Page 1
Information Theory of DNA Sequencing
David Tse
Dept. of EECS
U.C. Berkeley
LIDS Student Conference
MIT
Feb. 2, 2012
Research supported by NSF Center for Science of Information.
Guy Bresler Abolfazl Motahari
Page 2
DNA sequencing
DNA: the blueprint of life
Problem: to obtain the sequence of nucleotides.
…ACGTGACTGAGGACCGTGCGACTGAGACTGACTGGGTCTAGCTAGACTACGTTTTATATATATATACGTCGTCGTACTGATGACTAGATTACAGACTGATTTAGATACCTGACTGATTTTAAAAAAATATT…
courtesy: Batzoglou
Page 3
Impetus: Human Genome Project
1990: Start
2001: Draft
2003: Finished3 billion basepairs
courtesy: Batzoglou
Page 4
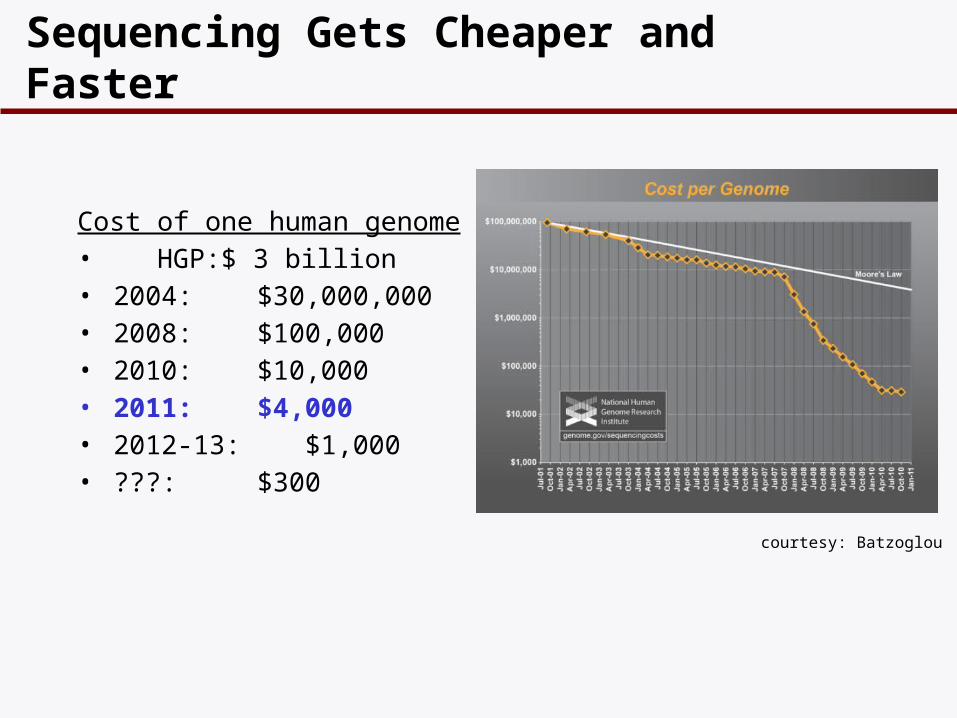
Sequencing Gets Cheaper and Faster
Cost of one human genome• HGP: $ 3 billion• 2004: $30,000,000• 2008: $100,000• 2010: $10,000• 2011: $4,000 • 2012-13: $1,000• ???: $300
courtesy: Batzoglou
Page 5
But many genomes to sequence
100 million species(e.g. phylogeny)
7 billion individuals (SNP, personal genomics)
1013 cells in a human(e.g. somatic mutations
such as HIV, cancer) courtesy: Batzoglou
Page 6
Whole Genome Shotgun Sequencing
Reads are assembled to reconstruct the original DNA sequence.
Page 7
Sequencing Technologies
• HGP era: single technology (Sanger)
• Current: multiple “next generation” technologies (eg. Illumina, SoLiD, Pac Bio, Ion Torrent, etc.)
• All provide massively parallel sequencing.
