Inducing Effect of Diamines on Transcription of the Cephamycin CGenes from the lat and pcbAB Promoters
in Nocardia lactamduransANA LUCIA LEITAO,1 FRANCISCO J. ENGUITA,2 JUAN LUIS DE LA FUENTE,2
PALOMA LIRAS,1,2 AND JUAN F. MARTIN1,2*
Institute of Biotechnology, 24006 Leon,1 and Area of Microbiology, Faculty of Biology,University of Leon, 24071 Leon,2 Spain
Received 10 November 1998/Accepted 25 January 1999
The diamines putrescine, cadaverine, and diaminopropane stimulate cephamycin biosynthesis in Nocardialactamdurans, in shake flasks and fermentors, without altering cell growth. Intracellular levels of the P7 protein(a component of the methoxylation system involved in cephamycin biosynthesis) were increased by diamin-opropane, as shown by immunoblotting studies. Lysine-6-aminotransferase and piperideine-6-carboxylatedehydrogenase activities involved in biosynthesis of the a-aminoadipic acid precursor were also greatlystimulated. The diamine stimulatory effect is exerted at the transcriptional level, as shown by low-resolutionS1 protection studies. The transcript corresponding to the pcbAB gene and to a lesser extent also the lattranscript were significantly increased in diaminopropane-supplemented cultures, whereas transcription fromthe cefD promoter was not affected. Coupling of the lat and pcbAB promoters to the reporter xylE gene showedthat expression from the lat and pcbAB promoters was increased by addition of diaminopropane in Streptomyceslividans. Intracellular accumulation of diamines in Nocardia may be a signal to trigger antibiotic production.
Polyamines are polycationic compounds that have a varietyof physiological effects in microorganisms and animal cells. Ingram-negative bacteria such as Escherichia coli, polyamineshave been shown to play an important role in chromosomestabilization by compacting DNA and neutralizing negativecharges (28). Polyamines affect the DNA synthesis by increas-ing the movement rate of the DNA replication fork (29). Fur-thermore, polyamines also stabilize functional ribosome struc-tures controlling the fidelity of the translation process,including specific steps such as aminoacyl-tRNA synthesis,binding, and initiation or elongation of the polypeptide chain(24). These compounds also appear to have a critical functionin membrane stability, suggested by their ability to decreasethe lysis of spheroplasts and protoplasts diluted in hypotonicmedia (30).
The molecule of cephamycin C is synthesized in Nocardialactamdurans from the three precursor amino acids, L-a-ami-noadipic acid, L-cysteine, and L-valine, which are initially con-densed to form the tripeptide d-(L-a-aminoadipyl)-L-cysteinyl-D-valine (LLD-ACV) by the large ACV synthetase multienzyme(4, 5). The tripeptide is later cyclized to isopenicillin N (7), andthis penicillin intermediate is converted into cephamycin C bya series of late modification reactions (1, 23). The N. lactam-durans genes encoding cephamycin biosynthesis enzymes havebeen cloned and sequenced and shown to be clustered (6) in a30-kb region of the genome of this microorganism (22). Thegenes of the cephamycin cluster are expressed as polycistronicmRNAs from three promoters, latp, pcbABp, and cefDp (11).
Production of cephamycin C by N. lactamdurans has beenimproved by strain selection and medium optimization. In-amine and Birmbaum (14) claimed in a Merck, Sharp &Dohme patent that cephamycin production in N. lactamdurans
is stimulated by diamines, although the mechanism of thisstimulation remains unknown.
Positive regulatory factors of antibiotic biosynthesis are ofgreat interest. The diamine effect on cephamycin biosynthesismight be exerted at the transcriptional level, or it might be dueto stimulation of the conversion of lysine into a-aminoadipicacid by the action of lysine-6-aminotransferase (5) and piperi-deine-6-carboxylate (P6C) dehydrogenase (8, 25).
In this article, we provide evidence showing that diamin-opropane and other diamines increase the levels of severalcephamycin biosynthetic enzymes by inducing transcription ofthe early cephamycin biosynthetic genes in N. lactamdurans.The inducer effect is clearly observed on transcription fromlatp and pcbABp, the two early promoters of the cephamycingene cluster.
