Influence of freezing stress on morphological alteration and biofilm formation by Listeria monocytogenes: relationship with cell surface hydrophobicity and membrane fluidity
Listeria monocytogenes is a eurythermal, osmotolerant, and pathogenic gram-positive rod. Its ability to grow at refrig-eration temperatures (najjar et al. 2007) and to persist to freezing stress (Miladi et al. 2008) is a constant challenge for food preservation. L. monocytogenes is responsible for foodborne disease in susceptible patients, including preg-nant women and immunocompromised patients. This spe-cies is considered as one of the most difficult pathogens to eradicate, due to its ubiquity in the environment (larpent 1995). L. monocytogenes is the causative agent of listeri-osis, a rare but severe illness with a mortality rate of 20–30 %. growth at refrigerated temperatures and survivability in adverse environments, including those conditions that occur in minimally processed foods, have made L. mono-cytogenes a challenge to control (gandhi and Chikindas 2007). The majority of infections caused by L. monocy-togenes is foodborne, mainly by non-cooked ready-to-eat food such as deli meat, raw milk products, sausages, or smoked fish (Swaminathan and gerner-Schmidt 2007). L. monocytogenes can enter and colonize a food-processing environment, its ability to grow at very low temperatures and survive for extended periods under adverse conditions have made this bacterium one of the hottest topics for the food industry (gandhi and Chikindas 2007). It has been shown that L. monocytogenes strains that persist in food industry environments form thicker biofilms than isolates found only sporadically (Di Bonaventura et al. 2008), indi-cating that biofilm formation is important for the survival
Abstract The morphological changes and adhesive prop-erty of three Listeria monocytogenes strains submitted to freezing stress (−20 °C) were studied. The atomic force micrographs showed a reduction in the cell size and an evolution to coccoid shape. The phenotypic slime produc-tion of L. monocytogenes and the expression of the adhe-sive gene were investigated before and after 10 months of incubation in salmon at −20°. Our results showed that after ten months, stressed stains become more adherent and able to produce slime. In addition, we noted that this pathogen presents same physiological changes to adapt to starvation conditions. The cellular fatty acids composition of adhered and floating cells of three L. monocytogenes strains was taken into consideration. The stressed strains presented different chain lengths and therefore an increase in the hydrophobicity level. Moreover, we noted that the adhesive property of L. monocytogenes strains affects the Benzalko-nium chloride bacterial sensitivity which increased after biofilm formation.
Communicated by erko Stackebrandt.
H. Miladi (*) · r. Ben Slama · A. Bakhrouf laboratory of Analysis, Treatment and Valorisation of environment Pollutants and Products, Faculty of Pharmacy, Monastir University, 5000 Monastir, Tunisiae-mail: [email protected]
H. Miladi · e. Ammar Ur Study and Management of Urban and Coastal environments, lArSen, national engineering School in Sfax, B.P. 1173, 3038 Sfax, Tunisia
n. Sakly laboratory of Physics and Chemistry of Interfaces, Faculty of Sciences, Monastir University, Monastir, Tunisia
Arch Microbiol
1 3
of L. monocytogenes in the food industry (rodriguez et al. 2008). L. monocytogenes is able to adjust to a wide range of environmental stresses to adapt to adverse conditions (najjar et al. 2007). In this context, to adapt to cold stress, the resistance to environmental stresses of this species is in part due to its ability to form biofilms on food-process-ing surfaces such as glass, PVC, or stainless steel (Weiler et al. 2013) and equipments which in turn serve as reser-voirs and primary sources of contamination (Blackman and Frank 1996; Wong 1998). Moreover, the attachment to surfaces and formation of biofilms may provide nutrition and protection against sanitation procedures and an adap-tive advantage for the attached microorganisms (Cox et al. 1997; gravani 1999).
However, this bacterium is able to persist extreme habi-tat conditions such as aridity, sanitizers, temperature, salt concentration, and pH stress in biofilms better than simi-lar planktonic cells (Frank and Koffi 1990). The condi-tion of cells in biofilm is profoundly different from that of the bacteria in the planktonic state. Bacterial biofilms are generally described as surface-associated bacterial com-munities forming microcolonies surrounded by a matrix of exopolymers (Costerton 1995). Their formation is gov-erned in part by physicochemical factors such as hydropho-bicity, van der Waals and lewis acid–base characteristics, and electrical properties (lewis 2000). However, in addi-tion to the surface physicochemical properties of the cells, in part depending on membrane phospholipid composition and their modulation, other factors come into play. Despite microbial biofilms demonstrate a decreased growth rate, variation in the transcribed genes (Donlan 2002) and induc-tion of the transcription of some genes following adhe-sion in L. monocytogenes have been reported (Taylor et al. 2002). In addition, an enhanced resistance to sanitizer, dis-infectants and antimicrobial agents has been investigated (robbins et al. 2005). However, Quaternary ammonium compounds (QACs) are cationic biocides that are com-monly used as disinfectants in both medical and food pro-duction environments. QACs are primarily active against gram-positive bacteria, but they are also active against gram-negative bacteria, some viruses, fungi, and protozo-ans. Benzalkonium chloride (BC) is a QAC that is widely used for medical disinfection and for sanitation in food-processing environments (To et al. 2002).
When L. monocytogenes is grown at low temperatures, the phospholipids of the cell membrane become more rigid and can interfere with normal functions of the membrane and membrane-associated proteins (Tasara and Stephen 2006; Schneiter and Toulmay 2007; najjar et al. 2007). To counteract this phenomenon, L. monocytogenes modi-fies its plasma membrane to maintain its fluidity at low temperature (Tasara and Stephen 2006); this is collec-tively termed homeoviscous adaptation (najjar et al. 2007).
low-temperature growth induces several levels of adjust-ments at the cell membrane. However, cell surface hydro-phobicity of bacteria can be altered by fatty acid and pro-tein compositional changes that occur in response to stress (Demel et al. 1996). An increase in short-chain hydrophilic fatty acids decreases cell hydrophobicity, whereas an increase in long-chain hydrophobic fatty acids increases cell hydrophobicity. Membrane hydrophobicity also affects bacterial attachment to surfaces (Van loosdrecht et al. 1987; Briandet et al. 1999) and the ability of metabolites to transit the cell membrane. Furthermore, one of the most frequently observed behaviors in the nutrient starvation response of bacteria is the size reduction and the cell mor-phology conversion from rod to coccoid shape (Kjelleberg et al. 1993). Despite the extended knowledge of molecular level about mechanism and regulation of nutrient starvation response, the morphology transition process from rod to coccoid shape is not yet well understood.
