Isolation of Insertion Sequence ISRLdTAL1145-1 from a
Rhizobium sp. (Leucaena diversifolia) and Distributionof Homologous Sequences Identifying Cross-
Inoculation Group RelationshipstDOUGLAS J. RICE,* PADMA SOMASEGARAN, KATHRYN MAcGLASHAN,
AND B. BEN BOHLOOLUniversity of Hawaii, NiJTAL Center, Paia, Maui, Hawaii 96779
Received 2 May 1994/Accepted 20 September 1994
Insertion sequence (IS) element ISRLdTAL1145-1 from Rhizobium sp. (Leucaena diversifolia) strain TAL1145 was entrapped in the sacB gene of the positive selection vector pUCD800 by insertional inactivation. Ahybridization probe prepared from the whole 2.5-kb element was used to determine the distribution ofhomologous sequences in a diverse collection of 135 Rhizobium and Bradyrhizobium strains. The IS probehybridized strongly to Southern blots of genomic DNAs from 10 rhizobial strains that nodulate both Phaseolusvulgaris (beans) and Leucaena leucocephala (leguminous trees), 1 Rhizobium sp. that nodulates Leucaena spp.,9 R. meliloti (alfalfa) strains, 4 Rhizobium spp. that nodulate Sophora chrysophylla (leguminous trees), and 1nonnodulating bacterium associated with the nodules of Pithecellobium dulce from the Leucaena cross-
inoculation group, producing distinguishing IS patterns for each strain. Hybridization analysis revealed thatISRLdTAL1145-1 was strongly homologous with and closely related to a previously isolated element, ISRmUSDA1024-1 from R. meliloti, while restriction enzyme analysis found structural similarities and differencesbetween the two IS homologs. Two internal segments of these IS elements were used to construct hybridizationprobes of 1.2 kb and 380 bp that delineate a structural similarity and a difference, respectively, of the two IShomologs. The internal segment probes were used to analyze the structures of homologous IS elements in otherstrains. Five types of structural variation in homolog IS elements were found. The predominate IS structuraltype naturally occurring in a strain can reasonably identify the strain's cross-inoculation group relationships.Three IS structural types were found in Rhizobium species that nodulate beans and Leucaena species, one ofwhich included the designated type IIB strain of R. tropici (CIAT 899). Weak homology to the whole IS probe,but not with the internal segments, was found with two Bradyrhizobiumjaponicum strains. The taxonomic andecological implications of the distribution of ISRLdTAL1145-1 are discussed.
Rhizobia, which are members of the family Rhizobiaceae, area genetically diverse and physiologically heterogeneous groupof soil bacteria which form nodules on the roots of leguminousplants. The bacteroidal form of the rhizobia in the nodules fixN2 in symbiosis with the host legume. Traditionally, rhizobiaare classified on the basis of their cultural characteristics andby the host legume-dependent cross-inoculation (CI) groupconcept (8), rather than their genetic relatedness. The inade-quacy of the CI group approach for classifying rhizobia hasbeen analyzed and criticized elsewhere (46), and variousattempts have been made to develop a taxonomic scheme forclassifying rhizobia (1, 21).
Recent advances in chemosystematic analyses have broad-ened the database on which taxonomic classifications andidentifications of microorganisms is based (28) and haveattracted the attention of rhizobiologists. For example, nif andnod gene DNA hybridization probes were instrumental in thedistinction of type I and type II rhizobia-nodulating beans(Phaseolus vulgaris) (25) and the recognition of two highly
tContribution from the NitTAL Center and MIRCEN, Departmentof Agronomy and Soil Science, University of Hawaii Journal Series no.
3996. This paper is dedicated to the memory of B. Ben Bohlool, whoseforesight brought rhizobial molecular genetics to the NifTAL Project.
divergent symbiotic types of Bradyrhizobium japonicum (41).Indeed, the members of the International Subcommittee forthe Taxonomy of Rhizobium and Agrobacterium state the needfor additional DNA probes derived from single-copy or reit-erated chromosomal DNA sequences from other strains toclarify taxonomic relationships (12).
Bacterial insertion sequences (IS) are mobile genetic ele-ments (DNA sequences of 800 to 2,500 bp), and their occur-
rence, variety and distribution have been reviewed by Galasand Chandler (9). The best known IS of rhizobia is ISRmlfrom Rhizobium meliloti (29). This IS was found to be presentin 80% of R. meliloti strains from North America (44, 45) andalso in 97% of a population of R. meliloti in a European fieldsoil (15). The potential of using IS elements as hybridizationprobes of Southern-blotted rhizobial genomic DNAs to pro-duce IS patterns for positive strain identification (45) and todetermine the genotype of a strain (35) has been demon-strated, but only for R. meliloti (35, 45).
Until recently, most of the IS reported for rhizobia were
accidental discoveries (5, 27, 29). Positive-selection, broad-host-range bacterial vectors such as pUCD800, pSUP104-rpsL,and pSUP104-pheS which facilitate the entrapment of ISelements are now available (10, 35). pUCD800 contains theconditionally lethal sacB gene from the gram-positive bacte-rium Bacillus subtilis (10). The sacB gene encodes levansucrasewhich catalyzes transfructorylation from sucrose to variousacceptors. The expression of levansucrase is induced by su-
4394
Vol. 60, No. 12
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
DISTRIBUTION OF INSERTION SEQUENCE ISRLdTALI 145-1 4395
TABLE 1. Rhizobial strains which cross-hybridized with ISRLdTAL1145-la
Strain (host legume) Other name(s) Origin Source or reference
B. japonicumTAL 102 (G. max) USDA 110 Florida USDANone (G. max) USDA 110; TAL 102 Florida NifTAL
R. melilotiTAL 380 (M. sativa) SU 47 New South Wales, Australia SUTAL 381 (Medicago sp.) SU 794 Australia SUTAL 920 (Medicago sp.) NZP 4008; B 18 New Zealand UHTAL 1165 (M. sativa) Allen 131 England UWTAL 1167 (M. indica) Allen 139 Hawaii UWTAL 1372 (M. sativa) POA 116 Brazil UFRGSTAL 1871 (M. sativa) Nam 1 Utah and Wyoming USUNone (Medicago sp.) NGR 185 35None (Medicago sp.) USDA 1024 35None (Medicago sp.) MV II Erlanger, Germany 35
Rhizobium sp.TAL 1436 (Acacia famesiana) ORS 911 Senegal, West Africa ORSTOMTAL 1 (C. calothyrsus) Original Nicaragua NifTALTAL 2 (C. calothyrsus) Original Nicaragua NifFALTAL 1145 (L. diversifolia) CB 3060; CIAT 1967; NA 4004-1 Australia CIATTAL 21 (L. leucocephala) Allen 711; Nit 94A1 Wisconsin NitraginTAL 82 (L. leucocephala) Original Hawaii NifTALTAL 583 (L. leucocephala) CB 948; NGR 8 New Guinea CSIROTAL 1861 (L. retusa) UPRM 8048 Puerto Rico UPRTAL 1910 (L. salvadorensis) Original Honduras NifTALTAL 78 (Phaseolus wrightii) Nit 127L3 Arizona NitraginTAL 9 (P. dulce) Original Nicaragua NiffALTAL 1445 (P. dulce) Original Hawaii NifFALTAL 600 (Prosopis chilensis) Original Hawaii NifTALTAL 603 (Prosopis chilensis) Original Hawaii NifTALTAL 1945 (S. chrysophylla) Original Hawaii NifFALTAL 1946 (S. chrysophylla) Original Hawaii NifTALTAL 1949 (S. chrysophylla) Original Hawaii NifTALTAL 1950 (S. chrysophylla) Original Hawaii NifTALTAL 1952 (S. chrysophylla) Original Hawaii NitTALNone (Sophora secundiflora inoculated NZP 5443; 146A2 United States NZP (19)
with rhizobia from Astragalus spp.)
R. tropiciTAL 1797 CIAT 899; ATCC 49672 CIAT
a USDA, Rhizobium culture collection, Beltsville Agricultural Research Center, Beltsville, Md.; NifTAL, NitTAL Project, University of Hawaii, Paia, Maui; SU,University of Sydney, Sydney, Australia; Nit, Nitragin Company, Wis.; CIAT, Centro Internacional Agriculturale Tropicale, Cali, Colombia; UH, University of Hawaii,Honolulu; UW, University of Wisconsin, Madison; UFRGS, Facultade Agronomia, Porto Alegre, Brazil; USU, Utah State University, Logan; ORSTOM, ORSTOM,Dakar, Senegal; CSIRO, Commonwealth Scientific and Industrial Research Organization, Brisbane, Australia; UPR, University of Puerto Rico, Mayaguez, PuertoRico; NZP, Department of Scientific and Industrial Research. Palmerston North, New Zealand.
crose and is lethal in the presence of 5% sucrose in the platingmedia of several gram-negative bacteria including rhizobia (R.meliloti) (10). Transposition of an IS into the sacB gene regioninactivates this gene and allows the positive selection of thebacterium and the entrapped IS.