MATERIALS AND METHODS
Microorganisms and culture conditions. N. lactamdurans MA4213, an im-proved cephamycin C producer strain (Merck, Sharp & Dohme, Rahway, N.J.)was used in this study. Mycelium was kept frozen in 20% glycerol at 220°C. Seedand antibiotic production cultures were grown in complex NYG medium (13).Triple-baffled flasks (500 ml) containing 100 ml of NYG medium were inoculatedwith 10 ml of a 48-h seed culture in the same medium and incubated at 24.5°Con a rotary shaker (New Brunswick Scientific) at 250 rpm. Batch fermentationswere carried out in NYG medium in a Biostat Q fermentor (Braun Biotech,Barcelona, Spain) with four identical vessels of 500 ml. The fermentors, contain-ing 270 ml of NYG medium, were inoculated with 1% (vol/vol) 48-h seed cultureand incubated at 24.5°C with stirring at 400 rpm and sparging with air at 0.15liter/min. Foaming was controlled by addition of antifoam MA24 DF 7960(Braun Biotech).
In experiments to study the effect exerted by different diamines on antibioticproduction, the NYG medium was supplemented with diaminopropane, pu-trescine, or cadaverine, at concentrations ranging from 0 to 5 g/liter.
Growth was estimated as cell DNA content, using the diphenylamine reagent(2) and salmon sperm DNA standard as described previously (18).
Antibiotic assays. Cephamycin C was routinely measured by bioassay using theagar diffusion method with E. coli ESS-2231 as an indicator strain. In addition,cephamycin C and four of its biosynthetic pathway intermediates were analyzedby high-pressure liquid chromatography (HPLC) as described elsewhere (17).Under these conditions, the retention times for these compounds were as fol-lows: deacetylcephalosporin C, 4 min; cephamycin C, 4.5 min; deacetoxycepha-
* Corresponding author. Mailing address: Area of Microbiology,Faculty of Biology, University of Leon, 24071 Leon, Spain. Phone:(34-987) 291505. Fax: (34-987) 291506. E-mail: [email protected].
losporin C, 10 min; 7-hydroxycephalosporin C, 14 min; and 7-methoxycephalos-porin C, 18 min.
Lysine-6-aminotransferase and P6C dehydrogenase activities. Lysine-6-ami-notransferase and P6C dehydrogenase activities were assayed in extracts of N.lactamdurans as described previously (7, 8). One unit of lysine-6-aminotransfer-ase or P6C dehydrogenase was defined as the activity that forms 1 nanomol ofproduct (P6C or NADH, respectively) per min.
SDS-PAGE and immunoblotting. Sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) analysis of proteins was performed as described byLaemmli (16), using a continuous buffer system (50 mM Tris, 150 mM glycine).
The P7 component of the methoxylation system (12) was detected by immu-noblotting. After SDS-PAGE, the proteins were transferred to a polyvinylidenedifluoride membrane (Immobilon; Millipore) in a semidry electroblotting system(LKB), using a buffer containing 50 mM Tris, 100 mM glycine, and 20% meth-anol, at a constant current of 0.8 mA/cm2. Free binding sites in the membraneswere blocked with 1% bovine serum albumin for 1 h at room temperature withgentle shaking. The membranes were then treated with the corresponding anti-bodies in a 50 mM sodium phosphate buffer (pH 7.6) containing 300 mM NaClfor 3 h at room temperature with shaking. The membranes were washed for 15min with 1 M NaCl and then treated with a commercial mouse anti-immuno-globulin G–alkaline phosphatase conjugate (Promega) at a concentration of 15U/ml in the same buffer for 1 h. After another membrane wash with 1 M NaClfor 15 min, the bands were developed with a commercial stabilized substratesolution of alkaline phosphatase (Promega) at room temperature until colorappeared.
