Interaction between satellite cells and skeletal muscle fibers
RICHARD BISCHOFF
Department of Anatomy and Neurobiology, Washington University School of Medicine, 4566 Scott Avenue, Saint Louis, MO 63110, USA
Summary
Single myofibers with attached satellite cells isolatedfrom adult rats were used to study the influence of themature myofiber on the proliferation of satellite cells.The satellite cells remain quiescent when cultured inserum containing medium but proliferate when exposedto mitogen from an extract of crushed adult muscle. Theresponse of satellite cells to mitogen was measured underthree situations with respect to cell contact: (1) in contactwith a viable myofiber and its basal lamina, (2) detachedfrom the myofiber by centrifugal force and deposited onthe substratum and (3) beneath the basal lamina of aMarcaine killed myofiber. The results show that satellitecells in contact with the plasmalemma of a viablemyofiber have reduced mitogenic response. Since inhi-biting growth may induce differentiation, I testedwhether satellite cells proliferating on the surface of a
myofiber would fuse. Although the satellite cell progenywere fusion competent, they did not fuse with themyofiber. To determine whether fusion competence ofthe myofiber changes with tune in culture, embryonicmyoblasts were challenged to fuse with myofibers thathad been stripped of satellite cells and cultured forseveral days. The myoblasts fused with pseudopodialsprouts growing from the ends of the myofiber, but didnot fuse with the original myofiber surface. These resultsindicate that contact with the surface of a maturemyofiber suppresses proliferation of myogenic cells butthe cells do not fuse with the myofiber.
Satellite cells of adult skeletal muscle are mononu-cleated cells that occupy shallow depressions in themyofiber. The cells have one surface in contact with thebasal lamina of the myofiber and the other surface incontact with its plasmalemma with a narrow gap ofabout 15 nm between the two cells (Ishikawa, 1966).Despite this intimate contact, there is no evidence ofelectrical coupling between the cells (Bader et al. 1988).Satellite cells of adult muscle have a heterochromaticnucleus and scant cytoplasm with few organelles butboth surfaces of the satellite cell show evidence ofspecialized activity in the form of abundant cell mem-brane caveolae (Schultz, 1976; Snow, 1977; Bischoff,1979). The function of these is unknown. Satellite cellsare mitotically quiescent in adult muscle but re-enterthe cell cycle following muscle injury to generate apopulation of myoblasts, which may subsequently fuseto form new myofibers (Bischoff, 1975; Konigsberg etal. 1975; Snow, 1977). In developing muscle, satellitecells are mitotically active and contribute virtually all ofthe nuclei in the secondary generation of myofibers(Kelly and Zacks, 1969; Cardasis and Cooper, 1975;Kelly, 1978; Ross et al. 1987). Identifying signals thatgovern satellite cell behavior is of central importance inunderstanding muscle growth and regeneration.
A study of the regulation of satellite cell division was
carried out using a tissue culture system consisting ofsingle myofibers with attached satellite cells from adultrats (Bischoff, 1986a, 19866). The satellite cells areretained beneath the basal lamina of these fibers andmost remain mitotically quiescent in vitro unless mito-gens are added to the culture medium. Effective growthstimulators include fibroblast growth factor and anextract of crushed adult muscle (Bischoff, 19866).Results of these studies show that proliferation ofsatellite cells is under positive control since death of acultured myofiber of itself had little effect on prolifer-ation of attached satellite cells, but mitogens extractedfrom whole crushed muscles were able to stimulate thegrowth of satellite cells on intact myofibers. Muscleregeneration is a complex phenomenon, however. Theregenerative response is proportional to the extent oftissue injury (McGeachie and Grounds, 1987) and thefinal outcome is influenced by innervation, vascularity,hormonal status and nutrition (Jirmanova and Thesleff,1972; Hansen-Smith and Carlson, 1979; d'Albis et al.1987; Phillips et al. 1987; Mulvaney et al. 1988). Thus itis likely that satellite cell growth is regulated by bothpositive and negative factors.
Among local factors that may influence satellite cellgrowth are the plasmalemma and the basal lamina ofthe myofiber since satellite cells have extensive area incontact with both of these surfaces. The basal lamina isthought to facilitate regeneration by providing a frame-
944 R. Bischoff
work upon which new myofibers can form and becomereinnervated (Vracko and Benditt, 1972; Sanes et al.1978). The mechanisms by which the basal laminainfluences regeneration are unknown, but may includestimulation of satellite cell growth and division.
The present study utilized myofibers with attachedsatellite cells in culture to evaluate separately theinfluence of the basal lamina and the plasmalemma ofthe myofiber on satellite cell proliferation induced byadding muscle extract to the culture medium. Resultsindicate that satellite cells in contact with the plasma-lemma of the myofiber have reduced sensitivity tomitogen stimulation.
Materials and methods
CulturesSingle isolated fibers and attached satellite cells were pre-pared from the flexor digitorum brevis muscles of 2- to 3-month-old male rats as described previously (Bischoff,1986a). The fibers were separated from non-muscle cells andcultured on coverslips in 35 mm plastic culture dishes. Severalmethods were used to attach fibers to the coverslips depen-ding upon the requirements of the experiment. In someexperiments, the fibers were embedded in a thin gel madefrom 50^1 rat tendon collagen solution containing about3 mg protein ml"1 0.1% acetic acid. After restoring salt andpH to physiological levels, the collagen was mixed with 50 /dof a suspension of myofibers and incubated at 37 °C for 10 minto induce polymerization (Elsdale and Bard, 1972). In exper-iments requiring the fibers to be free on the substratum,coverslips were coated with a thin layer of the rat tendoncollagen prepared as described and polymerized at 37°C. Thecoated coverslips were treated with 1 % glutaraldehyde (Elec-tron Microscopy Sciences, Fort Washington, PA) in phos-phate-buffered saline (PBS) for 1 h at room temperature,washed 5 times with PBS, treated with 0.5mgml"' polylysine(Sigma, St. Louis, MO) in PBS for 30 min and washed oncewith PBS. The treated coverslips were placed in 35 mmculture dishes and myofibers were attached by centrifugalforce (lOOg for 10 min) using a custom-designed dish carrier inan HN-S centrifuge (International Equipment Co., NeedhamHeights, MA). No satellite cells were detached from themyofibers at this centrifugal force (see below). Culturemedium was Eagle's MEM containing 10% horse serum and1% antibiotic-antimycotic mixture (GIBCO, Grand Island,NY). Culture medium was changed daily. In some recentexperiments, the 10% horse serum was replaced with 20%Controlled Process Serum Replacement No.2 (Sigma) plus1 % horse serum. Satellite cells remain viable, but fewproliferate under these conditions. A saline extract of crushedadult rat muscle prepared as described (Bischoff, 19866) wasadded to the culture medium to induce proliferation ofsatellite cells.
