered and both parts of the Major Project are combined.
The chapters in this book are support material for an introductory class in biomedical engineering1. They are intended to cover basic biomechanical and bioelectrical concepts in the field of bioengineering. Coverage of other areas in bioengineering, such as biochemistry, biomaterials and genetics, is left to a companion course. The chapters in this book are organized around several fundamental laws and principles underlying the biomechanical and bioelectrical foundations of bioengineering. Each chapter generally begins with a motivational introduction, and then the relevant principle or law is described followed by some examples of its use. Each chapter takes about one week to cover in a semester-long course; homework is normally given in weekly assignments coordinated with the lectures.
The level of this material is aimed at first-semester university students with good high-school preparation in math, physics and chemistry, but with little coursework experience beyond high school. Therefore, the depth of explanation and sophistication of the mathematics in these chapters is, of necessity, limited to that appropriate for entering freshman. Calculus is not required (though it is a class often taken concurrently); where needed, finite-difference forms of the time- and space-varying functions are used. Deeper and broader coverage is expected to be given in later classes dealing with many of the same topics.
Matlab is used as a computational aid in some of the examples in this book. W here used, it is assumed that the student has had some introduction to Matlab either from another source or from a couple of lectures in this class. In the first half of the cardiovascular Major Project discussed below,
1At the University of Utah, this course is entitled Bioen 1101, Fundamentals of Bioengineering I.

xvi PREFACE
Matlab is used extensively; therefore, the specific Matlab commands needed for this Major Project must be covered in class or in the lab if this particular part of the project is implemented.
A Major Project accompanies these chapters at the end of the book. The purpose of the Major Project, a semester-long comprehensive lab project, is to tie the various laws and principles together and to illustrate their application to a real-world bioengineering/physiology situation. The Major Project models the human systemic cardiovascular system. The first part of the problem takes approximately one-half of a semester to complete; it uses Matlab for computer modeling the flow and pressure waveforms around the systemic circulation. Finite-difference forms of the flow/pressure relationships for a lumped-element model are combined with conservation of flow equations, which are then iterated over successive cardiac cycles. The second half of the problem engages a physical electrical circuit to analyze the same lumped-element model and exploits the duality of fluid/electrical quantities to obtain similar waveforms to the first part. This Major Project covers about 80% of the topics from the chapter lectures; the lectures are given "just-in-time" before the usage of the concepts in the Major Project.
Although the Major Project included with this book deals with the cardiovascular system, other Major Project topics m aybe conceived and substituted instead. Examples include modeling human respiratory mechanics, the auditory system, human gait or balance, or action potentials in nerve cells. These projects could be either full- or half-semester assignments.
ACKNOWLEDGEMENTSThe overarching organizational framework of these chapters around fundamental laws and principles was conceived and encouraged by Richard Rabbitt of the Bioengineering Department at the University of Utah. Dr. Rabbitt also provided much of the background material and organization of Chapters 2 and 4. Angela Yamauchi provided the organization and concepts for Chapter 3. David Warren contributed to the initial organization of Chapter 8 . Their input and help was vital to the completion of this book.
Douglas A. Christensen University of Utah March 2009

C H A P T E R 8
a n d R e s i s t a n c e8.1 INTRODUCTIONBiological organisms rely upon myriads of electrical activities for proper functioning. These electrical processes occur continuously and are vital to life. For example, ion (charged particle) movement is responsible for all signal transmission along nerves and for all muscle contraction. Right now, as you read these words, several billions of ions are being rapidly transported back and forth across the cell membranes of the neurons in your retina, optic nerve, and brain, not to mention your heart muscle, kidney, blood vessels and all other cellular tissue. Ions commonly involved in biological activity include calcium (Ca++), potassium (K+), sodium (Na+), chloride (Cl- ) and bicarbonate (H C O 3 ). These ions flow through the fluid environment both inside and outside the cells, as shown in stylized form in Fig. 8.1.
Figure 8.1: Some common ions found in the fluid inside and outside biological cells.
A major component1 of the force causing these ions to move (or not move) across the cell wall is electrostatic, that is, the force produced on a charged particle by the presence of other nearby charged particles, some having the same electrical sign, some with opposite sign. The charge of a single electron is q = —1.60 x 10_19 C, where C is the symbol for coulomb, the SI unit of charge. The sign of the electron is negative, thanks in large part to choice by Benjamin Franklin. The charge of an ion is determined by how many electrons are missing from its atomic shell (a deficit of
'This is not the only component ot torce. Another important torce on the ions is a dittusional torce produced by concentration differences. The balancing of the electro-diffusional forces is covered in Chapter 14.

2 CHAPTER 8. OHM’S LAW: CURRENT, VOLTAGE AND RESI STANCE
electrons, giving a positive sign to the net charge) or are added to the atomic shell (an excess, giving a negative sign to the net charge). For example, since a sodium ion (Na+ ) is missing one electron, its charge is +1.60 x 10-19 C. The charge of a chloride ion (Cl- ), which has one excess electron, is —1.60 x 10-19 C, and the charge of a calcium ion (Ca++), which is missing two electrons, is +3.20 x 10“ 19 C.
Bare, mobile electrons also occur in nature (although not to any great extent in biological tissues). Mobile electrons are present in large concentrations only in metals and semiconductors. In these materials, they flow as electron currents and are an essential part of modern electronic devices and circuits. In general, an electric circuit is composed of a source and one or more elements connected together by metallic wires and leads (usually copper, but sometimes gold, silver or aluminum) to form a closed path. Figure 8.2 is an example of a simple circuit. The Major Project that models the human systemic circulation uses an analog electric circuit in its second half.
Figure 8.2: Example of an electric circuit consisting of a source, resistor and diode connected in a closed path by wires (assumed ideal, with zero resistance).
To quantitatively describe ion (and electron) movement, we first need to define the concepts of charge, electric field, current and voltage.
8.1.1 CHARGEFigure 8.3 shows two charged particles, <71 and <72 (both positive in this example) in close proximity to each other. It is well known that like charges repel and unlike charges attract. This phenomenon
1
2 2 force F that it experiences due to the presence of <71 is directed away from <71 (since <71 is positive);
1 22
is an equation relating these various quantities, and will be covered in more detail in Chapter 11. As mentioned earlier, the SI unit of charge is the coulomb, with symbol C.

8.1. INTRODUCTION 3
F
\
X t /
path/ %
E ^ ^ OX + q ,
F ^ — O " 1
+ q 2
Two charges q \ andg2 of the same sign repel. Thus, each charge experiences a force F (which can also be represented by an electric field E). As the charge q2 moves, it produces a current i and it may also change its potential energy, or voltage.
8.1.2 E L E C T R IC F IE L DThe existence of the force F on charge q 2 due to charge q \ can be represented by an electric field E at the position of q 2 , whose direction is the same as the force F (thus i i is a vector, with direction and magnitude) and whose magnitude is given by E = F f q 2 - Because of the division by q 2 , the magnitude of E is not dependent on the size of the test charge q 2 , but only on the size and sign of the charge q \ (and also inversely upon the distance between the charges, as will be evident when we cover Coulomb’s Law in Chapter 11). The electric field is a conservative field, as defined in Section 7.4.2 in Part I.
8.1.3 C U R R E N TNow suppose that the charge q 2 in Fig. 8.3 moves along the path shown by the arrows from the lower left to the upper right. The movement of charge produces a current (denoted by the symbol i). Current is the measure of the rate of flow of charge, and it has the SI uni t of ampere (A). I f an amount of charge Aq passes a certain point such as X during time A t , the current i is given by
i = A q / A t . (8.1)
In the limit as A t approaches zero, (8.1) becomes the differential equation
i = d q / d t . (8.2 )
By convention, electrons flow in the opposite direction to the flow of positive current, since the electron has a negative charge. This may be initially confusing, but it will become more comfortable with experience. For ions, positive current flows in the same direction as the flow of positively charged ions, but it flows in the opposite direction of negatively charged ions.

4 CHAPTER 8. OHM’S LAW: CURRENT, VOLTAGE AND RESI STANCE
8.1.4 V O LT A G EAs the charge <72 moves, it may move closer to q i (or further away). I f so, its movement is against (or with) the force F. As we have seen in the previous chapter, work— a change in energy— is equal to
21
force. The former case is shown in the example of Fig. 8.3. This form of energy is properly called2 1
energy into kinetic energy.I f we divide the potential energy of the charged particle by its charge, we obtain the voltage2
i'. Thus,
2
where E p is the charge’s potential energy at any given position. Voltage is the amount of work needed to move a unit charge between two points. A large voltage represents the potential to do a large amount of workJ . The SI unit of voltage is the volt, with symbol V. From (8.3) it can be seen that the unit of volt (V) is equivalent to a joule per coulomb (J/C).
21
relative-, often in circuits, one point on the circuit is chosen to be at zero voltage, and the voltages at all other points are measured in reference to this point.
tvoltage V
[VI
position
The voltage (proportional to potential energy) of the charge q2 in Fig. 8.3 as it moves along the path. Its voltage increases because it moves closer to q \.
2Voltage is sometimes referred to as “potential,” or “electrical potential,” which is understandable since it is related to potential energy.
3 But remember, voltage is work per unit charge. So a large amount of charge must flow (i.e., a large current) in order to achieve this amount of work. As we will see later, power equals voltage multiplied by current.

8.2. O H M ’S LAW 5
8.2 OHM’S LAWFigure 8.5 shows a particularly simple electrical circuit consisting of a voltage source connected across a single resistor with ideal wires. To a very good approximation, the current i flowing through the resistor is related to the voltage drop v across the resistor in a linear manner:
i R O hm ’s law (8.4)
where the proportionality constant is the resistance R of the resistor. This linear relationship between current and voltage is known as Ohm’s Law, named after German physicist Georg Simon Ohm. It holds for resistors in circuits, for resistive conductors and wires, and for the flow of electrolytes through solution, and even is used to help describe the electrical behavior of the cell wall (where the current is carried by ions)4. Any device or material that obeys Ohm ’s Law is termed “ohmic.”
, ideal w ire
Figure 8.5: Simple electrical circuit demonstrating Ohm’s Law for the resistor R.
W hen a plot is made of the voltage across an ohmic element as a function of the current through it, the linear relationship is obvious; see Fig. 8 .6 . Note that the line passes through the origin; that is, when the current reverses sign, the voltage also reverses sign. Also, if the current is zero, the voltage is zero. The slope of the line is constant and has a value equal to the resistance R of the element. R is always a positive number for passive elements like resistors. R has the SI unit of ohms, with the Greek symbol f2. From (8.4) it can be seen that an SI is equivalent to V / A. In some instances (for example, an ideal wire), the resistance is assumed to have a value of zero (a short circuit). On the other hand, when there is no conductive path at all between two points, the value of the resistance is infinite (an open circuit) and no current can flow.
Occasionally, the reciprocal of resistance, called the conductance G, is used instead of resistance. The SI unit of conductance is the siemen, or S. Conductance and resistance are related by
1 (8.5)
4 A useful electrical model of a cell membrane includes a capacitor in parallel with several resistors and voltage sources. The values of the resistors change dramatically as ion channels open and close, causing voltage spikes (action potentials) to propagate down nerves. This model is covered in Chapter 14.

6 CHAPTER 8. OHM’S LAW: CURRENT, VOLTAGE AND RESI STANCE
Figure 8.6: Voltage-current relationship for an ohmic element.
8.2.1 F L U ID A N A L O G IE SBy now, it should be obvious that there is a close analogy between fluid flow (where the flowing particles are molecules of fluid) and electrical current (where the flowing particles are electrons or ions). In fact, the equations below describing the linear relationship between the driving force and the flow rate are of exactly the same form for both fluids and electrical current.
* For fluid flow though a porous membrane (Chapter 2, Part I), the driving force is the pressure difference A P across the membrane, and the flow is the fluid passing through the membrane. Darcy’s Law,
A P = Q R , (2.5)
states that the pressure is proportional to the flow, with the proportionality constant called the hydraulic or fluid resistance R , given by (2.5).
* For fluid flow through a tube (Chapter 3, Part I), the driving force is the pressure difference A P between the two ends of the tube, and the flow is fluid flow through the tube. Poiseuille’s Law,
A P = Q R , (3.7)
shows that with laminar flow conditions, the pressure is again proportional to flow; the proportionality constant is similarly called hydraulic or fluid resistance R , given by (3.7).
* For electrical current (this chapter), Ohm ’s Law,
v = i R, (8.4)

8.3. SIGN CONVENTIONS FOR VOLTAGE AND CURRENT 7
states that the voltage drop across a resistive element is proportional to the electrical current, with the proportionality constant given by the resistance R .
Note the similar form of all three equations above. O f course the quantities are different, but their mathematical behavior is identical. Conservation laws also apply to both the conservation of fluid molecules in a fluid network and the conservation of electrical charge in an electrical circuit. This is the basis for using an electrical circuit as an analogy for a fluid circuit. In particular, in the second half of the Major Project that models the human systemic circulatory system, an electrical circuit is used to model the pressures and the blood flow.
8.3 SIGN CONVENTIONS FOR VOLTAGE AND CURRENTW hen doing an analysis of the resistors in circuits or resistive elements in biological models, it is important to keep accurate track of the signs of both the current and the voltage across the resistor. Figure 8.7 shows the symbol for a resistor used in schematic diagrams. A current i is assumed to flow through the resistor. The reference direction for positive current is the same as the direction of the arrow, shown to the side of the symbol. The voltage v across the resistor is given by the difference
1 2symbol A v should be used instead of v, but this becomes cumbersome after a while, so is usually shortened to just v.)
f
i ------ ►
Figure 8.7: Circuit symbol for a resistor o f value R. The order of the signs o f v and the direction o f i is given by the passive sign convention.
The order of the signs in Fig. 8.7 follows the passive sign convention (psc) which states that the positive sign fo r the voltage reference is p u t on the side o f the resistor where the positive current enters. We will always follow the passive sign convention in this book.
Although the arrow points from left to right in Fig. 8.7, that does not mean that the current i always flows in that direction. It merely sets the reference direction of flow for positive current; that is, when the sign of the current i is positive, it flows in the direction of the arrow, or from left to right in Fig. 8.7 (and— hold on— electrons flow from right to left). But when the current i has a negative value, it flows in the opposite direction of the arrow, or from right to left in Fig. 8.7 (and—you guessed it— electrons flow from left to right).
Similarly, the voltage v can have either a positive or negative value. I f v has a positive value,1 2
On the other hand, if v has a negative value, the voltage on the left is less than the voltage on the
\■ W

8 CHAPTER 8. OHM’S LAW: CURRENT, VOLTAGE AND RESI STANCE
1 2passive sign convention, when i is positive, v is also positive, and vice versa.
The direction initially chosen for the current arrow is usually arbitrary when setting up a circuit analysis. In every case, however, the passive sign convention—which relates the order of the signs of v to the direction of i— must be used in setting up the circuit references. For example, Fig. 8. 8 shows a case where the positive current direction has arbitrarily been chosen to be from right to left. Note that the order of the signs for v in Fig. 8. 8 has been reversed from Fig. 8.7 in order to follow the passive sign convention. Although the direction of the positive current arrow is often arbitrary, once that direction has been chosen, the order of the voltage signs must follow the passive sign convention!
v2 ~ V + V’l
M AR
Figure 8.8: The direction of the positive current flow through the resistor R has been reversed compared to Fig. 8.7. The order of the signs of v has also been reversed to follow the passive sign convention.
8.3.1 R E S IS T IV IT Y O F B U L K M A T E R IA L SIn the previous discussions, the resistive device was considered as a whole, possessing a net resistance R . In circumstances where microscopic behavior is o f more interest, it is useful to consider the resistive properties of a small incremental volume of the material making up the larger device. This property is called the resistivity p of the material, and it is specific to the material being considered, not to the particular geometry or size of the device. The SI unit of resistivity is ohm-meter (f2-m). This quantity is especially useful when analyzing bulk materials such as ionic solutions or insulating layers. An insulator such as quartz has a resistivity of about p = 1 0 x 10+17 f2-m. Normal saline
0 20p = 1.7 x 10- 8 f2-m.
For elements with certain common geometries, there exists a simple relationship between the bulk resistivity p (a material property) and the overall resistance R of the element. For example, if the cylindrical resistor shown in Fig. 8.9 is composed of a homogeneous material with resistivity p , the net resistance of the resistor is given by6
R = p i / A . (8.6)
Remember that R is always a positive number for resistors.6To practice consistency checking, use a units check and a ranging check on (8.6) to confirm that it has the correct form.

8.4. DIODES AND OTHER NON-OHMIC CIRCUIT ELEMENTS 9
Figure 8.9: A cylinder made from material with a bulk resistivity of p.
Sometimes the reciprocal of resistivity, called the conductivity a , is used. The two quantities are related by
or = 1 /p . (8.7)
The SI units of conductivity are siemen per meter, or S/m.
8.4 DIODES AND OTHER NON-OHMIC CIRCUIT ELEMENTS
Although many electrical elements follow Ohm’s Law at least over some range of their operation, other elements do not. Some circuit elements, such as the diode, are purposely made to behave in a non-ohmic manner.
The schematic symbol for a diode is shown in Fig. 8.10. Note that the voltage signs and current direction are set up to follow the passive sign convention.
anode ca thode
+ v
--------- ► i
Figure 8.10: The symbol for a diode. It allows easy current flow in the direction of the arrow, but blocks flow in the opposite direction.
A diode is used as a one-way “valve” of current. It allows easy flow in the direction of the arrow (the “forward” direction, from anode to cathode) when the voltage v is positive (i.e., when the voltage on the left side in Fig. 8.10, or anode side, is more positive than on the right side, or cathode side). But the diode is highly resistant to flow in the opposite direction (the “reverse” direction) when

10 CHAPTER 8. OHM’S LAW: CURRENT, VOLTAGE AND RESISTANCE
the voltage v is negative (i.e., when the right side has greater voltage than the left side in Fig. 8.10). This asymmetric current-voltage behavior is plotted in Fig. 8.11(a) for a typical diode. Note that the current is large and positive when v is positive, equivalent to a low forward resistance, but that the current is small and negative when v is negative, equivalent to a high reverse resistance.
a. t ♦ 1 b. 1 ♦current i
[Al | actual I / diode
current i [Al ideal
/ model
0 + voltage v [V] — ► 0 ,
voltage v
Figure 8.11: (a) The current-voltage relationship for a typical diode; (b) the idealized model of a diode, consisting of a short circuit in the forward direction and an open circuit in the reverse direction.
Often an ideal model of a diode is employed in circuit analysis. This makes the analysis considerably simpler. In the ideal model, it is assumed that in the forward direction, the diode possesses zero resistance (a short circuit); this means that there is no voltage drop across the diode regardless of the amount of current through it. However, in the reverse direction, the diode acts like an open circuit with infinite resistance, and no current flows regardless of the amount of voltage. This ideal model behavior is shown in the current-voltage plot of Fig. 8.11(b).
Several other important circuit elements are also non-ohmic. The transistor and the operational amplifier are examples of active devices that have gain and do not follow Ohm ’s Law. The operational amplifier is covered in Chapter 10.
8.5 POWER LOSS IN RESISTORSTo pass a current i through a resistor of value R requires an electrical force, measured by the voltage drop v across the resistor. Analogous to the formula for power loss in fluids [see Equation (3.9)], the electrical power loss in a resistor is given by the product of current and voltage:
P = iv , (8 .8)
in SI units of watts (W ). Equivalent expressions can be obtained by using Ohm ’s Law (8.4) in (8 .8):
P = i 2R = v2/ R . (8.9)

8.6 PROBLEMS8.1. A tungsten resistance wire is 100 m long with around cross section and a diameter of 0.20 mm.
W hen a variable voltage is applied between its ends, a current is measured as plotted below.
a. W hat is the resistance R of this wire?
[ans: R = 180 fi]
b. W hat is the resistivity p of this wire?
fans: p = 5.7 x 10_8fi • m]
8.6. PROBLEMS 11
voltage v (V)
Figure 8.12: Plot of current-voltage relastionship for wire analyzed in Problem 8.1.


13
C H A P T E R 9
K i r c h h o f f ’s
L a w s :
9.1 INTRODUCTION
e a n d C u r r e n t
s i s
As we have seen, fluid networks consist mainly of tubes, pipes and valves connected together and to pressure sources. The tubes have various degrees of resistance, and sometimes the tubes have compliance. An excellent example of a fluid network in biology is the cardiovascular system. Similarly, electrical networks— electrical circuits— are also composed of elements connected together. In this case, the basic elements are resistors, capacitors (discussed later), inductors (also discussed later), and voltage or current sources. We will start with the three basic elements whose symbols are shown in Fig. 9.1.
resistor independent
RV ( + ] voltage
s source
independent I ( A ) current
source
Figure 9.1: The symbols for three basic electrical elements.
The resistor’s symbol and its ohmic voltage/current behavior have already been introduced in Ohm ’s Law unit. The independent voltage source is a source whose voltage Vs across its terminals is always the same regardless of the current through the source (it is therefore called a “constant- voltage” source). This is an ideal model, never really found in practice, but batteries and voltage power supplies come close to this behavior for low to moderate currents. The independent current source is a mirror image of the voltage source: the current / s out of this source is always the same regardless of the voltage across its terminals (a “constant-current” source). Again this is an ideal model and is only an approximation of real current sources.
For every fluid network model, there is an equivalent electrical network model. For example, two very simple fluid networks are shown in Fig. 9.2 along with their electrical equivalents. The wide tubes are assumed to have negligible resistance, as are the wires.

14 CHAPTER 9. KIRCHHOFF’S VOLTAGE AND CURRENT LAWS: CIRCUIT ANALYSIS
FLUID ELECTRICAL
node 1
equiv to indep.
source
node 1
Figure 9.2: Two simple fluid networks on the left with their equivalent electrical circuits on the right.
The fluid network in the upper part of Fig. 9.2 is driven by a constant-pressure pump, such as an air-diaphragm pump; for this type of pump, Ps is constant but Q is not. Thus, if the resistance R of the narrow-tube section increases somehow, the flow rate Q will go down. Its electrical equivalent on the right, using an independent voltage source, shows the same behavior. The voltage Vs is always constant. The voltage is the same across both the source and the resistor, and the same current i goes through both the source and the resistor. By Ohm ’s Law, if the resistance R increases, the current i will decrease.
The fluid network in the lower part of Fig. 9.2 is driven by a constant-flow pump, such as a roller pump. The flow rate Q s out of this source is constant, but the pressure across it is not. (Although more complex than constant-pressure pumps, a constant-flow pump is often used in hydraulic elevators, where the rate of ascent and descent needs to be constant regardless of the load inside the elevator.) I f the resistance of the narrow-tube section increases, the pressure drop across the tube (and correspondingly the pump) goes up. The electrical equivalent, which uses an independent

9.2. KIRCHHOFF’S VOLTAGE LAW (KVL) IS
current source, has this same behavior. The voltage v is the same across both the source and the resistor, and the same current goes through both the source and the resistor.The current I s is always constant. By Ohm ’s Law, if the resistance R increases, the voltage v will increase.
9.2 KIRCHHOFF’S VOLTAGE LAW (KVL)Kirchhoff’s Voltage Law applies to all electrical circuits. I t states:
The algebraic sum o f voltages around any closed pa th (loop) is zero.
The application of this principle to circuits is straightforward. Going around any loop while summing its voltages is called taking a “Kirclilioff s T our” (K T).The tricky part is keeping track of the signs. To help with getting the signs correct, we must always use the passive sign convention (psc) for resistors (see Ohm ’s Law in Chapter 8), and then follow Rule 1:
Rule 1 - W hen using KVL around a loop, the sign of the voltage contribution across any element is the sign first encountered in the direction of the tour.
L et’s apply KVL to a simple example. In the upper-right electrical circuit of Fig. 9.2, start the KT at the part of the circuit labeled node 1. Go clockwise around the loop summing voltage contributions from each element (only two in this case) until you get back to the starting point. The first voltage crossed is +u; it is positive since the + sign was encountered first when going in a clockwise direction. The next voltage term is — V̂ ; it is negative because the — sign was encountered first. T hat gets us back to node 1, the starting point. KVL says that the sum of these voltages is zero, or
+ v - Vs = 0. (9.1)
Thus, v = V;, as is perhaps obvious (and was already anticipated in the figure labels). W e’ll use KVL for more complex examples later.
9.3 KIRCHHOFF’S CURRENT IAW (KCL)This law also applies to all electrical circuits. I t states:
The algebraic sum o f currents a t any node is zero-or- The currents entering any node equal the currents leaving.
The strict definition of a node is any point at which two or more circuit elements join, although in awhile we will be concerned only with nodes where more than two elements join (called essential nodes). In fluids, nodes are called junctions. Note that KCL is just a statement of the conservation of mass, identical to the rule for incompressible fluid volumetric flow. In fluids, the mass is composed of fluid molecules; in the electrical case, the mass is that of charged particles.
Once again, the application of KCL is straightforward as long as the signs of each current are accounted for correctly. L et’s apply KCL to the simple example shown in the lower-right part of

16 CHAPTER 9. KIRCHHOFF’S VOLTAGE AND CURRENT LAWS: CIRCUIT ANALYSIS
Fig. 9.2. At the (nonessential) node labeled node 1, the current entering is / v.T he current leaving isi. Using KCL,
/ = (9.2)
as is again perhaps intuitive (and already labeled as such in Fig. 9.2).
Example 9.1. Four-W ire NodeAnother example of KCL, applied to an essential node with four connections, is shown in Fig. 9.3.
Here KCL gives
i\ + 1 3 = 12 + 14,
' l + '3 - '2 - '4 = 0 . (9.3)
Figure 9.3: KCL applied to an essential node with four connections.
9.4 RESISTIVE CIRCUIT ANALYSIS USING THE BRANCH CURRENT METHOD
A somewhat more complex circuit is shown in Fig. 9.4. Kirchhoff’s Laws and Ohm ’s Law can be applied to solve for the voltages and currents in all elements of this circuit by one of three possible circuit analysis methods. Here we will use the Branch C urrent M ethod (the other two methods will be left for later classes). The circuit of Fig. 9.4 has two essential nodes (i.e., nodes with more than two connections) labeled node 1 and node 2. A branch is defined as a single path that connects one essential node to another. In this circuit there are three branches connecting essential node 1 to essential node 2 .