RNA purification. N. lactamdurans MA4213 total RNA from different fermen-tation conditions was isolated by the guanidinium salt protocol (19), with minormodifications. The cells were harvested by centrifugation at 10,000 3 g at 4°C for15 min. The pellet was rapidly frozen in liquid nitrogen, and the cells weredisrupted in a cold mortar with alumina. The disrupted cells were transferred toa clean tube and treated with equal volumes of phenol and ITCG solution (2 Mguanidinium isothiocyanate, 100 mM Tris-HCl [pH 7.6], 50 mM EDTA, 100 mMlauryl sarcosine, 50 mM b-mercaptoethanol). After incubation in a water bath at55°C for 30 min, the samples were extracted repeatedly with 1 volume of phenol-chloroform solution until the aqueous phase was clean. The RNA was precipi-tated with 0.3 volume of 8 M LiCl solution, and its purity was analyzed by UVspectrophotometry.
Low-resolution S1 nuclease RNA protection assays. Low-resolution S1 pro-tection assays were carried out as described by Sambrook et al. (26), using 250 mgof total RNA and 2 mg of the selected DNA probe for each reaction. Hybrid-izations between the RNA and DNA probes were performed at 68°C for 15 h,and the RNA-DNA hybrids were treated with S1 nuclease at 37°C for 1 h, using250 U of the commercial enzyme (Boehringer Mannheim) per ml. After S1nuclease digestion, the hybrids were precipitated with ethanol, electrophoresedin an agarose gel, and transferred to a nylon membrane. The membranes werehybridized at 68°C with the same digoxigenin labeled-DNA probe used for theprotection assay in a buffer containing 53 SSC (13 SSC is 0.15 M NaCl plus0.015 M sodium citrate), 0.1% (wt/vol) lauryl-sarcosine, and 0.02% SDS. Positivehybridization bands were visualized with an alkaline phosphatase-conjugatedantidigoxigenin antibody and the corresponding commercial detection system forthe conjugated enzyme (Boehringer Mannheim).
Coupling to the xylE reporter gene. To study if expression from latp andpcbABp was regulated by diaminopropane, DNA fragments containing each
promoter (11) were coupled in the appropriate orientation to the xylE in thepromoter-probe vector pIJ4083 (3).
RESULTS
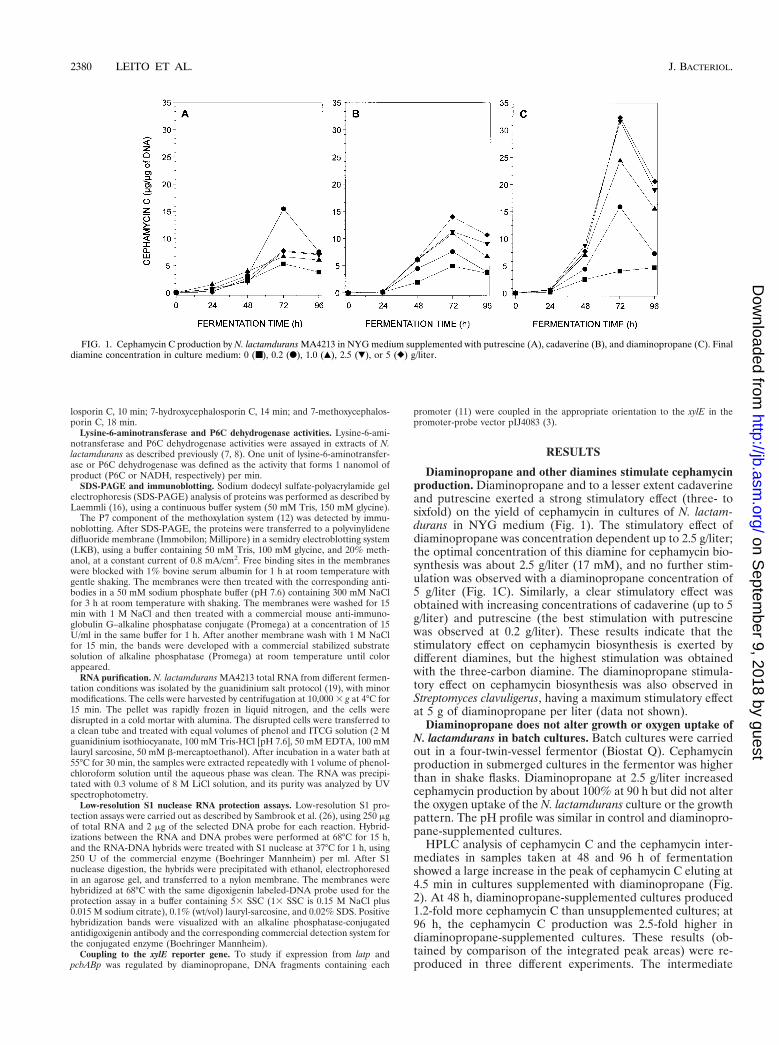
Diaminopropane and other diamines stimulate cephamycinproduction. Diaminopropane and to a lesser extent cadaverineand putrescine exerted a strong stimulatory effect (three- tosixfold) on the yield of cephamycin in cultures of N. lactam-durans in NYG medium (Fig. 1). The stimulatory effect ofdiaminopropane was concentration dependent up to 2.5 g/liter;the optimal concentration of this diamine for cephamycin bio-synthesis was about 2.5 g/liter (17 mM), and no further stim-ulation was observed with a diaminopropane concentration of5 g/liter (Fig. 1C). Similarly, a clear stimulatory effect wasobtained with increasing concentrations of cadaverine (up to 5g/liter) and putrescine (the