ORIGINAL ARTICLE Interaction between Nm23 and the tumor suppressor VHL Chih-Hung Lin & Vincent Dammai & Boris Adryan & Tien Hsu Received: 28 April 2014 /Accepted: 2 June 2014 # Springer-Verlag Berlin Heidelberg 2014 Abstract Among the anti-tumor genes (tumor suppressors and metastasis suppressors), the von-Hippel Lindau gene and the Nm23 family of genes are among the more intriguing ones. Both are small (long and short forms of VHL are 30 and 19 kD, respectively, and Nm23 is ~17 kD), and both possess diverse molecular and cellular functions. Despite extensive studies, the entire spectra of functions and the molecular function-phenotype correlation of these two proteins have not been completely elucidated. In this report, we present data showing these two proteins interact physically. We also sum- marize and confirm the previous studies that demonstrated the endocytic function of these two genes and further show that the endocytic function of VHL is mediated through the activ- ity of Nm23. These functional and molecular interactions are evolutionarily conserved from Drosophila to human. Keywords von Hippel-Lindau . Nm23 . Endocytosis . Tumor suppressor gene Introduction The von Hippel-Lindau tumor suppressor gene (VHL) muta- tions are the genetic defects in the familial VHL disease (Latif et al. 1993), which manifests in a limited number of organ- specific tumors (Maher et al. 2011), predominantly in kidney [clear-cell renal cell carcinoma (ccRCC)] and central nervous system (hemangioblastoma). Up to 70 % of the carriers of germ-line VHL mutations eventually develop either ccRCC or hemangioblastoma or both (Lonser et al. 2003). In addition, loss of VHL function, including somatic mutations and epige- netic defects, is found in 70–90 % of the sporadic ccRCC (Banks et al. 2006; Herman et al. 1994; Kim and Kaelin 2004). The pathophysiological mechanism for such strong organ specificity is currently unknown. The protein encoded by the VHL gene is best known as the substrate-binding subunit of an E3 ubiquitin ligase (Kamura et al. 1999; Lisztwan et al. 1999; Lonergan et al. 1998; Pause et al. 1998; Stehbens et al. 2006). However, accumulated evidence has indicated that many HIF-independent activities of VHL also exist (Frew and Krek 2007; Hsu 2012). Some of these functions are mediated through stabilizing VHL targets, contrary to its known E3 ligase activity. Therefore, VHL is a multifunctional adaptor protein that, depending on the interacting partners, can promote protein degradation or serve as a chaperon. Among these noncanonical functions is the evolutionarily conserved activity in regulating endocytosis (Hsu 2012). One important manifestation of the loss of this activity is the over-accumulation of fibroblast growth factor receptor (FGFR) on the cell surface. This mechanism has been demonstrated in Drosophila trachea morphogenesis (Hsouna et al. 2010), ccRCC cell motility (Hsu et al. 2006) and the angiogenic potential of mouse and human endothelial cells (Champion et al. 2008). This activity is the base of the recent clinical trial of FGFR antagonist for ccRCC patients (Angevin et al. 2013; Escudier et al. 2014; Sonpavde et al. 2014). The C.<H. Lin Department of Surgery, Cathay General Hospital, Taipei, Taiwan, Republic of China T. Hsu (*) Department of Medicine, Boston University School of Medicine, Boston, MA 02118, USA e-mail: [email protected]V. Dammai : B. Adryan Department of Pathology and Laboratory Medicine, Medical University of South Carolina, Charleston, SC 29425, USA C.<H. Lin : T. Hsu Graduate Institute of Systems Biology and Bioinformatics, National Central University, Chung-li, Taiwan 32001, Republic of China Present Address: B. Adryan Cambridge Systems Biology Center, Tennis Court Road, Cambridge, UK Naunyn-Schmiedeberg's Arch Pharmacol DOI 10.1007/s00210-014-1002-4
Transcript
ORIGINAL ARTICLE
Interaction between Nm23 and the tumor suppressor VHL
Chih-Hung Lin & Vincent Dammai & Boris Adryan &
Tien Hsu
Received: 28 April 2014 /Accepted: 2 June 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Among the anti-tumor genes (tumor suppressorsand metastasis suppressors), the von-Hippel Lindau geneand the Nm23 family of genes are among the more intriguingones. Both are small (long and short forms of VHL are 30 and19 kD, respectively, and Nm23 is ~17 kD), and both possessdiverse molecular and cellular functions. Despite extensivestudies, the entire spectra of functions and the molecularfunction-phenotype correlation of these two proteins havenot been completely elucidated. In this report, we present datashowing these two proteins interact physically. We also sum-marize and confirm the previous studies that demonstrated theendocytic function of these two genes and further show thatthe endocytic function of VHL is mediated through the activ-ity of Nm23. These functional and molecular interactions areevolutionarily conserved from Drosophila to human.
Keywords vonHippel-Lindau . Nm23 . Endocytosis .
Tumor suppressor gene
Introduction
The von Hippel-Lindau tumor suppressor gene (VHL) muta-tions are the genetic defects in the familial VHL disease (Latifet al. 1993), which manifests in a limited number of organ-specific tumors (Maher et al. 2011), predominantly in kidney[clear-cell renal cell carcinoma (ccRCC)] and central nervoussystem (hemangioblastoma). Up to 70 % of the carriers ofgerm-line VHLmutations eventually develop either ccRCC orhemangioblastoma or both (Lonser et al. 2003). In addition,loss of VHL function, including somatic mutations and epige-netic defects, is found in 70–90 % of the sporadic ccRCC(Banks et al. 2006; Herman et al. 1994; Kim and Kaelin2004). The pathophysiological mechanism for such strongorgan specificity is currently unknown.
The protein encoded by the VHL gene is best known as thesubstrate-binding subunit of an E3 ubiquitin ligase (Kamuraet al. 1999; Lisztwan et al. 1999; Lonergan et al. 1998; Pauseet al. 1998; Stehbens et al. 2006). However, accumulatedevidence has indicated that many HIF-independent activitiesof VHL also exist (Frew and Krek 2007; Hsu 2012). Some ofthese functions are mediated through stabilizing VHL targets,contrary to its known E3 ligase activity. Therefore, VHL is amultifunctional adaptor protein that, depending on theinteracting partners, can promote protein degradation or serveas a chaperon. Among these noncanonical functions is theevolutionarily conserved activity in regulating endocytosis(Hsu 2012). One important manifestation of the loss of thisactivity is the over-accumulation of fibroblast growth factorreceptor (FGFR) on the cell surface. This mechanism has beendemonstrated in Drosophila trachea morphogenesis (Hsounaet al. 2010), ccRCC cell motility (Hsu et al. 2006) and theangiogenic potential of mouse and human endothelial cells(Champion et al. 2008). This activity is the base of the recentclinical trial of FGFR antagonist for ccRCC patients (Angevinet al. 2013; Escudier et al. 2014; Sonpavde et al. 2014). The
C.<H. LinDepartment of Surgery, Cathay General Hospital, Taipei, Taiwan,Republic of China
T. Hsu (*)Department of Medicine, Boston University School of Medicine,Boston, MA 02118, USAe-mail: [email protected]
V. Dammai : B. AdryanDepartment of Pathology and Laboratory Medicine, MedicalUniversity of South Carolina, Charleston, SC 29425, USA
C.<H. Lin : T. HsuGraduate Institute of Systems Biology and Bioinformatics, NationalCentral University, Chung-li, Taiwan 32001, Republic of China
Present Address:B. AdryanCambridge Systems Biology Center, Tennis Court Road, Cambridge,UK
mechanism of this intriguing endocytic function has remainedunresolved, however.
