The dependen e of ma rophage phago yti eÆ a y onantibody on entrationNata�sa Ma ura� Tong Zhangy Arturo CasadevallyzNovember 24, 2006
1Department of Mathemati s, Trinity University, One Trinity Pla e, San Antonio, TX 782122Albert Einstein College of Medi ine, 1300 Morris Park Ave., Bronx, NY 104613Corresponding author, Email: asadeva�ae om.yu.edu, Tel: (718) 430-36651
Abstra tMa rophages ingest the fungus Crypto o us neoformans only in the presen e of op-sonins, and this provides a remarkably lean system for the detailed analysis of phago- ytosis. This system is also unusual in that antibody-mediated phago ytosis involvesingestion through both F and omplement re eptors in the absen e of omplement.Mathemati al modeling was used to analyze and explain the experimental data thatma rophage phago yti index in reased with in reasing doses of antibody despite sat-urating on entrations and de lined at high on entrations. A model was developedthat explains the in rease in phago yti index with in reasing antibody doses, di�er-entiates among the ontributions from F and omplement re eptors, and providesa tool for estimating antibody on entrations that optimize eÆ a y of phago ytosis.Experimental results and model al ulations revealed that blo king of F re eptors byex ess antibody aused a redu tion in phago yti index, but in reased phago ytosisthrough omplement re eptors rapidly ompensated for this e�e t. At high antibody on entrations further redu tion in phago yti index was aused by interferen e with omplement re eptor ingestion as a onsequen e of saturation of the fungal apsule.The ability of our model to predi t the antibody dose dependen e of the ma rophagephago yti eÆ a y for C. neoformans strongly suggest that the major variables thatdetermine the eÆ a y of this pro ess have been identi�ed. The model predi ts thatthe aÆnity onstant of the opsoni antibody for the F re eptor and the asso iation-disso iation onstant of antibody from the mi robial antigen are riti al parametersdetermining the eÆ a y of phago ytosis. 2
Introdu tion1 Phago ytosis is a pro ess by whi h ertain types of ells are able to ingest parti les and2 mi robes. Some protozoa like amoebae use phago ytosis for the a quisition of food. Among3 the metazoa, many animals have spe ialized ells for host defense that an ingest and destroy4 mi robes. In mammals, tissue ma rophages are highly spe ialized ells that ingest, destroy5 and digest mi robes and present peptide antigens to lympho ytes. Ma rophage phago y-6 tosis is dependent on ellular re eptors and an be enhan ed by the presen e of antibody7 or omplement opsonins. In many infe tious diseases the produ tion of opsoni antibody8 is asso iated with immunity. Therefore this phylogeneti ally an ient pro ess represents a9 riti al omponent of host defense against mi robial infe tions.10 Crypto o us neoformans is a pathogeni yeast that is a relatively frequent ause of life-11 threatening meningoen ephalitis, espe ially in immuno ompromised individuals [10℄. This12 fungus is unusual in that it has a polysa haride apsule that is antiphago yti . Conse-13 quently, the intera tion of C. neoformans and ma rophages rarely results in phago ytosis14 unless spe i� antibody and/or omplement-derived opsonins are present. Phago ytosis of15 C. neoformans by ma rophages has been studied with ells derived from various sour es16 in luding the J774 murine ma rophage-like ell line [8℄. In this system the phago yti index17 was shown to depend on several variables in luding the on entration and type of opsonin,18 the size of the apsule expressed by the C. neoformans strain, and the relative ratio of19 ma rophages to yeast ells [8℄. This system has several advantages for the study of phago y-20 3
tosis in luding the fa t that yeast ells are relatively large and an be easily ounted by light1 mi ros opy and the qualitative out ome of the intera tion whereby there is no signi� ant2 phago ytosis in the absen e of opsonins. Hen e, it is possible to de�ne the variables that3 a�e t the out ome of the intera tion between C. neoformans and ma rophages in a manner4 that would be very diÆ ult for other mi robial-ma rophage systems.5 We are parti ularly interested in the me hanisms of antibody-mediated prote tion against6 C. neoformans and the relationship between antibody dose and prote tive eÆ a y. Passive7 immunization with antibody to the apsule is prote tive but administration of large amounts8 of antibody abrogates prote tion [13, 15℄, and an a tually enhan e the ourse of infe tion.9 This phenomenon has been alled a \prozone-like" e�e t. While studying the intera tion10 of ma rophages and C. neoformans in vitro we noted that the phago yti index de lines at11 higher antibody on entrations [13, 15℄. Given that this observation ould be asso iated with12 the prozone-like phenomenon observed in passive prote tion experiments we de ided to study13 in more detail and onstru t a mathemati al model of phago ytosis of C. neoformans that14 would allow us to better understand the ontribution of the various parameters to opsoni 15 eÆ a y.16 There have been several attempts to generate mathemati al models of phago ytosis in the17 literature [11, 16℄. Phago ytosis is an attra tive pro ess for mathemati al modeling be ause18 many of the variables are relatively well understood. However, none of the models available19 have addressed the riti al ontribution of opsonin on entration and the subje t was last20 4
investigated almost two de ades ago.1 We propose and test a mathemati al model based on understanding of the underlying2 prin iples and me hanisms of phago ytosis. We identify the main variables and parameters of3 the model and analyze their impa t on the out omes of our experiments. Our mathemati al4 des ription of phago ytosis is based on the di�erential equation whi h des ribes the rate of5 hange of the total number of ingested mi robes PI as a fun tion of the population PF of non-6 ingested (free) mi robes and the rate of phago ytosis rT : The rate of phago ytosis depends7 on the amount of antibody bound to C. neoformans apsule and the number of re eptors8 available and we use this model to explore the dependen e of the phago yti index on the9 on entration of free antibody by ombining mathemati al analysis and experimental work.10 We dis uss the issues that arise from the analysis of the model and propose experimental11 and theoreti al questions for further re�nement of the model.12Materials and methods13 Experimental methods14 Phago ytosis assays. The assay used to study phago ytosis involved minor modi� a-15 tions to the previously des ribed methods ([8, 9℄). For all experiments we used the J774.1616 murine ma rophage ell line, whi h faithfully reprodu es the intera tion of C. neoformans17 with murine ma rophages [6℄. Brie y, on the day prior to the phago ytosis experiment ap-18 proximately 5 � 104 J774.16 ells were added to 96 well polystyrene tissue ulture plates19 5
