IN9B00141 CM to r «fi 6 " CD IGC-172 1996 Biological Indicators for Radiation Absorbed Dose : A Review Solomon FD, Paul,, P Venkatachalam and R H Jeevanram GOVERNMENT OF INDIA DEB^BTMENT OF ATOMC ENERGY INDIRA GANDHI CENTRE FOR ATOMIC RESEARCH KALMKKAM
Transcript
IN9B00141
CM to
r «fi
6 "CD
IGC-1721996
Biological Indicators for RadiationAbsorbed Dose : A Review
Solomon FD, Paul,, P Venkatachalam and R H Jeevanram
GOVERNMENT OF INDIA DEB^BTMENT OF ATOMC ENERGY
INDIRA GANDHI CENTRE FOR ATOMIC RESEARCH KALMKKAM
IGC-1721996
GOVERNMENT OF INDIADEPARTMENT OF ATOMIC ENERGY
BIOLOGICAL INDICATORS FOR RADIATIONABSORBED DOSE : A REVIEW
Solomon F.D. Paul, P. Venkatachalam and R.K. JeevanramHealth and Safety Division
Safety Research and Health Physics Group
Indira Gandhi Centre for A tomic ResearchKalpakkam - 603 102 (TN)
India
CONTENTS Page No.
1. Introduction 12. Qualities Required for Biological Indicators 13. Types of Indicator 4
•This report forms a part of Ph.D. thesis of Shri Solomon F.D.Paul to besubmitted to the University of Madras under the supervision ofDr. R.K. Jeevanram, Health and Safety Division, SR&HPG, IGCAR,Kal'pakkam603 102.
1.0 IntroductionThe changes in biochemical,
biophysical, immunological, cytogeneticaland molecular events taking place in a celldue to irradiation are used to measureradiation exposures. Exposure to ionisingradiation results in excitation of atoms,formation of ions and free radicals causingalterations in the arrangement ofmolecules. As described in Fig. 1, directeffect affects the sensitive molecules in the
DIRECT ACTION
Fig. I Direct and Indirect effect of IonisingRadiation
cells by the direct absorption of energyfrom radiation. In the case of the indirecteffect radiation induces the formation offree radicals which in turn affect theorganic molecules. Indirect effect isconsidered significant because nearly 70%of trie cell contains water.
Biologic.il dosimetry methodsmonitor changes in biomolecules in vivoand are important for monitoring persons
involved in radiation accidents, withassociated external and internal exposures.For while external exposures can also bemonitored with the help of physicaldosimeters, there are no such metersavailable for measuring internal exposures.Biological dosimetry therefore plays animportant role in monitoring personsinvolved in radiation accidents. Furthermore, biodosimetry can help to verify theresults derived from physical dosimetersduring improper use of these dosimeters.Determination of long term chronicexposure is another possibility. Presentlythe estimation of absorbed dose using suchbiological dosimeters is sensitive only atradiation levels above 0.10 Gy and doesnot therefore serve routine monitoringpurposes.
A number of biological indicatorshas been tried for estimating radiationdose. The main principle underlying thebiological indicators is that the radiationabsorbed dose leads to certain changes inbiomolecules particularly DNA, proteins,lipids etc. These changes alter theprodromal, hematological, physiological,biochemical, biophysical, immunological,cytogenetical and molecular events, thattake place in the organism. These changescan be used as an index to measure thedose. This report brings out the advantagesand disadvantages of various biologicalindicators that are available as on today.
2.0 Qualities required for biologicalindicators
Ideally a dosimeter for measuringradiation exposure should be sensitive,reproducible and practical (Muller andStreffer, 1991) as well as capable ofresponding to different energies and dose-rates. A biological indicator should reflecta measurable biological effect that isproportional to dose. Some importantcriteria are as follows:(a) The indicator should be specific toionising radiation.(b) The assessment should be rapid andreproducible.
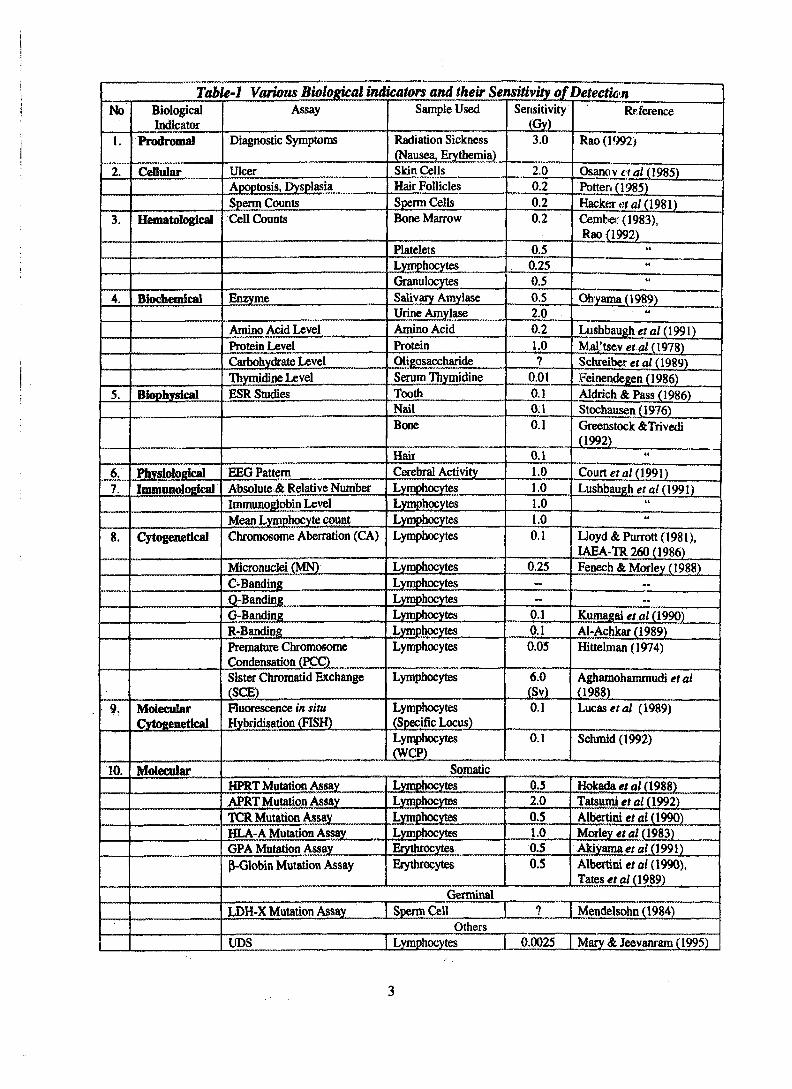
Table-1 Various Biological indicators and their Sensitivityjif DetectionNo
(c) It should have the capacity to yield ameasure of the dose even after a longperiod after exposure.(d) The method should be sensitive enoughto detect low doses of ionising radiation.(e) It should have the capacity todifferentiate between external and internalexposure, partial and whole body exposureand acute and chronic exposure.(f) The capability to differentiate betweenexposure due to high LET radiation fromthat of low LET radiation.
3.0 Types of indicatorsSeveral types of indicators have
been proposed to estimate radiationabsorbed dose. It is clear from the stringentrequirements and criteria for the suitabilityof a technique to perform as biodosimeter,that no single biological indicator cansatisfactorily meet all of these criteria(Muller and Streffer 1991 and Straume etal, 1992). The complete summary ofvarious biological indicators discussed inthis review is presented in Table-1. Theonset and fading time of each indicator isshown in Fig. 2. Following paragraphsdiscusses the various indicators in depth.
3.1 Prodromal IndicatorsProdromal response such as
anorexia, nausea, vomiting, diarrhoea,intestinal cramps, salivation, dehydrationand neuromuscular cramps are caused byacute whole body exposure. Theprodromal response appears during the first48 hours. The frequency of incidence ofprodromal syndromes, the time course oftheir appearance and their persistence aredose dependent. Whole body exposure to adose of approximately 2 Gy results in 50%of the exposed individuals showingprodromal effects with an average latentperiod of 3 hours. But partial bodyexposure do not eilict the same response.At a higher whole body dose, of about4 Gy, the prodromal syndromes manifestwithin an hour following exposure and lastfor a few months (Rao 1992).
3.2 Cellular indicatorsThe cells of the body such as skin,
blood, spleen, hair etc. undergo certainchanges due to radiation. These changescan be used to measure radiation dose.
3.2.1. Skin cellsThe skin and its constituent parts
consist of actively renewing cells, andhence they are sensitive to radiation. It hasbeen shown that the radiosensitivity ofepidermal cells is about 5 Gy. A thresholddose of 2 Gy induces skin erythema(Osanov et al, 1985). It is considered thatthe maximum dose of X-rays tolerated bythe skin with a single external exposure isabout 10 Gy. At higher doses, dermatitisappear and then ulcers. The erythemapersists for a period of few months.
3.2.2 Hair folliclesThe hair follicles during formation
contain rapidly proliferating cells. Onexposure to radiation the hair follicleundergoes dysplasia i.e., reduction in widthof the hair follicle and apoptosis (celldeath) at high doses. The above changesare used for biological dosimetry and havebeen described by Vladimirov and Smirnov(1978), Well (1981) and Potten (1985). Adecrease in width of the hair by 7% forevery Gy has been reported. Using thisprinciple a dose as low as 0.2 Gy can beestimated (Potten 1985). Dose dependentreduction of hair width for a dose range of0.5 to 5.0 Gy was best fitted using linearquadratic equation. The response time ofdysplasia is about 12 hours after localisedpartial body exposure to radiation and fallsback to normal in a few months time. Thismethod may be applicable for the earlydetection of persons exposed to highdoses.
3.2.3 Spermatogenic cellsIt has been shown that radiation
causes reduction in sperm counts andviability and also brings about changes inthe morphology of sperm cells. Thedecrease in sperm counts however, may
Fig. 2 Onset and Fading of Various Biological IndicatorsBiologicalIndicators
i1 I l l lSalivary Amylase l ^ ^ j ^ ^ | l i H | |
Urine Amylase 111 | 1 IBiffliitfiAmino Acid j f i f l iMProtein .flPilliifflllliffllllllflMBOliKOsaccharide | ?Serum Thymidine ifillMHBi 1 1 I 1 !
Tooth fifflfflffilHlilllHNail lMHIlMill'™l^^™™^^^%IMBone l l MHair JilllifHH^^Cerebral Activity | B J | i | M B l"" "T" 1Lymphocytes :^m^WmmlmMm 1 ILymphocytes ^WmmmBBBMBBBBK^BKBMLymphocytesLymphocytesLymphocytesLymphocytesLymphocytesLymphocytesLymphocytes
Lymphocytes —J Hi Hi Hi* Used for conformation ofDkentric chromosomesM, H, D, W, M, Y, I-Minutes, Hours, Days, Weeks, Months, Years and InfinityUS, S - Unstable aberration and Stable aberrationRSP, WCP - Repetitive Sequence Probes, Whole Chromosome Painting
not be observed until approximately 45days after exposure and lasts for a fewmonths before it returns to normalcy. Thisdelay allows one to determine the normalbaseline sperm counts in an individual ifsamples are collected well before the effectof radiation exposure is manifested. Inaddition to. this, the changes in the DNAcontent of the sperm cell using flowcytometric study enables the doseestimation (Gasinska and Wilson 1988,Hacker et al, 1981 and Hacker et al,1982). The sensitivity of this method wasfound to be 0.1 Gy. The restoration ofsperm number and fertility makes thisbiological parameter unsuitable lor longterm estimation of dose. None of thesemethods has reached any perfection as ontoday for use in biological dosimetry.