The endocytosis-based anti-tumor functions have beennoted. One prominent example is the tumor susceptibilitygene 101 (Tsg101) protein, which is a component of theESCRT (endosomal sorting complexes required for transport)involved in early endosome to late endosome transition(Gilbert and Moberg 2006; Vaccari et al. 2008). Anotherexample is the metastasis suppressor gene Nm23.
The Nm23 family of genes [or the unified name for thegenomic locus (NME)] encodes nucleoside diphosphate ki-nases (NDPKs), which generate nucleoside triphosphatesfrom cognate nucleoside diphosphates using ATP as the phos-phate source (Hartsough and Steeg 2000). In the late 1980s,mouse NME1 (Nm23M1) was identified as a potential metas-tasis suppressor gene when it was isolated as a cDNA clonedownregulated in metastatic derivatives of a murine melano-ma cell line K-1735 (Steeg et al. 1988). Subsequent studieshave identified a myriad of molecular and cellular functions,not unlike the multifunctional nature of VHL (Nallamothuet al. 2009). Interestingly, one of these functions is also theactivity in regulation of endocytosis (Hsu 2011).
In this report, we summarize the findings that link thesetwo noncanonical anti-tumor gene functions and show evi-dence that VHL and Nm23 interact molecularly andfunctionally.
Methods and materials
Yeast two-hybrid screen
We used the LexA fusion system (Fashena et al. 2000) previ-ously available from Clontech. The system uses bacterialoperators/regulators and allows for placement of the bait(LexA-VHL fusion) under control of the yeast inducibleGAL operon, such that the bait is expressed only in theabsence of glucose and in the presence of galactose/raffinose. Drosophila cDNA library in the pB42AD(TRP1) vector, yeast strain EGY48 [p8op-lacZ, URA3] (ura3,his3, trp1, and LexAop(x6)-LEU2), and the pGilda (HIS3) vectorwere purchased from Clontech.Drosophila VHL open-readingframe (ORF) was cloned in pGilda and fused to LexA undercontrol of the GAL1 promoter. Antibodies used for detectionand co-immunoprecipitation are: Anti-LexA rabbit polyclonal(at 0.1 μg/mL; EMD Millipore), high-affinity anti-hemagglutinin (HA)-peroxidase conjugate (at 5 mU/mL;Roche), protein A-horse radish peroxidase (HRP; at 1:2,500dilution; Life Technologies), and protein A-agarose (BioRad).Complete mini-protease inhibitor cocktail tablets (Roche,Indianapolis, IN, USA) were dissolved in Co-IP buffer (seebelow) to result in five times protease inhibitor cocktail stock.
After initial selection on minimal SD-His, SD-Ura, andSD-Trp (no leucine), the cotransformants were replica-platedonto permissive media (minimal SD media with SD-His, SD-Leu, SD-Ura, and SD-Trp dropout supplements) to select forLEU reporter gene expression. Selected clones that passedthese initial screening criteria were streaked onto either min-imal media glucose plates to repress bait expression (Fig. 1,Image 1) or onto minimal media galactose/raffinose plates toallow bait expression (Fig. 1, Image 2).
The positive yeast clones were grown in 5-mL permissivemedia till 1.5 OD600 and pelleted. Cell pellet was resuspendedin 400 μL of Co-IP buffer (50 mM Tris–HCl, pH 7.5/100 mMNaCl/10 % glycerol) and 100 μL of five times proteaseinhibitor cocktail. The suspension was sonicated 3×15 s with1 min cooling on ice between pulses. The lysate was spun at10,000×g for 10 min at 4 °C, and the supernatant (cleared celllysa te ) was then subjec ted to Western and co-immunoprecipitation (co-IP) analyses. Co-IP was performedusing anti-LexA antibody (at 0.1 μg/mL) followed by proteinA-agarose pull-down according to manufacturer 'sinstructions.
Drosophila genetics
y1, w67c23 was used as the wild-type stock in this study. ThedVHL and awd alleles have been described (Dammai et al.2003; Duchi et al. 2010). shi2 is a temperature-sensitive alleleof shibire that encodes Drosophila dynamin (Kitamoto 2001;Krishnan et al. 2001). Phenotypes were induced by incubatingembryos at 29 °C. Genetic recombination was used to incor-porate 1-eve-1, a transgenic line expressing the lacZ reportergene in all tracheal cells at all stages from the trachealess genepromoter (Perrimon et al. 1991) (a gift from A. Brand,University of Cambridge). The dVHL head-to-head duplexof the ORF (dVHLi) was cloned in the pCaSpe-hs vectorsfor hsp70-directed expression. The expression vector wereused to transform the y w flies.
Immunostaining of whole-mount Drosophila embryofollowed the standard protocol (Dammai et al. 2003;Nallamothu et al. 2008). Chicken polyclonal anti-β-Gal(1:500; AbCam), rabbit polyclonal anti-dVHL (Duchi et al.2010) and rabbit polyclonal anti-Awd antibody (Dammai et al.2003; Woolworth et al. 2009) were used. For colorimetricreaction, VectaStain (Vector) reagents were used. For immu-nofluorescence, AlexaFluor 488 or 546-conjugated secondaryantibodies (Invitrogen) were used at 1:200 dilution and incu-bation performed for 2 h at room temperature. Stained sampleswere examined with an Olympus BH20 bright field micro-scope or an Olympus IX70 microscope equipped with theFluoview 300 confocal capability. Digital images were proc-essed using the Photoshop software without biasedmanipulations.
Naunyn-Schmiedeberg's Arch Pharmacol
Cell lines and reagents
Human VHL null RCC line 786-O and VHL+ human embry-onic kidney HEK293 cells were from American Type CultureCollection (ATCC) and were usedwithin 8 passages. 786-Vec,786-EGFP and 786-VHL cells were generated by stable trans-fection of 786-O cells with pCDNA3.1, pCMV-EGFP, orpCMV-VHL (see below), respectively, and polyclonal selec-tion by G418 (Life Technologies). Cells were maintained inDMEM (high glucose) supplemented with 10% dialyzed fetalbovine serum (Invitrogen) and used within eight passages.G418 was removed in all assay conditions. Transfection ofHEK293 with Lipofectamine 2000 was performed in a six-well format according to the supplier's protocol (Invitrogen).Electroporation using Nucleofector (Lonza) was performed
with Solution T and program T-01 supplied by the vendor,which consistently achieved ~75 % transfection efficiency.
For FGFR receptor localization/activity assay, 200 ng/mLof recombinant human bFGF (Promega) was mixed with2.5 μg/mL heparin (Sigma) in serum-free media (with 1 %BSA) and added to serum-starved cells. Cells were incubatedat 4 °C for 2 h to allow ligand-receptor engagement withoutinternalization, washed with cold DMEM, and chase initiatedin pre-warmed DMEM (37 °C). At designated time, cells areprocessed for immunofluorescence or Western analysis.