(BD Fal on, Franklin Lakes, NY) in media onsisting of 10% heat-ina tivated fetal bovine1 serum (Gemini Bio-Produ ts, Woodland, CA), 10 % NCTC-109 (Gib o, Grand Island, NY),2 and 1% non-essential amino a ids (Mediate h, Herndon, VA) in Dulbe os Modi�ed Eagle3 Media (Gib o) ontaining 50 U/ml of IFN- : After overnight in ubation at 37ÆC the media4 was removed, the monolayer was washed 3� with fresh media, and 105 C. neoformans ells5 were added in media. All experiments used C. neoformans strain 24067 (Ameri an Type6 Tissue Colle tion, Ro kville, MD). Mono lonal antibody (mAb) 18B7 is a murine IgG1 that7 binds the C. neoformans apsular omponent Glu uronoxylomannan and has been exten-8 sively hara terized and used in a human lini al trial ([3, 5℄). mAb 18B7 was added to the9 C. neoformans suspension in variable amounts and the ma rophage-yeast suspension was10 in ubated for 2 h at 37ÆC. In this experimental system the ma rophages are immobilized11 at the bottom by virtue of their apa ity to grow as an adherent monolayer. The C. neo-12 formans is added in suspension, the yeast settle rapidly by gravity and phago ytosis begins13 within minutes. Afterwards the monolayer was washed 3� with phosphate bu�ered saline,14 �xed with old methanol and stained with Giemsa (Sigma-Aldri h, St. Louis, MO). The15 phago yti index was determined by ounting ingested yeast with an inverted mi ros ope at16 magni� ation 100 � q: Under these onditions it is very easy to distinguish atta hed from17 ingested yeast ells, sin e the latter reside in dis ernable intra ellular va uoles. The phago-18 yti index in the various onditions was determined by ounting internalized yeast ells19 per 100 ma rophages in various �elds (4 �elds were ounted). In our system and ondition20 6
antibody-mediated opsonization results in almost omplete ingestion su h that the over-1 whelming majority of yeast ells are inside ma rophages. In prior studies we have validated2 our ability to distinguish between atta hed and ingested yeast ells using oures en e dyes3 that stain only atta hed yeasts. Antibody-mediated opsonization in this system is highly4 eÆ ient and leads to ingestion. In all onditions more than 90% of the atta hed yeast ells5 internalized irrespe tive of antibody on entration.6 In some experiments yeast ells were in ubated with 18B7 for an hour or an hour and a7 half to obtain a near-saturation o upan y of the yeast apsule before adding the antibody-8 oated ells to the ma rophage monolayer. In some experiments we blo ked the omplement9 re eptors (CR) by adding antibody to CD18, CD11b, and CD11 (BD Bios ien es Pharmi-10 gen, San Jose, CA), whi h are expressed by J774.16 ells ([14℄). For these experiments the11 above proto ol was modi�ed by in ubating the ma rophage monolayer with blo king anti-12 bodies (50 �g/ml) for 1 h prior to the phago ytosis experiment. After washing and addition13 of the C. neoformans suspension and mAb 18B7 blo king was maintained by in luding the14 blo king antibodies (10 �g/ml) in the ma rophage-yeast suspension. It is noteworthy that15 this proto ol was arrived at after modeling results revealed that in ubation of J774 ells with16 10 �g/ml of blo king antibody was not suÆ ient to blo k the omplement re eptors.17 Mathemati al modeling. Our mathemati al model of phago ytosis is based on the di�er-ential equation dPIdt = rTPF (E-1 )7
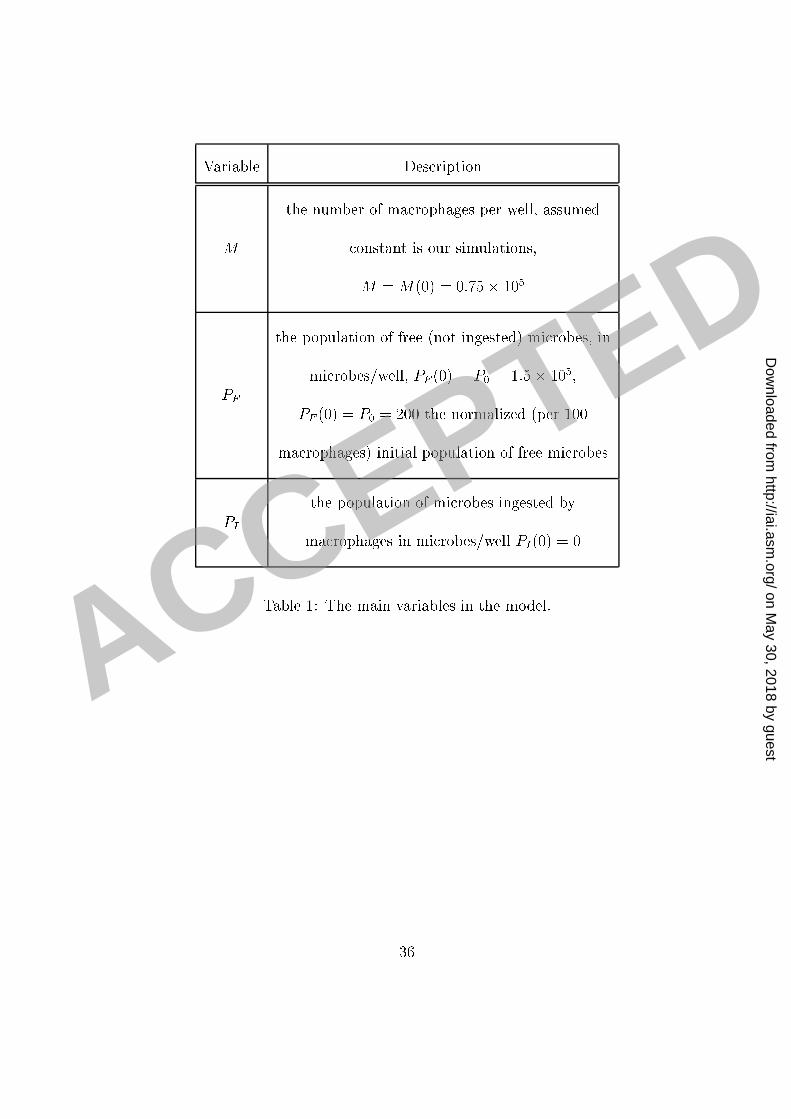
1 whi h des ribes the rate of hange of the total number of ingested mi robes PI as a fun tion2 of the population of free mi robes PF and the rate of phago ytosis rT . The detailed math-3 emati al analysis of pro esses involved in phago ytosis is given in the supplemental se tion4 where we onstru t a mathemati al model of this pro ess. The main variables in the model5 are listed and des ribed in Table 1. The main goal of this paper is to model the fun tion6 rT ; whi h we all the eÆ a y of phago ytosis and whi h depends on the amount of antibody7 bound to the C. neoformans apsule and the number of re eptors available.8 We built our mathemati al model of the phago yti eÆ a y on two main assumptions:9 the �rst assumption is that phago yti eÆ a y in reases as the number of binding sites on10 the C. neoformans apsule for the appropriate type of re eptors in reases and the se ond is11 that there is a maximum eÆ a y of phago ytosis for a given number of available re eptors.12 Both these assumptions were motivated and on�rmed by the experimental results.13 The simplest, and naturally the �rst, model to onsider is a model based on a linear14 in rease in the phago yti eÆ a y with the number of sites s on the C. neoformans ap-15 sule available to bind the appropriate re eptors on the ma rophage surfa e. This model is16 des ribed by the di�erential equation drT1ds = onst = k1; and is therefore based on the17 assumption that the rate at whi h the phago yti eÆ a y in reases does not depend on the18 amount of antibody already bound to the C. neoformans apsule. Moreover, the solution19 of this model, assuming rT1(0) = 0; is rT1 = k1s is proportional to the rate at whi h the20 8
antibody is binding to a given ( onstant) number of re eptors. Combining this model with1 the assumption that there is a maximum phago yti eÆ a y led to our �rst mathemati al2 model of phago yti eÆ a y:3 drT1ds = k1(RM1 � rT1) (E-2 )4 The onstant k1 is the initial growth rate of phago yti eÆ a y and the term (RM1 � rT1)5 represents the redu tion in the rate at whi h the eÆ a y of phago ytosis in reases as it6 approa hes the maximum phago yti eÆ a y RM1 for the given onditions. Both k1 and7 RM1 depend on the number of available re eptors and may also depend on the health of8 ma rophages and vary slightly depending on the experimental onditions.9 Our experimental results in the ase of fully blo ked omplement re eptors strongly sup-10 ported this model. The number of binding sites for F re eptors s is equal to the amount of11 antibody bound to the C. neoformans apsule AP and therefore the equation E-2 leads to12 the di�erential equation13 drT1dAP = k1(RM1 � rT1) (E-3 )14 whose solution is159
rT1 = RM1(1� e�k1AP ): (E-4 )1 The data of the phago yti eÆ a y versus the amount of antibody bound to the C. neofor-2 mans apsule indi ate that the same type of a model3 drT2ds = k2(RM2 � rT2) (E-5 )4 does not fully a ount for the in rease in the phago yti index when the omplement re ep-5 tors are involved. Consequently, to explain the eÆ a y of phago ytosis when the omplement6 re eptors are involved, we onsider a mathemati al model where the in rease in the phago-7 yti eÆ a y is a linear fun tion of the number of the omplement re eptor binding sites s8 on the C. neoformans apsule. The di�erential equation appli able in this ase is9 drT3ds = k3(R3 � rT3)s (E-6 )10 where s is the number of omplement re eptor binding sites on the C. neoformans apsule.11 This mathemati al model indi ates a possibility of ooperation among re eptors at high12 density of antibody bound to C. neoformans apsule. The general solution of the above13 di�erential equation is14 10
rT3 = R3 �Ke�k3s2=2; (E-7 )1 where K is a onstant. We ombined the two above models (E-5 and E-7) and obtained2 a mathemati al model that in orporates both e�e ts: the onstant in rease in phago yti 3 eÆ a y at lower on entrations and more rapid in rease in the eÆ a y of phago ytosis at4 higher density of antibody bound to C. neoformans apsule. We assumed that equation E-75 des ribes the rate of in rease of the maximum phago yti eÆ a y due to ooperation between6 re eptors. The onstant K was determined from the assumption that, when there is no7 ooperation (s = 0), the maximum phago yti eÆ a y is rT3 = R2M : We therefore obtained8 that rT3 = R2M (1+ R3R2M (1�e�k3s2=2):We repla ed the onstant maximum phago yti eÆ a y9 R2M in equation E-5 with rT3 and therefore modeled phago ytosis through omplement10 re eptors by the following equation11 rTC = (RM2(1� e�k2s))(1 +RM3(1� e�k3s2=2); (E-8 )12 where RM3 = R3R2M : Our experimental results also indi ated that, when both types of re-13 eptors are available, the phago ytosis through F and phago ytosis through omplement14 re eptors are not independent pro esses, that is, the umulative result is not simply the sum15 of their respe tive ontributions. Our model is set up so that this e�e t an be easily in or-16 11
