Individual variability in esterase activity and CYP1A levels inChinook salmon (Oncorhynchus tshawytscha) exposed to
esfenvalerate and chlorpyrifos
Craig E. Wheelocka, Kai J. Ederb,c, Inge Wernerb, Huazhang Huanga, Paul D. Jonesa,Benjamin F. Brammelld, Adria A. Elskusd,1, Bruce D. Hammocka,∗
a Department of Entomology and Cancer Research Center, University of California at Davis, Davis, CA 95616, USAb Aquatic Toxicology Program, Department of Anatomy, Physiology and Cell Biology, School of Veterinary Medicine,
University of California at Davis, Davis, CA 95616, USAc Animal Physiological Ecology, Zoological Institute, University of T¨ubingen, Konrad-Adenauer-Str. 20,
D-72072 Tubingen, Germanyd Department of Biology, University of Kentucky, Lexington, KY 40506, USA
Received 1 September 2004; received in revised form 9 May 2005; accepted 11 May 2005
Abstract
Acetylcholinesterase (AChE) activity has traditionally been monitored as a biomarker of organophosphate (OP) and/or car-bamate exposure. However, AChE activity may not be the most sensitive endpoint for these agrochemicals, because OPs cancause adverse physiological effects at concentrations that do not affect AChE activity. Carboxylesterases are a related family ofenzymes that have higher affinity than AChE for some OPs and carbamates and may be more sensitive indicators of environ-mental exposure to these pesticides. In this study, carboxylesterase and AChE activity, cytochrome P4501A (CYP1A) proteinla re acutelyt rifosa sue( e activityw ), whilee ot affectA n by OPs.C ct. Threet -i etectable.
0
evels, and mortality were measured in individual juvenile Chinook salmon (Oncorhynchus tshawytscha) following exposure ton OP (chlorpyrifos) and a pyrethroid (esfenvalerate). As expected, high doses of chlorpyrifos and esfenvalerate we
oxic, with nominal concentrations (100 and 1�g/l, respectively) causing 100% mortality within 96 h. Exposure to chlorpyt a high dose (7.3�g/l), but not a low dose (1.2�g/l), significantly inhibited AChE activity in both brain and muscle tis85% and 92% inhibition, respectively), while esfenvalerate exposure had no effect. In contrast, liver carboxylesterasas significantly inhibited at both the low and high chlorpyrifos dose exposure (56% and 79% inhibition, respectivelysfenvalerate exposure still had little effect. The inhibition of carboxylesterase activity at levels of chlorpyrifos that did nChE activity suggests that some salmon carboxylesterase isozymes may be more sensitive than AChE to inhibitioYP1A protein levels were∼30% suppressed by chlorpyrifos exposure at the high dose, but esfenvalerate had no effe
eleost species, Chinook salmon, medaka (Oryzias latipes) and Sacramento splittail (Pogonichthys macrolepidotus), were examned for their ability to hydrolyze a series of pyrethroid surrogate substrates and in all cases hydrolysis activity was und
Agrochemical usage practices are currently shift-ing, with a general movement away from organophos-phates (OPs) towards pyrethroid pesticides (Casida andQuistad, 1998). The ecological implications of thislarge-scale shift in pesticide application are unknown.Pyrethroids generally have low mammalian toxicity(Abernathy and Casida, 1973; Casida et al., 1983;Casida and Quistad, 1995), especially compared tomany OP pesticides. However, there have been sev-eral reports regarding the sensitivity of aquatic inverte-brates and some fish species to pyrethroids (Bradburyand Coats, 1989a; Werner et al., 2002; Denton et al.,2003). There is subsequently concern that the ecolog-ical consequences of increased pyrethroid applicationon aquatic ecosystems could be far-reaching.
Carboxylesterases are a class of enzymes thathydrolyze ester-containing compounds to the cor-responding alcohol and acid (hydrolysis products)(Satoh and Hosokawa, 1998; Wheelock et al., 2005).These enzymes are important in the metabolism andsubsequent detoxification of many xenobiotic andendogenous compounds, including pyrethroids andOPs (Fig. 1). Carboxylesterases reduce pyrethroid tox-icity by hydrolyzing these compounds to less toxicmetabolites (Abernathy and Casida, 1973; Wheelocket al., 2004). OPs are generally not esterase sub-strates; however they bind stoichiometrically to bothcarboxylesterases and acetylcholinesterases (AChE)( tad,2 nityo stedt thusp1 rvea Ps( -o rgistic
toxicity through OP-induced esterase inhibition render-ing the enzyme unable to hydrolyze and thus detoxifypyrethroids (Gaughan et al., 1980; Denton et al., 2003).
Mixed-function oxidases (MFOs), including thecytochrome P450 monooxygenases (CYP), comprisea superfamily of enzymes that can interact with bothOPs and pyrethroids (Casida and Quistad, 1995; Poetet al., 2003). CYP-mediated OP metabolism convertsthe thione (PS) to the corresponding oxon (PO)(Fukuto, 1990), which is the form that inhibits a numberof enzymes including AChE (Fulton and Key, 2001),carboxylesterase (Maxwell, 1992; Casida and Quistad,2004) and the CYPs themselves (Tang et al., 2002).For example, CYP-mediated desulfuration of chlor-pyrifos in rats produces chlorpyrifos-oxon (Fukuto,1990), in the process releasing the sulfur ion whichcan then suppress CYP activity through binding to theheme group (Tang et al., 2002). OPs also suppress CYPactivity in fish, including the widely used pollutantbiomarker enzyme, CYP1A (Flammarion et al., 1998).In contrast to OPs, in which CYP-mediated metabolismproduces toxic as well as nontoxic metabolites, CYPmetabolism of pyrethroids is exclusively a detoxifica-tion process (Casida and Quistad, 1995). Elevated CYPactivity is an important mechanism for insect resistanceto pyrethroids, including esfenvalerate (Scott, 1999).Unlike OPs, which have been reported to suppressCYPs, pyrethroids such as esfenvalerate can have vari-able effects, altering some CYP isoforms (Barry et al.,1995), but not others (Barry et al., 1995; Heder et al.,2
roidt tt rtlyr -i cines upone rathert k-
Sogorb and Vilanova, 2002; Casida and Quis004). Carboxylesterases have an increased affiver AChE for some OPs and it has been suggehat carboxylesterases act as a “sink” for OPs,rotecting the organism against OP toxicity (Maxwell,992). Carboxylesterase activity can therefore ses a detoxification route for both pyrethroids and OSogorb and Vilanova, 2002). Conversely, simultaneus exposure to OPs and pyrethroids causes syne
001).Some species of fish are very sensitive to pyreth
oxicity (Bradbury and Coats, 1989b); and it is thoughhat slow metabolism of the parent compound is paesponsible (Denton et al., 2003). However, few studes have examined pyrethroid metabolism in pispecies and these reports have generally reliedsterase measurements made on tissue pools,
han individual fish (Glickman and Lech, 1981; Glic
Fig. 1. Esterase detoxification mechanisms. (A) The pyrethroid esfenvalerate is hydrolyzed by esterase to the corresponding acid and alcohol.This process reduces the toxicity of the pyrethroid. (B) The organophosphate chlorpyrifos is converted by mixed-function oxidases (MFO) tothe active oxon form, which in turn inhibits esterase. After inhibition, the enzyme can go through two different pathways; regeneration, wherethe enzyme regains catalytic activity, or aging, in which catalytic activity is lost.
man et al., 1982). While useful, these data only pro-vide information on the average enzyme activity in apopulation or species and do not indicate the activityrange amongst individuals. Data derived from mea-surements in individuals are important to determineif some individuals metabolize pesticides slower thanothers, which could potentially correlate with increasedsensitivity to pyrethroid or OP exposure. To evalu-ate inter-individual variations in carboxylesterase and
AChE activity, as well as CYP1A protein levels, weevaluated these enzymes in tissue homogenates fromindividual fish exposed to esfenvalerate or chlorpyrifos.We further investigated esterase activity by measuringpyrethroid hydrolysis in three fish species to compareinter-species variability in activity and to evaluate therelationship between esterase activity level and pes-ticide toxicity. We hypothesized that fish with lowerlevels of esterase activity would be more sensitive to
pyrethroid and OP toxicity. Results from this work willbe valuable in understanding the mechanism of toxic-ity of two major classes of agrochemicals on multiplefish species. Additionally, these data will be useful forinterpreting the impact of increased pyrethroid usageupon aquatic ecosystems.
2. Materials and methods
2.1. Chemicals and equipment
All chemicals and reagents were purchased fromSigma–Aldrich Chemical Co. (St. Louis, MO), Invitro-gen (Carlsbad, CA), or Fisher Scientific (Pittsburg, PA)unless otherwise stated. Pesticide standards were pur-chased from Chem Service, Inc. (West Chester, PA).Porcine esterase was purchased from Sigma ChemicalCo. (catalog no. E-2884, lot no. 102K7062, 184 U/ml,10 mg/ml). The following esterase substrates werepreviously synthesized in our laboratory (Shan andHammock, 2001; Wheelock et al., 2003; Stok et al.,2004): �-cyano(6-methoxy-2-naphthyl)methyl acetate(see Fig. 2), �-cyano(6-methoxy-2-naphthyl)methylbutyrate (seeFig. 2), (R/S)-�-cyano(6-methoxy-2-naphthyl)methyl-(S)-(+)-2-(4-chlorophenyl)-3-methylbutanoate (�R/S)(2S)—compound1, (R/S)-�-cyano(6-methoxy-2-naphthyl)methyl-(R)-(−)-2-(4-chlorop-henyl)-3-methyl butanoate (�R/S)(2S)—compound2,(R/S) - � - cyano(6-methoxy-2-naphthyl)methyl-(R/S)-
cis/trans-3-(2,2-dichlorovinyl)-2,2-dimethylcyclopro-pane carboxylate—compound3, (R/S)-�–cyano(6-methoxy-2-naphthyl)methyl (cis/trans)-3-(2,2-dibro-movinyl)-2,2-dimethylcyclopropane carboxylate—compound4, (R/S)-�-cyano-(6-methoxy-2-naphthyl)methyl (trans)-3-(2-chloro-2-trifluoromethyl vinyl)-2,2-dimethylcyclopropane carboxylate—compound5,(R/S)-�-cyano-(6-methoxy-2-naphthyl)methyl (cis)-3-(2-chloro-2-trifluoromethyl vinyl)-2,2-dimethyl-cyclopropane carboxylate—compound6, (R/S)-�-cyano-(6-methoxy-2-naphthyl)methyl (cis/trans)-3-(2,2-dimethylvinyl)-2,2-dimethylcyclopropane carbo-xylate—compound 7, (R/S)-�-cyan(6-methoxy-2-naphthyl)methyl 2,2,3,3-tetramethylcyclopropanecarboxylate—compound8. The monoclonal antibodymade against scup CYP1A protein, MAb 1-12-3, wasa generous gift of Dr. John Stegeman (Woods HoleOceanographic Institution).