The aim of this work was to study morphological varia-tions in L. monocytogenes incubated 10 months in freshly sliced salmon stored at −20 °C and to assess its ability to form biofilms. Moreover, the possible correlation between changes in cell hydrophobicity and lipid composition of freezing stressed cells was examinated, in addition the study of the differences in BC sensitivity before and after adhesion on stainless steel surface.
Materials and methods
Bacterial strain isolation and biochemical characterization
Three strains of L. monocytogenes were used to artifi-cially contaminate cold salmon: one reference strain ATCC 19115 and two strains (S1 and S2) were isolated from meat according to the standardized French method nF V 08-055 (AFnOr 1999), for L. monocytogenes food detec-tion (Miladi et al. 2008). The three strains of L. monocy-togenes were separately cultured in brain heart infusion broth (BHIB) and used to surface inoculate 1 g of sterile salmon slices (108 CFU/g) (Miladi et al. 2008). The inocu-lated salmon were then incubated at −20 °C for 10 months of storage.
Determination of morphological changes by atomic force microscopy
In order to visualize any morphological changes in L. monocytogenes strains obtained after 10-month incu-bation in salmon at −20 °C, cells were examined by atomic force microscopy (AFM) (nanoscope IIIA, Digi-tal Instrument/Veeco, Santa Barbara, CA). For the experi-ments, the cells were collected, washed three times with
Arch Microbiol
1 3
phosphate-buffered saline (PBS), and centrifuged. The final pellet was resuspended in PBS placed on a round micro-scope cover slide and was simply dried in air according to the method previously described (Braga and ricci 1998; Ben Abdallah et al. 2008).
Phenotypic characterization of bacteria-producing slime
Biofilm formation was studied by culturing the isolated strains on Congo red Agar plate (CrA) made by mixing 36 g saccharose (Sigma Chemical Company, St. louis, MO) with 0.8 g Congo red in 1 l of brain heart infusion agar (Biorad, USA) and incubated at 37 °C for 24 h under aerobic conditions (Freeman et al. 1989). results were interpreted as follows: very black, black, and almost black or reddish-black colonies with a rough, dry, and crystal-line consistency on CrA were considered to be normal slime-producing strains, while very red, red, bordeaux, and smooth colonies were classified as non-slime-producing strains, as described previously (Arciola et al. 2002).
Molecular evaluation of adhesive gene of stressed cells
RNA isolation and reverse transcriptase polymerase chain reaction
Total rnA was isolated from control, and stressed cells was extracted by SV total rnA isolation system (Promega, France) according to the manufacturer’s instructions. The rnA was digested with Dnase I to eliminate residual DnA (Promega, France). rnA was electrophoresed in a formal-dehyde agarose gel to check the integrity of the samples. First-strand cDnA was produced by reverse transcription (rT) reactions, and a mixture of 100 ng total rnA, 1 μl of reverse primer (3 μg/μl; Invitrogen, Carlsbad, CA) named aad r; 5′- TCgTTgCCACgTTTTTTCAgg -3′ (AY561824), 2 μl of 10 mM dnTPs (Jena Bioscience), 4 μl of 5 × first-strand buffer (250 mM Tris–HCl at pH 8.3, 375 mM KCl, 15 mM MgCl2), 2.0 μl of 0.1 M dithi-othreitol (DTT), and 100 units of Moloney Murine leuke-mia Virus (MMlV) reverse Transcriptase (Promega) was brought up to a final volume of 20 μl with deionized water (dH2O) treated with diethylpyrocarbonate. The synthesis reaction was carried out for 45 min at 42 °C in a Multi-gene gradient Thermal Cycler (labnet International) and stopped by incubation at 75 °C for 10 min.
The cDnAs were amplified under relatively relaxed conditions as described by elabed et al. (2012). The 20-μl total reaction volume contained 6 μM MgCl2, 0.8 μl of rT reaction, 1.5 μM of aad r and forward primer (aad F; 5′- ATggCAATTAAAgAAAATgCgg -3′) (AY561824), 0.15 mM dnTPs, 2 μl of 10 × PCr buffer (200 mM Tris–HCl pH 8.4, 500 mM KCl) and 2.5 units of Taq
DnA polymerase (Promega, France). The amplification conditions comprised 5 min at 94 °C, 30 cycles of 1 min at 94 °C, 1 min at 56 °C and 1 min at 72 °C, and a final 10-min elongation at 72 °C. Primers for the 16S rRNA (Miladi et al. 2013) were used as controls. The reaction was performed in triplicate.
The PCr products (7 μl) were electrophoresed in a 1 % agarose gel in 1 × TBe buffer (0.089 M Tris–acetate, 0.089 M boric acid, 0.002 M ethylene diamine tetraacetic acid, pH 8.3) and stained with ethidium bromide (0.5 mg/ml) at 100 V for 45 min and viewed under ultraviolet transillumination. The amplification products were pho-tographed, and their sizes were determined with 100-bp molecular size marker (Promega). Quantitative analysis of DnA bands was performed using imaging software (gene Tools, Syngene, UK).
Bacterial hydrophobicity assay
The hydrophobicity of strains before and after 6 and 10 months of freezing stress (parent and stressed strains) was evaluated by the microbial adhesion to solvent (MATS) test (Bellon-Fontaine et al. 1996). It consisted on a com-parison between microbial cell affinity with a monopolar or apolar solvent. The monopolar solvent may be an electron-acceptor or electron-donor solvent, but both solvents must exhibit similar van der Waals surface tension components.
On this basis, we selected the following pairs of sol-vents: chloroform (sigma), an electron-acceptor solvent, with hexadecane (sigma), an apolar n-alkane, and ethyl acetate (sigma), a strong electron-donor solvent, with decane (sigma), an apolar n-alkane.
experimentally, 2.4 ml of a suspension containing approximately 108 cells in naCl 1.5 × 10−1 M was vortex-mixed for 60 s with 0.4 ml of the solvent under investiga-tion. The mixture was allowed to stand for 15 min to ensure complete separation of the two phases before a sample (1 ml) was carefully collected from the aqueous phase, and the optical density was measured at 400 nm. The percent-age of cells binding to each solvent was subsequently cal-culated using the equation: % affinity = 100 × [1 − (A1/A0)], where A0 is the optical density of the bacterial sus-pension measured at 400 nm before mixing, and A1 is the absorbance after mixing. each experiment was performed in triplicate by using three independently prepared cultures.