9.4. RESISTIVE CIRCUIT ANALYSIS USING THE BRANCH CURRENT METHOD 17
Figure 9.4: Example of a circuit to be analyzed by the Branch Current Method.
The Branch C urren t M ethod has four steps:
Step 1. Assign a current to each branch that does not have a current source. Give the current a name (using i symbols) and a direction. You can choose each direction arbitrarily. (Exception: for branches which contain a current source, the current i is always in the same direction and of the same value as specified by the source.)
Step 2. Assign a voltage to each element that does not have a specified voltage. Give the voltage a name (using v symbols) and arrange the signs of the voltage according to the passive sign convention. (Exception: sources, being active, do not have to follow the passive sign convention. The voltage value and polarity of voltage sources are always as specified by the source; the current direction through the voltage source, however, is arbitrary.)
Step 3. Apply KVL to each independent loop, Ohm ’s Law to each resistor, and KCL to each essential node. This results in a series of simultaneous algebraic equations.
S te p 4. Solve the simultaneous equations for each desired i> and i.
L et’s apply these steps to Fig. 9.4. After assigning names and directions by Steps 1 and 2, we have something that looks like Fig. 9.5. Note the correct application of the in Fig. 9.5.Now apply KVL of Step 3 to Fig. 9.5. The first KT we will take is around the left loop in the clockwise direction (denoted K T l) starting at the lower left comer. Summing voltages around this loop and following Rule 1, we get
- Va + ui + u3 = 0. (9.4)
Next, take a KT around the right loop starting at node 2 and going in the clockwise direction denoted KT2. Summing voltages, we get
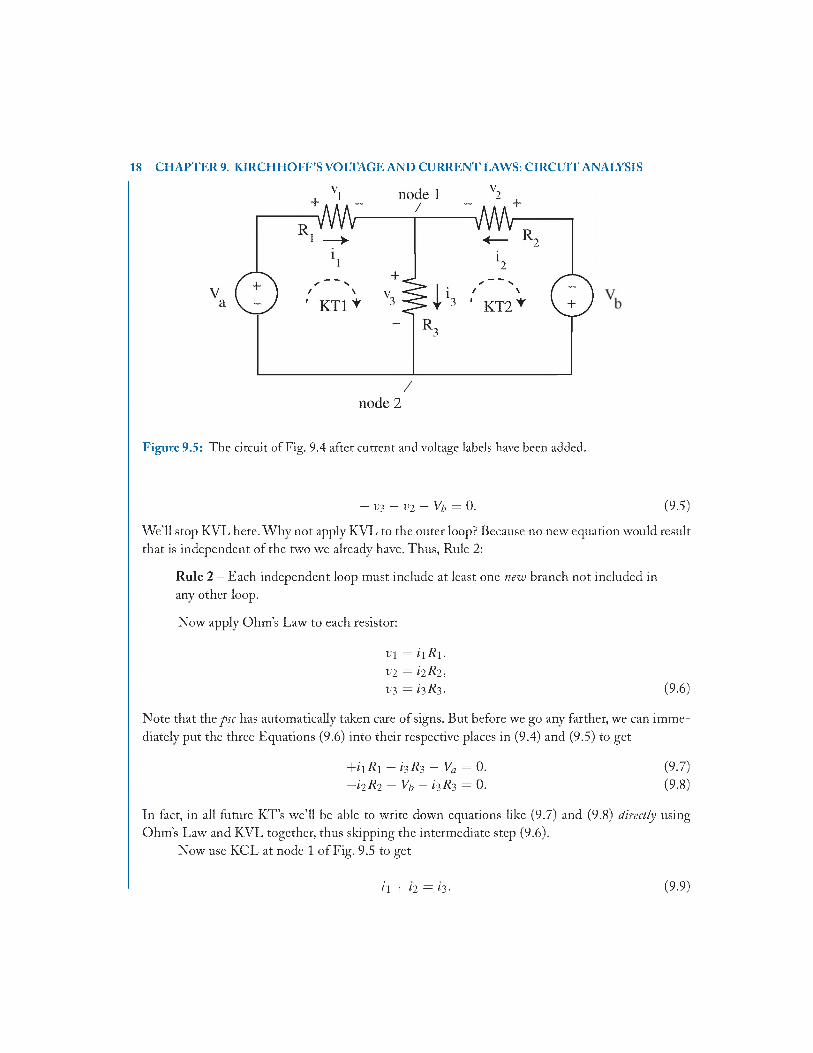
18 CHAPTER 9. KIRCHHOFF’S VOLTAGE AND CURRENT LAWS: CIRCUIT ANALYSIS
no d e 2
Figure 9.5: The circuit of Fig. 9.4 after current and voltage labels have been added.
- V 3 - V 2 - V b = 0. (9.5)
W e’ll stop KVL here. W hy not apply KVL to the outer loop? Because no new equation would result that is independent of the two we already have. Thus, Rule 2 :
Rule 2 - Each independent loop must include at least one new branch not included in any other loop.
Now apply Ohm ’s Law to each resistor:
v \ = i \ R \ ,
V2 = i i R i , V3 = i iR i - (9.6)
Note that the psc has automatically taken care of signs. But before we go any farther, we can immediately put the three Equations (9.6) into their respective places in (9.4) and (9.5) to get
+ ii/? i + i 3 / ? 3- Vfl = 0. (9.7)- h R i - V b - i3R3 = 0. (9.8)
In fact, in all future K T’s we’ll be able to write down equations like (9.7) and (9.8) directly using Ohm ’s Law and KVL together, thus skipping the intermediate step (9.6).
Now use KCL at node 1 o f Fig. 9.5 to get
i l + 12 = 13- (9.9)

9.4. RESISTIVE CIRCUIT ANALYSIS USING THE BRANCH CURRENT METHOD 19
W e’ll stop KCL here. W hy not apply KCL to the other node? Because no new equation would result that is independent of the one we already have. Thus, Rule 3:
Rule 3 - For n nodes, KCL gives (n — 1) independent equations.
Now we go to Step 4. We have three independent simultaneous Equations [(9.7), (9.8), and (9.9)] with three unknowns [ii, 12, and 13]. There are various ways to solve them (such as Kramer’s Rule and Matlab), but we’ll simply use substitution. Solving for i 1 from (9.9):
i i = j‘3 — i2.
Substituting this into (9.7) gives
2 1 3 1 32 2 3 3
1 2
2 3 1 3 1 1+ »2 + »3(fl3/fl2) = -Vfc/ / ?2.
2 3
. R i R 2
13 R 1 + R 3 R 3 ■ R i R 2
Now that we have an equation for 13, we could find the equations for i i m d 1*2 if desired. We could also find equations for v i , V2 , m d 113 easily by multiplying the currents by their respective resistances. Also if values have been given for the various components, we could calculate the values of all currents and voltages in the circuit. An example using another circuit is given next.
Example 9.2. A nother C ircuit Analysis Using K irchhoff’s LawsIn the circuit below,
2
2 2
(9.10)
(9.11)(9.12)
(9.13)(9.14)
(9.15)
I s = 1 mA /?i = 10 kQ
Va = Vh = 1 0 V R2 = 10 kQ R 3 = 2 0 k Q

20 CHAPTER 9. KIRCHHOFF’S VOLTAGE AND CURRENT LAWS: CIRCUIT ANALYSIS
V V,
Figure 9.6: Circuit analyzed in Example 9.2.
Solutiona. Steps 1 and 2 of the Branch Current M ethod have already been done in the figure above. Note
3in that branch. Now proceed with Step 3:KVL around the left loop:
KVL around the right loop:
KCL at upper node:
i i 3 0
V, + Is R3 - i2 R 2 - V b = 0.
(9.16)
(9.17)
h + h + h = 0. (9.18)
Step 4: Now solve these equations. Add (9.16) to (9.17), immediately. Several terms cancel, leaving
i
Put (9.20) into (9.19):
i i 2 2 0
i 2
(9.19)
(9.20)
Va - h R i - h ( R i + R 2) - V b = 0 (9.21)

9.5. PROBLEMS 21
i2 = - W a + Vh + Is R i ) / ( R i + R i) . (9.22)
b. Putting the values from Fig. 9.6 into (9.22) gives
i i = - ( 1 0 + 1 0 + 10)V/(10 + 10) kQ = - 3 / 2 mA = - 1 . 5 mA,
v 2 = i2R 2 = ( -1 .5 mA)(10 kQ) = - 1 5 V. (9.22)
W hat does the negative sign for i i mean? It says that the current in R i is really going in the opposite direction of the arrow, or left to right. We guessed wrong when we set up the current arrows, but it’s fine. The signs of the answers will give the correct final directions. W hat does the negative sign for i>2 mean? I t says that the magnitude of the voltage on the left side of R i is really larger than the voltage on the right side. Once again, the signs of the answers will give the correct polarity.
9.5 PROBLEMS9.1. Let R = 2.0 kQ in the circuit of Fig. 9.7. M ark on the figure the correct polarity for the voltage
v using the passive sign convention, then use KVL and Ohm ’s Law to solve for the current i and the voltage v.
T his can be solved directly for ii'.
Figure 9.7: Circuit to be analyzed in Problem 9.1.
fans: u = —10 V and i = —5.0 mA]
9.2. a. Use the Branch Current M ethod (KVL and Ohm ’s Law) to solve for the current i and the voltage V2 across the 1.0 kQ resistor in the circuit of Fig. 9.8.
fans: i = — 2 .0 mA and v i = —2 .0 V]
b. How many electrons will pass point a in the circuit during 5.0 seconds?

22 CHAPTER 9. KIRCHHOFF’S VOLTAGE AND CURRENT LAWS: CIRCUIT ANALYSIS
Tans: 6.3 x 1016 electrons]
c. Is the direction of the electron flow clockwise or counterclockwise?
Tans: clockwise]
point a
Figure 9.8: Circuit to be analyzed in Problem 9.2.
9.3. a. For the circuit shown in Fig. 9.9, use the Branch Current M ethod to find an expression for3
[ans: 13 = - V a (R i + + R 2 R 3 + R ^ 3 )]
b. Do at least two ranging checks on the answer of part a.
3
600
33
3 V b R l - V g ( *1 + Rl )
_ Ri R2 + R 1 R 3 + R 2 R 3 _3
b. Perform a units check on this equation.
c. Perform one ranging check on this equation.

9.5. PROBLEMS 23
Figure 9.9: Circuit to be analyzed in Problem 9.3.
R,
Vu
M rR ,
+ 1
+ A R,
-\MArV3 +
Figure 9.10: Circuit to be analyzed in Problem 9.4.


C H A P T E R 10
25
10.1 INTRODUCTION
There are several sources of electrical signals produced inside the body: signals from cardiac muscle (recorded as electrocardiograms, E C G or EKG), skeletal muscle (electromyograms, EM G), and brain activity (electroencephalograms, EEG ).These voltage signals can be measured with electrodes on the surface of the body, that is, noninvasively, to diagnose diseases and determine the physiological state of various organs. Rut the voltages are very weak by the time they propagate to the body’s surface— usually on the order of just a few microvolts (\.iV ). By the time they reach the measurement and recording devices, they are even smaller. Figure 10.1(a) models an E C G signal on the surface of the body as an independent voltage source Vecg. This model is simplified, because in reality the EC G signal is time-varying and not constant, but at any given time it may be approximated by this model. Electrodes and wires connect the source to a measurement device (for example, an oscilloscope or an
1and the input resistance (also known as the input impedance) of the measurement device is modeled
2
measuring
+
Figure 10.1: (a) Measurement of an internal electrical source Vecg with electrodes on the body’s surface. The wires and electrodes are modeled by R i , and the input impedance r f the measurement device by R 2 - The measured voltage is t’2 - (b) An amplifier with gain A (denoted by the triangle) is inserted between the source and the measurement device.
2the simple circuit of Fig. 10.1(a). KCL applied to any (nonessential) node around the circuit shows that the current i must be the same in all segments of the circuit. This has already been noted in the

26 CHAPTER 10. OPERATIONAL AMPLIFIERS
figure. KVL around the loop, starting at the lower-left corner and going clockwise, gives
- V ecg + i R i + i R 2 = 0, (10.1)
i 2
From Ohm ’s Law,
V2 = i R 2 = ( Vecg- (10.3)i 2
2
2 i 2voltage is applied to a string of resistors in series, and the measured voltage is taken across one
i
2 2 can be difficult to measure in the presence of noise. Some means of amplifying the source voltage is needed.
Figure 10.1(b) shows a simple amplifier with a voltage gain of A (and a very high input impedance) inserted between the source and the measuring device. The amplifier in this example has a single input, denoted by voltage i I t s output voltage v„ is related to the input voltage by
v0 = A v ,. (10.4)
The voltage gain A maybe large (up to perhaps i x i0 5). Thus, the measured voltage can be boosted with the amplifier to the neighborhood of a volt or a significant fraction of a volt, which is easy to measure and record.
10.2 OPERATIONAL AMPLIFIERSA very popular and versatile type of amplifier is the operational amplifier, or op amp. It forms the building block for a number of convenient amplifier circuits. The device itself consists of a tiny integrated circuit (IC) with many transistors, diodes, resistors and capacitors performing the amplifying function. We will not be concerned here with the internal electronics of the op amp. Instead we treat it as a module, and model it with an equivalent circuit that describes its overall electrical behavior.
The physical package of a typical op amp is much smaller than a postage stamp, and from the outside it looks like a “bug” with wire legs, at least in the popular dual-in-line (DIP) package, or “chip”. Figure 10.2 shows how an 8-pin D IP package looks from a top view. There is always some registration mark (a dot or a cutout) at one end to orient the numbering order of the pins. By convention, the pin numbers start from 1 on the left of the mark and increase in the counterclockwise

Device Symbol Simple Symbol
10.2. OPERATIONAL AMPLIFIERS 27
1 c ) 82 C 5 73 C 5 64 C
\5 5
8-pin DIPtop view
noninverting input ____
inverting" input
positive power supply
output
negative power supply
Figure 10.2: The physical layout of an example of an op amp package. Also shown are typical symbols used in op amp circuit diagrams.
direction as seen from the top, shown in Fig. 10.2. An 8-pin D IP package can actually house two separate op amps, which is common.
The complete symbol for one op amp is given at the center of Fig. 10.2. Note that the op amp has two inputs (labeled inverting and noninverting) and one output, so it is a type of dual-input, or differential, amplifier. Since the output voltage is often larger than the input voltage, the output power is often much larger than the input power. The conservation of energy principle requires that there be some other external source of power supplied to the amplifier to provide this increase in power. This takes the form of one or (usually) two power supplies that must be connected to the amplifier for it to function properly. These are labeled as the positive power supply and the negative power supply connections on the symbolic diagram.
An abbreviated symbol is often used in op amp circuit diagrams, shown on the right of Fig. 10 .2 . The location of the inverting input is denoted by a — sign, and the noninverting input by a + sign. It is important to note that the + and — signs inside the symbol for the two inputs have nothing to do with the actual polarity of the input voltages; they merely denote the inverting and noninverting nature of the two inputs, explained shortly. The power supply voltage leads are labeled V+ and V_. Here the + and — signs do denote the polarity of the supply voltages.
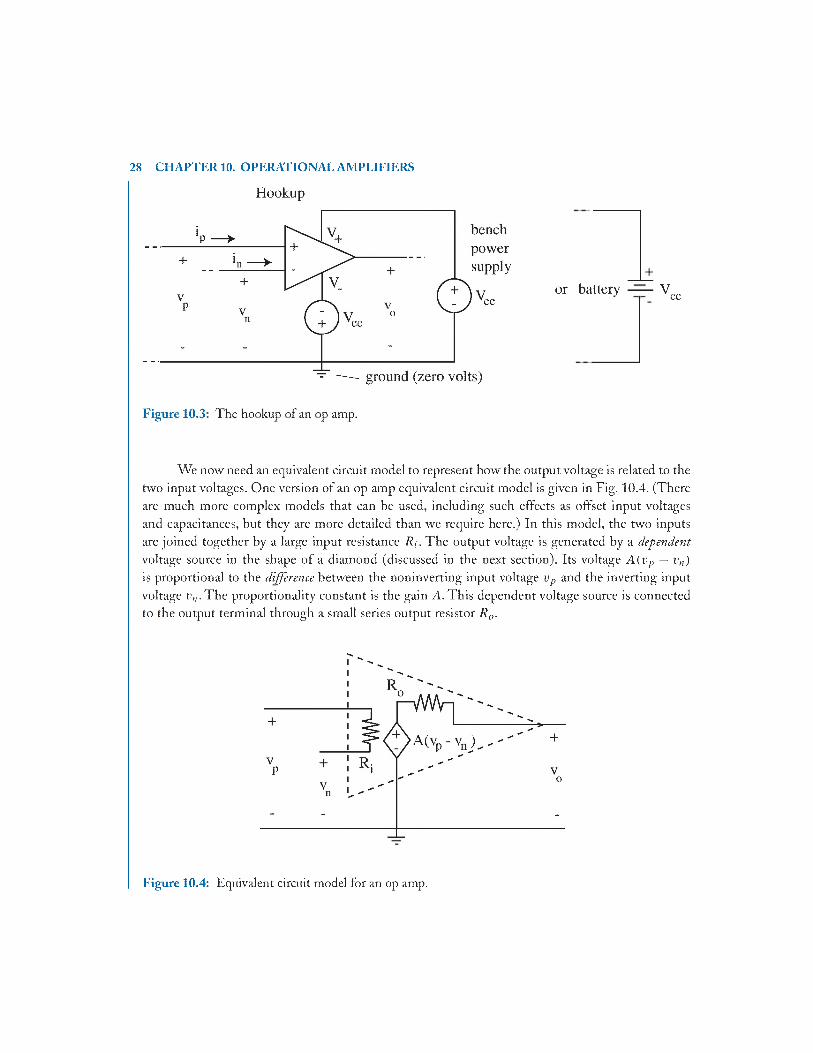
A diagram of how an op amp is hooked up is given in Fig. 10.3. The power supplies can be either bench-type voltage supplies or batteries (for portability); the positive supply requires a constant positive voltage (usually + 1 2 V or +15 V) and the negative supply requires a constant negative voltage (usually —12V o r —15 V). These are denoted + V CC and — Vcc.The input voltage to the noninverting terminal is labeled v f, and the input voltage to the inverting terminal is labeled v„. Both are referenced to the bottom wire, which is usually tied to ground at zero volts. The output voltage is v„, again referenced to ground. The ground is indicated by a series of short lines inside an inverted triangle.
28 CHAPTER 10. OPERATIONAL AMPLIFIERS
Hookup
or b a tte ry ------ V cc
We now need an equivalent circuit model to represent how the output voltage is related to the two input voltages. One version of an op amp equivalent circuit model is given in Fig. 10.4. (There are much more complex models that can be used, including such effects as offset input voltages and capacitances, but they are more detailed than we require here.) In this model, the two inputs are joined together by a large input resistance /?;. The output voltage is generated by a dependent voltage source in the shape of a diamond (discussed in the next section). Its voltage A { v p — vn) is proportional to the difference between the noninverting input voltage vp and the inverting input voltage i'„.T he proportionality constant is the gain A. This dependent voltage source is connected to the output terminal through a small series output resistor R 0.
Figure 10.4: Equivalent circuit model for an op amp.

10.3. DEPENDENT SOURCES 29
Typical values for a good op amp are listed in Table 10.1 along with simplifying approximations to these values. The input resistance R i is usually very large— several hundreds of megohms. Thus, it can be approximated as an infinite resistance, or open circuit. The output resistance R„ is usually small— less than 75 ohms— compared to other resistors in the circuit. Thus, it can be approximated as zero resistance, a short circuit or an ideal wire. The voltage gain A is very large. This leads to a further simplification in the model, as will be apparent shortly.
Table 10.1: Op Amp Parameters
Typical Values Approximate ValuesRi 1012 fi Ri ooR 0 < 75 fi R o ^ 0A = 105 — 106 A —*• oo
W hen these approximations are made in the equivalent circuit, the ideal op amp model of Fig. 10.5 results. In the ideal model, the inputs both terminate in open circuits in the amplifier; therefore no current can flow into these input terminals, regardless of the input voltage. Also the output is directly connected to the dependent voltage source (since R 0 = 0), so the output voltage is the same as the dependent source voltage. From now on, we will use this ideal model in the analysis of various amplifier configurations.
Figure 10.5: The ideal op amp equivalent circuit.
10.3 DEPENDENT SOURCESWe have just introduced a new type of electrical source, the dependent source. Independent sources were described in Chapter 9, where it was seen that an independent source (for example, an inde

30 CHAPTER 10. OPERATIONAL AMPLIFIERS
pendent voltage source) is characterized by the fact that its output voltage is fixed at a constant value. A battery is an example of this kind of source. Independent sources are identified by the shape of a circle or by a battery symbol.
D ependent sources, on the other hand, have outputs that are variable, depending upon the value of some voltage or current in another part of the circuit. Dependent sources are identified by a diamond shape. There are four possible types, categorized in Fig. 10.6.
Current-Controlled Voltage Source
(CCVS)
Voltage-Controlled Voltage Source
(VCVS)
Cu rrcnt-Control led Currcnt Source
(CCCS)
Voltage-Controlled Currcnt Source
(VCCS)
symbol
somewhere else in circuit vWV
q-v,
— M/V— —\AA/V— — M/V—
Figure 10.6: Dependent sources of four possible types.
The current-controlled voltage source (CCVS) is a dependent source whose output voltage is1
1with units of V/A. The voltage-controlled voltage source (VCVS) is a source whose output voltage is proportional to the voltage v i across some element in another part of the circuit. The output voltage of this source is q ■ v i regardless of the current through the source. The proportionality constant is q , with units of V/V, so it is dimensionless.The VCVS inside the ideal op amp equivalent circuit is an example of this type of source, extended such that the output voltage is proportional (with proportionality constant A ) to the difference (lip — v„) of two voltages on other parts of the circuit. To complete the possible dependent source configurations, Fig. 10.6 also shows a VCCS and a CCCS.
10.4 SOME STANDARD OP AMP CIRCUITS
Op amps are used in hundreds of different applications and configurations (for example, the cardiovascular Major Project utilizes one such specialized circuit: the capacitance-multiplier circuit), but there are three or four standard op amp configurations that are used again and again. They are described and analyzed next.

10.4. SOME STANDARD OP AMP CIRCUITS 31
10.4.1 IN V E R T IN G A M P L IF IE RThis arrangement is used when it is desired to amplify and invert an input signal so that the output voltage is larger than the input but of opposite polarity. Its circuit diagram is shown in Fig. 10.7(a).
with its ideal equivalent circuit.
Rs is the resistor connecting the source to the op amp. R f is a “feedback” resistor connecting the output back to the inverting input. The feedback resistor must always be connected back to the inverting input, not the noninverting input. Otherwise the amplifier will go unstable. Note in Fig. 10.7(a) that the inverting input is above the noninverting input; this order may vary from diagram to diagram.
To analyze this circuit, we always redraw the circuit, replacing the op amp symbol with its (ideal) equivalent circuit. We then can use the Branch Current M ethod or any appropriate method to find the output voltage in terms of the signal voltage. The redrawn circuit is shown in Fig. 10.7(b). vn , Vp, and va are identified at their respective terminals, labeled a, b, and c.
We first use KCL at the node near a. Its result is obvious: since no current can enter terminal a, the current entering the node must equal the current leaving, or
h = is- (10.5)
This result is so intuitive that from now on, whenever there is a continuous branch with multiple elements but containing no essential nodes (i.e., no places with a third wire branching off to the side), we will let the current be the same throughout that continuous branch.
KVL can be applied to the far-right loop— the one that includes the dependent source. This loop is indicated by the dotted line labeled KTx. Starting in the lower-left comer, we get
- A ( v p - vn ) + v0 = 0. (10.6)
soVp - V„ = v t, f A . (10.7)

Now a very important approximation can be made. Since in the ideal op amp model, A oo, (10.7) shows that
v p - vn % 0 , (10.8)
orEvery ideal op amp (10.9)
32 CHAPTER 10. OPERATIONAL AMPLIFIERS
Therefore, in the ideal model, the voltages at the inverting and noninverting input terminals are the same! We will employ this simplification from now on, and it makes the analysis of op amps much easier, as we will see. Also, when the relationship in (10.9) is used, we will never need (or want) to take a KT through the dependent source again.
Now apply KVL around the loop labeled K T l. Start at the lower left and go clockwise. The first voltage term encountered is the signal voltage V^The next term is the voltage across the resistor R s , which is + isR s . Now we need to get from terminal a (with voltage v„) to terminal b (with voltage V p ) . Since (10.9) states that V p = v,u there is no voltage drop encountered when going from point a to point b. So there is a zero contribution to the voltage sum here. T hat takes us to point b.There is a wire from this point back to the start of the loop (note that this means that v p = 0 in this circuit), so the total K T l tour results in:
V5 + is R s + 0 = 0, (10.10)
oris = Vs/ R s . (10.11)
There is one branch we have not yet used in KVL, the one containing R f , so we apply KVL to the loop labeled KT2. Note that KT2 avoids going through the dependent source to make things simpler, and instead goes through v0. Applying KVL to KT2 gives
+ is R f + v0 — 0 .
Putting (10.11) in (10.12) and solving for va gives
R r
* 1 v "Inverting amp
(10.12)
(10.13)
We check the units of (10.13) for consistency, and since both sides have the units of volts, we know that the derivation hasn’t gone terribly astray somewhere.
Equation (10.13) states that the output voltage of this amplifier is equal to the input voltage multiplied by the ratio of R f to /?s.This ratio gives the absolute value of the gain of this circuit, and is set by resistance values (and is therefore more stable compared to other electronic ways of setting the gain). The gain can be set to be low, moderate, or very high, as desired, by choosing the proper resistors, but cannot be larger than the native (open-loop) gain A of the op amp. The negative sign

10.4. SOME STANDARD OP AMP CIRCUITS 33
means that the polarity of the output is inverted from that of the input (thus the name “inverting” amplifier); for example, if R f / R s = 100 and V5 = + 6 mV, the output voltage is v0 = —600 mV.
10.4.2 N O N IN V E R T IN G A M P L IF IE RThis configuration is used when it is desired that the output voltage have the same polarity as the input voltage. The original circuit is shown in Fig. 10.8(a). The circuit redrawn with the ideal equivalent circuit substituted for the op amp symbol is shown in Fig. 10.8(b). This configuration is similar to the inverting amplifier, but the signal voltage source is now connected to the noninverting input rather than the inverting input.
Figure 10.8: (a) Noninverting amplifier; (b) redrawn circuit using ideal op amp equivalent circuit.
We don’t need to use KCL explicitly here, since there are no branching currents. KVL around K T l gives
0
sois = - V s/ R s . (10.15)
In getting (10.14) we have already used the fact that v p = v„. Next, we do not need or want to take a loop through the dependent source. W h a t’s left is KVL around KT2, giving
0
Solving for va and using (10.15) results in
Noninverting amp (10.17)
This result shows that the gain of the noninverting amplifier is also related to the ratio of resistances, as was the case with the inverting amplifier, but with a slightly different form due to the

first term in the parentheses.The gain of the noninverting amplifier is always greater than unity, even for small ratios of R f / R s ■ W hen R f > > R s , the gains of the two configurations are essential the same in absolute magnitude. Note that the sign of the gain for this noninverting amplifier is positive, so the input and the output voltages have the same sign (as the name “noninverting” indicates). For example, if R f / R s = 10 and Vs = + 6 mV, the output voltage is v 0 = + 6 6 mV.
10.4.3 V O LT A G E F O L L O W E RThis op amp configuration is the simplest possible one, and it is often used when it is necessary to avoid “loading down” a source. The term “loading” refers to the current that is required from a source when it is connected to a measuring or recording device, called the load. If the resistance of the load is low, then an appreciable amount of current will be drawn from the source. If the resistance of the source is not small (it often isn’t, especially in the case of chemical electrodes), then this current draw will lower the effective voltage that can be measured at the load. A demonstration of this loading effect was seen in the example of Fig. 10.1 (the voltage-divider phenomenon). For instance, if the internal resistance of a chemical electrode is R i = 1 MSI and the input resistance of the measuring device is R 2 = 1 kf2, then (10.3) shows that only about 1/1000 of the electrode voltage will actually be seen at the terminals of the measurement device. In order for the measured voltage to be nearly as large as the electrode voltage, R 2 must be > > Ri .
The voltage-follower configuration accomplishes this inequality while at the same time preserving the signal voltage in both sign and magnitude. In other words, the voltage follower has a very high input impedance and a voltage gain of positive unity. Figure 10.9(a) shows the op amp circuit and Fig. 10.9(b) is the redrawn circuit with the ideal equivalent circuit.
a. b.
34 CHAPTER 10. OPERATIONAL AMPLIFIERS
Figure 10.9: (a) Voltage follower circuit; (b) the circuit redrawn using the ideal equivalent circuit.
The key to analyzing this circuit is to note that since the branch that contains the signal source0
in the source branch. In turn, this means that there is no voltage drop (from Ohm ’s Law) across the

source resistance Rs (in fact, Rs has no effect at all on the output of this circuit). Then using KVL around K T l gives
10.4. SOME STANDARD OP AMP CIRCUITS 35
Vs + vo = 0 .
Vo = Kv Voltage follower
(10.18)
(10.19)
This result confirms that the voltage follower’s output voltage is exactly the same as the signal voltage, thus the name “voltage follower.” This circuit is also sometimes called a unity gain amplifier. In addition, due to the (approximately) infinite input impedance of the op amp, no current is drawn from the source and there are no loading effects. Therefore, this circuit is also often called a buffer amplifier.
A slightly more complex op amp configuration is treated in the next example.
Example 10.1. T h e Sum m ing Am plifierThis amplifier is used in situations where the voltages from two or more sources are to be added together, with perhaps different gains for each voltage. Its circuit is shown in Fig. 10.10.
b.
Figure 10.10: Summing Amplifier analyzed in Example 10.1.
SolutionSince we have an essential node at the node labeled e, KCL must be applied at this node to give
+ ia + ib ~~ i f = ° (10.20)
Now apply KVL around K Tl:

36 CHAPTER 10. OPERATIONAL AMPLIFIERS
0
Apply KVL around KT2:
0
Solving for ia and ib from (10.21) and (10.22), and substituting in (10.20) gives
i f = ( Va / Ra) + (V h /R h )• (10.23)
Now use KVL around KT3, which includes the remaining branch, to get
0
or _______________________________________
= —i fR f = —V{7 ( —— J — Vb ( —— J • Summing amp (10.25)\ R a J \ Rb )
Thus, the output voltage is the weighted sum of the input voltages (with inverted signs), where the weighting factors in the parentheses depend on the values o f the resistors chosen. For example, if R f = R a = R lu then v0 = ~ ( V a + Vb).
10.5 PROBLEMS10.1. Use the Branch Current M ethod to analyze the circuit in Fig. 10.11. Remember to redraw
the circuit using the ideal op amp equivalent circuit.
a. Based on the equivalent circuit and Ohm ’s Law, what is the voltage across the 330 £2 resistor?
0
b. W hat is the output voltage v0}
[ans: vp = —5.4 V]
10.2. Use the Branch Current M ethod to analyze the circuit in Fig. 10.12. (Remember to redraw the circuit.) W hat is the output voltage u„?
fans: v0 = —9 .0 V]

10.5. PROBLEMS 37
8700 Q.
Figure 10.11: Op amp circuit to be analyzed in Problem 10.1.
1.0 kQ .
Figure 10.12: O p amp circuit to be analyzed in Problem 10.2.