Plasmid constructs
pCMV-EGFP and pCMV-VHL were constructed by PCR-cloning EGFP and human VHL into EcoRV site of
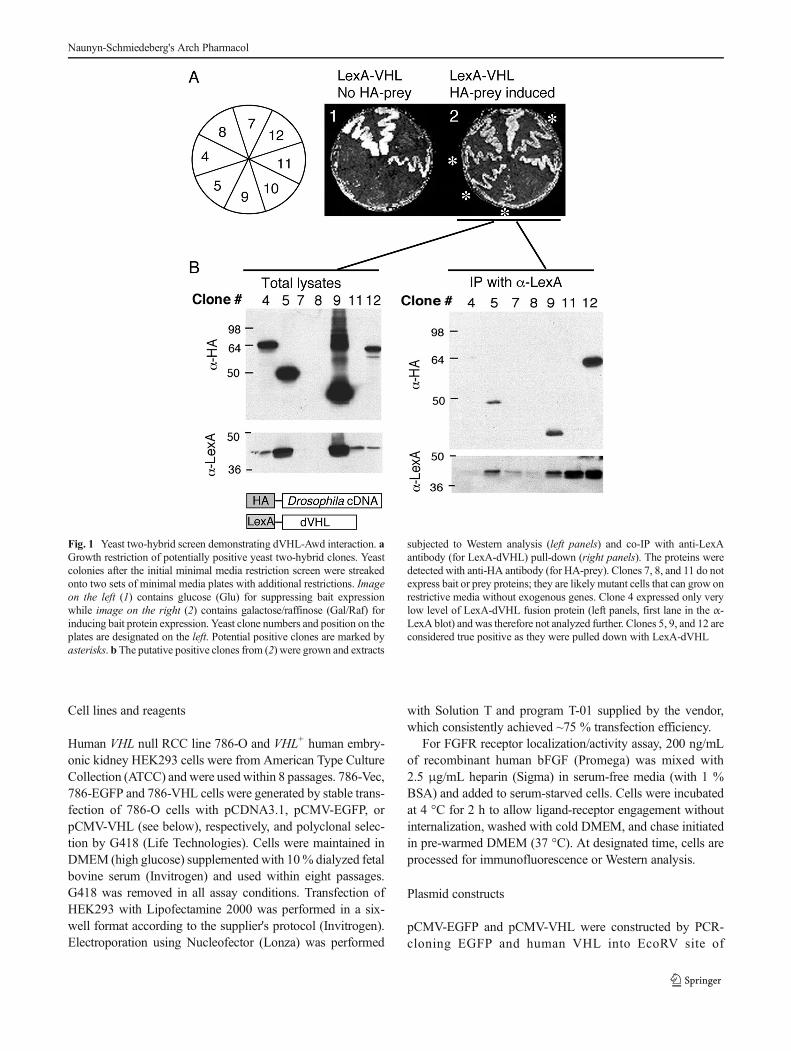
Fig. 1 Yeast two-hybrid screen demonstrating dVHL-Awd interaction. aGrowth restriction of potentially positive yeast two-hybrid clones. Yeastcolonies after the initial minimal media restriction screen were streakedonto two sets of minimal media plates with additional restrictions. Imageon the left (1) contains glucose (Glu) for suppressing bait expressionwhile image on the right (2) contains galactose/raffinose (Gal/Raf) forinducing bait protein expression. Yeast clone numbers and position on theplates are designated on the left. Potential positive clones are marked byasterisks. b The putative positive clones from (2) were grown and extracts
subjected to Western analysis (left panels) and co-IP with anti-LexAantibody (for LexA-dVHL) pull-down (right panels). The proteins weredetected with anti-HA antibody (for HA-prey). Clones 7, 8, and 11 do notexpress bait or prey proteins; they are likely mutant cells that can grow onrestrictive media without exogenous genes. Clone 4 expressed only verylow level of LexA-dVHL fusion protein (left panels, first lane in the α-LexA blot) and was therefore not analyzed further. Clones 5, 9, and 12 areconsidered true positive as they were pulled down with LexA-dVHL
Naunyn-Schmiedeberg's Arch Pharmacol
pCDNA3.1 (Invitrogen). For EGFP-VHL, human VHL wasPCR-cloned into XhoI-KpnI sites of pEGFP-C1 (Clontech).Plasmid-based shRNAs were constructed as follows: targetsequences (NM23H1-shRNA1 gtgagcgtaccttcattgc;NM23H1-shRNA2 ggtgaaatacatgcactca) and control (randomsequence ctactcagtatgcacgtcg) were cloned intopSuppressor-Neo according to instructions (Imgenex).The U6 promoter-shRNA cassette (BamHI-BglII frag-ment) was subcloned into BglII site of pCMV-EGFPand screened for promoters (U6 and CMV) oriented inthe opposite direction. GST fusions of human VHLwere prepared by first cloning VHL ORF intopGEX2T (BamH1-EcoR1) followed by PCR amplifica-tion of GST or GST-VHL and subcloning into BglII-EcoR1 of pIRES-Neo(3) (BD Biosciences). EGFP fu-sions of Nm23-H1 and Nm23-H2 were constructed byRT-PCR amplification of the ORFs and cloning intopEGFP-C1 (Xho1-Kpn1).
Indirect immunofluorescence
Cells were fixed in PBS+3.7 % formaldehyde for 20 min,quenched with PBS+10 mM Tris–HCl (pH 7.4) and perme-abilized with 0.15 % Saponin (Sigma) for 10 min at roomtemperature. Incubation with respective primary and second-ary antibodies was in PBS+1 % BSA (1 h at RT). Primaryantibodies were used at 1:100 dilution: mouse monoclonalanti-GFP (Santa Cruz), mouse monoclonal anti-p-FGFR1(Cell Signaling), mouse monoclonal anti-Nm23H1(Biomeda), and rabbit polyclonal anti-VHL (Cell Signaling).Secondary antibodies are highly cross-absorbed goat anti-rabbit-Alexa 546, goat anti-mouse-Alexa 546, goat anti-rabbit-Alexa 488, and goat anti-mouse-Alexa 488(Molecular Probes), which were used at 1:200 dilution.Confocal images were acquired with Olympus IX70(Fluoview 300).
Antibodies used in Western blotsand co-immunoprecipitation
All antibodies were used at recommended dilutions. Mousemonoclonal antibodies are: dp-ERK1/2 (Sigma-Aldrich), β-actin (Sigma), GFP (Santa Cruz), Nm23H1 (Biomeda), andGST (Santa Cruz). Rabbit polyclonal antibodies are: ERK1/2(Sigma) and VHL (Cell signaling). Goat polyclonal anti-GST(AbCam) was used for co-IP. HRPO-conjugated goat anti-mouse IgG (Sigma-Aldrich) and goat anti-rabbit IgG(Sigma-Aldrich) were used as secondary antibodies.
