1521-0103/350/2/435–443$25.00 http://dx.doi.org/10.1124/jpet.114.215145 THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 350:435–443, August 2014 Copyright ª 2014 by The American Society for Pharmacology and Experimental Therapeutics Inhibitory Effect of Baicalin on Collagen-Induced Arthritis in Rats through the Nuclear Factor–kB Pathway Hong-Zhi Wang, Hai-He Wang, Shi-Shun Huang, Hong Zhao, Yong-Gang Cao, Guang-Zhi Wang, Dong Wang, Zhi-Gang Wang, and Yan-Hong Liu Department of Laboratory Diagnosis (H.-Z.W., Y.-H.L.), The Second Affiliated Hospital of Harbin Medical University, Harbin, China; Departments of Laboratory Diagnosis (H.-Z.W., G.-Z.W.,), Department of Respiratory Medicine (H.Z.), and Department of Orthopaedic Surgery (D.W.), The Fifth Affiliated Hospital of Harbin Medical University, Daqing, China; College of Medical Laboratory Science and Technology and the Key Laboratory of Molecular Diagnosis in Laboratory Medicine (H.-H.W., S.-S.H., Z.-G.W.), and Department of Pharmacology (Y.-G.C.), Harbin Medical University, Daqing, China Received April 17, 2014; accepted May 29, 2014 ABSTRACT This study focused on the potential therapeutic effect of baicalin on collagen-induced arthritis (CIA) in rats and the underlying mechanisms. The CIA rats were injected with baicalin (50, 100, or 200 mg/kg) once daily for 30 days. The rats were monitored for clinical severity of arthritis, and joint tissues were used for radiographic assessment and histologic examination. We quantified tumor necrosis factor–a (TNF-a) and interleukin-1b (IL-1b) in experimental animals and used Western blots to assess levels of protein abundance, phosphorylation, and acetylation of nuclear factor (NF)-kB p65 and sirtuin 1 (sirt1) protein expression in joint tissues. Human fibroblast-like synoviocytes from rheumatoid arthritis (HFLS-RA) were adopted in further mechanistic investiga- tions. Baicalin intraperitoneal injection for 30 days dose-dependently blocked clinical manifestations of CIA, such as functional impair- ment and swollen red paws. Meanwhile, it alleviated collagen- induced joint inflammation injury and inhibited the secretion of TNF-a and IL-1b in both rat synovium and HFLS-RA. Further mechanistic investigations revealed that baicalin suppresses NF-kB p65 protein expression and phosphorylation in synovial tissue and human-derived synoviocytes. Moreover, the acetylation of NF-kB p65 was downregulated by baicalin, which negatively correlates with the baicalin-induced upregulation of sirt1 expression in the same conditions. The data indicate that CIA in rats can be alleviated by baicalin treatment via relieving joint inflammation, which is related to the suppression of synovial NF-kB p65 protein expression and the elevation of its deacetylation by sirt1. Introduction Rheumatoid arthritis (RA) is a common autoimmune disease characterized by chronic inflammation of synovial membranes and proliferation of the synovial lining, leading to synovial hyperplasia, vasculogenesis, cartilage and bone destruc- tion, and joint malformation (Yang et al., 2010; Luo et al., 2011). Currently, RA affects 1% of the adult population worldwide, leading to serious loss of quality of life (Feldmann et al., 1996). Several biologic agents, including tumor necrosis factor (TNF)-a inhibitors, interleukin (IL)-1 receptor antago- nists, and nonsteroidal anti-inflammatory drugs, have proved clinically effective in RA patients. However, given the cost of such biologic agents and their limited efficacy in some pa- tients (Genovese et al., 2002; Zink et al., 2005), there has been a great demand for the development of novel therapeutic agents with fewer toxic effects. Although the etiology of RA is not completely understood, synovial cells are known to play a fundamental role in joint damage during RA. It has been clearly demonstrated that nuclear factor (NF)-kB is highly activated and involved in the pathogenesis of RA and animal models of experimental arthritis. NF-kB activation may enhance recruitment of inflammatory cells and production of proinflammatory mediators (Tsao et al., 1997; Miagkov et al., 1998; Tak and Firestein, 2001). IL-1b and TNF-a play critical roles in the pathophysiology of RA (Ivashkiv, 1996; Han et al., 1998; Odeh, 1998; Lee et al., 2008). NF-kB controls the expression of gene products that affect important cellular responses, such as inflammation, immunity, cell proliferation, and apoptosis. The mammalian NF-kB protein family has five known members: p65 (RelA), p50 (NF-kB 1), p52 (NF-kB 2), Rel B, and c-Rel. The most prevalent form of NF-kB is the p65/p50 heterodimer. NF-kB is activated by specific stimuli, such as proinflammatory cytokines, and the liberated NF-kB translocates into the nucleus to regulate the expression of various target genes (Karin and Ben-Neriah, 2000; Tak and Firestein, 2001; Lee et al., 2008). Recent studies have revealed additional posttranslational modifications of p65, including reversible phosphorylation and acetylation, which modulate NF-kB transcriptional activity on target genes (Kim et al., 2012). An important site of phosphorylation of the p65 subunit is Ser536 (phospho-Ser536-p65), which promotes p300 recruitment This work was supported by the Heilongjiang Education Department, China [Grant 12541523]; and China Postdoctoral Scientific Foundation [Grants 20100471022 and 201104420]. dx.doi.org/10.1124/jpet.114.215145. ABBREVIATIONS: CIA, collagen-induced arthritis; ELISA, enzyme-linked immunosorbent assay; HFLS, human fibroblast-like synoviocytes; IL-1b, interleukin-1b; MTX, methotrexate; NF-kB, nuclear factor–kB; RA, rheumatoid arthritis; sirt1, sirtuin 1; TNF-a, tumor necrosis factor–a. 435 at ASPET Journals on March 7, 2020 jpet.aspetjournals.org Downloaded from
Transcript
1521-0103/350/2/435–443$25.00 http://dx.doi.org/10.1124/jpet.114.215145THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 350:435–443, August 2014Copyright ª 2014 by The American Society for Pharmacology and Experimental Therapeutics
Inhibitory Effect of Baicalin on Collagen-Induced Arthritis in Ratsthrough the Nuclear Factor–kB Pathway
Hong-Zhi Wang, Hai-He Wang, Shi-Shun Huang, Hong Zhao, Yong-Gang Cao,Guang-Zhi Wang, Dong Wang, Zhi-Gang Wang, and Yan-Hong LiuDepartment of Laboratory Diagnosis (H.-Z.W., Y.-H.L.), The Second Affiliated Hospital of Harbin Medical University, Harbin,China; Departments of Laboratory Diagnosis (H.-Z.W., G.-Z.W.,), Department of Respiratory Medicine (H.Z.), and Department ofOrthopaedic Surgery (D.W.), The Fifth Affiliated Hospital of Harbin Medical University, Daqing, China; College of MedicalLaboratory Science and Technology and the Key Laboratory of Molecular Diagnosis in Laboratory Medicine (H.-H.W., S.-S.H.,Z.-G.W.), and Department of Pharmacology (Y.-G.C.), Harbin Medical University, Daqing, China
Received April 17, 2014; accepted May 29, 2014
ABSTRACTThis study focused on the potential therapeutic effect of baicalinon collagen-induced arthritis (CIA) in rats and the underlyingmechanisms. The CIA rats were injected with baicalin (50, 100,or 200 mg/kg) once daily for 30 days. The rats were monitoredfor clinical severity of arthritis, and joint tissues were usedfor radiographic assessment and histologic examination. Wequantified tumor necrosis factor–a (TNF-a) and interleukin-1b(IL-1b) in experimental animals and used Western blots to assesslevels of protein abundance, phosphorylation, and acetylation ofnuclear factor (NF)-kB p65 and sirtuin 1 (sirt1) protein expression injoint tissues. Human fibroblast-like synoviocytes from rheumatoidarthritis (HFLS-RA) were adopted in further mechanistic investiga-tions. Baicalin intraperitoneal injection for 30 days dose-dependently
blocked clinical manifestations of CIA, such as functional impair-ment and swollen red paws. Meanwhile, it alleviated collagen-induced joint inflammation injury and inhibited the secretion ofTNF-a and IL-1b in both rat synovium and HFLS-RA. Furthermechanistic investigations revealed that baicalin suppressesNF-kB p65 protein expression and phosphorylation in synovialtissue and human-derived synoviocytes. Moreover, the acetylationof NF-kB p65 was downregulated by baicalin, which negativelycorrelates with the baicalin-induced upregulation of sirt1 expressionin the same conditions. The data indicate that CIA in rats can bealleviated by baicalin treatment via relieving joint inflammation,which is related to the suppression of synovial NF-kB p65 proteinexpression and the elevation of its deacetylation by sirt1.