2.2. Study species
Four- to five-month-old Chinook salmon (Onco-rhynchus tshawytscha) were supplied by the NimbusSalmon and Steelhead Hatchery (California Depart-ment of Fish and Game, Rancho Cordova, CA) andmaintained at the Center for Aquatic Biology andAquaculture at the University of California Davisaccording to University of California Davis animaluse protocols. Fish were acclimatized under flow-through conditions (15 l/min) for 14 days in a covered,
F steras -n thyl)me rethroids to prod aneouslyr . The o e( produc sis.
ig. 2. Hydrolysis mechanism for the fluorescent carboxyleaphthyl)methyl acetate (acetate) and�-cyano(6-methoxy-2-naphurrogates are shown inTable 6. The�-cyano ester is hydrolyzedearranges to the fluorescent aldehyde at neutral and basic pHPNPA), which has an acetate moiety coupled top-nitrophenol and
e substrates used in this study. The substrates�-cyano(6-methoxy-2thyl butyrate (butyrate) are shown here, while the remaining pyuce the corresponding acid and the cyanohydrin, which spontther carboxylesterase substrate used in this study wasp-nitrophenyl acetates the yellowp-nitrophenolate anion and acetic acid upon hydroly
4 ft tall circular FRP-tank (fiber reinforced plastic)with degassed, reoxygenated well water adjusted to15.2± 0.2◦C. Fish were fed a daily ration of soft-moistsalmon diet (3/32 in. pellet, Rangen Inc., Buhl, ID).
Medaka (Oryzias latipes) and Sacramento split-tail (Pogonichthysmacrolepidotus) were obtained fromthe University of California Davis Aquatic Toxi-cology Laboratory and were approximately 10 and12 months old at the time of sacrifice, respec-tively. Fish were reared in reconstituted water pre-pared according to U.S. EPA guidelines (U.S. EPA,1985). Fish were housed in a partially closed recir-culating plexiglass aquarium system equipped withwater pump, flow meter, biologic sand filter, par-ticle filter, activated charcoal filter, and ultravioletlight sterilizer. Water in the recirculating system wasmaintained at 80–100 mg/l CaCO3 (hardness), pH7.5± 2, dissolved O2 7.0± 1.0 mg/l, electrical conduc-tivity 300–400 mmho/cm, alkalinity 30–50 mg/l and25.0± 2.0◦C. Ammonia, nitrate and nitrite were keptbelow detectable levels by changing charcoal filtersweekly and replacing 20% of system water three timesa week. Fish were fed a purified casein-based diet(DeKoven et al., 1992).
2.3. Experimental design
Chlorpyrifos (99.5%) and esfenvalerate (AsanaTM,98%) stock solutions were prepared in HPLC grademethanol and used immediately. Nominal treatmentl intoi plesw afters forew rvedo rfi on,C d thes ctioncV ered ughe ncer with9 erea etry( .) att ra-
tory (Sacramento, CA) according to published methods(Crepeau et al., 2000).
Juvenile Chinook salmon were exposed to a rangeof concentrations of chlorpyrifos (1.0, 10 and 100�g/l)and esfenvalerate (0.01, 0.1 and 1�g/l) for 96 h.The light:dark ratio was 16 h:8 h. Solvent control fishreceived the highest concentration of methanol used(50�l MeOH/l, 0.005% final concentration). Fish (10per treatment) were exposed individually in 4.0 l, clearsoda-lime flint glass containers (Wheaton “800” Redi-Pak* Standard Wide Mouths, Fisher Scientific), eachfilled with 2 l of test solution. All exposure containerswere aerated (∼60 bubbles/min; Tetra DW96-2 TetraT-ech Air Pump, Tetra GmbH; Melle, Germany) and 75%of the test solution was replaced every 24 h. Water tem-perature was maintained at 14.8± 0.5◦C and waterquality was monitored daily for pH (8.4± 0.2), dis-solved oxygen (9.1± 0.8 mg/l) and electric conduc-tivity (680± 50�S/cm). Free ammonia was evaluatedcolorimetrically from three containers each day of theexposure using commercially available kits (EM Sci-ence, Gibbstown, NJ). Fish were not fed on the daypreceding initiation of the experiment or during thelength of the exposure. At the end of the 96 h expo-sure period, surviving fish were sacrificed by decapita-tion, dissected and tissue samples flash-frozen in liquidnitrogen and stored at−80◦C. Ten randomly sampledfish from the initial flow-through system were sacri-ficed and dissected the day before experiment initiationto serve as an unexposed control group.
evels were prepared by diluting stock solutionsndividual exposure containers. Two water samere taken from each treatment, one immediatelypiking the exposure container (0 h) and one just beater renewal (24 h). Water samples were presen ice and filtered through baked 0.7�m glass fibelters within 24 h (Advantec MFS, Inc., PleasantA). Terbuthylazine was added as a surrogate anamples were extracted using C8 solid-phase extraartridges (Varian Bond-Elut, 500 mg, 300 cm3 barrel;arian Inc., Walnut Creek, CA). The cartridges wried using a syringe to repeatedly force air throach cartridge and stored frozen until analysis. Oemoved from storage each cartridge was elutedml of ethyl acetate and levels of chlorpyrifos wnalyzed by gas chromatography/mass spectromSaturn 2000 GC/MS ion trap system, Varian, Inche U.S. Geological Survey California District Labo
.4. Acetylcholinesterase assays
Salmon brains were removed entirely, whereas mle samples consisted of one piece of epaxial wuscle taken from behind the head. Each sampleeighed, diluted 1:10 (mg:�l) in 0.1 M sodium phoshate buffer (pH 8.0) with 0.5% Triton X-100. Tissuere homogenized for 1 min using a glass dounce
ce. Homogenates were centrifuged at 4◦C for 10 mint 7000×g to remove large particulate material. Tupernatant fraction was transferred to a separatend the total protein concentration was determith the Biorad DC Protein Assay (Bio-Rad Lab
atories, Hercules, CA) using methods ofLowry etl. (1951). For the AChE assay, 0.1 M sodium phhate buffer (pH 8.0) with 0.5% Triton X-100 wdded to the supernatant fractions to produce final
tions of 1:500 (mg:�l) for muscle samples and 1:200(mg:�l) for brain samples. Assay optimization was per-formed with brain and muscle tissue from unexposedjuvenile Chinook salmon. Acetylthiocholine iodide(AtChI) concentrations between 0.1 and 5 mM weretested for optimal substrate concentration, and sam-ples were incubated with tetraisopropylpyrophospho-ramide (iso-OMPA, a selective AChE inhibitor) to mea-sure butyrylcholinesterase-mediated substrate hydrol-ysis. Results showed negligible butyrylcholinesteraseactivity in muscle tissue, therefore subsequent assayswere performed without the AChE inhibitor.
AChE activity in brain and muscle was analyzedusing modified methods ofEllman et al. (1961).AChE activity for each sample was determined byadding 30�l of diluted supernatant to a microplatewell (Costar 96 well EIA/RIA Plate; Corning Inc.,New York, NY) containing 250�l of 0.1 M sodiumphosphate buffer (pH 8.0), 10�l of 5,5′-dithiobis-2-nitrobenzoic acid (DTNB, 10.3 mM), and 30�l ofAtChI (21.4 mM). Final assay concentrations were0.32 mM DTNB and 2 mM AtChI. Final protein con-centrations ranged from 10.8 to 17.1�g/�l for muscleand 7.0 to 10.7�g/�l for brain. All assays were per-formed in triplicate. Absorbance at 412 nm was mea-sured at 2 min intervals for 10 min at 25◦C with anautomated microplate reader (Model EL3401; Bio-TekInstruments, Winooski, VT) and all samples were cor-rected for background hydrolysis. AChE activity wascalculated as�mol/min/g wet weight, and then nor-m ate(
2
ribeda ing5f la hos-pmL -f tes( ona ices,Sc -
ration. All assays were designed such that no more than10% of the substrate was hydrolyzed over the length ofthe assay and solvent content never exceeded 1% of thetotal assay volume. Reported results are all correctedfor background hydrolysis of the substrate. Activitywas monitored using a 2.0 min kinetic read at 405 nm.The amount of protein added in each assay varied byspecies and sample, but ranged from 29 to 158�g/wellfor the salmon liver, 10�g/well for splittail, 25�g/wellfor medaka and 4.4�g/well for porcine esterase. Pro-tein concentration was adjusted such that the assay waslinear over the reported time interval.
All assays with�-cyanoester substrates were per-formed as described inWheelock et al. (2003). Fluo-rescent assays were conducted with a Spectrafluor Plus(Tecan, Research Triangle, NC) running Magellan v.2.50 software. Assays were conducted in black 96-wellpolystyrene flat clear bottom microtiter plates (CorningInc.) at 30◦C. The total assay volume was 200�l, con-sisting of 180�l Tris buffer (pH 8.0, 20 mM) and 20�lof enzyme preparation. Substrate solutions were pre-pared in ethanol (10 mM) and assays were initiated bythe addition of 2�l substrate solution followed by shak-ing for 10 s. Production of 6-methoxynaphthaldehyewas monitored with excitation at 330 nm (bp 35) andemission at 465 (bp 35). All assays were performedwith the instrument gain set to 60. Assays were config-ured such that no more than 10% of the substrate washydrolyzed during the assay and solvent added neverexceeded 1% of the total assay volume. Reported activi-t achs ed bya dardc ydeq gen-e s ona withta y var-i 5.6t2e h thatt erieso sub-s d fore ciese ub-
alized to the amount of protein in the homogen�mol/min/mg protein).
.5. Carboxylesterase assays
Livers were excised and processed as descbove using Tris buffer (pH 8.0, 20 mM) containmM EDTA. Samples were centrifuged at 9000×g
or 20 min at 4◦C. Esterase assays withp-nitrophenycetate (PNPA) were performed using sodium phate buffer (pH 8.0, 0.1 M) at 30◦C according toethods ofWheelock et al. (2001)as adapted fromjungquist and Augustinsson (1971). Assays were pre
ormed in 96-well microtiter styrene flat bottom plaDynex Technologies, Chantilly, VA) and analyzedSpectramax 340PC plate reader (Molecular Devunnyvale, CA). The total assay volume was 200�l,onsisting of 180�l buffer and 20�l of enzyme prepa
ies were corrected for background hydrolysis. For epecies examined, standard curves were generatdding an equivalent amount of protein to each stanoncentration to account for protein-induced aldehuenching. It is important that standard curves arerated in the presence of authentic protein samplespecies-specific basis. Assays were performed
hree flashes and 10 cycles to give a∼3 min linearssay. The amount of protein added in each assa
ed with the species and sample, but ranged fromo 44�g/well for salmon liver, 10�g/well for splittail,5�g/well for medaka and 0.08�g/well for porcinesterase. Protein concentration was adjusted suc
he assay was linear over the reported time. A sf aliphatic and pyrethroid surrogate fluorescenttrates developed in our laboratory were screenesterase-mediated hydrolysis with the different spexamined in this study. A full description of these s
strates, including their synthesis and use as pyrethroidsurrogates, is described byStok et al. (2004).