porated: a natural extension of our model for phago ytosis through omplement re eptors1 is the following equation2 rT = �RM1(1� e�k1AP ) +RM2(1� e�k2s)�(1 +RM4(1� e�k4s2) (E-9 )3 where s is the number of omplement re eptor binding sites on the C. neoformans apsule.4 Note that we introdu ed RM4 and k4 in this model instead of RM3 and k3: While RM3 and5 k3 formally des ribed the e�e t of ooperation among omplement re eptors, RM4 and k46 a ount for the e�e t of ooperation among omplement re eptors as well as omplement7 and F re eptors. We assumed that the number of omplement re eptor binding sites on the8 C. neoformans apsule in reases quadrati ally with the amount of antibody bound to the9 apsule, rea hes a maximum and then de reases with binding of the additional antibody. The10 assumption of the quadrati in rease is justi�ed by the experimental results whi h show that11 the total in rease in phago yti eÆ a y is more rapid than the linear in rease in the number12 of binding sites s in model E-9 would imply. We were primarily interested in modeling13 the phago yti eÆ a y in the range of on entrations of free antibody where the number of14 omplement re eptor binding sites on C. neoformans apsule in reases. In that range our15 model leads to the following dependen e of the phago yti eÆ a y rT of phago ytosis on the16 amount of antibody bound to C. neoformans apsule1712
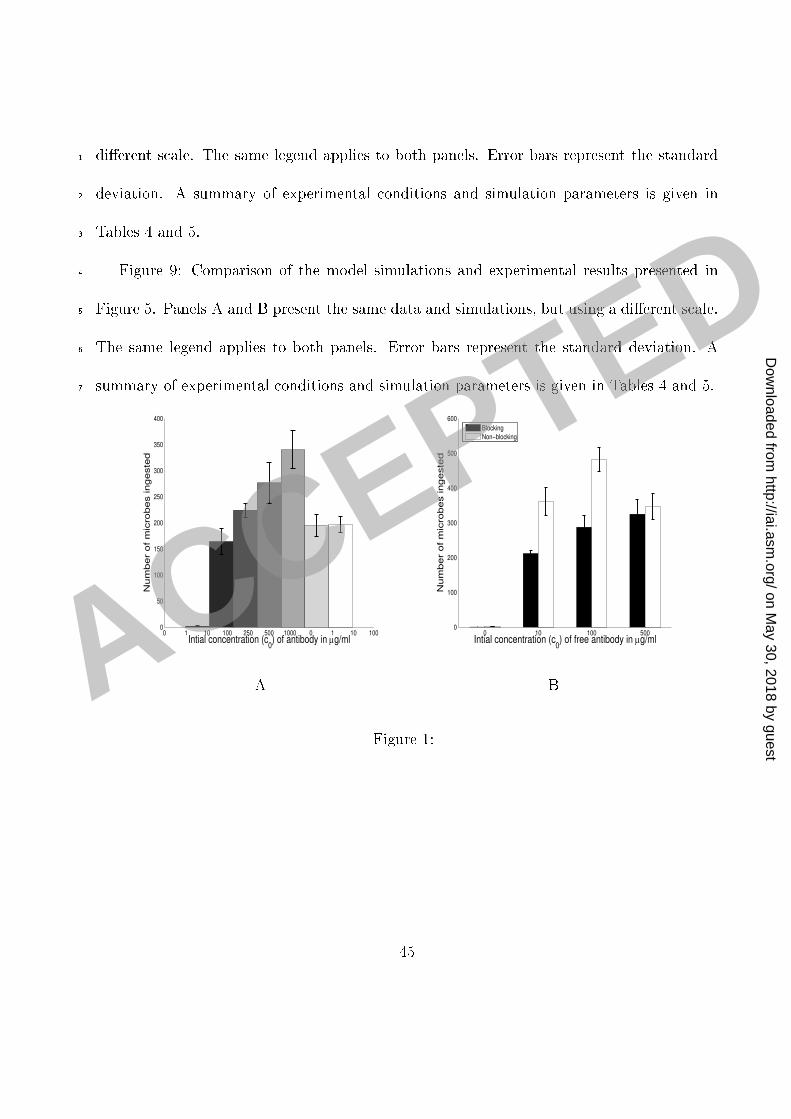
rT = �RM1(1� e�k1AP ) +RM2(1� e�k2A2P )�(1 +RM4(1� e�k4A4P ): (E-10 )Simulations of the model. We used MatLab 7.4 (The MathWorks, Nati k, MA) in simu-1 lations of our model.2Results3 Dependen e of phago ytosis on antibody on entration and time. The eÆ a y of phago yto-4 sis was studied as a fun tion of antibody on entration (Figure 1) and time (Figure 2) in5 the presen e and absen e of omplement re eptor blo kade. Although this system does not6 in lude omplement opsonins, phago ytosis an o ur through the omplement re eptor as7 a result of an antibody-mediated hange in the fungal polysa haride apsule that allows8 intera tion of polysa haride with the omplement re eptor. Hen e, in the absen e of om-9 plement re eptor blo kage, phago ytosis o urs through both F and omplement re eptors,10 whereas in the presen e of blo kage phago ytosis o urs only through the F re eptor. In11 the absen e of omplement re eptor blo kage the number of ingested mi robes in reased as12 a fun tion of antibody on entration until 250 �g/ml but sharply de reased thereafter (Fig-13 ure 2A). At most antibody on entrations studied, phago ytosis is essentially ompleted or14 onsiderably slowed down after 2 h.15 Mathemati al analysis of antibody binding to C. neoformans and F re eptors. C. neo-16 formans apsule has 1:1 � 106 binding sites [4℄ whi h implies that the 1:5 � 105 mi robes17 13
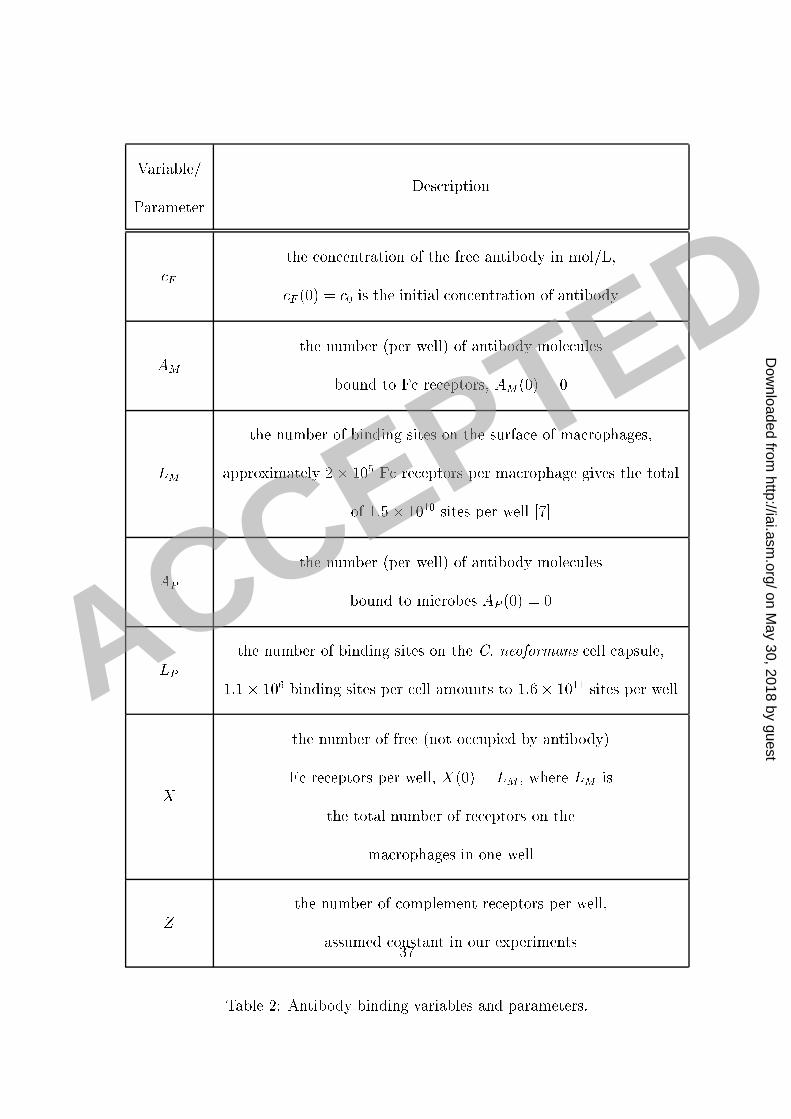
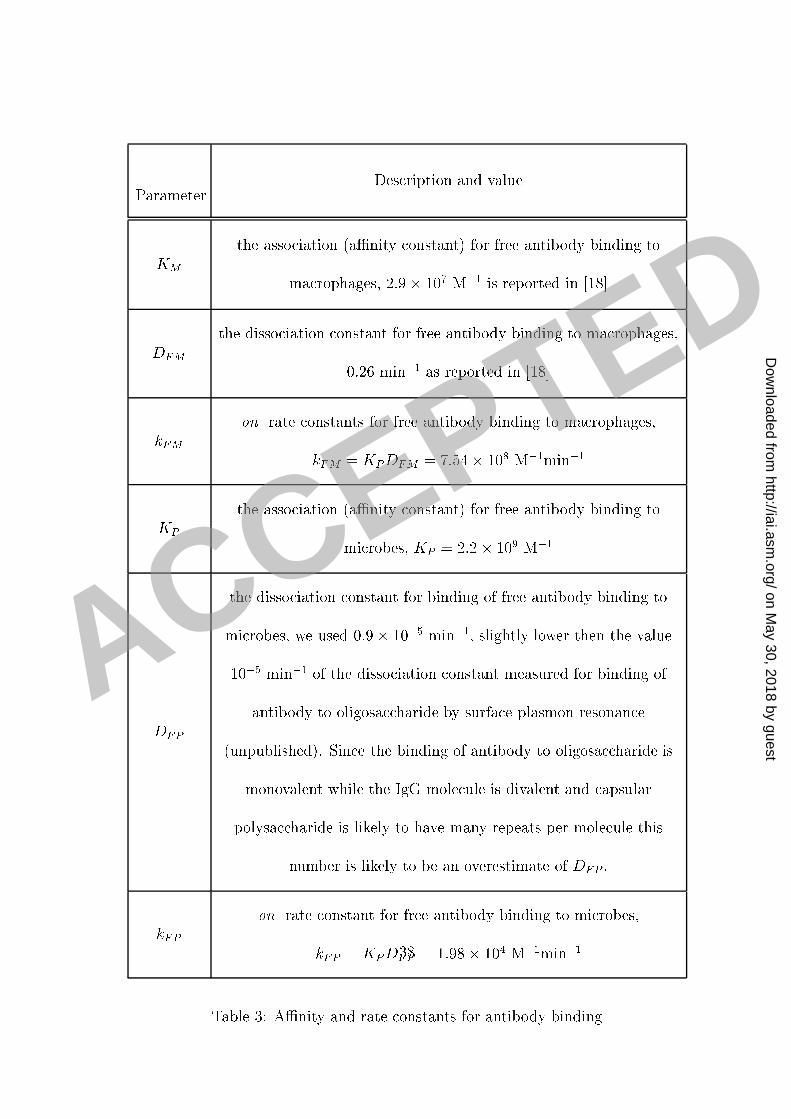
per well used in our experiments an bind 0.04 �g of IgG. It is known that ea h ma rophage1 ell has approximately 105 binding sites (see [18, 7℄) and therefore 0:75� 105 ma rophages2 would bind approximately 0.004 �g IgG (for 2� 105 binding sites per ma rophage as given3 in [7℄). This implies that in our experiments the amount of free antibody is well above the4 total (Crypto o us neoformans apsule and ma rophage together) apa ity for binding of5 antibody. In su h a ase, standard bio hemi al equations imply that the amount AM of the6 antibody bound to the F re eptors is given by the exponential fun tion7 AM = LM KM 0KM 0 + 1(1� e�DFM (KM 0+1)t): (E-11 )8 LM is the total apa ity (the number of re eptors) of F re eptors for binding the antibody,9 KM the aÆnity onstant and DFM the orresponding disso iation rate onstant. 0 is the10 initial on entration of free antibody. A summary of the variables and parameters as well11 as their values and units are given in Tables 2 and 3. A full derivation and a dis ussion of12 these equations is given in the supplemental se tion A mathemati al model of phago ytosis.13 Similarly, we obtain that the amount of the antibody AP bound to the Crypto o us14 neoformans apsule is given by:15 AP = LP KP 0KP 0 + 1(1� e�DFP (KP 0+1)t): (E-12 )16 14
LP is the total apa ity of mi robes for binding the antibody (number of binding sites in the1 apsule), KP the aÆnity onstant and DFP the orresponding disso iation rate onstant.2 From the above equations we an al ulate that the half-time for the antibody-ma rophage3 binding is S1=2 = 1DFM(KM 0 + 1) ln 2KM 0KM 0 � 1 and 90% of F re eptors are o upied after4 S90 = 1DFM(KM 0 + 1) ln 10KM 0KM 0 � 9 : This implies that 90% of F re eptors would be o u-5 pied in less than one minute at on entrations of 100, 250 and 500 �g/ml of free antibody,6 assuming that KM = 2� 107M�1: At 10 �g/ml approximately 80% of the total number of7 F re eptors are o upied in a long run and the saturation is rea hed within the �rst �ve8 minutes. The predi tions for the amount of free antibody bound to C. neoformans apsule9 are depi ted in Figure 3 and indi ate that this pro ess will rea h saturation within two hours.10 The above binding dynami s are onsistent with our experimental data on the time- ourse11 of phago ytosis at low (Figure 2A) as well as at high on entrations. There was no in rease12 in the number of ingested mi robes after one hour with high initial on entrations of free13 antibody (Figure 2B). However, the experimental data also revealed a progressive in rease in14 phago ytosis after 1 h for assays using an initial antibody on entration of 250 �g/ml. This15 was unexpe ted be ause at this on entration our al ulations showed that most of the F 16 re eptors on the ma rophage surfa e and antibody binding sites in the apsule are o upied17 (saturated) at that time.18 The in rease in the phago yti index at antibody on entrations of 100 and 250 �g/ml19 (Figure 1A) was also rather surprising, onsidering that most F re eptors are o upied very20 15
rapidly and that the number of sites on C. neoformans available for binding of antibody1 already bound to F re eptors is redu ed with higher on entrations. This was unexpe ted2 be ause at this on entration our al ulations showed that most of the F re eptors on the3 ma rophage surfa e and antibody binding sites in the apsule are o upied (saturated).4 It is noteworthy that these results indi ate that there is no repli ation of ingested yeast.5 That result is onsistent with the fa t that intra ellular yeast repli ation o urs several hours6 after ingestion [17℄, while the experiments studied and modeled here were limited to 2 h.7 The above experimental data and the analysis of the binding pro ess indi ated that the8 dynami s of binding in this experimental setup had a major in uen e on the out ome of9 these experiments. We hypothesized that phago ytosis at lower on entrations of antibody10 was fa ilitated by both F and omplement re eptors, that at higher on entrations the on-11 tribution of the omplement re eptors would be more signi� ant than at low on entrations,12 and the phago ytosis through F re eptors negligible sin e free antibody would blo k most of13 them very qui kly. Consequently, we proposed a mathemati al des ription of the eÆ a y of14 phago ytosis and designed several new experiments to analyze and quantify its dependen e15 on the amount of antibody and further develop the mathemati al formalism based on the16 experimental results obtained.17 We performed three additional sets of experiments, ea h set in luding two onditions, one18 with and the other without blo king omplement re eptors. In all three sets C. neoformans19 was in ubated with IgG1 for at least one hour to obtain near-saturation o upan y of the20 16