C H A P T E R 11
39
C o u l o m b ’s L a w , a n d
11.1 COULOMB’S LAWW e’ve seen earlier how electrical charges with the same sign repel each other and charges with opposite signs attract (this is usually stated “like charges repel and unlike charges attract”). W e’ll now add some detail in describing this phenomenon, and show how it leads to the concept of an electrical capacitor. Two charged particles are diagrammed in Fig. 11.1. The forces F exerted on each charge act in a direction along a line between the two charges. I f F has a positive value, the forces tend to push the particles apart. I f F has a negative value, the forces tend to pull the particles together.
Figure 11.1: The force F between two charges depends on their charge, their separation, and the medium between them.
Coulom b’s Law is a mathematical formulation of this principle:
F = g ig i A n s r 2 ’
i i r is the separation (m), and£ is the permittivity of the medium between the charges.
The permittivity of free space (a vacuum) is
(11.1)

40 CHAPTER 11. COULOMB’S LAW, CAPACITORS AND T H E FLUID/ELECTRICAL ANALOGY
eo = 8.854 x l O ^ C 1/ ^ • m1). (11.2)
The permittivity of other media, such as insulating materials called dielectrics, is larger than that of free space by a factor known as the relative permittivity s r of the material; s r is also called the relative dielectric constant. For example, the relative permittivity of polystyrene (for slowly varying voltages) is 2.551. For a material with a relative permittivity of sr , then s = so sr .
Note from Coulomb’s Law that the closer the charges are to each other (the smaller r), the greater the force. This is similar to the inverse distance relationship in Newton’s law of gravity. Also note that when <71 and q i are of opposite sign, the force is negative and inward; when <71 and q i have the same sign, the force is positive and outward. A popular demonstration of this is when a student places her hand on one electrode of a Van de Graaff generator (a generator of static electricity that is a source of excess electrons on one electrode). Because they repel each other, the excess electrons will rapidly spread out over her entire body—including her hair— causing her hair to literally “stand on end.”
11.2 CAPACITORSWe are now able to address the following question: Can we store excess charges in some chosen place in a circuit in spite of Coulomb’s Law, which says that they will repel each other, especially when packed densely with small r values? L et’s try to store excess electrons in a metal wire. A metal is composed of atoms whose outer electrons can escape easily from the atomic shell, and therefore can move under a force to produce a current. These negatively charged electrons are called “free” or “conduction” electrons. But they leave behind the immobile positively charged shell of the atoms (as ions) that are fixed in place by the lattice of the metal. Therefore, a free electron cannot move very far away from its fixed ion (due to the attractive force described by Coulomb’s Law) unless it is replaced by another free electron entering the vicinity of the shell to take its place. The whole metal wire is electrically neutral since the number of free electrons is equal to the number of fixed atomic cores.
Figure 11.2 shows such a metal wire, with — signs, or e symbols, representing free electrons and + signs representing the fixed metal ions. W hat would happen if we forced another (that is, an excess) electron into one end of the wire? Its negative charge is not balanced by a positive ion, so a Coulomb’s repulsive force would be exerted on neighboring electrons, which would move slightly and in turn exert forces down the wire on other neighboring electrons, so on down the wire. These forces would result in one electron being expelled out the other end of the wire (assuming the wire is connected to a conductive path), and current flows.
Because there was no increase in the number of free electrons in the wire, there was no storage. Essentially, excess charges cannot be stored in normal wires.
1 1 .iving tissue can also be characterized by permittivity and conductivity values, usually specified for alternating (ac) electromagnetic fields in the kHz to GHz frequency range. For example, at 1 MHz, muscle has a conductivity of about 0.5 S/m and a relative permittivity of about 500.

11.2. CAPACITORS 41
metal wire free electrons e '
e ------►+
-> e ’
fixed ions
Figure 11.2: A metal wire with an excess electron forced into one end, causing expulsion of an electron from the other end (a current).
The situation is different in the arrangement of Fig. 11.3, showing a parallel-plate capacitor. In this configuration, two metal plates of area A are facing each other separated by a very thin layer of insulating material of thickness d , such as polystyrene, mylar, or mica. Wires connect to each plate, but charges cannot pass across the insulating layer. Can we store excess charges on these plates? At first you might say no, because the concentration of charges would cause very large repulsion forces, prohibiting the storage of the charges close together as in the wire example discussed above. But if there was some nearby attraction forces, these would partially cancel out the repulsion forces and storage may be possible.
m etal plate .
+ r r ^ rV
[
excess charge q
t
excess charge q
d+"'+ + + I
I 1area A thin dielectric
insulating layer
tFigure 11.3: A parallel-plate capacitor (side view) that can store excess charge on one plate because the opposing plate holds the opposite charge.
Indeed, in the parallel-plate capacitor, when negative charges (electrons) are placed on one plate (the lower plate in Fig. 11.3), electrons are forced off the other (upper plate), passing through

the wire connected to this plate and producing a current i .The absence of electrons on the upper plate leaves behind a concentration of positively charged fixed ions on the upper plate, exactly matching the concentration of electrons on the lower plate. Since these two concentrations are close to each other (at a distance d ) and are of opposite charge, there is an attractive Coulombs force which partially balances the repulsive force due to the concentration of electrons and helps keep the charges in place!
Now think of adding one more electron (or a charge q through the lower wire and onto the lower plate. It would take some force (measured by voltage) to do so, because we would be adding to the concentration of electrons already stored there. But the voltage is not nearly as large as it would be if there were no compensating positive charges nearby. So we can postulate that the voltage v required to add another charge q might take the form
, = (11.3)As,
This form for the equation is justified by the following reasoning: The required voltage is proportional to the amount of charge q added, as is intuitive, and v is smaller if rf is smaller since the opposing charge is closer, v is also smaller if A is larger because the charge is not being concentrated as much on the plate. The permittivity s of the dielectric layer helps increase the attractive force across the layer, as (11.1) shows. The larger s , the less voltage that is required, or conversely, for a given voltage, the more charge can be stored.
The constants in (11.3) are defined by the geometry and dielectric material of the capacitor, and can be combined to define the capacitance C of the particular capacitor:
42 CHAPTER 11. COULOMB’S LAW, CAPACITORS AND T H E FLUID/ELECTRICAL ANALOGY
A sC = — . (11.4)
aThen (11.3) can be rearranged to give
Cu. (11.5)
which is the basic relationship between the charge on and the voltage across a capacitor. The units of C, are farads (in honor of scientist Michael Faraday) with the unit symbol F. From (11.5) the unit of F can be seen to be equivalent to C /V = C 2/(N-m). Also, from (11.4) it can be seen that permittivity s, can be expressed in units of F/m.
In practice, the unit of F is very large (a 1.0 F capacitor is huge!), so most electronic capacitors are measured in subdivisions of F, such as nF or jxF. Also, although all capacitors have two or more conducting plates facing each other as shown in the concept drawing of Fig. 11.3, to save space they are usually rolled up into cylinders or compressed by connecting many interleaved plates in parallel.
11.3 FLOW INTO AND OUT OF CAPACITORSThe symbol for a capacitor is simple: two lines close to each other representing the opposing plates of the capacitor. This is shown in Fig. 11.4. Also shown is a current i into one side (an identical

11.4. ANALOGY BETW EEN FLUID AND ELECTRICAL CIRCUITS 43
current i flows out the other side) and a voltage v across the capacitor related to the stored charge by (11.5).
+V
I 1
I 1Figure 11.4: The symbol for a capacitor.
W hen the charge changes by an amount Aq, the voltage will also change by an amount At). The capacitance C is constant, so (11.5) gives
A q = C Ai).
Now, from the definition of current in the Ohm ’s Law chapter [see (8.1)],
i = A q / A t .
Putting (11.6) into (11.7) yields
A i) A t
i
C '
(11.6)
(11-7)
(11-8)
showing that the current into (or out of) a capacitor causes a time rate-of-change of the voltage across0
differential form
dvd t
i
C '(11.9)
11.4 ANALOGY BETWEEN FLUID AND ELECTRICAL CIRCUITS
Equations (11.5) and (11.8) look familiar. We derived equations of similar form (but with obviously different variables) when we were dealing with fluids. L et’s exploit this similarity between fluids and electricity. Table 11.1 is a summary of some important fluid equations in the left column, and analogous electrical equations in the right column.
W ith a little study ofTable 11.1, we can start to pickout^tffVi ofquantities (a fluid quantity and its electrical counterpart) that appear in the same place in the analogous equations. The quantities
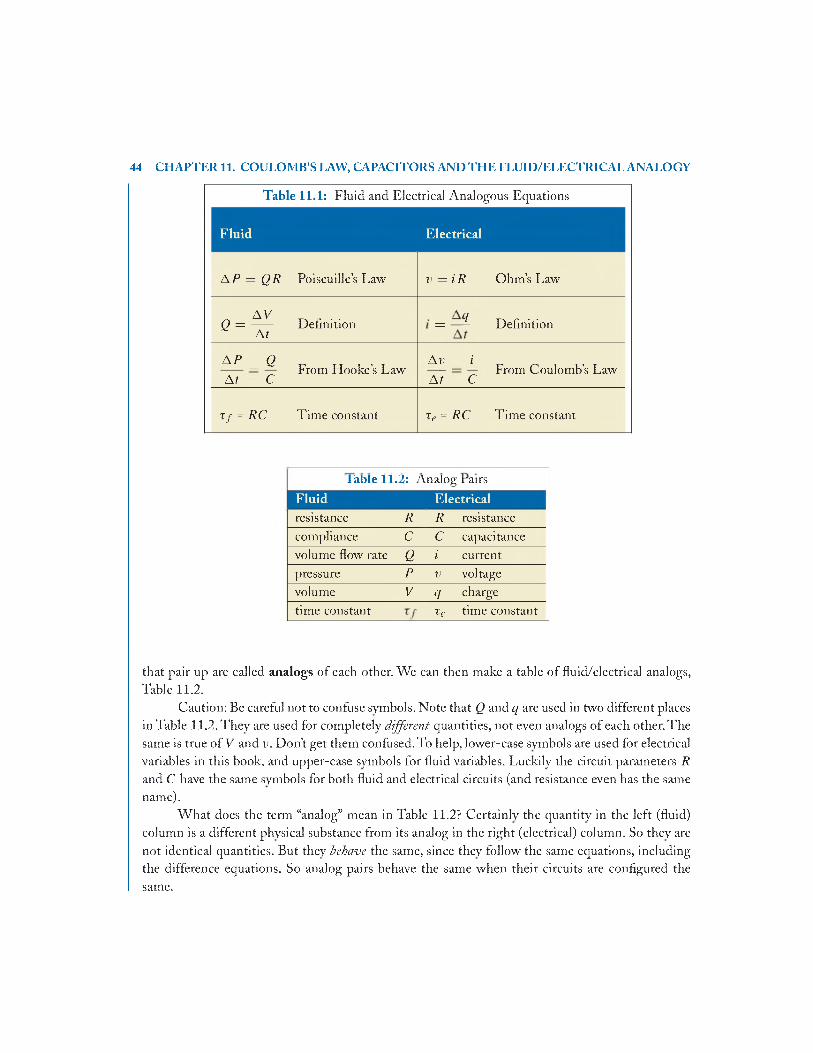
44 CHAPTER 11. COULOMB’S LAW, CAPACITORS AND T H E FLUID/ELECTRICAL ANALOGY
Table 11.1: Fluid and Electrical Analog;ous Equations
Fluid Electrical
A P = Q R Poiseuille’s Law v = iR Ohm ’s Law
A VQ = —A t
Definition II | ^ Definition
A P _ Q
A t CFrom Hooke’s Law
A i) i
a7 c From Coulomb’s Law
i f = R C Time constant Te = R C Time constant
Table 11..2: Analog rairsFluid Electricalresistance R R resistancecompliance C C capacitancevolume flow rate Q i currentpressure p V voltagevolume V <7 chargetime constant Te time constant
that pair up are called analogs of each other. We can then make a table o f fluid/electrical analogs, Table 11.2.
Caution: Be careful not to confuse symbols. Note that Q and q are used in two different places in Table 11.2. They are used for completely different quantities, not even analogs of each other. The same is true of V and v. Don’t get them confused. To help, lower-case symbols are used for electrical variables in this book, and upper-case symbols for fluid variables. Luckily the circuit parameters R and C have the same symbols for both fluid and electrical circuits (and resistance even has the same name).
W hat does the term “analog” mean in Table 11.2? Certainly the quantity in the left (fluid) column is a different physical substance from its analog in the right (electrical) column. So they are not identical quantities. But they behave the same, since they follow the same equations, including the difference equations. So analog pairs behave the same when their circuits are configured the same.

11.4. ANALOGY BETW EEN FLUID AND ELECTRICAL CIRCUITS 45
Therefore, it is possible— and indeed is an important engineering tool— to make an electrical circuit whose variables (various v, i , and q around the circuit) would mimic the behavior of the fluid variables ( P , Q , and V , respectively) in any fluid circuit being modeled. Thus, a compliant fluid vessel is modeled by a capacitor and a resistive fluid element is modeled by a resistor. The voltage v at a given node in the electrical circuit model represents the pressure P at the corresponding node in the fluid circuit. The cardiovascular Major Project exploits this analogy to model and analyze the human CV system.
11.4.1 SCALING TH E ANALOG PAIRSBut we still must deal with the translation of the resistance and compliance values from the fluid domain into resistance and capacitance values in the electrical domain. Is a 100 mmHg-s/L resistance in a fluid tube represented by a 100 fi resistor? Probably not. In fact, the units are different. Remember, the analog pairs are not identical quantities; they only behave the same. So in general, when translating from one domain to the other, we must use a scaling factor S. To translate a fluid resistance R f to an electrical resistance R e, we’ll use
R e = S / iR f , (11.10)
where the factor S r must have units such that the right-hand side of (11.10) has the units of fi. Similarly, for capacitance:
Ce = S c C f . (11.11)
The factor S c must have units such that the right-hand side of (11.11) has the units of F.Now, are we free to choose any values for the translating scale factors S r and S c ? Not without
impacting other scale factors. Note from Table 11.1 that
t f = R f C f (11.12)
andr , = R eC e . (11.13)
If we now make the reasonable decision that we want the time scale in the electrical domain to be the same as in the fluid domain2 (that is, that one second in the fluid circuit corresponds to one second in the electrical model), then from (11.10)—(11.13):
T /■ = R f C f = Te = R ec e = S RS c R f C f . (11.14)
Canceling R f C f terms in (11.14) gives
’There are some instances where it is desirable to slow down or speed up time in the analogous model compared to the system being analyzed, but we won’t do that for the cardiovascular Major Project since we want the electrical model to display the same heart rate as in real life.

46 CHAPTER 11. COULOMB’S LAW, CAPACITORS AND T H E FLUID/ELECTRICAL ANALOGY
or
S r S c = 1
S s = 1 / S C .
(11.15)
(11.16)
Once S c is chosen, S r is set by (11.16). It is often the case that the electrical capacitance size is limited by the largest value available conveniently, so that will determine the scale factor S c • Then S r is found using (11.16).
11.5 PROBLEMS11.1. In the circuit shown in Fig. 11.5, at time t = 0 the voltage V3 = +5 .0 V.
a. W hat is the charge q on the capacitor at t = 0?
[ans: q = 5.0 x 10^4 C]
b. Assuming the currents i i = 2.00 mA and i2 = 2 5 0 mA shown in Fig. 11.5 are constant, what is the voltage 1)3 on the capacitor at time t = 400 ms?
fans: V3 = + 3 .0 V]
i1 = 2.00 mA i9= 2.50 mA
VWVI+
V , —
V
= C =3 —
VWV
Figure 11.5: RC circuit analyzed in Problem 11.1.
11.2. The windkessel fluid circuit shown in Fig. 11.6 has the fluid element values: resistance R 1 = 145 mmHg-s/L, R 2 = 505 mmHg-s/L, and compliance C = 00700 L/mmHg.
You wish to model this circuit with an equivalent electrical circuit. To keep the resistor values from being too large, you want to use the largest capacitor value possible. The maximum capacitor value you have available is 100 /xF. Determine the two scaling factors that will find the electrical values corresponding to the fluid values, one for resistors and one for capacitors,

11.5. PROBLEMS 47
R1 = 145 mmHg.s/L R 2= 505 mmHg.s/L
vw v V\M r
C = 0.0700 L/ininHg
Figure 11.6: Windkessel circuit to be converted in Problem 11.2.
including the correct units for the conversions. Then calculate the values for the two equivalent resistors and the capacitor.
fans: S r = 700 (L-Q)/(mmIig-s), S c = 0.00143 (F-mmHg)/L, /?i = 102 kQ, = 354 kQ, and C = 100 /i ¥]


C H A P T E R 1249
a n d P a r a l l e l
C o m b i n a t i o n s o f R e s i s t o r s a n d
12.1 INTRODUCTION
We have encountered series and parallel combinations of resistive fluid elements in earlier units. We now consider series and parallel combinations of electrical resistors and of capacitors. We will initially use Kirchhoff s Laws and O hm ’s Law to analyze the combinations and to reduce them to single equivalent elements, but once formulas are derived for the value of the equivalent elements, we can use these formulas directly to simplify circuits. We will see that the equations for combining electrical components are identical to those for combining analogous fluid components.
12.2 RESISTORS IN SERIES
W hen two or more resistors are connected in a line, as shown in Fig. 12.1 for three resistors of different values, the same current i must flow through each resistor. This is a consequence of applying Kirchhoff s Current Law (KCL) at each (nonessential) node between the individual resistors.
R eq
\
Figure 12.1: A series combination of three resistors. The equivalent resistance is R eq

SO CHAPTER 12. SERIES AND PARALLEL COMBINATIONS
Kirchhoff’s Voltage Law (KVL) can be applied by taking a Kirchhoff’s Tour clockwise around the loop in Fig. 12.1, using Ohm ’s Law directly to express the voltage drop across each resistor. This gives
(12.1)
ori = ------------------- . (12.2)
i 2 3
Now the three resistors can be replaced by a single equivalent resistor R eq (as far as the effect at the outer terminals is concerned) if the current i is the same for the equivalent resistor as for the original series combination. Thus, we require
R,eq
Comparing (12.2) and (12.3) shows
Req = ^1 + ^2 + ^3 Series R (for N = 3)
for three resistors. In general, for N resistors,
(12.3)
(12.4)
Series R (any N ) (12.5)
Note from (12.4) or (12.5) that the equivalent resistance is always larger than the largest of the individual resistors.
12.3 RESISTORS IN PARALLELIn a parallel combination, the resistors are connected side-by-side, as shown in Fig. 12.2 for two resistors of different values. The currents ^ m d i 2 can be found by using Ohm ’s Law for each resistor:
v vii = — and i 2 = — .1 r 1 2 r 2
Applying KCL at node a and using (12.6),
v v ( 1 1 i — /1 in — -f- — — v 1 — — 1 .
1 2 1 2 1 2
(12.6)
(12.7)
Again we require that the current i is the same when the parallel combination is replaced (across the outer terminals) by an equivalent resistance R eq. Comparing (12.3) with (12.7) gives

o -
a/
12.3. RESISTORS IN PARALLEL 51
R eqR,
VO
R~
Figure 12.2: A parallel combination of two resistors having an equivalent resistance Rcq.
1 1+
1 iR eq R i R i R i R i
Inverting (12.8),
R eqR i R i
R i + R iParallel R (for N = 1 only)
(12.8)
(12.9)
for two resistors. In general, for N resistors in parallel,
1 ^ 1-----= V — Parallel R (any N )
1(12.10)
Note from (12.9) or (12.10) that R eC[ is always smaller than the smallest of the individual resistors. For example, for the simple case of two resistors of equal resistance R , (12.9) gives R eq = ( 1 /2 ) R .
Example 12.1. Resistor Netw orkFind the equivalent resistance of the resistor networkbetween the terminals markedX-X in Fig. 12.3.
SolutionFirst combine the three resistors that are in parallel (the lower three) using (12.10):
1 1 1 1+ _______+
R p 5 0 .0 Q 100.0 G 7 0 0 a0.0443/S^,
1
52 CHAPTER 12. SERIES AND PARALLEL COMBINATIONS
20.0 0
— W W -------------x
VWV70.0 0
50.0 0
—\/W\r100.0 0
■X
Figure 12.3: Parallel and series network of resistors analyzed in Example 12.1.
to obtainR p = 22.6 Q .
The network then reduces to two resistors in series (see Fig. 12.4), which can be replaced by a single R eq.
20.0 0
VWV- X
R p =22.6 0
X
R,eq
X--------------------------------X
Figure 12.4: The circuit of Fig. 12.3 simplified in steps.
Using (12.5), the overall equivalent resistance is
20 0 22 6
12.4 CAPACITORS IN SERIESFigure 12.5 shows two capacitors in series. Using KVL around the loop gives
1 2 (12.11)

12.4. CAPACITORS IN SERIES 53
Therefore, any change in the voltage across the capacitors A v divided by the increment in time A t is
1 iA t ^ A t + A t '
(12.12)
o -
r + i
C eq <
C, =!= v,
Co =±= v0
Vo -
Figure 12.5: Two capacitors in series. The effect is the same as a single equivalent capacitor Ceq.
The relationship between the change in voltage across each capacitor and the current into the capacitor is given by (11.8) from the Coulomb’s Law chapter:
(12.13)A v i
a7 c'Since the current i is the same into each capacitor, using (12.13) in the right-hand side of (12.12) gives
A v i i ( 1 1~Kt c 1 + c i
(12.14)
We can define an equivalent capacitance relating the current i to the change A v in the overall voltage
A v
A t C,eq
Comparing (12.15) with (12.14) shows that
1 1 1Ceq c [ + c i ’
c,eqC 1C i
C1 + C iSeries C (for N = i only)
(12.15)
(12.16)
(12.17)

54 CHAPTER 12. SERIES AND PARALLEL COMBINATIONS
for the case o f two capacitors in series. In general, for N capacitors in series,
Series C (any N ) (12.18)
Note that the formula for combining capacitors in series is the same as for combining resistors in parallel.
12.5 CAPACITORS IN PARALLELThe combination of three capacitors in parallel is shown in Fig. 12.6. Applying KCL to node a gives
i = i\ + i 2 + h - (12.19)
Ceq
O+
o
7 * ji |
c,"
' 2 111 cu
II ir
n UJII
Figure 12.6: Three capacitors in parallel, giving an equivalent capacitance C el{.
From (12.15), the equivalent capacitance can be found from
iCe
Putting (12.19) in (12.20):
C,11 12 13
/A iA ' / A iA ' / A dV
(12.20)
(12.21)
V A t ) \ A t ) \ A t )
Since the same voltage v appears across all capacitors, each term on the right-hand side of (12.21) is equal to [using (12.13)] the capacitance of the respective individual capacitors, giving
C eq = C i + C2 + C 3 Parallel C (for N = 3) (1 2 .2 2 )

12.5. CAPACITORS IN PARALLFX 55
for three capacitors in parallel. In general, for N capacitors in parallel,
N
£ Cj Parallel C (any N ) (12.23)1
Note that the formula for combining capacitors in parallel is the same as for combining resistors in series: the capacitances add. A good way to remember that capacitors in parallel add together, as (12.23) shows, is to recall that the equation for capacitance, (11.4), is
AeC = T '
(12.24)
where A is the area of the capacitor plates. Putting capacitors in parallel effectively adds their areas, so the equivalent capacitance is just the addition of the individual capacitances.
Example 12.2. Capacitor Netw orkFind the equivalent capacitance of the following network at the terminals markedX-X (see Fig. 12.7).
X
17 |uF —i—
= = i o mf
= = 1 0 MF
X
Figure 12.7: Capacitor network analyzed in Example 12.2.
SolutionFirst combine the two capacitors that are in series (on the right) using (12.18):
1 _ 1 1 _ 2 _ 1C s 10 (iF ^ 10 ( iF 10 5 f i F ’