Kinetic constants were measured using the assaysdescribed above for each substrate. A range of solu-tions of varying substrate concentration were preparedin ethanol. Total solvent never exceeded 1% of theassay volume. Kinetic constants were calculated usinga nonlinear curve fit with a minimum of nine substrateconcentrations as described bySegel (1976).
Assays to determine the concentration of inhibitorthat reduced enzyme velocity by 50% (IC50) wereperformed according to methods ofWheelock et al.(2004)and designed such that there were at least twodatum points above and below the IC50 value in the lin-ear range. Inhibitor solutions were prepared in ethanoland diluted as required for each assay. Solvent neverexceeded 1% of the assay volume and no solvent effectswere observed.
2.6. Native gel activity assay
Native polyacrylamide gel-electrophoresis (PAGE)analyses were performed using 12% tris-glycine gels(Invitrogen) according to the manufacturer’s instruc-tions. Gels were visualized with the carboxylesteraseactivity stain �-naphthyl acetate using methods ofHuang et al. (1993), and scanned with a UMAX Pow-erlook III flatbed scanner (UMAX Technologies Inc.,Dallas, TX).
2o
ngu -r hedpo( chw osem e,Nm ificb 3( zesC na ngC gG
as the secondary antibody (Jackson ImmunoresearchLaboratories Inc., West Grove, PA) and blots werescanned at 633 nm excitation/670 nm emission usinga Typhoon 8600 scanner (Molecular Dynamics, PaloAlto, CA) and quantified using Scion Image (NIST,http://www.nist.gov/lispix/imlab/labs.html). Livermicrosomes from trout treated with the CYP1A modelinducer,�-naphthoflavone (50 mg/kg, i.p.) were usedas positive controls and loaded in seven concentrations(0.1–7.0�g/well) to evaluate the linearity of theCYP1A signal on each blot. All samples were run inat least triplicate.
2.8. Statistical analyses
The significance of the effects of pesticide treatmentupon esterase assays was analyzed using the statisti-cal package in Microsoft Excel (Redmond, WA). Stu-dentst-test were performed, with significance reportedfor P< 0.05. Michaelis–Menton kinetic analyses wereperformed using two different methods, a double-reciprocal plot (Segel, 1976) and SigmaPlot (SystatSoftware Inc., Richmond, CA). CYP1A data were ana-lyzed using one-way analysis of variance (ANOVA)and the SAS statistical package (SAS, 1985) and homo-geneity of variances was determined by Levene’s test(Draper and Hunter, 1969). Separation of means wastested using Duncan’s multiple range test. All differ-ences were considered significant atP≤ 0.05.
3
3
urec a-s aa sys-t mi-n ults).E ofc denta at8 -i en-v y at1
.7. Immunoblotting procedures for determinationf CYP1A levels
CYP1A protein was quantified by immunoblottising a Bio-Dot SFTM microfiltration slot-blot appaatus (Bio Rad, Hercules, CA) according to publisrocedures (McArdle et al., 2004). Briefly, 20�gf S9 protein was suspended in 200�l of 1× TBSpH 7.5, 20 mM Tris, 0.5 M NaCl), loaded into eaell and vacuum transferred onto a nitrocellulembrane (0.45�M; Schleicher and Schull, KeenH). The membrane was incubated in 1× TBS-5%ilk for 1 h at room temperature to block non-specinding, followed by incubation with MAb 1-12-1:50), a monoclonal antibody which recogniYP1A in multiple vertebrate species (Stegemand Hahn, 1994). CYP1A signal was detected usiyTM 5-conjugated affinipure goat anti-mouse I
. Results
.1. Acute toxicity
Aqueous chlorpyrifos concentrations in exposontainers showed∼20–25% variability between meured and dosed concentrations (Table 1). These datgreed with previous results in similar exposure
ems that showed a <20% variability between noal and measured concentrations (unpublished resxposure of juvenile Chinook salmon to a rangehlorpyrifos concentrations elicited a dose-depencute response with 100% mortality observed1�g/l, 20% mortality at 7.3�g/l and 0% mortal
ty at 1.2�g/l. Fish exposure to a range of esfalerate concentrations resulted in 100% mortalit.0�g/l (nominal) or 0% mortality at 0.01 and 0.1�g/l
a Water concentrations are nominal unless specified. Values inparentheses were quantified using methods ofCrepeau et al. (2000).Unpublished work showed that esfenvalerate concentrations variedfrom nominal concentrations by∼20%.
b Juvenile Chinook salmon were exposed to the indicated concen-tration of pesticide for 96 h (n= 10).
(nominal). All subsequent enzyme assays were per-formed on fish that were exposed to sublethal pesticidedoses.
3.2. Acetylcholinesterase activity
AChE activity varied with pesticide concentrationand tissue type (Table 2). Solvent (methanol) expo-sure altered AChE activity in the brain (11% decrease,P< 0.001), but not in muscle, relative to untreatedcontrols. Significant suppression of AChE activity bychlorpyrifos treatment occurred only at the highestchlorpyrifos dose. Relative to solvent controls, expo-sure to low doses of chlorpyrifos (1.2�g/l) did notsuppress brain AChE, and even slightly elevated muscleAChE levels (112%,P< 0.05). In contrast, high dosechlorpyrifos exposure (7.3�g/l) reduced AChE activityby 85% (brain) and 92% (muscle) compared to solventcontrols, and by 84% (brain) and 93% (muscle) relativeto low dose fish (P< 0.001).
Esfenvalerate exposure did not affect AChE activityin either tissue at the lowest dose, 0.01�g/l. However,at 0.1�g/l, brain AChE activity increased by∼10%and muscle AChE activity decreased by∼10%. Thesedifferences, though slight, were significantly differentfrom solvent controls (0.05 >P> 0.01). Activities atthe two esfenvalerate exposure concentrations were notstatistically different from each other.
Inter-individual variability in AChE activity wasgenerally less in brain (∼10%) than in muscle (∼20%).T he
Table 2Acetylcholinesterase activity in pesticide-exposed juvenile Chinooksalmona
a Juvenile Chinook salmon were exposed to the indicated con-centration of pesticide for 96 h as described in Section2. Only thesalmon exposed to 7.3�g/l chlorpyrifos had significantly inhibitedacetylcholinesterase activity.
b All reported water concentrations are nominal values. Chlorpyri-fos concentrations were measured to be 1.2 and 7.3�g/l (as opposedto 1.0 and 10.0�g/l nominal). Esfenvalerate concentrations wereobserved to deviate by∼20% from nominal concentrations (unpub-lished results).
c Activity values are in units of nmol/min/mg and are the aver-age± the standard deviation (S.D.) for 10 fish (n= 10), except forthe 7.3�g/l chlorpyrifos exposure (n= 8). Assays were performed intriplicate and variability was less than 10%.
d Range values are given as the fold difference in activity betweenthe lowest and highest individuals.
e Control fish were not exposed to either pesticides or vehicle anddid not go through the experimental testing regimen.
f Solvent fish were exposed to the highest concentration of vehicle(0.005% MeOH) and went through the full 96 h testing regimen.
* Statistically different from the control, solvent-exposed, and the1.2�g/l chlorpyrifos-exposed salmon (P< 0.001).
highest level of activity to the individual with the lowestranged from 1.1 for esfenvalerate at 0.1�g/l in muscleto 2.2 at 0.01�g/l esfenvalerate in brain. The majorityof the exposures showed no significant effect upon therange of inter-individual activity, with the exception ofactivity in the muscle after chlorpyrifos exposure. Thissample showed a steady increase in the range of activitywith treatment. However, the overall change was stillrelatively small (from 1.2- to 2-fold).
he ratio in activity between the individual with t
Pesticide exposure affected carboxylesterase activ-ity in a compound and dose-dependent fashion(Table 3). All three substrates examined producedvery similar results, with hydrolysis activity profilesfor each substrate responding identically to pesti-cide exposure. Increasing concentrations of chlorpyri-fos caused significant decreases in carboxylesteraseactivity (Table 3). An identical inhibition pattern wasobserved using an�-naphthyl acetate carboxylesteraseactivity stain in a native gel (Fig. 3). Significant reduc-tions in carboxylesterase activity at the highest chlor-pyrifos dose (7.3�g/l) were detected by all three sub-strates, with reductions of 79% (PNPA), 52% (�-cyanoacetate) and 55% (�-cyano butyrate), relative to solventcontrols. Only one substrate, PNPA, detected a signif-icant reduction (56%) in carboxylesterase activity atthe low chlorpyrifos dose (1.2�g/l;P< 0.001). Solventexposure had a significant effect upon carboxylesteraseactivity for all three substrates, with reductions rang-ing from a 44% decrease in activity for the�-cyano
F os-t 2%t 1:c at1 ex-p
Table 3Carboxylesterase activity in liver cytosol from pesticide-exposedjuvenile Chinook salmona
a Juvenile Chinook salmon were exposed to the indicated concen-tration of pesticide for 96 h as described in Section2.
b Esfenvalerate concentrations are nominal values and chlorpyrifosconcentrations are measured.
c Activity values are in units of nmol/min/mg and are the aver-age± the standard deviation (S.D.) for 10 fish (n= 10), except forthe 7.3�g/l chlorpyrifos exposure (n= 8).
d Range values are given as the fold difference in activity betweenthe lowest and highest individuals.
e Control fish were not exposed to either pesticides or vehicle anddid not go through the experimental testing regimen.
f Carboxylesterase activity assays were performed with the sub-stratep-nitrophenyl acetate (PNPA).
g Carboxylesterase activity assays were performed with the sub-strate�-cyano(6-methoxy-2-naphthyl)methyl acetate (acetate).
h Carboxylesterase activity assays were performed with the sub-strate�-cyano(6-methoxy-2-naphthyl)methyl butyrate (butyrate).
i Solvent fish were exposed to the highest concentration of vehicle(0.005% MeOH) and went through the full 96 h testing regimen.
* Statistically different from the solvent-exposed salmon(P< 0.001).** The solvent-exposed (MeOH vehicle) salmon were statistically
different from the controls (P< 0.01).† The value is statistically different from the solvent-exposed
salmon (P< 0.05).
ig. 3. Carboxylesterase activity in liver cytosol from chlorpyrif
reated juvenile Chinook salmon. Activity was visualized in a 1ris-glycine native gel stained with�-naphthyl acetate. Lanehlorpyrifos-exposed at 10�g/l; lane 2: chlorpyrifos-exposed.0�g/l; lane 3: chlorpyrifos solvent control; lane 4: control unosed fish.
acetate to 23% for PNPA, relative to untreated con-trols. The�-cyano acetate and�-cyano butyrate sub-strates exhibited a wider range of inter-individualcarboxylesterase activities (2.4–3.0- and 1.3–3.9-fold,respectively), than did PNPA (1.3–1.8-fold).
Exposure to esfenvalerate had very little effectupon carboxylesterase activity. All concentrationstested were essentially identical to solvent controlvalues for all substrates, and no significant solventeffects were observed (Table 3). Exposure to 0.01and 0.1�g/l esfenvalerate inhibited carboxylesteraseactivities relative to untreated controls (P< 0.05) whenmeasured with the�-cyano acetate substrate, howevervalues were not significantly different from the solventcontrol. The range in inter-individual variation incarboxylesterase activity in response to esfenvalerateexposure was similar to that observed with chlor-pyrifos. Generally, the range in activity decreasedwith increasing pesticide concentration with all threesubstrates. The only exception occurred with thebutyrate substrate in which the lowest concentrationsof chlorpyrifos and esfenvalerate elicited the greatestrange of activity.