best stimulation with putrescinewas observed at 0.2 g/liter). These results indicate that thestimulatory effect on cephamycin biosynthesis is exerted bydifferent diamines, but the highest stimulation was obtainedwith the three-carbon diamine. The diaminopropane stimula-tory effect on cephamycin biosynthesis was also observed inStreptomyces clavuligerus, having a maximum stimulatory effectat 5 g of diaminopropane per liter (data not shown).
Diaminopropane does not alter growth or oxygen uptake ofN. lactamdurans in batch cultures. Batch cultures were carriedout in a four-twin-vessel fermentor (Biostat Q). Cephamycinproduction in submerged cultures in the fermentor was higherthan in shake flasks. Diaminopropane at 2.5 g/liter increasedcephamycin production by about 100% at 90 h but did not alterthe oxygen uptake of the N. lactamdurans culture or the growthpattern. The pH profile was similar in control and diaminopro-pane-supplemented cultures.
HPLC analysis of cephamycin C and the cephamycin inter-mediates in samples taken at 48 and 96 h of fermentationshowed a large increase in the peak of cephamycin C eluting at4.5 min in cultures supplemented with diaminopropane (Fig.2). At 48 h, diaminopropane-supplemented cultures produced1.2-fold more cephamycin C than unsupplemented cultures; at96 h, the cephamycin C production was 2.5-fold higher indiaminopropane-supplemented cultures. These results (ob-tained by comparison of the integrated peak areas) were re-produced in three different experiments. The intermediate
FIG. 1. Cephamycin C production by N. lactamdurans MA4213 in NYG medium supplemented with putrescine (A), cadaverine (B), and diaminopropane (C). Finaldiamine concentration in culture medium: 0 (■), 0.2 (F), 1.0 (Œ), 2.5 (�), or 5 (}) g/liter.
deacetylcephalosporin C (with a retention time of 4 min) wasdetected at low levels in control cultures at 48 and 96 h but wasnot found in diaminopropane-supplemented cultures, indicat-ing that deacetylcephalosporin C is more efficiently convertedto cephamycin C in supplemented cultures.
Diaminopropane increases lysine-6-aminotransferase andP6C dehydrogenase. The enzyme levels of lysine-6-aminotrans-ferase and P6C dehydrogenase (the first and second enzymesof the cephamycin pathway) were significantly stimulated incultures of N. lactamdurans supplemented with diaminopro-pane. P6C dehydrogenase activity was increased about sixfoldin diaminopropane-supplemented cultures at 48 h, and theactivity was kept at high levels at 72 h compared to control(unsupplemented) cultures (Fig. 3). The drastic increase inP6C dehydrogenase levels in diaminopropane-supplementedcultures suggests that the dehydrogenation of P6C to givea-aminoadipic acid may be a bottleneck in cephamycin biosyn-thesis and that stimulation of the P6C dehydrogenase levelsmay remove one of the bottlenecks of cephamycin biosynthe-sis.