Nitrogen-fixing tree legumes, like the grain and foragespecies of legumes, are also nodulated by Rhizobium spp. andBradyrhizobium spp. The rhizobia that nodulate tree legumesrepresent a new, unique, and diverse group of rhizobia whoseecology, taxonomy, and molecular biology have been thesubject of very limited research. The Rhizobium sp. (Leucaenadiversifolia) strain TAL 1145 (14) has been found to nodulateeight species in the genus Leucaena and a wide range of treelegumes which include Calliandra callothyrsus, Desmanthusvirgatus, Gliricidia sepium, Pithocellobium dulce, and Prosopisspp. (37a). In the present study, we utilized the pUCD800system to entrap an IS element (ISRLdTAL1145-1) from TAL1145 and studied its distribution in 135 strains of Rhizobiumand Bradyrhizobium spp. from various CI groups.
MATERIALS AND METHODS
Bacterial strains and plasmids. The rhizobial strains whichcross-hybridized with ISRLdTAL1145-1 are listed in Table 1.ISRLdTAL1145-1 did not hybridize with the following strains:from Glycine max, B. japonicum TAL 379 and TAL 1812; fromAcacia auriculaeformis, Bradyrhizobium sp. strain TAL 1446;from Acacia decurrens, Bradyrhizobium strain TAL 1881; fromAcacia mangium, Bradyrhizobium strains TAL 1867 and TAL1868; from Acacia meamsii, Bradyrhizobium strain TAL 1388;from Albizia falcataria, Bradyrhizobium strain TAL 45; fromAlbizia lebbek, Bradyrhizobium strain TAL 1536; from Arachishypogaea, Bradyrhizobium strains TAL 1000 and TAL 1371;from Cajanus cajan, Bradyrhizobium strains TAL 1127 andTAL 1132; from Crotolaria paulina, Bradyrhizobium strain TAL1380; from Enterolobium cyclocarpum, Bradyrhizobium strainTAL 1530; from Erythrina indica, Bradyrhizobium strain TAL749; from Macrotyloma afticanum, Bradyrhizobium strain TAL309; from Phaseolus acutifolius, Bradyrhizobium strain TAL
VOL. 60, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
APPL. ENvIRON. MICROBIOL.
644; from Vigna unguiculata, Bradyrhizobium strain TAL 169;from Phaseolus vulgaris, Rhizobium etli TAL 182 and TAL1865; from Trifolium sp., Rhizobium leguminosarum biovartrifolii TAL 1826, TAL 1827, and TAL 1828; from Lathyrushirsutus, R. leguminosarum biovar viceae TAL 634; from Pisumsativum, R. leguminosarum biovar viceae TAL 1236; from Cicerarietium, R. loti TAL 480, TAL 620, and TAL 1148; from Lotuscomiculatus, R. loti TAL 198 and TAL 234; from Medicagominima, R. meliloti TAL 142; from Medicago sativa, R. melilotiTAL 921, TAL 1149, TAL 1150, TAL 1153, TAL 1154, TAL1373, TAL 1408, TAL 1483, and TAL 1876; from Medicagoarabica, R. meliloti TAL 1175; from Medicago polymorpha, R.meliloti TAL 1479; from Medicago lupulina, R. meliloti TAL1498; from Acacia pennatula, Rhizobium strains TAL 41, TAL152, TAL 1595, TAL 1596, and TAL 1774; fromAcacia senegal,Rhizobium strains TAL 1428 and TAL 1429; from Calliandracalothyrsus, Rhizobium strain TAL 1801; from Calliandra sp.,Rhizobium strains TAL 1452 and TAL 1453; from Gliricidiamaculata, Rhizobium strain TAL 1788; from G. sepium, Rhizo-bium strain TAL 1768; from Lablab purpureus, Rhizobiumstrain NGR 234; from Leucaena leucocephala, Rhizobiumstrains TAL 582, TAL 590, TAL 762, TAL 996, TAL 1005,TAL 1006, TAL 1381, and TAL 1887; from Leucaena macro-phylla nelsonii, Rhizobium strains TAL 1943 and TAL 1951;from Leucaena retusa, Rhizobium strains TAL 1862, TAL 1863,and TAL 1864; from Leucaena salvadorensis, Rhizobium strainTAL 1909; from Leucaena sp., Rhizobium strain B 215; fromLeucaena shannoni, Rhizobium strain TAL 29; from Mimosainvisa, Rhizobium strains TAL 74, TAL 571, TAL 575, TAL576, TAL 586, and TAL 846; from Mimosa pigra, Rhizobiumstrains TAL 1001 and TAL 1784; from Mimosa pudica, Rhizo-bium strain TAL 1884; from P. vulgaris, Rhizobium strain TAL1383; from P. dulce, Rhizobium strains TAL 65 and TAL 1532;from Pithecellobium saman, Rhizobium strain TAL 1807; fromProsopis africana, Rhizobium strain TAL 1526; from Prosopisjuliflora, Rhizobium strains TAL 1523, TAL 1524, and TAL1525; from Prosopis pallida, Rhizobium strain TAL 70; fromSesbania longifolia, Rhizobium strain TAL 1042; from Sesbaniamacrocarpa, Rhizobium strain TAL 1123; from Sesbania ros-trata, Rhizobium strain TAL 674; from Sesbania sp., Rhizobiumstrains TAL 1113 and TAL 1114; from Sophora chrysophylla,Rhizobium strains TAL 1442, TAL 1947, and TAL 1948; fromSophora microphylla, Rhizobium strain NZP 5275; from So-phora tetraptera, Rhizobium strain NZP 5057; and from G. max,Sinorhizobium fredii TAL 1783 and TAL 1837.We used Escherichia coli HB101 transformed with pRK2013
(Kmr) (4, 7) and HB101 transformed with pUCD800 (Kmr)(10). pUHR9 is pLAFR1 with a 27-kb insert which includes thenodABCD host specificity genes and the NifAIJ symbioticregion of TAL 1145 (11). ISRmUSDA1024-1 was entrapped inthe rpsL gene of pSUP104-rpsL carried in E. coli S17-1 (35).Rhizobial CI group relationships were performed as describedpreviously (38).
Culture, mating, and selection conditions. E. coli strainswere grown on Luria-Bertani medium or Terrific broth sup-plemented with the appropriate antibiotics for plasmid main-tenance (30). Rhizobia were grown on yeast extract-mannitol(YM) medium (0.5 g liter-1 yeast extract) (42a), Y medium(YM without mannitol supplement), or tryptone-yeast (TY)medium (2).
Supplements to the growth media were used at the followingconcentrations: agar (Difco, Detroit, Mich.), 1.5% (wt/vol);carbenicillin, 10 ,ug ml-'; kanamycin, 10 ,ug ml-'; and sucrose,5% (wt/vol) (from a 50% solution of 0.22-,um-pore-size-filter-sterilized stock).TAL 1145 was determined to be intrinsically Cb' and Kms.
pUCD800 was transferred to TAL 1145 by conjugation pro-moted by triparental matings with pRK2013 (4). Patch matingwas performed for 1, 3, and 6 days at 31°C (34, 36). Conjuga-tion was disrupted by vigorous shaking in 0.85% NaCl-0.01%(wt/vol) Tween 80, and TAL 1145 containing pUCD800 wasselected on Y plates supplemented with carbenicillin and kana-mycin. Recipients were tested for the presence of a functionalsacB gene by replica plating individual colonies onto selectivemedia and selective media supplemented with sucrose. Indi-vidual colonies able to grow on selection media but unable togrow on selection media supplemented with sucrose wereselected for further use. Colonies selected for the isolation ofIS elements were confirmed to be strain TAL 1145 by thedirect fluorescence antibody technique as previously described(39).