IntroductionRheumatoid arthritis (RA) is a common autoimmune disease
characterized by chronic inflammation of synovial membranesand proliferation of the synovial lining, leading to synovialhyperplasia, vasculogenesis, cartilage and bone destruc-tion, and joint malformation (Yang et al., 2010; Luo et al.,2011). Currently, RA affects 1% of the adult populationworldwide, leading to serious loss of quality of life (Feldmannet al., 1996). Several biologic agents, including tumor necrosisfactor (TNF)-a inhibitors, interleukin (IL)-1 receptor antago-nists, and nonsteroidal anti-inflammatory drugs, have provedclinically effective in RA patients. However, given the cost ofsuch biologic agents and their limited efficacy in some pa-tients (Genovese et al., 2002; Zink et al., 2005), there has beena great demand for the development of novel therapeuticagents with fewer toxic effects.Although the etiology of RA is not completely understood,
synovial cells are known to play a fundamental role in joint
damage during RA. It has been clearly demonstrated thatnuclear factor (NF)-kB is highly activated and involved in thepathogenesis of RA and animalmodels of experimental arthritis.NF-kB activation may enhance recruitment of inflammatorycells and production of proinflammatory mediators (Tsao et al.,1997; Miagkov et al., 1998; Tak and Firestein, 2001). IL-1b andTNF-a play critical roles in the pathophysiology of RA (Ivashkiv,1996; Han et al., 1998; Odeh, 1998; Lee et al., 2008).NF-kB controls the expression of gene products that affect
important cellular responses, such as inflammation, immunity,cell proliferation, and apoptosis. Themammalian NF-kB proteinfamily has five known members: p65 (RelA), p50 (NF-kB 1), p52(NF-kB 2), Rel B, and c-Rel. Themost prevalent form of NF-kB isthe p65/p50 heterodimer. NF-kB is activated by specific stimuli,such as proinflammatory cytokines, and the liberated NF-kBtranslocates into the nucleus to regulate the expression of varioustarget genes (Karin and Ben-Neriah, 2000; Tak and Firestein,2001; Lee et al., 2008). Recent studies have revealed additionalposttranslational modifications of p65, including reversiblephosphorylation and acetylation, which modulate NF-kBtranscriptional activity on target genes (Kim et al., 2012).An important site of phosphorylation of the p65 subunit isSer536 (phospho-Ser536-p65), which promotes p300 recruitment
This work was supported by the Heilongjiang Education Department, China[Grant 12541523]; and China Postdoctoral Scientific Foundation [Grants20100471022 and 201104420].
to the p65 complex and contributes to p65 acetylation mainly atLys310 (K310). Acetylation of Lys310 is required for the fulltranscriptional activity of NF-kB (Chen et al., 2002; Lanzillottaet al., 2010; Kim et al., 2012). Activation of NF-kB depends onthe balance between p65 acetylation and deacetylation.Baicalin, whose chemical structure is shown in Fig. 1A, is a
predominant flavonoid isolated from the dry root of Scutellariabaicalensis Georgi (Huang-Qin, a medicinal plant). It is con-sidered to be one of the effective and safe drugs widely used inAsia for the treatment of a variety of diseases, such as braindiseases, hepatic disorders, inflammatory diseases, and so on.Furthermore, it has been reported that baicalin has multiplebiologic functions, including anti-inflammatory, antioxidant,antiapoptotic, and immune regulation properties (Xu et al.,2011; Yin et al., 2011; Hou et al., 2012). There is increasingevidence to support the notion that this compound may have
potential roles in anti-inflammation and immune regulation(Liu et al., 2008). Our previous studies showed that baicalinprotects hippocampal neurons by upregulating the expressionof brain-derived neurotrophic factor and inhibiting the expres-sion of caspase-3 in a global ischemic gerbil model (Cao et al.,2011). Lee and Kim (2010) found that baicalin displayed anantioxidant effect in rheumatoid arthritis. Although baicalinwas previously observed to inhibit NF-kB activation in acuteand chronic inflammation models, no further studies of theNF-kB signaling pathway in light of the anti-inflammatoryeffects of baicalin in rheumatoid arthritis have been reported.Thus, the exact mechanism of its action remains to be clarified.Regarding the experimental arthritis models, in contrast to
lipopolysaccharide- or adjuvant-induced arthritis model, collagen-induced arthritis (CIA) has been widely used as a model ofhuman rheumatoid arthritis, since the CIA model shares
Fig. 1. Blocking of the development and progression of collagen-induced arthritis by treatment with baicalin. (A) Chemical structure of baicalin(C21H18O11; molecular weight 446.4). (B) Representative examples of rat hind paws. Treatment began on day 16 after the primary immunization andcontinued for 30 days (to day 46). (a) Naive group: rats without CIA, received physiologic saline 0.1 ml/100 g. (b) Vehicle-treated group: rats with CIA,received physiologic saline 0.1 ml/100 g, once daily. (c) MTX-treated group: rats with CIA, received MTX 1 mg/kg every 3 days. (d–f) Baicalin-treatedgroups, respectively, received baicalin 50, 100, and 200 mg/kg once daily. (C) Body weight changes of the rats. (D) The severity of arthritis was evaluatedby clinical arthritic scoring. (E) Hind paw thickness changes of the rats. Data are expressed as the mean 6 S.D. *P , 0.05 compared with the vehicle-treated group.
a number of clinical, immunologic, and pathologic featureswith RA (Shou et al., 2006). Therefore, human fibroblast-likesynoviocytes (HFLS)–RA and the collagen-induced arthritismodel, a well studied animal model of RA that has proveduseful in the development of new therapies for RA (Brandet al., 2003; Shou et al., 2006) were adopted in this study todetermine the potential therapeutic action of baicalin andidentify its underlying molecular mechanism.