3.3 Hematological indicatorsLymphocytes are more radiation
sensitive among the blood cells as the sizeof their nucleus is large. The damagepersists in the body comparatively for alonger period of time as lymphocytes donot normally divide in circulation. Hencethey are ideal for studying absorbed dose.The changes occurring in chromosomes ofthese cells are useful in monitoring a doseas low as 0.05 Gy.
In addition to lymphocytes,counting of leukocytes and reticulocytesmay be of great help in assessing theabsorbed dose (Biender and Nothdurft1986, Messerschmidt 1985, Dehos andHinz 1984 and Feinendegen 1980). Stemcell, reticulocyte count is a quick andrather uncomplicated method that does notrequire any specialised laboratory (Flienderand Nothdurft 1979).
Chaudhuri and Messerschmidt(1982) have s'nown that the amount ofreticulocytes 'JI peripheral blood generallyreflects the, bone marrow status oferythropoiesis and may therefore be usefulin the evaluation of radiation injury. Theseinjuries are produced with a dose of
0.25 Gy and visible evidence of theirpresence lasts in the circulation for acouple of months. Dose dependentdecrease of reticulocytes was noticed. Theuse of cytometric quantification ofreticulocytes can well be employed forfaster results.
Erythrocytes can also be used forthe estimation of radiation absorbed dose.Butkowskji et al, (1987) used free flowelectrophoresis technique, in which the redblood cells are subjected to KMnCU andare subjected to electrophoresis. Theelectrophoretic behaviour of the cellmembrane is studied and is compared withthe normal cells. This technique cannot berelied upon for long term studies as thechanges in the behaviour persist for a shortperiod of time. The study of single gelelectrophoresis behaviour of lymphocytecells are also gaining lot of importance.The method is other wise called the Cometassay as the aberrated cells when run on aagar gel produce a Comet shaped tail. Thesize of the tail is directly proportional tothe amount of damage (Ashby et al,1995). The cloud of tail carry largenumber of broken DNA fragments. In thenormal cells the Comet shaped tail isabsent. On the whole, the size of the tailand the number of cells carrying the tail,would reflect the dose received by a person(Nocentini 1995). The image processingdevise enable the study of these Comets inease.
3.4 Biochemical indicatorsNoticeable increase or decrease in
biochemical levels have been observed incells following exposure to radiation. Theincrease or decrease in levels of thesebiochemicals have been proposed as anindicator for the assessment of dose. Thedisadvantage of this technique is that thesebiochemicals return to normal levels veryfast. Therefore this technique is useful onlywhere a rapid sorting of victims is requiredfollowing accidental exposures (Huebneretal, 1986).
3.4.1 Enzyme activityIt has been shown that the levels of
enzymes such as glutamic oxaloacetictransaminase, amylase, lacticdehydrogenase, and alkaline phosphatasepresent in serum could be used to estimateradiation dose (Martin et al, 1991, Boegl.efal, 1983, Boegl et al, 1984, Ohyama 1989and Hofmann et al, 1990). It has also beenshown that acid phosphatase and alkalinephosphatase in peripheral lymphocytes andgranulocytes can be used to estimate adose of the order of 0.01-1.90 Gy (Gu-Yuanxi and Cui-Yufang 1988).Biochemical indicators are in generalprecise but cannot be effectively employedto assess the dose, as the level of enzymedecreases with in few days and are restoredto normalcy.
3.4.1.1 Salivary amylaseIt has been found that persons
exposed to radiation in Chernobyl accidenthad increased levels of salivary amylasedue to the malfunction of liver and kidney(Ohyama 1989). 85 to 90% of the exposedpopulation were assessed for absorbeddose using this method. A dose rangingfrom 0.5 to 1.0 Gy could be estimated bythis method (Hofmann et al, 1990). Theincrease in amylase level is seen minutesafter exposure and lasts for few days. Thelimitations are that partial irradiation ofabdomen did not increase the amylase levelin the serum whereas the partial irradiationof the salivary gland (parotid gland)showed increased amylase level in theserum (Hennequin et al, 1989). Accordingto Hofmann el al, (1990) a rough estimatecould be obtained, with high exposure.Feinendegen (1986) felt that this method ispromising because of the simplicity,reproducibility and sensitivity under certainexposure conditions.
3.4.1.2 Urine amylaseSimilar to salivary amylase, urinary
amylase also indicates the absorbed dose.A linear dose dependent increase in the
urinary amylase has been observed(Ohyama 1989). This method has beenused for diagnosis, treatment as well asprognosis of the patients affected inChernobyl accident few days afterexposure (Ohyama 1989). As mentionedabove this method poses a lot of practicalproblems in calibration. As the amylaselevel reaches nonnalcy within few days ofexposure. This method may be useful inmass sorting of the persons within a fewdays after nuclear accidents.
3.4.2 Amino acid levelsA transient increase in the levels of
blood amino acids such as cystine, valine,leucine, phenylalanine and others have beenobserved in experimental animals exposedto radiation and also in humans followingradiation accidents.
Beta aminoisobutryic acid(BAIBA), a metabolite of thymidine, wasfound to be elevated as early as 3 daysafter a dose 0.22 to 3.65 Gy and persistedfor a period of 8 days in the high dosegroup. However a clear dose dependencycould not be obtained (Lushbaugh 1991).
3.4.3 Protein indicatorsIt has been shown that C-reactive
protein increases due to radiation exposure(Mal'tsev et al, 1978). The level of this C-reactive protein in the serum increaseslinearly during acute exposure (1-12 Gy)from tenth day and lasts for about a monthtime. This may be applicable for prognosisof acute radiation diseases. The increase inprotein levels however, could not beapplied as a successful dosimeter as it fadesin short period of time.
3.4.4 OligosaccharidesIncrease in lectin binding to the
platelets, lymphocytes and monocytes wasnoticed when exposed to doses between0.5 to 5 Gy (Schreiber et al, 1989). One ofthe methods of estimating lectin binding tocells is by using either tritium orfluorescence labelled concanavalin-A and
analysing labelled cells using either atritium counter or flow cytometryrespectively. The lectin binding to cellshowever is not suitable for measuringradiation dose as the binding property isnot reproducible nor significant.
3.4.5 Serum thymidineRadiation causes certain changes in
the structure, charge and fluidity of the cellmembrane, which results in a rapidaccumulation of thymidine in the serumeven at doses below 0.01 Gy. This appearsto be a temporary physiological equilibriumdisturbance and not related to cellulardamage. The transitory effect lasts forabout 4 hours and then falls back to thepre- irradiated level. The result obtainedfrom in vivo irradiation of mice could notbe seen in in vitro irradiation. This makesthe technique unsuitable for biodosimetry.
3.5 Biophysical indicators3.5.1 Electron Spin Resonance (ESR)
Free radicals formed by irradiationare trapped in solids, and their number isproportional to the absorbed dose. Thesefree radicals can be studied using ESR.The absorption and consequent emission ofelectromagnetic radiation by unpairedelectrons in certain types of atoms in thepresence of strong magnetic field, may aidin studying various biologicd materialswhich carry free radicals, ESR signals frombone (Greenstock and Trivedi 1992), teeth(Aldrich and Pass 1986), nails(Stockhausen 1976) jind hair (Greenstockand Trivedi 1992) have l>een used inbiological dosimetry. The sensitivity rangesbetween 0.5 and 5.0 Gy and is useful forestimating partial body exposure except forbone. Bone cells are useful in theestimation of whole body radiation(Greenstock and Trivedi 1992). Dentalenamel is the only living tissue which couldstore ot retain indefinitely the cumulativeradiation dose (Aldrich and Pass 1986) thedetection lii.hit is about 0.1 Gy. Poorreproducibilrty have limited their
applications. (Dalgarno and McClymont1989). ESR has been used initially infinding the age of mollusk shells (Anatoly1989). The dating was then converted tothe use of dosimetry to study a cumulativedose (Skinner 1989 and Rainer 1989). InESR, dating the sample under investigationacts as a dosimeter which record the dosesfrom all natural radioactive sources fromits immediate surroundings, as well ascomponent of cosmic rays (Rainer 1989).
It has been found that irradiatedDNA has shown significant levels ofradicals compared to unirradiated DNA(Stockhausen 1976).
In case of accidents, operators maycarry tablets, which consists ofcarbohydrates which are known to formradiation induced radicals with a reliablyhigh yield. Other tissue equivalent materialsare also being used to study the doseabsorbed using ESR. A simple aminoacidL-alpha-alanine with a linear dose responsehave also been used as a dosimeter materialin the range of 1 to 105 Gy dose (Ahlersand Schneider 1991; Onori et al, 1990).The radicals are quite stable for years andcan be used to study acute and chronicdoses.
3.6 Physiological indicatorsElectrophysiological changes such
as cerebral electrical activity, brain bloodflow, blood acid base equilibrium, heartand respiratory frequency etc. of animals(rodents, primates and humans) have beenproposed as biophysiological method ofestimation of the dose. In the brain, there isa regular, rhythmical change of electricalpotential, due to the rhythmical dischargeof energy by nerve cells. These changes canbe recorded graphically and the "brainwaves" examined. These recorded electro-encephalo-grams (EEG) are useful inidentifying diseased condition where thefrequency distribution of the waves ismodified. It has been reported that thesechanges are altered due to ionisingradiation. The EEG analysis makes it
possible to assess the absorbed dose with athreshold of about 1 Gy for whole bodydose which would reach normalcy withinfew days after irradation (Court et al,1991).
is currently available to assess cumulativeradiation exposure. This could be due tothe fact that information about cellularfactors involved in immune defensemechanisms is constantly evolving(Lushbaugh et al, 1991). The possibleimmunological indicators have beenreviewed by of Lushbaugh et al, (1991)and are listed below.
3.7.1 Absolute and relative numbersLymphocytes consist of a mixture
of different cellular subsets, Two broadclasses are B and T-lymphocytes. Thechanges in the ratio of B and Tlymphocytes following irradiation havebeen studied (Dehos et al, 1986) and alsochanges in the proportion of varioussubsets of T cells (Wassorman 1986) for adose of 1 Gy. It appears that evaluation ofa particular subpopulation of lymphocytesmight be used as an indicator of radiationdose for a period of days as repopulationoccurs rapidly in few days to weeks.
3.7.2 ImmunoglobuJin levelsImmunoglobulin levels were
monitored in lymphocyte culturesfollowing the stimulation of these cells withpoke weed r.nitogen (PWM). Little isknown abouv the time sequence betweenradiation exposure and in vitro Igproduction. It has been shown that duringand following radiation therapy, in vivoproduct .ion of three immunoglobulinclasses (i.e., igM, IgG, IgA) remainedsuppressed for 12 to 18 months. It isunlikely that this approach will proveussful in estimating dose years afterradiation exposure.
3.7.3 Mixed lymphocyte cultureMixture of two populations of
lymphocytes, from two different persons,have been found to stimulate growth ofthe responder cells. Measuring theincorporation of 3HTdR into the DNA ofthe responder cells would give the index toradiation insult. This test is at an earlystage of development in radiation doseassessment. Experience with radiationtherapy patients indicates that significantdepression of incorporation of 3HTdR inlymphocytes occurs in the dose range of 1to 6 Gy. The mechanism by which radiationaffects this function of the immune systemis not known. Currently, the sensitivity ofthis test requires doses greater than 1 Gy.As with the other tests, interferencefactors, individual variability, and lack ofstandardisation for good reproducibilityneed to be determined before this test canbe used for estimation of radiation dose.