Embryonic myoblasts were prepared from 19 day ratembryos by digestion of muscle tissue using 0.2 % crudetrypsin (Sigma, 1:250) in Earle's balanced salt solution for30 min at 37°C. Cells were dissociated by trituration andseparated from clumps and debris by filtration through 10/tmNitex cloth (TETCO, NY). Suspensions were enriched formyogenic cells by preplating (Konigsberg, 1979) and thepurified cells were cultured in Eagle's MEM with 10% horseserum, 5% embryo extract and antibiotics. The embryoextract was prepared from 11 day chick embryos (Bischoff andHoltzer, 1969). All other chemicals were from Sigma.
Separation of satellite cells and myofibersTo remove satellite cells from intact myofibers, a suspensionof myofibers in PBS was placed in 35 mm culture dishesbearing collagen-polylysine-treated coverslips. The disheswere centrifuged in an HN-S centrifuge at full speed (about1500g) for 10 min to allow attachment of the fibers to thesubstratum then the PBS was replaced with culture mediumand centrifugation was continued for another 20 min.
Satellite cells attached to the basal lamina in the absence ofthe myofiber were obtained by killing the myofibers with amyotoxic anaesthetic. Cultures were incubated for 10 min inPBS containing 0.05% Marcaine (Winthrop-Breon, NY),washed twice with PBS and returned to normal culturemedium. Myonuclei in Marcaine-killed myofibers completelylyse after overnight incubation. In some experiments, how-ever, it was necessary to remove the myonuclei more rapidlyand for this 33 j/gmF1 deoxyribonuclease I (Sigma, type IV)was added to the culture medium after Marcaine treatment.
Cell proliferationSatellite cell proliferation was measured by radioautographyafter incubating coverslip cultures in medium containing0.5^Cimr' [/ne//;y/-3H]thymidine ([3H]TdR, 6.7CimM-^New England Nuclear, Boston, MA) from 44 to 48h in vitro.Coverslips were fixed in alcohol, Formalin, acetic acid(20:2:1), attached to slides and dipped in NTB2 bulk emul-sion (Eastman Kodak, Rochester, NY) diluted 1,1 with water.After photographic processing according to the manufac-turer's recommendations, the cells were stained with Gill'shematoxylin (Polysciences, Warrington, PA). Labeled satel-lite cells were scored in at least 50 fibers from each treatment.
ImmunocytochemistryCultures were fixed with 4% formaldehyde in PBS, washedand incubated for 1 h in rabbit antibody against laminindiluted 1:200 with PBS (Bethesda Research Lab, Bethesda,MD). After washing, the cultures were incubated for lh influorescein-labeled goat anti-rabbit (GAR) IgG diluted 1:50(Miles, Elkhart, IN). Control cultures were incubated innormal rabbit serum and fluorescein-GAR IgG orfluorescein-GAR IgG alone.
MicroscopyA Zeiss microscope fitted with an epifluorescent condenserand halogen lamp was used for fluorescent microscopy.Specimens were mounted in glycerol:PBS (9:1), pH8.7. Forscanning electron microscopy, myofibers were centrifugedonto polylysine-coated coverslips and cultured for variousperiods. The cultures were fixed in 3% glutaraldehyde inPBS, dehydrated in methanol, critical point dried, andsputter-coated with 12-15 nm gold. Cultures were examinedwith a Philips 501 microscope.
Results
Experimental paradigmProliferation of satellite cells in response to muscleextract was measured by [3H]TdR labeling in radioau-tographs under three conditions with respect to cellcontact (Fig. 1). First, satellite cells were left in theirnormal location between the basal lamina and plasma-lemma of the myofiber. Second, satellite cells weredetached from the myofiber and basal lamina by centri-fugation and deposited on the culture substratum di-
Myoflbcr KUled
Fig. 1. Diagram of the experimental conditions used todetermine the effect of surface contact on satellite cellproliferation stimulated by muscle extract.
rectly beneath the myofiber. Third, satellite cells wereremoved from the influence of the plasmalemma, butleft in contact with the basal lamina by killing themyofiber. Because of technical limitations, experimentscomparing the three conditions were carried out separ-ately using the normal myofibers as control in each case.In evaluating the results, however, the three conditionsshould be considered together.
Attempts to provide cultures consisting of satellitecells on the myofiber plasmalemma in the absence ofthe basal lamina were unsuccessful. The basal laminacould be removed with trypsin or other enzymes(Bischoff, 1975; Bischoff, 1986a) but satellite cells didnot remain on the myofibers under these conditions.
Proliferation of satellite cells on vs off myofibersA suspension of myofibers with attached satellite cellswas centrifuged in culture dishes bearing coverslips withan adhesive surface (see Material and Methods). Ascentrifugal force was applied, the myofibers attached tothe coverslip within a few seconds, then, during thefollowing half hour of centrifugation, satellite cellsdetached from the myofibers and were deposited on thecoverslip beneath the myofibers. This technique avoids
Satellite cell-myofiber interaction 945
further enzymatic treatment and leaves the satellitecells close to their associated myofiber. Preliminaryexperiments showed that most cells were detached fromthe myofibers after lOmin at 1000g, but a higherforce/time was used routinely to ensure completeremoval of all cells. The myofiber itself appearedunchanged after centrifugation (Fig. 2) and remainedviable in culture. The satellite cells were often de-posited to one side of the myofiber. The unevendistribution may have resulted from the release of cellsbefore the dish holder reached a horizontal position inthe centrifuge. In addition to satellite cells, fibrillardebris and a few small vesicles were deposited on thesubstratum following centrifugation. This material wasnot identified further, but scanning electron microscopyreveals a few collagen fibrils and debris from thedissociation procedure adherent to the surface of thebasal lamina of isolated myofibers before centrifugation(Bischoff, 1986a).