Isolation of IS elements from TAL 1145. TAL 1145 cellscontaining pUCD800 with a functional sacB gene were grownto saturation in TY with kanamycin and carbenicillin and wereplated on selection media supplemented with sucrose. Colo-nies that as a result were able to grow in the presence ofsucrose were screened for altered mobility of pUCD800 by amodification of the rapid disruption of bacterial colony proto-col for the small-scale preparation of plasmid DNA (30). Asaturated TY culture (1 to 1.5 ml) in a Microfuge tube waspelleted by centrifugation in an Eppendorf Microfuge (model5415) for 1 min at 16,000 x g (subsequent references tocentrifugation will also be for 16,000 x g). The pellet (25 to 50,u) was resuspended in 200 ,u of 10 mM EDTA, pH 8.0, andan equal volume of alkaline lysis solution (0.2 N NaOH-0.5%sodium dodecyl sulfate-20% sucrose) was added with gentlemixing. The cells were heated for 5 min at 70°C in a water bath,and 2 volumes of phenol (unbuffered redistilled phenol-0.1%8-hydroxyquinoline and {chloroform-isoamyl alcohol [24:1]},1:1) was added and gently mixed until an emulsion formed.The emulsion was centrifuged for 5 min, and 250 RI of theaqueous phase was transferred to a new tube and extractedwith chloroform-isoamyl alcohol as per the phenol extraction.The supernatant (200 RI) was transferred to a new tube, a 0.5volume of 7.5 M ammonium acetate was added, and themixture was allowed to stand for 5 to 10 min at 25°C (roomtemperature [RT]). The sample was centrifuged for 10 min atRT, and the supernatant was transferred to a new tube. A totalof 2.5 volumes of 100% ethanol was added, and the mixturewas allowed to stand for 10 min at RT and was centrifuged for30 min. The pellet was washed with 1.0 ml of 76% ethanol-10mM ammonium acetate before being dried in a vacuum cen-trifuge (model SF-50; GeneVac, Ipswich, England). The pelletwas dissolved in 20 jil of TE (10 mM Tris-1 mM EDTA [pH8.0]), and half of the sample was analyzed for altered mobilityby 0.7% agarose gel electrophoresis in 40 mM Tris-acetate-1mM EDTA. Five microliters of the plasmid preparations fromselected colonies was used to transform HB101-competentcells (subcloning efficiency) (GIBCO-BRL, Gaithersburg, Md.)according to the supplier's instructions.DNA methods. Bacterial DNA was simply and rapidly
isolated from a wide range of rhizobia by modifying andcombining the acetone wash protocol described by Heath et al.(16) with the guanidine isothiocyanate lysis method describedby Saunders (31). A compact bacterial pellet was produced (asdescribed above for the miniplasmid preparation) and resus-pended in 0.75 ml of 50 mM Tris (pH 7.2) and centrifuged. Thesupernatant was discarded, and the pellet was resuspended inthe small amount of remaining supernatant before 0.75 ml ofice-cold acetone was added to the cell suspension and imme-diately mixed to keep the cells from clumping. The suspensionwas incubated on ice for 5 min and centrifuged as described
4396 RICE ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
DISTRIBUTION OF INSERTION SEQUENCE ISRLdTAL1145-1
A B C D
427 . -i:
__595\ -t~~~~~~~~~~~~~~~~~~~09
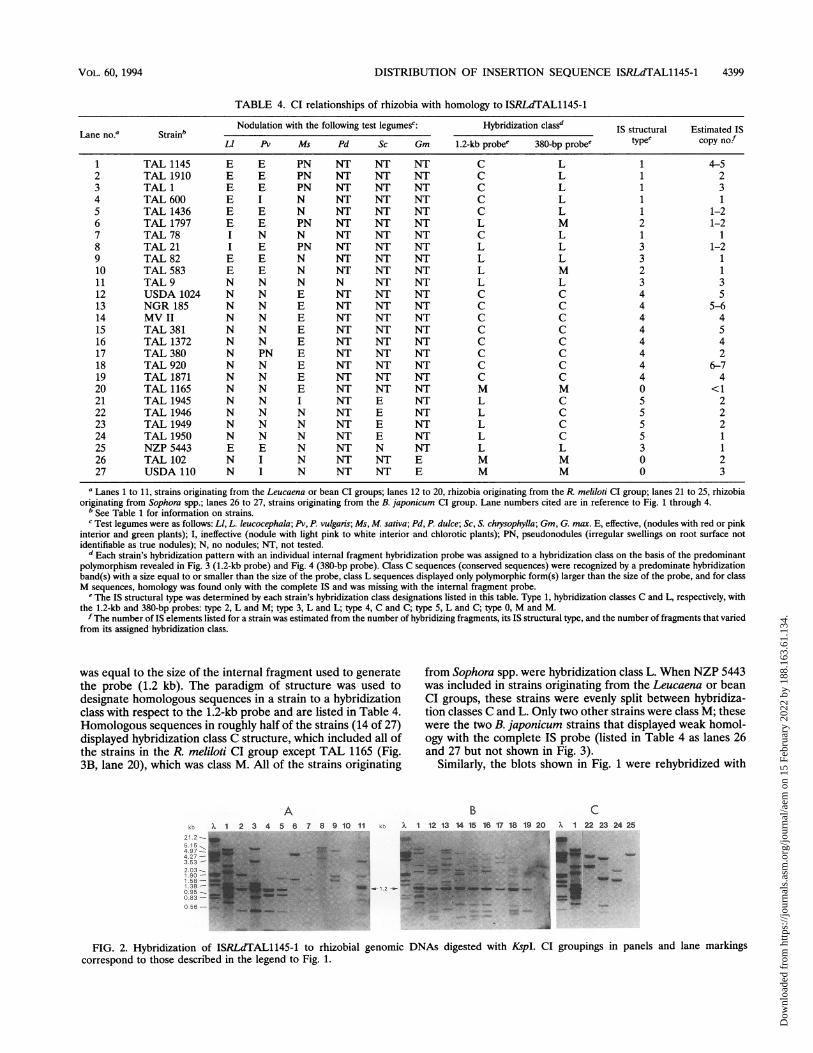
FIG. 1. Hybridization of ISRLdTAL1145-1 to rhizobial genomic DNAs digested with HindIll. To provide a comparative standard to judgehybridization intensity, TAL 1145 was included in lanes 1 of panels A, B, and C. A longer exposure was needed to distinctly visualize the TAL 102variants seen in panel D, and lane 25 shown in panel C is also shown in panel D to provide an exposure comparison. Strains originating from theLeucaena or bean CI groups (A), the R. meliloti CI group (B), the Sophora CI group (C), and the B. japonicum CI group (D) are shown. Gel lanenumbers correspond to rhizobial strains listed in Table 4. A lanes, molecular weight marker III labeled with Digoxigenin (Boehringer Mannheim).
above. The supernatant was aspirated, and the pellet was
allowed to air dry. The pellet was resuspended in 40 ,ul of TEand 60 RI of a freshly prepared solution lysozyme (16.7 mgml-' in 25% [wtlvol] sucrose-50 mM Tris-1 mM EDTA [pH8.0]) was added with mixing. The cells were incubated for 10min at RT before lysis by the addition of 200 [lI of 5 Mguanidine isothiocyanate-0.1 M EDTA, pH 7.0. The lysate wasimmediately mixed by inversion, and 150 ,ul of 7.5 M ammo-
nium acetate was added and mixed. Chloroform-isoamyl alco-hol (500 ,ul) was added, and an emulsion was formed byvortexing. The phases were separated by centrifugation for 4min. The aqueous phase (350 ,ul) was transferred to a freshmicrocentrifuge tube, and the DNA was precipitated by theaddition of 190 RI of isopropyl alcohol and incubation at RTfor 10 min. The DNA was collected by centrifugation for 10min, and the pellet was washed twice and dried as describedabove for the miniplasmid preparation. The nucleic acid pelletwas dissolved in 100 ,ul of TE and quantified by spectropho-tometry. Yields of 30 to 40 jig of total genomic DNA contam-inated with a variable amount of RNA were routinely ob-tained.
Restriction enzymes were obtained from Boehringer Mann-heim (Indianapolis, Ind.), GIBCO-BRL, and New EnglandBiolabs (Beverly, Mass.) and were used as directed by thesuppliers. A 2-,ug amount of total genomic DNA was digestedwith 12 U of activity in a 50-pI volume.
Restriction enzyme digests of genomic DNA were sizefractionated by electrophoresis in 0.7% agarose-0.5 x TBE (45mM Tris-borate-1 mM EDTA [pH 8]) at 5 V/cm. Gels were
vacuum blotted to GeneScreen Plus nylon membranes (NENResearch Products, Boston, Mass.) in a vacuum transferapparatus (Vacugene model 2016; Pharmacia LKB Biotech-nology, Piscataway, N.J.) according to the manufacturer'srecommendations.