3.8 Cytogenetical IndicatorsAn ideal and possible technique in
the estimation of radiation absorbed dosewould be the cytogenetical technique.Cytogenetic detection and measurement ofhuman exposures to ionising radiation havebecome well established during the past 2to 3 decades. The birth of cytogenetics asan experimental science might reasonablybe considered to have taken place in thelate 1930s with the studies of Karl Sax onthe induction of chromosome aberrations inTradescentia microspores by x-rays (Sax1938). The pioneering work oncytogenetics has evolved a long way fordetection of various clastogenic insult.
R, and Q-Bands)4. Premature Chromosomal Condensation
(PCC)5. Sister Chromatid Exchange (SCE)
3.8.1 Chromosomal aberrationThe damage caused by ionising
radiation to nuclear DNA of the cell whenit is at Go or d phase of the cell cycleresults in chromosomal aberration. Thischromosomal aberration results instructural changes which is seen asdicentric or ring chromosome. Fig. 3
levels of dicentric chromosome (1/1000cells) makes this technique comparativelymore sensitive. The sensitivity of thistechnique is 0.10 Gy. Dicentric frequencyfade with a half-life of about 2 years. Moredetailed review of various chromosomalaberration can be found in Paul et al,(1996).
INTERPHASE MITOSIS
PROPHASE METAPHASE
CELL
O2Oacxo
UNCOILED
INTERPHASE•
CHROMOSOME
ACENTRICCHROMOSOME
• 1
ELECTRON TRACKBREAKING THECHROMOSOME.
DAMAGED. _»
INTERPHASECHROMOSOME
OI/)o2oIo
MISREPAIRFUSION OF
CHROMOSOMES
1*2 METAPHASE
II
oaczUloo
I IACENTRIC
CHROMOSOME
Fig. 3 Radiation induced dicentric and acentric chromosomes
depicts the formation of dicentric and itsassociated acentric fragments caused dueto ionising radiation. The dicentric or ringchromosomes are seen clearly when thecells are in metaphase stage. The peripheralblood lymphocytes which are normally inGo stage can be made to undergo celldivision by adding a plant lectin such asphytoheraagglutinin and then arresting thecells at metaphase stage by addingcolcemid. These cells are stained withGbmsa, fixed on a microslide and viewedthrough microscope. The dicentricfrequency indicates the absorbed doses.Presently this is the most reliable method tomeasure riidiation absorbed dose duringaccidental conditions. The low background
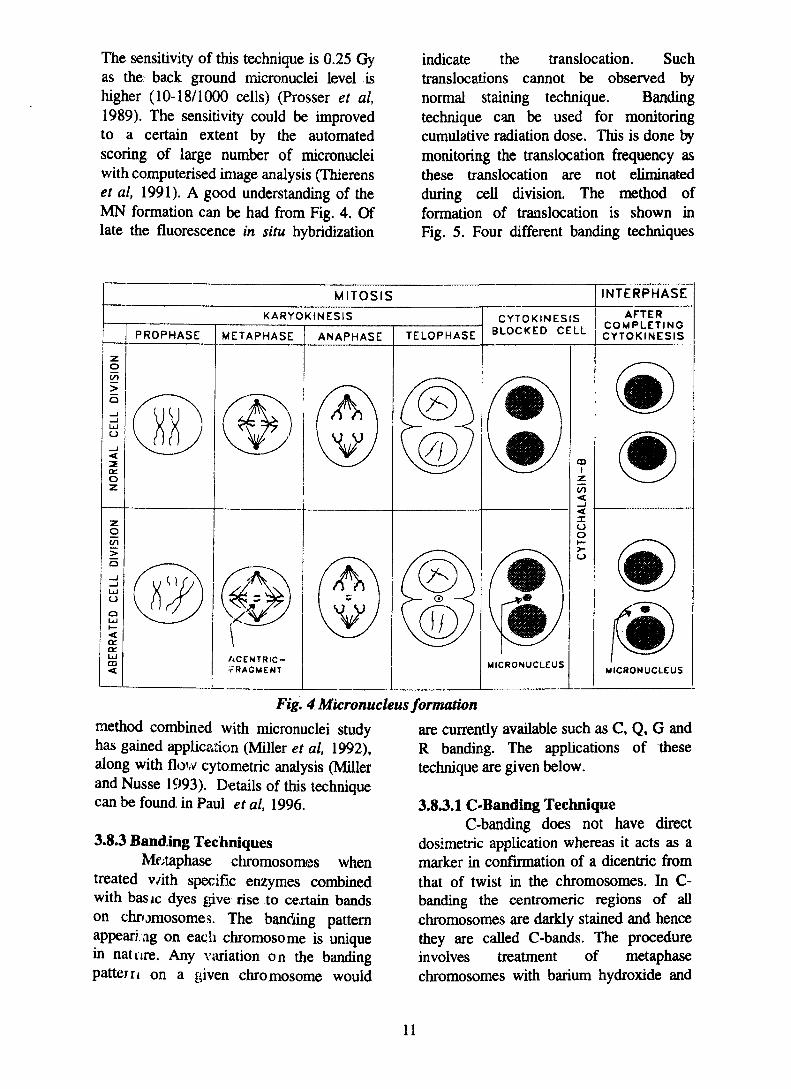
3.8.2 Micronuclei (MN)The other cytogenetical method is
the micronuclei technique. The mainadvantage of this technique is that scoringof micronuclei is faster and easier than theconventional dicentric chromosomeaberration technique (Fenech and Morley1986, Fenech et al, 1988, Prosser et al,1988). Micronuclei are the product ofbroken or lagging chromosome whichfailed to become incorporated into either ofthe two daughter nuclei during cell division(Prosser et al, 1989). As in dicentricchromosome aberration technique theblood is cultured with phytohemagglutinand the cells are arrested at cytokinesisstage by the addition of cytochalasin-B.
10
The sensitivity of this technique is 0.25 Gyas the back ground micronuclei level ishigher (10-18/1000 cells) (Prosser et al,1989). The sensitivity could be improvedto a certain extent by the automatedscoring of large number of micronucleiwith computerised image analysis (Thierenset al, 1991). A good understanding of theMN formation can be had from Fig. 4. Oflate the fluorescence in situ hybridization
indicate the translocation. Suchtranslocations cannot be observed bynormal staining technique. Bandingtechnique can be used for monitoringcumulative radiation dose. This is done bymonitoring the translocation frequency asthese translocation are not eliminatedduring cell division. The method offormation of translocation is shown inFig. 5. Four different banding techniques
MITOSIS INTERPHASE
KARYOKINESIS
PROPHASE METAPHASE ANAPHASE TELOPHASE
CYTOKINESISBLOCKED CELL
AFTERCOMPLETINGCYTOKINESIS
ACENTRIC-V'RACMENT MICRONUCLEUS
MICRONUCLEUS
Fig. 4 Micronucleus formation
method combined with micronuclei studyhas gained application (Miller et al, 1992),along with flow cytometric analysis (Millerand Nusse 1993). Details of this techniquecan be found in Paul etal, 1996.
3.8.3 Banding TechniquesMe^taphase chromosomes when
treated v/ith specific enzymes combinedwith basic dyes give rise to certain bandson chromosomes. The handling patternappearing on each chromosome is uniquein nature. Any variation on the bandingpattern on a given chromosome would
are currently available such as C, Q, G andR banding. The applications of thesetechnique are given below.
3.8.3.1 C-Banding TechniqueC-banding does not have direct
dosimetric application whereas it acts as amarker in confirmation of a dicentric fromthat of twist in the chromosomes. In C-banding the centromeric regions of allchromosomes are darkly stained and hencethey are called C-bands. The procedureinvolves treatment of metaphasechromosomes with barium hydroxide and
11
then staining with Giemsa. Lucas (1989)have used centromeric and teloraeric
are used as reference points or markers andalteration of the sequence of these is
iNTERPHASE MITOSIS
PROPHASE METAPHASE
2OV)
ooacxo
DAMAGED
MISREPAIRTRANSLOCATION '
OF CHROMOSOMES'*
ELECTRON TRACKBREAKING THECHROMOSOME
INTERPHASECHROMOSOME
Oo2oDCXu
1&2
CHROMOSOME
CONDENSATIONAT MITOSIS
METAPHASE
Fig. 5 Radiation
markers to identify translocation anddicentric in metaphase spreads.
3.8.3.2 Q-Banding TechniqueThis technique is very simple.
Chromosome preparations are incubated inan aqueous solution of the fluorescent dyesuch as quinacrine mustard dihydrochloride(Caspersson et al, 1968), washed andexamined using a fluorescence microscopy.Although this method is quite simple toperform the stain appears to fade onexcitation and analysis becomes difficult.Thus it is found that the application of Q-banding for biological dosimetry is verylimited.
3.8.3.3 G-Banding TechniqueThis is the most widely used
technique among the banding procedures.In simple terms the specific areas on thechromosome are digested using trypsin, aproteolytic enzyme, which produce specificbands on the chromosomes. These bands
induced translocationconsidered an aberrated chromosome. Theanalysis of banded chromosomes, howeverrequires substantial cytogeneticalknowledge and experience, and is timeconsuming. Chromosome aberrations in thelymphocytes of radiation workers werestudied by Kumagai et al, (1990) usingtranslocation frequency as an index. Thetrypsin banded chromosomes can beemployed for late effects of long termexposure to low doses of radiation.Kumagai et al, (1991) have compared thefrequency of translocations in controlsubjects as well as in radiation workers.The frequency was found to besignificantly higher in radiation workersthan in the control group. These authorshave found, that the translocations weremainly of the chromosomes 1,2,7 and 14in radiation workers and of 1,3,7 and 14 incontrol group. Similar distribution oftranslocation was observed in plutoniumworkers by Twan and Holdsworth (1995).They have pointed out possibility of
12
occurrence of hot spots in some regions ofchromosomes. Shuqing et al, (1988) haveshown that radiation induces almost equalamounts of stable and unstablechromosome aberrations. Shuqing et al,(1988) and Matsukoa et al, (1994) haveshown that the yield of unstablechromosome aberration decreases over aperiod of time or on completion of first cellcycle where as the stable aberration doesnot change.
3.8.3.4 R-Banding TechniqueThe staining pattern obtained with
R-banding is exactly opposite to thatobtained with Q and G-banding. This canbe achieved by incubating the chromosomepreparations either at higher temperatureor at a suitable pH. The slide containingchromosome preparation is placed in a hot(87°C) phosphate buffer saline for 20-60minutes at a pH 5.1. This is stained withGiemsa at 87°C rinsed and seen for bands.
R-banding technique has been usedto calculate the translocation frequency inblood samples exposed to radiation (Al-Achkar, 1987, 1989, Sabatier et al, 1988).Al-Achkar (1987) has studied the effect oflow dose (0.05 Gy) gamma radiation fromsamples obtained from healthy humanindividuals of various age. The sensitivityof detection of dose is poor in this methodbecause of high background frequencies.This technique may be applicable in youngadults and older people only when thedoses are more than 0.1 and 0.2 Gyrespectively. Al-Achkar has found thatthere is a significant increase in reciprocaltranslocaiion and dicentric yield anddecrease in deletions for a dose of 0.5 Gy.
3.8.4 Premature ChromosomeCondensation (PCC)
The genomic damage is usuallymeasured when the cells are in metaphase.The time taken by the damaged cells toattain mitosis may be delayed. Moreover,slow growing or non-dividing cells aredif/icult to study by conventional means.
Thus it becomes important to analyse thechromosomes at interphase stage.