3.4. Determination of carboxylesterase kineticconstants in different species
The carboxylesterase kinetic constants varied withspecies and substrate. It is not appropriate to directlycompare species in this study as the tissues were pre-p canb tail,w on,l car-b a par-t uri-fi cies,k . TheMlt rt mores∼ lit-t 7.8t ndp form on
Table 4Kinetic constants for substrates used in this studya
a Salmon data are from liver homogenates. Medaka and splittaildata are from whole body homogenates, and porcine esterase data arefrom a commercial partly purified preparation. Data are the averageof 3 independent determinations± the standard deviation.
b Kinetic constants were determined for the substratep-nitrophenylacetate (PNPA).
c Data are fromBarron et al. (1999)using rainbow trout liver.d Kinetic constants were determined for the substrate�-cyano(6-
methoxy-2-naphthyl)methyl acetate (acetate).e Kinetic constants were determined for the substrate�-cyano(6-
methoxy-2-naphthyl)methyl butyrate (butyrate).
and porcine enzymes were prepared from single organs(salmon) or were partially purified (porcine) and hadhigher specific activities.
In contrast to the Michaelis constant, the varia-tion in Vmax app was much greater among the differ-ent species examined. This variability tracked car-boxylesterase concentration in the preparation veryclosely, asVmax appvalues are normalized to unit pro-tein. The two samples that were prepared identically(whole body homogenates) had similarVmax app val-ues for PNPA, 198,000 nmol/min/mg for medaka and129,600 nmol/min/mg for splittail, whereas the sam-ples that contained a higher degree of specific activity(salmon liver or porcine esterase) hadVmax app val-ues that were up to 10-fold higher. Similar resultswere reported for studies with rainbow trout liverpreparations, which reported aVmax app for PNPA of672,000 nmol/min/mg (Barron et al., 1999). Similartrends were observed with both the�-cyanoacetate andbutyrate substrates.
ared differently and thus limited conclusionse drawn from the data. For medaka and splithole body homogenates were used, for salm
iver homogenates were used, and for porcineoxylesterase, measurements were made using
ially purified commercial esterase. Because ped carboxylesterases were not used for any speinetic constants are reported as apparent valuesichaelis constant (Km app) for PNPA varied from a
ow of 95�M for medaka to a high of 578�M for split-ail (a six-fold range) as shown inTable 4. Results fohe�-cyanoacetate and butyrate substrates wereimilar to each other than to PNPA.Km appvalues were30�M for the acetate for all species except sp
ail (5.8�M), whereas butyrate values ranged fromo 16.8�M. Interestingly, values for the salmon aorcine enzymes fell within the range observededaka, splittail and trout, even though the salm
The OP pesticides diazinon and chlorpyrifos, andtheir oxon-derivatives, were examined for their abil-ity to inhibit carboxylesterase activity in four speciesusing three different substrates (Table 5). The IC50’s(concentration of enzyme required to reduce enzymevelocity by 50%) for diazinon and chlorpyrifos weregreater than 100�M for all three substrates in all fourspecies, indicating that these two pesticides do notsignificantly inhibit carboxylesterase activity. In con-trast, the oxon forms of both pesticides were significantcarboxylesterase inhibitors, with IC50 values in thelow nM range for all substrates in all species exam-ined, except splittail. In splittails, diazinon-oxon didnot inhibit �-cyano acetate hydrolysis at any concen-tration examined (IC50 > 100�M) while chlorpyrifos-oxon mediated inhibition was as much as 1000-foldless potent than in the other species tested.
Of the two oxons tested, chlorpyrifos-oxon was themore potent carboxylesterase inhibitor, being on aver-age∼10-fold more potent than diazinon-oxon. How-ever, this number varied considerably with substrate
and species. For medaka and the porcine esterase, theIC50s for the two different oxons did not vary greatlywith substrate. However, for the Chinook salmonesterase, the two acetate-containing substrates, PNPAand �-cyano acetate, had IC50s more similar to oneanother than to the IC50 for the�-cyano butyrate sub-strate. This observation is in spite of the fact that thealcohol moiety of�-cyano acetate is quite differentfrom that of PNPA, being similar to�-cyano butyrate(seeFigs. 1 and 2for a description of acid and alcoholnomenclature and substrate structures).
3.6. Pyrethroid hydrolysis
Pyrethroid surrogate hydrolysis was not observedwith any of the fish species examined in this study.Only the porcine enzyme significantly hydrolyzed�-cyanoesters of pyrethroid acids as shown inTable 6. Anumber of assay conditions were varied in an attemptto measure hydrolysis activity. The temperature of theassay was increased up to 37◦C and the pH up to9; however none of these conditions was sufficient toincrease pyrethroid hydrolysis activity to quantifiable
Table 5Inhibition concentrations (IC50) for selected organophosphatesa
a All IC50 concentrations are in nM unless otherwise notedhole body homogenates, and porcine esterase data are frometerminations± the standard deviation.b IC50 determinations were performed with the substratep-nitropc All values for >100 are in�M.d IC50 determinations were performed with the substrate�-cyanoe IC50 values are in�M.f IC50 determinations were performed with the substrate�-cyano
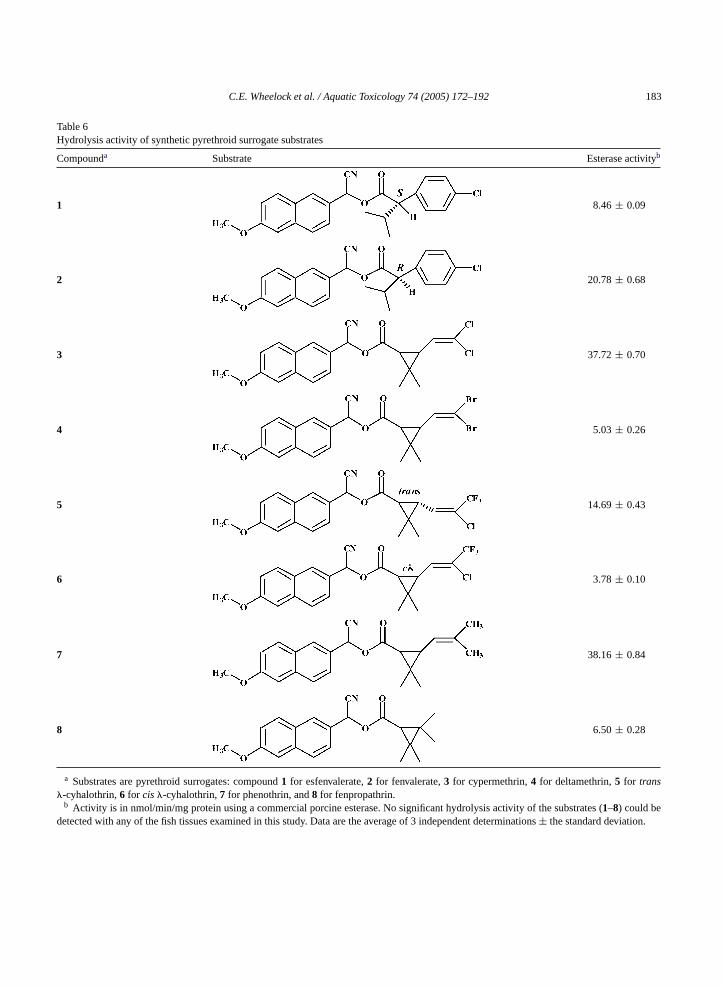
Table 6Hydrolysis activity of synthetic pyrethroid surrogate substrates
Compounda Substrate Esterase activityb
1 8.46± 0.09
2 20.78± 0.68
3 37.72± 0.70
4 5.03± 0.26
5 14.69± 0.43
6 3.78± 0.10
7 38.16± 0.84
8 6.50± 0.28
a Substrates are pyrethroid surrogates: compound1 for esfenvalerate,2 for fenvalerate,3 for cypermethrin,4 for deltamethrin,5 for trans�-cyhalothrin,6 for cis�-cyhalothrin,7 for phenothrin, and8 for fenpropathrin.
b Activity is in nmol/min/mg protein using a commercial porcine esterase. No significant hydrolysis activity of the substrates (1–8) could bedetected with any of the fish tissues examined in this study. Data are the average of 3 independent determinations± the standard deviation.
levels. Unless otherwise stated, the substrates were amixture of isomers. The hydrolysis rates of the eightdifferent pyrethroid surrogates examined did not varyby more than∼10-fold across all compounds (3.78-38.16 nmol/min/mg). For compound1, an esfenvaler-ate mimic, hydrolysis was∼2.5-fold slower than for thecorrespondingR-analog (compound2). The cyperme-thrin surrogate (compound3) was hydrolyzed at thesame rate as its dimethyl analog (synthesized fromchrysanthemic acid, compound7); however substitu-tion of the dichloro moiety by dibromo (deltamethrinsurrogate) decreased hydrolysis by∼7-fold (com-pound4). The�-cyhalothrin surrogates exhibited a 4-fold difference in hydrolysis rate, with thetransisomer(compound5) hydrolyzed faster than thecis (com-pound6).
3.7. CYP1A levels
CYP1A protein expression was slightly, but sig-nificantly (P< 0.05), suppressed (1.4-fold) in salmon
Table 7Relative levels of CYP1A protein in liver cytosol from pesticide-exposed Chinook salmona
Pesticide Concentration(�g/l)b
Average± S.D.c Ranged
Controle 0 1710± 310 1.7
Chlorpyrifos Solventf 2060± 440 2.6
E
ncen-tg
yrifosc
ddp
oteinl
andd
hicle( .
on(
liver cytosol from the high dose (7.3�g/l) chlorpyrifos-treated group relative to those treated with solvent car-rier (Table 7). However, there was no significant effectof chlorpyrifos at either dose relative to untreated con-trols. Esfenvalerate treatment had no effect on hepaticCYP1A protein expression at either dose (Table 7).No significant trends were observed in the range ofCYP1A levels in response to the different pesticideexposures. The range varied from a low of 1.6-fold forthe highest dose of esfenvalerate (similar to the con-trol value of 1.7) to a high of 4.3 for the highest doseof chlorpyrifos. However, variability in response to lowdose esfenvalerate (4.2) was similar to that provoked bythe high dose of chlorpyrifos (4.3), suggesting salmonresponse to these pesticides was generally similar. Thevariability in response to solvent exposure was moder-ate, with a range of 2.5 and 2.6 for esfenvalerate- andchlorpyrifos-exposed fish, respectively, ranges higherthan the response range for untreated controls (1.7).