Large-scale plasmid DNA was purified by following alkalinelysis and polyethylene glycol precipitation protocols (30).
Nonradioactive DNA labeling, hybridization, chemilumines-cence detection, and probe stripping were done with theGenius kit (Boehringer Mannheim) according to publishedmethods (17, 24). Hybridization probes were prepared fromtemplate DNA by random-primed labeling with Digoxigenin-dUTP (6). Portions (0.1 to 0.3 jug) of undigested plasmid DNAwere alkali denatured according to the method described byMierendorf and Pfeffer before labeling (26). Hybridizationprobes prepared from restriction enzyme digestion fragmentswere separated from vector DNA fragments by two rounds ofgel electrophoresis, and DNA fragment recovery was withglassmilk and reagents supplied by a Geneclean II kit (Bio 101,Inc., La Jolla, Calif.) according to the supplier's instructions.
The distribution of ISRLdTAL1145-1 was determined byhybridization of the labeled IS to either Southern blots or dotblots prepared in a Bio-Dot apparatus (Bio-Rad, Hercules,Calif.) according to the manufacturer's directions. Southernblots were prepared for all strains with positive dot blothybridizations. All strains screened for IS homology by South-ern blotting were stripped of the IS probe and rehybridizedwith a probe prepared from pUHR9 to spot check for geneticdiversity and the presence of a symbiotic gene region.
RESULTS
Isolation and characterization of an IS element from TAL1145. pUCD800 was transferred to TAL 1145 by conjugationat frequencies of 1 x i0-5, 3 x 10-5, and 1 X 10-3 transcon-jugants per final recipient for 1, 3, and 6 days of conjugation,respectively. The frequency of inactivation of the sacB gene inTAL 1145 transconjugants varied from 2 x 10-5 to 8 x 10-5.About 55% of the miniplasmid preparations from TAL 1145
colonies with a SacB- phenotype showed an electrophoreticmobility for pUCD800 slower than that for the wild-typecontrol. The rest of the SacB- colonies contained plasmidswith mobilities indistinguishable from that of pUCD800.The presence of IS elements within these plasmids was
investigated by Southern blot analysis. A control hybridizationdemonstrated that pUCD800 sequences were not homologousto that of TAL 1145 genomic DNA (data not shown). Hybrid-ization probes prepared from two of the larger SacB- pUCD800 plasmids produced IS patterns for TAL 1145 genomicDNA like those shown in Fig. 1, lane 1.The insertion size and location of the hybridizing elements
within pUCD800 were determined by restriction enzyme map-ping. One putative IS of about 2.5 kb, which was designatedISRLdTAL1145-1, inserted into a 41-bp region in the sacBgene bordered by the EcoRI and StuI sites. The other homol-ogous copy of ISRLdTAL1145-1 was of equivalent size and waslocated in an 818-bp StuI-KpnI fragment of the sacB gene.Neither isolated IS was digestible with BamHI, EcoRI, KpnI,PstI, or Stul, while both copies contained one HindIII and twoSacII and Sall sites. The IS located in the StuI-Kpnl fragmentwas either an identical or a closely related homologous copy ofISRLdTAL1145-1 and was not investigated further. ISRLdTAL1145-1, which was excised from pUCD800 by digestionwith EcoRI and StuI as a single 2.5-kb fragment bordered with41 bp of the sacB gene, was used to generate a hybridizationprobe representing the complete IS element to investigate itsdistribution in rhizobial strains.
Distribution of ISRLdTAL1145-1 in rhizobial strains. ISRLdTAL1145-1 was hybridized to genomic DNAs from 135
VOL. 60, 1994 4397
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
APPL. ENVIRON. MICROBIOL.
TABLE 2. Frequency of ISRLdTAL1145-1 hybridization withRhizobium and Bradyrhizobium species that define the
CI groups in which homology was found
Rhizobial specis*Fraction withCI group (host legume) homology(%L)
caena spp.Alfalfa R. meliloti 10/23 (43)Unassigned Rhizobium spp. (Sophora spp.) 6/11 (55)Soybeans B. japonicum 2/4 (50)
a Phaseolus strains are referred to as Rhizobium spp. (Phaseolus spp.) if theirspecies is uncertain (48).
strains of Rhizobium and Bradyrhizobium representing diversespecies both within and outside the Leucaena CI group (listedin Materials and Methods and Table 1). Southern blot analysiswith stringent wash conditions found homology with ISRLdTAL1145-1 in 24% of the strains tested (see Table 1 for alisting of 33 positive strains). The CI groups in which strainswere found to cross-hybridize with ISRLdTAL1145-1 and thefraction of strains that hybridized from each rhizobial speciesor host plant that defines the CI group are listed in Table 2. Nohybridization was found with strains originally isolated fromtwo of the host legumes in the Leucaena CI group or withstrains classified as R. etli in the bean CI group (Table 2).Strong hybridization was found in 27% (31 of 114) of strainsfrom the genus Rhizobium, and only weak cross-hybridizationwas found in strains from the genus Bradyrhizobium, represent-ing 9.5% (2 of 21) of this population.
Examination of the 33 IS profiles found 5 IS patterns thatwere identical with the patterns from one or two other strains(Table 3). Strains with matched IS profiles were also found tohave identical profiles when they were hybridized with pUHR9(not shown). Figure 1 shows 27 distinct IS profiles arrangedaccording to the host CI group from which the strains wereoriginally isolated.CI relationships of rhizobia with homology to ISRLdTAL
1145-1. The CI relationships of rhizobial strains in whichhomology was detected were investigated for nodulation spec-ificity and the effectiveness of nitrogen fixation with three orfour host legumes that define relevant CI groups (Table 4).TAL 9 did not nodulate any of the trial legume hosts and didnot nodulate even its host of origin. NZP 5443 did not nodulateits host of origin (Table 1) but effectively nodulated both
TABLE 3. Rhizobial strains with indistinguishable IS profilesRepresentative straina Identical IS profile(s)
TAL 1..... TAL2TAL 600..... TAL 603TAL 1145..... TAL 1445 and TAL 1861TAL 1165..... TAL 1167TAL 1950..... TAL 1952
a Only the representative strains are shown in the figures and in Table 4.
Leucaena species and beans, a trait common to most of thestrains with ISRLdTAL1145-1 homology originally isolatedfrom either Leucaena species or beans (see first 11 strains inTable 4).Homolog of ISRLdTAL1145-1. Since strong homology with
ISRLdTAL1145-1 was frequently found among strains fromthe R. meliloti CI group (Fig. 1), several IS elements that hadbeen entrapped from R. meliloti strains by Simon et al. (35)were tested for similarity with ISRLdTAL1145-1 by com-paring their IS hybridization patterns with the IS patternsproduced by ISRLdTAL1145-1 (35). Hybridization of ISRmUSDA1024-1 to Southern blots containing most of the strainsshown in Fig. 1 produced IS patterns indistinguishable fromthose produced by hybridization with ISRLdTAL1145-1 (notshown), indicating that the two IS elements were closelyrelated homologs.