Cellular fatty acids analysis
To analyze the total cellular fatty acids, control and stressed cells cultured after 6 and 10 months of freezing stress were recovered by centrifugation (4,000 rpm for 15 min), washed three times with distilled water (Hébraud and guzzo 2000), and transferred into extraction screw-cap
Arch Microbiol
1 3
tubes. The cellular FAs were saponified, methylated, extracted, and washed (Chihib et al. 2003). The resulting methyl esters were separated by gas chromatography (gC). The FAs were identified by gas chromatography coupled with mass spectrometry (gC–MS). The gC–MS analyses were carried out with gas chromatography (gC, Hewlett-Packard 6890 Series, Agilent Technology, Precision elec-tronique, Tunisia) equipped with a mass spectrometer (MS, Hewlett-Packard 5973 Mass Selective Detector, Agilent Technology). A HP-5MS-fused silica capillary column (60 m × 0.25 mm i.d., 0.25-μm film thickness, Agilent Technology) was used. The gC oven temperature was pro-grammed as follows: 100 °C held for 2 min, raised at 5 °C/min to 290 °C (held for 20 min). Helium was the carrier gas at a flow rate of 1.07 ml/min. The injection was set on a splitless mode at 250 °C. The injected volume was 10 ml, the solvent delay was 6.00 min, and total run time was 60 min.
Detection was conducted by a mass selective detector with ionization at 70 eV in selected ion monitoring mode. MS transfer line temperature was at 280 °C. The MS was operated in full scan in electron ionization mode with an electron multiplier voltage of 2,200 V. The mass scanning ranged from m/z 50 to 550. Mass spectra were compared with the reference compounds in Wiley 275 l mass spec-tral library and identified by comparison with the retention time of standards (Chihib et al. 2003).
Impact of adhesion on cell sensitivity to disinfectant
Microbial adhesion to AISI 316 stainless steel: solid surface and cleaning treatment
The solid support selected for this study was AISI 316 stainless steel (goodfellow, UK). Before physicochemical characterization and adhesion assays were begun, the steel was cut into rectangular chips (1 by 1 cm) and cleaned by soaking for 10 min at 50 °C in a 2 % (vol/vol) solution of the commercial detergent rBS 35 (Société des Traitements Chimiques de Surface, lambersart, France), rinsing five times for 5 min in tap water and five times for 5 min in demineralized water.
Adhesion experiments
Solid surfaces were immersed in 25 ml of bacterial sus-pension containing approximately 108 CFU/ml, and adhe-sion assays were performed by sedimentation for 3 h at 20 °C in a petri dish (diameter, 10 cm) containing 1-cm2 stainless steel chips. The stainless steel chips were then rinsed to remove the non-adhering bacteria, and the surface was rinsed five times in naCl 1.5 × 10−3 mol/l. The stain-less steel chips were immersed in a test tube containing
10 ml of sterile 1.5 × 10−3 M naCl. Bacterial cells were detached from the inert support by using a sonication bath (Ultrasonik) for 2 min at 40 kHz and 35 °C. CFUs were counted by using the serial dilution technique and the bac-terial suspension obtained after sonication. Counts were determined on Trypticase Soya agar-Yeast extract (TSA-Ye) (Biomérieux), and the preparations were incubated for 24 h at 37 °C. each experiment was performed in triplicate by using two independently grown cultures.
Disinfection test
Benzalkonium chloride (BC) (Acros organics, USA) com-monly used in the food industry was used in this study:
Test on planktonic cells
Cell suspensions (1 ml) containing approximately 108 CFU/ml were inoculated into 9 ml of 0.25 mg/ml of BC solution at room temperature (20 ± 1.5 °C) and incu-bated for 5 min. One milliliter of BC–cell mixture was then immediately transferred into 9.0 ml of the neutralizer solu-tion containing 0.53 % lecithin (Fisher), 3.75 % Tween 80, and 0.05 % KH2PO4 (pH 7.0) (ren and Frank 1993) and then incubated for 5 min. For control samples, phosphate-buffered saline (50 mM phosphate buffer, pH 7.0, 0.85 % naCl) was used instead of the sanitizer. Viable cells in neu-tralizer solution were enumerated using the serial dilution technique as described above.
Test on sessile cells
The assay for the determination of the activities of the BC on cell attachment and biofilm formation was performed by sedimentation for 3 h at 20 °C in a petri dish (diameter, 10 cm) containing 1-cm2 stainless steel chips as described above. The stainless steel chips were then immersed by 10 ml of a BC solution used above (sanitizer concentra-tions were chosen to allow measurable changes in cell num-bers). After 5-min incubation, the sanitizer was removed, and the stainless steel chips were immersed in 10 ml of the neutralizer solution for 5-min incubation. Bacterial cells were detached from the inert support by using a sonication bath, and viable cells were enumerated as described above.
Statistical analysis
each analysis was performed using the SPSS. 13.0 Win-dows statistic package. The differences in the biofilm for-mation degree (Bacterial hydrophobicity assay) and cells adherence potency were examined by the Friedman test, followed by the Wilcoxon signed ranks test. P-values of <0.05 were considered as significant. Other analysis were
Arch Microbiol
1 3
realized between the strain origin, the slime production, and the cell sensitivity to disinfectant.
Results and discussion
Morphological variations in stressed strain
On the TSA-Ye plate, before any incubation in salmon at −20 °C, all the studied strains gave characteristics of L. monocytogenes cells: smooth, small size, transparent, not pigmented, and regular shape. Therefore, after the incu-bation in salmon at −20 °C, the grown colonies obtained on TSA-Ye shrank dramatically. Our previous study inves-tigating survival of L. monocytogenes in frozen salmon reported a 3.69 log reduction in L. monocytogenes cells in the salmon inoculated with a relative high level of contam-ination (108 CFU/g) and stored at −20 °C for 10 months (Miladi et al. 2008). These results indicate that frozen prod-ucts may carry L. monocytogenes even after 10 months of frozen storage if the products were contaminated with L. monocytogenes before freezing. Therefore, freezing can only extend the shelf life of products and retard the growth of bacteria, but not eliminate contaminated pathogens. On the other hand, another study investigating changes in freshness during frozen storage of Farmed Coho salmon showed that after a prolonged period of storage at −18 °C (9 and 12 months), all groups of salmon samples showed acceptable values for biochemical freshness indicators (Ortiz et al. 2012).