Materials and MethodsAnimals. Female Wistar rats, 8 weeks old (180–200 g b.wt.), were
purchased from the Experimental Animal Center of Jilin University(Changchun, China). The animals were housed five per cage in a clearand ventilated environment maintained under laboratory conditions(temperature 22 6 1°C, relative humidity 50 to 70%, and 12-hour light/dark cycle). Standard food and water were provided ad libitum through-out the experiments. Animals were acclimated to their surroundingsover 5 days to eliminate the effect of stress prior to initiation of theexperiments. All animal experiments were performed according torelevant international experimental animal rules and ethical guidelines.
Induction of CIA in Rats and Baicalin Treatment. CIA wasinduced according to the method described previously with minormodification (Du et al., 2008). In brief, bovine type II collagen(Chondrex, Redmond, WA) was dissolved at 2 mg/ml in 0.05 M aceticacid by gently stirring overnight at 4°C. Collagen solutions wereemulsified with an equal volume of complete Freund’s adjuvant(Chondrex) with a homogenizer (Ronghua Instrument ManufacturingCo., JiangSu, China) on the ice water bath.We injected 0.2ml (collagen:200 mg) of the emulsion subcutaneously at the base of the tail,approximately 2 cm distal from the base. To ensure a high incidenceand severity of arthritis, a booster injection was given on day 7 afterinitial immunization. Freund’s incomplete adjuvant (Chondrex) wasapplied instead of Freund’s complete adjuvant to perform the secondaryimmunization. We prepared the collagen–incomplete Freund’s adju-vant emulsion as described above and injected 0.1 ml (100 mg collagen)of the emulsion subcutaneously into the tail (inserting the needle at3 cm from the base of the tail until the needle tip reached 1.5 cm fromthe base). In this model, the onset of arthritis in rats occurs within1 week after the second immunization. Starting on the day after thebooster injection, the rats were regularly monitored for the develop-ment and severity of paw inflammation. The primary immunizationday was defined as day 0. Body weight of rats was measured using anelectronic scale (Type ESJ200-4; Shenyang Longteng Electronic Co.,Shenyang, China) every 5 days from the primary immunization. Hindpaw thickness wasmeasured with electronic digital calipers (HanggongToolsManufacturing Co., Hangzhou, China) every 5 days, beginning onday 10. On day 16 after the primary immunization, rats displaying theonset of arthritis (arthritis index . 2) were randomly assigned to thefollowing groups (n 5 6): 1) vehicle-treated group: rats with CIA re-ceived physiologic saline (0.1 ml/100 g once daily i.p.); 2–4) baicalin-treated groups: rats with CIA were treated daily with intraperitonealbaicalin (Sigma-Aldrich, St. Louis, MO) with purity.95%, at the dosesof 50 mg/kg (group 2), 100 mg/kg (group 3), and 200 mg/kg (group 4); 5)methotrexate-treated group: rats with CIA were treated with metho-trexate (MTX) (Sigma-Aldrich), 1 mg/kg body weight every 3 days i.p.;and 6) naive group (untreated controls), six normal rats without CIAreceived physiological saline (0.1ml/100 g i.p.). Treatment continued for30 days. Baicalin was suspended in physiologic saline prior to ex-perimentation and injected intraperitoneally.
Clinical Assessment of Arthritis. Rats were inspected daily forthe onset of arthritis in the paws from the second immunization.Macroscopic signs of clinical arthritis were assessed by a qualitativeclinical score every 3 days beginning on the day when arthritic signswere first visible. Each paw was scored according to the followingcriteria: 0, normal; 1, mild redness and swelling of ankle or wrist joints;2, moderate redness and swelling of ankle or wrist joints; 3, severe
redness and swelling of the entire paw including digits; and 4, pawswith deformity or ankylosis (Alonzi et al., 1998). Themaximumscore fora single paw was 4 and for a single rat was 16; arthritis scores for allfour paws of each rat were summed as arthritis index. In a given group,the mean arthritis score for each group was calculated as the mean oftotal arthritis scores of all rats within the group. Arthritis index wasconducted under blinded conditions.
Radiographic Assessments. At day 46 after the first immuni-zation (i.e., day 30 after CIA rats were treated), the rats weresacrificed via anesthesia and the hind paw was obtained from thenormal and treated rats. The rats’ hind paw images were taken(MRAD-D50S RADREX-I; Toshiba Medical Manufacturing Co., Ltd.,Tokyo, Japan) to observe the radiologic changes. The X-ray param-eters were 40 kV, 100 mA, and 0.02 millisecond. Images were readindependently in a blinded fashion, and radiologic score was assessedaccording to the following criteria: 0) no radiologic changes were ob-served; 1) mild changes, with tissue swelling and edema; 2) moderatechanges, with joint erosion and disfiguration; and 3) severe changes,with bone erosion and osteophyte formation (Cai et al., 2007). Thetotal radiologic scores were calculated from the sum of both hind pawsof each rat; the maximum value was 6.
Histopathologic Assessments. For histologic analysis, the jointsof hind paw were removed and fixed in 4% paraformaldehyde for atleast 2 days. The joints of hind paw were then decalcified for 30 days in10% EDTA, the decalcification liquid was changed every 4 days.Afterward, the paws were embedded in paraffin blocks, longitudinallycut into 4-mm sections using microtome. To ensure extensive evaluationof the arthritic joints, at least three serial sections were cut, the sectionswere then splayed on water surface and mounted on microscope slides,and stained with H&E to study the degree of synovitis and bone erosionsby microscopic evaluation in a blinded manner. Histologic scores wereevaluated on the basis of infiltration of inflammatory cells, synovialhyperplasia, cartilage destruction, and bone erosions (Li et al., 2013). Therating criteria for studies on prognosis are as follows: 1) inflammatory cellinfiltration: 0 5 normal, 1 5 mild infiltration, 2 5 moderate infiltration,and 35 severe infiltration (large number of inflammatory cells wereobserved); 2) synovial hyperplasia: 0 5 no hyperplasia, 1 5 mildhyperplasia, 25moderate hyperplasia, and 35 severe hyperplasia;(3) destruction of cartilage: 05 no destruction, 15mild destruction,2 5 moderate destruction, and 3 5 severe destruction plus vas-culogenesis; and 4) erosions of bone: 0 5 no erosions, 1 5 milderosions, 2 5 moderate erosions, and 3 5 severe erosions (extendederosions and destruction of bone).