Diaminopropane increases the protein levels of the P7 com-ponent of the cephamycin methoxylation system. The availabil-ity of antibodies against the P7 component of the cephamycinmethoxylation system provided a useful tool for determiningwhether the stimulatory effect of diaminopropane on cepha-mycin biosynthesis was exerted on the synthesis of these pro-teins or whether it is simply due to stimulation of the activity ofsome of the biosynthetic enzymes.
Results of the immunoblotting (Fig. 4) showed that the levelof the P7 protein at 36, 48, 60, and 72 h was significantly higherin diaminopropane-supplemented cultures than in control cul-tures.
The diaminopropane effect is exerted at the transcriptionallevel. The stimulation of the P7 protein of the methoxylationsystem and the increased levels of lysine-6-aminotransferasewere consistent with the fact that these proteins are synthe-sized from long polycistronic mRNAs that include the lysine-6-aminotransferase, ACV synthetase, isopenicillin N synthase,methylcephem-39 hydroxylase, the P7 and P8 proteins (me-
FIG. 2. HPLC analysis of antibiotic production by diaminopropane (DAP)-supplemented and unsupplemented cultures of N. lactamdurans MA4213. Thecephamycin C peak eluted at a retention time of 4.5 min, and the deacetylcepha-losporin C peak eluted at 4 min.
FIG. 3. Stimulatory effect of diaminopropane on lysine-6-aminotransferaseand P6C dehydrogenase in N. lactamdurans MA4213. (■) Control (unsupple-mented) cultures; (Œ) diaminopropane (2.5 g/liter)-supplemented cultures.
FIG. 4. Immunoblotting detection of the P7 protein in control and diamin-opropane-supplemented N. lactamdurans MA4213 cultures. Numbers above thelanes indicate culture times (hours). Lanes D36 to D72 represent samples fromdiaminopropane-supplemented cultures taken at 36 to 72 h.
VOL. 181, 1999 INDUCTION OF CEPHAMYCIN BIOSYNTHESIS BY DIAMINES 2381
thoxylation system), and the 79-cephem carbamoyltransferase(11). The pcd gene (encoding P6C dehydrogenase), recentlycloned from S. clavuligerus (25), has not been located in the N.lactamdurans genome.
As shown in Fig. 5A, S1 nuclease protection studies indi-cated that the same fragment of the polycistronic transcriptwas protected with a 1,010-bp probe (internal to lat) in controland diaminopropane-supplemented cultures, but the intensityof the hybridization was 21% higher in RNA preparationsfrom diaminopropane-supplemented cultures. A high stimula-tion by diaminopropane (79%) was also observed when a1,083-bp probe corresponding to the 59 end of the pcbAB genewas used as the probe in S1 protection experiments (Fig. 5B).This result suggests that expression of the long transcript cor-responding to pcbAB and the late genes (11) is particularlyenhanced by diaminopropane. However, the band correspond-ing to the transcript of the cefD and cefE genes (protected bythe 1,392-bp probe) showed no difference in intensity, suggest-ing that expression from the cefD promoter is not stimulated bydiaminopropane.
Diaminopropane stimulates transcription initiation at thelat and pcbAB promoters. To confirm that the diaminopropaneeffect on transcription of the polycistronic mRNAs corre-sponding to lat, pcbAB, and the late genes of the pathway wasexerted at the transcriptional level, the two known promotersof this region (latp and pcbABp) were coupled to the xylEreporter gene. As shown in Fig. 6, a moderate catechol oxy-genase activity (yellow color) was observed in S. lividans trans-formants with the xylE gene under either the lat or pcbABpromoter. No formation of pigment was observed in S. lividanscultures transformed with the control plasmid (pIJ4083) with-out insert.
The pigment formed by the catechol oxygenase increasedconsiderably (to a reddish color) in plates supplemented withdiaminopropane (5 g/liter). The color formed in S. lividanstransformed with pIJ4083 under pcbABp was particularlystrong after only 30 min of incubation, whereas expressionfrom latp required a longer incubation time (24 h) to obtain thesame color. Although no precise quantification of reporter
gene activity is possible in solid medium, the increase in cate-chol oxygenase activity with respect to the promoterlesspIJ4083 constructions is at least twofold, in good correlationwith the stimulation observed in RNA studies.