The basal lamina was not removed from the myofiberby centrifugation as evidenced by the persistence ofstaining for laminin, a major component of the basallamina, following immunocytochemistry. Most myo-fibers had several small laminin-negative patches on thesurface (Fig. 3). These are not present in uncentrifugedmyofibers (Bischoff, 1986a) and were interpreted assites at which centrifugal force had removed satellitecells from the surface of the myofiber leaving tears inthe overlying basal lamina.
Satellite cells deposited on the substratum remainedrounded for several hours then spread out as plump,spindle-shaped cells. No satellite cells remained on themyofibers if the centrifugal force was 1500g, althoughat lower forces not all satellite cells were removed. Thecells did not grow in the absence of mitogen but, whenmuscle extract was added, a group of satellite cellprogeny surrounded the myofiber after 2 days in culture(Fig. 4). Although most satellite cells remained in thevicinity of the associated myofiber, some migrated up to100/im away. Only an occasional satellite cell migratedaway from its associated myofiber in control culturesthat were not centrifuged. When grown in 500 ̂ g ml"1
muscle extract for 2 days and labeled with [3H]TdR,48 % (437/905) of satellite cells on the substratum werelabeled while only 38% (410/1078) of myofiber-associ-
Fig. 2. Phase-contrast micrograph of living myofiber immediately after centrifugation at 1500g for 30min to detach satellitecells. Most of the material deposited on the polylysine-coated substratum is debris that adhered to the surface of themyofiber during dissociation. The satellite cells are rounded and cannot be reliably identified by phase-contrast microscopyalthough they can be identified by staining with hematoxylin (not shown). Bar, 25 ;/m.
946 R. Bischoff
B
Fig. 3. Fluorescent (A) and phase-contrast (B) micrographsof living myofiber immediately after centrifugation,incubated with rabbit antibody against laminin followed bygoat anti-rabbit IgG labeled with fluorescein. The unstainedspot (arrow) represents a tear in the basal lamina where asatellite cell was detached from the myofiber and depositedon the substratum below (arrowhead). After several hoursin culture, the satellite cells spread on the substratum.Control cultures incubated with non-immune serum orfluorescein-labeled second antibody alone did not stain.Bar, 50pan.
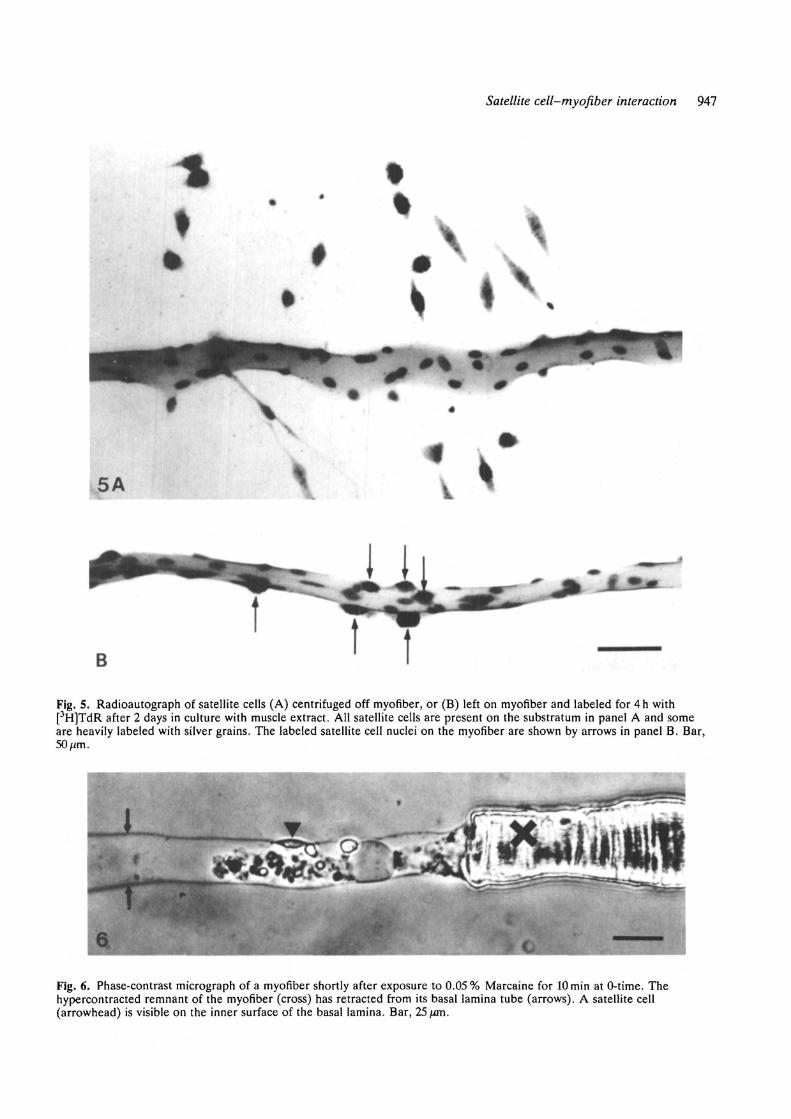
ated satellite cells were labeled in uncentrifuged com-panion cultures (Fig. 5). The number of labeled cellsper myofiber was not counted because the dispersion ofsatellite cells on the substratum during the 2 day cultureperiod made it difficult to reliably assign each satellitecell to a myofiber.