4. Discussion
Early studies suggested that fish metabolize xeno-biotics at much slower rates than mammals, if at all.More recently it has become clear that xenobioticmetabolism in fish species is often different from thatin mammals, but often very active nonetheless. Exten-sive work has examined cytochrome P450 (Stegemanand Lech, 1991; Goksoyr, 1995; Whyte et al., 2000),Aa ;B s.H il-a hesee ingp
ed inm tinga a,1 04;W ar-b icity( et ergis-t arn,1 s( rase
a Juvenile Chinook salmon were exposed to the indicated coration of pesticide for 96 h as described in Section2. All treatmentroups were statistically similar to untreated controls (P< 0.05).b Esfenvalerate concentrations are nominal values and chlorponcentrations are measured.c Relative CYP1A protein levels are the average± the standareviation (S.D.) for 10 fish (n= 10), except for the 7.3�g/l chlor-yrifos exposure (n= 8).d Range values are given as the fold difference in relative pr
evels between the lowest and highest individuals.e Control fish were not exposed to either pesticides or vehicleid not go through the experimental testing regimen.f Solvent fish were exposed to the highest concentration of ve
0.005% MeOH) and went through the full 96 h testing regimen* Statistically different from the solvent-exposed salmP< 0.05).
ChE (McKim et al., 1987; Fulton and Key, 2001),nd glutathione transferase (GST) (Parker et al., 1993ello et al., 2001) activity in a range of fish specieowever, to date relatively little information is avable on carboxylesterase activity, even though tnzymes interact with many agrochemicals includyrethroids, OPs, and carbamates.
Carboxylesterases have been intensively studiammalian systems due to their role in mediagrochemical-induced toxicity (Satoh and Hosokaw998; Sogorb and Vilanova, 2002; Stok et al., 20heelock et al., 2005). Research has shown that c
oxylesterases reduce pyrethroid-associated toxAbernathy and Casida, 1973) and that joint exposuro pyrethroids and/or carbamates can cause synic toxicity (Gaughan et al., 1980; Gupta and Dettb993). Early work on fish byKingsbury and Master1972)reported the presence of three carboxyleste
isozymes in rainbow trout with the detectionof one polymorphism. Since then a number ofresearchers have examined esterase activities in fish,but the work lags significantly behind that per-formed in mammalian systems (Boone and Chambers,1997; Huang et al., 1997; Sanchez-Hernandez et al.,1998; Barron et al., 1999; Al-Ghais, 2000; Wogram etal., 2001; Denton et al., 2003). This study expands thecurrent literature on carboxylesterases in several fishspecies and examines the appropriateness of multiplebiochemical endpoints as biomarkers of agrochemicalexposure and/or susceptibility.
4.1. Acute toxicity
Significant fish mortality at the highest concentra-tions of both pesticide exposures was anticipated and isconsistent with results from toxicity studies in relatedfish species. Since 96-h LC50 values for chlorpyrifosand esfenvalerate in juvenile Chinook salmon have notbeen reported in the literature, we chose concentra-tions that bracketed those used in studies with rainbowtrout, a related species. The 96-h LC50 for chlorpyri-fos has been reported to range from 9�g/l (Phippsand Holcombe, 1985) to 45�g/l (Kikuchi et al., 1996)in juvenile rainbow trout. For esfenvalerate, only two96-h LC50 studies have been published for rainbowtrout; 0.3�g/l (DuPont, 2002) and 0.07�g/l (U.S. EPA,2000). Based on these limited data, we chose a widerange of pesticide concentrations for our studies toi thatw ltsf or-t ngeo owt routh
4
t andt andpKt thee ilars ent
between the average AChE activity in brain of thecontrol group and mean control activity in a studywith juvenile steelhead trout, with 14.9�mol/min/gbrain tissue in salmon and 15.3�mol/min/g brain tis-sue in steelhead (expressed as wet weight) (Sandahl andJenkins, 2002).
Inhibition of AChE activity in fish is generally cor-related with increased mortality; however differencesamong species and between tissues make it difficult toidentify the lowest level of AChE inhibition that cancause mortality. The sensitivity of brain and muscleAChE, the relationship between tissue-specific AChEinhibition and mortality, as well as the presence andenzymatic activity of butyrylcholinesterase, all appearto be species-specific (Fulton and Key, 2001). Forexample, in most studies with estuarine fish, inhibi-tion levels in excess of 70% correlate with imminentmortality, but inhibition of AChE activity greater than80% in surviving fish is not uncommon (Coppage andMatthews, 1975; Coppage et al., 1975). This obser-vation might be due to species-specific differences intolerance to extremely high levels of brain AChE inhi-bition (Keizer et al., 1995). For this reason, it has beensuggested that muscle AChE inhibition, which exhibitsless species-specific differences than brain AChE inhi-bition, might be a more appropriate predictor of OP-related mortality (Fulton and Key, 2001). In our study,inhibition of brain and muscle AChE following expo-sure to 7.3�g/l chlorpyrifos was roughly equal. How-ever, at the lower chlorpyrifos dose (1.2�g/l) AChEi est-i OPi wellb ilars yi es7 %)a -i ina
erscec dfH elowl nd iti ifos
dentify doses that were acutely toxic and dosesould potentially elicit sublethal toxicity. Our resu
or chlorpyrifos- and esfenvalerate-induced acute mality identified concentrations within the general raf 96-h LC50 concentrations reported for rainb
rout, suggesting Chinook salmon and rainbow tave similar sensitivities to these pesticides.
.2. Acetylcholinesterase activity
Inhibition of AChE activity is linked directly withhe mechanism of toxic action of OP insecticides,hus is often used as an indicator of OP exposurehysiological effect in exposed animals (Fulton andey, 2001). The AChE activity data (Table 2) from
his study suggest either that multiple isoforms ofnzyme do not exist, or, if they do, that all have simensitivity to chlorpyrifos. We found good agreem
nhibition was only observed in the brain, suggng that this tissue is a more sensitive indicator ofnsecticide exposure than muscle at concentrationselow those causing mortality. Results from simtudies (Fulton and Key, 2001) suggest that mortalits likely to occur when brain AChE inhibition reach0–80%. However, we observed little mortality (20t chlorpyrifos doses (7.3�g/l) that substantially inhib
ted AChE activity (85% and 92% inhibition in brand muscle, respectively).
OP insecticide concentrations in California rivontinue to exceed water quality standards (Wernert al., 2000; Phillips et al., 2004), and chlorpyrifosoncentrations as high as 3.2�g/l have been reporteor the Central California Coast (Hunt et al., 2003).owever, measured concentrations are generally b
evels that cause mortality in laboratory studies, as uncertain to what extent environmental chlorpyr
concentrations affect AChE activity. A number of stud-ies reported a link between AChE inhibition in fish andsublethal behavioral and physiological effects, such asreduced swimming stamina (Post and Leasure, 1974;Van Dolah et al., 1997), altered feeding (Wildish andLister, 1973; Bull and McInerney, 1974), and alteredsocial interactions (Symons, 1973). In mammalian andavian systems, it has been suggested that AChE mayplay a direct role in the development of the nervous sys-tem (Brimijoin and Koenigsberger, 1999; Lauder andSchambra, 1999). It is therefore possible that inhibi-tion of AChE activity at sublethal levels is having anadverse effect upon fish health and ecological viability.
4.3. Carboxylesterase activity
Carboxylesterase activity exhibited a dose–response relationship, with activity decreasing withincreasing chlorpyrifos concentration (Tables 2 and 3).However following exposure to 7.3�g/l chlorpyrifos,a significant amount of carboxylesterase activity(∼65%) was still observed with the�-cyano acetateand butyrate substrates (as opposed to PNPA, whichhad 21% remaining). This observation suggests thepresence of multiple isozymes, with a significant levelof activity (∼40–50%) not sensitive to chlorpyrifosinhibition. These data agree with results reported byDenton et al. (2003)who showed that exposure todiazinon resulted in a maximum of∼50% inhibitionof carboxylesterase activity in fathead minnows inv anys ityi the1 %.T as ap asei t ones ge ofv itivei hisv r-m ure,b e top ingc
gc andd sub-
strate to examine activity of crude tissue homogenate.Given that there are likely multiple esterase isoformspresent in the preparation, it is necessary to have arange of reporters of activity for full characteriza-tion. Generally the substrate PNPA, or other analogsusing p-nitrophenol as a reporter, is employed dueto its ease of use, availability and colored hydrolysisproduct. However, it is likely that there are additionalesterase isozymes that do not hydrolyze this substrate.Correlation analyses performed on PNPA hydroly-sis and pyrethroid hydrolysis in human liver showedvery little correlation between the hydrolytic profiles(r2 = 0.29 for a fenvalerate surrogate) (Wheelock et al.,2003). These results suggest that different enzymesare involved in the hydrolysis of the different sub-strates. Therefore monitoring of PNPA activity, or thatof other general substrates, may not provide an accu-rate account of pyrethroid hydrolysis activity. In thisstudy, PNPA appeared to be a more sensitive indica-tor of activity/inhibition than the�-cyano substrates,reporting significant inhibition at the lowest level ofchlorpyrifos examined. However, the�-cyano acetateand butyrate substrates exhibited a wider range in activ-ities than PNPA, suggesting that they are hydrolyzedby more isozymes than PNPA.
All of the fish species examined in this study wereunable to hydrolyze the pyrethroid surrogate substratesshown inTable 6at significant levels. Substrates suchas those developed byRiddles et al. (1983), whichcouple the leaving groupp-nitrophenol to pyrethroida roidh beu talc toh ento gatess er-ca oids d beu
tor-i theI it-t thatd al-u rc with
ivo. Even though esfenvalerate did not causeignificant inhibition of carboxylesterase activn our studies, the range in activities amongst0 fish did significantly drop by as much as 50his observation suggests that esfenvalerate hreviously unknown effect upon carboxylester
sozyme abundance. These data also suggest thaet of pesticide sensitive isozymes has a wide ranariability, but that a second set of pesticide insenssozymes has a narrower range of variability. Tariability in activity could be important in deteining the effects of OP and/or carbamate exposecause some individuals may be more sensitivesticide exposure due to lower levels of detoxifyarboxylesterases or possible polymorphisms.
The data inTable 3show the effect of measurinarboxylesterase activity with different substratesemonstrate that it is not appropriate to use a single
cids may be improved general reporters of pyrethydrolysis activity compared to PNPA. They couldseful tools for determining if specific environmenontaminants interfered with an organism’s abilityydrolyze pyrethroids, versus an overall measuremf general esterase activity. However, these surrotill vary greatly in the alcohol portion from commial pyrethroids and the substrates reported byStok etl. (2004)are probably more appropriate pyrethrurrogates. Ultimately, the actual pesticide shoulsed to test for activity.
The importance of substrate choice for moning esterase activity was further demonstrated byC50 data inTable 5. Inhibition assays with the splail homogenate showed a very striking result iniazinon-oxon was a potent inhibitor (nM IC50 ves) when assays were run with either PNPA o�-yano butyrate. However, when assays were run
�-cyano acetate, no inhibition was observed. Simi-lar results were observed with the chlorpyrifos-oxonassays. These data strongly suggest that a battery ofsubstrates should be employed when measuring car-boxylesterase activity to ensure that an accurate indi-cation of enzyme activity is obtained.
