Interaction of Gibberellic Acid and Indole-3-acetic Acid in theGrowth of Excised Cuscuta Shoot Tips in Vitro'
Received for publication February 5, 1979 and in revised form August 23, 1979
RAMESH MAHESHWARI2, C. SHAILINI, K. VELUTHAMBI, AND S. MAHADEVANDepartment of Biochemistry, Indian Institute of Science, Bangalore-560 012, India
ABSTRACT
Gibberellic acid (GA3) induced a marked elongation of 2.5-centimetershoot tips of Cuscuta chinensis Lamk. cultured in vitro. In terms of theabsolute amount of elongation, this growth may be the largest reported foran isolated plant system. The response to hormone was dependent on anexogenous carbohydrate supply. The hormone-stimulated growth was dueto both cell division and cell elougation. The growth response progressivelydecreased if GA3 was given at increasingly later times after culturing, butthe decreased growth response could be restored by the application ofindole-3-acetic acid (IAA) to the apex. Explants deprived of GA3 graduallylost their ability to transport IAA basipetally, but this ability was alsorestored by auxin application. The observations are explained on the basisthat: (a) the growth of Cuscuta shoot tip in vitro requires, at least, both anauxin and a gibberellin; and (b) in the absence of gibberellin the culturedshoot tip explants lose the ability to produce and/or transport auxin.
In our efforts to determine the chemical requirements for thegrowth and maintenance of the excised shoot tip of the angio-spermic parasite, Cuscuta, as an indeterminate meristem in in vitroculture, it was observed that GA3 had a marked promotive effecton its growth. The increased elongation in the presence of hor-mone was at times 20-fold greater than the control growth (-GA3)in about 6 to 8 weeks. The magnitude of this response suggestedthat the excised shoot tip of Cuscuta may be a promising materialfor studying the control of cell division and elongation by gibber-ellin. Such studies with other angiospermic plants have beendifficult because of the tendency of the explants to root in cultureand thus revert to the whole plant status. In Cuscuta, this problemwas eliminated because it lacks roots. The isolated Cuscuta shoottips afforded the further advantage that they remained alive inmineral salts solution for quite a long period (1, 10). This permittedinvestigation of the temporal sequence of interactions of growthhormones in regulating shoot elongation and lateral bud devel-opment. This plant vegetatively propagated itself as a stem para-site, and thus the experimental material was abundantly available.The lack of appendages (true leaves) made it easy to collect andsurface-sterilize a large number of shoot tips for experimentation.In this paper we describe the in vitro response of Cuscuta shoottips to GA3 and its interaction with IAA.
MATERIALS AND METHODS
Plant. The experimental material was from Cuscuta chinensisLamk. (dodder) vines parasitizing Tecoma stans (Bignoniaceae)
'This work was supported by a research grant from the UniversityGrants Commission, New Delhi to S. M. and R. M.'To whom correspondence should be addressed.
on the campus of the Indian Institute of Science. Shoot tipsapproximately 8.0 cm long were cut and brought to the laboratoryand placed under artificial light for 16 h at 25 ± 2 C with the cutends dipped in water. During this period, the segments increasedin length by 20 to 30%o and turned visibly green. Shoot tips frommaterial treated this way responded more uniformly. However,this preculture treatment was not essential for the GA3 response.
Culture Methods. Prior to culturing, the shoot tips were reducedin length to 3 to 4 cm and surface-sterilized with successivetreatments of 0.5% Polysan3 (1-2 min), 10%1o sodium hypochlorite(5 min), and 0.1% mercuric chloride (5 min). The segments wererinsed with three changes of sterile distilled H20 between treat-ments and finally with several more changes of distilled H20. Thisprocedure of surface-sterilization kept contamination below 5 to10%1o. The segments were further reduced in length and shoot tips2.5 cm long were placed upright, singly per tube containing aliquid medium.The composition ofthe BM4 used was as follows: macronutrients
(mg/i): 360 MgSO4.7H20, 260 Ca(NO3)2 .4H20, 200 Na2SO4, 200NaH2PO4, 80 KNO3, 65 KCI, 37.3 Na2EDTA, 27.8 FeSOV7H20; micronutrients (mg/i): 3 MnSO4.4H20, 0.5 ZnSO4-7H20, 0.5 H3B03, 0.026 CuSO4.5H20, 0.025 Na2MoO7.2H20,0.025 CoCl2; amino acid (mg/i): 2 glycine; vitamins: (mg/i): 0.5thiamine hydrochloride, 0.5 nicotinic acid, 0.5 pyridoxine hydro-chloride, 0.5 calcium panthothenate, 100 myo-inositol; carbonsource: 5% sucrose (w/v). The pH of the medium was adjusted to5.5 before autoclaving. Hormones were filter-sterilized using Mil-lipore (0.25 ,um) filters and aseptically added to autoclaved BM.In experiments wherein the dose-response of a chemical was'studied, the medium was also sterilized by filtration.