Proliferation of satellite cells on live vs killedmyofibersThe satellite cell is in contact with both the plasma-lemma and the basal lamina of the myofiber. Toseparate the effects of these two surfaces on satellite cellreplication, the myofiber was killed with the myotoxicanaesthetic Marcaine (Hall-Craggs, 1974) leaving thesatellite cells in contact with the basal lamina tube butremoving them from the influence of the myofiber
plasmalemma (Fig. 6). When provided with mitogen,the satellite cells proliferated and most remained withinthe basal lamina tube (Fig. 7). The live and Marcaine-killed myofibers were compared for response to muscleextract (Fig. 8). Satellite cells associated with the killedmyofibers were about 40 % more responsive to a givendose of muscle extract than satellite cells associatedwith live myofibers. Marcaine alone had no effect onsatellite cell proliferation, confirming earlier studies(Schultz and Lipton, 1978). The protease inhibitors hadno effect on satellite cell proliferation, but myonucleiand debris from the Marcaine-killed myofibers per-sisted for at least several days within the basal laminatube. In the absence of protease inhibitors, nucleiwithin the killed myofiber became pyknotic and dis-solved after a few hours.
To obtain additional information on the effect of theintact myofiber, I examined the dose-response ofmitogen-stimulated satellite cell proliferation on killedand living myofibers. In these experiments, a mixture ofkilled and live myofibers was obtained in the sameculture dish by killing a fraction (10-50%) of themyofibers just prior to plating. The myofibers are verysensitive to mechanical damage and variable numberscan be killed by vigorous trituration or by trituratingwith a pipet having a tip diameter less than the myofiberlength (lmm). These cultures were grown in 0-1.8 mgml"' muscle extract for 2 days and [ H]TdR was addedduring the last 4h. Labeled satellite cells were countedseparately in killed and live myofibers. The killedmyofibers were easily identified by hypercontraction oftheir myofibrils and the absence of myonuclei. Therewere substantially more labeled satellite cells associatedwith the killed myofibers over the entire range ofmuscle extract, even at concentrations in the plateauregion of the dose-response curve (Fig. 9).
Fusion of myogenic cells with myofibersInhibition of proliferation may induce myogenic cells todifferentiate (Lathrop et al. 1985; Linkhart et al. 1982).Since the mature myofibers suppress proliferation ofattached myogenic cells, I tested whether the cellswould fuse with the myofibers they contacted. Previousstudies have shown that progeny derived from satellitecells on single myofibers are fusion competent and formmyotubes by fusion among themselves when the associ-ated mature myofiber is killed (Bischoff, 1979; Bischoff,1980).
Satellite cellsThis experiment was designed to challenge labeledsatellite cells to fuse with their associated myofiber and
Fig. 4. Phase-contrastmicrograph of living myofibergrown in culture with muscleextract for 2 days aftercentrifugation. The detachedsatellite cells have migratedaway from the myofiber andare proliferating on thesubstratum. Bar, 50/mi.
Satellite cell-myofiber interaction 947
B
Fig. S. Radioautograph of satellite cells (A) centrifuged off myofiber, or (B) left on myofiber and labeled for 4h with[3H]TdR after 2 days in culture with muscle extract. All satellite cells are present on the substratum in panel A and someare heavily labeled with silver grains. The labeled satellite cell nuclei on the myofiber are shown by arrows in panel B. Bar,50fim.
Fig. 6. Phase-contrast micrograph of a myofiber shortly after exposure to 0.05% Marcaine for lOmin at 0-time. Thehypercontracted remnant of the myofiber (cross) has retracted from its basal lamina tube (arrows). A satellite cell(arrowhead) is visible on the inner surface of the basal lamina. Bar, 25 fan.
948 R. Bischoff
Fig. 7. Micrograph of satellite cells growing in the basal lamina tube of a myofiber killed by exposure to Marcaine as inFig. 6. Cultures were incubated for 2 days in medium containing muscle extract. The basal lamina is not readily visible inthis hematoxylin-stained preparation, although it can be seen after staining with anti-laminin (Bischoff, 1986a). The darklystained mass in the center is the remnant of the original myofiber. The only nuclei present are those of satellite cells; themyonuclei have dissolved. Bar, 50/xm.
MuscleExtract
BasalMedium
+ Protease Inhibitors
-—\^
—i D Killed MyofibersI D Live Myofibers
110 20
Labeled Satellite Cells per Myofiber30
Fig. 8. Effect of muscle extract on satellite cell proliferationin live and killed myofibers. Isolated myofibers were eitherleft intact or killed by a lOmin exposure to 0.05 %Marcaine in EBSS at 0-time. Cultures were incubated inbasal medium (MEM+10% serum) with or without0.5mgml~' muscle extract. Some cultures also receivedO.lmgml"1 each of aprotinin and leupeptin to inhibitproteases released from the killed myofibers. All cultureswere labeled with O.S^Ciml"1 [3H]TdR at 44 h in vitro andkilled at 48 h. Labeled satellite cells were counted inradioautographs. The error bars represent the standarderror of the mean (SEM) of counts of at least 50 myofibersfrom duplicate cultures.
1 2Muscle Extract (mgAnl)
Fig. 9. Dose-response of satellite cell proliferationstimulated by mitogen on live and killed myofibers. Aportion of the myofibers were killed by vigorous triturationduring isolation so that the cultures contained a mixture oflive and killed myofibers. The cultures were incubated inbasal medium containing various concentrations of muscleextract and labeled as described for Fig. 8. The mean±SEMis shown for counts of at least 50 myofibers from duplicatecultures.
then to detect the resulting labeled myonuclei by theirdisappearance after Marcaine treatment. Althoughsatellite cells and myonuclei can be distinguished micro-scopically in whole mounts of myofibers (Bischoff,1986a), the overlying radioautographic silver grainsmakes identification uncertain in some cases. Singlemyofibers were cultured in medium containing a lowconcentration of muscle extract (100figml~ ) plusO.ljiCimr1 [3H]TdR for 68h. Half the cultures werethen treated with 0.05 % Marcaine to kill myofiberswhile the other half was left as control. Both groupswere incubated with medium containing 0.32 ITIM unla-beled thymidine and 33/igmP1 deoxyribonuclease andsacrificed after 4h, by which time the myonuclei werelysed in the killed myofibers. If any of the satellite cells
had fused during the 3 day period, the resulting labeledmyonuclei would be eliminated by Marcaine and thecontrol myofibers would therefore contain more labelednuclei than the Marcaine-treated myofibers. Countsmade of total labeled nuclei per myofiber in radioauto-graphs revealed no significant difference between thekilled and living myofibers (killed: 16.7±0.8, live:17.5±1). Therefore, although contact with the myo-fiber inhibits proliferation of associated satellite cells, itdoes not induce them to fuse with the mature myofiberduring a 3 day period.