• Each technology has different read lengths, noise profiles, etc
Page 8
Assembly Algorithms
• Many proposed algorithms.
• Different algorithms tailored to different technologies.
• Each algorithm deals with the full complexity of the problem while trying to scale well with the massive amount of data.
• Lots of heuristics used in the design.
Page 9
A Basic Question
• What is the minimum number of reads needed to reconstruct with a given reliability?
• A benchmark for comparing different algorithms.
• An algorithm-independent basis for comparing different technologies and designing new ones.
Page 10
Coverage Analysis
• Pioneered by Lander-Waterman
• What is the minimum number of reads to ensure there is no gap between the reads with a desired prob.?
• Only provides a lower bound.
• Can one get a tight lower bound?
Page 11
Communication and Sequencing: An Analogy
Communication:
Sequencing:
Page 12
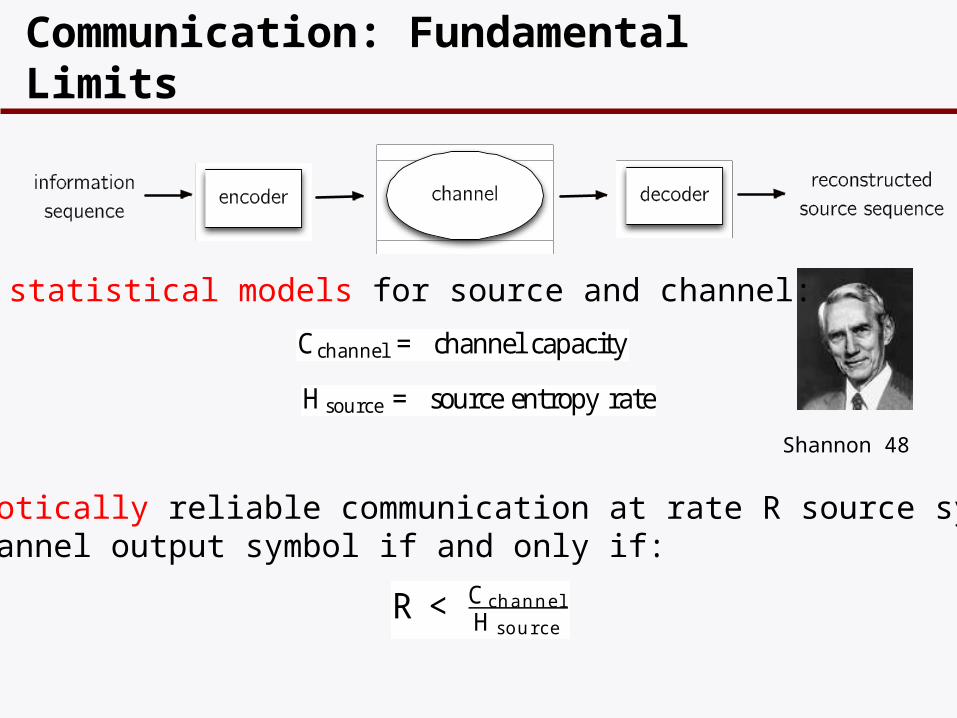
Communication: Fundamental Limits
Cchannel = channel capacity
Hsource = sourceentropy rate
Shannon 48
Asymptotically reliable communication at rate R source symbols per channel output symbol if and only if:
R < C channelH source
Given statistical models for source and channel:
Page 13
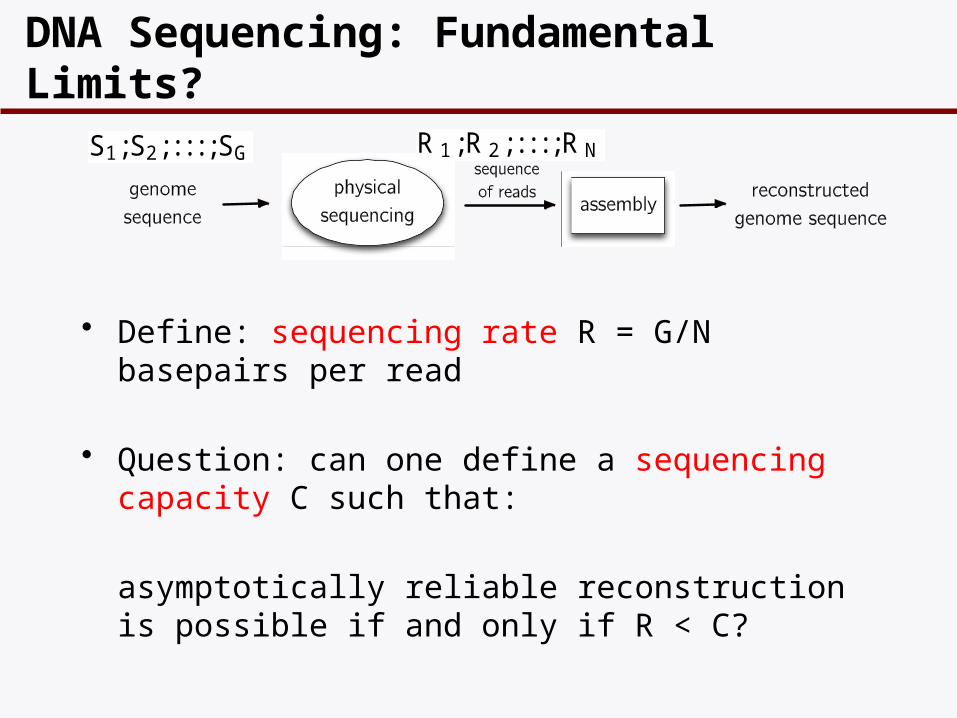
DNA Sequencing: Fundamental Limits?
S1;S2; : : : ;SG R 1;R 2; : : : ;R N
• Define: sequencing rate R = G/N basepairs per read
• Question: can one define a sequencing capacity C such that:
asymptotically reliable reconstruction is possible if and only if R < C?
Page 14
A Simple Model
• DNA sequence: i.i.d. with distribution p.
• Starting positions of reads are i.i.d. uniform on the DNA sequence.
• Read process is noiseless.
Will extend to more complex source model and noisy read process later.
Page 15
The read channel
• Capacity depends on
– read length: L
– DNA length: G
• Normalized read length:
• Eg. L = 100, G = 3 £ 109 :
read channel
AGCTTATAGGTCCGCATTACC AGGTCC
¹L :=L
logG
L ") C "
G ") C #
¹L = 4:6
Page 16
Result: Sequencing Capacity
H2(p) = ¡ log4X
i=1
p2i
Renyi entropy of order 2
C = 0
C = ¹L
Page 17
Coverage Constraint
TL
Starting positions of reads ~ Poisson(1/R)
E [# of gaps]= N ¢P [T > L]= Ne¡ LR
R = GN
E [# of gaps] ! 0
, R < ¹L
G
N reads
Page 18
No-Duplication Constraint
E [# of duplicated pairs]¼N 2 ¢
Ã4X
i=1
p2i
! L
L L L L
E [# of duplicated pairs] ! 0
, ¹L >2
H2(p)
= N 2e¡ L H 2(p)
The two possibilities have the same set of length L subsequences.