In the year 1970, Johnson and Raohave reported that the cells in mitotic phasecould induce the condensation ofchromatin of interphase cells on fusion.This is called premature chromosomecondensation. Presently Chinese HamsterOvary cells (CHO) are fused withlymphocytes using polyethylene glycol(PEG). Earlier sendai virus was used forfusing the cells and replacement of thesame with PEG has improved the PCCmethodology (Pantelias 1984). The factorspresent in the mitotic cells, which inducecondensation of chromosomes are not yetwell characterised, however much progresshas recently been made in this area(Dumphy and Newport 1988). Themorphology of the PCC reflects the stageof the interphase cell at the time of cellfusion. Cells fused at G| phase give rise toPCC with a single chromatid perchromosome, while cells fused at G2 giverise to PCC with two chromatids perchromosome. Cells fused during DNAsynthesis yield PCC with a pulverisedappearance.
Cells at Gi and G2 phase are mostsuitable for chromosome aberration studiesbecause the PCC are discrete and can becounted and analysed for structuralalterations. For example, PCC with Gi cellsgives chromosome fragments (Waldren andJohnson 1974) while G2 cells givechromatid gaps, breaks, and exchanges(Hittelman and Rao 1974).
PCC offers several advantages, andit is presented as an alternative to theconventional cytogenetic technique. First,the chromosomal aberration damage maybe visualised in about 2 hours. Second, thechances for inteiphase cell death orchromosomal repair are decreased. Third,for PCC analysis of about 100mononuclear blood cells are sufficient toestimate radiation damage even at lowdoses. Fourth, the linear dose- responserelationships may be standardised easily.
13
Finally the inter laboratory variation inresults are reduced in PCC as the variablesintroduced by the stimulation and cultureof lymphocytes required for conventionalcytogenetic techniques are not present inthis technique. This approach thus providesa direct and most sensitive cytogeneticalmethod for the estimation of radiationinjury and repair. A dose of 0.05 Gy can bedetected using this method. Of late PCChas been effectively combined with in situhybridisation method to achieve moresensitivity (Brown et al,. 1992).
3.8.5 Sister Chromatid Exchange (SCE)SCE are reciprocal exchanges
between sister chromatids. Although DNAbreakage and reunion is presumablyinvolved, the exact molecular mechanismremains unknown. They are producedduring DNA replication. In order to seeSCE, cells in culture are allowed toreplicate twice in bromodeoxyuridine(BrdU). Incorporation of BrdU into newlysynthesised DNA in place of thymidinemodifies the staining properties of thechromatids. For example, the fluorescentstain, Hoechst 33258, stains only theparticular chromatid where in only onestrand is substituted with BrdU. If a sisterchromatid exchange (SCE) has occurred,this is readily recognised by the bright anddim fluorescence patterns along thechromatids. Another technique is thefluorescence plus giernsa staining (FPG)which involves, the differentiaUy staining ofthe BrdU treated culture with Giemsa. Thestained slide can be studied under anordinary microscope unlike the above,which requires a fluorescence microscope.
SCFi are most efficiently induced bysubstances that form covalent adducts withthe DNA., or interfere with DNA precursormetabolism or repair. Where as potentclasto^ens such as bleoniycin and ionisingradiation show poor response to induceSCxi indicating SCE are unrelated tosixuctural chromosome aberrations anddiese mechanisms involved in the types of
lesion are obviously dissimilar (Twan andHoldsworth 1992, Perry and Evans 1975).However, it has been reported that highLET radiation like neutrons (Savage andHolloway 1987, 1988) and alpha particlesare capable of producing SCE when cellspasses into S-phase (Aghamohammadi etal, 1988,1989). This technique finds someapplication in low level radiation exposure.The application of SCE in biologicaldosimetry has been discussed by Mary andJeevanram 1995.
3.9 Molecular cytogenetics indicatorsThe technique such as fluorescent
in situ hybridisation is useful in studyingthe cumulative radiation dose. The in situhybridisation provides a means to locatetranslocations visually by identifying thepresence of certain DNA sequence of onechromosome on another chromosome. Inthis method the DNA of metaphasechromosome is first denatured at 70°C,dehydrated and air dried. Similarly thebiotin labelled DNA probe is alsodenatured at 70°C, mixed with denaturedmetaphase chromosome and incubatedovernight. The biotin labelled probes aredetected on metaphase chromosomes bytreating sequentially with avidin,conjugated to fluorescein, a monoclonalantibody against avidin labelled with biotinand finally a second layer of avidinconjugated to fluorescein. This gives theprobe a bright fluorescence and is easilyseen on the metaphase chromosome. Oflate more than one chromosome probe withdifferent colours are used for translocationstudies. The multicoloured chromosomesare seen visually by using multiple bandfilters which are called chromosomepainting (Popp and Cremer 1992 Schmid etal, 1992 and Yoshida 1992). This methodof chromosome painting or specificrepetitive in situ hybridisation enables theeasy study of chromosomal damagesinduced by ionising radiation (Lucas et al,1989, Gray et al, 1991, Lucas et al, 1992,Natarajan et al, 1992, Buchinger et al,
14
1993 and Matsuoka 1994). A moreelaborate discussion of in- situhybridisation has been discussed elsewhereVenkatachalam et al,. (1996). Presentlyfluorescence in situ hybridization offers amethod of choice for estimating cumulativeradiation exposure.
3.10 Molecular indicators3.10.1 Mutational assay
Ionising radiation has also beenshown to induce specific locus mutations.Measurement of the somatic mutationfrequency would serve as an indicator ofhuman exposure to radiation (Kushiro etal, 1992). However, methods for thequantitative detection of these mutations inman are very limited. Recently, severalmethods for the detection of mutations at
cell system used in these assays at presentare the red blood cells and the T-lymphocytes. Somatic mutation in theRBCs can arise in any of the nucleatedprecursor cells. T-lymphocytes arenucleated white cells that are widelydispersed in peripheral blood, lymph nodesand most of the body tissues. Maturelymphocytes arise from stem cells in thebone marrow and migrate to the thymus,where they differentiate. Mutation in thesecells are expressed as alteration in theirproduct. Mutation of a single gene is arelatively rare event, most experimentalsystem are therefore designed to select outcells carrying mutation. These assays arebroadly classified as somatic and germinalmutation assay (See Flow Chart-1. Thevarious mutational assays which are
Flow Chart-1 Classification of various mutational assays
MUTATIONAL ASSAY
SOMATIC MUTATION GERMINAL MUTATION
Lymphocyte Erythrocyte Sperm Cell
HPRT Mutation assay
(Hypoxanthine PhosphoribosylTransferase)
APRT Mutation assay
(Adenine PhosphoribosylTransferase)
TCR Mutation assay
(T'Cell Antigen Receptor)
HLA-A Mutation assay(Human Leukocyte Antigen-A)
specific loci have been established. A pre-requisite for these mutational assays is toselect a suitable gene and suitable cell todetermine the mutational frequency. The
GPA Mutation assay
(Glycophorin-A)
fi-Globin Mutation assay
(Beta-Gbbin)
LDH'X Mutation assay
(Lactate Dehydrogenase-X)
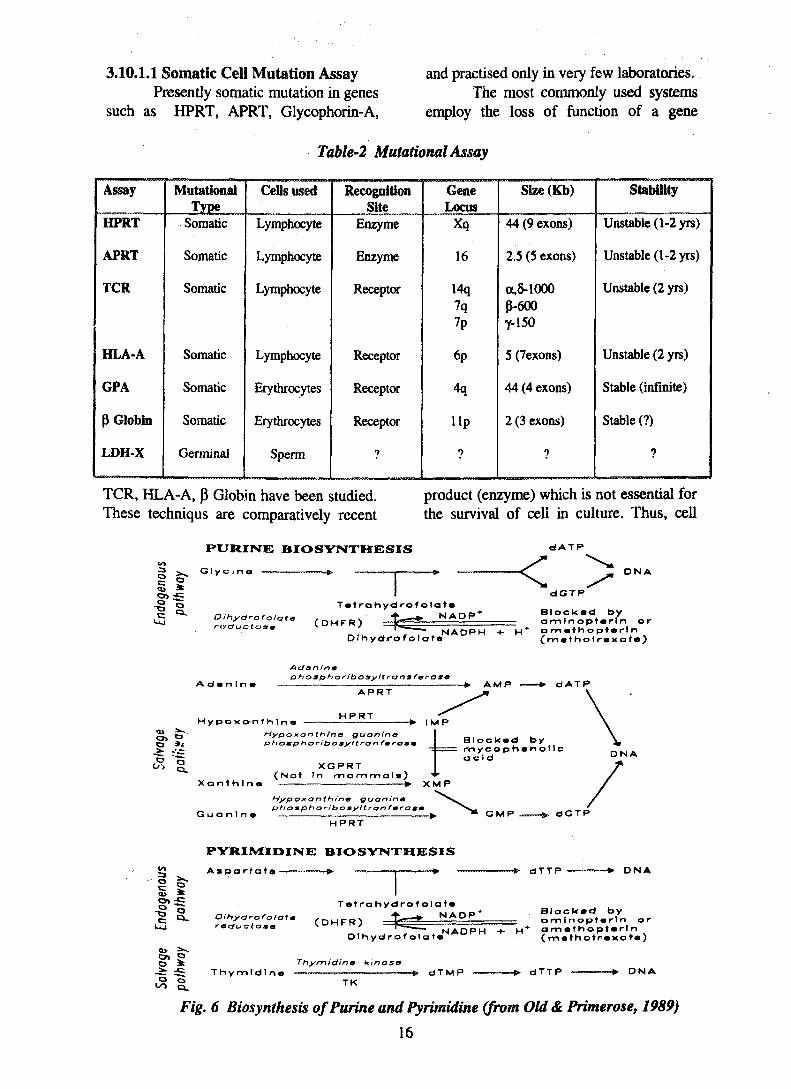
available currently are tabulated in Table-2.The gene locus and the span length of eachgene are also compared.
15
3.10.1.1 Somatic Cell Mutation AssayPresently somatic mutation in genes
such as HPRT, APRT, Glycophorin-A,
and practised only in very few laboratories.The most commonly used systems
employ the loss of function of a gene
Table-2 Mutational Assay
Assay
HPRT
APRT
TCR
HLA-A
GPA
p Globin
LDH-X
MutationalType
Somatic
Somatic
Somatic
Somatic
Somatic
Somatic
Germinal
Cells used
Lymphocyte
Lymphocyte
Lymphocyte
Lymphocyte
Erythrocytes
Erythrocytes
Sperm
RecognitionSite
Enzyme
Enzyme
Receptor
Receptor
Receptor
Receptor
?
GeneLocus
Xq
16
14q7q7p
6p
4q
lip
?
Size (Kb)
44 (9 exons)
2.5 (5 exons)
a.8-1000p-600y-150
5 (7exons)
44 (4 exons)
2 (3 exons)
7
Stability
Unstable (1-2 yrs)
Unstable (1-2 yrs)
Unstable (2 yrs)
Unstable (2 yrs)
Stable (infinite)
Stable (?)