Cold starvation may cause changes in bacterial metabo-lism. In this study, no change in biochemical characteris-tics of tested pathogens was observed during three-month frozen storage (Miladi et al. 2008). So, no modification of biochemical characters of L. monocytogenes in fresh salmon was observed during the first three months of fro-zen storage (−20 °C). However, after ten months of freez-ing, we noticed that the number of bacteria begins to report
a significant diminution and evolutions of bacterial bio-chemical reactions was observed, which caused a false-negative identification of L. monocytogenes based on the analysis of biochemical characteristics (Miladi et al. 2008). However, the atypical L. monocytogenes isolate was identi-fied by PCr assay. Therefore, the presence of VBnC cells creates a major public health problem since they cannot be detected by traditional culturing methods and the cells remain potentially pathogenic under favorable conditions.
The impact of freezing on bacterial cells is multifold and has a general purpose in the food industry of suppressing bacterial growth although bacteria reside in a dormant state while being frozen. The response to starvation remains quite intricate and involves multiple changes within the cell. numerous morphological physiological changes in the cell may be attributed to the increase in protein production. The change to starvation state may induce a production of genes and proteins within the cell (Herbert and Foster 2001).
Alterations in cell morphology and topography due to starvation stress were examined via AFM (Fig. 1). The con-trol L. monocytogenes cells, whose length is about 2 μm, have a normal rod shape with a smooth surface (Fig. 1a), whereas the cells obtained after 10-month incubation in salmon at −20 °C have a coccoid shape <1 μm (Fig. 1b). It has been proposed that under limited exogenous nutri-ent conditions, member of L. monocytogenes can undergo a number of morphological and physiological changes to adapt to starvation conditions (roszak and Colwell 1987). Starvation survival has been defined as “the process of sur-vival in the absence of energy-yielding substrates” (Morita 1982). According to Morita (1993), several bacteria can survive for a long period under stressing environmental conditions owing to gradual changes in cellular physiol-ogy and morphology. So bacterial cells become smaller and more rounded during starvation. Similarly, during long-term survival, L. monocytogenes cells appear small and rounded as well. This rounding and shrinking may increase
Fig. 1 Atomic force micro-graphs of resuscitate Listeria monocytogenes cells. a control; b cells obtained after 10-month incubation in salmon at −20 °C
Arch Microbiol
1 3
the ability of the cell to absorb nutrients (Wen et al. 2009). Bakhrouf et al. (1994) showed that several bacteria such as Salmonella can survive in natural soil microcosm during 13 years with many morphological and metabolic modi-fications, among which the reduction in the cells size and their evolution to coccoid shapes. In fact, several gram-neg-ative species of bacteria, which usually live in oligotrophic environments, were usually found to have coccoid-shaped cells in their natural habitat (Kjelleberg et al. 1993). The reduction in the bacteria size, such as during the stress, is a strategy of survival to minimize the cell needs for nutrients (Jiang and Chai 1996). This reduction in the cells size is a result of the cytoplasmic contraction as well as the reduc-tion in the bacterial periplasm volume (Huisman et al. 1996). The metabolic activity of bacterial cells in a food-deficiency state can reach 0.05 % compared to the activity measured in the exponential phase (Kolter et al. 1993).
Determination of slime production
Phenotypic slime production was assessed by culturing the investigated strains on CrA plates. Among the three L. monocytogenes strains before freezing were no slime pro-ducer since they showed red colonies for reference strain and white colonies for S1 and S2 on CrA plates. After 10-month incubation in fresh minced salmon at −20 °C, different strains used in this study were a slime producer developing black colonies (Fig. 2). Indeed, slime produc-tion plays an important role in the pathogenesis of infec-tions caused by different microorganisms (Alcaráz et al. 2003) and is considered to be a significant virulence fac-tor for some staphylococci (Mack et al. 2000) as well as for Aeromonas spp, which indicates the high-risk source contamination (Sechi et al. 2002). Slimes are generally polysaccharidic materials, in spite of the probable pres-ence of other polymers. They are probably involved in the
protection of microbial cells. However, these molecules are also important in the biofilm formation on surfaces (Chae and Schraft 2000).
effect of freezing stress on aad mrnA abundance
After freezing stress, we observed an increase in the expres-sion level of aad gene in L. monocytogenes, and we noted that the expression level of this gene shows a significant increase (P < 0.05) after 10-month incubation in salmon at −20 °C (Fig. 3). These results confirm the qualitative study of the L. monocytogenes strains adhesion mentioned above.
Listeria adhesion protein has been established as an important adhesion factor for L. monocytogenes pathogen-esis (Jaradat et al. 2003), and several studies showed that the expression of Listeria adhesion protein (lAP) in which the genetic identity was determined to be alcohol acetalde-hyde dehydrogenase (aad) is influenced by growth temper-ature (Santiago et al. 1999) and nutrient-limiting conditions (Jaradat and Bhunia 2002).
Microbial adhesion to solvent
The affinity of the bacteria studied for the solvents was performed to evaluate the surface properties of each strain before and after 6 and 10 months of freezing stress between parent strains and their corresponding stressed strains after 6 and 10 months of freezing stress (Table 1). The higher affinity of parent strains to the acidic compound chloro-form when compared to hexadecane was an indicative of the predominance of basic properties on the cell surface indicating the hydrophilic nature of three parent strains. The affinity for decane was higher than the affinity for the basic compound ethyl acetate, showing the weak electron-accepting properties of L. monocytogenes in general. The results showed that all the parent strains had cell surfaces
Fig. 2 Colorimetric scale for colony analysis of slime production by L. monocytogenes using Congo red agar assay. a non-producing strain (red), b slime-producing strain (black) (color figure online)
Arch Microbiol
1 3
that were characterized as strongly basic and less acidic properties.
The percentage of bound cells to hexadecane was used to evaluate the hydrophobicity. According to Chae et al. (2006), cells could be strongly hydrophobic, when the bound cells to hexadecane were higher than 55 %, mod-erately hydrophobic (30–54 %), moderately hydrophilic (10–29 %), and strongly hydrophilic (<10 %). L. monocy-togenes showed the predominance of basic properties in parent strains. After 6-month incubation, we found no dif-ference of the adhesive ability of bacteria concerning their affinity for the solvents. When the bacterium was after 10-month incubation in salmon at −20 °C, the affinity of stressed strains to hexadecane was significantly (P < 0.05) increased. This was probably due to the hydrophobicity of stressed strain. However, it did not meant a prevalence of acidic properties, as the adhesion in ethyl acetate was lower when compared to decane. Although considered hydropho-bic in three stressed strains, the affinity for hexadecane was higher after incubation in fresh minced salmon at −20 °C for 10 months.