Cells and Culture Conditions. Human fibroblast-likesynoviocytes–rheumatoid arthritis were purchased from Cell Applica-tions, Inc. (San Diego, CA). Cells were cultured in synoviocyte growthmedium (Cell Applications) supplemented with 100 IU/ml penicillinand 100 mg/ml streptomycin at 37°C in a humidified incubator with 5%carbon dioxide and 95%air.HFLS-RA between passages four and sevenwere used for subsequent experiments, during which time they werea homogeneous population of synoviocytes.
Cell Treatment. The synoviocyte growth medium was replacedwith serum-free medium for an additional 24-hour culture beforefurther treatments. HFLS-RA were suspended at 2 � 106 cells/ml forexperiments. In the mechanism experiments, HFLS-RA treated withserum-free medium only served as the control group. HFLS-RA weretreated with different concentrations of baicalin (10, 20, or 30 mM) inDulbecco’s modified Eagle’s medium for 24 hours.
HFLS-RA Proliferation Assay by 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyl-2H-Tetrazolium Bromide. HFLS-RA were cul-tured using the above method and suspended at 2� 106 cells/ml. Cellswere seeded into 96-well plates (100 ml/well) and incubated for24 hours before various concentrations of baicalin (10, 20, 30, 40, 50,or 60 mM) were added for 48 hours. According to the manufacturer’srecommendations, 20 ml of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide solution (0.5 mg/ml in phosphate-bufferedsaline) (Sigma-Aldrich) was added to eachwell and incubated with cellsunder standard conditions for 4 hours. Subsequently, the formazan
Therapeutic Effects of Baicalin on CIA in Rats 437
crystals in each well were dissolved with dimethylsulfoxide after themedium was removed. Finally, the optical density was measured withthe enzyme-linked immunosorbent assay (ELISA) microwell reader(Bio-Rad, Hercules, CA) at 490 nm, and the results were expressed asthe mean of triplicate wells.
Measurements of TNF-a and IL-1b Levels in Serum andCulture Supernatants by ELISA. At day 46 after the first im-munization, the rats were anesthetized with an intraperitonealinjection of 10% chloral hydrate (0.3 ml/100 g). Serum samples werecollected and stored at 280°C until used. To determine cytokine levelsin vitro, HFLS-RA were treated with or without baicalin using theabovemethod; supernatants were harvested from eachwell. TNF-a andIL-1b levels in serum and culture supernatants were measured usingrat TNF-a and IL-1b ELISA kits (R&D Systems, Minneapolis, MN)according to the manufacturer’s instructions.
Western Blotting Detections. Western blot analysis was per-formed as previously described (Wang et al., 2010) for measuringNF-kB p65, phospho-NF-kB p65 (Ser536), acetyl-NF-kB p65 (Lys310),and sirtuin 1 (sirt1). Briefly, total proteins were extracted fromthe synovium of rats and HFLS-RA lysates, thereafter separated bySDS-PAGE and transferred to polyvinylidene fluoride membranes.The membranes were blocked in 5% skim milk in phosphate-bufferedsaline–Tween at room temperature for 2 hours, and probed withanti–NF-kB p65, anti–phospho-NF-kB p65, anti–acetyl-NF-kB p65,and anti-sirt1 (Cell Signaling Technology, Danvers, MA). Horseradishperoxidase–conjugated secondary antibodies (Sigma-Aldrich) and anenhanced chemiluminescence substrate kit were used in detection ofspecific proteins. b-Actin was used as the loading control.
Statistical Analysis. Data were expressed as mean 6 S.D. andanalyzed with GraphPad Prism version 5.0 (GraphPad Software, LaJolla, CA). Comparisons were by one-way analysis of variance with theTukey post hoc test. P , 0.05 was regarded as statistically significant.
ResultsSignificant Blocking of the Progression of CIA by
Treatment with Baicalin. In this study, we used thecollagen-induced arthritis model to define the therapeuticeffects of baicalin. Relative to other experimental arthritismodels, CIA has been widely used as a model of humanrheumatoid arthritis, since the CIA model shares a numberof clinical, immunologic, and pathologic features with RA(Brand et al., 2003; Shou et al., 2006).The onset of arthritis in rats occurs within 1 week after the
second immunization. Treatment began on day 16 after theprimary immunization and continued until day 46. It consistedof intraperitoneal injections of 50, 100, and 200 mg/kg baicalin,1 mg/kg methotrexate, or physiological saline. Clinical mani-festations, such as functional impairment and swollen redpaws, were observed (Fig. 1B). Some infected rats exhibitedlusterless hair and slow body weight gain (Fig. 1C). Arthritisstarted to develop and worsen over time in vehicle-treated CIArats. As shown in Fig. 1D, the therapeutic effect of baicalin wasdose-dependent, baicalin (100 and 200 mg/kg once daily)markedly reduced arthritic scores in the baicalin-treated grouprats comparedwith the vehicle-treated CIA rats, as assessed byclinical arthritic score. CIA rats treated with vehicle continuedto develop severe arthritis, and reached a plateau between days28 and 34. In contrast, CIA rats treated with baicalin showeda progressive decrease in the severity of arthritis; thisdifference was statistically significant (P , 0.01) at the end ofthe experiment (when rats were killed), especially for rats given200 mg/kg baicalin. A similar reduction in arthritic score wasobserved with 100 mg/kg baicalin and 1 mg/kg MTX in ratswith CIA. Similar changes were also observed in hind paw
thickness (Fig. 1E). Thus, our results suggest that baicalin canblock the progression of inflammatory arthritis.Significant Suppression of Collagen-Induced Joint
Inflammation Injury by Treatment with Baicalin. After30 days of treatment, the rats were sacrificed and hind limbswere obtained from both normal and treated rats. X-rayradiographs and histologic examinations were carried out tofurther assess the therapeutic effects of baicalin. Under X-rayimaging conditions, soft tissue swelling, cartilage and bonedestruction, joint narrowing, and bone loss were observed. Asshown in Fig. 2A, the rats without arthritis (naive group, Fig.2A-a) showed normal soft tissue, joint structure, and jointspace (arrow). The CIA rats treated with vehicle showed
Fig. 2. Therapeutic effects of baicalin on paw swelling and bone destructionin rats with collagen-induced arthritis. On day 46 after primary immuni-zation the rats were sacrificed, and hind limbs were obtained from normaland treated rats. (A) Representative macroradiographs of rat hind paws.Top arrows show the joint structure and joint space, and bottom arrowsshow the soft tissue. Neither paw swelling nor joint erosion were observed innormal rats (a). Severe paw swelling and bone erosion were seen in CIA ratstreated with vehicle (b). Moderate change was observed in rats treated withbaicalin 50mg/kg (d).Mild changewas observed in rats treated withMTX (c)and baicalin 100 mg/kg (e), but the damage was much less in rats treatedwith baicalin 200 mg/kg (f). (B) The radiologic scores of bone erosion intreated CIA rats were evaluated as described underMaterials andMethods.Data are expressed as mean6 S.D. (n = 6 per group). **P , 0.01 comparedwith vehicle-treated rats.