DISCUSSION
Although the importance of diamines in biological systems iswell documented (9, 15), very little is known about their mech-anism of action, particularly with respect to the biosynthesis ofsecondary metabolites. In E. coli, putrescine, cadaverine, andspermidine are formed through the action of basic amino aciddecarboxylases (30). Recently Sekowska et al. (28) reportedthat in Bacillus subtilis there is a single pathway to polyaminesstarting from arginine with agmatine as an intermediate. How-ever, it is unknown whether a similar biosynthetic pathwayoccurs in Nocardia and other gram-positive bacteria.
Polyamines are used as nitrogen (and carbon) sources bydifferent microorganisms. In S. lividans, lysine is known to bedecarboxylated to cadaverine that is later further oxidized to2-ketoglutarate (21). However, in the b-lactam producers N.lactamdurans and S. clavuligerus, lysine is not significantly de-graded to cadaverine (10) but instead is converted to a-ami-noadipic acid by the lysine-6-aminotransferase (20).
In the present work, we report that the biosynthesis ofcephamycin C was stimulated by several diamines, in a mech-anism that involves a positive effect on the levels of somebiosynthetic enzymes, including lysine-6-aminotransferase andP6C dehydrogenase and the P7 protein of the methoxylationsystem. Unfortunately, the gene encoding the N. lactamduransP6C decarboxylase is not located in the cephamycin cluster andhas not been cloned. Therefore, we cannot conclude whetherthe substantial stimulation of the P6C dehydrogenase by dia-minopropane is exerted at the transcriptional level. Our resultsclearly indicate that diaminopropane acts by inducing expres-sion of the mRNAs for cephamycin biosynthesis at the pcbABpromoter and to a lesser extent at the lat promoter. These twopromoters are used for expressing the early (lat, pcbAB, andpcbC) and late (cefF, cmcH, cmcI, and cmcJ) genes of the
FIG. 5. Low-resolution S1 protection study of selected regions of the cephamycin C cluster from N. lactamdurans, in diaminopropane-supplemented (lanes 2) andunsupplemented (lanes 1) cultures. Total RNA was extracted from cells harvested at 48 h and hybridized with the following probes: (A) SalI-NotI fragment (1,010 bp),internal to the lat gene; (B) PstI fragment (1,083 bp), corresponding to the initial region of the pcbAB gene; (C) XmnI-BamHI fragment (1,392 bp), covering part ofcefD and cefE genes. A map of the cephamycin biosynthetic genes and the corresponding transcriptional units is shown at the top. The protected bands are indicatedby arrows. The results are very similar in repeated experiments with an approximate 5% error in band intensity, using always the same RNA/probe DNA ratio.
cephamycin pathway (11). Interestingly, no significant stimu-lation of the expression from the cefDE promoter (which islocated in opposite orientation to the lat and pcbAB promot-ers) was observed.
The stimulatory effect could be due to a stabilization of
mRNA. However, the finding that diaminopropane also stim-ulates expression from the lat and pcbAB promoters whencoupled to the xylE reporter gene strongly supports an induc-ing effect on gene expression.
To some extent, the enhancing effect of diaminopropane on
FIG. 6. Effect of diaminopropane on xylE gene (encoding catechol oxygenase) from promoters pcbABp (A) and latp (B) of the N. lactamdurans cephamycin Ccluster. S. lividans containing the indicated plasmids was grown on Trypticase soy agar plates with 25 mg of thiostrepton per ml for 36 h, and the plates were sprayedwith 1 M catechol solution for color developing. Right, control (unsupplemented) plates; left, diaminopropane (DAP) (2.5 g/liter)-supplemented plates. pcbABp isindicated as pACVs.