TCR, HLA-A, P Globin have been studied. product (enzyme) which is not essential forThese techniqus are comparatively recent the survival of cell in culture. Thus, cell
to
PURINE BIOSYNTHESIS dATP
dGTP
D N A
Oihydrofolate
Tatrahyd rofolata•f. », N A D P *
Dthydrofolat«NAOPH
Blocked byamlnopUrln orom«thop)«rln(ma'holraxat*)
Adflnlnt
AtJanintfphosphoribosyltransferaso
Hypoxonth ln i
APRT
HPRT
-•• AMP dATP
-* • I M P
O 9*Hypoxonthino guaninaphompHorib omyttr&n fmramti
XGPRT(Not In mammala) *'
Xanthlna • XMP
Blocked bytnycophenollcacid
G u a n 1 n
Hypoxa nthino Quaninptiom ph oriboayltrtjn fmr
HPRT
D N A
G M P dGTP
PYRI1VIIDINE BIOSYNTHESIS
Aspartata—" J.̂ ......,». .—_ -f» — * • - * - dTTP D N A
Oihydrofolota . ,
Tetrahydrofolot*•» > NADP'
~ S Thymldlna
DIhydrof olataj
Thymidino kinasa
Blockad byamlnoptarin or
* amathoptarln(mafhatrcxata)
dTMP dTTP D N AT K
Fig. 6 Biosynthesis ofPurine and Pyrimidine (from Old& Primerose, 1989)
16
may be challenged with a mutagen (toxicdrug or ionising radiation) that wouldnormally react with fatal consequences. Ifmutation renders the gene producing thespecific enzyme ineffective, the cell willsurvive, and thus the mutation frequencycan be obtained by measuring thesurvivors. The most common of this typeare the HPRT and APRT. In HPRTsystem, the loss of enzyme, hypoxanthinineguanine phosphoribosyl transferase,renders the cell resistant to the drug 6-thioguanine (6-TG) and the enzymethymidine kinase (TK) gives resistance totrifluorthymidine (TFT). Adeninephosphoribosyl transferase (APRT)mutants are resistant to 2,6-diamino purine(DAP). Resistance enable mutant cells togrow in TG medium where as wild type(normal cells) are not able to survive inthese medium. The various mutants, theirresistance property and inhibitors availablefor cell culture is shown below. Fig. 6shows the purine and pyrimidine
The gene HPRT, an enzymeresponsible for the synthesis of purine islocated on Xq chromosome arm. HPRTgene has 44 kb and contains 9 exons andhence it has a "large target" for mutation.As mentioned earlier, HPRT-deficientmutant cell can be readily recognized bytheir resistance to the purine analogue, 6-thioguanine (TG). This is done by cloningof lymphocytes which are able to grow inthioguanine (Morley et al, 1983 andAlbertini et al, 1990). HPRT can beassayed by two methods the variantfrequency or the mutant frequency. Thevariant frequency involves the cloning oflymphocytes in presence of TG mutantcells. The growth induced is eithermeasured by BrdU fluorescence labelling
Xq
Xq
HPRT"VARIANT FREQUENCY
HPRT'
Normal
PHA Stimulated+
Thioguanine (TG)+
3HTdR or BrdU
PHA Stimulated+
Thioguanine (TG)
Cell grows
(TG")DNA
Synthesis
Cell killed DNA
Increase IncreaseJHTdR ~* counts
Increase IncreaseBrdU •""*• fluorescence
Decrease Decrease/ 'HTdR -*" counts
(TG1) SynthesisHTdR or BrdU
MUTATIONAL FREQUENCY
^Decrease DecreaseBrdU ~ * fluorescence
PHA Stimulated
Thioguanine (TG) ~
IL-2 + Feeder cells
PHA Stimulated
Thioguanine (TG) ~"
IL-2 + Feeder cells
Various cone.
In titer plates
Increase
titer value
Various cone.
In titer plates
Decrease
titer value
Fig. 7 HPRT mutational assay
increase
colony
efficiency
Decreasecolonyefficiency
17
or by autoradiography (Fig. 7). In mutationfrequency method, the lymphocytes aregrown along with feeder cells, in titerplates in presence of TG and the efficiencyis determined by increased titer valueindexing for more mutation and vies versa.Several laboratories have studied "background" HPRT mutation frequencies innormals with the T-cell cloning assay, therange of value for adults is 3.1- 9.5 x 10'6
and the mean value for a total of 252individuals is 5.3 (±2.7) x 10"6 (Albertini etal, 1990). A clear age related increase inmutational frequency has been reported. Aswith the short term assay, chemo andradiotherapy in cancer patients causesignificant rise in mutation frequency hasbeen reported. Atomic bomb survivors, 43years after exposure showed significant butsmall response as a function of theirestimated A-bomb radiation dose (Hakodaet al,. 1988). It has been shown that the
mutant response in the hprt assay decaywith time (Ammenheuser et al,. 1938),Thus it has been said that this method isunlikely to be useful for measuringcumulative dose.
Adenine phosphoribosyltrausferase(APRT), catalyses the phosphorylation ofadenine. This enzyme is encoded by a genepresent on chromosome 16. The patientwith "APRT deficiency," suffers fromkidney or ureter stone composed of 2,8-dihydroxy adenine. Tatsumi et al,, (1992)used EB virus to create immortal Blymphocyte population to assay andquantify the forward mutation fromAPRT1 to APRT"'" that is selectablebecause of its resistance to an adenineanalogue 2,6-diaminopurine (DAPR).APRT is assayed similar to HPRT assay
VARIANT FREQUENCY
APRT"
16
Normol
APRT"
16
Normal
PHA Stimulated+
Diamino purlne(DAP)
+5HTdR or BrdU
PHA Stimulated+
Diamino purine.(DAP)
Cell grows DNA
(DAP") Synthesis
Cell killed
(DAP1) "DNA
Synthesis
HTdR or BrdU
MUTATIONAL FREQUENCY
Increase IncreaseHTdR counts
Increase IncreaseBrdU ~~* fluorescence
Decrease DecreaseHTdR counts
Decrease Decrease
BrdU fluorescence
PHA Stimulated+
Diomino purine(DAP)
IL-2 + Feeder cells
PHA Stimulated+
Diamino purine(DAP)
Various cone.
in titer plates
Increase
titer value
Various cone,
in titer plates
Decrease
titer value
Increase
colony
efficiency
Decrease
colony
efficiency
IL-2 + Feeder cells
Fig. 8 APRT mutational assay
18
mentioned above (Fig. 8). Tatsumi et al,(1992) showed that the mutation increasesin a dose dependent manner. Themeasurement of adenine phosphoribosyletransferase (APRT) mutation in CHO celldid not show any changes below 2.0 Gy(Miles and Meuth 1989). Fading of themutants due to the selection of the cells
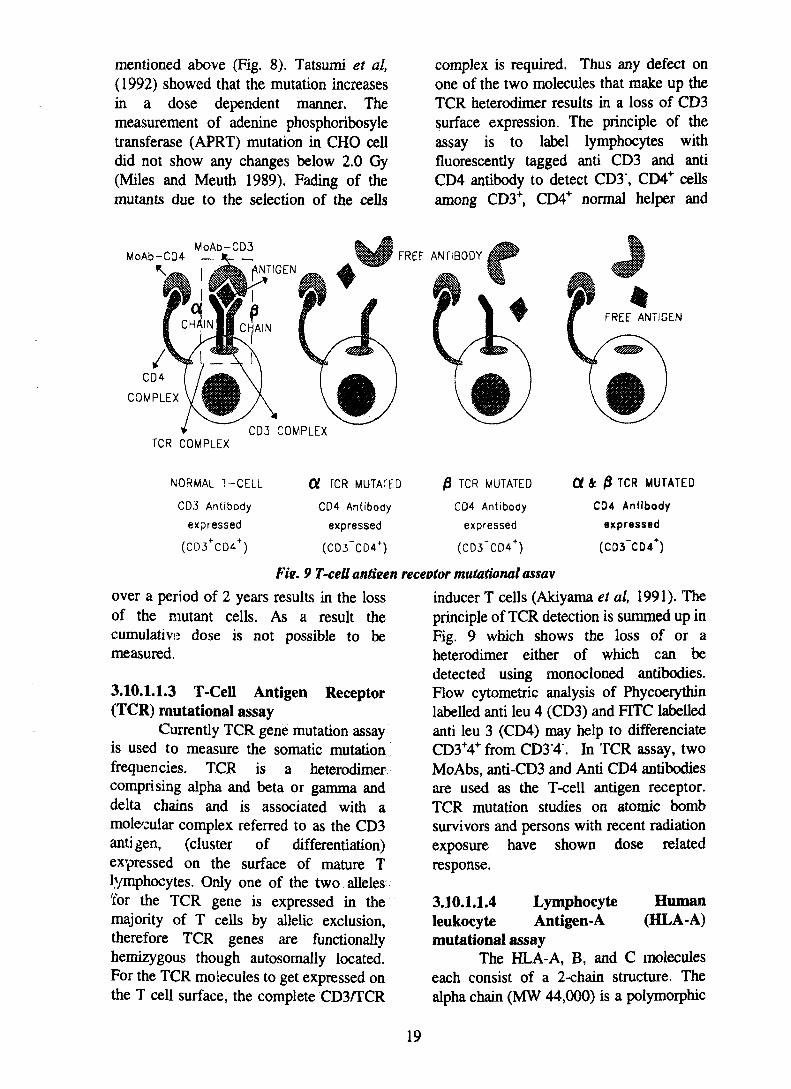
complex is required. Thus any defect onone of the two molecules that make up theTCR heterodimer results in a loss of CD3surface expression. The principle of theassay is to label lymphocytes withfluorescently tagged anti CD3 and antiCD4 antibody to detect CD3\ CD4+ cellsamong CD3+, CD4+ normal helper and
MoAb-CQ4MoAb-CD3
CD3 COMPLEX
FREF ANTIBODY
FREE ANTIGEN
TCR COMPLEX
NORMAL T-CELL
CD.'5 Antibody
expressed
(CD3+CD4+)
01 TCR MUTATED
CD4 Antibody
expressed
(CD3~CD4+)
|3 TCR MUTATED
CD4 Antibody
expressed
(CD3~CD4+)
Fie. 9 T-cell antieen receptor mutational assav
over a period of 2 years results in the lossof the mutant cells. As a result thecumulative dose is not possible to bemeasured.
Currently TCR gene mutation assayis used to measure the somatic mutationfrequencies. TCR is a heterodimercomprising alpha and beta or gamma anddelta chains and is associated with amole'cular complex referred to as the CD3antigen, (cluster of differentiation)expressed on the surface of mature Tlymphocytes. Only one of the two alleles'for the TCR gene is expressed in themajority of T cells by allelic exclusion,therefore TCR genes are functionallyhemizygous though autosomally located.For the TCR molecules to get expressed onthe T cell surface, the complete CD3/TCR
inducer T cells (Akiyama et al, 1991). Theprinciple of TCR detection is summed up inFig. 9 which shows the loss of or aheterodimer either of which can bedetected using monocloned antibodies.Flow cytometric analysis of Phycoerythinlabelled anti leu 4 (CD3) and FITC labelledanti leu 3 (CD4) may help to differenciateCD3+4+ from CD3'4". In TCR assay, twoMoAbs, anti-CD3 and Anti CD4 antibodiesare used as the T-cell antigen receptor.TCR mutation studies on atomic bombsurvivors and persons with recent radiationexposure have shown dose relatedresponse.