56 CHAPTER 12. SERIES AND PARALLEL COMBINATIONS
to obtainCs = 5 f i F.
The network then reduces to two capacitors in parallel, which can be replaced by a single Ceq (see Fig. 12.8), and, using (12.23), the overall equivalent capacitance is
C eq = H + 5 = 22 iiF .
1 7 | i F ^ =
X
=!= C
X
cq
Figure 12.8: The circuit of Fig. 12.7 simplified in steps.
12.6 VOLTAGE DIVIDERIt is often desirable in electrical circuits to reduce the voltage from a source to a smaller value than the original source voltage. A circuit that will accomplish this is the voltage divider, shown in Fig. 12.9. Here a chain of resistors (two or more) in series is attached to the source, and the voltage across one of the resistors, say v i , is measured.
The equivalent resistance of the series network is found from (12.4), R eq = R \ + R 2 + R 3 , so the current i from Ohm ’s Law is
i = — = --------—-------- . (12,25)Req R l + ^2 + ^3
Using Ohm’s Law again, the voltage V2 equals i R i , or
V2 = VsR i
R i + R 2 + R 3Voltage divider (for N = 3) (12.26)
Notice that the voltage Vi has been reduced by the fraction in the parenthesis of (12.26). This fraction is the ratio of the resistance across which the voltage of interest is measured {R i) to the

12.7. CURRENT DIVIDER 57
Figure 12.9: A voltage-divider network.
sum o f all the resistances in series ( R 1 + R i + ^ ^ .T h u s , the larger the resistor R i , the greater thei
12.7 CURRENT DIVIDERW hen the current from a current source is split among two or more branches, this configuration is known as a current divider, shown in Fig. 12.10.
Figure 12.10: A current-divider network.
To find the common voltage v across the parallel combination of resistors, we use the equivalent resistance from (12.9) in conjunction with Ohm ’s Law:
'The number oi resistors in the chain oi the voltage divider is arbitrary. There could be two, or tour, or more in series. In every case, the numerator o f (12.26) remains the same, while the denominator is the sum of all the resistances in the chain.
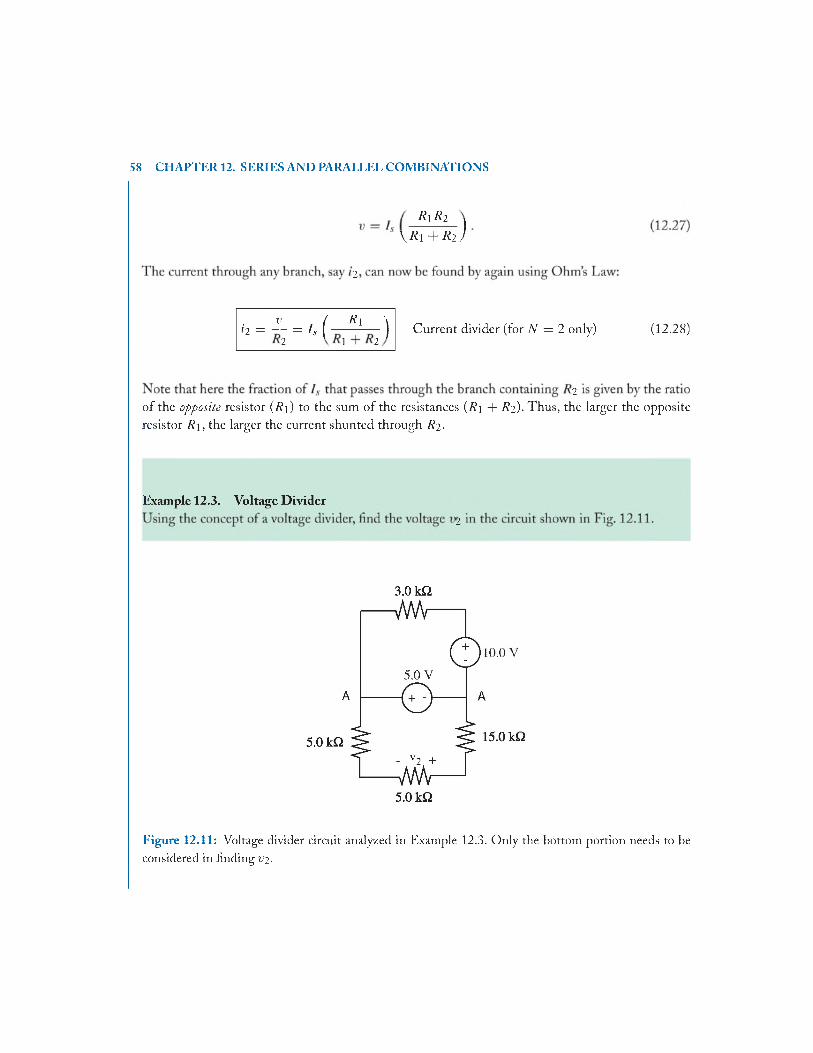
58 CHAPTER 12. SERIES AND PARALLEL COMBINATIONS
R i R 2 j
\ ^ + ^ 2 /
2
fe = JL = , , ( _ 5 i _ ' ) 2 2 i 2
Current divider (for N = 2 only) (12.28)
2of the opposite resistor (R i) to the sum of the resistances (R i + R 2). Thus, the larger the opposite resistor R i , the larger the current shunted through /?2-
Example 12.3. Voltage Divider2
3.0 k£2
Figure 12.11: Voltage divider circuit analyzed in Example 12.3. Only the bottom portion needs to be considered in finding t'2 -

The voltage source across the terminals marked A -A is an independent voltage source with a value of —5.0 V. Notice that it has a negative sign when it is referenced to the positive side of the voltage v i . This source voltage is also across the three resistors at the bottom of the circuit. (The upper part of the circuit does not affect the voltage across A -A since the 5.0 V source is a constant voltage source.)
ican be found easily using (12.26):
Tr / 5.0 \ / 5.0 \v i = - 5 .0 V ------------------- --------------- — = -5 .0 V ----- = -1 .0 V.i
12.8. PROBLEMS 59
Solution
Example 12.4. C urren t D ivider1
Figure 12.12: Current divider circuit analyzed in Example 12.4.
Solution1
The sum of the two resistances is 3.0 k£2. Thus, using (12.28) and the reasoning following it,
/ 2.0 k£2 \ A / 2.0 \ i\ = 6.0 mA I --------------------— I = 6.0 mA I — I = 4.0 mA.
\ 1.0 k £2 + 2.0 k£2/ \3 .o y
12.8 PROBLEMS12.1. a. Calculate the equivalent resistance across the terminals X-X in the circuit on the left of
Fig. 12.13.[ans: R eq = 16.8 k£2]
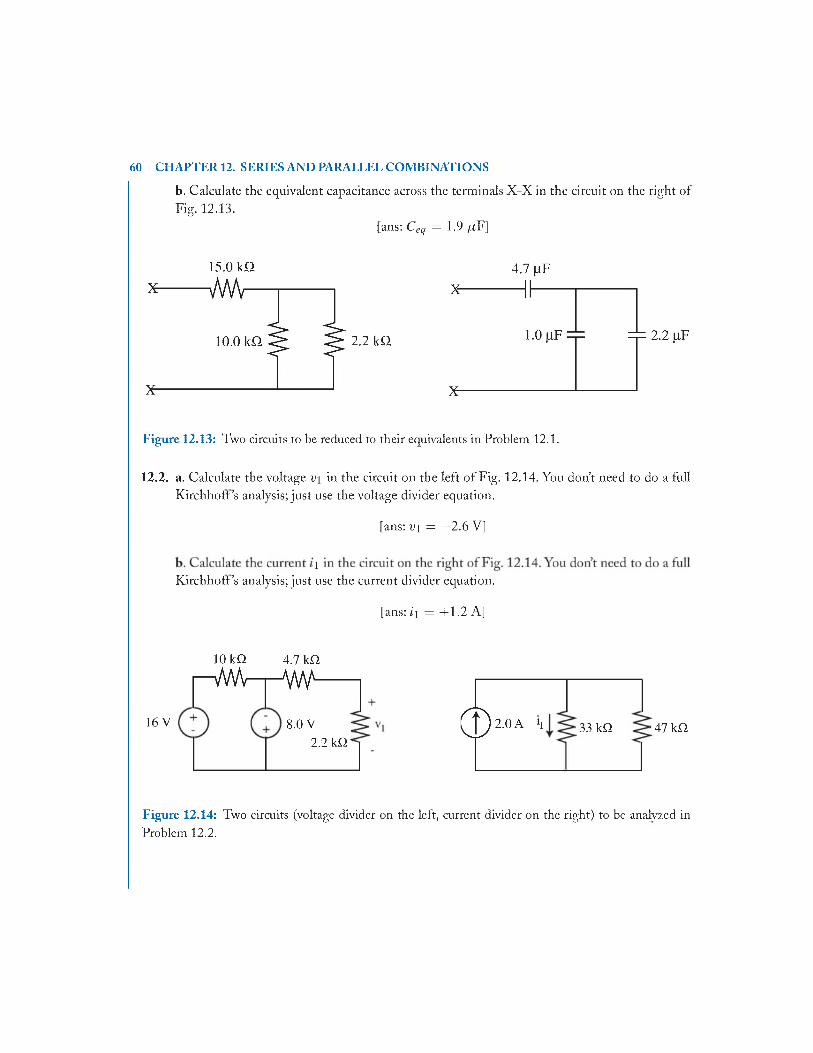
60 CHAPTER 12. SERIES AND PARALLEL COMBINATIONS
b. Calculate the equivalent capacitance across the terminals X-X in the circuit on the right of Fig. 12.13.
fans: Ceq = 1.9 /iF]
15.0 k a 4.7 (iF
2.2 k Q
Figure 12.13: Two circuits to be reduced to their equivalents in Problem 12.1.
12.2. a. Calculate the voltage i>i in the circuit on the left of Fig. 12.14. You don’t need to do a full Kirchhoff’s analysis; just use the voltage divider equation.
fans: v i = —2.6 V]
iKirchhoff’s analysis; just use the current divider equation.
fans: i i = + 1 .2 A]
10k£l
— VWV-4.7 kQ.
A W -
16 V 8.0 V ;2.2 k i r
© 2.0 A ii ■ 33 kQ. ■47 kQ
Figure 12.14: Two circuits (voltage divider on the left, current divider on the right) to be analyzed inProblem 12.2.

C H A P T E R 1361
a n d F i r s t - 1
C o n s t a n t s
13.1 THEVENIN EQUIVALENT CIRCUITSIt is possible, and often convenient, to transform a source from one form into another without changing its effect on the remainder of the circuit at all. One example o f this is shown in Fig. 13.1. On the left is an example of a source consisting of an independent current source in parallel with a source resistance R s (this arrangement is commonly called a Norton source) and connected to a load resistor R i . On the right, we have transformed the source into a Thevenin equivalent circuit, which consists of an independent voltage source in series with the same value of source resistance R s . The value of the Thevenin voltage source is Vs = ISR S.
Norton T hevenin
----------------------------------* — equivalent — w — *—+ to Rs
Q vs = isRs
Figure 13.1: The Thevenin circuit on the right is equivalent to the Norton source on the left.
To show that this transformation has no effect on the voltage or current going to the load resistor, let’s find the current coming from the Norton source (on the left in Fig. 13.1) into R l . The load resistor and the source resistor are in parallel, so R eq = R i R s/ { R i + R s). Since the current into the parallel combination is fixed at Is , the voltage v i is
= = ( m > Now consider the Thevenin equivalent circuit on the right in Fig. 13.1. The resistors are now
in series, so the voltage across the load can be found from the voltage-divider formula:

62 CHAPTER 13. THEVENIN EQUIVALENT CIRCUITS
V L = VsR l
IsR l R s (13.2)
R l + R s J \ R l + Rs . which is the same value as found in (13.1) using the Norton source.Thus, the voltage (and current by Ohm ’s Law) across the load resistor is unchanged by the source transformation.
Actually, this principle is much more general. The transformation of a source into a simpler Thevenin equivalent can be done for any linear source, no matter how complex, and the transformation will not change the state of the remaining circuit, no matter how complex. This is shown schematically in Fig. 13.2 for any source (circuit A) and any load (circuit R).
a------*------—► +
any i anycircuit V circuitA B
------x-----b
can be rcplaccd
Figure 13.2: Replacing a general source by its Thevenin equivalent circuit.
Since the Thevenin equivalent circuit contains only two elements (an independent voltage source Vj/, and a series resistor /?//,), it only takes two steps to find the Thevenin equivalent for a general circuit: first determining V//7 and then determining /?//,.The procedure is as follows:
Step 1. F ind ing V//7 - I f Thevenin’s principle is true for any circuit R in Fig. 13.2, it will be true if circuit R is an open circuit, as shown in Fig. 13.3. In Fig. 13.3, no current can flow through R th since it ends in an open circuit. Thus, there is no voltage drop across /?//, for this situation, and Vth = voc. So Step 1 can be summarized:
Step 1 - L eave the terminals o f the original circuit open a n d fin d the voltage across them. This open-circuit voltage, voc , is the Thevenin voltage; therefore V//7 = voc.
Step 2. F ind ing R th - Here it is necessary to look back into the original circuit at its open terminals and calculate the equivalent single resistance appearing across those terminals. Refore doing so, you must deactivate all independent sources.This means setting all independent voltage sources to a short circuit (since theirvoltage does not depend upon current) and setting all independent current sources to an open circuit (since their current does not depend upon voltage).Then the remaining resistances can be combined into a single equivalent resistance, which is /?//,. So Step 2 can be summarized:
Step 2 - D eactivate the independent sources (see Table 13.1) and look back into the original circuit at its open terminals. Combine the resistances into one equivalent resistance R eq. Then
Rfh = Req-

13.2. FXECTRICAL BEHAVIOR OF CAPACITORS 63
open oc circuit
Figure 13.3: The open-circuit voltage is the same as theThevenin voltage.
Table 13.1: Deactivating Sources Independent voltage source —»• short circuit Independent current source —»• open circuit
13.2 ELECTRICAL BEHAVIOR OF CAPACITORSResistors and capacitors behave much differently from each other with regard to their voltage and current interactions. For resistors, Ohm ’s Law (v = iR ) shows a direct proportional relationship between the current through the resistor and the voltage across it. Capacitors, on the other hand, follow a more complex relationship. From (11.9) in Coulomb’s Law chapter, in differential form the current-voltage relationship is
- = 03.3)d t C
showing that the tim e rate-of-change of voltage, not the voltage directly, is proportional to the current in a capacitor. Figure 13.4 indicates the polarities of the current through a capacitor and the voltage across it.
Figure 13.4: Current and voltage polarities for a capacitor.
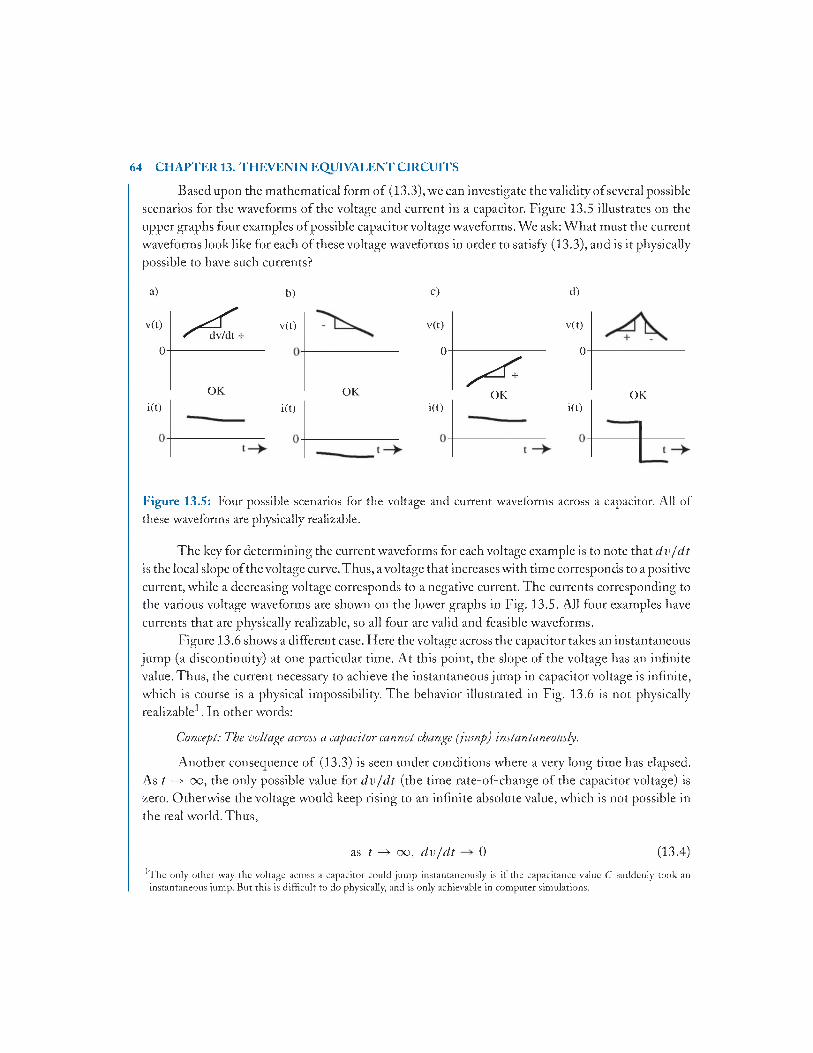
Based upon the mathematical form of (13.3), we can investigate the validity of several possible scenarios for the waveforms of the voltage and current in a capacitor. Figure 13.5 illustrates on the upper graphs four examples of possible capacitor voltage waveforms. We ask: W hat must the current waveforms look like for each of these voltage waveforms in order to satisfy (13.3), and is it physically possible to have such currents?
64 CHAPTER 13. THEVENIN EQUIVALENT CIRCUITS
b)
vft)
OK
d)
vft)
0-
i(t) i(t) i(t)OK
if t)OK
Figure 13.5: Four possible scenarios for the voltage and current waveforms across a capacitor. All of these waveforms are physically realizable.
The key for determining the current waveforms for each voltage example is to note that d v / d t is the local slope ofthe voltage curve.Thus, a voltage that increases with time corresponds to apositive current, while a decreasing voltage corresponds to a negative current. The currents corresponding to the various voltage waveforms are shown on the lower graphs in Fig. 13.5. All four examples have currents that are physically realizable, so all four are valid and feasible waveforms.
Figure 13.6 shows a different case. Flere the voltage across the capacitor takes an instantaneous jump (a discontinuity) at one particular time. A t this point, the slope of the voltage has an infinite value. Thus, the current necessary to achieve the instantaneous jump in capacitor voltage is infinite, which is course is a physical impossibility. The behavior illustrated in Fig. 13.6 is not physically realizable1. In other words:
Concept The voltage across a capacitor cannot change (jump) instantaneously.
Another consequence of (13.3) is seen under conditions where a very long time has elapsed. As t —*• oo, the only possible value for d v / d t (the time rate-of-change of the capacitor voltage) is zero. Otherwise the voltage would keep rising to an infinite absolute value, which is not possible in the real world. Thus,
as f -> oo, d v / d t 0 (13.4)
1The only other way the voltage across a capacitor could jump instantaneously is it the capacitance value C suddenly took an instantaneous jump. But this is difficult to do physically, and is only achievable in computer simulations.

13.3. RC TIM E CONSTANTS 65
Figure 13.6: An instantaneous jump in capacitor voltage requires an infinite current, which is not physically possible.
for a capacitor. Using (13.4) in (13.3) shows that the current into the capacitor must also go to zero for long times. This means that the capacitor acts like an open circuit. In other words:
Concept: A fte r a long tim e has elapsed (between changes in the source voltage j, a capacitor acts like an open circuit.
The two concepts just presented will greatly facilitate the analysis of circuits containing capacitors. But note carefully: These concepts apply only to capacitors and capacitor voltages, not to other electrical quantities in a circuit. It is possible, for example, for the current into a capacitor to jump instantaneously. Also, the voltage across a resistor can change instantaneously. In fact, they often do.
13.3 RC TIME CONSTANTSW hen a capacitor is connected with a resistor to a source, a simple “R C ” circuit is formed, as in Fig. 13.7. I f the source voltage v \ changes abruptly (which is possible; remember that it is a source voltage, not the voltage across a capacitor), then the output voltage, labeled i>(0> changes more slowly. Being the voltage across a capacitor, v( t ) cannot change instantaneously. Rather it moves in an exponential manner from its initial value to some final value. Let the source voltage v\ jump abruptly from a value of Va to a value of Vg at some point in time, as indicated in Fig. 13.7. We will assume that the source has been at the original value Va for a long time prior to its jump. We can also assign the time of the jump to occur at t = 0, since the time reference is arbitrary.
L et’s analyze the circuit in Fig. 13.7 using Kirchhoff’s and Ohm ’s Laws. Using KVL around the loop,
— v \ + i R + v { t ) = 0. (13.5)

66 CHAPTER 13. THEVENIN EQUIVALENT CIRCUITS
V B
VA
Tt = 0 t — ►
- w -
\ c 4= v(t)
< '
+
Figure 13.7: A simple RC circuit for analyzing the time behavior of the voltage across the capacitor when the source voltage vi changes.
(Notice that we have written the voltage across the capacitor as v( l ) to show explicitly that it is a function of time.) Putting (13.3) into (13.5) and rearranging results in
v( t ) + R C d v / d t = v \ . (13.6)
Equation (13.6) is a first-order differential equation describing the behavior of this circuit. You will get plenty of experience in solving equations such as this in later courses. Here we will just offer the general solution to this equation, without proof, and then use it in examples.
The general form of the solution to (13.6) is
v ( t ) = (Vj — V f ) e !^T + V f (13.7)
where
and
Vi is the initial value of v( l ) (at t = 0), same as i>(0), v f is the final value of v( l ) (as t —*• oo), same as v(oo ), x is the first-order time constant for voltage.
Substituting (13.7) back into (13.6) gives
R C Time Constant (13.8)
so the time constant of the circuit is given by the product of the resistance and the capacitance2.L et’s check to make sure that the designations Vj for initial value and V f for final value are
correct in the formula of (13.7).
Initial value: W hen t = 0, e t! r = e° = 1, so (13.7) becomes
u(0) = (Vi - v f ) 1 + v f = Vi, (13.9)
'■Note that the exponential nature o f the voltage waveform is identical to that found in Chapter 6 (Part I) for the pressure in a resistive and compliant fluid system [see (6,15)]; this is not surprising since the differential equation is the same. The fluid time constant is also o f identical form, being the product o f the resistance and the compliance [see (6.16)].

which shows that the value of v( l ) as t —*• oo is the final value Vf , as the subscript f indicates.
In order to plot the waveform of (13.7) for the source voltage of Fig. 13.7, we need to find values for Vj and v f . Here the concept of a capacitor acting like an open circuit after a long time has elapsed will help greatly.
1. F ir s t fin d Vj. Since the voltage v \ has been at Va for a long time before 1 = 0 , the capacitor looks0 0
0
2. N o w fin d v f . Since the final voltage is approached by waiting a very long time (actually many time constants) after the source voltage has jumped, the capacitor again looks like an open circuit. This
0v j = Vb -
We can now plot Equation (13.7). The waveform is shown in Fig. 13.8. Note that v ( t ) is0 0
0 0
closer and closer to Vb for longer times, but the rate of rise decreases as v ( t ) approaches Vb A measure of how rapidly the voltage is changing is given by the value of the RC time constant
r . A t a time that is one time constant after the jump in the source voltage (that is, at t = r) , (13.7) gives
d ( t) = (vj — V f ) e ^ ^ T + v f = { Vi — V f ) e ^ 1 + Vf = Vf — Vj + Vj — V f e ^ l + V j e ^ 1 = Vi + ( V f - V i ) ( 1 - e ^ 1 )
= Vi + (V f - Vi)(0.63). (13.11)
Since Vj is the starting value of v( l ) , and ( v f — Vj) is the extent of the change (the “excursion”) in v( l ) , then (13.11) leads to the following conclusion:
0
This is shown graphically on the plot in Fig. 13.8. We will use the general formula (13.7) to solve a slightly more complex RC circuit containing two resistors in next example.
J Afterfiv e time constants it — 5 t), the voltage has gone 99.3% of its way to the final value.
13.3. RC TIM E CONSTANTS 67
0
0
0
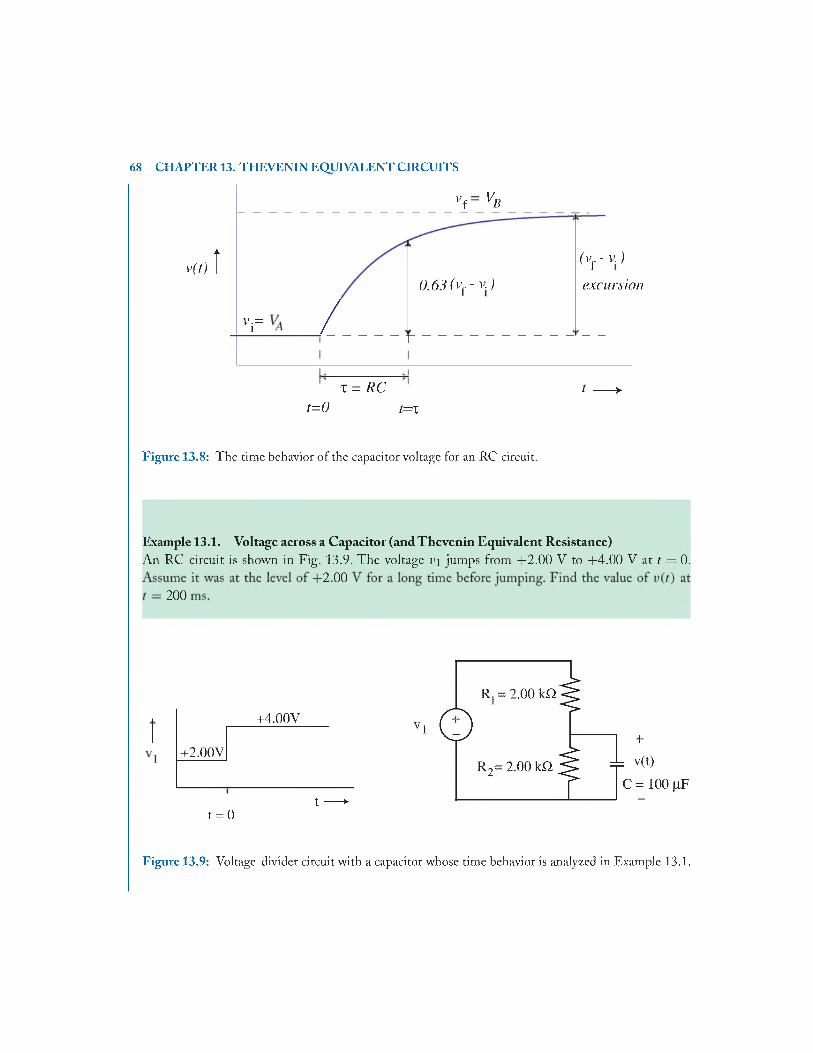
68 CHAPTER 13. THEVENIN EQUIVALENT CIRCUITS
v f “ VB
A
v(t)
v i= /
0.63 (vf - v.)
f
(vf - v . )
excursion
% = R C
r=0 t=xt — >-
Figure 13.8: The time behavior o f the capacitor voltage for an RC circuit.
Example 13.1. Voltage across a Capacitor (and T hevenin Equivalent Resistance)An RC circuit is shown in Fig. 13.9. The voltage vi jumps from +2.00 V to +4.00 V at t = 0.
2 00 200
+4.00V
+2,00 V
t = 0t — ►
Figure 13.9: Voltage-divider circuit with a capacitor whose time behavior is analyzed in Example 13.1.