binding sites on the C. neoformans apsule. This provided us with a set of data where the1 amount of antibody bound to C. neoformans apsule did not hange signi� antly over the2 two hours during whi h the phago ytosis experiments were ondu ted and was therefore3 suitable for modeling phago yti eÆ a y and validating our models and assumptions.4 Experiments designed to investigate phago yti eÆ a y. In the �rst set of experiments5 C. neoformans ells were in ubated with antibody for one hour and then separated from re-6 maining free antibody present in the solution and added to the ma rophage monolayer in the7 fresh medium. This experimental setup eliminated the possibility of blo king of F re eptors8 by ex ess free antibody. In this set we ondu ted experiments without blo king omplement9 re eptors as well as experiments with (partial) blo king of omplement re eptors. The exper-10 imental results are shown in Figure 4. We blo ked the omplement re eptor using antibodies11 to the omplement re eptor subunit omponents CD11b, CD11 , and CD18. These antibod-12 ies do not a�e t the F re eptor but inhibit phago ytosis through the omplement re eptor.13 The experimental methodology used had been developed in a prior study that explored me h-14 anisms by whi h IgM promoted phago ytosis in the absen e of omplement [14℄. Initially we15 applied those onditions to the system studied but the inability of our modeling equations to16 adequately des ribe the dependen e of the phago yti index on the antibody on entration17 suggested that this proto ol whi h had been devised to blo k IgM-mediated phago ytosis18 through the omplement re eptor was inadequate for fully inhibiting IgG-mediated phago-19 ytosis through that re eptor. Hen e, the proto ol was modi�ed to in rease the amount20 17
of antibody in initial blo king step and to maintain blo king by adding antibody to the1 omplement re eptor to the phago ytosis onditions. Phago ytosis data generated with the2 enhan ed blo king onditions was well simulated by our proposed model. Hen e, the model3 proposed here was predi tive of in omplete blo king and generated a hypothesis that ould4 be tested by altering the onditions of the experiment.5 In the se ond set of experiments C. neoformans ells were again in ubated with IgG16 and for one hour to obtain near-saturation o upan y of the binding sites on the C. ne-7 oformans apsule. C. neoformans ells were then separated from remaining free antibody8 present in the solution and added to ma rophage monolayer in the fresh medium. We �rst9 ondu ted a ontrol experiment without blo king of omplement re eptors and under the10 same experimental onditions as the experiment without blo king omplement re eptors in11 the �rst set. The experiment with blo king of omplement re eptors was also ondu ted un-12 der the same experimental onditions as the orresponding experiment in the �rst set, ex ept13 for the enhan ed blo king of omplement re eptors. For these experiments the proto ol of14 the experiment with blo king was modi�ed by in ubating the ma rophage monolayer with15 ( omplement) blo king antibodies (50 �g/ml) for 1 h prior to the phago ytosis experiment.16 After washing and addition of the C. neoformans suspension and mAb 18B7 omplement17 re eptor blo king was maintained by in luding the blo king antibodies (10 �g/ml) in the18 ma rophage-yeast suspension. The experimental results are shown in Figure 4, Panel B.19 The experimental results show onsiderable redu tion in phago yti index ompared to the20 18
experiment with blo king in the �rst set and, in parti ular, almost no in rease in phago yti 1 index with in reasing on entrations of free antibody above 10 �g/ml. In our simulations2 we assume that omplement re eptors in this experiment were fully blo ked.3 In the third set of experiments we in ubated antibody with C. neoformans for an hour4 and a half. Mixture of antibody and C. neoformans was added to ma rophages and phago-5 ytized C. neoformans ells ounted after two hours. In this set of experiments we again6 examined onditions with and without blo king of omplement re eptors. The experiment7 with blo king of omplement re eptors was done under the same onditions as the experi-8 ment without blo king ex ept for the (partial) blo king of omplement re eptors (by adding9 10 �g/ml omplement blo king antibodies, as dis ussed above). The experimental results10 are shown in Figure 5. Our model reveals that a slight, but noti eable, drop in the number11 of ingested mi robes observed in experimental results at 10 �g/ml is a result of blo king F 12 re eptors by ex ess free antibody. A summary of the experimental onditions in the above13 des ribed three sets of experiments is given in Table 4.14 Modeling the eÆ a y of phago ytosis and simulations of the model. We used our pro-15 posed mathemati al model and the above des ribed experimental results to investigate the16 dependen e of the phago yti eÆ a y on the antibody bound to the apsule and to generate17 simulations that ould be ompared to the experimental data. The main variables and pa-18 rameters of the model are listed in Tables 1, 2 and 3 and a detailed mathemati al derivation19 is given as supplemental material.20 19
In our experiments C. neoformans was in ubated with IgG1 for at least one hour to1 obtain near-saturation o upan y of the binding sites on the C. neoformans apsule. This2 provided us with a set of data where the amount of antibody bound to C. neoformans3 apsule did not hange signi� antly over the two hours during whi h the phago yti eÆ a y4 was observed. In addition to that, sin e the number of ingested mi robes per ma rophage is5 small ompared to the number of mi robes a ma rophage an ingest, we an also assume that6 the number of re eptors does not hange signi� antly in our �rst two sets of experiments.7 Consequently, we an onsider that the phago yti eÆ a y rT does not hange during the8 time of the experiments (120 minutes) and solve the main equation in our model (equation E-9 1: dPIdt = rTPF ) analyti ally. Therefore, after 120 minutes, the number of ingested mi robes10 (per 100 ma rophages) is given by11 PI = �P0(1� e�120rT ): (E-13 )12 In our simulations of the model we will assume that the initial population of mi robes is13 �P0 = 200 whi h orresponds to the population of mi robes (200) per 100 ma rophages in our14 experiments, therefore normalizing the number of ells involved with respe t to the number15 of ma rophage.16 We were primarily interested in modeling the phago yti eÆ a y in the range of on-17 entrations of free antibody where the number of omplement re eptor binding sites on18 20