VOL. 181, 1999 INDUCTION OF CEPHAMYCIN BIOSYNTHESIS BY DIAMINES 2383
transcription is promoter specific. The strong induction ofpcbABp by diaminopropane makes this promoter useful forinducible expression at will of genes in N. lactamdurans, in S.lividans, and probably in other actinomycetes.
Induction by the diamines following their accumulation maybe a metabolic signal to trigger antibiotic production. Intracel-lular accumulation of diamines may regulate the lysine cata-bolic pathway, therefore affecting the lysine supply for cepha-mycin biosynthesis. However, it is unclear whether there is adiamine biosynthetic pathway in N. lactamdurans. Alterna-tively, the presence of extracellular diamines may indicate thatdiamine-producing bacteria are growing and competing fornutrients in the Nocardia habitat, signaling that cephamycinbiosynthesis must be triggered as a defense mechanism.
Addition of diamines at high concentration may provoke astress response resulting in up-regulation of stress-controlledpromoters through sS. However, no evidence for or againstthis hypothesis is available at this time.
ACKNOWLEDGMENTS
This project was supported by grants BIO97-0289-CO2 from theCICYT and by a Generic Project of the Agencia de Desarrollo ofCastilla and Leon (10-2/98/LE/0003). A. L. Leitao was supported by aPRAXIS XXI fellowship (BD13931/97) (Portugal), and F. J. Enguitareceived a fellowship from the PFPI Program, Ministry of Educationand Culture (Spain).
We thank M. Corrales, M. Mediavilla, and R. Barrientos for excel-lent technical assistance.
REFERENCES
1. Aharonowitz, Y., G. Cohen, and J. F. Martın. 1992. Penicillin and cephalo-sporin biosynthetic genes: structure, organization, regulation and evolution.Annu. Rev. Microbiol. 46:461–495.
2. Burton, K. 1956. A study of the conditions and mechanism of the diphe-nylamine reaction for the colorimetric estimation of deoxyribonucleic acid.Biochem. J. 62:315–323.
3. Clayton, T. M., and M. J. Bibb. 1990. Streptomyces promoter-probe plasmidsthat utilize the xylE gene of Pseudomonas putida. Nucleic Acids Res. 18:1077.
4. Coque, J. J. R., J. L. de la Fuente, P. Liras, and J. F. Martın. 1996.Overexpression of the Nocardia lactamdurans a-aminoadipyl-cysteinyl-valinesynthetase in Streptomyces lividans. The purified multienzyme uses cystathi-onine and 6-oxopiperidine 2-carboxylate as substrates for synthesis of thetripeptide. Eur. J. Biochem. 242:264–270.
5. Coque, J. J. R., P. Liras, L. Laiz, and J. F. Martın. 1991. A gene encodinglysine 6-aminotransferase, which forms the b-lactam precursor a-aminoadi-pic acid, is located in the cluster of cephamycin biosynthetic genes in No-cardia lactamdurans. J. Bacteriol. 173:6258–6264.
6. Coque, J. J. R., P. Liras, and J. F. Martın. 1993. Genes for a b-lactamase, apenicillin-binding protein and a transmembrane protein are clustered withthe cephamycin biosynthetic genes in Nocardia lactamdurans. EMBO J.12:631–639.
7. Coque, J. J. R., J. F. Martın, J. G. Calzada, and P. Liras. 1991. Thecephamycin biosynthetic genes pcbAB, encoding a large multidomain pep-tide synthetase, and pcbC of Nocardia lactamdurans are clustered together inan organization different from Acremonium chrysogenum and Penicilliumchrysogenum. Mol. Microbiol. 5:1125–1133.
8. De la Fuente, J. L., A. Rumbero, J. F. Martın, and P. Liras. 1997. D-1-Piperideine-6-carboxylate dehydrogenase, a new enzyme that forms a-ami-noadipate in Streptomyces clavuligerus and other cephamycin C-producingactinomycetes. Biochem. J. 327:59–64.
9. De la Vega, A. L., and A. H. Delcour. 1996. Polyamines decrease Escherichiacoli outer membrane permeability. J. Bacteriol. 178:3715–3721.