Embryonic myoblastsSince single myofibers undergo morphological andbiochemical changes after several days in culture
Satellite cell-myofiber interaction 949
IpssflSEgg
(Bischoff, 1980; Glavinovicef a/. 1983; Bischoff, 1986a),I wished to determine whether the fusion competenceof the myofibers for myogenic cells changes with time.The proliferating satellite cells in single myofiber cul-tures, however, begin to fuse among themselves beforesubstantial changes in the myofibers occur (Bischoff,1986a). Since this activity would complicate the assay,embryonic myoblasts were challenged to fuse withmyofibers that had been stripped of satellite cells andcultured for several days. Single myofibers were treatedwith trypsin as described (Bischoff, 1986a) to removethe basal lamina and satellite cells. The fibers wereattached to polylysine-treated coverslips by centrifu-gation and cultured with daily feeding of MEM with10% horse serum and 5% chick embryo extract.Myogenic cells were prepared from 19 day rat embryosas described and labeled overnight with O.ljiiCiml"1
[3H]TdR. Radioautographs showed that this procedureresults in the labeling of 95±1.6% of the cells. The cellswere dissociated with trypsin, counted with a hemocyt-ometer and suspended in the same medium containing0.32 mM unlabeled thymidine. Myogenic cells (3x10per 35 mm dish) were added to the single myofibercultures at 6 or 7 days in vitro. By this time, most of themyofibers had pseudopodial outgrowths from the myo-tendon ends (Fig. 10). Some myogenic cells were alsogrown in fresh polylysine-treated dishes as a control forfusion competence of the cells. The cultures were fixedat 24, 48 and 72 h after adding labeled cells andradioautographs were prepared. The labeled myogeniccells fused with each other and with the sprouting endsof the mature myofibers beginning at 24h, but did not
Fig. 10. Pseudopodial sprouts growingfrom the myotendon ends of the myofiberafter prolonged culture. Myofibers wereincubated in medium containing 10~5Mcytosine arabinoside to kill satellite cells.(A) Scanning electron micrograph of themyotendon end of a myofiber cultured for3 days. A short sprout is beginning toelongate on the substratum. Bar, 10 ̂ m.(B) Phase-contrast micrograph of livingfiber after 6 days in culture. Two sproutsarise from the end of the myofiber; theupper sprout contains several myonucleithat have migrated from the originalportion of the myofiber (at left). Bar,50/im.
fuse with the original portion of the mature myofibers atany time period tested (Fig. 11).
Discussion
Proliferation of satellite cells following muscle injury isinitiated by the release of diffusible growth factors fromthe damaged muscle (Bischoff, 1986b). In addition tothis positive stimulation, the present results indicatethat satellite cell growth is regulated by negative fac-tors, including contact with the surface of a viablemyofiber. Satellite cells on the culture substratum or onthe myofiber basal lamina both proliferated to a greaterextent than cells on intact myofibers. Although it ispossible that the substratum stimulated proliferation,this seems unlikely since satellite cells on the sub-stratum were not stimulated to proliferate withoutadded mitogen. Furthermore, cells on the intact myo-fiber were suppressed relative to those on the basallamina alone and the only difference in this case was thepresence of the myofiber plasmalemma.
A growth strategy based upon both positive andnegative elements affords a greater degree of finetuning of muscle regeneration since satellite cell re-sponse depends upon the balance between positive andnegative regulators. For example, death of a singlemyofiber, or a small group, would release its associatedsatellite cells from contact inhibition and allow the cellsto proliferate in response to the small amount of growthfactor liberated from the degenerating myofiber. At thesame time, satellite cells on neighboring viable myo-
950 R. Bischoff
•r*
m
11i A0
Fig. 11. Fusion of embryonic myoblasts with the sprouting end of a myofiber. Myogenic cells were obtained from ratembryos and labeled with [3H]TdR as described. The cells were added to isolated myofibers cultured in cytosine arabinosidefor 6 days and killed 48 h later. The pseudopodial sprout is comparable to that shown in Fig. 10B and contains a mixture oflabeled nuclei from the added cells and unlabeled nuclei from the original myofiber at the right. Bar, 10/tm.
fibers would be prevented from responding to thegrowth factor by contact inhibition and remain quiesc-ent.
The present results show that with a low dose ofmuscle extract it is possible to stimulate a moderatelevel of satellite cell proliferation on killed myofibersbut virtually none on live myofibers (Fig. 9). There isabundant evidence in the literature for local control ofregeneration. Small foci of regenerating muscle, involv-ing one or a few myofibers, are often observed follow-ing strenuous exercise (Giddings et al. 1985; Darr andSchultz, 1987; Irintchev and Wernig, 1987) and inmuscular dystrophy (Engel, 1977). Contact-dependentcommunication between satellite cells and myofibermay also explain mobilization of satellite cells upstreamor downstream from the actual site of injury (Mazanetet al. 1982; Klein-Ogus and Harris, 1983). Here, theresponse might include migration of satellite cells aswell as proliferation, but the response would be muchgreater in the axial than the transverse dimension of themuscle. Unfortunately, the reaction of satellite cellsdistant from a wound has not been investigated quanti-tatively. Finally, contact inhibition may be important inmuscle development as a mechanism for coordinatingmitotic quiescence of satellite cells. As the myofiberreaches a certain level of maturity, changes in theplasmalemma might signal attached satellite cells towithdraw from the cell cycle. It would be of interest tomeasure the degree of contact inhibition as a function ofmyofiber age or maturity.