Thus, this study indicates that a change in cell surface hydrophobicity does appear to be involved in the adapta-tion response of L. monocytogenes to freezing stress. These results were in agreement with the data of giovannacci et al. (2000) that showed that the growth temperature and the phase of growth may influence the cell wall composi-tion, thereby modifying the surface electrical properties,
hydrophobicity, and electron-donor or electron-acceptor character of the bacteria. Thus, Ben Slama et al. (2012)founded that slime production of L. monocytogenes isolates is highly correlated with other properties responsible for its firm adherence such as hydrophobicity.
Changes in cellular fatty acid composition
Cell surface properties including cell hydrophobicity and membrane fluidity as impacted by fatty acid (FA) com-position can be altered through exposure to various envi-ronmental stresses. In addition, the ability of bacteria to change their membrane fluidity in response to different environmental stresses affects critical biochemical reac-tions (li et al. 2002; Yuk and Marshall 2003). Cell hydro-phobicity reflects the aggregate charge of the fatty acids and proteins at the cellular membrane. In this study, fatty acid profiles (Table 2) were determined for three parents and stressed L. monocytogenes strains before and after 6 and 10 months of freezing stress exposure in fresh minced salmon at −20 °C. The parent strains were characterized by a high proportion of iso- and anteiso-branched FAs having 15 and 17 carbon atoms. On the other hand, the stressed strains of 6 months of incubation do not show a large dif-ference, but the stressed strains of 10-month incubation in salmon at −20 °C presented a different FA profile charac-terized by a prevalence of straight FAs such as long-chain hexadecanoic acid (16:0), heptadecanoic acid (17:0), and
Fig. 3 Agarose gel analysis (1 %) of aad expression of Listeria monocytogenes before and after freezing stress in fresh cut salmon. (a) M, 100 bp DnA ladder (Promega); nC, negative control; 1, 2, and 3: strains before freezing stress; 4, 5, and 6: strains after freezing stress. The relative expression of aad gene. (b)
Table 1 Hydrophobicity results for parent and stressed strains of Listeria monocytogenes before and after 6 and 10 months of freezing stress exposure in salmon
SD Standard deviation, Listeria monocytogenes strains (ATCC 19115, S1 and S2)
octadecanoic acid (18:0), which were the principal fatty acids occurring in the strains after freezing stress exposure, and presented a percentage up to 75 % for reference strain L. monocytogenes ATCC 19115 and up to 80 % for isolated strains S1 and S2.
When cells of three strains were incubated for ten months in fresh minced salmon at −20 °C, the extent of branched fatty acid decreased and the membrane was less fluid, as indicated by decreases in the anteiso C15:0 fatty acids. In addition, we noted that freezing stress expo-sure reduced short-chain hydrophilic FA accumulation, but increased the long-chain hydrophobic FA (Table 2). Hence, decreased membrane fluidity by a significant reduction in anteiso fatty acids of stressed L. monocy-togenes cells might increase hydrophobicity (Table 1), which explains the adherent character of stressed strains. These results demonstrate that L. monocytogenes responds to freezing stress with an adjustment of its membrane fluidity. Mykytczuk et al. (2007) evaluated the cell mem-brane adjustments of various microorganisms in response to temperature stress. The authors found that cells adjust to different stresses through branching, unsaturation, and through changes in the length of their membrane fatty acids.
It has become clear in recent years that bacteria in nature do not prevalently exist in free floating cells but as biofilm (Costerton 1999). In effect, the primary source of food product contamination with pathogens appears to be the processing environments instead of the raw materials (Kathariou 2002). However, knowledge of the microbial physiology, and in particular the cell FA composition or stress response, is principally based on studies regarding planktonic cells. It has been frequently reported that the FA composition of L. monocytogenes is characterized by
the prevalence of branched FAs (Annous et al. 1997; Chi-hib et al. 2003). In particular, anteiso C15:0, iso C15:0, and anteiso C17:0 are the most abundant FAs. In fact, the psychrotrophic attitude of L. monocytogenes has been attributed to its relevant proportion of branched fatty acids, which presumably greatly contributes to maintain appropri-ate membrane fluidity at temperature near to 0 °C (Juneja and Davidson 1993; Püttman et al. 1993). The dramatic dif-ferences between the FA composition of stressed cells of the three strains taken into consideration suggests that FA membrane composition changes after freezing stress expo-sure, resulting the character of biofilm formation of strains. However, the almost total prevalence in adhered cells of straight FAs suggests that this peculiar physiological trait is functional to the new state of adhered cells. Due to the novelty of these findings, this phenomenon finds no basis in the current literature. The increase in straight FAs in adhered cells could be associated with a cell adaptation to a new environment (Baysse et al. 2005). It has been reported that the adhesion of L. monocytogenes to abiotic surfaces is associated with hydrophobic bonds between the food-pro-cessing plant surface and outer surface components of the cells (Brown 1996).
The importance of the freeze temperature on the adher-ent ability can be associated with the hydrophobicity and the hydrophobic/hydrophilic properties of free fatty acids. In fact, the adhesion of cells significantly increased when they were inoculated in fresh minced salmon at −20 °C for ten months. The FAs released presented different chain lengths and therefore different hydrophobicity lev-els. It is interesting to observe that the adhesive ability of stressed strains specifically released long-chain FAs like stearic acid (C18:0) and more increased hydrophobicity in general.
Table 2 Mean concentration and standard deviation of selected fatty acids of three parents and stressed strains of Listeria monocytogenes before and after 6 and 10 months of freezing stress exposure in salmon
Impact of adhesion on cell sensitivity to disinfectant
The adherence ability of L. monocytogenes strains after 3 h of contact between 108 CFU/ml of bacterial suspensions and stainless steel surface is shown in Fig. 4. For three tested strains, bacterial cells were bound more effectively to the stainless steel in which adhesion was greatest than 5 log CFU/cm2. Furthermore, for the reference strain L. monocytogenes ATCC 19115, adhesion was more impor-tant (5.65 log UFC/cm2) than the two food isolated strains S1 and S2 (P < 0.05), which are 5.20 log UFC/cm2 and 5.13 log UFC/cm2, respectively (Fig. 4).