Restriction enzyme analysis of the two homologs foundstructural differences as well as similarities between them.ISRmUSDA1024-1 is about 200 bp longer than ISRLdTAL1145-1 and contains one extra HindIII site and one extra Sallsite (35). Digestion of ISRmUSDA1024-1 with HindIll re-leased an internal segment of about 380 bp (not shown), thesame size as that of the smallest fragment shown for theHindlIl digest of strain USDA 1024 (Fig. 1B, lane 12). NeitherISRLdTAL1145-1 (not shown) nor any of the homologouscopies of ISRLdTAL1145-1 found in TAL 1145 released a380-bp internal segment when digested with Hindlll (Fig. 1,lane 1).Both ISRLdTAL1145-1 and ISRmUSDA1024-1 completely
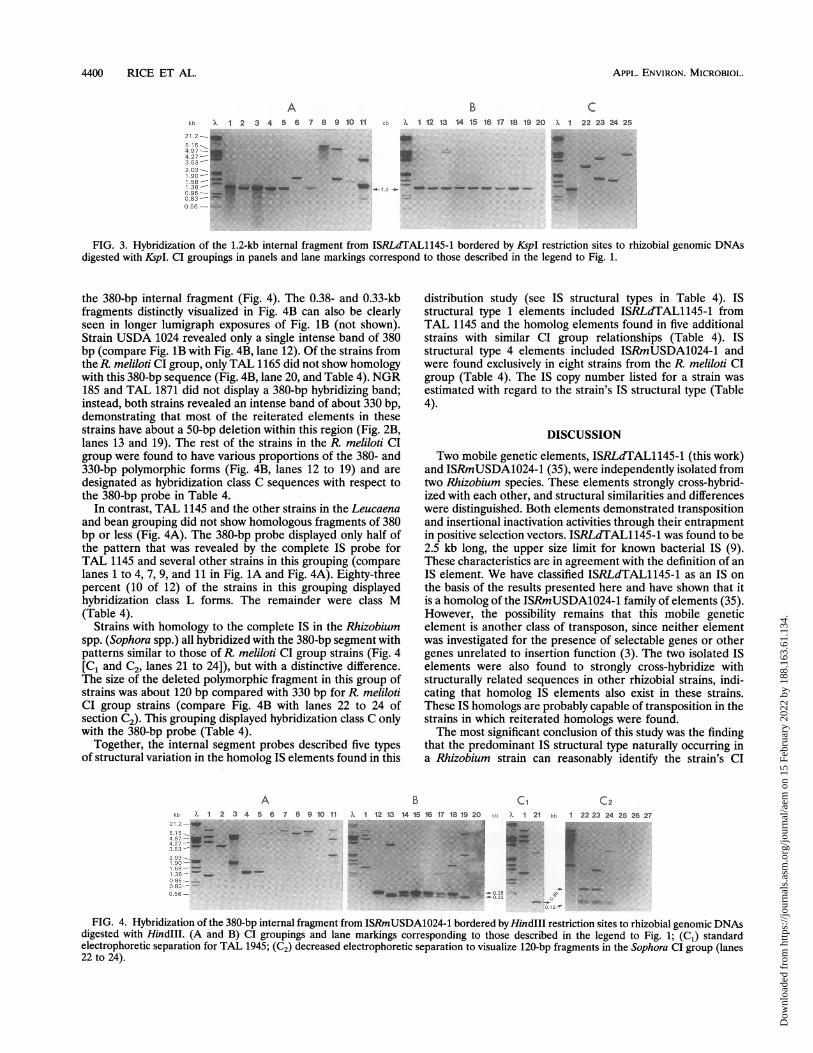
released a 1.2-kb internal segment when digested with SacII orits isoschizomer, KspI (data not shown). This common 1.2-kbDNA fragment was revealed by Southern blot analysis whengenomic DNAs from strains TAL 1145 and USDA 1024 weredigested with KspI and hybridized with ISRLdTAL1145-1 (Fig.2B, lanes 1 and 12, respectively). K:spl was chosen for thedigestion of rhizobial genomic DNA because hybridizationanalysis demonstrated that KspI digested genomic DNA fromTAL 1145 and some rhizobial strains more completely thanSacII, even with enzyme activity-to-DNA ratios of as high as 50U/,ug (not shown).
Conservation of DNA sequences homologous to ISRLdTAL1145-1 and ISRmUSDA1024-1. The 1.2-kb and the 380-bp inter-nal fragments of ISRLdTAL1145-1 and ISRmUSDA1024-1,respectively, delineate a structural similarity and a structuraldifference between the two IS homologs. The structures ofhomologous sequences in other strains were investigated withhybridization probes derived from these internal IS segmentsusing the following paradigm. If all of the IS copies in a straincontain the same internal segment that was used to prepare thehybridization probe, then hybridization analysis of its genomicDNA digested with the same restriction enzyme specificityused for probe production will display only the same sizefragment as the internal segment, irrespective of the number ofIS copies in the strain. IS copies with deletions within theinternal segment will be displayed as smaller fragments. Largerpolymorphic fragments could be the result of homologoussequences not having one or both of the restriction sites usedto release the internal segment or could result from insertionswithin this region. No cross-hybridization with the internalsegment probe indicates that this region is missing in thesequences homologous to the complete IS in this strain.The blots prepared with rhizobial DNA digested with KspI
were washed free of the previous hybridization probe andrehybridized with the 1.2-kb internal segment (compare Fig. 2with Fig. 3). Both TAL 1145 and USDA 1024 (Fig. 3B, lanes 1and 12, respectively) revealed only one intense band whose size
4398 RICE ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
DISTRIBUTION OF INSERTION SEQUENCE ISRLdTAL1145-1 4399
TABLE 4. CI relationships of rhizobia with homology to ISRLdTAL1145-1
Nodulation with the following test legumesc: Hybridization classd IS structural Estimated ISLane no.' Strain' yee oynLl Pv Ms Pd Sc Gm 1.2-kb probee 380-bp probee type copy no.
1 TAL1145 E E PN NT NT NT C L 1 4-52 TAL1910 E E PN NT NT NT C L 1 23 TAL1 E E PN NT NT NT C L 1 34 TAL600 E I N NT NT NT C L 1 15 TAL1436 E E N NT NT NT C L 1 1-26 TAL1797 E E PN NT NT NT L M 2 1-27 TAL78 I N N NT NT NT C L 1 18 TAL21 I E PN NT NT NT L L 3 1-29 TAL82 E E N NT NT NT L L 3 110 TAL583 E E N NT NT NT L M 2 111 TAL9 N N N N NT NT L L 3 312 USDA1024 N N E NT NT NT C C 4 513 NGR185 N N E NT NT NT C C 4 5-614 MVII N N E NT NT NT C C 4 415 TAL381 N N E NT NT NT C C 4 516 TAL1372 N N E NT NT NT C C 4 417 TAL380 N PN E NT NT NT C C 4 218 TAL920 N N E NT NT NT C C 4 6-719 TAL1871 N N E NT NT NT C C 4 420 TAL1165 N N E NT NT NT M M 0 <121 TAL1945 N N I NT E NT L C 5 222 TAL1946 N N N NT E NT L C 5 223 TAL1949 N N N NT E NT L C 5 224 TAL1950 N N N NT E NT L C 5 125 NZP5443 E E N NT N NT L L 3 126 TAL102 N I N NT NT E M M 0 227 USDA11O N I N NT NT E M M 0 3
a Lanes 1 to 11, strains originating from the Leucaena or bean CI groups; lanes 12 to 20, rhizobia originating from the R. meliloti CI group; lanes 21 to 25, rhizobiaoriginating from Sophora spp.; lanes 26 to 27, strains originating from the B. japonicum CI group. Lane numbers cited are in reference to Fig. 1 through 4.
b See Table 1 for information on strains.c Test legumes were as follows: Ll, L. leucocephala; Pv, P. vulgaris; Ms, M. sativa; Pd, P. dulce; Sc, S. chrysophylla; Gm, G. max. E, effective, (nodules with red or pink
interior and green plants); I, ineffective (nodule with light pink to white interior and chlorotic plants); PN, pseudonodules (irregular swellings on root surface notidentifiable as true nodules); N, no nodules; NT, not tested.
d Each strain's hybridization pattern with an individual internal fragment hybridization probe was assigned to a hybridization class on the basis of the predominantpolymorphism revealed in Fig. 3 (1.2-kb probe) and Fig. 4 (380-bp probe). Class C sequences (conserved sequences) were recognized by a predominate hybridizationband(s) with a size equal to or smaller than the size of the probe, class L sequences displayed only polymorphic form(s) larger than the size of the probe, and for classM sequences, homology was found only with the complete IS and was missing with the internal fragment probe.
e The IS structural type was determined by each strain's hybridization class designations listed in this table. Type 1, hybridization classes C and L, respectively, withthe 1.2-kb and 380-bp probes: type 2, L and M; type 3, L and L; type 4, C and C; type 5, L and C; type 0, M and M.fThe number of IS elements listed for a strain was estimated from the number of hybridizing fragments, its IS structural type, and the number of fragments that varied
from its assigned hybridization class.
was equal to the size of the internal fragment used to generatethe probe (1.2 kb). The paradigm of structure was used todesignate homologous sequences in a strain to a hybridizationclass with respect to the 1.2-kb probe and are listed in Table 4.Homologous sequences in roughly half of the strains (14 of 27)displayed hybridization class C structure, which included all ofthe strains in the R. meliloti CI group except TAL 1165 (Fig.3B, lane 20), which was class M. All of the strains originating
A
from Sophora spp. were hybridization class L. When NZP 5443was included in strains originating from the Leucaena or beanCI groups, these strains were evenly split between hybridiza-tion classes C and L. Only two other strains were class M; thesewere the two B. japonicum strains that displayed weak homol-ogy with the complete IS probe (listed in Table 4 as lanes 26and 27 but not shown in Fig. 3).