Contact inhibition is not absolute and can be over-come by increasing the concentration of muscle extractin vitro. Similarly, in certain situations in vivo it may beadvantageous for satellite cells to proliferate on viablemyofibers. There are examples of this during compen-satory hypertrophy (Hanzlfkova' et al. 1975; Schiaffinoet al. 1976), ischemia (Jennische, 1986) and toxin-stimulated regeneration (Maltin et al. 1983). Some ofthe satellite cell progeny may fuse with the myofibers(Schiaffino et al. 1976). It is not known whether these
cases represent increased release of growth factors orreduced contact inhibition by the myofiber, or both.
The muscle basal lamina survives various types oftissue injury and is believed to have a beneficial effecton regeneration (Le Gros Clark, 1946; Vracko andBenditt, 1972; Carlson and Faulkner, 1983), but its rolehas not been systematically investigated. Unfortu-nately, in the present experiments it was not possible todirectly test the effect of the basal lamina on satellitecell proliferation. The basal lamina can be removedfrom single isolated myofibers with a low concentrationof trypsin or Pronase (Bischoff, 1986a), but the satellitecells invariably become detached from the myofibersand thus their response to mitogen cannot be tested onfibers with and without a basal lamina covering. In theMarcaine experiments, the satellite cells were in contactwith basal lamina in both the live and killed myofibers,yet were more responsive to mitogen in the absence of alive myofiber. Thus it seems that if basal laminacomponents, such as laminin, stimulate proliferation ofmyogenic cells, as has been shown in previous studiesusing monqlayer cultures (Podleski et al. 1979; Foster etal. 1987; Ocalan et al. 1988), the effect can be sup-pressed by contact with the viable myofiber.
Inhibiting the proliferation of attached satellite cellsby contact with a myofiber does not increase thelikelihood that the cells will fuse with the myofiber. Infact, there was no instance of fusion of either satellitecells or embryonic myoblasts with the surface of amature myofiber. The enzymatic treatment used todisaggregate tissue into single myofibers may havedamaged surface receptors for fusion. In the case ofembryonic myoblasts, however, the single fibers hadbeen in culture for up to 7 days, allowing ample time forrecovery. These results, together with earlier work,suggests that the plasma membrane becomes refractoryto fusion as part of the terminal differentiation ofmyofibers. Bischoff and Holtzer (1969) found thatfusion-competent myogenic cells failed to fuse withwell-differentiated myotubes in monolayer cultures and
Satellite cell-myofiber interaction 951
speculated that the block to fusion might reside in thebasal lamina or extracellular matrix. This possibility canbe ruled out in the present study. The proliferatingsatellite cells on single myofiber are beneath the basallamina, yet do not fuse. In the experiments withembryonic myoblasts, all extracellular coats wereremoved prior to challenging the myofibers with fusion-competent cells. Trypsin-treated myofibers lack a basallamina and the openings of the transverse tubules areclearly visible with the scanning electron microscope(Bischoff, 1986a).
A fusion block might function to regulate the nucleardensity of myofibers during myogenesis. Most mononu-cleated myogenic cells adhere to the surface of myo-tubes during development in vivo (Tello, 1922) and invitro (Bischoff and Lowe, 1974). Unfused myogeniccells on the surface of a myotube that became closed tofurther fusion would be encouraged to fuse amongthemselves and thus initiate the formation of a newgeneration of myotubes. The fusion block may also beinvolved in the formation of Go satellite cells byinducing myogenic cells to withdraw from the cell cycle,perhaps as the concentration of growth factors becomeslimiting during the late stages of development. Accord-ing to this scheme, quiescent satellite cells could existon mature myofibers at the same time as dividingsatellite cells on young myofibers nearby. Satellite cellproliferation would.be regulated both by concentrationof growth factors and by the surface properties of themyofiber. Further studies are needed to investigate thefusion properties of myofibers. Lipton and Schultz(1979) reported that myogenic cells injected into adultmuscle can fuse with mature myofibers. Fusion of theinjected cells among themselves was noted by 4 daysand many of the injected cells had migrated to aposition beneath the basal lamina of the myofibers bythis time, but the cells did not fuse with maturemyofibers until 7 days after transplantation. There is noexplanation for the delay in fusion. Since there wasmyofiber degeneration and regeneration at the injectionsite, it is possible that the implanted cells first fused herewith the ends of injured myofibers and the myonucleirequired several days to migrate into undamaged re-gions of the muscle.
Although myogenic cells did not fuse with maturemyofibers in the present experiments, they readilyfused with nucleated sprouts growing from the ends ofthe mature myofibers. The sprouts consist of pseudopo-dial extensions that begin to appear after 4 or 5 days inculture. Eventually the entire myofiber is transformedinto an elongate, sometimes branched, myotube-likestructure (Bischoff, 1980; Bischoff, 1986fl). Althoughmyonuclei migrate from the original myofiber into thesprouts, it seems unlikely that labeled cells first fusedwith the original myofiber then their nuclei migratedcentrifugally, for labeled nuclei were never found in theoriginal myofiber.