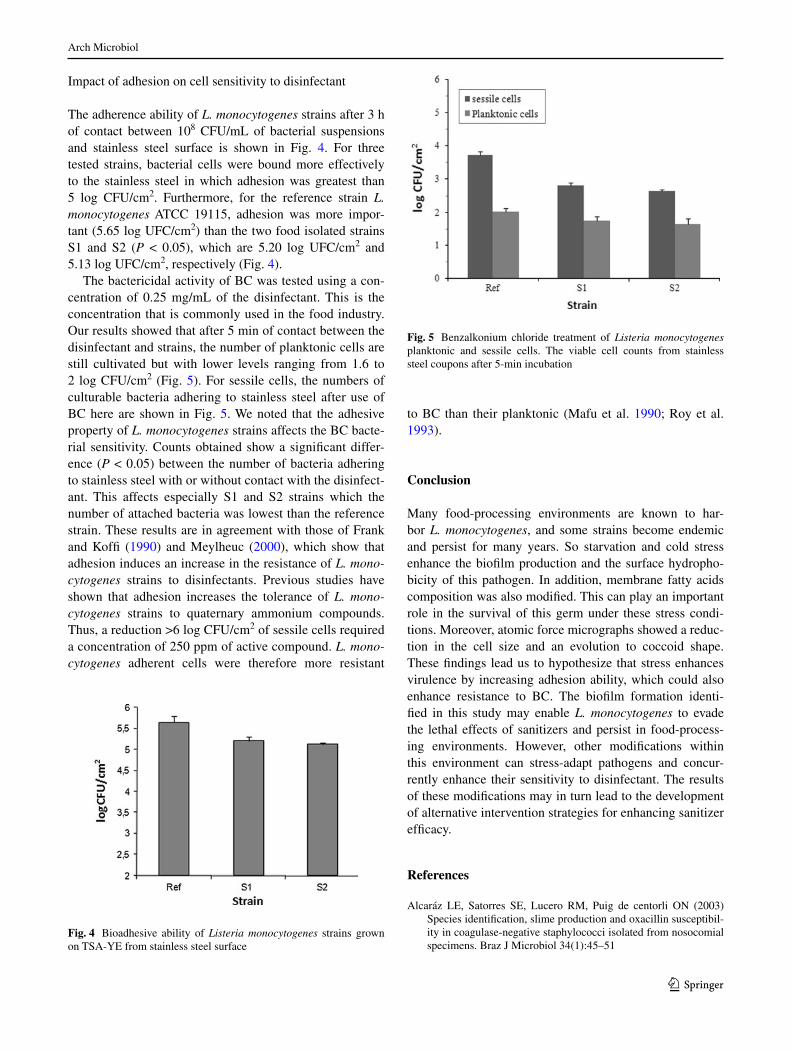
The bactericidal activity of BC was tested using a con-centration of 0.25 mg/ml of the disinfectant. This is the concentration that is commonly used in the food industry. Our results showed that after 5 min of contact between the disinfectant and strains, the number of planktonic cells are still cultivated but with lower levels ranging from 1.6 to 2 log CFU/cm2 (Fig. 5). For sessile cells, the numbers of culturable bacteria adhering to stainless steel after use of BC here are shown in Fig. 5. We noted that the adhesive property of L. monocytogenes strains affects the BC bacte-rial sensitivity. Counts obtained show a significant differ-ence (P < 0.05) between the number of bacteria adhering to stainless steel with or without contact with the disinfect-ant. This affects especially S1 and S2 strains which the number of attached bacteria was lowest than the reference strain. These results are in agreement with those of Frank and Koffi (1990) and Meylheuc (2000), which show that adhesion induces an increase in the resistance of L. mono-cytogenes strains to disinfectants. Previous studies have shown that adhesion increases the tolerance of L. mono-cytogenes strains to quaternary ammonium compounds. Thus, a reduction >6 log CFU/cm2 of sessile cells required a concentration of 250 ppm of active compound. L. mono-cytogenes adherent cells were therefore more resistant
to BC than their planktonic (Mafu et al. 1990; roy et al. 1993).
Conclusion
Many food-processing environments are known to har-bor L. monocytogenes, and some strains become endemic and persist for many years. So starvation and cold stress enhance the biofilm production and the surface hydropho-bicity of this pathogen. In addition, membrane fatty acids composition was also modified. This can play an important role in the survival of this germ under these stress condi-tions. Moreover, atomic force micrographs showed a reduc-tion in the cell size and an evolution to coccoid shape. These findings lead us to hypothesize that stress enhances virulence by increasing adhesion ability, which could also enhance resistance to BC. The biofilm formation identi-fied in this study may enable L. monocytogenes to evade the lethal effects of sanitizers and persist in food-process-ing environments. However, other modifications within this environment can stress-adapt pathogens and concur-rently enhance their sensitivity to disinfectant. The results of these modifications may in turn lead to the development of alternative intervention strategies for enhancing sanitizer efficacy.