severe swelling of the soft tissues, bone erosion, and anarrowed joint space (Fig. 2A-b). Moderate change was ob-served in rats treated with baicalin 50 mg/kg, and mildchange was observed in rats treated with MTX and baicalin100 mg/kg. The X-ray films of rats in the baicalin 200 mg/kggroup revealed that the soft tissue of each immunized pawwas slightly swollen, and less joint destruction was evident incomparison with other treatment groups. Radiologic analysisrevealed severe joint erosion in the CIA rats, as shown in Fig.2B. The mean radiologic score in the vehicle-treated groupwas significantly higher than the scores for rats receiving50 mg/kg baicalin, 100 mg/kg baicalin, or MTX. The CIA ratsreceiving 200 mg/kg baicalin exhibited significant protection,with the lowest scores for bone erosion. The radiographicarthritic scores indicate that 200 mg/kg baicalin markedlysuppressed bone erosions and destruction of the joints.Further evidence to support the inhibitory effects of baicalin
on CIA was obtained by histopathology analysis of joints. Thenonimmunized rats showed normal architecture of the jointwith normal appearance of the cartilage lining, joint space,and the underling bones, without inflammatory infiltrates inthe synovial tissue (Fig. 3A-a). In vehicle-treated CIA rats,the histologic appearance of the joint was highly abnormal,with pronounced synovial hyperplasia, inflammatory cell
infiltration, pannus formation, and extensive erosion changes incartilage and bone (Fig. 3A-b). High-power views of the boxedarea show pannus formation with extensive articular cartilagedestruction as compared with the naive group. In contrast, CIArats treated with baicalin were largely spared the chronicinflammation of synovial tissue. These rats showed dose-dependent, significant reductions in inflammatory cell infiltra-tion, pannus formation, cartilage destruction, and bone erosioncompared with the vehicle-treated group (Fig. 3, A and D–F).The histologic scores of CIA rats that received baicalin weresignificantly lower than the scores of CIA rats treated withvehicle. These data demonstrate that baicalin relieved jointinflammation injury in CIA rats.Baicalin Inhibits the Secretion of TNF-a and IL-1b in
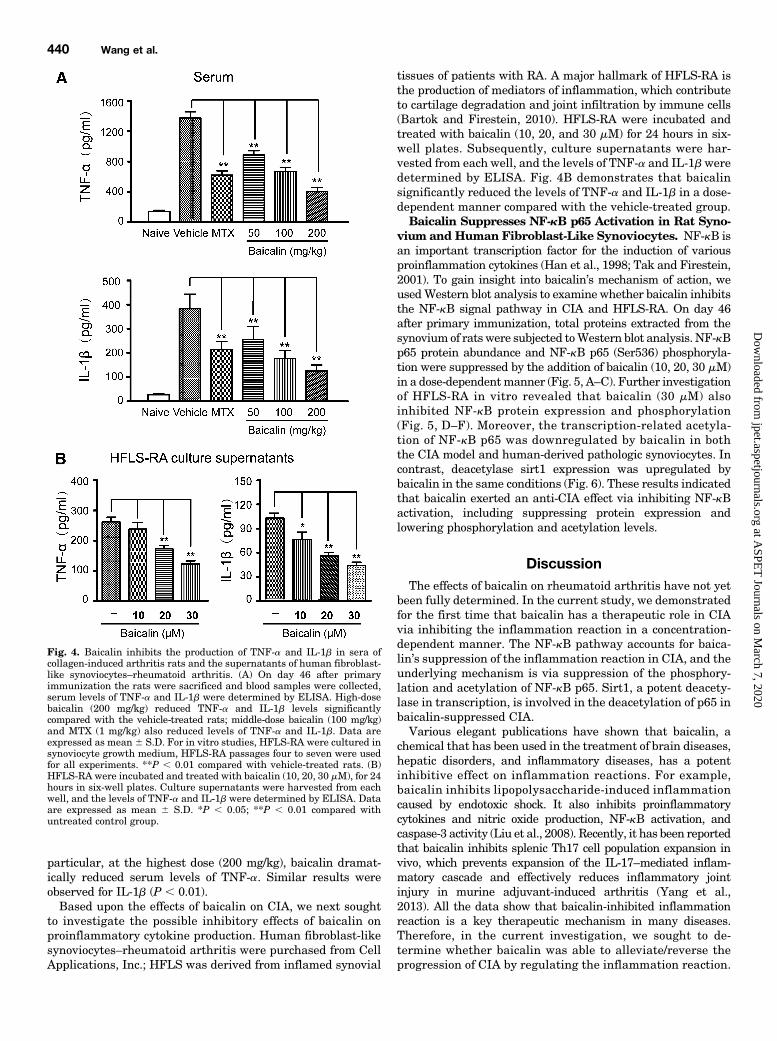
Rat Serum and Human Fibroblast-Like Synoviocytes.To investigate whether baicalin modulates the inflammatoryprocess by regulating the secretions of cytokines in vivo, wemeasured the serum levels of TNF-a and IL-1b in rats byELISA. As shown in Fig. 4A, substantial increases in pro-inflammatory cytokine levels were found in the serum samplesof vehicle-treated CIA rats on day 46 after primary immuni-zation. In contrast, TNF-a levels were significantly and dose-dependently lower in baicalin-treated groups (50, 100, and200 mg/kg) than in the vehicle-treated group (P , 0.01). In
Fig. 3. Therapeutic effects of baicalin on synovial inflammation and cartilage-bone destruction in rats with collagen-induced arthritis. On day 46 afterprimary immunization, the joints of hind paws were treated as described underMaterials and Methods. (A) The sections were stained with hematoxylinand eosin to study the degree of synovitis and cartilage-bone erosions. Histologic features of representative joints are shown for each group of rats.Further magnification of the black-bordered box (top) shows the typical inflammatory injuries (bottom). Naive group (a) shows normal cartilage, bone,and synoviumwithout inflammation. CIA rats treated with vehicle (b) showed pronounced synovial hyperplasia, inflammation, cell infiltration, cartilagedestruction, bone erosion, and pannus formation. A high-power view of the boxed area shows pannus formation with extensive articular cartilagedestruction. CIA rats treated with baicalin 200 mg/kg (f) exhibited well-preserved joint spaces and articular cartilage surfaces, with minimal pannusformation compared with the vehicle-treated group, indicating that the administration of baicalin directly correlated with a reduction in diseaseseverity. Mild changes in joint pathology were seen in CIA rats treated with MTX (c) and baicalin 100 mg/kg (e). Moderate change was observed in ratstreated with baicalin 50 mg/kg (d). bo, bone; ca, cartilage; sy, synovium; js, joint space; sh, synovial hyperplasia; ce, cartilage erosion; ic, inflammatorycells; pf, pannus formation. Original magnification, 10�; 20� in high-power views. (B) Histologic score was evaluated from the joints of CIA rats treatedwith or without baicalin on the basis of infiltration of inflammatory cells, synovial hyperplasia, pannus formation, cartilage destruction, and boneerosion. Data are expressed as mean 6 S.D. (n = 6 per group). *P , 0.05 compared with vehicle-treated rats.