The HLA-A, B, and C moleculeseach consist of a 2-chain structure. Thealpha chain (MW 44,000) is a polymorphic
19
glycoprotein determined by genes in theHLA complex on chromosome 6. This isnoncovalently linked to a nonpolymorphic
mutant cells when exposed with 100 kVpx-rays. Studies on the atomic bombsurvivors showed no variation compared to
Cells
Normal
Antibody
'HLA-A, -
,HLA-Aj -
Antigen
[Presence of AnfigenJ
>>HLA-A2
Resell
-• Killed
Immuno-selection->HLA-A3 >> Killed
Mutant'HLA-A2
,HLA-A3
[Absence of Antigen]
• HLA-A2 —— ->• Survives
Immuno-selection->HLA-Aj 1 Survives
Fig. 10 HLA-A mutational assay
protein (MW 12,000), p2-microglobulin,determined by a gene on chromosome 15.The mutation on the HLA-A on theperipheral blood lymphocyte can bedetected by autoradiography and cloningversion as well. The lymphocyte HLA-Alocus mutation assay uses monoclonalantibodies specific to HLA-A2 and A3 plusits complements to kill normal cellexpressing HLA-A2 and A3 antigen on thesurface, but mutant cells lacking theantigen may survive. Morley et al, 1983have observed a linear increase in the
the control. The exact mechanism by whichthe mutants are eliminated in vivo isunknown. The HLA assay is not suitablefor use as a lifetime dosimeter. Howeverdepending on the half life of the mutants invivo it may be useful for evaluation ofrecent exposure. The principle of the assayhas been described in Fig. 10.
Glycophorin-A is the major cellsurface sialoglycoprotein of human
COO"
a.
o
BLOO
D
MM
MN
NN
AMINO1
Sar
S a r
L»u
Lau
ACIDS
Gly
Gly
Glu
Glu
Fig. 11 Structure of GPA, from red blood cells.(Note the variation in aminoacids at two positions in M and N blood groups)
20
erythrocyte and is present in about 5 x 105
copies per cell (Race and Sanger (1975),Anstee (1980), Langlosis et al,. (1985) andMerry et al,. (1986)). GPA occurs in twoallelic forms. The two forms M and N areidentical except for variation in aminoacidsat two positions in amino terminals(Furthmayer 1978). The amino acids inindividuals with blood group M have serineand a glycine at position 1 and 5, while
of the pure variant cells. The occurrence ofthe variant cell frequency is 1 x 10'5 innormal donors. Langlois et al, (1986). haveshown a significant dose-related increase inmutational frequency. Similar increase invariant cell frequencies are seen in cancerpatients after exposure to chemotherapyLanglosis et al, (1986). This method isapplicable only in 50% of populationcarrying the heterozygous allele of M and
MN BLOOD GROUPWITH 0 MUTATION
S-SERINEG-GLYCINE
L-LEUCINEE-CLUTAMIC ACID
0-NULL MUTATIONMM,MM,MN-SLOOD CROUPS
Fig. 12 Erythrocytes with homozygous and heterozygous allelefor GPA-A
those in group N individuals are leucineand glutamic acid, respectively (Fig. 11).The individuals with MN blood group, theglycophorin-A for M and N aileles areexpressed codominantly. Using mutationalassay, erythrocyte lacking expression of anallelic form of glycophorin-A can beenumerated (Fig. 12) (Bigbee et al, 1990).The loss of expression on erythrocytes forglycophorin-A is presumably due to in vivosomatic mutational events in erythroidprecursor cells (Awa et al, 1978, Bucktonet al, 1978, Langlosis et al, 1986 andBigbee et al, 1990). Monoclonal antibodiesspecific for M and N glycophorin-A allelesare used to identify variant cells that lackthe expression of one allele in bloodsamples from MN heterozygotes. Flowcytometric sorting enables the enumeration
N as shown in Fig. 12. As the damage inthe gene is on the precursor cell which iscontinuously replenishing RBC's carryingthese mutants, these methods can be usedto detect cumulative exposure inoccupational radiation workers.
3.10.1.1.6 Beta globin (p-globin)mutational assay
p-globin method detects rare redblood cells that contain significantquantities of a pre-selected specific mutanthemoglobin. It was originally designed forsickle cell hemoglobin, using fluorescentantibodies that can identify such cellsmicroscopically (Papayannopoulou et al,1976). The detection of such alteredhemoglobin has recently been refined byusing automated scanning microscopy and
21
Bata globin — A (8*)
1 1p
Normal Call H Polyclonad Absenca of
Slckla call (8')
antibody B' fluorasownc*
Dataclionf lowcytometry
Mutotad Callrll Polyclonad Prasenca of
antibody B* (luormctnci
Fig. 13 Normal and mutated gene sequence of $-globin
image analysis of a large number of RBGs(Tates et al, 1989). This method employsthe treatment of RBCs fixed on glass slidewith fluorescent cells and defines thehemoglobin variant frequency. Because theinvolved imitations result from singlespecific base changes (the smallest possibletarget in the genome), the mutants areexceedingly rare, on the order of the threemutants per one hundred million cells. Fig.13 shows the normal and mutated genesequence of P-globin. This techniqueneeds further validation and availabilitybefore it can be applied for biodosimetry.The sensitivity of the technique may beincreased by the simultaneous use ofseveral antibodies specific for other mutanthemoglobins.
3.10.1.2 Germinal cell mutationThis mutation is only one of the
kind so far used in the estimation of thedose using germinal cell expression. Thishas a disadvantage compared to thesomatic mutation expression that it wouldbe studied only in the male population whoare exposed to radiation.
cell mutation assay was employed byMendelsohn (1984) for biologicaldosimetry. This method is still in the initialstage of development, and the literaturepertaining to this is found to be meagre.
3.10.2 Unscheduled DNA synthesis as amolecular indicator
The biological effects of low levelradiation are not clearly understood. Theeffects such as chromosomal aberrationsare perceptible only at levels above 50mGy. There is considerable interest inidentifying possible physico-chemicaleffects at lower exposure levels forapplication for dosimetry purposes. It hasbeen shown that persons receiving lowlevels of ionising radiation show enhancedrate of DNA repair of their lymphocytes.This has been studied using UnscheduledDNA synthesis (UDS). The intrinsiccapacity to repair UV induced thyminedimer formation has been calibrated andquantified in vitro by Kang (1988). Tomeasure UDS, Tritiated thymidine(3HTdR) is used as an indicator. 3HTdRgets incorporated into the newlysynthesised DNA during repair process.The extent of incorporation of 3HTdR intoDNA depends on the repair capacity of thecell. 3HTdR is incorporated in DNA also
22
during semiconservative DNA synthesis.But the addition of hydroxyurea in theculture medium inhibits semiconservativeDNA synthesis, and helps in discriminatingthe incorporated 3HTdR in newlysynthesised DNA due to UDS. Thismethod of UDS has been found to be asuitable method in the estimation of lowradiation dose. Various methods areavailable for the measurement of UDS likeautoradiography, liquid scintillationcounting, BND (benzoylated naphthoylatedDEAE) cellulose method, equilibriumsedimentation technique and fluorescencetechnique has been reviewed by Mary andJeevanram (1995) in depth.
4.0 ConclusionFrom the review of various
biological indicators for measuringradiation dose, it is that no single biologicalindicator can satisfactorily meet all therequirements explained in section 2.0.Prodromal indicators are evident in highdose exposure as in Goiania accidentwhere people showed radiation sickness.These cannot be used to detect low levelsof radiation. Hematological indicators arealso mainly used in accidental exposuresand are applicable for doses of the order of0.5 Gy and above. Spermatogenesisindicators are not suitable for long termbiodosimetry as the restoration of fertilityreaches normalcy very fast. The changes inthe levels of biochemicals in cells due toradiation too have limited application asthere are problems of establishing a cleardose effect relationship. Prodromal andcellular indicators lack sensitivity andreprodudbility. Physiological changes arerestored quite fast. Immume responsevaries from person to person and hencethese indicators are not ideal for dosimetry.The review clearly indicates thatcytogenetic studies are the ones which canbe relied on the estimation of radiationabsorbed dose. Currently, FISH assayappears to be more promising to evaluatethe dose received during a long period oftime. It is yet not within reach of all
laboratories. Mutational assays may besensitive but refined methods to identifyvarious mutational genes need to bedeveloped. Except for GPA mutationalassay, all the other assays present particulardifficulties in studies of the effect of doserate when exposures can be spread overvarying period of time. GPA is confined tothe estimate of dose to 50% of theheterozygote population. The ESR studyof free radicals is quite stable and sensitiveand can estimate the cumulative dose. Ingeneral no biodosimetry technique canfulfil all the criteria for the ideal dosimeter.One has to resort to combination of severalbiological indicators to achieve bestsensitive estimate of doses.
5.0 AcknowledgementWe wish to express our sincere
thanks to Shri L.V. Krishnan, Director,Safety Research and Health Physics. Groupand to Shri A.R. Sundararajan, HeadHealth and Safety Division for theencouragement given during the course ofthe study. We would also like to thankDr. P. Rodriguez Director IGCAR, forestablising the biodosimetry group atIGCAR. The authors would like to extendtheir thanks to Mr. R. Mathiyarasu forbringing out an excellent piece of desk toppublication.