C. neoformans apsule in reases. It that range our model, mathemati ally derived in the1 Methods, leads to the following expression of the phago yti eÆ a y rT of phago ytosis2rT = �RM1(1� e�k1 APLP ) +RM2(1� e�k2(APLP )2)�(1 +RM4(1� e�k4(APLP )4); (E-14 )3 where AP=LP is the fra tion of antibody binding sites on C. neoformans apsule that are4 o upied. The units for rT ; RM1 and RM2 are min�1 and the parameters k1; k2; k4 and RM45 are dimensionless parameters. We refer the reader to Table 2 for the des ription, values and6 units of AP and LP :7 This model is based on the assumption that there is a maximum phago yti eÆ a y8 RM1 for F as well as for omplement re eptors (RM2), but that high on entrations of9 antibody bound to C. neoformans apsule and the presen e of omplement re eptors provide10 an additional in rease in both the maximum phago yti eÆ a y and the rate at whi h this11 maximum is rea hed. The �rst two terms des ribe the in rease in phago yti eÆ a y due12 to F (RM1(1� e�k1 APLP )) and omplement re eptors (RM2(1� e�k2(APLP )2)) when there is no13 ooperation among re eptors. The rates k1 and k2 quantify the rates at whi h the eÆ a y14 of phago ytosis in reases with the in reasing amount of antibody bound the C. neoformans15 apsule. The fa tor RM4(1� e�k4(APLP )4) a ounts for the in rease in the phago yti eÆ a y16 due to ooperation among di�erent re eptors.17 We assumed that a maximum number of omplement re eptor binding sites is rea hed for18 21
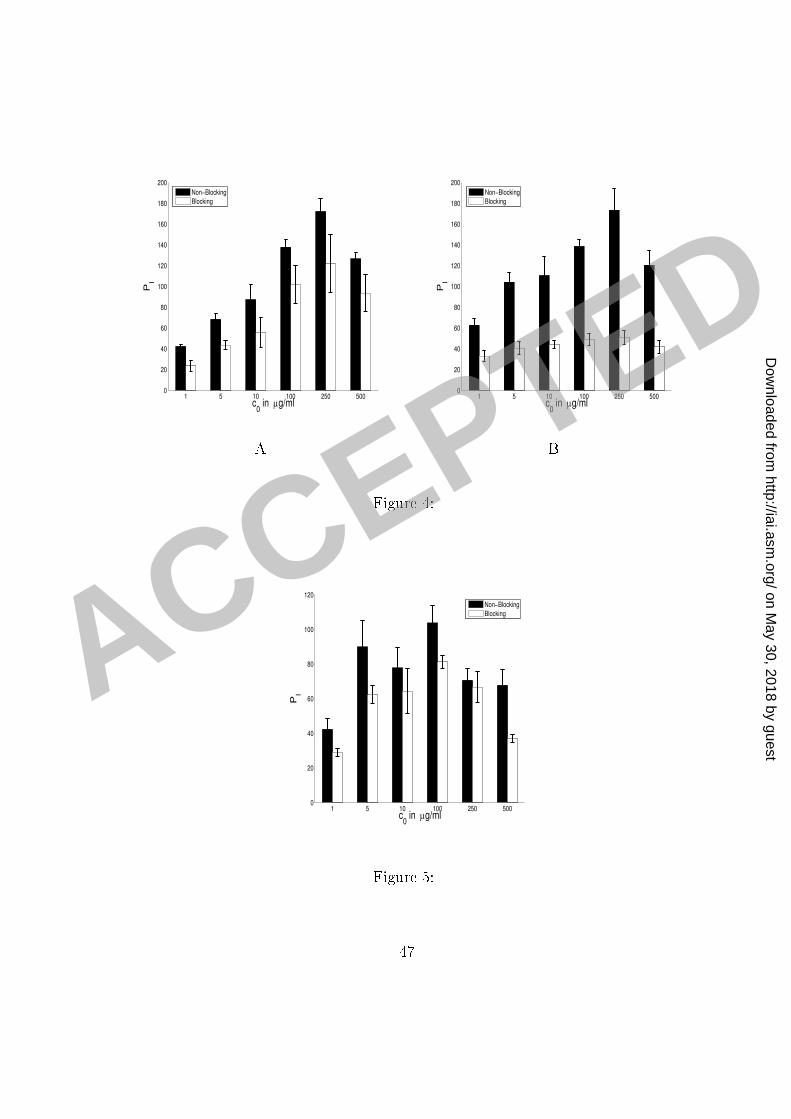
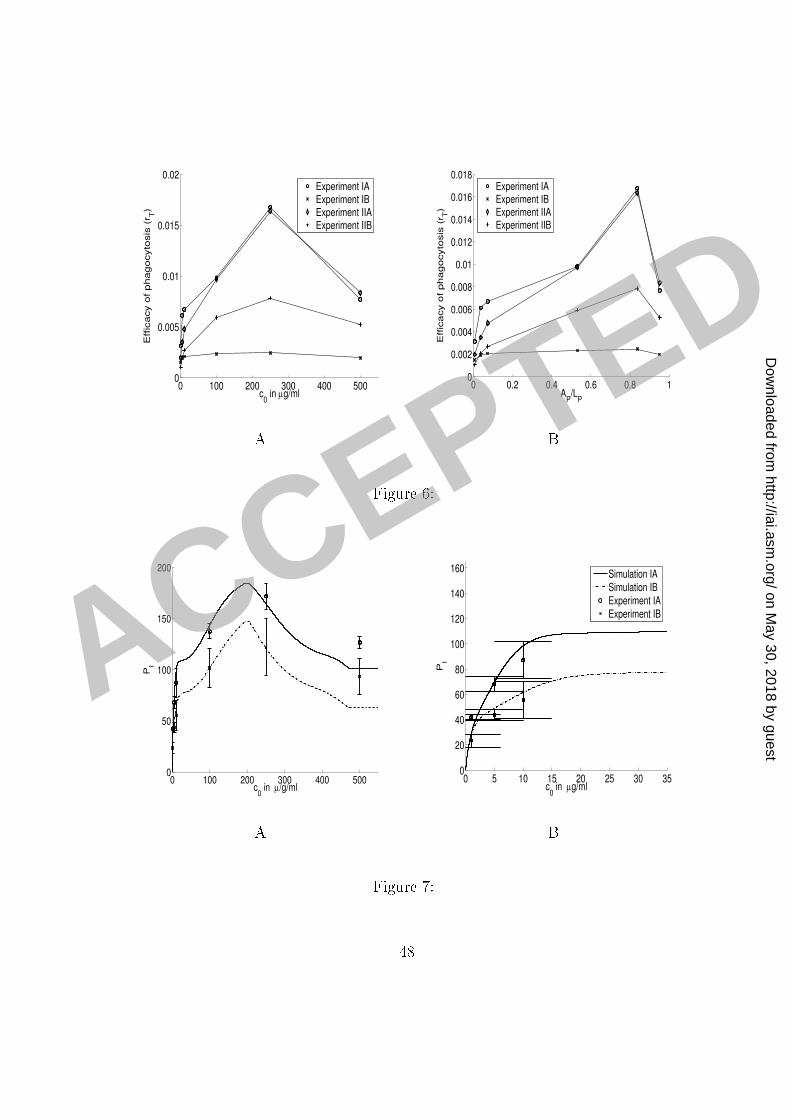
a ertain o upan y of C. neoformans apsule and that, after that point, the number of sites1 de reases quadrati ally with the in reasing amount of the antibody bound to the apsule.2 Our model provides an estimate for the fra tion of antibody binding sites on a C. neoformans3 apsule o upied that results in the maximum phago yti eÆ a y of phago ytosis through4 omplement re eptors: data presented in Figure 6 ( omputed from the experimental results5 presented in Figures 4 and 5) indi ate that the maximum number of omplement re eptor6 binding sites on C. neoformans apsule is rea hed when between 70 and 80% of the sites are7 o upied. However, we do not have enough data to set up a simple model for the events that8 o ur after the maximum eÆ a y is rea hed.9 We therefore use the model E-14 to des ribe the in rease in phago yti eÆ a y involving10 both F and omplement re eptors and in the range in whi h the number of omplement11 re eptor binding sites on the C. neoformans apsule in reases. We provide an approximate12 model, based on equation E-9 and assuming a quadrati de rease in the number of om-13 plement re eptor binding sites on the C. neoformans apsule after the maximum number is14 rea hed. This approximation �ts well to the data. The experimental results and simulations15 are presented in Figures 7, 8 and 9.16 Simulation parameters. The data for rT ; omputed from the experimental results and17 presented in Figure 6, determine the maximum eÆ a y of phago ytosis as well as the appro-18 priate rates of in rease. We �rst determined the parameters RM1 and k1 by �tting the model19 to the data-points depi ted in Figure 6, (Experiment IIB: experiment with (full) blo king in20 22
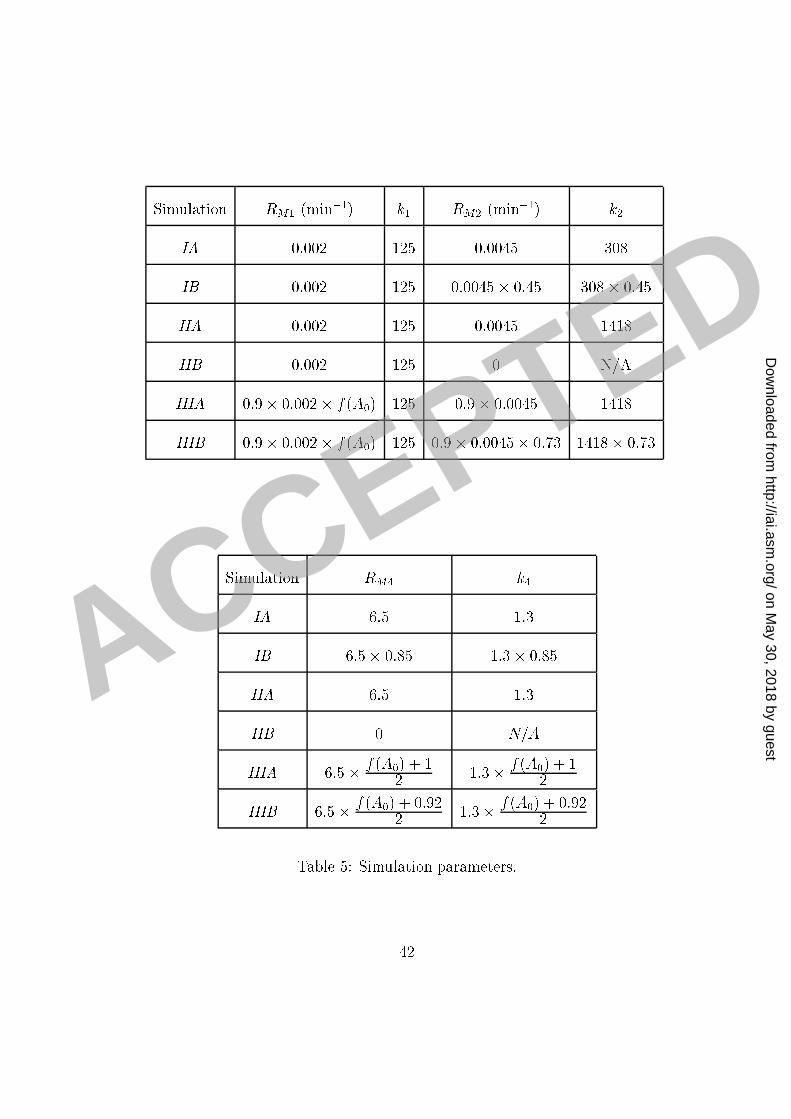
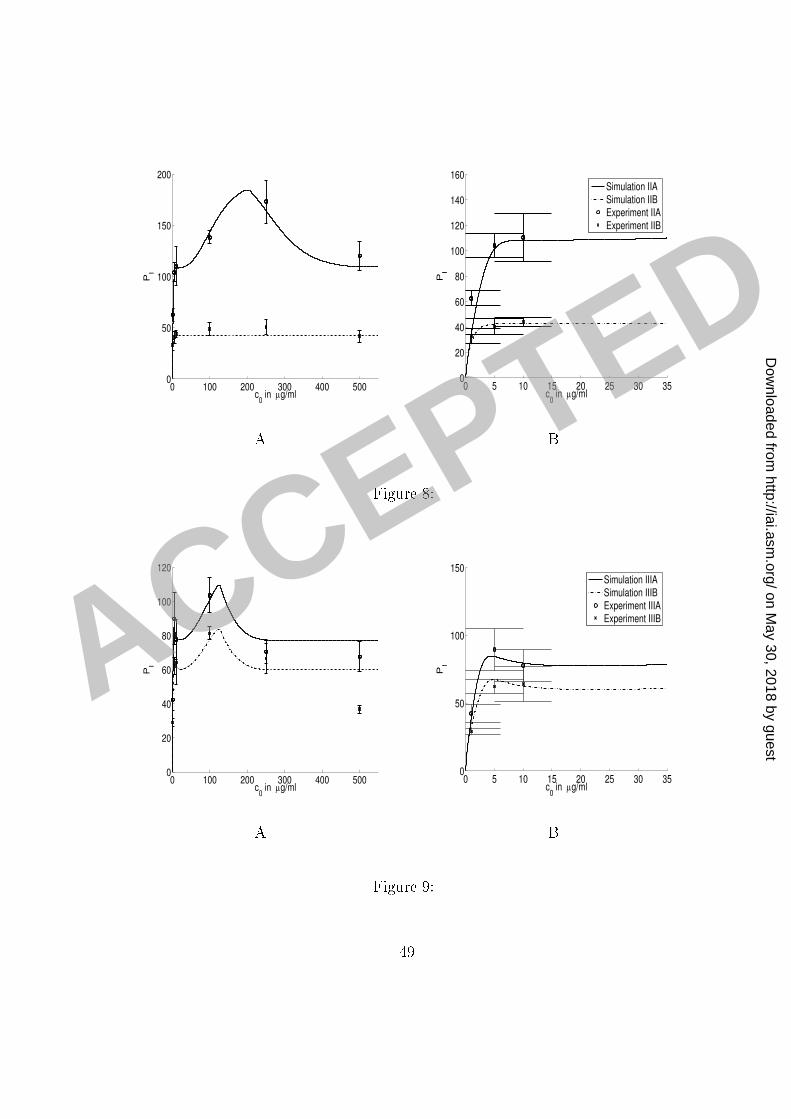
our se ond set of experiments). The model and experimental results for phago ytosis through1 F re eptors and with full omplement re eptors ompletely blo ked are presented in Figure2 8. The parameters RM2; RM4; k2 and k4 were then determined by �tting the model to the3 data-points omputed from the experimental results (Figure 4, Panel B) for phago ytosis4 without blo king omplement re eptors (in the same set) and depi ted in Figure 6 (Experi-5 ment IIA). The model and experimental results for phago ytosis through F re eptors and6 without omplement re eptors blo king are also presented in Figure 8.7 Simulations of the model for the experimental onditions in our �rst set of experiments8 presented in Figure 7 were obtained by redu ing the appropriate parameters in our model9 for the phago ytosis through omplement re eptors, as indi ated in Tables 4 and 5. We note10 that the out omes of the experiments without blo king, whi h were ondu ted under the11 same experimental onditions, were slightly di�erent. We attributed this di�eren e to small12 variations in the experimental pro edure; for example, state of a tivation of ma rophages,13 di�erent media lots, et and a ounted for those variations �rst by adjusting the parameters14 (k2) of the model to �t the experimental results of the ontrol experiment. Similar adjustment15 was done when �tting the parameters to simulate the results of the experiments in our16 third set, where we redu ed the total maximum eÆ a y of phago ytosis by a fa tor 0.9.17 Results (not shown) of earlier experiments ondu ted under the same onditions justify this18 adjustment and its attribution to the experiment-to-experiment variation.19 Simulations of the results of the experiments in our third set, ondu ted without sepa-20 23
rating ex ess free antibody from the suspension ontaining mi robes, are presented in Figure1 9. The model parameters in this ase were obtained by introdu ing the dependen e of the2 parameters RM1 and RM4; on the initial amount of free antibody. While the assumption3 that the number of re eptors does not signi� antly hange in the ourse of our experiments4 is reasonable in our �rst two sets of experiments, it does not apply to the third set, sin e our5 omputations showed that a large fra tion of F re eptors were blo ked by ex ess free anti-6 body. Sin e the onditions in our experiments were uniform and ontrolled, we know that7 most yeast ells rea h the ma rophage layer within minutes. Moreover, our omputations8 indi ated that most F re eptors are blo ked by ex ess antibody also within minutes at all9 initial on entrations of free antibody used in our experiments ex ept the lowest one. There-10 fore we assumed that the phago yti eÆ a y in our experiment was mostly in uen ed by11 the number of F re eptors that were available at the moment when the yeast ells rea hed12 the ma rophage layer. Consequently, when applying our model on the experimental results13 of the results of our third set of experiments, we a ounted for this redu tion in the number14 of F re eptors by making an appropriate adjustment to phago yti eÆ a y. We therefore15 introdu ed a de rease in the maximum phago yti eÆ a y through F re eptors RM1 by a16 fa tor f( 0) = e�0:23 0 (where 0 is in �g/ml). This de rease orresponds to the in rease17 in blo king of F re eptors as the on entration of free antibody is in reasing. We also re-18 du e RM4 by a fa tor whi h hara terizes the ontribution of F re eptors to the in rease in19 phago yti eÆ a y due to ooperation among the re eptors.20 24
This is a very rough approximation but it gives surprisingly good �t to the experimental1 data. The rationale for su h an estimate is that the fra tion of the F re eptors that will bind2 free antibody in reases exponentially as a fun tion of the initial on entration and that the3 phago yti eÆ a y is onsiderably redu ed at the point when a large fra tion of F re eptors4 are o upied very early in the pro ess.5 The summary of experimental onditions in the experiments modeled and the orrespond-6 ing adjustments of the parameters in the simulations is given in the Table 4. The values of7 model parameters used in simulations are given in Table 5.8Dis ussion9 Experimental studies of dependen e of the phago yti index on IgG on entration revealed a10 pe uliar aspe t of this pro ess that was diÆ ult to explain: while phago yti index in reased11 steadily in the 10-100 �g/ml IgG range, at antibody on entrations greater than 250 �g/ml12 there was a onsistent net redu tion in phago yti index. This phenomenon is alled the13 prozone-like e�e t and it is noteworthy to mention that a similar type of non-linear dose14 dependen e is observed in the experiments with survival rate of mi e subje t to passive15 immunization, where the high doses of antibody are asso iated with redu ed eÆ a y and16 prote tion [13℄. Moreover, the higher on entrations used in our experiments and studied17 in our mathemati al analysis are omparable to those found when antibody is administered18 passively or therapeuti ally. The lower on entrations of antibody used in our studies are19 25