Therapeutic Effects of Baicalin on CIA in Rats 439
particular, at the highest dose (200 mg/kg), baicalin dramat-ically reduced serum levels of TNF-a. Similar results wereobserved for IL-1b (P , 0.01).Based upon the effects of baicalin on CIA, we next sought
to investigate the possible inhibitory effects of baicalin onproinflammatory cytokine production. Human fibroblast-likesynoviocytes–rheumatoid arthritis were purchased from CellApplications, Inc.; HFLS was derived from inflamed synovial
tissues of patients with RA. A major hallmark of HFLS-RA isthe production of mediators of inflammation, which contributeto cartilage degradation and joint infiltration by immune cells(Bartok and Firestein, 2010). HFLS-RA were incubated andtreated with baicalin (10, 20, and 30 mM) for 24 hours in six-well plates. Subsequently, culture supernatants were har-vested from each well, and the levels of TNF-a and IL-1bweredetermined by ELISA. Fig. 4B demonstrates that baicalinsignificantly reduced the levels of TNF-a and IL-1b in a dose-dependent manner compared with the vehicle-treated group.Baicalin Suppresses NF-kB p65 Activation in Rat Syno-
vium and Human Fibroblast-Like Synoviocytes. NF-kB isan important transcription factor for the induction of variousproinflammation cytokines (Han et al., 1998; Tak and Firestein,2001). To gain insight into baicalin’s mechanism of action, weusedWestern blot analysis to examine whether baicalin inhibitsthe NF-kB signal pathway in CIA and HFLS-RA. On day 46after primary immunization, total proteins extracted from thesynovium of ratswere subjected toWestern blot analysis. NF-kBp65 protein abundance and NF-kB p65 (Ser536) phosphoryla-tion were suppressed by the addition of baicalin (10, 20, 30 mM)in a dose-dependentmanner (Fig. 5, A–C). Further investigationof HFLS-RA in vitro revealed that baicalin (30 mM) alsoinhibited NF-kB protein expression and phosphorylation(Fig. 5, D–F). Moreover, the transcription-related acetyla-tion of NF-kB p65 was downregulated by baicalin in boththe CIA model and human-derived pathologic synoviocytes. Incontrast, deacetylase sirt1 expression was upregulated bybaicalin in the same conditions (Fig. 6). These results indicatedthat baicalin exerted an anti-CIA effect via inhibiting NF-kBactivation, including suppressing protein expression andlowering phosphorylation and acetylation levels.
DiscussionThe effects of baicalin on rheumatoid arthritis have not yet
been fully determined. In the current study, we demonstratedfor the first time that baicalin has a therapeutic role in CIAvia inhibiting the inflammation reaction in a concentration-dependent manner. The NF-kB pathway accounts for baica-lin’s suppression of the inflammation reaction in CIA, and theunderlying mechanism is via suppression of the phosphory-lation and acetylation of NF-kB p65. Sirt1, a potent deacety-lase in transcription, is involved in the deacetylation of p65 inbaicalin-suppressed CIA.Various elegant publications have shown that baicalin, a
chemical that has been used in the treatment of brain diseases,hepatic disorders, and inflammatory diseases, has a potentinhibitive effect on inflammation reactions. For example,baicalin inhibits lipopolysaccharide-induced inflammationcaused by endotoxic shock. It also inhibits proinflammatorycytokines and nitric oxide production, NF-kB activation, andcaspase-3 activity (Liu et al., 2008). Recently, it has been reportedthat baicalin inhibits splenic Th17 cell population expansion invivo, which prevents expansion of the IL-17–mediated inflam-matory cascade and effectively reduces inflammatory jointinjury in murine adjuvant-induced arthritis (Yang et al.,2013). All the data show that baicalin-inhibited inflammationreaction is a key therapeutic mechanism in many diseases.Therefore, in the current investigation, we sought to de-termine whether baicalin was able to alleviate/reverse theprogression of CIA by regulating the inflammation reaction.
Fig. 4. Baicalin inhibits the production of TNF-a and IL-1b in sera ofcollagen-induced arthritis rats and the supernatants of human fibroblast-like synoviocytes–rheumatoid arthritis. (A) On day 46 after primaryimmunization the rats were sacrificed and blood samples were collected,serum levels of TNF-a and IL-1b were determined by ELISA. High-dosebaicalin (200 mg/kg) reduced TNF-a and IL-1b levels significantlycompared with the vehicle-treated rats; middle-dose baicalin (100 mg/kg)and MTX (1 mg/kg) also reduced levels of TNF-a and IL-1b. Data areexpressed as mean 6 S.D. For in vitro studies, HFLS-RA were cultured insynoviocyte growth medium, HFLS-RA passages four to seven were usedfor all experiments. **P , 0.01 compared with vehicle-treated rats. (B)HFLS-RA were incubated and treated with baicalin (10, 20, 30 mM), for 24hours in six-well plates. Culture supernatants were harvested from eachwell, and the levels of TNF-a and IL-1b were determined by ELISA. Dataare expressed as mean 6 S.D. *P , 0.05; **P , 0.01 compared withuntreated control group.
Consistent with our hypothesis, the results showed thatbaicalin exerts a therapeutic role in CIA via suppressing theinflammation reaction, including alleviating redness andswelling of the ankle and decreasing the secretion of keycytokines in pathologic synovium. Radiologic and histologicanalyses also revealed that baicalin significantly alleviatedjoint damage and blocked the progression of inflammatoryarthritis. Other possible mechanisms mediated by baicalinin RA, such as its antiapoptotic role in the brain (Cao et al.,
2011) and its antioxidant effect in RA (Lee and Kim, 2010),will be addressed in future studies.The conventional inflammation reaction is mediated by
many cytokines, such as TNF-a, IL-1b, IL-6, TGF-b, IL-8, IL-l0,and so on. In the current study, we found that baicalin loweredcirculating TNF-a and IL-1b levels in plasma in the CIA ratmodel. Further investigations involving the pathologic RA cellmodel and human fibroblast-like synoviocytes from rheuma-toid arthritis showed that baicalin also decreased the secretion
Fig. 5. Effects of baicalin on NF-kB signaling pathways both in vivo and in vitro. (A) For in vivo studies, on day 46 after primary immunization the ratswere sacrificed and total proteins extracted from the synovium were separated by SDS-PAGE as described in Materials and Methods. Proteins weresubjected to Western blot analysis for NF-kB p65 and phospho-NF-kB p65 using specific antibodies. (B and C) Quantitation of Western blots of NF-kBp65 and phospho-NF-kB p65 in synovium of rats from different groups. (D) HFLS-RA were incubated and treated with baicalin (10, 20, 30 mM) for 24hours in six-well plates. Protein was subjected to Western blot analysis for NF-kB p65 and phospho-NF-kB p65. (E and F) Quantitation of Western blotsof NF-kB p65 and phospho-NF-kB p65 in HFLS-RA from different groups. Data are expressed as mean 6 S.D.; b-actin was used as the loading control.*P , 0.05 compared with the control group;
Fig. 6. Effects of baicalin on NF-kB signaling pathways both in vivo and in vitro. (A) For in vivo studies, on day 46 after primary immunization the ratswere sacrificed and total proteins extracted from the synovium were separated by SDS-PAGE as described in Materials and Methods. Protein sampleswere subjected toWestern blot analysis for acetyl-NF-kB p65 and sirt1 using specific antibodies. (B and C) Quantitation of Western blots of acetyl-NF-kBp65, and sirt1 in the synovium of rats from different groups. (D) HFLS-RA were incubated and treated with baicalin (10, 20, 30 mM) for 24 hours in six-well plates. Protein samples were subjected to Western blot analysis for acetyl-NF-kB p65 and sirt1. (E and F) Quantitation of Western blots of acetyl-NF-kB p65, and sirt1 in HFLS-RA from different groups. Data are expressed as mean6 S.D; b-actin was used as the loading control. *P, 0.05 comparedwith the control group.