Similarly, the blots shown in Fig. 1 were rehybridized with
B Ckb
21.2-5.15 N4.97 -4.27 -3.53 -2.03 -
1.90-1.58-1.38-0.95 -0.83 -0.56 -
FIG. 2. Hybridization of ISRLdTAL1145-1 to rhizobial genomic DNAs digested with KspI. CI groupings in panels and lane markingscorrespond to those described in the legend to Fig. 1.
VOL. 60, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
APPL. ENVIRON. MICROBIOL.
A B CX 1 2 3 4 5 6 7 8 9 10 11 kb
FIG. 3. Hybridization of the 1.2-kb internal fragment from ISRLdTAL1145-1 bordered by KspI restriction sites to rhizobial genomic DNAsdigested with KspI. CI groupings in panels and lane markings correspond to those described in the legend to Fig. 1.
the 380-bp internal fragment (Fig. 4). The 0.38- and 0.33-kbfragments distinctly visualized in Fig. 4B can also be clearlyseen in longer lumigraph exposures of Fig. 1B (not shown).Strain USDA 1024 revealed only a single intense band of 380bp (compare Fig. 1B with Fig. 4B, lane 12). Of the strains fromthe R. meliloti CI group, only TAL 1165 did not show homologywith this 380-bp sequence (Fig. 4B, lane 20, and Table 4). NGR185 and TAL 1871 did not display a 380-bp hybridizing band;instead, both strains revealed an intense band of about 330 bp,demonstrating that most of the reiterated elements in thesestrains have about a 50-bp deletion within this region (Fig. 2B,lanes 13 and 19). The rest of the strains in the R. meliloti CIgroup were found to have various proportions of the 380- and330-bp polymorphic forms (Fig. 4B, lanes 12 to 19) and aredesignated as hybridization class C sequences with respect tothe 380-bp probe in Table 4.
In contrast, TAL 1145 and the other strains in the Leucaenaand bean grouping did not show homologous fragments of 380bp or less (Fig. 4A). The 380-bp probe displayed only half ofthe pattern that was revealed by the complete IS probe forTAL 1145 and several other strains in this grouping (comparelanes 1 to 4, 7, 9, and 11 in Fig. 1A and Fig. 4A). Eighty-threepercent (10 of 12) of the strains in this grouping displayedhybridization class L forms. The remainder were class M(Table 4).
Strains with homology to the complete IS in the Rhizobiumspp. (Sophora spp.) all hybridized with the 380-bp segment withpatterns similar to those of R. meliloti CI group strains (Fig. 4[C1 and C2, lanes 21 to 24]), but with a distinctive difference.The size of the deleted polymorphic fragment in this group ofstrains was about 120 bp compared with 330 bp for R. melilotiCI group strains (compare Fig. 4B with lanes 22 to 24 ofsection C2). This grouping displayed hybridization class C onlywith the 380-bp probe (Table 4).
Together, the internal segment probes described five typesof structural variation in the homolog IS elements found in this
lA
B
k 1%44 1 .% A. a41 04%fl . ..". .
distribution study (see IS structural types in Table 4). ISstructural type 1 elements included ISRLdTAL1145-1 fromTAL 1145 and the homolog elements found in five additionalstrains with similar CI group relationships (Table 4). ISstructural type 4 elements included ISRmUSDA1024-1 andwere found exclusively in eight strains from the R. meliloti CIgroup (Table 4). The IS copy number listed for a strain wasestimated with regard to the strain's IS structural type (Table4).
DISCUSSION
Two mobile genetic elements, ISRLdTAL1 145-1 (this work)and ISRmUSDA1024-1 (35), were independently isolated fromtwo Rhizobium species. These elements strongly cross-hybrid-ized with each other, and structural similarities and differenceswere distinguished. Both elements demonstrated transpositionand insertional inactivation activities through their entrapmentin positive selection vectors. ISRLdTAL1 145-1 was found to be2.5 kb long, the upper size limit for known bacterial IS (9).These characteristics are in agreement with the definition of anIS element. We have classified ISRLdTAL1145-1 as an IS onthe basis of the results presented here and have shown that itis a homolog of the ISRmUSDA1024-1 family of elements (35).However, the possibility remains that this mobile geneticelement is another class of transposon, since neither elementwas investigated for the presence of selectable genes or othergenes unrelated to insertion function (3). The two isolated ISelements were also found to strongly cross-hybridize withstructurally related sequences in other rhizobial strains, indi-cating that homolog IS elements also exist in these strains.These IS homologs are probably capable of transposition in thestrains in which reiterated homologs were found.The most significant conclusion of this study was the finding
that the predominant IS structural type naturally occurring ina Rhizobium strain can reasonably identify the strain's CI
C. C2kb A 1 2 3 4 0 17 8 Y 1V 11 A l1;1 14 1:5 1(l 18 19ZU kb I 1 i2 kb 1 ZZZJ Z425 26 Z7
5:.15 -| *
4.27 _3.536-
1.90 _ ,_|, 1__ns1.58
FIG. 4. Hybridization of the 380-bp internal fragment from ISRmUSDA1024-1 bordered by HindlIl restriction sites to rhizobial genomic DNAsdigested with HindIll. (A and B) CI groupings and lane markings corresponding to those described in the legend to Fig. 1; (C1) standardelectrophoretic separation for TAL 1945; (C2) decreased electrophoretic separation to visualize 120-bp fragments in the Sophora CI group (lanes22 to 24).
4400 RICE ET AL.
A BD
ownl
oade
d fr
om h
ttps:
//jou
rnal
s.as
m.o
rg/jo
urna
l/aem
on
15 F
ebru
ary
2022
by
188.
163.
61.1
34.
DISTRIBUTION OF INSERTION SEQUENCE ISRLdTAL1145-1 4401
group relationships. Five IS structural types were described onthe basis of the characteristic polymorphism(s) displayed withone or both internal segment hybridization probes (Table 4).With one exception (TAL 9), the individual IS structural typeswere found to be partitioned within groups of rhizobia havingsimilar CI group relationships. Rhizobium spp. that nodulateand fix nitrogen on both beans and Leucaena species werefound to contain IS structural types 1, 2, and 3 (Table 4). Twostrains with type 1 homologs (TAL 78 and TAL 600) did notdisplay all of these symbiotic properties but could at leastnodulate one or both hosts (Table 4). Structural type 4 and 5homologs were found exclusively in the R. meliloti species andthe tree legume-nodulating Rhizobium spp. (Sophora chryso-phylla), respectively (Table 4). Strain NZP 5443 (Table 4) wasfound to contain a type 3 homolog, and its CI relationshipswere found to match two other Rhizobium spp. (TAL 21 andTAL 82) with type 3 structures rather than its isolation historyas outlined in Table 1 (19). A structural type 3 homolog wasfound in TAL 9, a nonnodulating gram-negative rod-shapedbacterium associated with the nodules of Pithecellobium dulcefrom the Leucaena CI group (Table 4). Southern blot analysiswith symbiotic genes from TAL 1145 (pUHR9) detected nohomology with TAL 9 genomic DNA, indicating that this strainwas not a Rhizobium or Bradyrhizobium strain (not shown). Wedo not know the relationship of this bacterium to the Rhizo-bium strains that share this IS structural type but speculate thatTAL 9 could be a rhizosphere-associated bacterium bound tothe outside of the nodule from which it was isolated or that itcould have been a Rhizobium strain from the bean andLeucaena CI group that lost its symbiotic gene region aspostulated for other bacteria related to Rhizobium species (20,33, 37).Type 0 sequences were not CI group specific. Type 0
sequences cross-hybridized to the complete ISRLdTAL1145-1element; however, no structural relatedness to either internalsegment probe was found. The less intense band shown for R.meliloti TAL 1165 (Fig. 1, lane 20) may represent a portion ofa type 4-like sequence, and its presence as a single fragmentsuggests that it is nontransposable. The very weakly hybridizingtype 0 sequences shown for B. japonicum TAL 102 and itsvariant (USDA 110) may represent distantly related elementswhich are capable of transposition, since these strains displayvariable numbers of related fragments (Fig. 1D, lanes 26 and27).Although not all of the phylogenetic relationships of strains