Results
Drosophila Nm23 is a VHL-interacting protein
In order to analyze the function of VHL, we conducted ayeast-two-hybrid (Y2H) screen for VHL-interacting proteins.For this purpose, we reasoned that using a genetically simplesystem such asDrosophilawould allow us to identify the coreactivity of this evolutionarily conserved protein (Hsu 2012).In this screen, the full-length Drosophila VHL (dVHL) cDNAwas tagged with the DNA-binding LexA peptide readingframe (the bait), and the prey protein coding sequences,consisting of the repertoire protein ORFs generated from aDrosophila full-length cDNA library, were fused with a tran-scription activation domain and tagged with the hemaggluti-nin (HA) peptide coding sequence (the preys). The Y2Hscreen is based on the principle that when the bait proteininteracts with the prey, a bimodular (DNA-binding+transcrip-tion activation domains) transcription factor is formed. Thebimodular bait+prey can activate the reporter gene that allowsfor growth on restrictive media. Figure 1 shows the second-round screen of the initial positive clones. In this screen,clones 7, 8, and 11 were false positives as they could growwithout induction of the prey protein genes (Fig. 1a). Theseclones also did not express any prey proteins (Fig. 1b, leftpanels), indicating that they carry intrinsic mutations thatbypassed the growth restriction. Clone 10 failed to grow ininduced or uninduced prey samples (Fig. 1a) and was notanalyzed further. Among the four positive clones, clone 4failed to show strong interaction with the bait (dVHL) insubsequent pull-down assay (Fig. 1b, right panels), probablydue to the low expression level of LexA-VHL fusion protein.This clone was therefore set aside. The remaining three truepositive clones that also showed strong interaction with LexA-VHL by co-immunoprecipitation (Fig. 1b, right panels) weresequenced. Interestingly, clone 9 turned out to be theDrosophila Nm23 homolog, awd (Biggs et al. 1990;Nallamothu et al. 2008).
Drosophila awd and VHL functionally interact
Both dVHL and awd have been shown to modulate trachealtubule migration during embryogenesis. In embryo, trachealmorphogenesis follows a stereotypical branching pattern thatis mainly guided by the chemotactic signals provided by theFGFR signaling system (Sutherland et al. 1996). In dVHL andawdmutants, the tracheal tubule system is disrupted, showingectopic branching morphogenesis (Adryan et al. 2000;Dammai et al. 2003; Hsouna et al. 2010). In awd mutant,FGFR accumulates on the tracheal cell surface, which leads toincreased and persistent MAPK signaling and ectopic tubulemigration (Dammai et al. 2003). The phenotype is similar tothat of the dynamin mutant shibire (shi). In addition, reduction
Naunyn-Schmiedeberg's Arch Pharmacol
of the gene dosage of the FGFR or the downstream signalingpathway component pnt (Drosophila Ets transcription factorgene) could alleviate the awd phenotype. Conversely, awd-shiheterozygous-hemizygous double mutant could exacerbatethe phenotypes of the single mutants. Therefore, awd modu-lates the tracheal tubule migration by downregulate the sur-face FGFR level via endocytosis.
We confirmed these results by performing additional ge-netic interactions between dVHL, awd and shi mutants. Asshown in Fig. 2, compared with wild-type (Fig. 2a), dVHL andawd homozygous mutants exhibit severe tracheal tubulebreakdown with the appearance of tracheal cell detached fromthe tracheal epithelia. These cells also extend cellular process-es suggesting increased motility (Fig. 2b, c). The penetranceof these homozygous mutants is >85 % (Fig. 2g). Suchphenotype is similar to that of the shi hemizygous males orhomozygous females (shi is a X-linked gene) (Fig. 2d). dVHLheterozygous mutants, on the other hand, showed moderate
Fig. 2 Genetic interaction between dVHL and awd. a–f All strains werecombined with the 1-eve-1 transgenic marker that expresses β-Gal in thetrachea (from the trachealess promoter). Stages 14 and 15 embryos werestained with anti-β-Gal antibodies. Empty arrowheads point to breaks inthe main tracheal trunks. Sharp arrows point to mesenchymal cells thatare dissociated from the epithelia. g Quantification of the tracheal pheno-types in designated genotypes. Ectopic migration is defined as the ap-pearance of ectopic branches and mesenchymal tracheal cells. Moderatephenotype denotes fewer than five tubule breaks and detached cells(combined). Severe phenotype is above 5 of these defects. Statisticalanalysis used Student's t test. *p≤0.05 is considered statisticalsignificant. Bars are 50 μm
Fig. 3 dVHL interacts with Awd functionally and molecularly in vivo. aWild-type (yw; 1-eve-1/+) or dVHL1 (yw; dVHL1/dVHL1; 1-eve-1/+)embryos were processed for immunofluorescence detection of the pro-teins indicated on top. Awd level is diminished in dVHL mutant, con-comitant with disrupted tracheal tubules. Bars are 50 μm. b yw embryosor yw containing one copy or two copies of hsp-dVHLi (expressing dVHLduplex under heat-shock promoter control) were untreated or heat-treatedtwice a day for 30 min each at 37 °C for 3 days. Embryonic extracts wereWestern blotted for the indicated proteins. dVHL levels are knockeddown by dVHLi in a dosage-dependent manner and Awd levelsare reduced in step with the reduced dVHL levels. c Embryoscarrying btl-Gal4; UAS-dVHL::HA (dVHL-HA fusion expressed inthe btl-positive tracheal cells) were collected at 29 °C and extractsprepared; 50 μg of extracts were incubated with rabbit polyclonalanti-Awd or goat polyclonal anti-HA as indicated on top. Immu-noprecipitation was performed using protein A-agarose beads. Thepull-down fraction was then Western blotted using rabbit anti-dVHL antibody, chicken anti-human Nm23 (which cross-reactswith Awd), or mouse anti-actin antibodies
Naunyn-Schmiedeberg's Arch Pharmacol
disruption of tracheal tubules with ectopic branches and oc-casional mesenchymal cells with moderate penetrance(Fig. 2e). awd heterozygotes showed similar haploinsufficientphenotypes (Dammai et al. 2003) (Fig. 2g). When dVHL andawd heterozygotes were combined, the transheterozygousdouble mutants showed severe tracheal phenotypes indistin-guishable from the cognate homozygous mutants with a 97 %penetrance (Fig. 2f, g). Similar genetic interaction was ob-served between dVHL and shi (Fig. 2g). The severity of awd-shi double mutant phenotype is similar to those in dVHL orawd homozygotes and shi hemizygotes (Fig. 2b–d) but with ahigher penetrance. Thus, the genetic interaction suggests anagonistic functional relationship between dVHL and awd inregulating epithelial tubule cell migration, likely via regula-tion of endocytosis.