Therapeutic Effects of Baicalin on CIA in Rats 441
of TNF-a and IL-1b in cellmedium in vitro. All the data showedthat baicalin suppressed the inflammation reaction by re-ducing the secretion of the key inflammatory cytokines inHFLS-RA and the CIA rat model.It is well established that TNF-a and IL-1b, the main
inflammation cytokines, exert their proinflammatory effects byinteracting with their own receptors in cell membranes andregulating the NF-kB pathway. The NF-kB pathway is the keymediator of the conventional inflammation reaction. The anti-inflammatory activity of baicalin has been associated with NF-kB, as shown in various acute and chronic inflammationmodels;it attenuates inflammation by inhibiting NF-kB activationin Staphylococcus aureus–induced mastitis and a cigarettesmoke–induced inflammatory model (Lixuan et al., 2010; Guoet al., 2013). Animal models of experimental arthritis supportthe notion that NF-kB activation plays a pathogenic role in theinflammation reaction. For example, increased synovial NF-kBbinding precedes the development of clinical joint involvementin CIA and gradually increases during the evolution of disease(Han et al., 1998; Tak and Firestein, 2001). The important roleof NF-kB in inflammation has also been shown in rats withstreptococcal cell wall–induced arthritis and adjuvant-inducedarthritis (Tsao et al., 1997; Miagkov et al., 1998). Consistentwith these published findings, we found that baicalin sup-pressed the protein expression of NF-kB p65, not only insynovium in CIA rats but also in HFLS-RA. Moreover, thephosphorylation levels of NF-kB p65 were reduced by baicalintreatment in a dose-dependent manner. All the data indicatethat the NF-kB pathway participates in baicalin’s inhibitionof the progression of CIA.Since the protein expression and phosphorylation of NF-kB
were lowered by baicalin treatment in both HFLS-RA and thesynovium in the CIA model, we sought to determine whetherits transcriptional activity is affected by baicalin. It is welldocumented that transcriptional activation of NF-kB corre-lates closely with NF-kB p65 acetylation (Lys310) (Chen et al.,2002; Lanzillotta et al., 2010; Kim et al., 2012). In the currentstudy, we found that baicalin dose-dependently reduced theacetylation levels of NF-kB p65 in synovium in CIA rats. Theinhibitive effect of baicalin on p65 acetylation was observed inHFLS-RA in vitro as well. Likewise, NF-kB transcriptionalactivity can be inhibited by the NAD1-dependent class IIIhistone deacetylase sirtuins. Indeed, it has been reported thatsirtuin 1, the best characterized and most well studied of thesirtuins, is a potent inhibitor of NF-kB transcription throughthe deacetylation of p65. Sirt1 represses NF-kB gene ex-pression by deacetylating RelA/p65 at lysine 310 (Yeung et al.,2004). In the current investigation, baicalin upregulated sirt1expression levels in both the synovium of CIA rats and HFLS-RA. All the data suggest that baicalin lowers the NF-kB p65acetylation level, and the possible mechanism involves in-duction of deacetylase sirt1 expression.In general, the data in our paper strongly demonstrate that
baicalin, an important component isolated from the dry root ofS. baicalensis Georgi, has therapeutic potential in CIA viainhibiting the inflammation reaction; the underlying mecha-nism is involved in the suppression of NF-kB p65 proteinexpression and the elevation of its deacetylation by sirt1.
Authorship Contributions
Participated in research design: H.-Z. Wang, H.-H. Wang,Z.-G. Wang, Liu.
Performed data analysis: H.-Z. Wang, Huang, Zhao, Z.-G. Wang.Wrote or contributed to the writing of the manuscript: H.-Z. Wang,
Z.-G. Wang.
References
Alonzi T, Fattori E, Lazzaro D, Costa P, Probert L, Kollias G, De Benedetti F, Poli V,and Ciliberto G (1998) Interleukin 6 is required for the development of collagen-induced arthritis. J Exp Med 187:461–468.
Bartok B and Firestein GS (2010) Fibroblast-like synoviocytes: key effector cells inrheumatoid arthritis. Immunol Rev 233:233–255.
Brand DD, Kang AH, and Rosloniec EF (2003) Immunopathogenesis of collagen ar-thritis. Springer Semin Immunopathol 25:3–18.
Cai X, Zhou H, Wong YF, Xie Y, Liu ZQ, Jiang ZH, Bian ZX, Xu HX, and Liu L (2007)Suppression of the onset and progression of collagen-induced arthritis in rats byQFGJS, a preparation from an anti-arthritic Chinese herbal formula. J Ethno-pharmacol 110:39–48.
Cao Y, Mao X, Sun C, Zheng P, Gao J, Wang X, Min D, Sun H, Xie N, and Cai J (2011)Baicalin attenuates global cerebral ischemia/reperfusion injury in gerbils via anti-oxidative and anti-apoptotic pathways. Brain Res Bull 85:396–402.
Chen LF, Mu Y, and Greene WC (2002) Acetylation of RelA at discrete sites regulatesdistinct nuclear functions of NF-kappaB. EMBO J 21:6539–6548.
Du F, Lü LJ, Fu Q, Dai M, Teng JL, Fan W, Chen SL, Ye P, Shen N, and Huang XF,et al. (2008) T-614, a novel immunomodulator, attenuates joint inflammation andarticular damage in collagen-induced arthritis. Arthritis Res Ther 10:1–11.
Feldmann M, Brennan FM, and Maini RN (1996) Rheumatoid arthritis. Cell 85:307–310.
Genovese MC, Bathon JM, Martin RW, Fleischmann RM, Tesser JR, Schiff MH,Keystone EC, Wasko MC, Moreland LW, and Weaver AL, et al. (2002) Etanerceptversus methotrexate in patients with early rheumatoid arthritis: two-year radio-graphic and clinical outcomes. Arthritis Rheum 46:1443–1450.
Guo M, Zhang N, Li D, Liang D, Liu Z, Li F, Fu Y, Cao Y, Deng X, and Yang Z (2013)Baicalin plays an anti-inflammatory role through reducing nuclear factor-kB andp38 phosphorylation in S. aureus-induced mastitis. Int Immunopharmacol 16:125–130.
Han Z, Boyle DL, Manning AM, and Firestein GS (1998) AP-1 and NF-kappaB reg-ulation in rheumatoid arthritis and murine collagen-induced arthritis. Autoim-munity 28:197–208.