References
Alcaráz le, Satorres Se, lucero rM, Puig de centorli On (2003) Species identification, slime production and oxacillin susceptibil-ity in coagulase-negative staphylococci isolated from nosocomial specimens. Braz J Microbiol 34(1):45–51
Fig. 4 Bioadhesive ability of Listeria monocytogenes strains grown on TSA-Ye from stainless steel surface
Fig. 5 Benzalkonium chloride treatment of Listeria monocytogenes planktonic and sessile cells. The viable cell counts from stainless steel coupons after 5-min incubation
Arch Microbiol
1 3
Annous BA, Becker lA, Bayles DO, labeda DP, Wilkinson BJ (1997) Critical role of anteiso-C15:0 fatty acid in the growth of Listeria monocytogenes at low temperatures. Appl environ Microb 63:3887–3894
Arciola Cr, Campoccia D, gamberini S, Cervellati M, Donati e, Montanaro l (2002) Detection of slime production by means of an optimised Congo red agar plate test based on a colourimetric scale in Staphylococcus epidermidis clinical isolates genotyped for ica locus. Biomaterials 23:4233–4239
Bakhrouf A, Zaafrane S, Mzoughi r, Maatoug K, gauthier MJ (1994) Phenotypic alteration of Salmonella isolated from healthy carri-ers in seawater. Marine life 4:3–8
Baysse C, Cullinane M, Dénervaud V, Burrowes e, Maxwell Dow J, Morrissey JP, Tam l, Trevors JT, O’gara F (2005) Modulation of quorum sensing in Pseudomonas aeruginosa through alteration of membrane properties. Microbiology 151:2529–2542
Bellon-Fontaine M, rault J, van Oss C (1996) Microbial adhesion to solvents: a novel method to determine the electron-donor/electron acceptor or lewis acid-base properties of microbial cells. Col-loids Surf B Biointerfaces 7:47–53
Ben Abdallah F, lagha r, Bakhrouf A (2008) resuscitation and mor-phological alterations of Salmonella bovismorbificans cells under starvation in soil. World J Microbiol Biotechnol 24:1507–1512
Ben Slama r, Bekir K, Miladi H, noumi A, Bakhrouf A (2012) Adhesive ability and biofilm metabolic activity of Listeria mono-cytogenes strains before and after cold stress. Afr J Biotechnol 11(61):12475–12482
Blackman IC, Frank JF (1996) growth of Listeria monocytogenes as a biofilm on various food-processing surfaces. J Food Protect 59:827–831
Braga PC, ricci D (1998) Atomic force microscopy: application to investigation of Escherichia coli morphology before and after exposure to cefodizime. Antimicrob Agents Chemother 42:18–22
Briandet r, Meylheuc T, Maher C, Bellon-Fontaine M-B (1999) Lis-teria monocytogenes Scott A: cell surface charge, hydrophobic-ity, and electron donor and acceptor characteristics under dif-ferent environmental growth conditions. App environ Microbiol 65:5328–5333
Brown M (1996) Studies on the adhesion of Listeria monocytogenes to leaf and model surfaces. Ph.D thesis. University of Manches-ter, Manchester
Chae MS, Schraft H (2000) Comparative evaluation of adhesion and biofilm formation of different Listeria monocytogenes strains. Int J Food Microbiol 62(1–2):103–111
Chae MS, Schraft H, Hansen lT, Mackereth r (2006) effects of phys-icochemical surface characteristics of Listeria monocytogenes strains on attachment to glass. Food Microbiol 23:250–259
Chihib ne, ribeiro da Silva M, Delattre g, laroche M, Federighi M (2003) Different cellular fatty acid pattern behaviours of two strains of Listeria monocytogenes Scott A and Cnl 895807 under different temperature and salinity conditions. FeMS Microbiol lett 218:155–160
Costerton JW (1995) Overview of microbial biofilms. J Ind Microbiol 15:137–140
Costerton JW (1999) Introduction to biofilm. Internat J Antimicrob Ag 11:217–221
Cox nA, Bailey JS, Mauldin JM, Blankenship lC (1997) The pres-ence of Listeria monocytogenes in the integrated poultry industry. J Appl Poultry res 6:116–119
Demel rA, Peelen T, Siezen rJ, De Kruijff B, Kuipers OP (1996) nisin Z, mutant nisin Z and lacticin 481 interactions with anionic lipids correlate with antimicrobial activity. A monolayer study. eur J Biochem 235:267–274
Di Bonaventura g, Piccolomini r, Paludi D, D’Orio V, Vergara A, Conter M, Ianieri A (2008) Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact
surfaces: relationship with motility and cell surface hydrophobic-ity. J Appl Microbiol 104:1552–1561
Donlan rM (2002) Biofilms: microbial life on surfaces. emerg Infect Dis 8(9):881–890
elabed H, Bakhrouf A, Hamza r, Azaiez M, gaddour K (2012) evi-dence of the adaptive response in Pseudomonas aeruginosa to 14 years of incubation in seawater. Ann Microbiol 62:1385–1394
Frank JF, Koffi rA (1990) Surface-adherent growth of Listeria mono-cytogenes is associated with increased resistance to surfactant sanitizers and heat. J Food Protect 5:550–554
Freeman DJ, Falkiner Fr, Keane CT (1989) new method for detect-ing slime production by coagulase negative staphylococci. J Clin Pathol 42:872–874
gandhi M, Chikindas Ml (2007) Listeria: a foodborne pathogen that knows how to survive. Int J Food Microbiol 113:1–15
giovannacci I, ermel g, Salvat g, Vendeuvre Jl, Bellon-Fontaine Mn (2000) Physicochemical surface properties of five Listeria monocytogenes strains from a pork-processing environment in relation to serotypes, genotypes and growth temperature. J Appl Microbiol 88(6):992–1000
gravani r (1999) Incidence and control of Listeria in food-processing facilities. In: ryser eT, Marth eH (eds) Listeria, listeriosis and food safety, 2nd edn. Marcel Dekker, new York, pp 657–709
Hébraud M, guzzo J (2000) The main cold shock protein of Listeria monocytogenes belongs to the family of ferritin-like proteins. FeMS Microbiol lett 190:29–34
Herbert KC, Foster SJ (2001) Starvation survival in Listeria monocy-togenes: characterization of the response and the role of known and novel components. Microbiology 147:2275–2284
Huisman gW, Siegele DA, Zambrano MM, Kolter r (1996) Morpho-logical and physiological changes during stationary phase. In: neidhardt FC, Curtiss III r, Ingraham Jl, lin eCC, low KB, Magasanik B, reznikoff WS, riley M, Schaechter M, Umbarger He (eds). Escherichia coli and Salmonella: cellular and molecu-lar biology. 2nd ed. Washington. p 1672–1682
Jaradat ZW, Bhunia AK (2002) glucose and nutrient concentra-tions affect the expression of a 104-kilodalton Listeria adhe-sion protein in Listeria monocytogenes. Appl environ Microbiol 68:4876–4883