13.3. RC TIM E CONSTANTS 69
We always start by writing the general formula (13.7) for the voltage across a capacitor when the0
Solution
v( t ) = (vj — Vf ) e >/ T + Vf . (13.7)
The task is to find the values of Vj , V f , and r for the particular circuit being analyzed.
2 00 00
Figure 13.10: The state of the circuit just before t = 0.
This is a simple voltage divider. Using the voltage-divider formula,
„ = = B .0 0 V ) ( 2 0 ^ = i .0 0 v .1 \ R i + R 2 j V 4.00k£2/
0 i 4 00
2 00
2 00 v f = (400 V) I — = 2 0 0 V.
4 00
F in d t :This is more complicated than the earlier example in Fig. 13.7 since we have tw o resistorsi 2
resistance at the terminals of the capacitor. We deactivate the independent voltage source (replace it with a short circuit) and look back into the circuit at the terminals o f the capacitor. This situation is shown in Fig. 13.11Note that the resistors are in parallel, so the equivalent resistance is
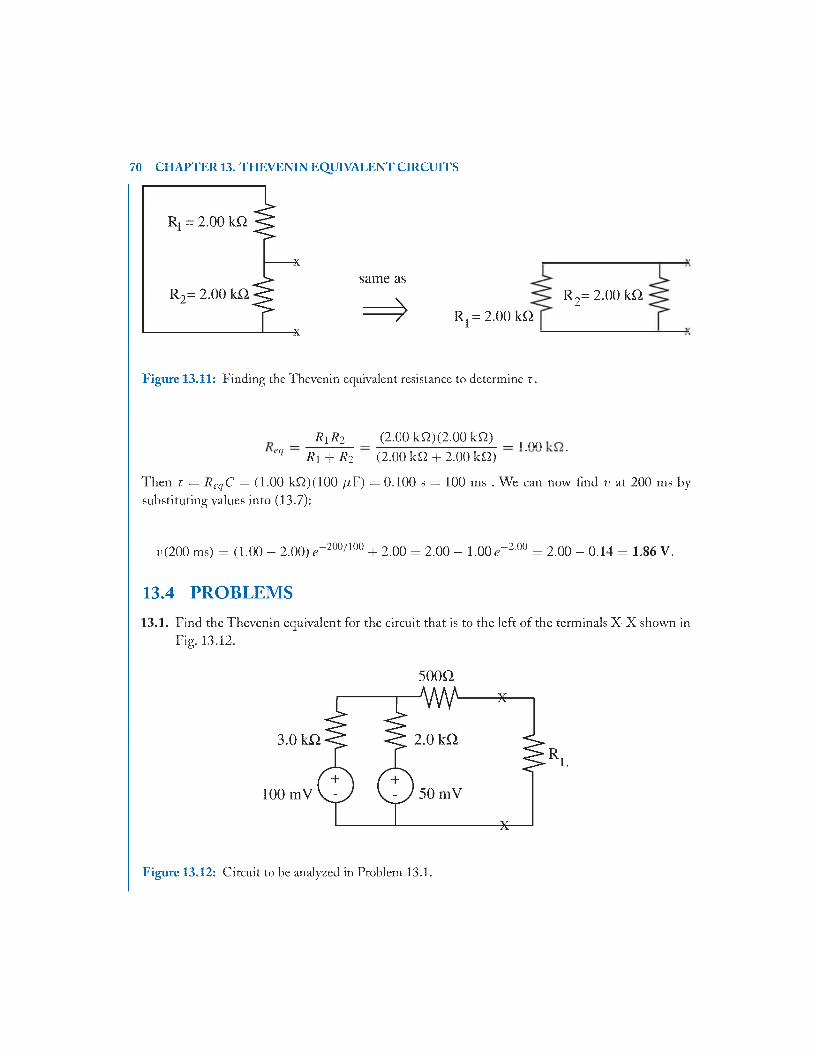
70 CHAPTER 13. THEVENIN EQUIVALENT CIRCUITS
same as
*R 2= 2.00 k Q
R i = 2.00 k Q
Figure 13.11: Finding the Thevenin equivalent resistance to determine r.
_ R i R 2 (2.00 kQ)(2.00 kQ) _ 1 00 eq R i + R 2 (2 0 0 k Q + 2 0 0 k£2) ' '
Then x = R eqC = (1.00 kQ)(100 /xF) = 0.100 s = 100 ms . We can now find v at 200 ms by substituting values into (13.7):
i>(200 ms) = (1.00 - 2.00) <^200/100 + 2.00 = 2.00 - 1 .00e^2'00 = 2.00 - 0 1 4 = 1.86 V.
13.4 PROBLEMS13.1. Find the Thevenin equivalent for the circuit that is to the left of the terminals X-X shown in
Fig. 13.12.
soon
R L
Figure 13.12: Circuit to be analyzed in Problem 13.1.

13.4. PROBLEMS 71
fans: see Fig. 13.13]
Figure 13.13: Answer to Problem 13.1.
13.2. (Lab Experim ent) In the lab, set up the following circuit using a prototyping board (Fig. 13.14). 1
1
from function generator center connector
(red) +
from function generator ground connector
(black)
2.2 k Q.
"VWV—100 J.IF
+v-,
+ 15V
-15V
LF353
to oscilloscope probe center (channel 1)
to oscilloscope probe center (channel 2)
to oscilloscope probe ground wire
v l
(V)
250 500 750 1000 1250
t (ms)
1500
------ ►
Figure 13.14: Circuit to be set up in lab for measuring voltages.

voltage follower and the second channel of the oscilloscope to simultaneously observe the voltage V2 . By hand and with a ruler, plot both the v \ voltage waveform and the V2 voltage waveform on the same graph axes as a function of time for a cycle or two. Label the vertical axis in units of V, but also label the vertical axis (that is, use a dual label) with “equivalent” blood pressure— assuming that 10 V is equivalent to 100 m m Hg of pressure.
Use a sheet of green ‘engineering’ paper for this graph (it will be provided). Follow proper graphing techniques when doing this graph; points will be deducted from your score if proper techniques are not used. (The graph is all you need to turn in as homework for this problem, one graph per student.)
13.3. In Problem 13.2 the function generator charged a single capacitor through a resistor when its voltage Dx was at its high value of +10 V, then discharged it when the voltage was 0 V. The
2
a. Assuming the voltage V2 at the beginning of the charging period was +0.30 V, calculate the voltage at the end of the charging period.
fans: 1)2 at 0.250 s = +6.9 V]
b. Compare your answer in part a with the value you actually measured and plotted in Problem 13.2. I f they are not identical, give one reason why.
13.4. The voltage V\ in the circuit below has been +10 V for a long time, then drops to +5.0 V at t = 0, as shown in Fig. 13.15. Find the voltage V2 at ? = 35 ms.
fans: i>2(35 ms) = +2.9 V]
72 CHAPTER 13. THEVENIN EQUIVALENT CIRCUITS
33 kO
10 V
■5.0 V
t = 0 t — ►
Figure 13.15: Circuit to be analyzed in Problem 13.4.
v 21
0

C H A P T E R 1473
N e r n s t P o t e n t i a l : C e l l
M e m b r a n e
14.1 INTRODUCTIONWe have seen in the Darcy’s Law chapter how important the cell membrane is to the proper functioning of the cell in its various roles in the body. An excellent example is the neuron, the cell of the nervous system, which triggers, controls, and transmits signals to and from all parts of the body. The neuron uses action potentials— electrical impulses that travel down the length of the axon of the neuron— to transmit signals within the interconnected network of the nervous system. The ion permeability properties of the neuron produce the membrane’s electrical state, and changes in that electrical state initiate and propagate the action potential. Modeling the cell membrane will employ of many of the electrical concepts covered in the previous chapters, in particular Kirchhoff’s Laws (Chapter 9), the Thevenin equivalent circuit (Chapter 13), and the behavior of capacitors in circuits (Chapters 11 and 13).
14.2 CELL MEMBRANE STRUCTUREThe membrane of the neuronal cell, like that of all cells, is not an impervious barrier, as first discussed in Chapter 2 of Part I. Nutrients can enter the cell and waste products can be excreted; otherwise the cell would die. Moreover, the membrane selectively allows various ions (charged atoms or molecules) to cross its walls, setting up an electrical potential (voltage) across the membrane. This voltage, vm , is a key element in the production of action potentials in neurons.
Figure 14.1 is a drawing of the cross-section of the cell wall of a typical neuron1. The microstructure of the membrane reveals two major features responsible for its selective permeability to ions and other molecules:
1. Channels, - These are tiny water-filled pores formed by membrane-spanning proteins that extend across the thickness of the membrane, allowing movement of ions from inside to outside the cell, and vice versa. They generally are partially selective to passing a specific ion or ion type, selecting either by size or by charge. For example, there are potassium-ion (K+ ) channels, sodium-ion (Na+) channels, and chloride-ion (Cl- ) channels.The channels, in turn, can be put into two categories:
11n drawings in this book, the membrane segment is oriented such that the region above the membrane is inside the cell while the region below is outside the cell. This is because the voltage outside the cell is usually set to be the reference (ground) voltage, and by convention, circuit diagrams usually have the ground voltage located at the bottom of the diagram (see Fig. 14.6, for example).

74 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT
© ©inside cell
+ + gated Na1 channel
carrier protein: Na-K pump
[Na ]o(greater)
© ©
[Cl ] ./greater)
gated K 4 channel
outside of cell
Figure 14.1: Stylized drawing of the channels (open and gated) and carrier proteins responsible for transporting ions across the cell membrane. Only a section o f the neuronal cell wall is shown here.
a. Open, or passive, channels - These channels are almost always open to allow ion passage (at least for the species of ions that the channel is selective for).There is, however, some resistance that must be overcome as the ions pass through the channels.
b. Gated, or active, channels - These channels change their state (with electrical, mechanical, or chemical stimulus) from being either open or closed to the opposite, thereby modulating the flow of the specific ions. For instance, gated N a+ channels that open when the membrane voltage reaches above a threshold value are responsible for generation o f neuronal action potentials.
2. Carrier proteins, or ion pum ps - Some of the proteins that span the membrane actively transport specific ions across the membrane barrier. These ion pumps consume metabolic energy to transport the ions against the forces that are simultaneously causing the ions to flow through open channels in the opposite direction. The carrier proteins are specific for a particular ion or pair of ions. For example, there is a Na+ -K+ pump in the membrane of neurons that transports Na+ ions out of the cell while at the same time transporting K+ ions into the cell.

14.2.1 MECHANISMS OFTRANSPORTThere are three main mechanisms that can cause the transportation of ions and other molecules across membranes:
1. Hydraulic pressure-driven transport - We have covered this topic in the Darcy’s Law chapter. Pressure-driven transport plays an important role in regulating blood volume in the capillaries and water balance in the kidneys, but it is normally not involved in the flow of ions across cell walls, and will not be considered further here.
2. Electro-chemical diffusion-driven transport - This is the major determinant of ion transport across cell walls. As the name implies, there are two components:
a. Electrically-driven flow - Coulomb’s Law describes the electrostatic force that causes like charges to repel and unlike charges to attract. I f there is an excess accumulation of one species of ions on one side of the membrane, such as shown in Fig. 14.2, an electric field will be set
electric field
14.2. CELL MEMBRANE STRUCTURE 75
..... * © — O n Ou o
+ V -
drift current----- ►
charges © ©
© ©
Figure 14.2: An accumulation of charge sets up an electric field, causing a drift current of ions toward the opposite charge.
up across the membrane which exerts a force on similarly charged ions, pushing them in a direction across the membrane away from the accumulation and toward the oppositely charged ions. This flow of ions under the influence of an electric field is termed drift current.
b. Concentration-driven flow - Due to the random motion of atoms and molecules at a finite temperature (Brownian motion), whenever there is a non-uniform concentration of any species within a volume, such as shown in Fig. 14.3 for a concentration difference across a membrane, the species tends to spread out to eventually produce a uniform concentration. Fick’s Law describes the flow of atoms or molecules under the influence of a concentration gradient. It applies to charged ions as well. This flow of ions is termed diffusion current.
3. A ctive Transport - This mechanism describes the transport of ions or other molecules across the membrane by the carrier proteins, as discussed in the previous section. I t is responsible (in combination with electro-chemical diffusion) for setting up and maintaining ionic and other concentrations

selectively permeable
76 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT
membrane
^ diffusion current *
/ 1
^ [C1‘
f ^ ^ -
■ J ' - V f^ — V
[ C ] 2 ^ ^
/ / [ 1
beaker
Figure 14.3: The higher concentration of C molecules on the right side of the beaker will result in a diffusion current across the selectively permeable membrane. The current is proportional to the concentration gradient (i.e., the difference between the concentration of the right side [C]2 and the left side[C]i).
across the membrane. Active transport requires metabolic energy, since often the movement is against voltage or concentration gradients.
14.3 NERNST POTENTIAL
Consider the situation shown in Fig. 14.4, representative of the ion distribution for a simple cell with channels selective to only one ion species. The fluid inside the cell (the intracellular fluid, or cytoplasm) is separated from the fluid outside the cell (the extracellular fluid) by the selectively permeable membrane. A t least one species of ions (for example, K+) has unequal concentrations in the two compartments— a higher concentration of K+ inside than outside the cell, say. The open channels for K+ allow ion flow in both directions depending on the prevailing forces. This means that there is a diffusion current flowing through the channels from the side that has the higher concentration (the inside) toward the lower concentration (the outside).
There is also a background of negatively charged ions (Cl- if KC1 was the original compound dissociated in the fluid) whose concentration almost balances the concentration of K+ in the two regions to maintain approximate charge neutrality in each region. But there is a slight excess of C l- ions (not balanced by K+ ions) in a layer immediately next to the inside of the membrane, and an equal slight excess of K+ ions (not balanced by Cl- ions) in a layer immediately outside

14.3. NERNST POTENTIAL 77
© inside cellopenK channels [K lj (greater)
membranevoltage
+v„
+ + +
Figure 14.4: Example of a simple membrane with open K+ channels. The concentration of K+ inside the cell [K+ ](- is larger that on the outside [K+ ]„ causing a diffusion current o f K+ out o f the cell. However, positive charges on the outside of the membrane produce an electrical force that causes a drift current of K+ into the cell. W hen the membrane voltage vm is at the Nernst potential, these opposing currents just cancel.
the membrane. This slight imbalance is initially set up by the diffusion current described in the previous paragraph. In turn, this excess charge causes a negative voltage vm across the membrane. The magnitude of the unbalanced charge is tiny compared to the total charge in each region (less than 1 part in 10,000), but its effect in producing vm is major. (Note that the reference polarity of vm is taken with respect to the outside fluid, as shown by the + / — signs of vm in Fig. 14.4. A negative vm then means that the inside voltage is lower than the outside voltage.)
The presence of the membrane voltage means there is an electric field whose force tends to push K+ ions from the outside to the inside of the cell. This K+ drift current will flow through the open K+ channels, opposing the diffusion current in the other direction. A t a certain membrane voltage, called the N ernst potential, the two opposing currents will just cancel, so the net K+ current is zero and the cell is in equilibrium with respect to K+ ions. This special voltage is given (for K+ ) by the formula2
’This formula will be derived formally later in courses in Biology, Biophysics, or Physiology. It is sometimes written in a form where the ratio in the argument o f the In function is inverted, with the outside concentration in the numerator and the inside concentration in the denominator; the negative sign then disappears.

78 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT
VkK T [K+l
------1n — -j—q [K+l
Nernst Potential for K+ (14.1)
where K is Boltzmann’s constant,T is temperature in Kelvin, q is the charge of the ion, including sign,1n is the natural logarithm,[K+l; is the concentration of the K+ ion inside the cell, and [K+l,, is the concentration of the K+ ion outside the cell.
26
[K
at room temperature. For N a+ ions, the Nernst equation is
fK+ l; Tr ,VK = - 2 6 1n mV (14.2)
^N‘i — —2 6 1n -— T^-m V , (14.3)[Na 1„
and for C l- ions, the Nernst equation is
r c r i -y _ _j_261n--------- mV (14.4)
[CU !,,
where the positive sign is due to the negative charge of the chloride ion.It is important to note that the Nernst potential for any ion depends on the two concentrations
of the ion across the membrane, and will change if the concentration changes. Also, the Nernst potential is not usually the actual voltage vm across the membrane, especially when there are two or more ion species for which the membrane is permeable, as will be seen shortly. This is because vm is influenced by other factors in addition to the Nernst voltages, including the permeability values for each ion species and the existence of ion pumps in the membrane. Examples of finding the actual membrane voltage using the membrane equivalent circuit are given in the next section.
14.4 EQUIVALENT CIRCUIT FORTHE MEMBRANEFor a simple membrane such as shown in Fig. 14.4 that has only channels for one species of ions, an equivalent circuit can be constructed like Fig. 14.5. It consists of an independent voltage source of Nernst potential magnitude, V k , in series with a resistor representing the electrical resistance of the channels to K+ -ion flow. The top terminal is at the potential of the intracellular fluid, while the bottom terminal is at the potential of the extracellular fluid, usually taken at zero voltage as the reference.

+
v = V„m K
14.4. EQUIVALENT CIRCUIT FOR THE MEMBRANE 79
inside
at equ ilib rium / +V K N ern st po ten tia l
ooutside
Figure 14.5: Equivalent circuit for the ion flow across a simple membrane (such as Fig. 14.4) with channels permeable only to K+ ions.
Since K+ -ion flow provides the only current through this simple membrane, there are no other branches to its equivalent circuit. W hen the net current is zero (at equilibrium), there is no voltage drop across the resistor and the membrane voltage vm is equal to the Nernst potential V k - This single-ion equilibrium situation is one of the few cases where the membrane potential is equal to the Nernst voltage.
For a more general membrane, such as was shown earlier in Fig. 14.1 for a neuron, the equivalent circuit is more complicated. Each ion forwhich the membrane has channels is represented by a separate branch with its own Nernst voltage source in series with a unique resistor, as shown in Fig. 14.6. The upper wire represents the cytoplasm inside the cell (which has relatively high conductivity compared to the channels and whose resistance can therefore be ignored), and the lower wire represents the extracellular fluid (which also has high conduct! vity)J .
Although the + / — polarity of all the sources shown in Fig. 14.6 is oriented in a direction such that the reference voltage is at the bottom (i.e., outside the cell), individual Nernst voltages may have negative values depending upon the particular concentration differences and sign of the ion charge. Also, the values of the resistances can, and will, change as the respective gated channels open and close. In addition, to represent the capacitance of the membrane with its opposite charges separated across a barrier, a capacitor Cm has been added to the equivalent circuit in Fig. 14.6.
For the squid giant axon, a much-studied neuron, the values of the ion concentrations and equivalent resistances are listed in Table 14.1. The concentration values can be used to find the Nernst potentials for each ion using (14.2)—(14.4), and these are also listed in Table 14.1.
°The simple equivalent circuit shown in Fig, 14,6 assumes that the ion current through each branch is linearly related to the voltage across that branch, which is approximately correct for the channels in a squid giant axon membrane. O ther cells are better described by a slighdy more complex, and nonlinear, model defined by the Goldman-Hodgkin-Katz (or simply the Goldman) equation. However, the qualitative behavior o f the ion currents and membrane voltage is similar in both models.

80 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT
cj) ^ inside
+ '—
v rm —i- v-
outside
Figure 14.6: Equivalent circuit for the membrane of a typical neuronal cell, as illustrated in Fig. 14.1, which is permeable to three ions. Also included is a capacitor representing the capacitance of the membrane.
Table 14.1: Values for a Squid Giant Axon at Rest
^NemSt CffK+ 397 mM 20 mM 2.7 kQ -7 8 mVNa+ 50 mM 437 mM 30.0 kQ +56 mV 1.0 / iFc r 40 mM 556 mM 3.3 kQ —68 mV
The values from Table 14.1 can then be put into the equivalent circuit, as done in Fig. 14.7. A study of this figure shows that for the general case, the membrane voltage is not the same as any of the Nernst potentials, since in general current must be flowing through the resistors, even at equilibrium. The actual membrane voltage is found by a circuit analysis of the complete circuit; this will be done in an example next.
But first some observations can be made here: The magnitudes of the Nernst voltage sources are all different, and in fact, some have opposite signs. The overall membrane voltage will be determined, therefore, by the relative magnitudes of the series resistors. The smaller the resistance, the closer the membrane voltage will be to the Nernst voltage of that branch; the larger the resistance, the more isolated the membrane voltage is from the Nernst voltage of that branch. Since in Fig. 14.7 the K+ branch and the C l- branch have much smaller resistances (corresponding to larger permeabilities) than the Na+ branch, the membrane voltage will be negative, close to (but not quite as negative as) the Nernst voltage for K+ and for C l- . In fact, as will be seen in the example next, the resting membrane voltage for the squid giant axon is about —67 mV.

14.4. EQUIVALENT CIRCUIT FOR TH E MEMBRANE 81
Figure 14.7: Equivalent circuit with three ion channels for the squid giant axon at rest.
Also, a careful study of the equilibrium condition shown in Fig. 14.7 reveals a seeming inconsistency A t equilibrium, no net current flows out of the top of the complete circuit. This means that the current flowing down the K+ branch, composed of K+ ions flowing from inside to outside the cell, must equal the current flowing up in the N a+ branch, composed of Na+ ions flowing from outside to inside the cell. (Note: It will be seen shordy that the C l- current is nearly zero.) But these two currents carry different ions, and after a while, if left alone, these currents would lead to changes in the respective concentrations inside and outside the cell, a violation of the equilibrium condition. The answer to this enigma is that there is an active Na-K pump across the membrane— not shown for simplicity in the equivalent circuit but illustrated in Fig. 14.1— that pumps K+ ions into the cell and Na+ ions out of the cell, thus maintaining the concentrations at their equilibrium values.
Finally, a note about units. In practice, the magnitude of the membrane branch resistances for the total cell are often given in terms of conductance G , which is the inverse of the resistance R; so G = l / R , in units of l/ohm=siemens (S). Even more, the membrane conductance is usually
2G = g ■ A m, where A m is the total surface area of the membrane under consideration. (Similarly, the
2In this case, r = l / g and R = r / A m .) The capacitance Cm is also often given as a capacitance per
2Then if the branch conductance g (or resistance r) and capacitance cm values are specified per
unit surface area, they can be inserted into the equivalent circuit of Fig. 14.7 in one of two ways:
• Each g and cm value can be adjusted by multiplying it by the total surface area of the membranel
C m are pu t in the circuit; or

* The r = l / g and cm values can be inserted directly as they stand into the equivalent circuit, in which case the equivalent circuit represents a unit area of the membrane (and the branch currents are then found per unit area). This latter case is illustrated in a homework problem at the end of this chapter.
In either case the resulting membrane voltage and membrane time constant are the same.
82 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT
Example 14.1. Resting Potential Across Cell M em brane
For the membrane properties of a squid giant axon given in Table 14.1:a. Calculate the Nernst potential for each ion.b. Calculate the resting membrane voltage using only the K+ and N a+ branches.c. Find the RC time constant of the membrane.
Solutiona. Equations (14.2)-(14.4) can be used to find the Nernst potential at room temperature. Straightforward math gives the values shown in Table 14.1.The equivalent circuit is then given by Fig. 14.7.b .T o find the resting membrane potential, we redraw the equivalent circuit of Fig. 14.7 using only the K+ and N a+ branches. (The reason why the C l- branch is not included here will be explained shortly.) The capacitor is not included since it acts like an open circuit in the steady-state or resting condition. The redrawn circuit is shown in Fig. 14.8.
Figure 14.8: Equivalent circuit of squid giant axon membrane to find the resting membrane voltage.
The branch-current method can then be used to solve for vm. Using KVL around the loop labeled K T l gives

14.4. EQUIVALENT CIRCUIT FOR TH E MEMBRANE 83
- ( -7 8 mV) + i (2.7 t o ) + i (30 k£2) + 56 mV = 0, (14.5)
i = ( -1 3 4 mV)/32.7 k£2 = -4 .1 f iA . (14.6)
Using KVL around the loop labeled KT2 gives
30 56 0
Putting the value for i from (14.6) into (14.7) gives the resting membrane voltage:
vm = 56 mV + (—4.1 ^A )(30 k ^ ) = 56 mV — 123 mV = —67mV. (14.8)
Note that the value of the resting membrane voltage is very close to the Nernst potential for the C l- ions as given in Table 14.1. This means that there is essentially no current flowing through the C l- branch, and it has little effect on the remainder of the circuit. I t is therefore valid to exclude it from the equivalent circuit when calculating the membrane voltage. The basic reason for this is that C l- is passively distributed across the membrane by open channels only with no ion pump for C l- involved, and the C l- concentrations on both sides therefore adjust themselves to match the membrane voltage set by the K+ and Na+ concentrations.c. The time constant is given by the product of R eqC m, where C m is the capacitance (1.0 f iF) of the membrane and R eq is the equivalent resistance of the membrane at rest. This resistance is found by calculating the Thevenin equivalent resistance of the circuit as seen from the terminals o f the capacitor. The circuit includes all branches of the equivalent circuit. After disabling the independent voltage sources (replacing with shorts), the circuit looks like Fig. 14.9.
equiv. to
Figure 14.9: Finding the Thevenin equivalent resistance to determine the membrane time constant.
The equivalent resistance is the parallel combination of the three resistors, so
1 / R eq = 1 /2 .7 kto + 1/30 kQ + 1/3.3 k Q . (14.9)

Thus,R eq = l . 4 k Q , (14.10)
andr = R eqC m = 1.4 ms. (14.11)
Note: I f it remained constant, this RC time constant of 1.4 ms would be problematic (as shown in Chapter 13 and Fig. 13.8), since the voltage across the membrane capacitance, vm, cannot change at a rate faster than the time constant. In fact, it reaches only 63% of its excursion during one time constant and thus takes several time constants (perhaps 5 to 6 ms for a time constant of 1.4 ms) to accomplish an appreciable change. Yet nerve action potentials require a much faster rate of rise and fall than this; the complete rise and fall phases of the membrane voltage that comprise an action potential take less than about 3 ms. Luckily, as will be seen in the next example, the RC time constant of the membrane decreases significantly during the course of an action potential.
14.5 ACTION POTENTIALSThe situation shown in Table 14.1 and Fig. 14.7 holds for a resting axon only. W hen the cell is resting, the gated K+ and N a+ channels are in their resting position, having not been stimulated to change their state. But under stimulation through its synapse with neighboring neurons, the cell’s membrane voltage and the state of the gated channels change dramatically. W hen the membrane voltage is raised above a threshold (about —40 mV for the squid giant axon) by stimulation from adjacent cells, many gated Na+ channels open rapidly with a major effect. The resistance of the Na+ branch declines precipitously to a very low value, much lower than the resistance of the K+ branch. This rapidly moves the membrane voltage to a value close to the N a+ Nernst voltage, which is positive at +56 mV.
The gated K+ channels also begin to open, but at a slower rate than the N a+ channels. This delayed action moves the membrane voltage back from its positive value again toward the K+ Nernst voltage at —78 mV. The overall result is a voltage spike in the membrane voltage, called an action potential, which begins at the resting voltage, rises rapidly to a positive voltage as the N a+ channels open, then declines back toward the resting voltage (actually overshooting in the negative direction) as the K+ channels open and the Na+ channels close again. The K+ channels then close shortly after, returning the cell to its resting state ready for the next stimulation.
The situation leads to action potentials as plotted in Fig. 14.10. The upper graph shows the time behavior of the conductance G ofboth the K+ and Na+ channels just after the stimulus, when the membrane voltage rises above threshold. (It is easier to graph conductance G than resistance
lconductance of the Na+ channels increases rapidly, causing the membrane voltage— shown in the lower graph— to rise abruptly toward the positive N a+ Nernst potential. The rise in K+ conductance is delayed slightly, so by the time it starts to increase, the Na+ conductance is decreasing. This causes the membrane voltage to fall back toward the negative K+ Nernst potential, producing the declining
84 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT

14.5. ACTION POTENTIALS 85
Figure 14.10: W hen the gated Na+ channels rapidly open as the membrane voltage crosses the threshold value in response to a stimulus, the N a+ conductance Gjs;., rises (upper graph) and the membrane voltage rises (lower graph) toward the positive N a+ Nernst potential, starting the action potential. I t falls when the N a+ conductance Gjs;., falls and the K+ conductance G k rises, slightly later.
segment of the action potential. The increased K+ conductance during this phase actually results in a more negative voltage than at rest, so the declining segment dips below the resting value. It slowly returns to the resting potential when the K+ conductance declines to its resting value. This completes the cycle of the action potential, and the nerve is in a state to again be stimulated.
The generation of the action potential, with its accompanying transmission down the nerve axon, is the major electrical event responsible for the signaling and control functions of the nervous system. As we have seen, it is made possible by the unique ion permeability characteristics of the neuronal cell membrane.
Example 14.2. R ising Phase o f the A ction PotentialFor the squid giant axon analyzed in the previous example:a. Assume that the Na+ channels now open up rapidly upon stimulation such that the Na+ conductance rises to G Na = 0.12. Using the equivalent circuit from the previous example with a new

Figure 14.11: Equivalent circuit for estimating the peak voltage of an action potential.
Following steps similar to before, KVL around loop K T l gives
78 2 7 8 3 56 0
i = (-1 3 4 mV)/2.7083 kQ = -4 9 .5 f i A . (14.13)
Then KVL around loop KT2 gives
8 3 56 0
56 49 5 8 3 56 0 4
which is very close to the N a+ Nernst voltage of +56 mV. The peak voltage of the action potential50
Gnu ar|d an increase in G k ) cause it to drop back again, completing the action potential spike.