omparable to those found after an immune response or va ination.1 To understand the me hanisms of phago ytosis and, in parti ular, those responsible for2 the above des ribed in rease and subsequent drop in phago yti index, we turned to math-3 emati al modeling whi h allows quantitative analysis of ausal relationships and the testing4 of assumptions. There are several mathemati al models of phago ytosis in the literature5 [11, 16℄, but none has onsidered the me hanism of atta hment of mi robes to ma rophages6 and the dependen e of the pro esses involved in both atta hment and ingestion on the7 amount of antibody and the number of available re eptors. We des ribed the phenomenon8 of IgG-mediated phago ytosis by a system of seven di�erential equations. We started with9 very general assumptions that a biologi al pro ess of this type would satisfy and onstru ted10 a model that reprodu ed the experimental results very well.11 The results of our mathemati al analysis of the dependen e of phago yti index on the12 on entration of antibody in our initial experiments brought to light a perplexing phe-13 nomenon: the on entrations where the in rease in the phago yti index was observed in our14 initial experiments were onsiderably higher than the binding apa ity of both ma rophages15 and mi robes and there was a high likelihood that both antigen binding sites in the rypto-16 o al apsule and F re eptors were saturated. This suggested additional experiments that17 provided insights into these e�e ts and enabled us to onstru t a mathemati al model that18 reprodu ed the experimental data.19 More pre isely, our al ulations indi ated that in our initial set of experiment F re eptors20 26
would be rapidly saturated at on entrations higher than 10 �g/ml. That is, we obtained1 from equation E-11 that at on entrations of 100, 250 and 500 �g/ml of free antibody half2 of the F re eptors are o upied within a minute and 90% of them are o upied in less then3 5 minutes. As we already pointed out in our initial experiments we observed an in rease in4 phago yti index through the range of on entration up to 250 �g/ml of free antibody.5 Our mathemati al analysis indi ated that the onditions in the experiments, whose out-6 omes we analyzed were quite extreme: the on entrations of free antibody were mu h higher7 than the binding apa ity of not only ma rophages but also mi robes. The fa t that the range8 of on entrations studied is well above the binding apa ity of mi robes and ma rophages9 and yet we still observed an in rease in phago yti index over that range put an emphasis on10 understanding the dynami s of phago ytosis and, in parti ular, the ontribution of phago-11 ytosis through omplement re eptors to the phago yti index. Our analysis of binding of12 antibody to the C. neoformans apsule predi ted that the timing of mixing the antibody13 with mi robes and ma rophages was essential for the out omes of our experiments.14 More pre isely, the analysis indi ated that the time ne essary for the binding sites on15 C. neoformans apsule to rea h saturation was omparable to the time during whi h the16 phago ytosis was taking pla e and was observed (e.g. 2h). Consequently, this indi ated that17 the amount of antibody bound to C. neoformans apsule hanged onsiderably throughout18 the duration of our experiments. To understand and model the dependen e of phago yti 19 index on the amount of antibody bound to C. neoformans apsule we designed and ondu ted20 27
a new set of experiments where the amount of antibody would not hange signi� antly1 during the time when phago ytosis is in progress and used their results to onstru t our2 mathemati al model.3 In parti ular, we introdu ed and studied the eÆ a y of phago ytosis, whi h we de�ne as4 the fra tion of the free mi robes that is ingested as a fun tion of the amount of antibody5 bound to C. neoformans apsule and the number of re eptors available (see E-1). We dis-6 ussed and tested mathemati al models for this fun tion and onstru ted a mathemati al7 model for phago yti eÆ a y through F and omplement re eptors separately as well as for8 phago yti eÆ a y when both types of re eptors are available.9 By studying the redu tion in phago yti index that resulted from blo king omplement10 re eptors by antibodies to CD11 and CD18 we distinguished among the ontributions from11 the two di�erent types of re eptors: F re eptors and omplement re eptors. In fa t, our12 modeling simulations predi ted that the blo king proto ol used in earlier studies [13, 14℄ was13 inadequate to fully inhibit phago ytosis through the omplement re eptor at higher antibody14 on entrations. Consequently, the proto ol for blo king omplement re eptors was modi�ed15 to use higher amounts of blo king antibody, leading to improved on ordan e between ex-16 perimental results and mathemati al predi tions. The ability of the mathemati al model to17 dis riminate between re eptor binding and to yield insight on the relative ontribution of18 the two re eptors to phago ytosis provides on�den e in the relevan e and orre tness of the19 proposed equations.20 28
We onsidered IgG on entration e�e ts on both the mi robe and the ma rophage. Our1 model predi ts that more free antibody results in more rapid binding to the yeast apsule,2 produ ing a higher saturation level and therefore rapidly enhan es phago ytosis through3 F re eptors. However, too mu h antibody rapidly redu es the number of available F 4 re eptors. Phago ytosis through omplement re eptors rapidly ompensates for this e�e t,5 and after a slight but noti eable drop in the phago yti eÆ a y (at the on entration of6 free antibody around 10 �g/ml in our experiments), phago yti index ontinues to in rease.7 However, eventually there is too mu h antibody bound to the apsule, whi h onsiderably8 redu es phago ytosis through omplement re eptors, possibly through steri e�e ts and/or9 additional hanges to the apsule. Additional experimental information that ould ontribute10 to the re�nement of the model in lude more quantitative data on the involvement of di�erent11 types of re eptors, the extent to whi h there is ooperation between them and a better12 understanding of the intera tion between antibody-modi�ed apsular polysa haride and13 the omplement re eptors. Parti ularly interesting would be experiments with di�erent14 biologi al systems whereby antibody-mediated phago ytosis o urs through more than one15 ellular re eptor.16 Given that antibody-mediated phago ytosis through the omplement re eptor has re-17 ently been shown to be asso iated with prote tion [19℄, interferen e with this me hanism18 by high antibody on entrations may ontribute to the de line in eÆ a y observed in passive19 prote tion studies with large antibody doses [13, 15℄. We expe t that in all onditions the20 29