Jaradat ZW, Wampler Jl, Bhunia AK (2003) A Listeria adhesion pro-tein-deficient Listeria monocytogenes strain shows reduced adhe-sion primarily to intestinal cell lines. Med Microbiol Immunol 192:85–91
Jiang X, Chai TJ (1996) Survival of Vibrio parahaemolyticus at low temperatures under starvation conditions and subsequent resus-citation of viable nonculturable cells. Appl environ Microbiol 62:1300–1305
Juneja VK, Davidson PM (1993) Influence of temperature on the fatty acid profile of Listeria monocytogenes. J rapid Meth Aut Mic 2:73–81
Kathariou S (2002) Listeria monocytogenes virulence and patho-genicity, a food safety perspective. J Food Protect 65:1811–1829
Kjelleberg S, Flfirdh K, nystrom T, Moriarty DJW (1993) growth limitation and starvation of bacteria. In: Ford T (ed) Aquatic microbiology: an ecological approach. Blackwell, Oxford, pp 289–320
Kolter r, Siegele DA, Tormo A (1993) The stationary phase of the bacterial life cycle. Annu rev Microbiol 47:855–874
larpent JP (1995) les Listeria. In: lavoisier (ed) Tec & Doc, Paris p 140
lewis K (2000) Programmed death in bacteria. Microbiol Mol Biol r 64:503–514
li J, Chikindas Ml, ludescher rD, Montville TJ (2002) Tempera-ture- and surfactant-induced membrane modifications that alter Listeria monocytogenes nisin sensitivity by different mecha-nisms. Appl environ Microbiol 68:5904–5910
Arch Microbiol
1 3
Mack D, rohde H, Dobinsky S, riedewald J, nedelmann M, Kno-bloch JK, elsner HA, Feucht HH (2000) Identification of three essential regulatory gene loci governing expression of the Staphy-lococcus epidermidis polysaccharide intercellular adhesion and biofilm formation. Infect Immun 68(7):3799–3807
Mafu AA, roy D, goulet J, Magny P (1990) Attachment of Listeria monocytogenes to stainless steel, glass, polypropylene and rubber surfaces after short contact times. J Food Prot 53:742–746
Meylheuc T (2000) Influence de biosurfactants sur l’adhésion de Lis-teria monocytiogenes à des surfaces inertes—Conséquences sur la désinfection. Université Paris XI, Thèse de doctorats 191p
Miladi H, Chaieb K, Bakhrouf A, elmnasser n, Ammar e (2008) Freezing effects on survival of Listeria monocytogenes in artificially contaminated cold fresh-salmon. Ann Microbiol 58(3):471–476
Miladi H, Chaieb K, Snoussi M, Ammar e, Bakhrouf A (2013) rT-PCr assays for the evaluation of the expression of Listeria mono-cytogenes virulence genes after cold and freezing shock. Afr J Microbiol res 7(8):612–619
Morita rA (1982) Starvation survival of heterotrophs in the marine environment. Adv Microb ecol 6:171–198
Morita rY (1993) Bioavailability of energy and the starvation state. In: Kjelleberg S (ed) Starvation in Bacteria. Plenum Press, new York, pp 1–23
Mykytczuk nCS, Trevors JT, leduc lg, Ferroni gD (2007) Fluo-rescence polarization in studies of bacterial cytoplasmic mem-brane fluidity under environmental stress. Prog Biophys Mol Bio 95:60–82
najjar MB, Chikindas M, Montville TJ (2007) Changes in Listeria monocytogenes membrane fluidity in response to temperature stress. Appl environ Microb 73:6429–6435
Ortiz J, Vivanco JP, Quitral V, larrain MA, Concha g, Aubourg SP (2012) Changes in freshness during frozen storage of farmed coho salmon: effect of replacement of synthetic antioxidants by natural ones in fish feeds. n Am J Aquacult 74:224–229
Püttman M, Ade n, Hof H (1993) Dependence of fatty acid com-position of Listeria spp. on growth temperature. res Microbiol 144:279–283
ren T, Frank JF (1993) Susceptibility of starved planktonic and bio-film Listeria monocytogenes to quaternary ammonium sanitizer as determined by direct viable and agar plate counts. J Food Prot 56:573–576
robbins JB, Fisher CW, Moltz Ag, Martin Se (2005) elimination of Listeria monocytogenes biofilms by ozone, chlorine, and hydro-gen peroxide. J Food Prot 68(3):494–498
rodriguez A, Autio Wr, Mclandsborough lA (2008) effects of con-tact time, pressure, percent relative humidity (%rH), and mate-rial type on Listeria biofilm adhesive strength at a cellular level using atomic force microscopy (AFM). Food Biophys 3:305–311
roszak DB, Colwell rr (1987) Survival strategies of bacteria in the natural environment. Microbiol rev 51:365–379
roy B, Ackermann HW, Pandian S, Picard S, goulet J (1993) Biolog-ical inactivation of adhering Listeria monocytogenes by listeri-aphages and a quaternary ammonium compound. Appl environ Microb 59:2914–2917
Santiago nI, Zipf A, Bhunia AK (1999) Influence of temperature and growth phase on expression of a 104- kilodalton Listeria adhe-sion protein in Listeria monocytogenes. Appl environ Microbiol 65:2765–2769
Schneiter r, Toulmay A (2007) The role of lipids in the biogen-esis of integral membrane proteins. Appl Microbiol Biotechnol 73:1224–1232
Sechi lA, Deriu A, Falchi MP, Fadda g, Zanetti S (2002) Distribu-tion of virulence genes in Aeromonas spp. isolated from Sardin-ian waters and from patients with diarrhoea. J Appl Microbiol 92(2):221–227
Swaminathan B, gerner-Schmidt P (2007) The epidemiology of human listeriosis. Microbes Infect 9:1236–1243
Tasara T, Stephen r (2006) Cold stress tolerance of Listeria mono-cytogenes: a review of molecular adaptive mechanisms and food safety implications. J Food Prot 69:1473–1484
Taylor CM, Beresford M, epton HAS, Sigee DC, Shama g, Andrew PW, roberts Ian S (2002) Listeria monocytogenes relA and hpt mutants are impaired in surface-attached growth and virulence. J Bacteriol 184:621–628
To MS, Favrin S, romanova n, griffiths MW (2002) Postadaptational resistance to benzalkonium chloride and subsequent physico-chemical modifications of Listeria monocytogenes. Appl environ Microbi 68(11):5258–5264
Van loosdrecht MC, lyklema J, norde W, Schraa g, Zehnder AJ (1987) The role of bacterial cell wall hydrophobicity in adhesion. Appl environ Microbiol 53:1893–1897
Weiler C, Ifland A, naumann A, Kleta S, noll M (2013) Incorporation of Listeria monocytogenes strains in raw milk biofilms. Int J Food Microbiol 161:61–68
Wen J, Anantheswaran r, Knabel S (2009) Changes in barotoler-ance, thermotolerance, and cellular morphology throughout the life cycle of listeria monocytogenes. Appl environ Microb 75:1581–1588
Wong AC (1998) Biofilms in food processing environments. J Dairy Sci 81(10):2765–2770
Yuk Hg, Marshall Dl (2003) Heat adaptation alters Escherichia coli O157:H7 membrane lipid composition and verotoxin production. Appl environ Microbiol 69:5115–5119