6.0 ReferencesAghamohammadi, S.Z.; D.T. Goodhead
and J.R.K. Savage (1988), Induction ofsister chromatid exchanges (SCE) in Golymphocytes by plutonium-238 alpha-particles. InL J. Radiat. Biol 53 : 909-915
Aghamohammadi, S.Z.; D.T. Goodheadand J.R.K. Savage (1989), Productionof chromosome aberrations, micronucleiand sister-chromatid. exchanges by 24-KeV epithermal neutrons in human Golymphocytes. Mutat. Res. Ill: 225-230
Ahlers, F.J. and C.C.J. Schneider (1991),Alanine ESR dosimetry: An assessmentof peak-to-peak evaluation. Radiat.Prot.Dosi.2n. 117-122
23
Akiyama, M.; Y.Kusunoki; S. Umeki; Y.Hirai; N. Nakamura and S. Kyoizumi(1991), Evaluation of four somaticmutation assays as biological dosimeterin humans. In: Radiation Research-A2$h century perspective, Volume-ILCongress Proceeding of the &n
International Congress on RadiationResearch, Editors W.C. Dewey; M.Edington; R.J.M. Fry, AcademicPress, pp. 177-182
Al-Achkar, W. (1987), Development of adouble method for the detection of sisterchromatid exchanges on the "R"banding pattern application and thestudy of radio-induced chromosomeaberrations in human lymphocytes.Aalam-Al-Zarra 4 : 67-68
Al-Achkar, W. (1989), Chromosomalaberrations induced by low dose gammairradiation study of R-bandedchromosomes of human lymphocytes.Muiat Res. 212 : 167-172
Albertini, R.J.; J.A. Nicklas; J.P. O'Neilland S.H. Robison (1990), In vivosomatic mutations in humans:Measurement and analysis. In: AnnualReview of Genetics, Editors A.Campbell; B.S. Baker; E.W. Jones,Annual Review Inc, Palo Alto,California, 24:305-326
Aldrich, J.E. and B. Pass (1986), Dentalenamel as an in vivo radiationdosimeter: separation of the diagnosticX-ray dose from the dose due to naturalsource. Radial. Prot. Dost 17 : 175-179
Ammenheuser, M.M.; J.B. Jr. Ward; E.B.Jr. Whorton; J.M. Killian; M.S. Legator(1988), Elevated frequencies of 6-thio-guanine-resistant lymphocytes inmultiple sclerosis patients treated withcyclophosphamide: A prospective study.Mutat. Res. 204 : 509-520
Anatoly, M. (1989), The problem of long'term fading of absorbed palaeodose onESR dating of quaternary mollusc shells.Appl. Radiat, Isotopes 40 : 1087-1093
Anstee, DJ. (1980), Blood group MNSs-active silaoglycoproteins of the human
erythrocyte membrane. In:Immunobiology of the erythrocyte,Editors S.G. Sandier; J. Neesbaciherandj. Schanfield, Lh;s, New York, pp67-98
Ashby, J.; H. Tinwell; P.A. Lefevere; andM.A Browne (1995), The single cellgell electrophoresis assay for inducedDNA damage (comet assay)measurement of tail length andmovement. Mutagenesis 10 : 85 -90
Awa, A.A.; T. Sofuni; T. Hond*; M. Itoh;S. Neriishi and M. Otake (1978), Therelationship betv/een the radiaion doseand chromosome aberrations in atomicbomb survivors of Hiroshima andNagasaki. Radiat. Res. 19 : 126-140
Bauchinger, M.; E. Schmid; H.Zitzelberger; H. Braselmann and UNahrsted (1993), Radiation-inducedchromosome aberrations analysed bytwo-colour fluorescence in situhybridization with composite wholechromosome-specific DNA probes and apancentromeric DNA probe. Int. J.Radiat. Biol. 64 : 179-184
Bigbee, W.L.; A.J. Wyrobek; R.G.Langlois; R.H. Jensen and R.B. Everson(1990), The effect of chemotherapy onthe in vivo frequency of glycophorin-A"null" varient erythrocytes. Mutat. Res.240:165-175
Boegl, W.; A. Stamm; L. Heide; E. Reichand E. Stumpf (1983), Biochemicalindicators of radiation exposure.Experimental studies on the erythrocytemembrane, the activities of acidphosphatase and amylase, and DNAsynthesis. In: Proceedings of thesixteenth annual meeting of theFachverband fuer Strahlenschuty,Germany; Radiation Protection,measurements and methods, EditorsF.V. Strahlenschutze; G. Strahlen andH. Umweltforschung, Germany,pp.565-568
Boegl, W.; A. Stamm; L. Heide; N. Willichand J. Lissner (1984), The significanceof acid serum phosphates and serumamylase as biochemical indicators of
24
radiation exposure. In Radiation RiskProtection, Editors A. Kaul, R. Neider;J. Pensko; F.E. Stieve and H.Brunner, IRAP, Germany, pp.397-400
Brown, J.M.; J. Evans and M.S. Kovacs(1992), The prediction of human tumourradiosensitivity in situ : an approachusing chromosome aberrations detectedby fluorescence in situ hybridization.Int. J. Radial. Oncol 24 : 279-286
Buckton, K.E.; G.E. Hamilton; L. Patonand A.O. Langlands (1978),Chromosome aberrations in irradiatedankylosing spondylitis. In: Mutageninduced chromosome damage in man,University Press, London, pp. 142-150
Butkowskji, W.T.; A. Spiegelberg and W.Boegl (1987), Investigation of theradiation induced changes ofelectrophoretic mobility of humanerythrocytes. In: Abstract,Bundesministerium fuer Umwelt,Naturschutz and Reaktorsicherheit,Bonn, Germany, pp.61
Caspersson, T.; S. Farber; G.E. Foley; J.Kudynowsky; E.J. Modest; E.Simonsson; U. Wach and L. Zech(1968), Chemical differentiation alongmetaphase chromosomes. Exp. Cell.Res. 49: 219-222
Cember, H. (198.3), Biological effects ofradiation. In: Introduction to HealthPhysics-Second Edition, PergamonPress, pp. 177-199
Chaudhuri, J.P. and O. Messerschmidt(1982), Amount of peripheralreticulocytes as biologic dosimetry ofionizing radiation. Acta. Radiologica.Oncology Radiation Therapy Physicsand Biology 2 1 : 177-179
Court, L.; M. Fatome; P. Gourmelon andJ.C. Mestrics (1991), Contribution ofelectrophysiology to the assessment of awholebody acute or semi-chronicexposure. Radioprotection 26 : 223-230
Dalgarno, B.G. and J.D. McClymont(1989), Evaluation of ESR as a radiationaccident dosimetry technique. Appl.Radkit. Isotopes 40 : 1013-1020
Dehos, A. and G. Hinz (1984), Changes for
lymphocyte populations and their role asa biological indicator of ionizingradiation. In: Proceedings of the SixthInternational Congress of theInternational Radiation ProtectionAssociation and exibition; RadiationRisk Protection, Vol-I, Verl, TUEV,Rheinland, Germany, pp.413-415
Dehos, A.; G. Hinz and E.R. Schwarz(1986), Changes in number and functionof the lymphocyte populations as abiological indicator for ionisingradiation. In: Proceeding of theInternational workshop on biologicalindicators for radiation doseassessment; Indicators for RadiationDose Assessment, Editors A. Kaul; A.Dehos; W. Boegl; G. Hinz; F. Kossel;E.R. Schwarz; A. Stamm and G.Stephan, MMV Medizin Verl,Germany, pp.298-301
Dumphy, W.G. and J.W. Newport (1988),Unraveling of mitotic controlmechanisms. Cell 55 : 925-928
Feinendegen, L.E. (1980), Biologicalindicators for the assessment of aradiation exposure. In: Proceeding ofthe Joint Radiation ProtectionConference; Incidents in Industry andRadiation Exposure, Editors O.Messerschmidt; L.E. Feinendegen andW. Hunzinger, Germany, pp.207-223
Feinendegen, L.E. (1986), Biochemicalindicators. In: Proceeding of theInternational workshop on biologicalindicators for radiation doseassessment; Biological Indicators forRadiation Dose Assessment, Editors A.Kaul; A. Dehos; W. Boegl; G. Hinz; F.Kossel; E.R. Schwarz; A. Stamm andG. Stephan, Germany, MMV, Medizin,Verl, pp.70-84
Fenech, M. and A.A. Morley (1986),Cytokinesis block micronucleus methodin human lymphocytes: effect of in vivoaging and low dose x-irradiation. Mutat.Res. 161: 193-198
Fenech, M.; A.A. Morley and J. Denham(1988), The cytokinesis blockedmicronuclei assay: a sensitive technique
25
for measuring radiation inducedchromosome damage. In: HealthEffects of Low dose IonisingRadiation, Proceeding of InternationalConference, British National Society,BNES, London, pp.29-33
Fliender, T.M. and W. Nothdurft (1979),Blood stem cells as an indicator ofradiation exposure. In: Proceeding ofthe International meeting ofinvestigators on biological indicatorsof radiation effects in man; Biologicalindicators of radiation effects in man,Editor M. Bauchinger, Germany,pp.22
Fliender, T.M. and W. Nothdurft (1986),Cytological indicators: Haematopoeticeffects. In: Proceeding of theInternational workshop on biologicalindicators for radiation doseassessment; Biological Indicators forRadiation* Dose Assessment, Editors A.Kaul;A. Dehos; W. Boegl; G. Him; F.Kossel; E.R. Schwarz; A. Stamm andG. Stephan, MMV Median VerlGermany, pp. 123-156
Furthmayer, H. (1978), Structuralcomparison of glycophorins andimmunochemical analysis of geneticvariants. Nature 111: 519-524
Gasinska, A. and G.D. Wilson (1988),Flow cytometric evaluation of theeffects of 2.3 MeV neutrons and 240KV x-rays on mouse spermatogenic S-phase cells using bromodeoxyuridineincorporation. Brit. J. Radiol. 61 : 133-139
Gray, J.W.; J. Lucas; O. Kallioniemi; A.Kallioneimi; W.L. Kuo; T. Straume; D.Tkachuk; T. Tenjin; H.U. Weier and D.Pinkel (1991), Applications offluorescence in situ hybridization inbiological dosimetry and detection ofdiseases-specific chromosomeaberrations. In: New Horizons inBiological Dosimetry, Wiley- Liss, Inc,pp. 399-411
Greenstock, C.L. and A. Trivedi (1992),Free radical measurement in bio-organicsubstances using an electron spin
resonance technique. In : IRPA8,International Radiation ProtectionAssocation, Proceedings, 1: 455-4'58
Gu-Yuanxi and Cui-Yufang (1988), Doseestimation with biological indicators inacute radiation excident. J. Radial,. Res.and Radiation Processing 6 : 17-21
Hacker, U.; J. Schumann; W. Goebde andK. Mueller (1981), Mammalianspermatogenesis as a biologic dosimeterfor radiation. A eta. Radiologica.Oncology Radiation Therapy Physicsand Biology 20 : 279-282
Hacker, U.; J. Schumann and W. Goehde(1982), Mammalian spermatogenesis asa new system for biologic dosimetry ofionizing radiation. Acta. Radiologica.Oncology Radiation Therapy Physicsand Biology 21: 349-3551
Hakoda, M.; M. Akiyama; S. Kyoizumi;A.A. Awa; M. Yamakido and M. Otake(1988), Increased somatic cell mutantfrequency in atomic-bomb survivors.Mutat. Res. 201 : 39 48
Hennequin, C ; J.M. Cosset; P.E. Cailleux;T. Girinsky; G. Ganem; E. Comoy andJ. Dutreix (1989), Amylasemia: abiological marker for irradiationaccidents. Bulletin du Cancer 76 : 617-624
Hittelman, W.N. and P.N. Rao (1974),Premature chromosome condensation-I.Visualization of X-ray inducedchromosome damage in interphase cells.Mutat. Res. 23:251-258
Hofmann, R.; W. Hendriks; G.A. Schreiberand K.W. Boegl (1990), Blood amylasea biochemical radiation indicator? In:Abstract, Bundesant fuerstrahlenschutz, Neuherberg, Germanypp.64
Huebner, K.F.; L.G. Littlefield and R.K.Dufrain (1986), Experience in thepractical application of biologicalindicators. In: Proceeding of theInternational workshop on biologicalindicators for radiation doseassessment; Biological Indicators forRadiation Dose Assessment, Editors A.Kaul; A. Dehos; W. Boegl; G. Hinz; F.