It is possible that the ability to detoxify pyrethroidsvia hydrolysis is inversely correlated with pyrethroidtoxicity, but there are not currently enough data avail-able in the literature to fully examine this issue. Car-boxylesterase activity is most likely important forpyrethroid detoxification in some species of fish, butwork by Glickman and coworkers showed that themost important factor in rainbow trout sensitivity topermethrin was target site sensitivity at the sodiumchannel (Glickman et al., 1981; Glickman and Lech,1981, 1982). Rainbow trout had lower esterase activ-ity than rats, and thus a decreased ability to hydrolyzepermethrin. However, after inhibiting all measurableesterase activity in both rainbow trout and rat usingan esterase inhibitor, the rainbow trout were still moresensitive to permethrin toxicity. It is still possible thatin some cases, different species will have greater lev-els of esterase activity, which could affect the ability todetoxify agrochemicals.Glickman et al. (1979)showedthat carp had higher levels of esterase activity and agreater ability to hydrolyze permethrin than rainbowtrout, which could potentially account for observedinter-species differences in pyrethroid toxicity.
Carboxylesterase activity may be a more sensitivem tiv-i ofA als,m rba-m y,2 ay,2a po-s fr easeda hatc verA ba-m rte,1 4I forO orkb of
both cholinesterase and carboxylesterase activityin the musselMytilus galloprovincialis to monitorpollution in an agricultural region of Spain withknown applications of OPs and carbamates. Theyfound that carboxylesterase activity was more sensitiveto pesticide exposure and that seasonal variation incarboxylesterase activity correlated with pesticide loadin the organism. Based upon their observations, theypostulated that carboxylesterase activity could serveas a protective mechanism for OP inhibition of AChE.A similar result was reported byWogram et al. (2001)using the three-spined stickleback (Gasterosteusaculeatus), who found that carboxylesterase was13–17-fold more sensitive to paraoxon than AChE.O’Neill et al. (2004)stated that both cholinesteraseand carboxylesterase activity declined in responseto exposure to sewage effluent discharge in north-west England; however carboxylesterase was moreseverely inhibited. These studies support the conceptof using carboxylesterase activity as a biomarker ofOP/carbamate exposure either alone or in combinationwith AChE activity.
We had originally hoped to examine correlationsbetween carboxylesterase activity and pyrethroid andOP toxicity. However, there is not enough informationavailable in the literature to draw direct correlationsbetween toxicity (such as LC50) and esterase activity.One could envision a linear free energy relationshipthat correlated carboxylesterase and/or AChE activityto a toxicity endpoint for a given pesticide or classo nede e) ino on-d ientd nos rvea cals.A lti-p ta hati ark-e oneo teds ow-e achi e. Ak E orc ment
arker for agrochemical exposure than AChE acty. Many different groups have studied the useChE as a biomarker of exposure to agrochemicostly OPs, but some work has focused on caates as well (Sturm et al., 2000; Fulton and Ke001; Galloway et al., 2002; Rickwood and Gallow004; Bonacci et al., 2004). It is inconclusive if AChEctivity alone is an appropriate biomarker of OP exure (Rickwood and Galloway, 2004). A number oesearchers have shown that many OPs have incrffinity for carboxylesterase over AChE, indicating tarboxylesterase would be preferentially inhibited oChE following exposure to OPs and potentially carates (Gupta and Dettbarn, 1993; Escartin and Po997; Wogram et al., 2001; O’Neill et al., 200).
n other words, carboxylesterases act as a sinkPs, thereby rescuing AChE from OP toxicity. Wy Escartin and Porte (1997)examined the use
f pesticides. It would be useful if studies examisterase activity (both AChE and carboxylesterasrganisms for which toxicity assays were being cucted in the hopes of eventually collecting sufficata for the correlation analyses. It is likely thatingle biological measurement or biomarker will ses a universal reporter of exposure to agrochemilogical approach would be the integration of mu
le biological or physiological endpoints.Galloway el. (2004)developed a multibiomarker approach t
ncorporates a suite of ecologically relevant biomrs. They found that carboxylesterase activity wasf the most discriminating markers among polluites, with correlation coefficients as high as 0.93. Hver, one problem with this multibiomarker appro
s that the analysis is labor intensive and expensivey advantage of a single end-point such as ACharboxylesterase activity is the ease of measure
and low cost. It may be necessary to have multiple lay-ers of testing that depend upon the available funds andlabor as well as the rigor of the answer required. How-ever, with the increasing availability of assays designedfor 96- or 384-well applications and the advent ofrobotics, one could design experiments to look at multi-ple enzymes in several species under different exposureconditions in a cost and time effective manner.
4.4. CYP1A levels
Induction of CYP1A is extensively used as an indi-cator of exposure and response to organic pollutants inteleost fish and other vertebrates (Stegeman and Hahn,1994). However, suppression of CYP1A can compli-cate and limit its utility as a biomarker. For example,metals (Fent and Bucheli, 1994) and hormones (Elskuset al., 1992; Elskus, 2004) have been shown to suppressCYP1A expression in fish, making it imperative thatreproductive stage, gender and the presence of metalsbe taken into consideration when interpreting CYP1Adata. It is possible that pesticides can co-occur withCYP1A-inducing chemicals, making it critical that reg-ulation of CYP1A by pesticides is understood.
As expected, CYP1A protein was not induced injuvenile Chinook salmon exposed to either chlorpyrifosor esfenvalerate. Aromaticity and planarity are gener-ally considered two structural features important forchemical induction of CYP1A (Safe, 1990). Inducerswith these features include planar chlorinated aromatich o-d ingi esepic aro-m itya 1A.
int yri-f ctso noe th-e P1Amode rate
affects CYP forms in fish other than CYP1A (Barryet al., 1995), suggesting its effects may be isozyme-specific.
Chlorpyrifos had little effect on CYP1A proteinlevels in salmon, suggesting OPs may not be strongregulators of CYP1A in fish. We know of no other stud-ies examining chlorpyrifos effects on CYP1A proteinin vertebrates. However, studies with the OP methi-dathion found that it slightly increased CYP1A proteinlevels in cyprinids after 4 days of aqueous exposure(Flammarion et al., 1998). This observation is in con-trast to our present study in Chinook salmon in whicha 4-day exposure to chlorpyrifos suppressed CYP1Aprotein levels (Table 7). However, even though bothstudies found OPs altered CYP1A protein levels infish, these alterations were slight, and suggest that OPsare, at best, weak regulators of CYP1A protein in thefish studied. Rather, the main effect of OPs on CYP1Aappears to be catalytic suppression (Flammarion et al.,1996, 1998; Paolini et al., 1997; Tang et al., 2002).Organophosphate effects on CYPs occur as a resultof CYP-mediated metabolism, where OPs, such aschlorpyrifos, undergo desulfuration by CYPs leadingto release of a free sulfur ion, which binds to theCYP heme and inhibits catalytic activity (Fukuto, 1990;Tang et al., 2002).
It is unlikely that reduced CYP1A protein expres-sion in chlorpyrifos-treated fish would significantlyaffect chlorpyrifos toxicity. Induced CYP1A activitylevels do not affect either the activation (via desulfura-t osi ,t thato 4)a ta areu
5
are wasi lso elso ante ndi-v as
ydrocarbons, such as 2,3,7,8-tetrachlorodibenzp-ioxin and certain polychlorinated biphenyls, mak
t one of the most widely used biomarkers of thollutants in vertebrates (Buchneli and Fent, 1995),
ncluding salmonids (Whyte et al., 2000). While bothhlorpyrifos and esfenvalerate have chlorinatedatic rings (Fig. 1), neither chemical exhibits planarnd would therefore not be expected to induce CYP
Although CYP1A expression was not inducedhe pesticide exposed fish in this study, chlorpos and esfenvalerate did exhibit differential effen salmon CYP1A. We found esfenvalerate hadffect on CYP1A protein levels, in keeping with ors who report esfenvalerate had no effect on CYRNA levels in rat hepatocytes (Heder et al., 2001),r on CYP1A catalytic activity (ethoxyresorufin-O-eethylase, EROD) levels in adult rainbow fish (Barryt al., 1995). There is some evidence that esfenvale
ion) or detoxification (via dearylation) of chlorpyrifn channel catfish (Straus et al., 2000). These findingsogether with studies in humans demonstratingther CYP isoforms (CYP2B6, CYP2C19, CYP3Are responsible for chlorpyrifos metabolism (Tang el., 2001), suggest alterations in teleost CYP1Anlikely to affect the toxicity of chlorpyrifos in fish.
. Conclusion
This study showed that activity in two similnzymes systems, carboxylesterase and AChE,
nhibited in vivo by exposure to chlorpyrifos. Levef CYP1A were slightly suppressed at higher levf chlorpyrifos, but esfenvalerate had no significffect upon CYP1A levels or esterase activities. Iidual variability in all three enzymes examined w
fairly narrow, suggesting that it is appropriate to reportenzyme activity from homogenates prepared from tis-sues of combined individuals. However, the small sam-ple size (n= 10) may preclude the identification of indi-viduals with decreased enzyme activity if it occurs withlow frequency in the population. Of particular interestis the observation that solvent-exposed fish evidencedsignificant inhibition of carboxylesterase activity, eventhough the level of solvent was only 0.005%. Thisobservation could have implications for in vivo stud-ies on carboxylesterase activity. The use of multiplesubstrates to examine carboxylesterase activity showedsubstrate-specific responses, with PNPA detecting inhi-bition of activity at low chlorpyrifos doses that wasnot observed with other substrates. This observationcombined with the lack of correlation between dif-ferent esterase substrates suggests that it is necessaryto use a battery of substrates when measuring car-boxylesterase activity. Inhibition studies with splittailcarboxylesterase activity further supported this find-ing, with IC50 values varying over several ordersof magnitude for different substrates examined. Thelack of observed hydrolysis activity of�-cyanoestersof pyrethroid acids for all fish species examined inthis study suggests that these fish have very littleesterase-mediated pyrethroid metabolism. This obser-vation could account for some of the extreme sensitivityof fish to pyrethroid toxicity. Results indicated thatChinook salmon have multiple liver carboxylesteraseisozymes, whose activity can be measured with a com-b
s ofc E,s thanA e useo po-s e ofu s itsr ea-s e toO sus-c rm iona onA it isa hise activ-i r of
OP exposure is that results are integrative. Exposure tomultiple OPs should result in a concomitant decreasein enzyme activity, indicating total effects upon theexposed organism. However, there is little informa-tion in the literature on carboxylesterase activity andfurther work should attempt to determine constitutivelevels in species important for biomonitoring projects.Carboxylesterase activity could serve as a key indica-tor of an organism’s exposure to agrochemicals or as acomponent of a comprehensive monitoring program toexamine overall ecosystem health.