14.5. ACTION POTENTIALS 87
b. The new time constant is much shorter than its resting value due to the reduced resistance of the membrane. The new Thevenin equivalent resistance is given by
1 / R eq = 1/2.7 t o + 1/8.3 fi + 1/3.3 kfi, (14.16)
R eq = 8.25 fi, (14.17)
T = R eqCm = (8.25 fi)(1 .0 x 10^6F ) = 8.25 /as. (14.18)
This time constant (reached when G ^ a is at its maximum value) is now short enough not to restrict the rapid rise and fall of the action potential voltage, whose timing is really determined by the changes in ion conductance.
In obtaining the peak voltage during the action potential in the previous example, it was assumed that the N ernst voltages for Na+ and K+ did not change during the course of the action potential. But when the gated Na+ channels open up (and the delayed K+ channels also), ions of both species will cross the membrane at rates greater than at rest, potentially changing their concentrations inside and outside the cell, thereby upsetting the Nernst potentials, which are based upon these concentration values. It is shown next, however, that the relative concentration changes that occur during an action potential are very small compared to the resting concentrations.
We first find the number of ions that cross the membrane during a typical action potential rise where the initial membrane voltage is — 67 mV (resting value) and the peak voltage is +50 mV. The voltage change is A v = 50 mV — (—67 mV) = 117 mV, According to Eq. (11.6) and using
1 0
A q = C A v = (1.0 x 10“ 6 F)(117 x 10^3 V) = 1.2 x 10^7 C. (14.19)
Since the charge of each ion is q = 1.6 x 10-19 C, this means that (1.2 x 10_7)/(1 .6 x 10-19) = 7 5 1011 ions cross the membrane. This seems like a large number, except when compared to the number of ions already in solution. Assuming an equilibrium concentration of Na+ ions, for example, of 50 mM, a squid giant axon cell volume of about 1 x 10-5 L, and using the Avogadro constant, the number of N a+ ions inside the cell at equilibrium is
N = (molarity) (Avogadro constant) (volume in L)= (50 x 10_3)(6.02 x 1023)(1 x 10“ 5) = 3.0 x 1017. (14.20)
or about 3 parts per million, or 0.0003%, which is insignificant.
7 5 1011 3 0 1017) = 2.5 X 10 6

88 CHAPTER 14. NERNST POTENTIAL: CELL MEMBRANE EQUIVALENT CIRCUIT
14.6 PROBLEMS14.1. The table below gives the characteristics of a typical frog muscle cell. Note that the ion
conductance and membrane capacitance values are givenper u n it area:
Values for Frog Skeletal M uscle a t Rest
[conc], [conc]t, S ^NernstK+Na+c r
140 mM 13 mM 3.0 mM
2.5 mM 110 mM 90 mM
0.21 mS/cm2 0.023 mS/cm2 0.11 mS/cm2
2.1 /zF/cm2
a. Calculate the Nernst potentials at room temperature for each of the ions, and draw the equivalent circuit for the cell membrane at rest. Let this equivalent circuit represent a unit area of the membrane surface (i.e., 1 cm2). Remember that conductance g is the inverse of
1
T ans: VK = -1 0 5 mV, VNa = +56 mV, VC1 = -8 8 mV]
b. Using the equivalent circuit with only the K+ and Na+ branches, calculate the membrane voltage at rest.
89
c. Calculate the RC time constant of the membrane at rest.
61
14.2. M uscle Action Potential (M A P )
a. Now assume that the gated Na+ channels in the muscle cell of Problem 14.1 quickly open2
Calculate the resulting membrane voltage. (This represents the voltage toward which the action potential is rising. Use the equivalent circuit from Problem 14.1 again with only the Na+ and K+ branches but with the new r for the N a+ branch.)
50 2 54
b. Calculate the new RC time constant of the membrane under the condition of the new r for the opened Na+ channels.
fans: x = 0 .1 0 ms]

14.6. PROBLEMS 89
c. In 1-2 sentences, explain why is it essential for the membrane time constant found in this problem to be so much lower than that of Problem 14.1 (at rest) in order to allow a muscle action potential spike.


91
C H A P T E R 15
F o u r i e r T r a n s f o r m s :
C u r r e n t s ( A C ) a n d t h e
15.1 INTRODUCTION - SINUSOIDSMany of the voltages we have encountered so far have been constant, or nearly constant. An example is a battery modeled as an independent voltage source. Such steady sources are called dc signals, for “direct current” (even though they may be voltages or pressures, not strictly currents). But some important signals are time-varying, such as the aortic pressure pulse and the action potential in neurons. I t is useful to describe these time-varying signals not only by their time waveforms but by a complementary viewpoint: their frequency content. Signals that vary in time are called ac signals, for “alternating current.”
A very important and common time-varying waveform is the sinusoid. A voltage that has a sinusoidal time behavior can be written in general as1
v = V o Q m ( 2 7 r f t - 4 > ) , (15.1)
where cos is the cosine function,Vo is the peak voltage (V),/ is the frequency (hertz, or Hz, equivalent to 1/s), t is time (s), and<t> is a phase angle (radians or degrees, dimensionless).Figure 15.1 plots the sinusoidal waveform of (15.1) in two ways: (a) as a function of the
argument of the cosine term, and (b) as a function of time. The phase angle <t> represents the offset0
case when <t> = 7t/ 2 (or 90°), the cosine wave can be written as
v = Vo cos(2k f t - jt/2 ) = Vo sin (2 jr/f). (15.2)
In the time plot, the sinusoid repeats with period T . This occurs whenever its argument increases by 2x . So from (15.1), the relationship between period T and frequency f is 2 n f T = 2 n ,or
f = l / T . (15.3)
1 General sinusoidal waves, like (15.1), are usually called “sine” waves for short, even though they may be written in terms of cosines.
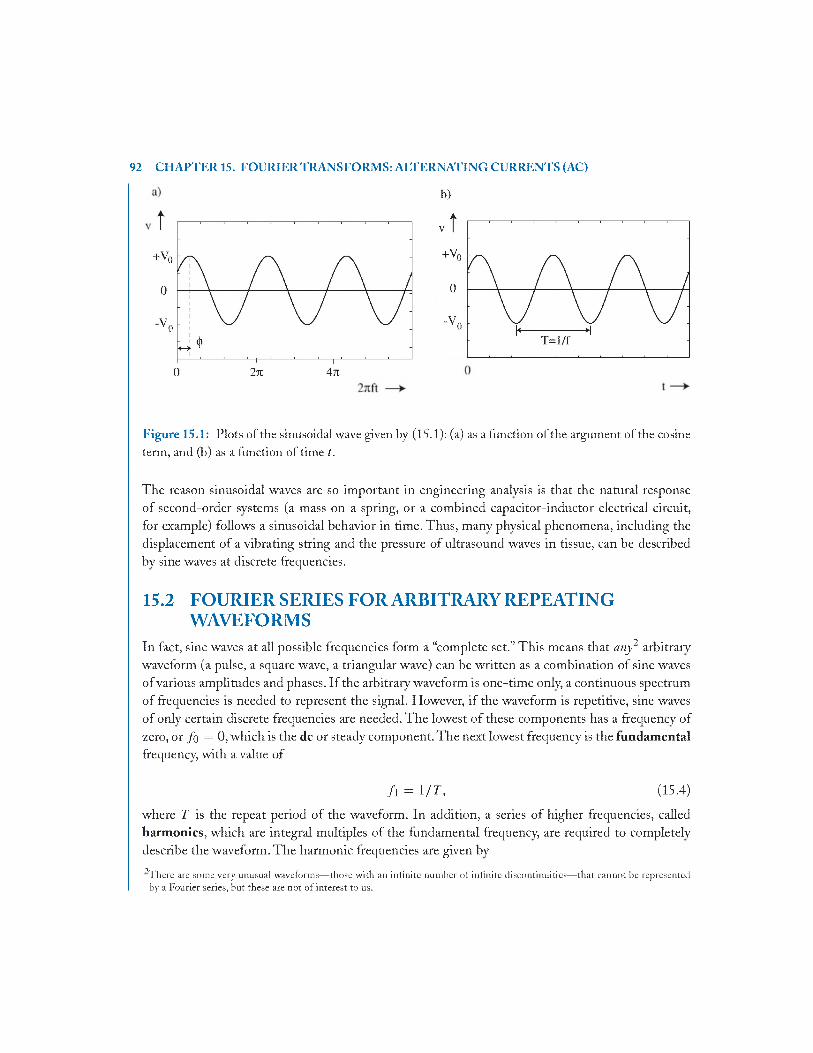
92 CHAPTER 15. FOURIER TRANSFORMS: ALTERNATING CURRENTS (AC)
Figure 15.1: Plots of the sinusoidal wave given by (15.1): (a) as a function of the argument of the cosine term, and (b) as a function of time t.
The reason sinusoidal waves are so important in engineering analysis is that the natural response of second-order systems (a mass on a spring, or a combined capacitor-inductor electrical circuit, for example) follows a sinusoidal behavior in time. Thus, many physical phenomena, including the displacement of a vibrating string and the pressure of ultrasound waves in tissue, can be described by sine waves at discrete frequencies.
15.2 FOURIER SERIES FOR ARBITRARY REPEATING WAVEFORMS
In fact, sine waves at all possible frequencies form a “complete set.” This means that any2 arbitrary waveform (a pulse, a square wave, a triangular wave) can be written as a combination of sine waves of various amplitudes and phases. I f the arbitrary waveform is one-time only, a continuous spectrum of frequencies is needed to represent the signal. However, if the waveform is repetitive, sine waves of only certain discrete frequencies are needed. The lowest of these components has a frequency of zero, or f o = 0, which is the dc or steady component. The next lowest frequency is the fundam ental frequency, with a value of
f l = 1 / r , (15.4)
where T is the repeat period of the waveform. In addition, a series of higher frequencies, called harmonics, which are integral multiples of the fundamental frequency, are required to completely describe the waveform. The harmonic frequencies are given by
’There are some very unusual waveforms—those with an infinite number o f infinite discontinuities—that cannot be represented by a Fourier series, but these are not o f interest to us.

15.2. FOURIER SERIES FOR ARBITRARY REPEATING WAVEFORMS 93
f n = n f l = n / T for n = 0,1)2.... (15.5)
where n is the harmonic number. For example, /2 is the second harmonic, /5 is the fifth harmonic,0
1Mathematically, any arbitrary repetitive signal g( t ) with fundamental period T can be written
as a Fourier series:
(15.6)n=0
where A„ is the amplitude of the n ,h harmonic component,<t>„ is the phase of the n ,h harmonic component, and f n is the harmonic frequency given by (15.5).
The individual terms in the series (15.6) are called the Fourier components.The “sharper” the features of the signal, the larger the amplitudes A n of the higher harmonics making up the waveform. For example, a sharp spike contains a large number of significant high-frequency Fourier components. On the other hand, a smooth single sinusoidal wave has only one component: the fundamental.
As a simple example of a repetitive waveform, consider the rectangular waveform shown in Fig. 15.2 as a function of time (that is, in the “time domain”). Its “on” time is 250 ms, its “off” time
tg(t)
250 800 1600 t(ms)
Figure 15.2: A repetitive rectangular waveform.
is 550 ms, and it repeats with a period of 800 ms. Its Fourier frequency components are shown in Fig. 15.3 (in the “frequency domain”). Roth the amplitude A n and phase <t>„ are plotted for each frequency component. Note that the fundamental frequency is 1 / T = 1 /(800 ms) = 125 Hz, and that the higher harmonics are integral multiples of 1.25 Hz. The amplitude of the dc term is just the average value of the signal, which can be calculated in another way for a rectangular waveform by finding the peak value multiplied by the ratio of the “on” time to the period, or1 x (250 ms/800 ms) = 0.3125.

94 CHAPTER 15. FOURIER TRANSFORMS: ALTERNATING CURRENTS (AC)
Figure 15.3: The amplitude A„ and phase tp„ of the Fourier components of the rectangular wave shown in Fig. 15.2.
It is important to realize that Figs. 15.2 and 15.3 are just two different ways of representing the same signal. Roth contain all the information inherent in the signal. In fact, the time waveform— Fig. 15.2— can be reconstructed by adding together all its Fourier terms using the amplitudes and phases from Fig. 15.3 for all frequency components. The result of this progressive summation at different stages (for increasing numbers S of terms added in) is shown in Fig. 15.4, starting with the first two terms (5 = 2 ,fo rn = O andn = 1), and ending with all the terms (S = 16) added together.
I t ’s evident from the S = 2 curve in Fig. 15.4 that the fundamental term sets the overall period of the wave, and the higher harmonics represent the sharper details of the wave. As the
16
15.3 FFT: CALCULATING THE DISCRETE FOURIER TRANSFORM
There are several formulas available for finding the frequency components o f a general time waveform. Mathematically they involve integrals over the time variable of the product of the waveform and sine waves. You will use one or more of these mathematical formulas in later courses (math and engineering classes), but for this book we will use the computer to find the frequency components, in particular by using Matlab as our programming and analysis tool.
The algorithm that most software programs employ to find frequency components is the FFT algorithm, which stands for “Fast Fourier Transform.” To use this method, the time waveform first needs to be sampled at discrete, uniformly spaced time intervals. According to the Nyquist criterion, the sampling interval A t should be small enough that there are at least tw o samples per period 7/, of the highest frequency component contained in the original waveform (we’ll see why shortly), or
2 1
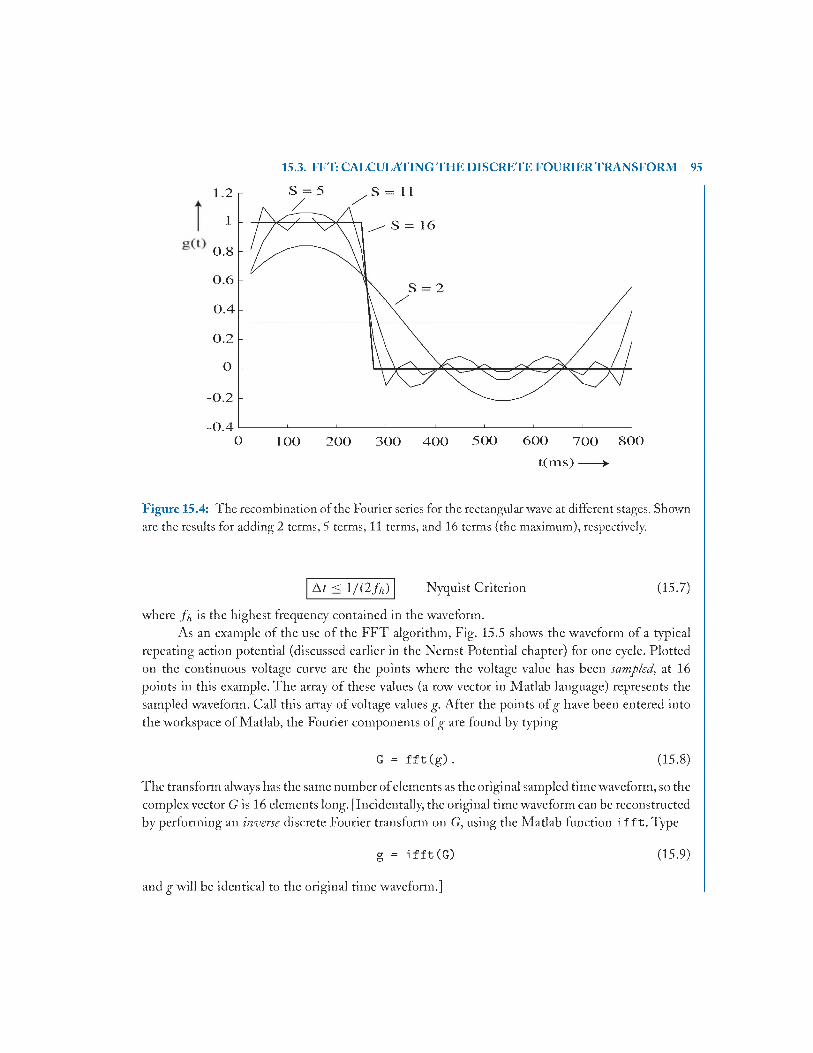
15.3. F F T CALCULATING TH E DISCRETE FOURIERTRANSFORM 95
0 100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 7 0 0 8 0 0
t ( m s ) ----- ►
Figure 15.4: The recombination of the Fourier series for the rectangular wave at different stages. Shown are the results for adding 2 terms, 5 terms, 11 terms, and 16 terms (the maximum), respectively.
A t < 1 /(2 //,) Nyquist Criterion (15.7)
where / , is the highest frequency contained in the waveform.As an example of the use of the F F T algorithm, Fig. 15.5 shows the waveform of a typical
repeating action potential (discussed earlier in the Nernst Potential chapter) for one cycle. Plotted on the continuous voltage curve are the points where the voltage value has been sampled., at 16 points in this example. The array of these values (a row vector in Matlab language) represents the sampled waveform. Call this array of voltage values g. After the points of g have been entered into the workspace of Matlab, the Fourier components of g are found by typing
G = f f t ( g ) . (15.8)
The transform always has the same number of elements as the original sampled time waveform, so the complex vector G is 16 elements long. [Incidentally, the original time waveform can be reconstructed by performing an inverse discrete Fourier transform on G, using the Matlab function i f f t . Type
g = i f f t (G) (15.9)
and g will be identical to the original time waveform.]
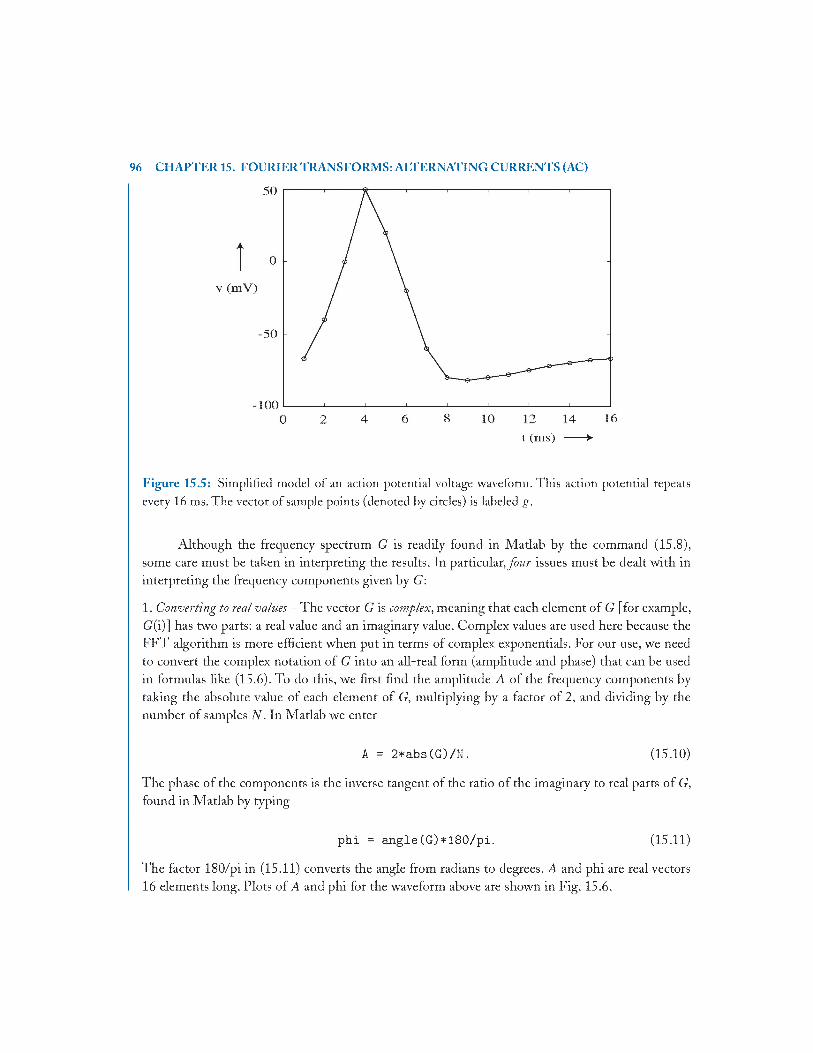
96 CHAPTER 15. FOURIER TRANSFORMS: ALTERNATING CURRENTS (AC)
t (ms) ----- ►
Figure 15.5: Simplified model of an action potential voltage waveform. This action potential repeats every 16 ms. The vector of sample points (denoted by circles) is labeled g.
Although the frequency spectrum G is readily found in Matlab by the command (15.8), some care must be taken in interpreting the results. In particular, fo u r issues must be dealt with in interpreting the frequency components given by G:
1. Converting to real values — The vector G is complex, meaning that each element of G [for example, G(i)] has two parts: a real value and an imaginary value. Complex values are used here because the F F T algorithm is more efficient when put in terms of complex exponentials. For our use, we need to convert the complex notation of G into an all-real form (amplitude and phase) that can be used in formulas like (15.6). To do this, we first find the amplitude A of the frequency components by taking the absolute value of each element of G, multiplying by a factor of 2, and dividing by the number of samples N . In Matlab we enter
A = 2*abs(G )/N . (15.10)
The phase of the components is the inverse tangent of the ratio of the imaginary to real parts of G, found in Matlab by typing
p h i = an g le (G )* 1 8 0 /p i. (15.11)
The factor 180/pi in (15.11) converts the angle from radians to degrees. A and phi are real vectors 16 elements long. Plots of A and phi for the waveform above are shown in Fig. 15.6.