The agonistic relationship between VHL and Nm23 isinconsistent with the role of VHL as an E3 ubiquitin ligase.However, besides the canonical E3 ubiquitin ligase function,VHL has been shown to act as a chaperon that stabilizes otherproteins, including p53, JADE-1, and kinesin-2 (Chitaliaet al. 2008; Lolkema et al. 2007; Roe and Youn 2006).We therefore examined the expression level of Awd indVHL mutant embryos. As shown in Fig. 3a, the overallAwd protein expression in the embryo was reduced indVHL mutant. The reciprocal experiment showed thatdVHL protein expression was not affected in awd mu-tant (data not shown; also see below). To exclude thepossibility that the reduced expression of Awd was anindirect effect due to degeneration of the mutant embry-os, we examined lysates from embryos expressing short-
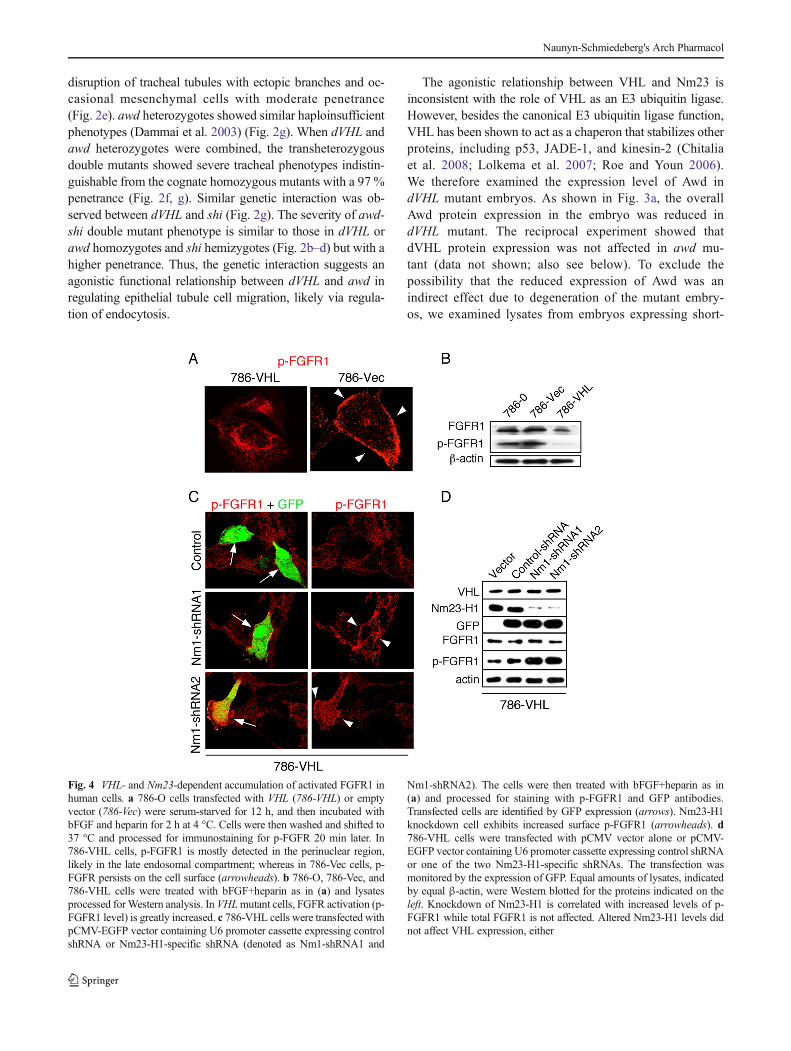
Fig. 4 VHL- and Nm23-dependent accumulation of activated FGFR1 inhuman cells. a 786-O cells transfected with VHL (786-VHL) or emptyvector (786-Vec) were serum-starved for 12 h, and then incubated withbFGF and heparin for 2 h at 4 °C. Cells were then washed and shifted to37 °C and processed for immunostaining for p-FGFR 20 min later. In786-VHL cells, p-FGFR1 is mostly detected in the perinuclear region,likely in the late endosomal compartment; whereas in 786-Vec cells, p-FGFR persists on the cell surface (arrowheads). b 786-O, 786-Vec, and786-VHL cells were treated with bFGF+heparin as in (a) and lysatesprocessed for Western analysis. In VHLmutant cells, FGFR activation (p-FGFR1 level) is greatly increased. c 786-VHL cells were transfected withpCMV-EGFP vector containing U6 promoter cassette expressing controlshRNA or Nm23-H1-specific shRNA (denoted as Nm1-shRNA1 and
Nm1-shRNA2). The cells were then treated with bFGF+heparin as in(a) and processed for staining with p-FGFR1 and GFP antibodies.Transfected cells are identified by GFP expression (arrows). Nm23-H1knockdown cell exhibits increased surface p-FGFR1 (arrowheads). d786-VHL cells were transfected with pCMV vector alone or pCMV-EGFP vector containing U6 promoter cassette expressing control shRNAor one of the two Nm23-H1-specific shRNAs. The transfection wasmonitored by the expression of GFP. Equal amounts of lysates, indicatedby equal β-actin, were Western blotted for the proteins indicated on theleft. Knockdown of Nm23-H1 is correlated with increased levels of p-FGFR1 while total FGFR1 is not affected. Altered Nm23-H1 levels didnot affect VHL expression, either
Naunyn-Schmiedeberg's Arch Pharmacol
hairpin dVHL sequence under heat-shock promoter con-trol (RNA interference). Consistent with the whole-mount immunohistochemistry analysis, knockdown ofdVHL was accompanied by reduced expression of Awdin a dosage-dependent manner (Fig. 3b).
Since the Y2H screen suggests a physical interaction be-tween dVHL and Awd, we sought to confirm the interaction inthe embryos. For this experiment, we used a transgenic systemto express tagged dVHL specifically in the trachea: btl-Gal4;UAS-dVHL::HA, as currently available dVHL (or mammalianVHL) antibodies are not suitable for immunoprecipitation.Weshow that Awd and dVHL indeed interact either directly orindirectly as they can be pulled down with either HA or Awdantibodies from whole embryonic extracts (Fig. 3c).
Functional and molecular interaction between human Nm23and VHL
We next examined whether the functional and molecularinteractions between VHL and Nm23 were conserved inhuman cells. We have shown previously that in humanccRCC cell line 786-O, which is VHL null, FGFR1 accumu-lates on the cell surface as assayed by surface biotinylation
and immunofluorescence, and that the accumulation of sur-face FGFR1 results in elevated MAPK signaling andincreased cell motility (Hsu et al. 2006). The samemechanism was also demonstrated in Drosophila andin mouse primary kidney tubule cells (Hsouna et al.2010). Therefore, the VHL-FGFR-MAPK axis that mod-ulates epithelial cell motility appears to be conservedfrom flies to human.