Page 19
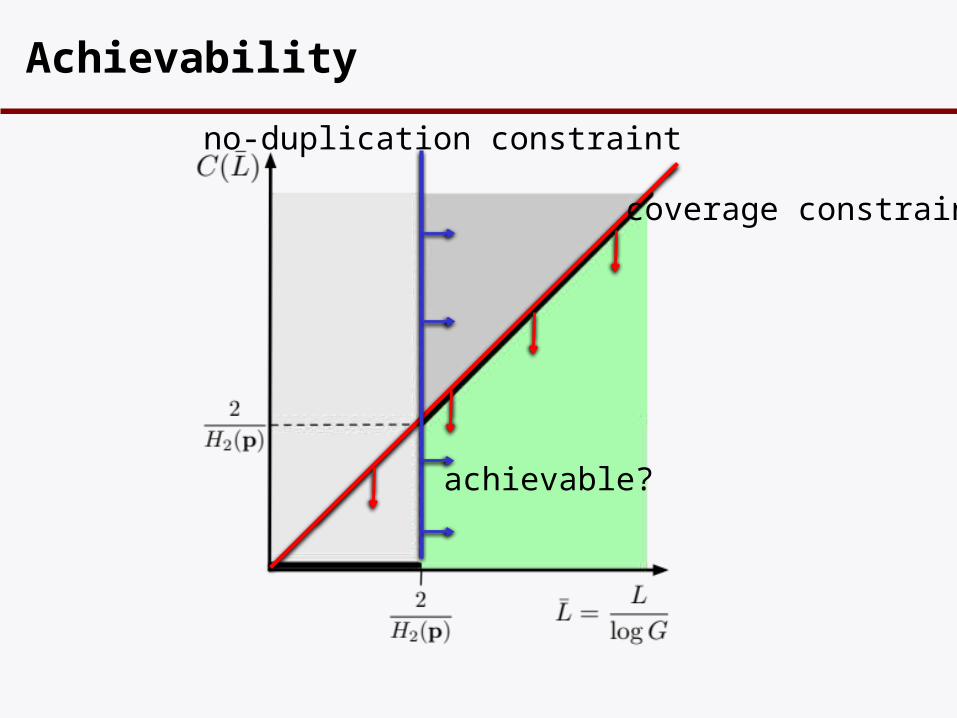
Achievability
coverage constraint
no-duplication constraint
achievable?
Page 20
Greedy Algorithm
Input: the set of N reads of length L
1. Set the initial set of contigs as the reads.
2. Find two contigs with largest overlap and merge them into a new contig.
3. Repeat step 2 until only one contig remains or no more merging can be done.
Algorithm progresses in stages:
at stage
merge reads at overlap `= L ¡ 1;L ¡ 2;: : : ;0
`
Page 21
Greedy algorithm: the beginning
gap
Most reads have large overlap with neighbors
Expected # of errors in stage L-1:
probability two disjoint reads are equal
Very small since no-duplication constraint is satisfied.
Page 22
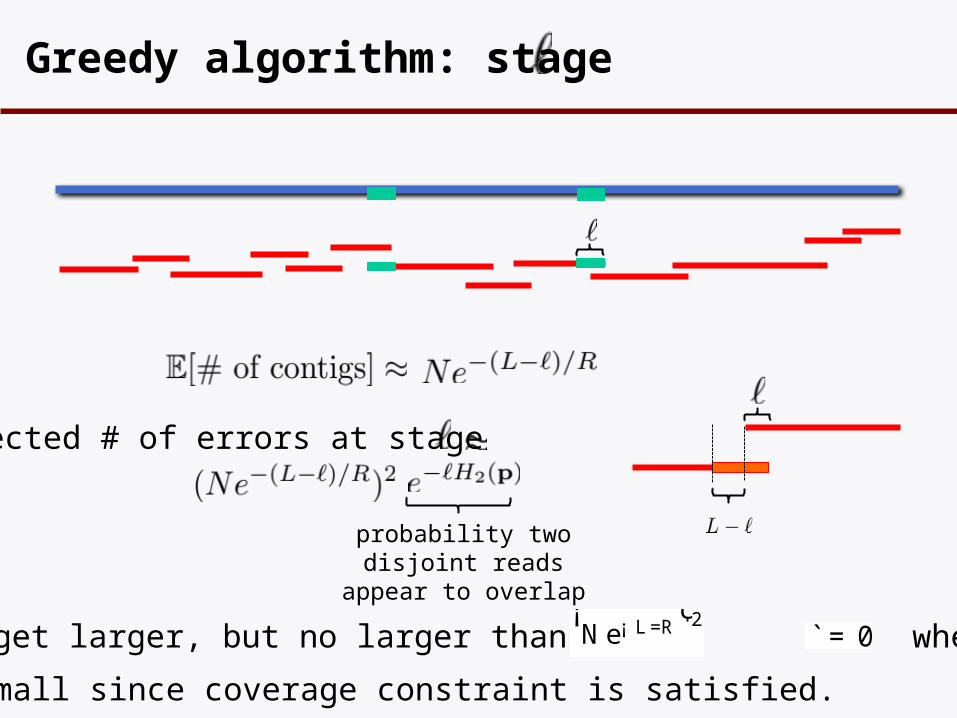
Greedy algorithm: stage
probability two disjoint reads appear to overlap
Expected # of errors at stage
This may get larger, but no larger than when¡Ne¡ L =R
¢2
Very small since coverage constraint is satisfied.