26
Kossel; E.R. Sckwarz; A. Stamm andG. Stephan, MMV Medizin Verl,Germany, pp. 17-34
Kang, W. (1988), Estimation of DNAsynthesis in lymphocyte as biologicalindicators. IAEA /R-4535/F, pp. 1-8
Kumagai, E.; R. Tanaka; T. Kumagai; M.Onomichi and S. Sawada (1990),Effects of long-term radiation exposureon chromosomal aberrations inradiological technologist Radiat. Res.31 : 270-279
Kumagai, E.; M. Onomichi; R. Tanaka andS. Sawada (1991), Effect of long termlow dose radiation relationship betweenchromosomal translocation and thecellular oncogenes. Rinsho- Byori. 39 :639-644
Kushiro, J.; Y. Hirai; Y. Kusumoki; S.Kyoizumi; Y. Kodama; A. Wakisaka; A.Jeffreys; J.B. Cologne; K. Dohi; N.Nakamura and M. Akiyama (1992),Development of a flow-cytometricHLA-A locus mutation assay for humanperipheral blood lymphocytes. Mutat.Res. 272: 17-29
Langlois, R.G.; W.L. Bigbee and R.H.Jensen (1985), Flowcytometriccharacterization of normal and variantcell with monoclonal antibodies specificfor glycophorin-A. J. Immunol. 134 :4009-4017
Langlois, R.G.; W.L. Bigbee and R.H.Jensen (1986), Measurements of thefrequency of human erythrocytes withgene expression loss phenotypes at theglycophorin-A locus. Hum. Genet. 74 :353-362
Lucas, J.N.; T. Tenjin; T. Straume; D.Pinkel; D. Mooreii; M. Litt and J.W.Gray (1989), Rapid human chromosomeaberration analysis using fluorescencs in
situ hybridization. Int J. Radiat. Biol.56:35-44
Lucas, J.N.; A.A. Awa; T. Straume; M.Poggensee; Y. Kodama; M. Nakano; K.Ohtaki; H.U. Weier; D. Pinkel; J. Grayand G. Littlefield (1992), Rapidtranslocation frequency analysis inhuman decades after exposure toionising Radiation. Int. J. Radiat Biol.62:53-63
Lushbaugh, C ; G. Eisele; W. Burr, Jr.; K.Hubner and B. Wachholz (1991),Current status of biological indicators todetect and quantify previous exposuresto radiation. In : Assessment oftechniques to determine previousradiation exposures (Emphasizingexposures occurring decades earlier)Health Phy. Sup-l 60 : 103-109
Mal'tsev, V.N.; V.A. Strel'nikov and A.A.Ivanov (1978), C-reactive protein ofblood serum as an indicator of radiationinjury heaviness. Doklady AkademiiNauk SSSR, USSR 239 : 750- 752
Martin, C ; E. Multon and S. Martin(1991), Biological dosimetry:Biochemical and cellular parameters.Radioprotection 26 : 205-221
Mary, M and R.K. Jeevanram (1995),Biological Monitors for low levels ofionising radiation. IGC-167 pp 1-38
Matsuoka, A.; J.D. Tucker; M. Hayaski; N.Yamazabi and T. Sofuni (1994),Chromosome painting analysis of X-rayinduced aberrations in humanlymphocytes in vitro. Mutagenesis 9 :151-155
Mendelsohn, M.L. (1984), Prospects forcellular mutational assay in humanpopulations. In: Abstract, Of theSymposium on assessment of risk fromlow level exposure in radiation andchemicals, Upton, New York, USA.pp.11
Merry, A.H.; C. Hodson; E. Thompson; G.Mallinson and D.J. Anstee (1986), Theuse of monoclonal antibodies toquantitate the levels ofsilaoglycoproteins alpha, delta andvariant silaoglycoproteins in human
27
erythrocyte membrane. Siochem. J. 233: 93-98
Messerschmidt, 0 . (1985), The influence ofmodifying factors on radiation effect innuclear accident. In: Proceedings of theRadiation Protection Conference-25tn
annual conference; 25 Years ofMedical Radiation Protection,Radiation protection: History andoutlook, Editors HA. Ijadner; C.Reiners; W. Boerner and J. Schuetz,Germany, pp.31-37
Miles, C. and M. Meuth (1989), DNAsequence determination of y-radiationinduced mutations of the hamster APRTlocus. Mutat. Res. Ill: 97-102
Miller, B.M.; T. Werner; H.U. Weier andM. Nusse (1992), Analysis of radiation-induced micronuclei by fluorescence insitu hybridization (FISH) simultaneouslyusing telomeric and centromeric DNAprobes. Radiat. Res. 131: 177-185
Miller, B.M. and M. Nusse (1993),Analysis of micronuclei induced by 2-chlorobenzylidene molonitrile (CS)using fluorescence in situ hybridisationwith telomeric and centromeris DNAprobes, and flow cytometry.Mutagenesis 8: 35-41
Morley, A.A.; K.J. Trainor; R. Seshadri;R.B. Ryall (1983), Measurement of invivo mutation in human lymphocytes.Nature 302: 155-156
Muller, W.U. and C. Streffer (1991),Biological indicators for radiationdamage. Int. J. Radiat. Bio I. 59 : 863-873
Natarajan, A.T.; R.C. Vyas; F. Durroudiand S. Vermeulen (1992), Frequenciesof X-ray induced chromosometranslocations in human peripherallymphocytes as detected by in situhybridization using chromosome-specific DNA libraries. Int. J. Radiat.Biol. 61 : 199-203
Nocentini, S. (1995), Comet assay analysisof repair of DNA strand breaks innormal and deficient human cellsexposed to radiation and chemicals.Evidence for a repair pathway specificity
of DNA ligation. Radiat. Res. 144 :170-180
Ohyama, H. (1989), Radiation inducedbiochemical changes in blood and urine:A Review In: Proceedings of theNational Institute of RadiologicalScience, (NIRS) symposium; ExposedDose Evaluation and Medical Care ofRadiation Accident, Editor N. Isamu,Chiba, Japan, pp. 161 -169
Old, R.W. and S.B. Primrose (1992),Introducing genes into animal cells. In:Principles of Gene Manipulation, AnIntroduction to Genetic Engineering.Editors R.W. Old and S.B. Primrose,Blackwell Scientific Publications, pp.225-295
Onori, S.; A. Bartolotta; B. Caccia; P.L.Indovina; F. Milano; R. Renzi; G.Scarpa; C. Caporali and M. Moscati(1990), Dosimetric characteristics ofalanine-based ESR detectors in electronbeams used in radiotherapy. Radiat.Prot. Dosi. 34 : 287-289
Osanov, D.P.; V.P. Panova and A.T. Shaks(1985), Multilayer detector for skinabsorbed dose measuring, Pribory-i-Tekhnika Ehksperimenta, USSR No: 3pp.86-89
Pantelias, G.E. and H.D. Maillie (1984),The use of peripheral bloodmononuclear cell pematurely condensedchromosomes for biological dosimetry.Radiat. Res. 99 : 140-150
Papayannopoulou, Th.; R.C. McGuire; G.Lim; E. Grazel and G.Stamatoyannopoulos (1976),Identification of hemoglobin S in redcells and normoblasts using fluorescentanti-Hbs antibodies. Brit. J. Haematol.34:25-31
Paul, S.F.D, P. Venkatachalam andR.K. Jeevanram (1996), Chromosomeaberration analysis for biologicaldosimetry - A Review. IGC-173
Paul, S.F.D; P. Venkatachalam and R.K.Jeevanram (1996), Micronuclei analysisfor biological dosimetry - A Review.IGCAR Report (under preparation)
Perry, P.E. and H.J. Evans (1975),28
Cytogenetic detection of mutagen-carcinogen exposure by sister chromatidexchange. Nature 258 : 121-125
Popp, S. and T. Cremer (1992),Development of a biological dosimeterfor translocation scoring based on two-color fluorescence in situ hybridizationof chromosome subset. / . Radiat. Res.Sup 3 3 : 61-70
Potten, C.S. (1985), Cell death (apoptosis)in hair follicles and consequent changesin the width of hair after irradiation ofgrowing follicles. Int. J. Radiat. Biol.Relat. Stud. Phys. Chem. Med. 48 :349-360
Prosser, J.S.; J.E. Moquet; D.C. Lloyd andA.A. Edwards (1988), Radiationinduction of micronuclei in humanlymphocytes Mutat. Res. 199: 37-45
Prosser, J.S.; D.C. Lloyd and A.A.Edwards (1989), A comparison ofchromosomal and micronuclear methodsfor radiation accident dosimetry.Radiation Protection-NRPB pp. 133-136
Race, R.R. and R. Sanger (1975)", Bloodgroup in man, Sixth edition, Blackwell,Oxford, pp.92-138
Rainer, G. (1989), Present status on ESRdating. Applied Radiation and Isotopes40: 1045-1055
Rao, B.S. (1992), Clinical and BiologicalDosimetry. In: INDRP, BARC,Trombay, India, pp. 1-32
Sabatier, L.; B. Dutrillaux and W. Al-Achkar (1988), Transmission ofradiation induced rearrangementsthrough cell divisions. Mutat. Res. 198 :191-198
Savage, J.R.K. and M. Holloway (1987),Induction of SCE by d(42MeV)-Beneutrons in unstimulated human bloodlymphocytes. Int. J. Radiat. Biol. 52 :483
Savage, J.R.K. and M. Holloway (1988),Induction of SCE by d(42 MeV)-Beneutrons in unstimulated human bloodlymphocytes. Brit. J. Radiol 61 :231-234
Sax, K. (1938), Chromosome aberrations
induced by X-Rays. Genetics 23 : 494-516
Schmid, E.; H. Zitzelsberger; H.Braselmann; J.W. Gray and M.Bauchinger (1992), Radiation-inducedchromosome aberrations analysed byfluorescence in situ hybridization with atriple combination of composite wholechromosome-specific DNA probes. Int.J. Radiat. Biol. 62 : 673-678
Schreiber, G.A.; A. Spiegelberg; R.Hofmann; K.W. Boegl and N. Wfflich(1989), Changes in oligosaccharide ofhuman blood cells measured by tritiumand FITC labeled lectin after in vitroand in vivo irradiation lectin binding as abiological dosimeter. Int. J. Radiat.Biol. 55:843-859
Shuqing, S.; W. Guoyin and X. Yi (1988),Comparison of stable and unstablechromosome aberrations induced by x-rays in human lymphocytes. ChineseJournal of Radiological Medicine andProtection 8:403-406
Stockhausen, K. (1976), ESR studies onthe effects of ionizing radiation inbiological media. In: Proceeding of the15tn annual meeting of theVereinigung Deutscher Strahlen-schutzaerzte, Karlsruhe; Theauthorised physician as stipulated inradiation protection legislation.Special contributions of medicine toradiation and environmentalprotection, Editors H. Broun; H.A.Ladner; J. Mehl; O. Messerschmidt;G. Moehrle and F.E. Stieve, Germany,pp.99-119
Straume, T.; J.N. Lucas; J.D. Tucker; W.L.Bigbee and R.G. Langlois (1992),Biodosimetry for a radiation workerusing multiple: assays. Health Phy. 63 :421-426
Tates, A.D.; L.F. Bernini; A.T. Natarajan;J.S. Ploem; N.P. Verwoerd; J. Cole;M.H.L. Green; C.F. Arlett and P.N.Norris (1989), Detection of somatic
29
mutants in man : HPRT mutations inlymphocytes and hemoglobin mutationsin erythrocytes. Mutat. Res. 213 : 73-82
Tatsumi, K.; A. Fujimori; A. Tachibana andY. Houki (1992), Concurrent analysesof mutagenesis by ionizing radiation atthe APRT and HPRT loci in humanlymphoblastoid cells. In : Low DoseIrradiation and Biological DefenseMechanisms, Proceedings of theInternational Conference on Low DoseIrradiation and Biological DefenseMechanisms, Kyoto, Japan, Editors,T.Sugahara; LA. Sagan and T.Aoyatna, Elsevier Science Publishers,Netherlands, pp. 3 31 -3 34
Technical Report (1986), Biologicaldosimetry, Chromosomal aberrationanalysis for dose assesment. In:Technical Report Series No: 260,IAEA, Vienna, pp. 1-69
Twan, E.J. and D. Holdsworth (1992),Mutagen induced chromosome damagein human lymphocytes. In : Humancytogenetics - A practical approach,Vol-II, Malignancy and acquiredabnormalities, Second edition, EditorsD.E. Rooney and B.H. CzepulkowsM,1RL Press, Oxford, pp. 189-208
Thierens, H.; A. Vral and L. de, Riddcr
(1991), Biological dosiroeuy using themicronucleus assay for lymphocytes:interindividual differences in doseresponse. HerJth Phy. 6 1 : 623-630
Venkatachalam, P, 'S.F.D. Paul and R.K.Jeevanram (1996), Fluorescence in situhybridisation (V-ISH) w i t h reference tobiological do< ,imetry-A Review. IGC-174
Vladimirov, V.G. and A.D. Srnirnov(1978), 'Siological dosimetry atexposure ' ,o ionising radiation. Voenno-Medtisinskij-Zhurnal-USSR 2:76-79
Waldren, C.A. and R.T. Johnson (1974),Analysis of interphase chromosomedamage by means of prematurechromosome condensation after X andultraviolet irradiation. Proc. Natl. Acad.Sci.ll: 1137-1141