15.3. FFT: CALCULATING TH E DISCRETE FOURIERTRANSFORM 97
Iphi(deg!
f ( Hz ) f (Hz) ■
Figure 15.6: The amplitude and phase of the frequency components of the action potential shown in Fig. 15.5, before the redundancy is removed.
2. Rem oving redundancy in A and p h i — Because the time waveform we started with in Fig. 15.5 is real (as are nearly all of the waveforms of interest in practical signals), the Fourier transform is H erm itian. T hat means that the right half of both the A and phi vectors in Fig. 15.6 are mirror images of the left half, and all necessary information is contained in the first half (9 elements, including the first, or dc, term) of each3. Thus, without loss of information we can cut the 16-element A and phi vectors in Fig. 15.6 nearly in half, preserving only the first 9 elements. The results are shown in Fig. 15.7.
3. Special treatm ent o f the dc term — In Step 2 above we removed the right half of the spectrum representing the redundant negative frequencies. Since cos(«) = COS(—a ) , these negative frequencies were compensated for by using the factor of 2 in the multiplication of (15.10). Rut the amplitude of the dc component is given by the first element of A , which is A(1). This dc term, having zero frequency, should not be multiplied by 2 since it is not included as part of the negative frequencies. Instead we should multiply by a factor of 1. Thus, A(1) is calculated in Matlab by typing
A( l ) = a b s ( G ( l ) ) / N . (15.12)
Also note that the dc term (i.e., for / = 0) is located in the first element (j = 1) of A , not the “zeroeth” element, since Matlab starts with the index i = 1; it does not allow an index i = 0.
4. D eterm in ing thefrequency scale - The last step is determining the correct scale for the frequencies of the Fourier components. The vector g describing the original time waveform does not contain any information about the actual time scale. That is, it is merely an indexed array of sampled values: t?(1)> g(2) , g(3) , etc. It is up to the user to know that the time interval A t between the samples has a
3The right half o f the complex vector G actually represents negative frequencies, needed in the general case of a complex time signal. But since we are dealing here only with real signals, the right half o f G (and A and phi) is redundant with the left half.
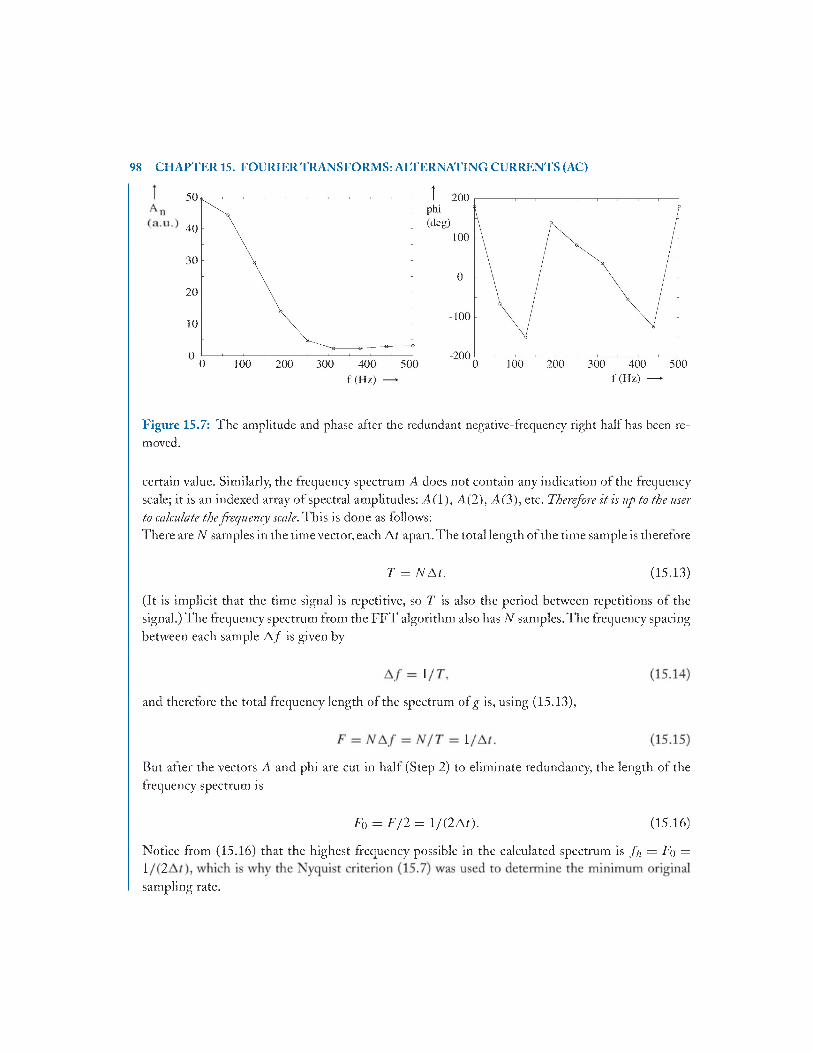
98 CHAPTER 15. FOURIER TRANSFORMS: ALTERNATING CURRENTS (AC)
Figure 15.7: The amplitude and phase after the redundant negative-frequency right half has been removed.
certain value. Similarly, the frequency spectrum A does not contain any indication of the frequency scale; it is an indexed array of spectral amplitudes: A( 1) , A( 2 ) , A( 3 ) , etc. Therefore i t is up to the user to calculate the frequency scale. This is done as follows:There are N samples in the time vector, each A t apart. The total length of the time sample is therefore
T = N A t . (15.13)
(It is implicit that the time signal is repetitive, so T is also the period between repetitions of the signal.) The frequency spectrum from the F F T algorithm also has N samples. The frequency spacing between each sample A f is given by
1
and therefore the total frequency length of the spectrum of g is, using (15.13),
1
But after the vectors A and phi are cut in half (Step 2) to eliminate redundancy, the length of the frequency spectrum is
Fo = F / 2 = 1/(2A f). (15.16)
Notice from (15.16) that the highest frequency possible in the calculated spectrum is f h = Fo =1 2sampling rate.
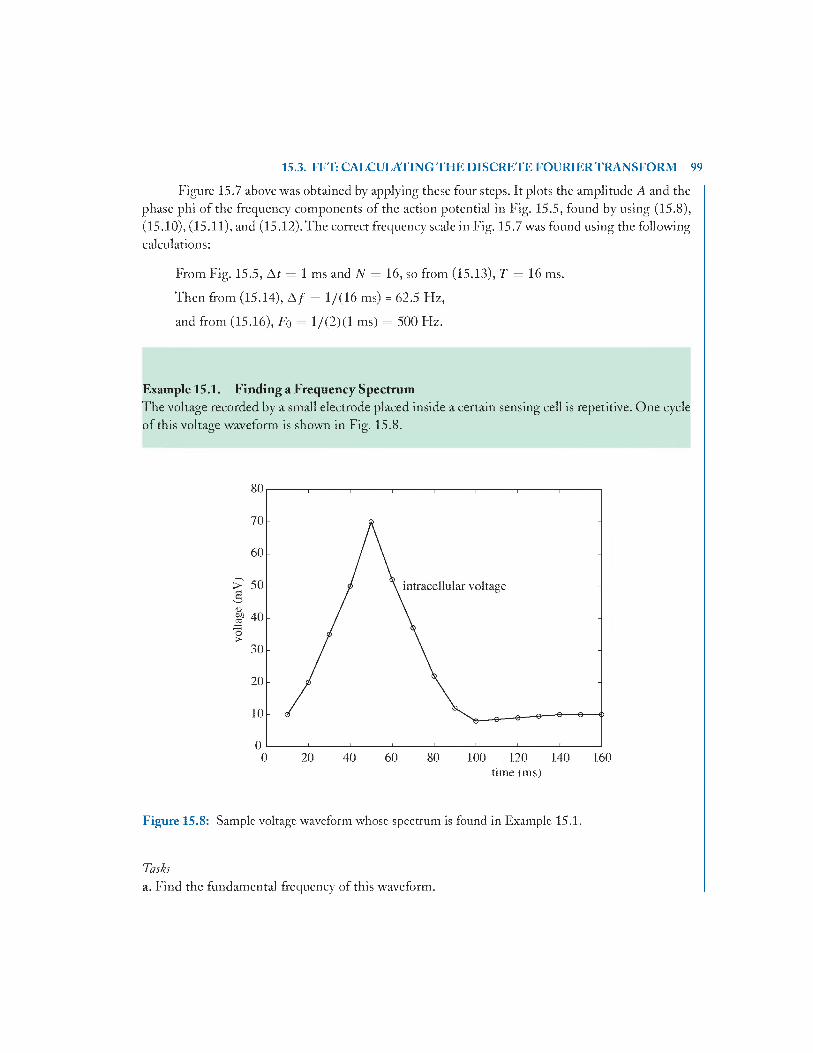
15.3. FFT: CALCULATING TH E DISCRETE FOURIERTRANSFORM 99
Figure 15.7 above was obtained by applying these four steps. It plots the amplitude A and the phase phi of the frequency components of the action potential in Fig. 15.5, found by using (15.8), (15.10), (15.11), and (15.12). The correct frequency scale in Fig. 15.7 was found using the following calculations:
From Fig. 15.5, A t = 1 ms and N = 16, so from (15.13), T = 16 ms.
Then from (15.14), A f = 1/(16 ms) = 62.5 Hz,
and from (15.16), Fo = 1/(2)(1 ms) = 500 Hz.
Example 15.1. F inding a Frequency SpectrumThe voltage recorded by a small electrode placed inside a certain sensing cell is repetitive. One cycle of this voltage waveform is shown in Fig. 15.8.
time (ms)
Figure 15.8: Sample voltage waveform whose spectrum is found in Example 15.1.
Tasksa. Find the fundamental frequency of this waveform.

b.Use the F FT function in Matlab to find and plot the amplitude and phase of the Fourier frequency components of the waveform.
Solutiona. From (15.4), f i = 1 / T = 1/160 ms = 6.25 Hz.b. First we enter in the command window of Matlab a 16-element vector^ of equally spaced samples of the voltage (obtaining the value at each sample point from the graph above), and the number of samples N:
g = [10 20 35 50 70 52 37 22 12 8 8 .5 9 9 .5 10 10 10];N = 16;
Then we find the complex Fourier spectrum of g using (15.8):
G = f f t ( g ) ;
To calculate the real amplitude A and phase phi of the frequency components, we follow Steps 1-3 above using (15.10), (15.11), and (15.12). Therefore, concerning ourselves with only the first half (9 elements) of A and phi, we enter
A = 2 * a b s ( G( l : 9 ) ) / N ;p h i = an g le (G ( 1 : 9 ) ) * 1 8 0 /p i ;A(1) = a b s ( G ( l ) ) / N ;
To plot A and phi, we need to find the correct frequency scale (Step 4). Since T = 160 ms and A t = (160 ms)/16 = 10 ms from the original waveform, (15.14) gives A f = 1/160 ms = 6.25 Hz,
0 1 2 10 50the frequency scale by entering
f = [ 0 : 6 . 2 5 : 5 0 ] ;
and plot A and phi by entering
p l o t ( f , A , f , A , ’o ’ ) ; x l a b e l ( ’freq u en c y ( Hz ) ’ ) ; y l a b e l ( ’am p litu d e A’ ) ; f i g u r ep l o t ( f , p h i , f , p h i , ’o ’ ) ; x l a b e l ( ’freq u en c y ( Hz ) ’ ) ; y l a b e l ( ’p hase ( d e g ) ’ ) ;
The results are shown in Fig. 15.9.
100 CHAPTER IS. FOURIER TRANSFORMS: ALTERNATING CURRENTS (AC)
15.4. PROBLEMS 101
Figure 15.9: Spectrum of sample waveform in Example 15.1.
15.4 PROBLEMS15.1. m. An electrocardiogram (EKG) recording for a healthy heart has the approximate waveform
shown in Fig. 15.10.
It repeats every 800 ms for a resting heart.
a. Calculate the fundamental frequency of this waveform.
fans: f \ = 125 Hz]
b. Using Matlab, find the amplitude A and the phase O of the frequency components of this wave. Sample the waveform at 16 uniformly spaced points. Then plot the amplitude A and phase O as a function of frequency. You only have to turn in these plots as your homework answer. Re sure your frequency scale is correct!
fans: see plots below]

102 CHAPTER IS. FOURIER TRANSFORMS: ALTERNATING CURRENTS (AC)
0 100 2 00 3 0 0 4 0 0 5 0 0 6 0 0 7 0 0 800
t (ms) --------►
Figure 15.10: EKG waveform analyzed in Problem 15.1.

15.4. PROBLEMS 103
frequency (Hz)
frequency (Hz)
Figure 15.11: Spectrum results o f Problem 15.1.


A P P E N D I X C
N ote on O rganization o f M ajor ProjectThis project can be arranged into two halves:
Computer modeling only (1/2 semester) - Section C .l (Background) and Section C.2 (MatlabModel).
Electrical circuit modeling only (1/2 semester) - Section C .l (Background) and Section C.3(Electrical Circuit Analog).
For a full-semester project, all sections are covered.
C .l BACKGROUND (AND PRELIMINARIES)C .1.1 M O D E L IN G T H E C A R D IO V A SC U L A R SY STEMModels are used extensively in engineering analyses. This is sometimes because experimenting with the real thing is too expensive or cumbersome (e.g., determining the failure modes of a large bridge during an earthquake, or finding the most likely fracture planes of a hip-joint replacement fixture), and sometimes because experimenting would be unethical (testing chemical warfare agents in public places, or testing the effects of zero blood flow to the kidneys in humans). In such cases, computer models, animal models or electrical models can be used to simulate the procedure and predict its various effects.
NASA is interested in finding how the human cardiovascular (CV) system behaves under the unusual environmental conditions that astronauts will encounter. These include the high acceleration forces (up to 4 G) that astronauts experience during lift-off, and the zero gravity (0 G) environment they encounter in orbit and outside the earth’s gravity field. Gravity plays an important role in determining the distribution of blood volume in various parts of the body, so changing gravity conditions (especially when protracted) can alter the functioning of the entire CV system.
To study the effects of various gravity conditions on the heart, vessels, blood flow, and blood pooling, NASA relies upon computer models. For example, a study at the University of Utah led by Dr. Keith Sharp (then a research associate professor in the Department of Bioengineering) and performed by graduate student Kristy Peterson developed an extensive computer model of the human CV system1. This research predicted the changes in blood distribution for several posture positions that the astronauts might take during lift-off and under zero-gravity exposure. O ur computer model
1K. Peterson, A Numerical Simulation o f the Cardiovascular System to Investigate Changes in Posture and Gravitational Acceleration, M aster of Science Thesis, University of Utah, Dec. 1999.

for this major project will be similar to their model (though much simpler), but without any gravity effects. However, it will still give valuable insight into the behavior of the human circulation system for both healthy and diseased physiology
C .1.2 O V E R V IE W O F M A JO R P R O JE C TA major goal of this project is to use engineering modeling and simulation tools for examining the dynamic behavior of the human CV system, which is shown in a simplified anatomical drawing in Fig. C .l. Although we will focus on the CV system, the techniques you will learn can be applied to a vast variety of other bioengineering problems, such as orthopedic modeling, imaging, biosensing, and biomaterials behavior.
We will use a lumped-element model of the CV systemic system, explained in much more detail in Section C .l.5. It has been simplified in many regards, such as ignoring the pulmonary circulation and using only a single-chambered heart in order to be more easily analyzed, but it still retains enough detail to display and predict many of the essential features of the CV system. It is essential in our model to keep the two valves that form the entrance port (the mitral valve) and exit port (the aortic valve) of the left ventricle. The arrangement of these two valves around the left ventricle is depicted near the top in Fig. C .l.
The body of this project is comprised of two major sections:
M atlab (Section C.2) - In this part, you write equations relating the variables of the CV model to each other, then numerically solve these equations using the Matlab computer language by stepping through time in small increments during a heart cycle. As input parameters to the Matlab program, you will need to calculate typical values for a healthy CV system. You will plot pressures and other essential quantities during successive heart cycles for a healthy circulation. Then you will model three disease states to see their effects on CV performance.
Electrical Analog (Section C.3) - In this part, you will see that electrical circuits follow the same equations as fluid-mechanical circuits (such as the CV system), once pairings that relate electrical quantities to corresponding fluid quantities are made. The electrical circuit is identical to the Matlab model, except that a special op amp circuit is needed to model the left ventricle and an extra branch is needed to add or bleed “blood volume” to the circuit. You will determine the values of the circuit components, then assemble the circuit, run it, and measure equivalent CV variables (such as the voltage waveform representing the aortic blood pressure waveform). You will initially model a healthy heart, then change the circuit to represent two more disease states to see their effects on CV performance.
C .1.3 N O T E B O O K R E Q U IR E M E N T SOne hallmark of a good engineer is that she or he always keeps complete and accurate records of derivations and experimental observations. The record should be so complete that another engineer can follow the derivation or repeat the experiment several years later based upon this written record
106 APPENDIX C. MAJOR PROJECT
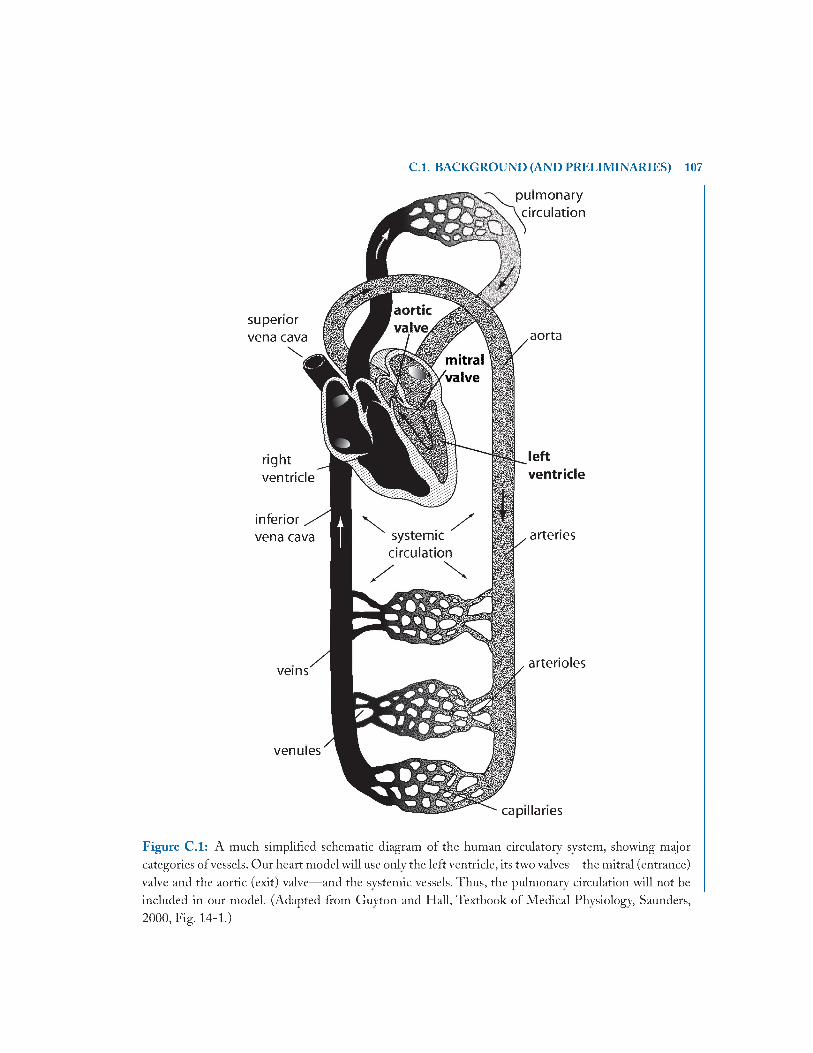
C.l. BACKGROUND (AND PRELIMINARIES) 107
superior vena cava
\
rightventricle
inferior vena cava
veins
venules
pulm onarycirculation
aorta
leftventricle
arteries
arterioles
capillaries
Figure C .l: A much simplified schematic diagram of the human circulatory system, showing major categories of vessels. Our heart model will use only the left ventricle, its two valves—the mitral (entrance) valve and the aortic (exit) valve—and the systemic vessels. Thus, the pulmonary circulation will not be included in our model. (Adapted from Guyton and Hall, Textbook of Medical Physiology, Saunders, 2000, Fig. 14-1.)

alone. This means that it needs to contain all the detail that was originally needed to solve the problem, including even false starts and mistakes!
Another reason for keeping a complete record is for patent purposes. Many organizations have a “patent notebook” policy requiring engineers to keep a legal record of all work as proof of inventorship. This involves keeping a contemporary record (on-the-spot entries) in a bound notebook, in ink, with all pages numbered, dated, and signed. For this project, you will gain experience keeping this type of notebook. All written work related to the project must be recorded in this notebook. The rules for keeping the notebook are:
1. The notebook must be about 8 1/2 x 11” in size with fastened-inpages', no folders or loose-leaf binders. This means that no pages can be added or taken away from the original. A spiral- bound notebook will suffice for us. (Actually, true patent notebooks should have sewn-in pages, but these are somewhat expensive for us.)
2. A ll entries must be in ink. I f you make a mistake, cross it out with an X and start again. Do not try to erase or obliterate.
3. All pages must be numbered in order from the front of the notebook. Do this in ink by hand. You can use one or both sides of the page (use one side if your writing shows through the paper).
4. Make all your entries immediately in the notebook. Do not do them first on a loose piece of paper to transfer later—you won’t have time. Your notebook does not have to be overly neat, just complete and readable by someone else like a fellow engineer or the teaching assistant. Don’t spend a lot of time making the tables and drawings fancy, just complete and clear.
5. For purposes here, you can tape in computer printouts and graph printouts on blank pages of the notebook to keep a record of all your work.
6. After you have filled a page, date it in ink at the top and sign each page.
C .1.4 G R A D IN G A N D C H E C K O F F D A T E SThere are four checkoff periods: two for the Matlab section and two for the electrical circuit section. You will be required to show your work and resulting data to the teaching assistant four times during the semester in your scheduled lab session within the weeks shown in the schedule given in the class syllabus. Your grade on the major project will be determined by how well you have followed and completed the tasks given in this handout, as evidenced by your notebook entries and records, your filled-in tables, your printouts, your assembled circuit, measurements on your circuit, and answers to questions by the teaching assistant.
No checkoffs of any Major Project section will be allowed after the scheduled week for that section. I f you have any questions about how you are graded, please first see the teaching assistant, then the instructor if you still have unresolved questions. Table C .l shows the four checkoff sections, what items will be graded, and how many points are assigned to each item.
108 APPENDIX C. MAJOR PROJECT

C.l. BACKGROUND (AND PRELIMINARIES) 109
Table C.l:
«...
......
..Mat
lab
Mod
el..
Checkoff To Be Completed Points
#1
□ Good notebook practice (guidelines followed).□ Filled-in Table 2.□ Filled-in Table 4.□ Calculations of all fluid R & C values (in notebook).□ Equations relating CV variables (in notebook).□ Vectorized and Euler's form of these equations (in notebook).□ Matlab m-file with FOR loops started (display or printout) -
[Note: At this stage, the program does not have to be finished,ju st started. ]
* 15
► 15
#2
□ Matlab m-file completed (printout in notebook).□ Matlab simulation for healthy CV parameters:
demo of program to teaching assistant,printout of graph of pressure waveforms (in notebook),printout of parameter file for healthy CV (in notebook).
□ Matlab simulations of three diseases; for each disease:demo of program to teaching assistant,printout of graph of pressure waveforms (in notebook),printout of parameter file for each disease (in notebook).
□ Relevant boxes in Table 5 filled in.
*• 15
► 20
-4.E
lect
rica
l Ci
rcui
t M
odel
......
.....►
#3
□ Filled-in Table 6.□ Calculations of all electrical R & C values (in notebook).□ Circuit diagram for left ventricular module (in notebook).□ Assembly of left ventricle module, including two voltage followers:
demo of working circuit to teaching assistant, dual plot—by hand—of voltage waveforms for both excitation and vh, with vertical axis in units of both V and equiv mmHg (in notebook). [Note: points will be deducted i f graph is not done carefully and completely. ]
} ‘ 5
* 20
#4
□ Assembly of entire systemic CV circuit.□ Measurements of electrical waveforms for healthy CV system:
demo of circuit to teaching assistant,dual plot of voltage waveforms for vh and v (in notebook)—see comments above regarding grading of dual waveform plot, record of cardiac output (in notebook).
□ Measurements of affected electrical variables for two diseases:demo of circuit to teaching assistant, graphs of voltage waveforms for vh and v (in notebook), record of cardiac output, percentage change, and resulting BP for each disease (in notebook).
□ Table 5 completed.
* 20
► 15

110 APPENDIX C. MAJOR PROJECT
C .1.5 O U R M O D E LA schematic of the CV model we will use for both the Matlab simulation and the electrical circuit simulation is given in Fig. C.2. It is obvious that several simplifying approximations have been made, as can be seen by comparing our model to even a simple anatomical drawing of the actual human CV system. Rut our model does have enough components to give the correct overall blood flow and pressure behavior, and to predict the effects of many of the common CV disease states, such as the high blood pressure that results from atherosclerosis.
Your first task will be to calculate from measured data of the healthy human CV system the values of the components in Fig. C.2, namely the resistance elements (R) that represent the pressure drop across major vessels and the capacitance elements (C) that represent the compliance of these windkessel vessels. Note in Fig. C.2 that we have lumped major categories of vessels together (for example, large and small arteries and arterioles) in the model for simplicity. This lumped-element approximation loses some information about spatial details, but helps keep the number of equations to a manageable level. The labels in Fig. C.2 show which elements correspond to the various segments of the CV system. W ith some study of the model, you will see that once a certain blood volume is inside the system, it just keeps circulating around the loop with no leakage out (unless some is bled away or added on purpose).
Also note in Fig. C.2 that in addition to the model parameters R and C, there are several variables which represent the state of the system at any given time. These include the pressure P of various vessels, the volume V in these vessels, and the volumetric flow rate Q through the vessels. These are the variables you will solve for using Matlab during time increments of a heart cycle. It is very important to keep track of the traditional clinical units used when measuring these variables and to consistently use these same units throughout the problem. To help keep track, fill in Table C.2 below with the clinical units you will use for each variable. (Note: The entire class should decide on common units to make communication and cross-checking easier.)
Table C.2: Traditional Clinical Units to be Used in SolutionVariable C linical Units
Pressure P Volume V Volumetric Flow Q
To keep the number of variables reasonable, we have modeled the heart by only one chamber— the left ventricle—which is the major pump responsible for ejecting blood into the aorta. Its pumping action is modeled by a time-varying compliance; the ventricle’s compliance changes from a higher value Cfrj during diastole to a lower value C/,.v during systole in an exponential manner, as shown in Fig. C.3. This exponential behavior mimics reasonably well the compliance changes during an actual cardiac cycle, and is similar to the compliance waveform that will be generated by the electrical circuit in the second half of the project.

for e
lectric
al
C.l. BACKGROUND (AND PRELIMINARIES) 111
Figure C.2: Simple model of cardiovascular system.
aorta
, arte
ries,
arter
ioles
, ca
pillar
ies
, and
ve
na
cava