Here, we verify the result by comparing the expressionpattern of activated FGFR1 (p-FGFR) at 20 min after bFGFstimulation, at which time the endocytic process normallyshould proceed past early endosomes and into late endosomes(Marchese et al. 1998; Maxfield and McGraw 2004). Asshown in Fig. 4a, in 786-O cells transfected with VHL, p-FGFR mostly localizes to the perinuclear region, likely inthe late endosomes. By contrast, VHL mutant 786-Ocells transfected with empty vector show persistent p-FGFR accumulation on the surface. The VHL action onFGFR activity is verified as VHL mutant cells (786-Oand 786-Vec) show significantly increased p-FGFRlevels but only moderately increased total FGFR1 levels(Fig. 4b). Significantly, the FGFR accumulation pheno-type could be recreated when Nm23-H1 was knocked
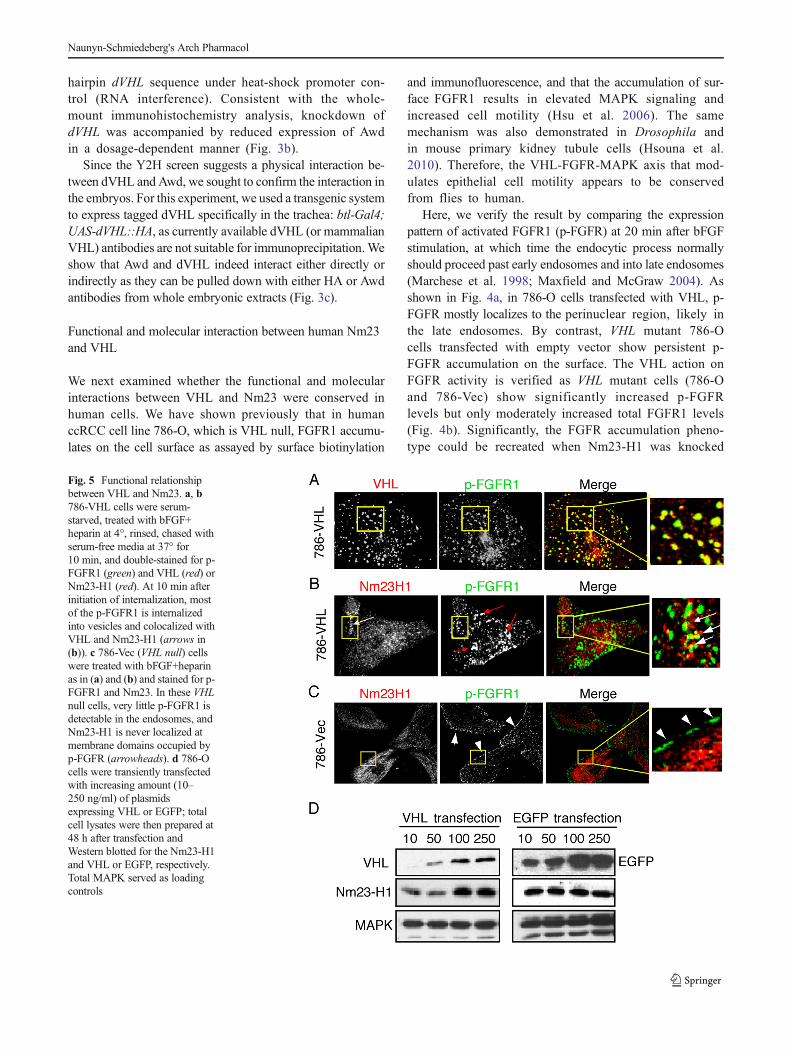
Fig. 5 Functional relationshipbetween VHL and Nm23. a, b786-VHL cells were serum-starved, treated with bFGF+heparin at 4°, rinsed, chased withserum-free media at 37° for10 min, and double-stained for p-FGFR1 (green) and VHL (red) orNm23-H1 (red). At 10 min afterinitiation of internalization, mostof the p-FGFR1 is internalizedinto vesicles and colocalized withVHL and Nm23-H1 (arrows in(b)). c 786-Vec (VHL null) cellswere treated with bFGF+heparinas in (a) and (b) and stained for p-FGFR1 and Nm23. In these VHLnull cells, very little p-FGFR1 isdetectable in the endosomes, andNm23-H1 is never localized atmembrane domains occupied byp-FGFR (arrowheads). d 786-Ocells were transiently transfectedwith increasing amount (10–250 ng/ml) of plasmidsexpressing VHL or EGFP; totalcell lysates were then prepared at48 h after transfection andWestern blotted for the Nm23-H1and VHL or EGFP, respectively.Total MAPK served as loadingcontrols
Naunyn-Schmiedeberg's Arch Pharmacol
down in 786-VHL cells (Fig. 4c). Consistent with theVHL mutant phenotype, knockdown of Nm23-H1 alsoresults in increased FGFR1 activity (p-FGFR1 levels)with little effect on the total FGFR1 levels (Fig. 4d).These results indicate that VHL function requiresNm23-H1, as loss of Nm23-H1 after reexpression ofVHL in 786-O cells remain defective in FGFR1 inter-nalization and FGFR1 signal processing.
The endocytic role of VHL and Nm23H1 is supported bythe demonstration that in VHL positive cells, p-FGFR1 colo-calizes with VHL and Nm23-H1 in the early endosomes at10 minutes after induction of endocytosis (Fig. 5a, b). We notethat the extent of association between Nm23 and p-FGFR1 isless than that of VHL and p-FGFR1. This is probably becauseeither Nm23 has other cellular functions than endocytosis, orthe association betweenNm23 and endosome is transient.Mostimportantly, the association of Nm23-H1 with endosomes islost in VHLmutant 786-O cells (Fig. 5c). In agreement with theDrosophila system, increasedVHL protein levels are correlatedwith increased Nm23-H1 levels (Fig. 5d). Taken together, theseresults indicate that the endocytic role of VHL is mediated viastabilization and proper localization of Nm23. To examinewhether VHL and Nm23-H1 interact in human cells, weexpressed tagged Nm23 (either H1 or H2 isoform) and VHL
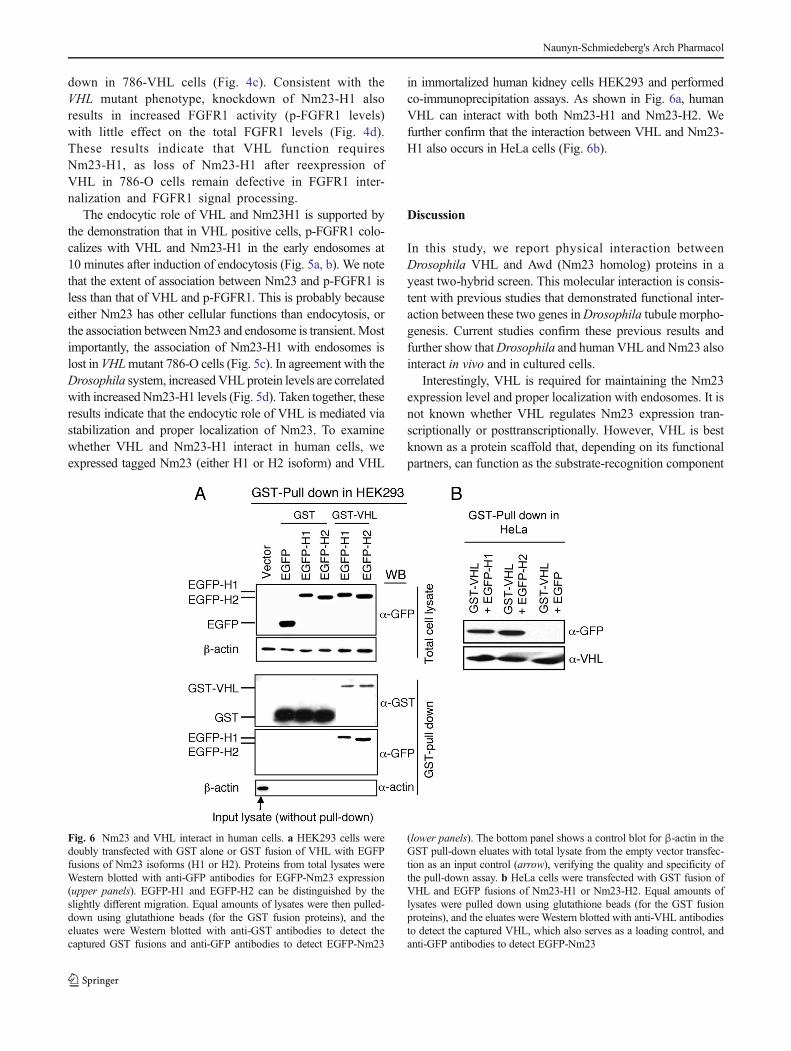
in immortalized human kidney cells HEK293 and performedco-immunoprecipitation assays. As shown in Fig. 6a, humanVHL can interact with both Nm23-H1 and Nm23-H2. Wefurther confirm that the interaction between VHL and Nm23-H1 also occurs in HeLa cells (Fig. 6b).
Discussion
In this study, we report physical interaction betweenDrosophila VHL and Awd (Nm23 homolog) proteins in ayeast two-hybrid screen. This molecular interaction is consis-tent with previous studies that demonstrated functional inter-action between these two genes inDrosophila tubule morpho-genesis. Current studies confirm these previous results andfurther show thatDrosophila and human VHL and Nm23 alsointeract in vivo and in cultured cells.