10. Domınguez, H., and J. F. Martın. Unpublished results.11. Enguita, F. J., J. J. R. Coque, P. Liras, and J. F. Martın. 1998. The nine
genes of the Nocardia lactamdurans cephamycin cluster are transcribed intolarge mRNAs from three promoters, two of them located in a bidirectionalpromoter region. J. Bacteriol. 180:5489–5494.
12. Enguita, F. J., P. Liras, A. L. Leitao, and J. F. Martın. 1996. Interaction ofthe two proteins of the methoxylation system involved in cephamycin Cbiosynthesis. Immunoaffinity, protein cross-linking, and fluorescence spec-troscopy studies. J. Biol. Chem. 271:33225–33230.
13. Ginther, C. L. 1979. Sporulation and serine protease production by Strepto-myces lactamdurans. Antimicrob. Agents Chemother. 15:522–526.
14. Inamine, E., and J. Birnbaum. August 1976. U.S. patent 3,977,942.15. Koshi, P., and M. Vaara. 1991. Polyamines as constituents of the outer
membranes of Escherichia coli and Salmonella tryphimurium. J. Bacteriol.173:3695–3699.
16. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
17. Leitao, A. L., F. J. Enguita, and J. F. Martın. 1997. Improved oxygen transferin cultures of Nocardia lactamdurans maintains the cephamycin biosyntheticproteins for prolonged times and enhances the conversion of deacetylcepha-losporin into cephamycin C. J. Biotechnol. 58:39–50.
18. Leitao, A. L., F. J. Enguita, J. L. De la Fuente, P. Liras, and J. F. Martın.1996. Allophane increases the protein levels of several cephamycin biosyn-thetic enzymes in Nocardia lactamdurans. Microbiology 142:3399–3406.
19. MacDonald, R. J., G. H. Swift, A. E. Przybyla, and J. M. Chirgwin. 1987.Isolation of RNA using guanidinium salts. Methods Enzymol. 152:219–227.
20. Madduri, K., S. Shapiro, A. C. DeMarco, R. L. White, C. Stuttard, and L. C.Vining. 1991. Lysine catabolism and a-aminoadipate synthesis in Streptomy-ces clavuligerus. Appl. Microbiol. Biotechnol. 35:358–363.
21. Madduri, K., C. Stuttard, and L. C. Vining. 1989. Lysine catabolism inStreptomyces sp. is primarily through cadaverine: b-lactam producers alsomake a-aminoadipate. J. Bacteriol. 171:299–302.
22. Martın, J. F. 1998. New aspects of genes and enzymes for b-lactam antibioticbiosynthesis. Appl. Microbiol. Biotechnol. 50:1–15.
23. Martın, J. F., and P. Liras. 1989. Organization and expression of genesinvolved in the biosynthesis of antibiotics and other secondary metabolites.Annu. Rev. Microbiol. 43:173–206.
24. Mitsui, K., K. Igarashi, T. Kakegawa, and S. Hirose. 1984. Preferentialstimulation of the in vivo synthesis of a protein by polyamines in Escherichiacoli: purification and properties of the specific protein. Biochemistry 23:2679–2683.
25. Perez-Llarena, F. J., A. Rodrıguez-Garcıa, F. J. Enguita, J. F. Martın, and P.Liras. 1998. The pcd gene encoding piperideine-6-carboxylate dehydroge-nase involved in biosynthesis of a-aminoadipic acid is located in the cepha-mycin cluster of Streptomyces clavuligerus. J. Bacteriol. 180:4753–4756.
26. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
27. Sawada, Y., T. Konomi, N. A. Solomon, and A. L. Demain. 1980. Increase inactivity of b-lactam synthetases after growth of Cephalosporium acremoniumwith methionine or norleucine. FEMS Microbiol. Lett. 9:281–284.
28. Sekowska, A., P. Bertin, and A. Danchin. 1998. Characterization of poly-amine synthesis pathway in Bacillus subtilis 168. Mol. Microbiol. 29:851–858.
29. Tabor, C. W., and H. Tabor. 1984. Polyamines. Annu. Rev. Biochem. 53:749–790.
30. Tabor, C. W., and H. Tabor. 1985. Polyamines in microorganisms. Microbiol.Rev. 49:81–99.