Acknowledgements
CEW was supported by NIH Post Doctoral traininggrant T32 DK07355-22 and a UC TSR&TP GraduateFellowship. PDJ was supported by NIH Post Doctoraltraining grant T32 DK07355-22. This work was sup-ported in part by the Calfed Ecosystem Restoration Pro-gram Project #99-N08, NIEHS Grant R37 ES02710,NIEHS Superfund Grant P42 ES04699, NIEHS Centerfor Environmental Health Sciences Grant P30 ES05707and NIH/NIAID Grant U01 AI058267. The authorsthank the Nimbus Fish Hatchery, the Center for AquaticBiology and Aquaculture, the staff of the Aquatic Tox-icology Laboratory, and Paul Lutes, John Hendersonand Gina Lee. The CYP1A work was supported bythe Department of the Interior, U.S. Geological Surveyand the University of Kentucky Research Foundation,G en-t iewsa se oft ssar-i sedo
R
A terase236.
A c-fish.
B is-icol.
B ay,surerain-
ination of substrates.Carboxylesterase activity was inhibited at level
hlorpyrifos that did not significantly affect AChuggesting a greater affinity of carboxylesterasesChEs for some OPs. This observation supports thf carboxylesterase activity as a biomarker of exure to agrochemicals. The additional advantagsing carboxylesterase activity as a biomarker iole in detoxifying pyrethroids. Therefore one murement provides both a biomarker of exposurPs and/or carbamates as well as a biomarker ofeptibility to pyrethroid toxicity. It is likely that eithearker alone would not provide enough informatnd given both the large amount of existing dataChE levels as well as the ease of the assay,ppropriate to continue monitoring the activity of tnzyme system. An advantage of using esterase
ty (both carboxylesterase and AChE) as a reporte
rant Agreement No. 01HQGR0133, through the Kucky Water Resources Research Institute. The vnd conclusions contained in this document are tho
he authors and should not be interpreted as necely representing the official policies, either expresr implied, of the U.S. Government.
eferences
bernathy, C.O., Casida, J.E., 1973. Pyrethroid insecticides: escleavage in relation to selective toxicity. Science 179, 1235–1
l-Ghais, S.M., 2000. Differential inhibition of xenobiotimetabolizing carboxylesterases by organotins in marineEcotoxicol. Environ. Safety 46, 258–264.
arry, M.J., Ohalloran, K., Logan, D.C., Ahokas, J.T., HoldwD.A., 1995. Sublethal effects of esfenvalerate pulse-expoon spawning and non-spawning Australian crimson-spotted
Bello, S.M., Franks, D.G., Stegeman, J.J., Hahn, M.E., 2001.Acquired resistance to Ah receptor agonists in a populationof Atlantic killifish (Fundulus heteroclitus) inhabiting a marinesuperfund site: in vivo and in vitro studies on the inducibility ofxenobiotic metabolizing enzymes. Toxicol. Sci. 60, 77–91.
Bonacci, S., Browne, M.A., Dissanayake, A., Hagger, J.A., Corsi,I., Focardi, S., Galloway, T.S., 2004. Esterase activities in thebivalve molluscAdamussium colbeckias a biomarker for pollu-tion monitoring in the Antarctic marine environment. Mar. Pollut.Bull. 49, 445–455.
Boone, J.S., Chambers, J.E., 1997. Biochemical factors contributingto toxicity differences among chlorpyrifos, parathion, and methylparathion in mosquitofish (Gambusia affinis). Aquat. Toxicol. 39,333–343.
Bradbury, S.P., Coats, J.R., 1989b. Toxicokinetics and toxicodynam-ics of pyrethroid insecticides in fish. Environ. Toxicol. Chem. 8,373–380.
Brimijoin, S., Koenigsberger, C., 1999. Cholinesterases in neuraldevelopment: new findings and toxicologic implications. Envi-ron. Health Persp. 107, 59–64.
Buchneli, T., Fent, K., 1995. Induction of cytochrome P450 as abiomarker for environmental contamination in aquatic ecosys-tems. Crit. Rev. Environ. Sci. Technol. 25, 201–268.
Bull, C.J., McInerney, J.E., 1974. Behavior of juvenile coho salmon(Oncorhynchus-kisutch) exposed to Sumithion (fenitrothion),an organophosphate insecticide. J. Fish. Res. Board Can. 31,1867–1872.
Casida, J.E., Gammon, D.W., Glickman, A.H., Lawrence, L.J., 1983.Mechanisms of selective action of pyrethroid insecticides. Annu.Rev. Pharmacol. Toxicol. 23, 413–438.
C m ofrumford
C earch:
C logy:rgets.
C inhi-ures) in
C 75.is of
hys-
C sises inhro-
urvey
California district organic chemistry laboratory, 1996–1999. U.S.Geological Survey Open-File Report 00-229, 19 pp.
DeKoven, D.L., Nunez, J.M., Lester, S.M., Conklin, D.E., Marty,G.D., Parker, L.M., Hinton, D.E., 1992. A purified diet formedaka (Oryzias latipes): refining a fish model for toxicolog-ical research. Lab. Anim. Sci. 42, 180–189.
Denton, D.L., Wheelock, C.E., Murray, S., Deanovic, L.A., Ham-mock, B.D., Hinton, D.E., 2003. Joint acute toxicity of esfen-valerate and diazinon to fathead minnow (Pimephales promelas)larvae. Environ. Toxicol. Chem. 22, 336–341.
Ellman, G.L., Courtney, K.D., Andres Jr., V., Feather-Stone, R.M.,1961. A new and rapid colorimetric determination of acetyl-cholinesterase activity. Biochem. Pharmacol. 7, 88–95.
Elskus, A., Pruell, R.J., Stegeman, J.J., 1992. Endogenously-mediated, pretranslational suppression of cytochrome P450IAexpression in PCB-contaminated flounder. Mar. Environ. Res.34, 97–101.
Elskus, A.A., 2004. Estradiol and estriol suppress CYP1A expressionin rainbow trout primary hepatocytes. Mar. Environ. Res. 58,463–467.
Escartin, E., Porte, C., 1997. The use of cholinesterase and car-boxylesterase activities fromMytilus galloprovincialisin pol-lution monitoring. Environ. Toxicol. Chem. 16, 2090–2095.
Fent, K., Bucheli, T.D., 1994. Inhibition of hepatic-microsomalmonooxygenase system by organotins in-vitro in fresh-water fish.Aquat. Toxicol. 28, 107–126.
Flammarion, P., Migeon, B., Garric, J., 1996. Joint effects of cop-per sulphate and methidathion on rainbow trout (Oncorhynchusmykiss) EROD and AChE activities. Bull. Environ. Contam. Tox-icol. 56, 440–445.
Flammarion, P., Migeon, B., Urios, S.B., Morfin, P., Garric, J.,1998. Effect of methidathion on the cytochrome P-450 1A inthe cyprinid fish gudgeon (Gobio gobio). Aquat. Toxicol. 42,
F s and4.
F inphos-hem.
G 02.sure intiv-
G we,kerhnol.
G tions:, tox-estic.
G im-out.
asida, J.E., Quistad, G.B., 1995. Metabolism and synergispyrethrins. In: Casida, J.E., Quistad, G.B. (Eds.), PyrethFlowers: Production, Chemistry, Toxicology, and Uses. OxUniversity Press, New York, NY, pp. 258–276.
asida, J.E., Quistad, G.B., 1998. Golden age of insecticide respast, present, or future? Annu. Rev. Entomol. 43, 1–16.
oppage, D.L., Matthews, E., 1975. Brain acetylcholinesterasebition in a marine teleost during lethal and sublethal exposto 1,2-dibromo-2,2-dichloroethyl dimethyl phosphate (Naledseawater. Toxicol. Appl. Pharmacol. 31, 128–133.
oppage, D.L., Matthews, E., Cook, G.H., Knight, J., 19Brain acetylcholinesterase inhibition in fish as a diagnosenvironmental poisoning by malathion,O,O-dimethyl S-(1,2-dicarbethoxyethyl) phosphorodithioate. Pestic. Biochem. Piol. 5, 536–542.
repeau, K.L., Baker, L.M., Kuivila, K.M., 2000. Method of analyand quality-assurance practices for determination of pesticidwater by solid-phase extraction and capillary-column gas cmatography/mass spectrometry at the U.S. Geological S
93–102.ukuto, T.R., 1990. Mechanism of action of organophosphoru
alloway, T.S., Brown, R.J., Browne, M.A., Dissanayake, A., LoD., Jones, M.B., Depledge, M.H., 2004. A multibiomarapproach to environmental assessment. Environ. Sci. Tec38, 1723–1731.
aughan, L.C., Engel, J.L., Casida, J.E., 1980. Pestcide interaceffects of organophosphorus pesticides on the metabolismicity, and persistence of selected pyrethroid insecticides. PBiochem. Physiol. 14, 81–85.
lickman, A.H., Hamid, A.A., Rickert, D.E., Lech, J.J., 1981. Elination and metabolism of permethrin isomers in rainbow trToxicol. Appl. Pharmacol. 57, 88–98.
Glickman, A.H., Lech, J.J., 1981. Hydrolysis of permethrin, apyrethroid insecticide, by rainbow trout and mouse tissuesin vitro: a comparative study. Toxicol. Appl. Pharmacol. 60,186–192.
Glickman, A.H., Lech, J.J., 1982. Differential toxicity oftrans-permethrin in rainbow trout and mice. II. Role of target organsensitivity. Toxicol. Appl. Pharmacol. 66, 162–171.
Glickman, A.H., Shono, T., Casida, J.E., Lech, J.J., 1982. In vitrometabolism of permethrin isomers by carp and rainbow trout livermicrosomes. J. Agric. Food Chem. 27, 1038–1041.
Glickman, A.H., Weitman, S.D., Lech, J.J., 1982. Differential tox-icity of trans-permethrin in rainbow trout and mice. I. Role ofbiotransformation. Toxicol. Appl. Pharmacol. 66, 153–161.
Goksoyr, A., 1995. Use of cytochrome P450 1A (CYP1A) in fish asa biomarker of aquatic pollution. Arch. Toxicol. Suppl. 17, 80–95.
Gupta, R.C., Dettbarn, W.D., 1993. Role of carboxylesterases inthe prevention and potentiation ofN-methylcarbamate toxicity.Chem. Biol. Interact. 87, 295–303.
Heder, A.F., Hirsch-Ernst, K.I., Bauer, D., Kahl, G.F., Desel, H.,2001. Induction of cytochrome P4502B1 by pyrethroids in pri-mary rat hepatocyte cultures. Biochem. Pharmacol. 62, 71–79.
Huang, T.L., Obih, P.O., Jaiswal, R., Hartley, W.R., Thiyagarajah,A., 1997. Evaluation of liver and brain esterases in the spottedgar fish (Lepisosteus oculatus) as biomarkers of effect in thelower Mississippi River Basin. Bull. Environ. Contam. Toxicol.58, 688–695.
Huang, T.L., Szekacs, A., Uematsu, T., Kuwano, E., Parkinson, A.,Hammock, B.D., 1993. Hydrolysis of carbonates, thiocarbonates,carbamates, and carboxylic esters of�-naphthol,�-naphthol, andp-nitrophenol by human, rat, and mouse liver carboxylesterases.Pharm. Res. 10, 639–648.