eÆ a y of phago ytosis will involve a subtle balan e between enough free antibody available1 to bind to the yeast apsule and yet not so mu h as to saturate the apsule and F re eptors.2 We established that with higher initial on entrations of free antibody the aÆnity on-3 stant of F re eptors for binding of free antibody is an essential parameter of the model and4 might in uen e how the phago yti index depends on the on entrations of free antibody.5 This in turn implies that modeling the eÆ a y of phago ytosis will strongly depend on the6 type of the phago yti ell and the types, aÆnities and numbers F re eptors involved. Fur-7 thermore, the model predi ts that the magnitude of the aÆnity and disso iation onstants8 of the antibody from binding sites on the mi robe is a major ontributor to the phago yti 9 eÆ a y, espe ially for the phago ytosis through the omplement re eptors.10 In summary, we propose a mathemati al model for the phago ytosis of C. neoformans11 that an simulate and explain experimental results obtained through a range of antibody12 on entrations. The ability of our model to predi t the experimental out ome on the ba-13 sis of the assumed parameters provides on�den e that the major variables and parameters14 in this system are well understood. The model proposed su essfully predi ts the depen-15 den e of the phago yti index on the antibody on entration and explains the paradoxi al16 redu tion in phago yti index at high antibody on entrations. Hen e, our model provides17 an explanation for the pro esses involved, establishes the relative importan e of those vari-18 ables and parameters and provides a starting point for in orporating additional omplexity.19 Although the equations presented were derived to explain the experimental data for C. ne-20 30
oformans they ould serve as a basis for future mathemati al modeling of phago ytosis of1 other mi robes, espe ially those systems in whi h opsonization and/or ooperation among2 di�erent types of re eptors play a signi� ant role. The development of a more a urate model3 for C. neoformans phago ytosis would require additional information that is not urrently4 available, in luding the rates of mi robe-ma rophage atta hment as a fun tion of antibody5 on entration, ma rophage re eptor density and mi robial binding sites. In this regard,6 the mathemati al modeling has identi�ed gaps in the urrent knowledge of C.neoformans7 phago ytosis, and alls for further studies, experimental and theoreti al, of the me hanisms8 of atta hment and ingestion of this and other mi robe to ma rophages, the signaling path-9 ways involved and the potential for ooperation between di�erent types of opsoni re eptors.10
A knowledgment: A. Casadevall is supported by NIH grants AI033142, AI033774 and1 HL059842.2 N. Ma ura would like to thank W. We kesser for many valuable dis ussions. The authors3 would also like to thank The Mathemati al Bios ien es Institute at Ohio State University for4 organizing the interdis iplinary workshop on Host-Pathogen Intera tions in Disease Models5 in June 2004 where we began our ollaboration. We are also grateful to the reviewers for6 their valuable omments.7Referen es8 [1℄ Camner, P. and M. Lundborg and L. Lastbom and P. Gerdeand N. Gross9 and C. Jarstrand. Experimental and al ulated parameters on parti le phago ytosis10 by alveolar ma rophages. J. Appl. Physiol., 92(6):2608{16, 2002.11 [2℄ Casadevall, A. The methodology for determining the eÆ a y of antibody-mediated12 immunity. J. Immunol. Methods., 291(1-2):1{10, 2004.13 [3℄ Casadevall, A. and W. Cleare and M. Feldmesser and A. Glatman-Freedman14 and D. L. Goldman and T. R. Kozel and N. Lendvai and J. Mukherjee and15 L. Pirofski and J. Rivera and A. L. Rosas and M. D. S har� and P. Valadon16 and K. Westin and Z. Zhong. Chara terization of a murine mono lonal antibody to17
Crypto o o us neoformans polysa haride that is a andidate for human therapeuti 1 studies. Antimi rob. Agents Chemotherap., 42:1437{46, 1998.2 [4℄ Dada hova, E. and R. A. Bryan and C. Apostolidis and A. Morgenstern and3 T. Zhang and T. Moadel and M. Torres and X. Huang and E. Revskaya and4 A. Casadevall. Intera tion of radiolabeled antibodies with fungal ells and omponents5 of the immune system in vitro and during radioimmunotherapy for experimental fungal6 infe tion. J. Infe t. Dis., 193(10):1427{36, 2006.7 [5℄ Larsen, R. A. and P. G. Pappas and J. R. Perfe t and J. A. Aberg and A.8 Casadevall and G. A. Cloud and R. James and S. Filler and W. E. Dismukes.9 A phase I evaluation of the safety and pharma odynami a tivity of a murine-derived10 mono lonal antibody 18b7 in subje ts with treated rypto o al meningitis. Antimi rob.11 Agents Chemother., 49(3):952{58, 2005.12 [6℄ Luo, Y. and S. C. Tu ker and A. Casadevall. F - and omplement-re eptor a ti-13 vation stimulates ell y le progression of ma rophage ells from G1 to S. J. Immunol.,14 174(11):7226{33, 2005.15 [7℄ Mellman, I. S. and H. Plutner and R. M. Steinman and J. C. Unkeless and Z.16 A. Cohn. Internalization and degradation of ma rophage F re eptors during re eptor-17 mediated phago ytosis. J. Cell Biol., 96(3):887{95, 1983.1833
[8℄ Mukherjee, S. and M. Feldmesser and A. Casadevall. . J774 murine ma rophage-1 like ell intera tions with Crypto o us neoformans in the presen e and absen e of op-2 sonins. J. Infe t. Dis., 173(5):1222{31, 1996.3 [9℄ Mukherjee, S. and S. C. Lee and A. Casadevall. Antibodies to Crypto o us4 neoformans glu uronoxylomannan enhan e antifungal a tivity of murine ma rophages.5 Infe t. Immun., 63(2):573{79, 1995.6 [10℄ Perfe t, J.R. and A. Casadevall. Crypto o osis. Infe t. Dis. Clin. North Am.,7 16(4):837{74, v{vi, 2002.8 [11℄ Petri, I. and R. Egerer and A. Stelzner and J. Suss and H. S hutz. Devel-9 opment of mathemati al models for an in vitro-phago ytosis test system. Zentralbl.10 Bakteriol. Mikrobiol. Hyg. [A℄., 267(2):217{27, 1987.11 [12℄ Rivera, J. and A. Casadevall. Mouse geneti ba kground is a major determinant of12 isotype-related di�eren es for antibody-mediated prote tive eÆ a y against Crypto o -13 us neoformans. J. Immunol., 174(12):8017{26, 2005.14 [13℄ Taborda, C.P. and A. Casadevall. Immunoglobulin M eÆ a y against Crypto o us15 neoformans:me hanism, dose dependen e, and prozone-like e�e ts in passive prote tion16 experiments. J. Immunol., 166(3):2100{7, 2001.1734
[14℄ Taborda, C.P. and A. Casadevall. . CR3 (CD11b/CD18) and CR4 (CD11 /CD18)1 are involved in omplement-independent antibody-mediated phago ytosis of Crypto o -2 us neoformans. Immunity, 16(6):791{802, 2002.3 [15℄ Taborda, C.P and J. Rivera and O. Zaragoza and A. Casadevall. More is not4 ne essarily better: prozone-like e�e ts in passive immunization with IgG. J. Immunol.,5 170(7):3621{30, 2003.6 [16℄ Tran, C.L. and A. D. Jones and K. Donaldson. Mathemati al model of phago y-7 tosis and in ammation after the inhalation of quartz at di�erent on entrations. S and.8 J. Work Environ. Health., 21 Suppl 2:50{4, 1995.9 [17℄ Tu ker, S.C. and A. Casadevall. Repli ation of Crypto o us neoformans in10 ma rophages is a ompanied by phagosomal permeabilization and a umulation of vesi-11 les ontaining polysa haride in the ytoplasm. Pro . Natl. A ad. S i., 99(5):3165{70,12 2002.13 [18℄ Unkeless, J.C. and H. N. Eisen. Binding of monomeri immunoglobulins to F 14 re eptors of mouse ma rophages. J. Exp. Med., 142(6):1520{33, 1975.15 [19℄ Wang, F. and A. Nakouzi and M. Alvarez and O. Zaragoza and R. H. An-16 geletti and A. Casadevall. Stru tural and fun tional hara terization of gly osyla-17 tion in an immunoglobulin G1 to Crypto o us neoformans glu uronoxylomannan. Mol.18 Immunol., 3.(7):987{8, 2006.19 35
Variable Des riptionM the number of ma rophages per well, assumed onstant is our simulations,M = M(0) = 0:75� 105PF the population of free (not ingested) mi robes, inmi robes/well, PF (0) = P0 = 1:5� 105;�PF (0) = �P0 = 200 the normalized (per 100ma rophages) initial population of free mi robesPI the population of mi robes ingested byma rophages in mi robes/well PI(0) = 0Table 1: The main variables in the model.