`= 0
Page 23
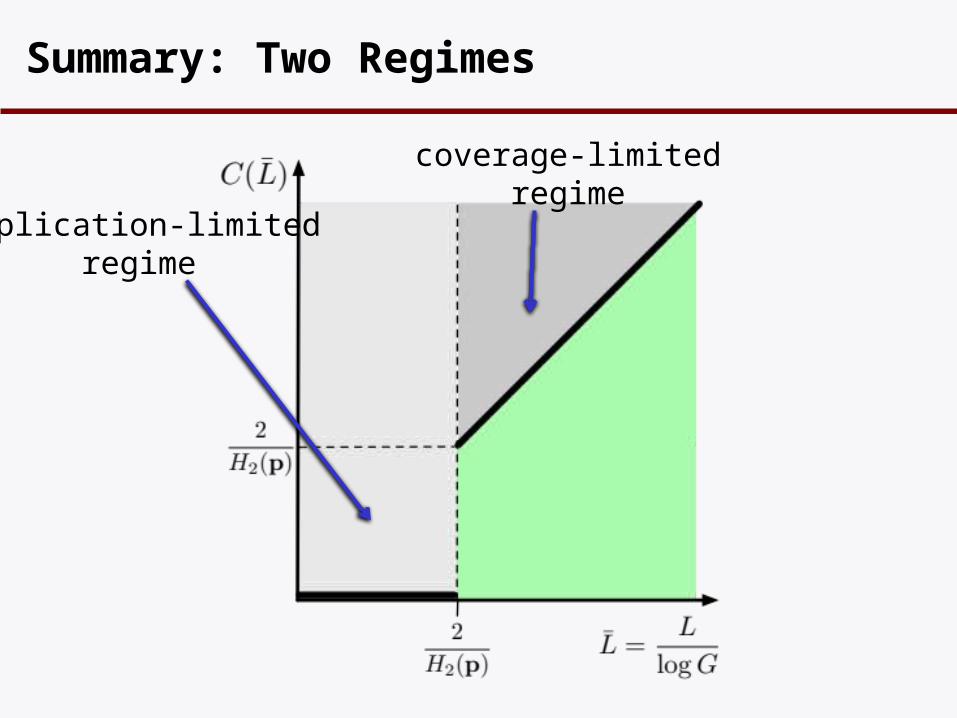
Summary: Two Regimes
coverage-limitedregime
duplication-limitedregime
Page 24
Relation to Earlier Works
• Coverage constraint: Lander-Waterman 88
• No-duplication constraint: Arratia et al 96
• Arratia et al focused on a model where all length L subsequences are given (seq. by hybridization)
• Our result: the two constraints together are necessary and sufficient for shotgun sequencing.
Page 25
Rest of Talk
• Impact of read noise.
• Impact of repeats in DNA sequence
Page 26
Read Noise
Model:
discrete memoryless channel defined by transition probabilities
ACGTCCTATGCGTATGCGTAATGCCACATATTGCTATGCGTAATGCGTTATACTTA
Page 27
Modified Greedy algorithm
Y
X
This is a hypothesis testing problem!
We observe two strings: X and Y.
Are they noisy versions of the same DNA subsequence?