with strong homology to ISRLdTAL1145-1 are understood, theknown relationships suggest that the group of structurallyrelated IS homologs are distributed in the genus Rhizobium inclosely related rhizobial species. Recently, the phylogeneticrelationships of strain TAL 1145 to several of the rhizobialspecies in which ISRLdTAL1145-1 homologs were found weredetermined by 16S rRNA gene sequencing (11, 42, 47). Thesephylogenetic trees show that TAL 1145 is distinct from alldesignated species of Rhizobium but closely related to R.leguminosarum, R meliloti, and R tropici. To resolve TAL1145's taxonomic position, it has been suggested to identifyTAL 1145 to the species level as a third type of R. tropici (42).The three IS structural types found in the Rhizobium spp. thatnodulate both beans and Leucaena species may aid in thedesignation of subtypes in the R. tropici species, since a type 2structure was found in TAL 1797 (CIAT 899) (Table 4), whichis the designated type IIB strain of the R. tropici species (25),and TAL 1145 contains a structural type 1 homolog (Table 4).Further investigation is needed to determine if the IS struc-tural types correlate with R tropici subtypes.The distribution patterns of individual IS structural types
more closely match the previously reported distribution pat-terns of rhizobial IS elements than does the sum of thedistribution patterns found for all ISRLdTAL1145-1 homologtypes. The general picture that has emerged from the fewdistribution studies of rhizobial IS elements is that a variablenumber of strongly cross-hybridizing sequences are found inmany to most strains within the species of IS isolation (27, 35,43, 44), with infrequent strains in related species showing a fewsignificant but less intense bands (5, 35, 43, 44), or that nocross-hybridization is found outside a closely related group ofstrains (27). The narrower distribution of strongly homologouselements reported for other rhizobial IS elements may resultfrom most of these studies not focusing on finding relatedelements outside the species of IS isolation (5, 27, 35), as eventhe most extensive distribution studies tested a relatively lownumber of strains outside the species of isolation comparedwith the present study (43, 44). The distribution study ofISRml most closely parallels our results by finding cross-hybridization in a more distantly related diazotrophic bacte-rium and in two unclassified rhizobial strains that nodulateboth beans and alfalfa (44). More distant relationships of ISelements ISRml, ISRm3, and ISRm4 to diverse, distantlyrelated bacteria are found through the similarity of amino acidsequences derived through comparisons of nucleotide se-quence data (23, 40, 43). Alternatively, the novel distributionpattern of all of the ISRLdTAL1145-1 homolog types inrhizobia may be due to their belonging to a particular class ofmobile genetic elements which will best be resolved throughnucleotide sequencing studies.Our distribution study of ISRLdTAL1145-1 in rhizobia
implies a pattern of horizontal gene exchange. The distributionof individual IS structural types within groups of rhizobia withrelated CI group relationships suggests that an IS type isexchanged relatively frequently within the CI group and that itsstructure is conserved in the group through homologousrecombination. The partial structural relatedness of the differ-ent IS homologs suggests that horizontal transfer occurredbetween CI groups but was less frequent than exchange withinCI groups. The IS structural type 5 elements hint that thishomolog is a mosaic of the class L patterns seen with the 1.2-kbprobe of structural type 3 elements and the class C patternsseen with the 380-bp probe of IS structural type 4 elements(Table 4). These types of horizontal transfer and recombina-tion events of IS elements follow the example reported in aDNA sequence variation study of three IS elements in entericbacteria (22).The use of bacterial IS elements in strain identification and
to spot underlying genetic relationships of rhizobial strains iswell established (13, 18, 32, 35, 43, 45). We found several setsof strains with identical (Table 3) and related IS patterns suchas TAL 600 and TAL 1436 (Fig. 1 and 2, lanes 4 and 5) andTAL 381 and TAL 1372 (Fig. 1 and 2, lanes 15 and 16). Mostof the sets of strains with identical IS patterns reported herehave close accession numbers and were independent isolatesfrom the same soil and host plant. The two strains with ISpatterns identical to that of TAL 1145 (Table 3) are likelyreisolated strains of this highly competitive and widely distrib-uted inoculant strain. Neither the fluorescence antibody tech-nique nor hybridization analysis with the pUHR9 probe (notshown) could differentiate these three strains. They wereisolated from different geographical locations (Table 1) anddifferent host legumes in the same CI group; however, TAL1445 was isolated from soil at NifTAL after TAL 1145 hadbeen introduced to NifTAL soil, and TAL 1861 was isolatedafter TAL 1145 had been sent to the University of Puerto Rico.The genealogy of most of the strains with identical profiles
VOL. 60, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
APPL. ENVIRON. MICROBIOL.
supports the contention that they are the same or are closelyrelated strains. Variations in patterns are due to differences inthe restriction enzyme fragments of the bacterial genome,differences in the genomic locations of the IS determined bytransposition, and differences in the restriction enzyme siteswithin individual IS elements.
This investigation extends the usefulness of IS elements asDNA probes for positive strain identification in culture collec-tions, inoculant quality control, and rhizobial ecological stud-ies. ISRLdTAL1145-1 can be used both to identify a strain andto identify CI relationships of rhizobia that harbor one of thefive IS structural types. This study is an abundant resource forfurther studies of population genetics and dynamics, systemat-ics, and the horizontal transmission of genes in bacteria.
ACKNOWLEDGMENTS
This research was supported by the U.S. Agency for InternationalDevelopment Cooperative agreement DAN-4177-A-00-6035-00, DAN-4177-A-00-1077-00 and subgrant SM-CRSP-023 under grant no. DAN-1311-G-00-1049-00.We thank Dulal Borthakur, C. I. Kado, and Reinhard Simon for the
gifts of pUHR9, pUCD800, and ISRmUSDA1024-1, respectively. Wealso thank Wendy Asano, Megan Loomis Powers, and Bruce Martinfor technical assistance.
REFERENCES1. Allen, 0. N., and E. K. Allen. 1957. Family II. Rhizobiaceae Conn,
1938, p. 285-296. In R. S. Breed, E. D. G. Murray, and N. R.Smith (ed.), Bergey's manual of determinative bacteriology. Wil-liams and Wilkins Co., Baltimore.
2. Beringer, J. 1974. R factor transfer in Rhizobium leguminosarum. J.Gen. Microbiol. 84:188-198.
3. Campbell, A., D. Berg, D. Botstein, E. Lederberg, R. Novick, P.Starlinger, and W. Szybalski. 1977. Nomenclature of transposableelements in prokaryotes, p. 15-22. In A. I. Bukhari, J. A. Shapiro,and S. L. Adhya (ed.), DNA insertion elements, plasmids, andepisomes. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
4. Ditta, G., S. Stanfield, D. Corbin, and D. R. Helinski. 1980. Broadhost range DNA cloning system for gram-negative bacteria: con-struction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad.Sci. USA 77:7347-7351.
5. Dusha, I., S. Kovalenko, Z. Banfalvi, and A. Kondorosi. 1987.Rhizobium meliloti insertion element ISRm2 and its use foridentification of the fixX gene. J. Bacteriol. 169:1403-1409.
6. Feinberg, A. P., and B. Vogelstein. 1983. A technique for radiola-beling DNA restriction endonuclease fragments to high specificactivity. Anal. Biochem. 132:6-13.
7. Figurski, D. H., and D. R. Helinski. 1979. Replication of anorigin-containing derivative of plasmid RK2 dependent on aplasmid function provided in trans. Proc. Natl. Acad. Sci. USA76:1648-1652.
8. Fred, E. B., I. L. Baldwin, and E. McCoy. 1932. Root nodulebacteria and leguminous plants. University of Wisconsin Press,Madison.
9. Galas, D. J., and M. Chandler. 1989. Bacterial insertion se-quences, p. 109-162. In D. E. Berg and M. M. Howe (ed.), MobileDNA. American Society for Microbiology, Washington, D.C.
10. Gay, P., D. Le Coq, M. Steinmetz, T. Berkelman, and C. I. Kado.1985. Positive selection procedure for entrapment of insertionsequence elements in gram-negative bacteria. J. Bacteriol. 164:918-921.
11. George, M. L. C., J. P. W. Young, and D. Borthakur. 1994. Geneticcharacterization of Rhizobium sp. strain TAL 1145 that nodulatestree legumes. Can. J. Microbiol. 40:208-215.
12. Graham, P. H., M. J. Sadowsky, H. H. Keyser, Y. M. Barnet, R. S.Bradley, J. E. Cooper, D. J. D. Ley, B. D. W. Jarvis, E. B. Roslycky,B. W. Strijdom, and J. P. W. Young. 1991. Proposed minimalstandards for the description of new genera and species of root-and stem-nodulating bacteria. Int. J. Syst. Bacteriol. 41:582-587.