Variable/Parameter Des ription F the on entration of the free antibody in mol/L, F (0) = 0 is the initial on entration of antibodyAM the number (per well) of antibody mole ulesbound to F re eptors, AM(0) = 0LM the number of binding sites on the surfa e of ma rophages,approximately 2� 105 F re eptors per ma rophage gives the totalof 1:5� 1010 sites per well [7℄AP the number (per well) of antibody mole ulesbound to mi robes AP (0) = 0LP the number of binding sites on the C. neoformans ell apsule,1:1� 106 binding sites per ell amounts to 1:6� 1011 sites per wellX the number of free (not o upied by antibody)F re eptors per well, X(0) = LM ; where LM isthe total number of re eptors on thema rophages in one wellZ the number of omplement re eptors per well,assumed onstant in our experimentsTable 2: Antibody binding variables and parameters.37
Parameter Des ription and valueKM the asso iation (aÆnity onstant) for free antibody binding toma rophages, 2:9� 107 M�1 is reported in [18℄DFM the disso iation onstant for free antibody binding to ma rophages,0.26 min�1 as reported in [18℄kFM on rate onstants for free antibody binding to ma rophages,kFM = KPDFM = 7:54� 108 M�1min�1KP the asso iation (aÆnity onstant) for free antibody binding tomi robes, KP = 2:2� 109 M�1DFP
the disso iation onstant for binding of free antibody binding tomi robes, we used 0:9� 10�5 min�1, slightly lower then the value10�5 min�1 of the disso iation onstant measured for binding ofantibody to oligosa haride by surfa e plasmon resonan e(unpublished). Sin e the binding of antibody to oligosa haride ismonovalent while the IgG mole ule is divalent and apsularpolysa haride is likely to have many repeats per mole ule thisnumber is likely to be an overestimate of DFP :kFP on rate onstant for free antibody binding to mi robes,kFP = KPDFP = 1:98� 104 M�1min�1Table 3: AÆnity and rate onstants for antibody binding38
Experiment Simulation and the parametersFigure 4, Panel A, Experiment IA:Phago ytosis without blo king omplementre eptors, mi robes in ubated with antibodyfor one hour, ex ess free antibody removedprior to adding antibody oated mi robes toma rophages.Figure 7, Simulation IAMaximum values for RM1; k1; RM2; RM4and k4: k2 redu ed due to experimentalvariations.
Figure 4, Panel A, Experiment IB:Phago ytosis with partial blo king of omplement re eptors, mi robes in ubated withantibody for one hour and ex ess free antibodyremoved prior to adding antibody oatedmi robes to ma rophages.Figure 7: Simulation IBMaximum values for RM1; k1. k2 redu eddue to experimental variations.Complement and ooperation parametersRM2; k2; RM4 and k4 redu ed.Figure 4, Panel B, Experiment IIA:Phago ytosis without blo king omplementre eptors, mi robes in ubated with antibodyfor one hour, ex ess free antibody removedprior to adding antibody oated mi robes toma rophages.Figure 8, Simulation IIAMaximum values for RM1; k1; RM2; k2;RM4 and k4:1 40
Experiment ParametersFigure 4, Panel B, Experiment IIB:Phago ytosis with omplete blo king of omplement re eptors, mi robes in ubated withantibody for one hour and ex ess free antibodyremoved prior to adding antibody oatedmi robes to ma rophages.Figure 8, Simulation IIBMaximum values for RM1; k1. RM2 = 0 andRM4 = 4 due to the omplete blo king of omplement re eptors. and ooperationparameters.Figure 5, Experiment IIIA: Phago ytosiswithout blo king omplement re eptors,mi robes in ubated with antibody for one and ahalf hour, suspension ontaining mi robes andex ess free antibody added to ma rophages.Figure 9, Simulation IIIAValues of RM1; and RM2; redu ed by fa tor0.9 due to experimental variation. RM1 andRM4 (by a fa tor f( 0)) redu ed due to theblo king of F re eptors by ex ess freeantibody.
Figure 5, Experiment IIIB: Phago ytosis withpartial blo king of omplement re eptors,mi robes in ubated with antibody for one and ahalf hour, suspension ontaining mi robes andex ess free antibody added to ma rophages.
Figure 9: Simulation IIIBValues of RM1; and RM2; redu ed by fa tor0.9 due to experimental variation. RM1 andRM4 redu ed (by a fa tor f( 0)) due to theblo king of F re eptors by ex ess freeantibody. Complement parameters redu eddue to the blo king of omplementre eptors.Table 4: Summary of experiments designed to model eÆ a y of phago ytosis and the orre-sponding simulations 41
Figure Legends1 Figure 1: Experimental ounts of the number of phago ytised mi robes (per 100 ma rophages)2 as a fun tion of antibody on entration (Panel A) and in the setting of blo ked omplement3 (C3R) re eptors (Panel B). Error bars represent the standard deviation.4 Figure 2: Experimental ounts of the number of phago ytised mi robes (per 100 ma rophages)5 as a fun tion of time for antibody on entrations of 10 and 100 �g/ml (Panel A) and 0,250,6 500 and 100 �g/ml (Panel B). Error bars represent the standard deviation.7 Figure 3: Graphs of the antibody binding to the mi robe apsule: the amount bound8 after 2 h as a fun tion of the initial on entration (Panel A) and the dynami s of the binding9 (Panel B).10 Figure 4: Experimental ounts of the number of phago ytised mi robes PI (per 10011 ma rophages) as a fun tion of the initial antibody on entration 0. IgG1 was in ubated12 with C. neoformans ells for one hour to obtain near-saturation o upan y of the binding13 sites on the C. neoformans apsule. C. neoformans ells were then separated from antibody14 and added to ma rophage monolayer in the fresh medium (see the se tion Methods). The15 two sets of results represent experiments without and with blo king of omplement re eptors.16 The results of experiments with blo king represented in Panel A were obtained with partial17 blo king of omplement re eptors and in Panel B with omplement re eptors fully blo ked.18 Error bars represent the standard deviation.19 Figure 5: Experimental ounts of the number of phago ytised mi robes PI (per 10020 43
ma rophages) as a fun tion of the initial antibody on entration 0: IgG1 was in ubated1 with C. neoformans ells for an hour and half to obtain near-saturation o upan y of the2 binding sites on the C. neoformans apsule. C. neoformans (in the solution) were then3 added to ma rophage monolayer (see the se tion Methods). The two sets of results represent4 experiments without and with (partial) blo king of omplement (C3R) re eptors. Error bars5 represent the standard deviation.6 Figure 6: Plots of eÆ a y of phago ytosis omputed from the experimental data. We7 omputed the eÆ a y using the formula rT = � ln(1� PI=200)=120 and experimental data8 from the three sets of experiments presented in Figure 4 and Figure 5. The eÆ a y of9 phago ytosis is plotted as a fun tion of the initial on entration 0 of antibody (Panel A)10 and as a fun tion of the fra tion (AP=LP ) of the antibody binding sites on the C.neoformans11 apsule o upied by IgG (Panel B). A summary of experimental onditions is given in Table12 4.13 Figure 7: Comparison of the model simulations and experimental results presented in14 Figure 4, Panel A. Panels A and B present the same data and simulations, but using a15 di�erent s ale. The same legend applies to both panels. Error bars represent the standard16 deviation. A summary of experimental onditions and simulation parameters is given in17 Tables 4 and 5.18 Figure 8: Comparison of the model simulations and experimental results presented in19 Figure 4, Panel B. Panels A and B present the same data and simulations, but using a20 44
di�erent s ale. The same legend applies to both panels. Error bars represent the standard1 deviation. A summary of experimental onditions and simulation parameters is given in2 Tables 4 and 5.3 Figure 9: Comparison of the model simulations and experimental results presented in4 Figure 5. Panels A and B present the same data and simulations, but using a di�erent s ale.5 The same legend applies to both panels. Error bars represent the standard deviation. A6 summary of experimental onditions and simulation parameters is given in Tables 4 and 5.7
0 1 10 100 250 500 1000 0 1 10 1000
50
100
150
200
250
300
350
400
Intial concentration (c0) of antibody in µg/ml
Num
ber
of m
icro
bes ingeste
d
0 10 100 5000
100
200
300
400
500
600
Intial concentration (c0) of free antibody in µg/ml