Sprouting has also been observed in injured muscle invivo, where it is believed to contribute to a processtermed continuous regeneration. Immature myotubeswith clustered nuclei and sparse myofibrils are often
found near the wound edge several days after injury, inapparent continuity with surviving mature myofibers(Le Gros Clark, 1946; Hall-Craggs, 1974). The natureof these myotubes is difficult to determine in lightmicroscope studies, but electron microscopic examin-ation clearly shows cytoplasmic continuity with maturemyofibers (Shafiq and Gorycki, 1965). The sprouts mayarise from myofibers that were transected by injury andsubsequently healed over (Echeverria et al. 1987). Thisprocess offers several advantages to muscle regener-ation. By elongating across the wound area, the sproutsbridge the gap to join myofibers from the opposite edge(Gay and Hunt, 1954; Hudgson and Field, 1973; Webb,1973) The growth rate of the sprouts is rapid enough toestablish electrical continuity before satellite-cell-derived myotubes have had much opportunity to form(Stuart etal. 1981). Sprouting involves dedifferentiationof the myofibers, a process that includes the reappear-ance of embryonic isozymes of creatine kinase (Bisch-off, 1986a). Another aspect of dedifferentiation, asdemonstrated in the present study, is the reacquisitionof a surface membrane compatible with fusion. Thiswould allow transected myofibers to restore their orig-inal nuclear complement by fusion with satellite-cell-derived myoblasts, or alternatively to fuse with satel-lite-cell-derived myotubes or with sprouting myofibersfrom the opposite wound edge.
I thank Kym Hallsworth for competent assistance withthese experiments.
This research was supported by a grant from the NationalInstitutes of Health (AM36853).
References
BADER, C. R., BERTRAND, D., COOPER, E. AND MAURO, A. (1988).
Membrane currents of rat satellite cells attached to intact skeletalmuscle fibers. Neuron 1, 237-240.
BISCHOFF, R. (1975). Regeneration of single skeletal muscle fibersin vitro. Anot. Rec. 182, 215-236.
BISCHOFF, R. (1979). Tissue culture studies on the origin ofmyogenic cells during muscle regeneration in the rat. In MuscleRegeneration (ed. A. Mauro), pp. 13-29. New York: RavenPress.
BISCHOFF, R. (1980). Plasticity of the myofiber-satellite cellcomplex in culture. In Plasticity of Muscle (ed. D. Pette) pp.119-129. Berlin: De Gruyter and Co.
BISCHOFF, R. (1986a). Proliferation of muscle satellite cells onintact myofibers in culture. Devi Biol. 115, 129-139.
BISCHOFF, R. (1986b). A satellite cell mitogen from crushed adultmuscle. Devi Biol. 115, 140-147.
BISCHOFF, R. AND HOLTZER, H. (1969). Mitosis and the processes ofdifferentiation of myogenic cells in vitro. J. Cell Biol. 41,188-200.
BISCHOFF, R. AND LOWE, M. (1974). Cell surface components andthe interaction of myogenic cells. In Exploratory Concepts inMuscular Dystrophy 11. (ed. A. T. Milhorat) pp. 17-29.Amsterdam: Excerpta Medica.
CARDASIS, C. A. AND COOPER, G. W. (1975). An analysis of nuclearnumbers in individual muscle fibers during differentiation andgrowth: A satellite cell-muscle fiber growth unit. J. exp. Zool.191, 347-358.
CARLSON, B. M. AND FAULKNER, J. A. (1983). The regeneration ofskeletal muscle fibers following injury, a review. Med. Sci. SportsExercise 15, 187-198.
D'ALBIS, A., WEINMAN, J., MIRA, J - C , JANMOT, C. AND
952 R. Bischoff
COUTEAUX, R. (1987). Role regulateur des hormonesthyroidiennes dans la myogenese. Analyse des isoformes de lamyosine dans la regeneration musculaire. C.r. hebd. Stanc Acad.Sci. Paris HI 305, 697-702.
DARR, K. C. AND SCHULTZ, E. (1987). Exercise-induced satellitecell activation in growing and mature skeletal muscle. J. Appl.Physiol. 63, 1816-21.
ECHEVERKIA, O. M., NlNOMIYA, J. G. AND VAZQUEZ-NlN, G. H.
(1987). Microscopical and electrophysiological studies on thehealing-over of striated fibers of cremaster muscle of the guineapig. Ada Anal. 128, 274-280.
ELSDALE, T. AND BARD, J. (1972). Collagen substrate for the studyof cell behavior. J. Cell Biol. 41, 298-311.
ENGEL, W. K. (1977). Integrative histochemical approach to thedefect of Duchenne muscular dystrophy. In Pathogenesis ofHuman Muscular Dystrophies (ed. L. P. Rowland), pp. 277-309.Amsterdam: Excerpta Medica.
FOSTER, R. F., THOMPSON, J. M. AND KAUFMAN, S. J. (1987). A
laminin substrate promotes myogenesis in rat skeletal musclecultures, analysis of replication and development usingantidesmin and anti-BrdUrd monoclonal antibodies. Devi Biol.122, 11-20.
GAY, A. J. AND HUNT, T. E. (1954). Reuniting of skeletal musclefibers after transection. Anal. Rec. 120, 853-871.
GIDDINGS, C. J., NEAVES, W. B. AND GONYEA, W. J. (1985).
Muscle fiber necrosis and regeneration induced by prolongedweight-lifting exercise in the cat. Anal. Rec. 211, 133-141.
GLAVINOVIC, M. I., MILEDI, R. AND NAKAJIMA, Y. (1983).
Morphological and physiological changes in dissociated adult frogmuscle fibers after prolonged culturing. Proc. Roy. Soc. Lond. B219, 91-101.
HALL-CRAGGS, E. C. B. (1974). The regeneration of skeletal musclefibers per continuum. J. Anat. 117, 171-178.
HANSEN-SMITH, F. M. AND CARLSON, B. M. (1979). Cellularresponses to free grafting of the extensor digitorum longusmuscle of the rat. J. Neurol. Sci. 41, 149-173.
HANZLJKOVA, V., MACKOVA, E. V. AND HNfc, P. (1975). Satellitecells of the rat soleus muscle in the process of compensatoryhypertrophy combined with denervation. Cell Tissue Res. 160,411-421.
HUDGSON, P. AND FIELD, E. J. (1973). Regeneration of muscle. InStructure and Function of Muscle. Vol 11, 2nd ed. (ed. G.Bourne) pp. 312-363. New York: Academic Press.
IRINTCHEV, A. AND WERNIG, A. (1987). Muscle damage and repairin voluntarily running mice, strain and muscle differences. CellTissue Res. 249,509-521.