Or from two different locations?
Y
Do we merge the two reads at overlap ?
(merge)
(do not merge)
Page 28
Impact on Sequencing Rate
MAP rule: declare H0 if
• Hypothesis test:
H0: noisy versions of the same DNA subsequence (merge)
H1: from disjoint DNA subsequences (do not merge)
Y
X Y
Two types of error:
• missed detection (new type of error)
• false positive (same as before)
Page 29
coverage constraint
no-duplication constraint
obtained by optimizing MAP threshold
Impact on Sequencing Rate
Page 30
More Complex DNA Statistics
• i.i.d. is not a very good model for the DNA sequence.
• More generally, we may want to model it as a correlated random process.
• For short-scale correlation, H2(p) can be replaced by the Renyi entropy rate of the process.
• But for higher mammals, DNA contains long repeats, repeat length comparable or longer than reads.
• This is handled by paired-end reads in practice.
H2 = lim`! 1
¡1`logP (x` = y`)
Page 31
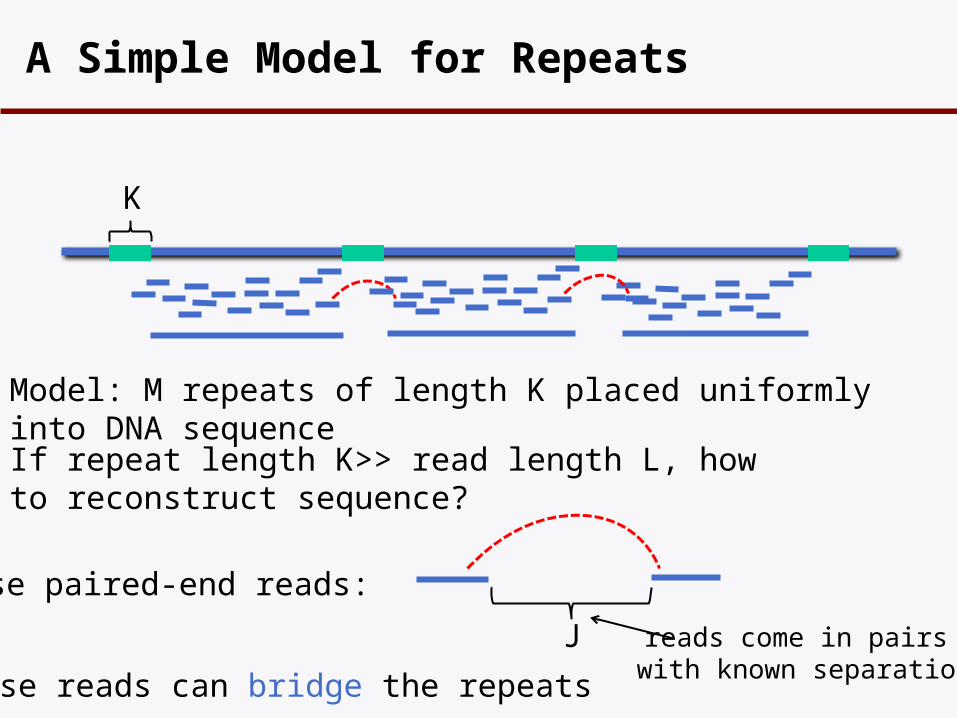
A Simple Model for Repeats
K
Model: M repeats of length K placed uniformly into DNA sequence
If repeat length K>> read length L, how to reconstruct sequence?
Use paired-end reads:
J reads come in pairs with known separation
These reads can bridge the repeats
Page 32
coverage constraint
no-duplication constraint
Impact on Sequencing Rate
coverage of repeatsconstraint
If J > 2d + Kthen capacityis the same aswithout repeats
constant indep of K
K= repeat lengthJ = paired-end
separation
Page 33
Conclusion
• DNA sequencing is an important problem.
• Many new technologies and new applications.
• An analogy between sequencing and communication is drawn.
• A notion of sequencing capacity is formulated.
• A principled design framework?