13. Green, L., R. D. Miller, D. E. Dykhuizen, and D. L. Hartl. 1984.Distribution of DNA insertion element IS5 in natural isolates of
Eschenichia coli. Proc. Natl. Acad. Sci. USA 81:4500-4504.14. Halliday, J., and P. Somasegaran. 1983. Nodulation, nitrogen
fixation, and Rhizobium strain affinities in the genus Leucaena, p.27-32. In A. Chouinard (ed.), Leucaena research in the Asian-Pacific region. Unipub, New York.
15. Hartmann, A., and N. Amarger. 1991. Genotypic diversity of anindigenous Rhizobium meliloti field population assessed by plasmidprofiles, DNA fingerprinting, and insertion sequence typing. Can.J. Microbiol. 37:600-608.
16. Heath, L. S., G. L. Sloan, and H. E. Heath. 1986. A simple andgenerally applicable procedure for releasing DNA from bacterialcells. Appl. Environ. Microbiol. 51:1138-1140.
17. Holtke, H. J., G. Sagner, C. Kessler, and G. Schmitz. 1992.Sensitive chemiluminescent detection of digoxigenin-labeled nu-cleic acids: a fast and simple protocol and its applications.BioTechniques 12:104-113.
18. Hu, M., and R. C. Deonier. 1981. Comparison of IS1, IS2, and IS3copy number in Escherichia coli strains K-12, B and C. Gene16:161-170.
19. Jarvis, B. D. W. 1983. Genetic diversity of Rhizobium strains whichnodulate Leucaena leucocephala. Curr. Microbiol. 8:153-158.
20. Jarvis, B. D. W., L. J. H. Ward, and E. A. Slade. 1989. Expressionby soil bacteria of nodulation genes from Rhizobium leguminosa-rum. Appl. Environ. Microbiol. 55:1426-1434.
21. Jordan, D. C., and 0. N. Allen. 1974. Genus II, Rhizobium, p.262-264. In R. E. Buchanan and N. E. Gibbons (ed.), Bergey'smanual of determinative bacteriology. Williams and Wilkins Co.,Baltimore.
22. Lawrence, J. G., H. Ochman, and D. L. Hartl. 1992. The evolutionof insertion sequences within enteric bacteria. Genetics 131:9-20.
23. Mariani, F., E. Piccolella, V. Colizzi, R. Rappuoli, and R. Gross.1993. Characterization of an IS-like element from Mycobacteriumtuberculosis. J. Gen. Microbiol. 139:1767-1772.
24. Martin, R., C. Hoover, S. Grimme, C. Grogan, J. Holtke, and C.Kessler. 1990. A highly sensitive, nonradioactive DNA labelingand detection system. BioTechniques 9:762-768.
25. Martinez, E., M. A. Pardo, R. Palacios, and M. A. Cervallos. 1985.Reiteration of nitrogen fixation gene sequences and specificity ofRhizobium in nodulation and nitrogen fixation in Phaseolus vul-garis. J. Gen. Microbiol. 131:1779-1786.
26. Mierendorf, R. C., and D. Pfeffer. 1987. Direct sequencing ofdenatured plasmid DNA, p. 556-562. In S. L. Berger and A. R.Kimmel (ed.), Guide to molecular cloning techniques. AcademicPress, Inc., San Diego.
27. Priefer, U. B., J. Kalinowski, B. Ruger, W. Heumann, and A.Puhler. 1989. ISR1, a transposable DNA sequence resident inRhizobium Class IV strains, shows structural characteristics ofclassical insertion elements. Plasmid 21:120-128.
28. Priest, F., and B. Austin. 1993. Modern bacterial taxonomy.Chapman & Hall, London.
29. Ruvkun, G. B., S. R. Long, H. M. Meade, R. C. van den Bos, andF. M. Ausubel. 1982. ISRml: a Rhizobium meliloti insertion se-quence that transposes preferentially into nitrogen fixation genes.J. Mol. Appl. Genet. 1:405-418.
30. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y.
31. Saunders, N. A. 1989. Analysis of restriction fragment lengthpolymorphisms for epidemiological tracing of bacteria using non-radioactive probes. Focus 11:47-49.
32. Schoner, B., and R. Schoner. 1981. Distribution of IS5 in bacteria.Gene 16:347-352.
33. Segovia, L., D. Pifiero, R. Palacios, and E. Martinez-Romero.1991. Genetic structure of a soil population of nonsymbioticRhizobium leguminosarum. Appl. Environ. Microbiol. 57:426-433.
34. Simon, R. 1984. In vivo genetic engineering: use of transposableelements in plasmid manipulation and mutagenesis of bacteriaother than E. coli, p. 125-140. In A. Puhler and K. N. Timmis (ed.),Advanced molecular genetics. Springer-Verlag, New York.
35. Simon, R., B. Hotte, B. Klauke, and B. Kosier. 1991. Isolation andcharacterization of insertion sequence elements from gram-nega-tive bacteria by using new broad-host-range, positive selectionvectors. J. Bacteriol. 173:1502-1508.
4402 RICE ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 15
Feb
ruar
y 20
22 b
y 18
8.16
3.61
.134
.
DISTRIBUTION OF INSERTION SEQUENCE ISRLdTAL1145-1 4403
36. Simon, R., M. O'Connell, M. Labes, and A. Piihier. 1986. Plasmidvectors for the genetic analysis and manipulation of rhizobia andother gram-negative bacteria, p. 640-659. In A. Weissbach and H.Weissbach (ed.), Plant molecular biology. Academic Press, Inc.,Orlando, Fla.
37. Soberon-Chavez, G., and R. Najera. 1989. Isolation from soil ofRhizobium leguminosarum lacking symbiotic information. Can. J.Microbiol. 35:464-468.
37a.Somasegaran, P. Unpublished data.38. Somasegaran, P., and H. J. Hoben. 1985. Methods in legume-
Rhizobium technology. NifTAL Project, University of Hawaii,Paia, Maui.
39. Somasegaran, P., and R. B. Martin. 1986. Symbiotic characteris-tics and Rhizobium requirements of a Leucaena leucocephala xLeucaena diversifolia hybrid and its parental genotypes. Appl.Environ. Microbiol. 52:1422-1424.
40. Soto, M. J., A. Zorzano, J. Olivares, and N. Toro. 1992. Sequenceof ISRm4 from Rhizobium meliloti strain GR4. Gene 120:125-126.
41. Stanley, J., G. G. Brown, and D. P. S. Verma. 1985. Slow-growingRhizobium japonicum comprises two highly divergent symbiotictypes. J. Bacteriol. 163:148-154.
42. van Berkum, P., R. B. Navarro, and A. A. T. Vargas. 1994.Classification of the uptake hydrogenase-positive (Hup+) beanrhizobium as Rhizobium tropici. Appl. Environ. Microbiol. 60:554-561.
42a.Vincent, J. M. 1970. A manual for the practical study of the rootnodule bacteria. Blackwell Scientific Publications, Ltd., Oxford.
43. Wheatcroft, R., and S. Laberge. 1991. Identification and nucle-otide sequence of Rhizobium meliloti insertion sequence ISRm3:similarity between the putative transposase encoded by ISRm3 andthose encoded by Staphylococcus aureus IS256 and Thiobacillusferrooxidans IST2. J. Bacteriol. 173:2530-2538.
44. Wheatcroft, R., and R. J. Watson. 1988. Distribution of insertionsequence ISRml in Rhizobium meliloti and other gram-negativebacteria. J. Gen. Microbiol. 134:113-121.
45. Wheatcroft, R., and R. J. Watson. 1988. A positive strain identifi-cation method for Rhizobium meliloti. Appl. Environ. Microbiol.54:574-576.
46. Wilson, J. K. 1944. Over five hundred reasons for abandoning thecross-inoculation groups of legumes. Soil Sci. 58:61-69.
47. Young, J. P. W. 1993. Molecular phylogeny of rhizobia and theirrelatives, p. 587-592. In R. Palacios, J. Mora, and W. E. Newton(ed.), New horizons in nitrogen fixation. Kluwer Academic Pub-lishers, Dordrecht, The Netherlands.
48. Young, P., E. Martinez, Y. Barnet, J. Cooper, and K. Lindstrom.1993. Report from the taxonomy meeting, subcommittee onAgrobacterium and Rhizobium, p. 777-778. In J. Palacios, J. Mora,and W. E. Newton (ed.), New horizons in nitrogen fixation.Kluwer Academic Publishers, Dordrecht, The Netherlands.