Hou J, Wang J, Zhang P, Li D, Zhang C, Zhao H, Fu J, Wang B, and Liu J (2012)Baicalin attenuates proinflammatory cytokine production in oxygen-glucose de-prived challenged rat microglial cells by inhibiting TLR4 signaling pathway. IntImmunopharmacol 14:749–757.
Ivashkiv LB (1996) Cytokine expression and cell activation in inflammatory arthritis.Adv Immunol 63:337–376.
Karin M and Ben-Neriah Y (2000) Phosphorylation meets ubiquitination: the controlof NF-[kappa]B activity. Annu Rev Immunol 18:621–663.
Kim YJ, Kim HC, Ko H, Amor EC, Lee JW, and Yang HO (2012) Inhibitory effects ofaurentiacin from Syzygium samarangense on lipopolysaccharide-induced in-flammatory response in mouse macrophages. Food Chem Toxicol 50:1027–1035.
Lanzillotta A, Sarnico I, Ingrassia R, Boroni F, Branca C, Benarese M, Faraco G,Blasi F, Chiarugi A, and Spano P, et al. (2010) The acetylation of RelA in Lys310dictates the NF-kB-dependent response in post-ischemic injury. Cell Death Dis 1:e96.
Lee JH and Kim GH (2010) Evaluation of antioxidant and inhibitory activities fordifferent subclasses flavonoids on enzymes for rheumatoid arthritis. J Food Sci 75:H212–H217.
Lee SJ, Nam WD, Na HJ, Cho YL, Ha KS, Hwang JY, Lee H, Kim SO, Kwon YG,and Kim YM (2008) CT20126, a novel immunosuppressant, prevents collagen-induced arthritis through the down-regulation of inflammatory gene expression byinhibiting NF-kappaB activation. Biochem Pharmacol 76:79–90.
Li Y, Wang S, Wang Y, Zhou C, Chen G, Shen W, Li C, Lin W, Lin S, and Huang H,et al. (2013) Inhibitory effect of the antimalarial agent artesunate on collagen-induced arthritis in rats through nuclear factor kappa B and mitogen-activatedprotein kinase signaling pathway. Transl Res 161:89–98.
Liu LL, Gong LK, Wang H, Xiao Y, Wu XF, Zhang YH, Xue X, Qi XM, and Ren J(2008) Baicalin inhibits macrophage activation by lipopolysaccharide and protectsmice from endotoxin shock. Biochem Pharmacol 75:914–922.
Lixuan Z, Jingcheng D, Wenqin Y, Jianhua H, Baojun L, and Xiaotao F (2010) Bai-calin attenuates inflammation by inhibiting NF-kappaB activation in cigarettesmoke induced inflammatory models. Pulm Pharmacol Ther 23:411–419.
Luo X, Zuo X, Mo X, Zhou Y, and Xiao X (2011) Treatment with recombinant Hsp72suppresses collagen-induced arthritis in mice. Inflammation 34:432–439.
Miagkov AV, Kovalenko DV, Brown CE, Didsbury JR, Cogswell JP, Stimpson SA,Baldwin AS, and Makarov SS (1998) NF-kappaB activation provides the potentiallink between inflammation and hyperplasia in the arthritic joint. Proc Natl AcadSci USA 95:13859–13864.
Odeh M (1998) Role of cytokines in rheumatoid arthritis. Drug News Perspect 11:331–341.
Shou J, Bull CM, Li L, Qian HR, Wei T, Luo S, Perkins D, Solenberg PJ, Tan SL,and Chen XY, et al. (2006) Identification of blood biomarkers of rheumatoid ar-thritis by transcript profiling of peripheral blood mononuclear cells from the ratcollagen-induced arthritis model. Arthritis Res Ther 8:1–14.
Tak PP and Firestein GS (2001) NF-kappaB: a key role in inflammatory diseases.J Clin Invest 107:7–11.
Tsao PW, Suzuki T, Totsuka R, Murata T, Takagi T, Ohmachi Y, Fujimura H,and Takata I (1997) The effect of dexamethasone on the expression of activatedNF-kappa B in adjuvant arthritis. Clin Immunol Immunopathol 83:173–178.
Wang Z, Yao T, and Song Z (2010) Involvement and mechanism of DGAT2 upregulationin the pathogenesis of alcoholic fatty liver disease. J Lipid Res 51:3158–3165.
Xu XF, Cai BL, Guan SM, Li Y, Wu JZ, Wang Y, and Liu B (2011) Baicalin induceshuman mucoepidermoid carcinoma Mc3 cells apoptosis in vitro and in vivo. InvestNew Drugs 29:637–645.
Yang M, Xiao C, Wu Q, Niu M, Yao Q, Li K, Chen Y, Shi C, Chen D, and Feng G, et al.(2010) Anti-inflammatory effect of Sanshuibaihu decoction may be associated withnuclear factor-kappa B and p38 MAPK alpha in collagen-induced arthritis in rat.J Ethnopharmacol 127:264–273.
Yang X, Yang J, and Zou H (2013) Baicalin inhibits IL-17-mediated joint in-flammation in murine adjuvant-induced arthritis. Clin Dev Immunol 2013:268065.
Yeung F, Hoberg JE, Ramsey CS, Keller MD, Jones DR, Frye RA, and Mayo MW(2004) Modulation of NF-kappaB-dependent transcription and cell survival by theSIRT1 deacetylase. EMBO J 23:2369–2380.
Yin F, Liu J, Ji X, Wang Y, Zidichouski J, and Zhang J (2011) Baicalin prevents theproduction of hydrogen peroxide and oxidative stress induced by Ab aggregation inSH-SY5Y cells. Neurosci Lett 492:76–79.
Zink A, Listing J, Kary S, Ramlau P, Stoyanova-Scholz M, Babinsky K, von HinueberU, Gromnica-Ihle E, Wassenberg S, and Antoni C, et al. (2005) Treatment con-tinuation in patients receiving biological agents or conventional DMARD therapy.Ann Rheum Dis 64:1274–1279.
Address correspondence to: Dr. Yanhong Liu, Department of LaboratoryDiagnosis, The Second Affiliated Hospital of Harbin Medical University, 246Xuefu Road, Nangang District, Harbin, 150001, P. R. of China. E-mail:[email protected]
Therapeutic Effects of Baicalin on CIA in Rats 443











![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/234/3/local/back...effect, 708 Blockade, reticuloendothelial, enzyme-al-bumin conjugates, chronic adininis-tration](https://static.documents.pub/doc/80x56/60757ab7f966210d5e51d2f2/index-jpet-jpet-effect-708-blockade-reticuloendothelial-enzyme-al-bumin.jpg)



![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/230/3/local/back-matter.pdf · histrionicotoxin effects (frogs), 619 ... distribution kinetics ana- ... and myocardium](https://static.documents.pub/doc/80x56/5b7ac0067f8b9ae1328d73ab/index-jpet-jpet-histrionicotoxin-effects-frogs-619-distribution-kinetics.jpg)