Hunt, J.W., Anderson, B.S., Phillips, B.M., Nicely, P.N., Tjeerdema,R.S., Puckett, H.M., Stephenson, M., Worcester, K., De Vlaming,V., 2003. Ambient toxicity due to chlorpyrifos and diazinon in
ess.
K Vit-nonlec-71,
K ofbow
K eighterase58,
L etyl-
L per-ur. J.
L Pro-hem.
Maxwell, D.M., 1992. The specificity of carboxylesterase protec-tion against the toxicity of organophosphate compounds. Toxicol.Appl. Pharmacol. 114, 306–312.
McArdle, M.E., McElroy, A.E., Elskus, A.A., 2004. CYP1A, CYP3Aand estradiol 2-hydroxylase in fish exposed to organic pollutantsand sewage effluent. Environ. Toxicol. Chem. 23, 953–959.
McKim, J.M., Bradbury, S.P., Niemi, G.J., 1987. Fish acute toxi-city syndromes and their use in the QSAR approach to hazardassessment. Environ. Health Persp. 71, 171–186.
O’Neill, A.J., Galloway, T.S., Browne, M.A., Dissanayake, A.,Depledge, M.H., 2004. Evaluation of toxicity in tributaries ofthe Mersey estuary using the isopodAsellus aquaticus(L.). Mar.Environ. Res. 58, 327–331.
Paolini, M., Pozzetti, L., Sapone, A., Mesirca, R., Perocco, P.,Mazzullo, M., Cantelli-Forti, G., 1997. Molecular non-geneticbiomarkers of effect related to acephate cocarcinogenesis: sex-and tissue-dependent induction or suppression of murine CYPs.Cancer Lett. 117, 7–15.
Parker, L.M., Lauren, D.J., Hammock, B.D., Winder, B., Hinton,D.E., 1993. Biochemical and histochemical properties of hepatictumors of rainbow trout,Oncorhynchus mykiss. Carcinogenesis14, 211–217.
Phillips, B.M., Anderson, B.S., Hunt, J.W., Nicely, P.A., Kosaka,R.A., Tjeerdema, R.S., de Vlaming, V., Richard, N., 2004. Insitu water and sediment toxicity in an agricultural watershed.Environ. Toxicol. Chem. 23, 435–442.
Phipps, G.L., Holcombe, G.W., 1985. A method for aquatic multi-ple species toxicant testing acute toxicity of 10 chemicals to 5vertebrates and 2 invertebrates. Environ. Pollut. A 38, 141–158.
Poet, T.S., Wu, H., Kousba, A.A., Timchalk, C., 2003. In vitro rathepatic and intestinal metabolism of the organophosphate pesti-cides chlorpyrifos and diazinon. Toxicol. Sci. 72, 193–200.
Post, G., Leasure, R.A., 1974. Sublethal effect of malathion to threesalmonid species. Bull. Environ. Contam. Toxicol. 12, 312–319.
R hibi-
oxi-
R pli-
the132,
Senvi-t theox-
S i, S.,focusiver
S sesti-col.
a central California coastal watershed. Environ. Monit. Ass82, 83–112.
eizer, J., D’Agostino, G., Nagel, R., Volpe, T., Gnemi, P.,tozzi, L., 1995. Enzymological differences of AChE and diazihepatic metabolism: correlation of in vitro data with the setive toxicity of diazinon to fish species. Sci. Total Environ. 1213–220.
ikuchi, M., Miyagaki, T., Wakabayashi, M., 1996. Evaluationpesticides used in golf links by acute toxicity test on raintrout. Bull. Jpn. Soc. Sci. Fish. 62, 414–419.
ingsbury, N., Masters, C.J., 1972. Heterogeneity, molecular winterrelationships and developmental genetics of the estisoenzymes of the rainbow trout. Biochim. Biophys. Acta 2455–465.
auder, J.M., Schambra, U.B., 1999. Morphogenetic roles of accholine. Environ. Health Persp. 107, 65–69.
jungquist, A., Augustinsson, K.B., 1971. Purification and proties of two carboxylesterases from rat-liver microsomes. EBiochem. 23, 303–313.
owry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951.tein measurement with the Folin phenol reagent. J. Biol. C193, 265–275.
ickwood, C.J., Galloway, T.S., 2004. Acetylcholinesterase intion as a biomarker of adverse effect. A study ofMytilus edulisexposed to the priority pollutant chlorfenvinphos. Aquat. Tcol. 67, 45–56.
iddles, P.W., Schnitzerling, H.J., Davey, P.A., 1983. Apcation of trans andcis isomers of p-nitrophenyl-(1R,S)-3-(2,2-dichlorovinyl)-2,2-dimethylcyclopropanecarboxylate toassay of pyrethroid hydrolyzing esterases. Anal. Biochem.105–109.
afe, S., 1990. Polychlorinated biphenyls (PCBs), dibenzo-p-dioxins(PCDDs), dibenzofurans (PCDFs), and related compounds:ronmental and mechanistic considerations which suppordevelopment of toxic equivalency factors (TEFs). Crit. Rev. Ticol. 21, 51–88.
anchez-Hernandez, J.C., Fossi, M.C., Leonzio, C., Focard1998. Use of biochemical biomarkers as a screening tool tothe chemical monitoring of organic pollutants in the Biobio Rbasin (Chile). Chemosphere 37, 699–710.
andahl, J.F., Jenkins, J.J., 2002. Pacific steelhead (Oncorhynchumykiss) exposed to chlorpyrifos: Benchmark concentrationmates for acetylcholinesterase inhibition. Environ. ToxiChem. 21, 2452–2458.
Segel, I.H., 1976. Biochemical Calculations, 2nd ed. John Wiley &Sons Inc., New York, p. 441.
Shan, G., Hammock, B., 2001. Development of sensitive esteraseassays based on�-cyano-containing esters. Anal. Biochem. 299,54–62.
Sogorb, M.A., Vilanova, E., 2002. Enzymes involved in the detoxi-fication of organophosphorus, carbamate and pyrethroid insecti-cides through hydrolysis. Toxicol. Lett. 128, 215–228.
Stegeman, J.J., Hahn, M.E., 1994. Biochemistry and molecular biol-ogy of monooxygenases: current perspectives on forms, func-tions, and regulation of cytochrome P450 in aquatic species.In: Malins, D.C., Ostrander, G.K. (Eds.), Aquatic Toxicology:Molecular, Biochemical, and Cellular Perspectives. Lewis, AnnArbor, pp. 87–206.
Stegeman, J.J., Lech, J.J., 1991. Cytochrome P-450 monooxygenasesystems in aquatic species: carcinogen metabolism and biomark-ers for carcinogen and pollutant exposure. Environ. Health Persp.90, 101–109.
Stok, J., Huang, H., Jones, P.D., Wheelock, C.E., Morisseau, C.,Hammock, B.D., 2004. Identification, expression and purifica-tion of a pyrethroid hydrolyzing carboxylesterase from mouseliver microsomes. J. Biol. Chem. 279, 29863–29869.
Straus, D.L., Schlenk, D., Chambers, J.E., 2000. Hepatic microso-mal desulfuration and dearylation of chlorpyrifos and parathionin fingerling channel catfish: lack of effect from Aroclor 1254.Aquat. Toxicol. 50, 141–151.
Sturm, A., Wogram, J., Segner, H., Liess, M., 2000. Differ-ent sensitivity to organophosphates of acetylcholinesterase and
-l.
Shate
T ein,anliver
T lismby
U.S. EPA, 1985. Short-term methods for estimating the chronictoxicity of effluents and receiving waters to freshwater organ-isms. EPA/600/4/85/014. Environmental Protection Agency, pp.58–75.
Van Dolah, R.F., Maier, P.P., Fulton, M.H., Scott, G.I., 1997. Compar-ison of azinphosmethyl toxicity to juvenile red drum (Sciaenopsocellatus) and the mummichog (Fundulus heteroclitus). Environ.Toxicol. Chem. 16, 1488–1493.
Werner, I., Deanovic, L.A., Connor, V., de Vlaming, V., Bailey,H.C., Hinton, D.E., 2000. Insecticide-caused toxicity toCeri-odaphnia dubia(Cladocera) in the Sacramento-San JoaquinRiver Delta, California, USA. Environ. Toxicol. Chem. 19, 215–227.
Werner, I., Deanovic, L.A., Hinton, D.E., Henderson, J.D., deOliveira, G.H., Wilson, B.W., Krueger, W., Wallender, W.W.,Oliver, M.N., Zalom, F.G., 2002. Toxicity of stormwater runoffafter dormant spray application of diazinon and esfenvalerate(Asana) in a French prune orchard, Glenn county, California,USA. Bull. Environ. Contam. Toxicol. 68, 29–36.
Wheelock, C.E., Miller, J.L., Miller, M.G., Shan, G., Gee, S.J., Ham-mock, B.D., 2004. Development of toxicity identification eval-uation (TIE) procedures for pyrethroid detection using esteraseactivity. Environ. Toxicol. Chem. 11, 2699–2708.
Wheelock, C.E., Wheelock,A.M., Zhang, R., Stok, J.E., LeValley, S.E., Green, C.E., Hammock, B.D., 2003. Evalua-tion of �-cyanoesters as fluorescent substrates for examin-ing interindividual variation in general and pyrethroid-selectiveesterases in human liver microsomes. Anal. Biochem. 315, 208–222.
Wheelock, C.E., Shan, G., Ottea, J., 2005. Overview of car-ides.
W 00.a
570.W on
0,
W ectsrase,sol.
butyrlcholinesterase from three-spined stickleback (Gasterosteus aculeatus) application in biomonitoring. Environ. ToxicoChem. 19, 1607–1615.
ymons, P.E.K., 1973. Behavior of young Atlantic salmon (Salmosalar) exposed to or force-fed fenitrothion, an organophospinsecticide. J. Fish. Res. Board Can. 30, 651–655.
ang, J., Cao, Y., Rose, R.L., Brimfield, A.A., Dai, D., GoldstJ.A., Hodgson, E., 2001. Metabolism of chlorpyrifos by humcytochrome P450 isoforms and human, mouse, and ratmicrosomes. Drug Metab. Dispos. 29, 1201–1204.
ang, J., Cao, Y., Rose, R.L., Hodgson, E., 2002. In vitro metaboof carbaryl by human cytochrome P450 and its inhibitionchlorpyrifos. Chem. Biol. Interact. 141, 229–241.
boxylesterases and their role in the metabolism of insecticJ. Pestic. Sci. 30, 75–83.
hyte, J.J., Jung, R.E., Schmitt, C.J., Tillitt, D.E., 20Ethoxyresorufin-O-deethylase (EROD) activity in fish asbiomarker of chemical exposure. Crit. Rev. Toxicol. 30, 347–
ildish, D.J., Lister, N.A., 1973. Biological effects of fenitrothiin the diet of brook trout. Bull. Environ. Contam. Toxicol. 1333–339.
ogram, J., Sturm, A., Segner, H., Liess, M., 2001. Effof parathion on acetylcholinesterase, butyrylcholinesteand carboxylesterase in three-spined stickleback (Gasterosteuaculeatus) following short-term exposure. Environ. ToxicChem. 20, 1528–1531.