112 APPENDIX C. MAJOR PROJECT
tLeftVentricleCompliance
systole diastole
Figure C.3: Exponential model for the left ventricular compliance.
During the systolic segment, the ventricular compliance follows the equation2
Ch = (Ch d - C hs) e - ' ^ + C h s , (1)
where xs is the time constant representing the onset of systole (r5 = 30 ms fits our model best). During the diastolic segment, the compliance follows the equation
Ch = (Chs - Chd) e + Chll, (2)
where td is the time constant representing the onset of diastole (r^ = 60 ms fits best here).To help you determine the compliance (C ) and resistance (R ) values of each of the vessel
groups shown in Fig. C.2, you will need some important properties of a typical healthy human CV system given in Table C.3, including the blood volume contained in each compliant vessel category as well as the (estimated) residual volume in each compartment. Note that the total blood volume in the systemic loop (not including the pulmonary circulation) is 5.0 L — 0.8 L = 4.2 L. In addition, Fig. C.4 shows typical pressures around the systemic circulation, and Fig. C.5 shows the pressure waveforms of the left ventricle and aorta during one cardiac cycle. You will use these data sources to calculate the values of the model parameters in Section C.7.
C .1.6 A P P R O X IM A T IO N SAny engineering model (mathematical, electrical or otherwise) must make some simplifying approximations to be useful; otherwise, it is so complex that it is impossible to solve. But these approximations can’t go too far or the essence of the problem will be lost. For example, in our CV model we have reduced the four-chambered heart to a single chamber to keep the number of equations smaller, but we have to keep at least one chamber lest there would be no pumping action in the heart.
'■In these equations, the time variable t is referenced to zero at the beginning ot each segment. In the actual com puter program, each segment must be shifted to match the systole and diastole tim ing shown in Fig. C.5.
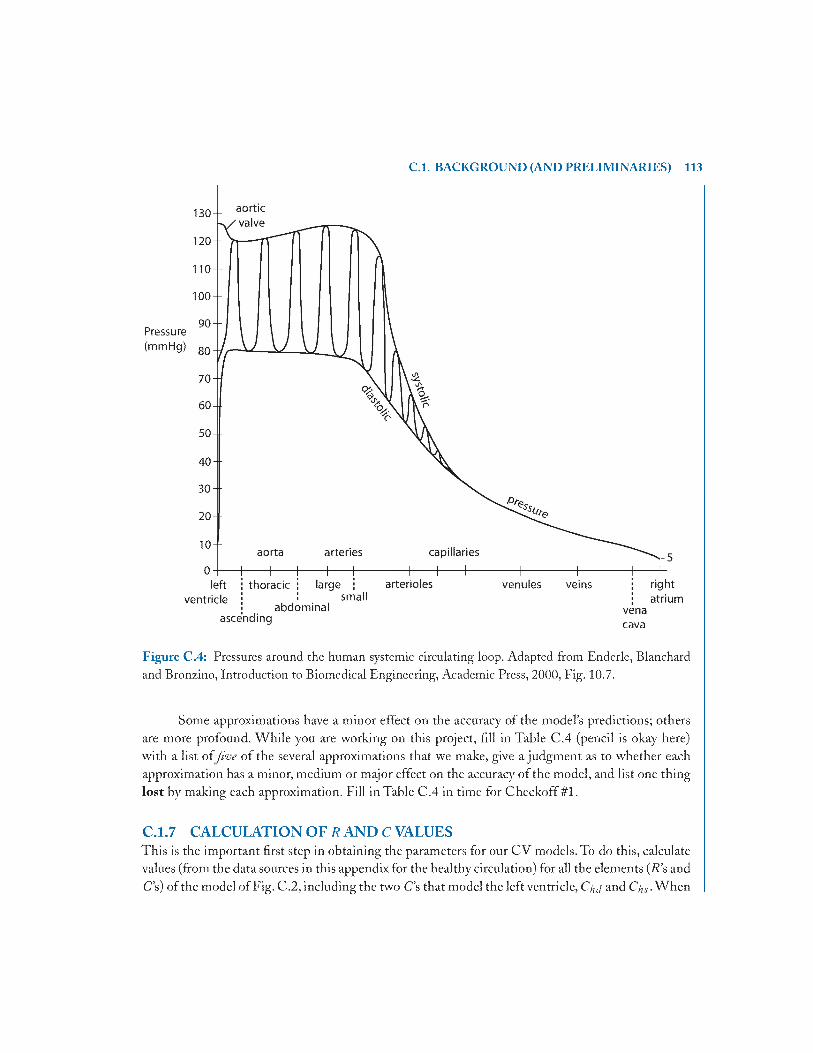
C.l. BACKGROUND (AND PRELIMINARIES) 113
Figure C.4: Pressures around the human systemic circulating loop. Adapted from Enderle, Blanchard and Bronzino, Introduction to Biomedical Engineering, Academic Press, 2000, Fig. 10.7.
Some approximations have a minor effect on the accuracy of the model’s predictions; others are more profound. W hile you are working on this project, fill in Table C.4 (pencil is okay here) with a list of five of the several approximations that we make, give a judgment as to whether each approximation has a minor, medium or major effect on the accuracy of the model, and list one thing lost by making each approximation. Fill in Table C.4 in time for Checkoff #1.
C . l .7 C A L C U L A T IO N O F R A N D C VALUESThis is the important first step in obtaining the parameters for our CV models. To do this, calculate values (from the data sources in this appendix for the healthy circulation) for all the elements (R ’s and C’s) of the model of Fig. C.2, including the two C’s that model the left ventricle, C/,,/ and Cils- W hen

114 APPENDIX C. MAJOR PROJECT
Pressure(mmHg)
last of previousventriculardiastole
ventricularsystole
beginning of nextventriculardiastole
0 100 200 300 400
Time (ms)
500 600 700 800
Figure C.5: Wiggers diagram showing the relationship between various blood pressures and ventricular blood volume during a cardiac cycle. Adapted from D. Silverthorn, Human Physiology, an Integrated Approach, Prentice Hall, 1998, Fig. 14-27.
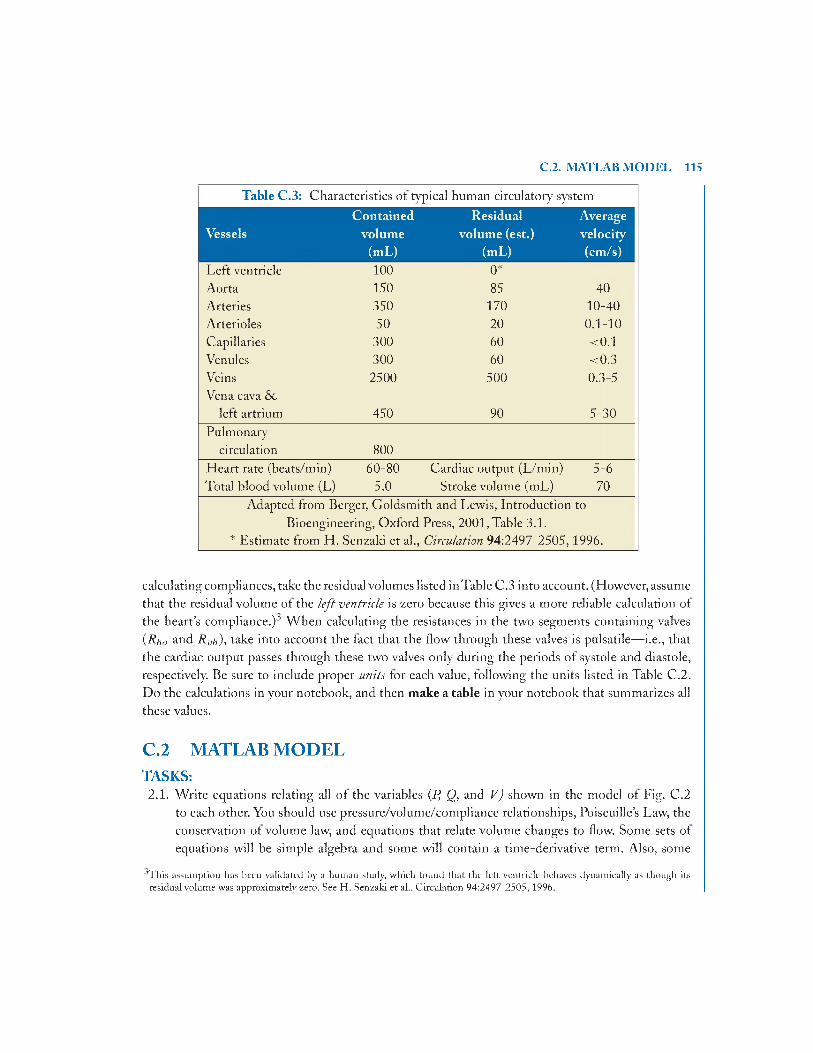
C.2. MATLAB MODEL 115
Table C.3: Characteristics of typical human circulatory systemContained Residual Average
Vessels volume volume (est.) velocity(mL) (mL) (cm/s)
Left ventricle 100 0*Aorta 150 85 40Arteries 350 170 10-40Arterioles 50 20 0.1-10Capillaries 300 60 <0.1Venules 300 60 <0.3Veins 2500 500 0.3-5Vena cava &
left artrium 450 90 5-30Pulmonary
circulation 800Heart rate (beats/min) 60-80 Cardiac output (L/min) 5-6Total blood volume (L) 5.0 Stroke volume (mL) 70
Adapted from Berger, Goldsmith and Lewis, Introduction to Bioengineering, Oxford Press, 2001, Table 3.1.
* Estimate from H. Senzaki et al., Circulation 94:2497-2505,1996.
calculating compliances, take the residual volumes listed in Table C.3 into account. (However, assume that the residual volume of the left ventricle is zero because this gives a more reliable calculation of the heart’s compliance.)J W hen calculating the resistances in the two segments containing valves (Rho and R vh), take into account the fact that the flow through these valves is pulsatile— i.e., that the cardiac output passes through these two valves only during the periods of systole and diastole, respectively. Be sure to include proper units for each value, following the units listed in Table C.2. Do the calculations in your notebook, and then make a table in your notebook that summarizes all these values.
C.2 MATLAB MODELTASKS:
2.1. W rite equations relating all of the variables (P, Q, and V) shown in the model of Fig. C.2 to each other. You should use pressure/volume/compliance relationships, Poiseuille’s Law, the conservation of volume law, and equations that relate volume changes to flow. Some sets of equations will be simple algebra and some will contain a time-derivative term. Also, some
"This assumption has been validated by a human study, which found that the left ventricle behaves dynamically as though its residual volume was approximately zero. See H . Senzaki et al.. Circulation 94:2497-2505,1996.
116 APPENDIX C. MAJOR PROJECT
Table C.4:
i>O
O—
C\3xiH5D3HS3O
S3.2S3S
*soS-HOhQh<d
i>
~s3CdO5DS33
T3Oo
5D.Sooo«
1>T31)£* 'uc3S3 TS C ’"Oa s -I scu
£.3'-3I
T31>
W)1)S3
Oh-2S3O
C\3S3O3Cu
©HIX
m

C.2. MATLAB MODEL 117
equations will be conditional (with an IF statement) describing the flow through the valves. You should end up with about 22 final equations, not counting the equations that generate the waveform for C/,. Do all the mathematics in your notebook.
2.2. Break the period of one heart cycle into many increments, each with time length A?. Then apply Eulers method to approximate by forward differences the time derivatives of those equations developed in Task 2.1 that contain such derivatives.
Then write all of the final equations of Task 2.1 in vector notation. That is, let all the variables (for example: Pu or V,,) be represented by row vectors, whose elements Pu (i) or V;, O') are the values of the variable at each increment of time. For those equations that use Euler’s method, solve for the value at the next increment of time Va O' + 1) in terms of the value at the current time step Vu (;'). This will allow your Matlab program to step through time during a heart cycle. Since you will be coding these equations in Matlab in the next task, collect them all (after you’ve finished putting them in vectorized format) on one or two pages of your notebook.
2.3. W rite a Matlab script m-file that contains two FO R loops. The inner FO R loop (the major one) steps through the time increments during one heart cycle (say 800 steps) solving for the time behavior of all the variables (P ’s, Q ’s, and F ’s) shown in Fig. C.2. This uses the equations developed in Task 2.2. Also inside this loop will be some conditional sections using IF statements to handle the conditional flow through the valves.
The outer FO R loop repeats the heart cycle again and again for as many times as necessary to reach steady state (perhaps 10 or more cycles). Be sure to “glue” the start of each cycle to the
1values at the very end of the previous cycle Va (801).
Also, figure out away to calculate the cardiac output (CO) of each cycle. CO is defined as the blood volume being pumped out of the left ventricle per minute (so its units are L/min), but it can be calculated on a beat-to-beat basis. You will write out the CO after each heart cycle in the next task, along with writing out the blood volume in the venous side and plotting all the pressure waveforms after each cycle.
Before entering into the outer FO R loop for the first time, your program must first read in all the parameters for your model (for example, the R and C values for a healthy circulation, all timing parameters, and the residual volumes) and the initial distribution of the blood volume in the various compliant vessels. (Im portant: The distribution of blood volume throughout the system will surely adjust during the simulation run, so exactly what the distribution is when you start is not very important. But the total blood volume will remain constant, so make sure you have the proper total starting volume, which is 4.2 L for the normal CV systemic system.) It is convenient to use a separate m-file which contains all those parameters for a healthy circulation and to read in this separate file by “inputting” its name (without the .m extension) at the beginning of the run of your main script file. Then other abnormal circulation states (e.g., hypovolemia) can have their own parameter files.

Also, before entering the outer FO R loop for the first time, you should calculate a row vector representing the left ventricular compliance waveform C/,. Use the equations found in Section 1.5, with each segment (systole and diastole) shifted appropriately to correspond to the timing found in Fig. C.5.
2.4. In your main script m-file, put the capability to p lo t out on the same graph on the screen the following waveforms updated after each heart cycle:
* Left ventricular pressure.
* Aortic pressure.
* Arterial pressure.
* Capillar}' pressure.
* Venous pressure.
On the graph axes, put labels and correct units. To identify each pressure, use separate line styles and colors for each of the five lines. Also, use the T E X T command to write inside the graph the values of: a) CO for the heart cycle, and b) total venous blood volume (venules, veins and vena cava) at, say, the start of that heart cycle. Put these values in a text line (somewhere inside the graph) that looks like this:
C O = 5.3 L /m in Venous Vol = 3.3 L.
At the end of the plot section, put a PAUSE command [e.g., pause(0.5)] to allow time for the program to print the graphs on the computer screen before continuing the FO R loop. Print out a copy of the final version of your main m-file and tape it in your notebook.
2.5. Run your script m-file using healthy circulation parameters for several cycles, until steady state is reached. On the graph on the screen, watch the changes in the pressure waveforms as the total blood volume redistributes itself throughout the various compartments of the system. Watch for changes in CO and venous volume. After steady state is reached, stop your program and print out the last graph, representing the behavior of a healthy circulation, and tape it in your notebook along with a copy of the healthy parameter file. Be prepared to demonstrate this program and answer questions about it during Checkoff #2.
2.6. Several CV diseases have been observed and categorized in humans. A few of the more important ones are listed in Table C.5. In this task, you are to simulate three of them— anaphylactic shock, left-heart failure (sometimes called congestive heart failure), and hypovolemia—by using your Matlab program to see what effects these pathologies have on CV performance. To see how each disease state should be modeled, consult relevant books or explore the internet to learn about each disease. Then make new parameter m-files representing each disease state with its modified values. Run your Matlab program for each of these abnormalities, and note
118 APPENDIX C. MAJOR PROJECT

C.3. ELECTRICAL CIRCUIT ANALOG 119
the major effects on certain pressures, CO and venous volume. Print out a representative graph to tape in your notebook along with its corresponding parameter file for each pathology. Then fill in (pencil is okay) the various columns in Table C.5 for these three diseases. (The last two diseases will be simulated with the electrical circuit, next.)
C.3 ELECTRICAL CIRCUIT ANALOGAs you learned earlier, there is a one-to-one correspondence between the differential equations describing major fluid-mechanical components and the equations describing electrical circuits once the electrical variables are identified which correspond to the respective fluid variables. Thus, the model shown in Fig. C.2 can be directly implemented as an electrical circuit, and pressures and flows can be measured as electrical quantities from this circuit. This is the overall goal of Section C.3 of this project.
TASKS:3.1. Fill in Table C.6 with the units you used in Table C.2 for the fluid CV variables, then with
the names and units of the electrical variables that correspond to the fluid variables.
3.2. Translate all of the fluid-element values that you previously calculated in Section C.1.7 for a healthy circulation (all R ’s and C’s) into their respective electrical-element values. To find the translation scaling factor for the C’s, use the fact that that the largest compliance value found in Section C.1.7 should correspond to the largest practical capacitance size available in the lab (100 n F). Then, to find the translation scaling factor for the R ’s, use the fact that tim e is the same in both systems (fluid and electrical); the result is that the scaling factor for the R ’s will depend on the scaling factor found for the C ’s. Be sure to include units in your scaling equations. Then make a table of all the electrical R and C values for Fig. C.2 for a healthy circulation, with proper units. Do this work and record your values in your notebook. (Note: Another scaling equation— relating fluid pressure to electrical voltage—will be left unspecified until later, when it can be adjusted to keep the voltages of the circuit within a reasonable range.)
3.3. The left ventricle poses a special challenge for the electrical-circuit model. The capacitance of capacitor C/7 must change from a high value C m during diastole to a low value C/1S during systole w ithout changing the electrical charge (which is analogous to ventricular blood volume) stored on the capacitor during the rapid changeover— this corresponds to isovolumic contraction of the ventricle.
An active circuit that can achieve this by using two op amps and an analog switch is shown in Fig. C.6. The portion containing the two op amps is called a "capacitance multiplier" circuit since the effective capacitance seen across the input terminals (marked by X-X) can be larger than the physical capacitance value C \. It can be shown that, using the branch current method and an ideal op amp model, the effective capacitance is given by

120 APPENDIX C. MAJOR PROJECT
Table C.5:ac5
60 O 1/1Oh>j
a9.ou
u£P «-• § O6 U
CCSGOOftP
03GO
OftP
CD"O£ o -(U03 ^ GH G CS <3 *rt xJ&H O □605 g $ fe: -= l> U
U cai .a « Q ■§
CDX ) ^
at !£ I— M S' *g r̂t wj*i > gj03> CD w'S* .r t
r t CDcs G X O
o>of t;>>
X
cfS P
X3 0> 0̂3 CjlJ *£3 T5
s_
-rt ^•S ~ -23 £ ES 4J 1)>>>2- o1 - u O1 2 £< •§ - a
Gj>1 3
M
►>o jap
3o Oii
<CD

C.3. ELECTRICAL CIRCUIT ANALOG 121
Table C.6: Fluid/Electrical Pairs (Note: pencil is 0 kay here)
F luidQ uantity
Symbol F lu idUnits
Corresponding Electrical Q u an tity
Symbol ElectricalUnits
Resistance element R R
Complianceelement
C C
Pressure P V
Volume V 9
Volumetric flow Q i
C# - ( 1 + R l ) C l '
In Fig. C.6, the analog switch is in the down position (open) during systole and is up (closed) during diastole; the resulting changes in C eff- are used to model the desired changes in C/7. During diastole, for example, the circuit resistance R i = 10 kQ, but during systole the circuit
1your notebook, write two expressions for C/, (that is, C ef f ) in terms of Ci , S ^ ^ d R 2 : write one for C/w (systole— switch open) and one for C/,d (diastole—switch closed). Then use these expressions and the values for C/w and C/,d you derived in Task 3.2 above to derive the values
1 2
(Note: The 4.7 /iF capacitor is added here only so the variation in C/7 follows a more gradual exponential waveform rather than an abrupt change between systole and diastole. Ignore the presence of the 4.7 /iF capacitor in your calculations above.)
3.4. Redraw the complete left ventricular module diagram of Fig. C.6 in your notebook. Show pin numbers on your diagram by referring to the pin-out diagrams of the op amps (Fig. C.7) and analog switch (Fig. C.8). Show the power connections to the op amp chip and the switch chip. Also show values for the capacitors and resistors to give the desired values for C/„/ and C/7 S as determined in Task 3.3.
3.5. In the lab, assemble the left ventricle circuit you designed in Task 3.4 using the left half of a prototyping board, jum per wires, op amp chip, analog switch chip, and a function generator. Connect apower supply to the circuit and adjust the function generator to represent the cardiac

122 APPENDIX C. MAJOR PROJECT
Figure C.6: Left ventricular module consisting of a capacitance multiplier circuit and an analog switch. The effective capacitance seen across terminals X-X represents C/,, which changes exponentially between a high value during diastole and a low value during systole.
cycle of Fig. C.5 with the proper systolic and diastolic periods. Ifyou use electrolytic capacitors, be sure to observe the proper polarity of the leads by orienting the “+ ” lead toward the positive voltage and the ” lead toward ground.
The voltage across the effective capacitance represents left ventricular pressure, and it will be a function of how much charge (representing blood volume) is stored on the capacitor. The scale factor relating circuit voltage to blood pressure can now be set. A good signal voltage range for the op amp is 0 to 4 V, so a recommended scale factor for translating between voltage and pressure is one such that 100 m m H g pressure is represented by a voltage o f 4.0 V. Thus if the oscilloscope vertical scale is set at 2 V per division, then a blood pressure of 100 m m Hg is represented by a height of 2 divisions (2 “boxes”) on the oscilloscope screen.
Run your left ventricle circuit and observe the changing voltage across the effective capacitor with an oscilloscope. To inject a proper amount of charge onto the capacitor in your circuit, use the charge injector arrangement shown in the dotted box in Fig. C.2. Inject enough charge into the capacitor to get a reasonable systolic blood pressure for a healthy left ventricle using the scale factor above. By hand, plot the voltage waveforms for both the excitation voltage (i.e., the voltage from the function generator driving the analog switch) and for the ventricular voltage Vh. Overlay the two waveforms on the same graph with the same horizontal axis (time) and the same vertical axis. Label the vertical axis with two scales: one in voltage (units of V) and one in equivalent blood pressure (units of mmHg) using the scale factor above. The graph

C.3. ELECTRICAL CIRCUIT ANALOG 123
Connection Diagram LF353 Dual Operational Amplifier
Figure C.7: Connection diagram for the LF353 op amps.
must be neat and complete, or points will be deducted from your score! Do the plot in your notebook, and be prepared to demonstrate this circuit during Checkout #3.
Im portan t note: An oscilloscope has a large but not infinite input impedance, meaning that a small amount of current will be drained off the capacitor when the oscilloscope is connected, bleeding away most of the charge after a minute or two. This has undesirable consequences, both from an electrical point of view and a physiological point of view (obviously, excessive bleeding of a patient in order to take many repeated measurements of blood gases can lead to hypovolemia and eventual death). Therefore, we need to use an impedance “buffer” between the oscilloscope and the circuit to be measured. The arrangement in Fig. C.9 shows two voltage followers, and takes advantage of the extremely large (1012 £2) input impedance of an op amp. Connect up two voltage followers on the top side of your prototyping board (using one additional op amp chip) between your circuit and the oscilloscope. Refer to the pin-out diagram of Fig. C.7 for correct connections. You can reuse the voltage follower setup that you have already employed in Problem 13.2. Connect one channel of the two-channel oscilloscope to the excitation (function generator) voltage and the other channel to the left ventricle (capacitor) voltage.
3.6. After the left ventricle module is working, you can assemble the entire circuit of Fig. C.2 on the prototyping board. Use the R and C values for a healthy heart found in Tasks 3.2 and 3.3. Inject enough charge into the circuit to get the correct systolic peak pressure in the aorta (this

124 APPENDIX C. MAJOR PROJECT
Connection Diagram DG419 Analog Switch
source: Vishay Siliconix Databook
Switching Conditions
Vin s, S2
OV on off
5 V off on
Figure C.8: Connection diagram for the DG419 analog switch.
Figure C.9: Two voltage followers act as buffers between the electrical CV model circuit and the oscilloscope to avoid draining charge off the capacitors.
is close to the blood pressure measured in your upper arm) as given in Fig. C.5. There are several seconds of time lag between the time that charge is injected and the time the voltage settles to a final value, so have patience when injecting the charge. (The time lag represents the redistribution of blood within the various compartments of the body.) In order to measure the voltages iv, and v0 simultaneously, use the two voltage followers from the previous task connected to the channels of a two-channel oscilloscope. By hand, plot the voltage waveforms for both the ventricular voltage iv, and the aortic voltage vQ. Overlay the two waveforms on the

C.3. ELECTRICAL CIRCUIT ANALOG 125
same graph with the same horizontal axis (time) and the same vertical axis. Label the vertical axis with two scales: one in voltage (units of V) and one in equivalent blood pressure (units of mmHg) using the scale factor above. The graph must be neat and complete, or points will be deducted from your score! Do the plot in your notebook, and compare this graph to Fig. C.5. Be prepared to demonstrate your circuit performance and answer questions from the teaching assistant in Checkoff #4.
Since blood volume and electrical charge are analogs, cardiac output (CO) is represented by the average current flow around the circuit. To measure CO, you could insert an ammeter in any link of the circuit, but in practice the current is so low that most ammeters won’t read it accurately So use Ohm ’s Law to relate the voltage across one of the resistors to the current through it, and connect a hand-held voltmeter across this resistor to measure dc voltage. (Rcv is a convenient place to measure CO since the flow is rather steady here.) Record this voltage proportional to CO in your notebook after reaching steady state.
3.7. Now model the two remaining diseases in Table C.5, atherosclerosis and aortic valve regurgitation, using your electrical circuit. Determine which electrical components need to be changed (using only resistors) to represent each disease, then modify your circuit accordingly For each disease, run the circuit, plot in your notebook the waveforms for i>/, and vot record the new blood pressures in the ventricle and aorta, and record the new voltage proportional to CO. Calculate and record the percentage change in the CO compared to the healthy system. W hen adding, removing or changing resistors, keep the circuit running (“hot swap” the resistors) and avoid draining any charge off the capacitors while changing the resistors. Fill in the remaining boxes in Table C.5 based upon your results. You will be asked to demonstrate one of these pathologies during Checkoff #4.
G O O D LUCK (and remem ber to take care o f your cardiovascular system during your lifetime)!


[1] Berger, S.A., Goldsmith, W . and Lewis, E.R. 1996. Introduction to Bioengineering, Oxford: Oxford University Press.
[2] Berne, R.M. and Levy, M .N. 1993. Physiology, St. Louis: Mosby-Year Book, Inc.
[3] Darcy, H . 1856. Les Fountaines de la Ville de Dijon, Dalmont, Paris.
[4] Durney, C .H . 1973. “Principles of Design and Analysis of Learning Systems,” Engineering Education, March 1973,406-409.
[5] Durney, C .H . and Christensen, D.A. 2000. Basic Introduction to Bioelectromagnetics, Boca Raton: CRC Press.
[6] Eide, A.R., et al. 1997. Engineering Fundamentals and Problem Solving, 3rd Edition, Boston: W CB M cGraw-Hill.
[7] Enderle, J., Blanchard, S. and Bronzino, J. 2000. Introduction to Biomedical Engineering, San Diego: Academic Press.
[8] Folkow, B. and Neil, E. 1971. Circulation, New York: Oxford University Press.
[9] Fung, Y.C. 1984. Biomechanics: Circulation, New York: Springer-Verlag.
[10] Fung, Y.C. 1990. Biomechanics: Motion, Flozu, Stress, and Growth, New York: Springer-Verlag.
[11] Guyton, A.C. 1974. Function o f the H uman Body, Philadelphia: Saunders.
[12] Guyton, A.C. and Hall, J.E. 2000. Textbook o f Medical Physiology, Philadelphia: Saunders.
[13] Keener, K. and Sneyd, J. 1998. M athematical Physiology, New York: Springer.
[14] Mars Climate Orbiter, on NASA web site: www.NASA.gov.
[15] Matlab Help Desk, The M ath Works Inc., Natick, MA.
[16] Nillson, J.W. and Riedel, S.A. 1996. Electric Circuits, 5th Edition, Reading, MA: Addison- Wesley.
[17] Palm, W.J. III. 1999. Matlab fo r Engineering Applications, Boston: W CB McGraw-Hill.

128 BIBLIOGRAPHY
[18] Peterson, K. 1999. A Numerical Simulation o f the Cardiovascular System to Investigate Changes in Posture and Gravitational Acceleration, MS Thesis, University of Utah.
[19] Senzaki, IL , Chen, C -II. and Kass, D.A. 1996. “Single-Beat Estimation of End-Systolic Pressure-Volume Relation in Humans,” Circulation, 94: 2497-2505.
[20] Silverthorn, D. 1998. Human Physiology, an Integrated Approach, Upper Saddle River: Prentice- Hall.
[21] Starling, E.H . 1918. The Linacre Lecture on the Law o f the Heart, London: Longmans, Green.
[22] W ithers, P.C. 1992. Comparative A n im al Physiology, Fort Worth: Saunders College Publishing.

