Interestingly, VHL is required for maintaining the Nm23expression level and proper localization with endosomes. It isnot known whether VHL regulates Nm23 expression tran-scriptionally or posttranscriptionally. However, VHL is bestknown as a protein scaffold that, depending on its functionalpartners, can function as the substrate-recognition component
Fig. 6 Nm23 and VHL interact in human cells. a HEK293 cells weredoubly transfected with GST alone or GST fusion of VHL with EGFPfusions of Nm23 isoforms (H1 or H2). Proteins from total lysates wereWestern blotted with anti-GFP antibodies for EGFP-Nm23 expression(upper panels). EGFP-H1 and EGFP-H2 can be distinguished by theslightly different migration. Equal amounts of lysates were then pulled-down using glutathione beads (for the GST fusion proteins), and theeluates were Western blotted with anti-GST antibodies to detect thecaptured GST fusions and anti-GFP antibodies to detect EGFP-Nm23
(lower panels). The bottom panel shows a control blot for β-actin in theGST pull-down eluates with total lysate from the empty vector transfec-tion as an input control (arrow), verifying the quality and specificity ofthe pull-down assay. b HeLa cells were transfected with GST fusion ofVHL and EGFP fusions of Nm23-H1 or Nm23-H2. Equal amounts oflysates were pulled down using glutathione beads (for the GST fusionproteins), and the eluates were Western blotted with anti-VHL antibodiesto detect the captured VHL, which also serves as a loading control, andanti-GFP antibodies to detect EGFP-Nm23
Naunyn-Schmiedeberg's Arch Pharmacol
of the ubiquitin ligase, or as a chaperon that stabilizes otherproteins (Frew and Krek 2007; Hsu 2012). We show here thatboth VHL and Nm23 are associated with early endosomesduring active endocytosis of activated FGFR1. Importantly,the association of Nm23 with endosomes is disrupted in VHLmutant cells. Nm23/Awd has been suggested as a GTP sup-plier for dynamin or stabilization of other small GTPasesinvolved in endocytosis such as Rab5 (Ignesti et al. 2014;Krishnan et al. 2001; Woolworth et al. 2009). It is thereforepossible that VHL acts as an adaptor protein that brings Nm23to endosomes. ReducedNm23 level inVHLmutant may be anindirect result of improper protein localization.
It is also worth noting that both VHL and Nm23 have beenlinked to microtubule (MT) function. Direct association ofmammalian Nm23 with MTwas the base of the first purifica-tion of the Nm23 protein (at the time known as NDPK)(Nickerson and Wells 1984). Functional and physical associ-ation of VHL and microtubules has also been demonstrated inDrosophila and in mammalian systems (Duchi et al. 2010;Hergovich et al. 2003; Kuehn et al. 2007; Lolkema et al.2007). It will be interesting to determine whether the func-tional partnership between VHL and Nm23 also extends tothis cellular system.
VHLmutations are the main genetic defects in familial andsporadic renal cell carcinoma of the clear-cell type. AlthoughNm23 has been implicated in epithelial maintenance in gen-eral (Boissan et al. 2010), no specific roles of Nm23 in renaldevelopment and renal cell carcinoma progression have beendemonstrated. However, reduced expression of Nm23 hasbeen linked with renal cell carcinoma in several studies(Ayhan et al. 1998; Bosnar et al. 1997; Leone et al. 1991;Nakagawa et al. 1998; Ohba et al. 2005).
Both Nm23 and VHL are multifunctional proteins. VHLmainly serves as a protein scaffold that interacts with an arrayof functionally diverse proteins (Frew and Krek 2008; Hsu2012). Nm23, on the other hand, possess multiple enzymaticactivities, including NDPK, nuclease, histidine-dependentprotein kinase, transcription factor, and secreted growth factor(Steeg et al. 2011). It is curious how such diverse functions arecoordinated subcellularly. Our current finding may suggestthat Nm23 exerts its diverse functions by localizing to differ-ent subcellular loci, which in turn requires association withdifferent protein partners such as VHL.
Acknowledgments This work was supported by a grant from theNational Institutes of Health, USA, # RO1CA109860, to T.H.
References
Adryan B,Decker HJ, Papas TS, Hsu T (2000) Tracheal development andthe von Hippel-Lindau tumor suppressor homolog in Drosophila.Oncogene 19:2803–2811
Angevin E, Lopez-Martin JA, Lin CC, Gschwend JE, Harzstark A,Castellano D, Soria JC, Sen P, Chang J, Shi M, Kay A, Escudier B(2013) Phase I study of dovitinib (TKI258), an oral FGFR, VEGFR,and PDGFR inhibitor, in advanced or metastatic renal cell carcino-ma. Clin Cancer Res 19:1257–1268
Ayhan A, Usubutun A, Ozen H, Yasui W, Tahara E (1998) nm23 proteinexpression in renal cell tumors: the role of the cell type. Oncol Rep5:979–983
Banks RE, Tirukonda P, Taylor C, Hornigold N, Astuti D, Cohen D,Maher ER, Stanley AJ, Harnden P, Joyce A, Knowles M, Selby PJ(2006) Genetic and epigenetic analysis of vonHippel-Lindau (VHL)gene alterations and relationship with clinical variables in sporadicrenal cancer. Cancer Res 66:2000–2011
Biggs J, Hersperger E, Steeg PS, Liotta LA, Shearn A (1990) ADrosophila gene that is homologous to a mammalian gene associat-ed with tumor metastasis codes for a nucleoside diphosphate kinase.Cell 63:933–940
BoissanM,DeWever O, Lizarraga F,WendumD, Poincloux R, ChignardN, Desbois-Mouthon C, Dufour S, Nawrocki-Raby B, Birembaut P,BrackeM, Chavrier P, Gespach C, LacombeML (2010) Implicationof metastasis suppressor NM23-H1 in maintaining adherens junc-tions and limiting the invasive potential of human cancer cells.Cancer Res 70:7710–7722
Bosnar MH, Pavelic K, Hrascan R, Zeljko Z, Krhen I, Marekoyic Z,Krizanac S, Pavelic J (1997) Loss of heterozygosity of the nm23-H1gene in human renal cell carcinomas. J Cancer Res Clin Oncol 123:485–488
Champion KJ, Guinea M, Dammai V, Hsu T (2008) Endothelial functionof von Hippel-Lindau tumor suppressor gene: control of fibroblastgrowth factor receptor signaling. Cancer Res 68:4649–4657
Chitalia VC, Foy RL, Bachschmid MM, Zeng L, Panchenko MV, ZhouMI, Bharti A, Seldin DC, Lecker SH, Dominguez I, Cohen HT(2008) Jade-1 inhibits Wnt signalling by ubiquitylating beta-cateninand mediates Wnt pathway inhibition by pVHL. Nat Cell Biol 10:1208–1216