ISHIKAWA, H. (1966). Electron microscopic observations of satellitecells with special reference to the development of mammalianskeletal muscles. Z. Anat. Entwicklungsgesch. 125, 43-63.
JENNISCHE, E. (1986). Rapid regeneration in postischaemic skeletalmuscle with undisturbed microcirculation. Acta Physiol. Scand.128, 409-414.
JIRMANOVA, I. AND THESLEFF, S. (1972). Ultrastructural study ofexperimental muscle degeneration and regeneration in the adultrat. Z. Zellforsch. Mikrosk. Anat. 131, 77-97.
KELLY, A. M. (1978). Satellite cells and myofiber growth in the ratsoleus and extensor digitorum longus muscles. Devi Biol. 65,1-10.
KELLY, A. M. AND ZACKS, S. (1969). The histogenesis of ratintercostal muscle. J. Cell Biol. 42, 135-153.
KLEIN-OGUS, C. AND HARRIS, J. B. (1983). Preliminaryobservations of satellite cells in undamaged fibres of the ratsoleus muscle assaulted by a snake-venom toxin. Cell Tissue Res.230, 671-676.
KONIGSBERG, I. R. (1979). Skeletal myoblasts in culture. Meth.Enzymol. 58, 511-527.
KONIGSBERG, U. R., LIPTON, B. H. AND KONIGSBERG, I. R. (1975).
The regenerated response of single mature muscle fiber isolatedin vitro. Devi Biol. 45, 260-275.
LATHROP, B., OLSON, E. AND GLASER, L. (1985). Control by
fibroblast growth factor of differentiation in the BC3H1 musclecell line. J. Cell Biol. 100, 1540-1547.
LE GROS CLARK, W. E. (1946). An experimental study of theregeneration of mammalian striped muscle. J. Anat. 80, 24-36.
LINKHART, T. A., CLEGG, C. A. AND HAUSCHKA, S. D. (1982).
Myogenic differentiation in permanent clonal mouse cell lines,Regulation by macromolecular growth factors in the culturemedium. Devi Biol. 86, 19-30.
LIPTON, B. H. AND SCHULTZ, E. (1979). Developmental fate ofskeletal muscle satellite cells. Science 205, 1292-1294.
MALTIN, C. A., HARRIS, J. B. AND CULLEN, M. J. (1983).
Regeneration of mammalian skeletal muscle following theinjection of the snake-venom toxin, taipoxin. Cell Tissue Res.232, 565-77.
MAZANET, R., REESE, B. F., FRANZINI-ARMSTRONG, C. AND REESE,
T. S. (1982). Variability in the shapes of satellite cells in normaland injured frog sartorius muscle. Devi Biol. 93, 22-27.
MCGEACHIE, J. K. AND GROUNDS, M. D. (1987). Initiation andduration of muscle precursor replication after mild and severeinjury to skeletal muscle of mice. An autoradiographic study.Cell Tissue Res. 248, 125-130.
MULVANEY, D. R., MARPLE, D. N. AND MERKEL, R. A. (1988).
Proliferation of skeletal-muscle satellite cells after castration andadministration of testosterone propionate. Proc. Soc. exp. Biol.med. 188, 40-45.
OCALAN, M., GOODMAN, S. L., KOHL, U., HAUSCHKA, S. D. AND
VON DER MARK, K. (1988). Laminin alters cell shape andstimulates motility and proliferation of murine skeletalmyoblasts. Devi Biol. 125, 158-167.
PHILLIPS, G. D., LU, D. AND CARLSON, B. M. (1987). Survival ofmyogenic cells in freely grafted rectus femoris and extensordigitorum longus muscles. Amer. J. Anat. 180, 365-372.
PODLESKl, T . R . , GREENBERG, I . , SCHLESS1NGER, J. AND Y A M A D A ,
K. M. (1979). Fibronectin delays the fusion of L6 myoblasts.Expl Cell Res. 122, 317-326.
Ross, J. J., DUXSON, M. J. AND HARRIS, A. J. (1987). Formation ofprimary and secondary myotubes in rat lumbrical muscles.Development 100, 383-394.
SANES, J. R., MARSHALL, L. M. AND MCMAHAN, U. J. (1978).
Reinnervation of muscle fiber basal lamina after removal ofmyofibers, Differentiation of regenerating axons at the originalsynaptic site. J. Cell Biol. 78, 176-198.
SCHIAFFINO, S., PIEROBON BORMIOU, S. AND ALOISI, M. (1976). The
fate of newly formed satellite cells during compensatory musclehypertrophy. Virchows Arch. B Cell Path. 21, 113-118.
SCHULTZ, E. (1976). Fine structure of satellite cells in growingskeletal muscle. Am. J. Anat. 147, 49-70.
SCHULTZ, E. AND LIPTON, B. H. (1978). The effect of Marcaine onmuscle and non-muscle cells in vitro. Anal. Rec. 191, 351-370.
SHAFIQ, S. A. AND GORYCKI, M. A. (1965). Regeneration in skeletalmuscle of mouse, some electron-microscope observations. J.Pathol. Bacteriol. 90, 123-127.
SNOW, M. H. (1977). Myogenic cell formation in regenerating ratskeletal muscle injured by mincing. II. An autoradiographicstudy. Anat. Rec. 188, 210-218.
STUART, A., MCCOMAS, A. J., GOLDSPINK, G. AND ELDER, G.
(1981). Electrophysiological features of muscle regeneration.Expl Neurol. 74, 148-159.
TELLO, J. F. (1922). Die Entstehung der motorischen undsensibelen Nervendungen. I. Z. Anat. Ennvicklungsgesch. 64,348-.
VRACKO, R. AND BENDITT, E. P. (1972). Basal lamina, the scaffoldfor orderly cell replacement. Observations on regeneration ofinjured skeletal muscle fibers and capillaries. J. Cell Biol. 55,406-419.
WEBB, P. (1973). The effect of innervation, denervation, andmuscle type on the reunion of skeletal muscle. Brit. J. Surg. 60,180-182.
(Accepted November 1989)(Received in Cambridge 30 April 1990)