Dammai V, Adryan B, Lavenburg KR, Hsu T (2003)Drosophila awd, thehomolog of human nm23, regulates FGF receptor levels and func-tions synergistically with shi/dynamin during tracheal development.Genes Dev 17:2812–2824
Duchi S, Fagnocchi L, Cavaliere V, Hsouna A, Gargiulo G, Hsu T (2010)Drosophila VHL tumor-suppressor gene regulates epithelial mor-phogenesis by promoting microtubule and aPKC stability.Development 137:1493–1503
Escudier B, Grunwald V, Ravaud A, Ou YC, Castellano D., Lin CC,Gschwend JE, Harzstark A, Beall S, Pirotta N, Squires M, Shi MM,Angevin E (2014) Phase II Results of Dovitinib (TKI258) in PatientsWith Metastatic Renal Cell Cancer. Clin Cancer Res 20:2012–2022
Hergovich A, Lisztwan J, Barry R, Ballschmieter P, Krek W(2003) Regulation of microtubule stability by the vonHippel-Lindau tumour suppressor protein pVHL. Nat CellBiol 5:64–70
Herman JG, Latif F, Weng Y, LermanMI, Zbar B, Liu S, Samid D, DuanDS, Gnarra JR, Linehan WM et al (1994) Silencing of the VHLtumor-suppressor gene by DNA methylation in renal carcinoma.Proc Natl Acad Sci U S A 91:9700–9704
Naunyn-Schmiedeberg's Arch Pharmacol
Hsouna A, Nallamothu G, Kose N, Guinea M, Dammai V, Hsu T (2010)Drosophila von Hippel-Lindau tumor suppressor gene function inepithelial tubule morphogenesis. Mol Cell Biol 30:3779–3794
Hsu T (2011) NME genes in epithelial morphogenesis. NaunynSchmiedeberg’s Arch Pharmacol 384:363–372
Hsu T (2012) Complex cellular functions of the von Hippel-Lindau tumorsuppressor gene: insights from model organisms. Oncogene 31:2247–2257
Hsu T, Adereth Y, Kose N, Dammai V (2006) Endocytic function of vonHippel-Lindau tumor suppressor protein regulates surface localiza-tion of fibroblast growth factor receptor 1 and cell motility. J BiolChem 281:12069–12080
Ignesti M, Barraco M, Nallamothu G, Woolworth JA, Duchi S, GargiuloG, Cavaliere V, Hsu T (2014) Notch signaling during developmentrequires the function of awd, the Drosophila homolog of humanmetastasis suppressor gene Nm23. BMC Biol 12:12
Kamura T, Koepp DM, Conrad MN, Skowyra D, Moreland RJ,Iliopoulos O, Lane WS, Kaelin WG Jr, Elledge SJ, Conaway RC,Harper JW, Conaway JW (1999) Rbx1, a component of the VHLtumor suppressor complex and SCF ubiquitin ligase. Science 284:657–661
Kim WY, Kaelin WG (2004) Role of VHL gene mutation in humancancer. J Clin Oncol 22:4991–5004
Kitamoto T (2001) Conditional modification of behavior in Drosophilaby targeted expression of a temperature-sensitive shibire allele indefined neurons. J Neurobiol 47:81–92
Krishnan KS, Rikhy R, Rao S, Shivalkar M, Mosko M, Narayanan R,Etter P, Estes PS, Ramaswami M (2001) Nucleoside diphosphatekinase, a source of GTP, is required for dynamin-dependent synapticvesicle recycling. Neuron 30:197–210
Kuehn EW, Walz G, Benzing T (2007) Von hippel-lindau: a tumorsuppressor links microtubules to ciliogenesis and cancer develop-ment. Cancer Res 67:4537–4540
Latif F, Tory K, Gnarra J, Yao M, Duh FM, Orcutt ML, Stackhouse T,Kuzmin I, Modi W, Geil L et al (1993) Identification of the vonHippel-Lindau disease tumor suppressor gene. Science 260:1317–1320
Leone A, McBride OW, Weston A, Wang MG, Anglard P, Cropp CS,Goepel JR, Lidereau R, Callahan R, Linehan WM et al (1991)Somatic allelic deletion of nm23 in human cancer. Cancer Res 51:2490–2493
Lisztwan J, Imbert G,Wirbelauer C, GstaigerM, KrekW (1999) The vonHippel-Lindau tumor suppressor protein is a component of an E3ubiquitin-protein ligase activity. Genes Dev 13:1822–1833
LolkemaMP,Mans DA, Snijckers CM, van Noort M, van BeestM, VoestEE, Giles RH (2007) The von Hippel-Lindau tumour suppressorinteracts with microtubules through kinesin-2. FEBS Lett 581:4571–4576
Lonergan KM, Iliopoulos O, OhhM, Kamura T, Conaway RC, ConawayJW, Kaelin WG Jr (1998) Regulation of hypoxia-inducible mRNAsby the von Hippel-Lindau tumor suppressor protein requires bindingto complexes containing elongins B/C and Cul2. Mol Cell Biol 18:732–741
Lonser RR, Glenn GM, Walther M, Chew EY, Libutti SK, Linehan WM,Oldfield EH (2003) von Hippel-Lindau disease. Lancet 361:2059–2067
Maher ER, Neumann HP, Richard S (2011) von Hippel-Lindau disease: aclinical and scientific review. Eur J Hum Genet 19:617–623
Maxfield FR,McGraw TE (2004) Endocytic recycling. Nat RevMol CellBiol 5:121–132
Nakagawa Y, Tsumatani K, Kurumatani N, Cho M, Kitahori Y, KonishiN, Ozono S, Okajima E, Hirao Y, Hiasa Y (1998) Prognostic valueof nm23 protein expression in renal cell carcinomas. Oncology 55:370–376
Ohba K, Miyata Y, Koga S, Kanda S, Kanetake H (2005) Expression ofnm23-H1 gene product in sarcomatous cancer cells of renal cellcarcinoma: correlation with tumor stage and expression of matrixmetalloproteinase-2, matrix metalloproteinase-9, sialyl Lewis X,and c-erbB-2. Urology 65:1029–1034
Pause A, Lee S, Lonergan KM, Klausner RD (1998) The von Hippel-Lindau tumor suppressor gene is required for cell cycle exit uponserum withdrawal. Proc Natl Acad Sci U S A 95:993–998
Perrimon N, Noll E, McCall K, Brand A (1991) Generating lineage-specific markers to study Drosophila development. Dev Genet 12:238–252
Roe JS, Youn HD (2006) The positive regulation of p53 by the tumorsuppressor VHL. Cell Cycle 5:2054–2056
Sonpavde G, Willey CD, Sudarshan S (2014) Fibroblast growth factorreceptors as therapeutic targets in clear-cell renal cell carcinoma.Expert Opin Investig Drugs 23:305–315
Steeg PS, Bevilacqua G, Kopper L, Thorgeirsson UP, Talmadge JE,Liotta LA, Sobel ME (1988) Evidence for a novel gene associatedwith low tumor metastatic potential. J Natl Cancer Inst 80:200–204
Steeg PS, ZolloM,Wieland T (2011) A critical evaluation of biochemicalactivities reported for the nucleoside diphosphate kinase/Nm23/Awdfamily proteins: opportunities and missteps in understanding theirbiological functions. Naunyn Schmiedeberg’s Arch Pharmacol 384:331–339
Stehbens SJ, Paterson AD, Crampton MS, Shewan AM, Ferguson C,Akhmanova A, Parton RG, Yap AS (2006) Dynamic microtubulesregulate the local concentration of E-cadherin at cell-cell contacts. JCell Sci 119:1801–1811
Sutherland D, Samakovlis C, Krasnow MA (1996) branchless encodes aDrosophila FGF homolog that controls tracheal cell migration andthe pattern of branching. Cell 87:1091–1101
Vaccari T, Lu H, Kanwar R, Fortini ME, Bilder D (2008) Endosomalentry regulates Notch receptor activation in Drosophilamelanogaster. J Cell Biol 180:755–762