Cultures were initiated in sterile tubes measuring 10 x 1 cm or12.5 x 1.2 cm (i.d.) containing 0.5 or I ml of medium per tube, sothat only 0.5 to 1.0 cm of the base of the explant was immersed insolution. Sterile medium was replenished periodically and when-ever necessary the growing shoot tip was transferred into largertubes (20.5 x 2.5 cm), containing 10 ml medium per tube. Cultureswere maintained at 25 ± 2 C with a 16-h light and 8-h darkregimen. Illumination was provided from fluorescent lamps (cool-white, 40 w) placed about 50 cm from the cultures which yieldeda light intensity of about 150 lux at the level of the cultures.Growth measurement was by increase in length. Measurement
ofexplant in vitro was made by overlapping a thread and recordingthe contour length. This method was accurate within 5%. Data arebased on a minimum of 20 to 25 replicates.
Nitrogen estimation was by the method of Kjeldahl. The tissuewas digested with salicylic-sulfuric acid mixture to ensure conver-sion of all nitrate-nitrogen to ammonia (3).
3An iodophor germicide-detergent containing 1.6% iodine (w/v) andalkyl phenoxypolyethylene ethanol, a product of Polypharm Private Ltd.,Bombay, India.
I3HjThymidine Incorporation into DNA. Shoot tips were cul-tured in upright position in tapered tubes containing 0.2 ml ofmedium and 4 ,liCi of [3H]thymidine (Bhabha Atomic ResearchCentre, Trombay, Bombay, India, 12.6 Ci/mmol). Medium wasreplenished when necessary.
After the experiment, the shoot tips were washed with waterand homogenized with 5% cold trichloroacetic acid (w/v) in achilled mortar. The homogenate was filtered through a glass fiberWhatman filter using suction. The sample was washed successivelywith 20 mm unlabeled thymidine in cold 5% trichloroacetic acid,95% ethanol, ethanol-chloroform-ether (2:1:2, w/v/v) and finallywith ether. The filter circle containing the sample was heated in10%o HC104 for 30 min in a boiling water bath, cooled andcentrifuged, and an aliquot of the supernatant was counted in ascintillation fluid containing 0.2% PPO and 0.002% POPOP inmeth,anol-toluene (1: 1).
Transport of Auxin. Basipetal transport of radioactive IAA instem segments cut from shoot tips was measured using the donorblock-tissue-receiver set-up. Carboxyl-[14CJIAA with a specificradioactivity of 57 mCi/mmol (Radiochemical Centre, Amer-sham, England) was used. Donor blocks were prepared containingabout 10 pM IAA (around 18,000 cpm) in 1.5% agar in I mmpotassium citrate buffer (pH 4.9). Transport was for 3 h in ahumid chamber. The radioactivities in the receiver and in thetissue were separately counted in a scintillation fluid containing500 mg PPO and 25 mg POPOP in 100 ml toluene-dry ethanol (3:1).Inorganic chemicals and sucrose were of analytical reagent
grade. Growth hormones and other biochemicals were purchasedfrom Sigma Chemical Co. Lanolin (Mallinckrodt) was a gift ofDr. Larry D. Nooden, University of Michigan.
RESULTS
Nature of Shoot Tip Growth in the Absence and in the Contin-uous Presence of GA3. In the absence of GA3, shoot tips elongated
EU
I~-
0
to 4 to 4.5 cm in about 2 weeks and thereafter no appreciablegrowth change was seen till about 3 to 6 weeks when the basalbud in several of these tips started elongation growth. Onceinitiated, the growth of this lateral shoot was quite rapid.
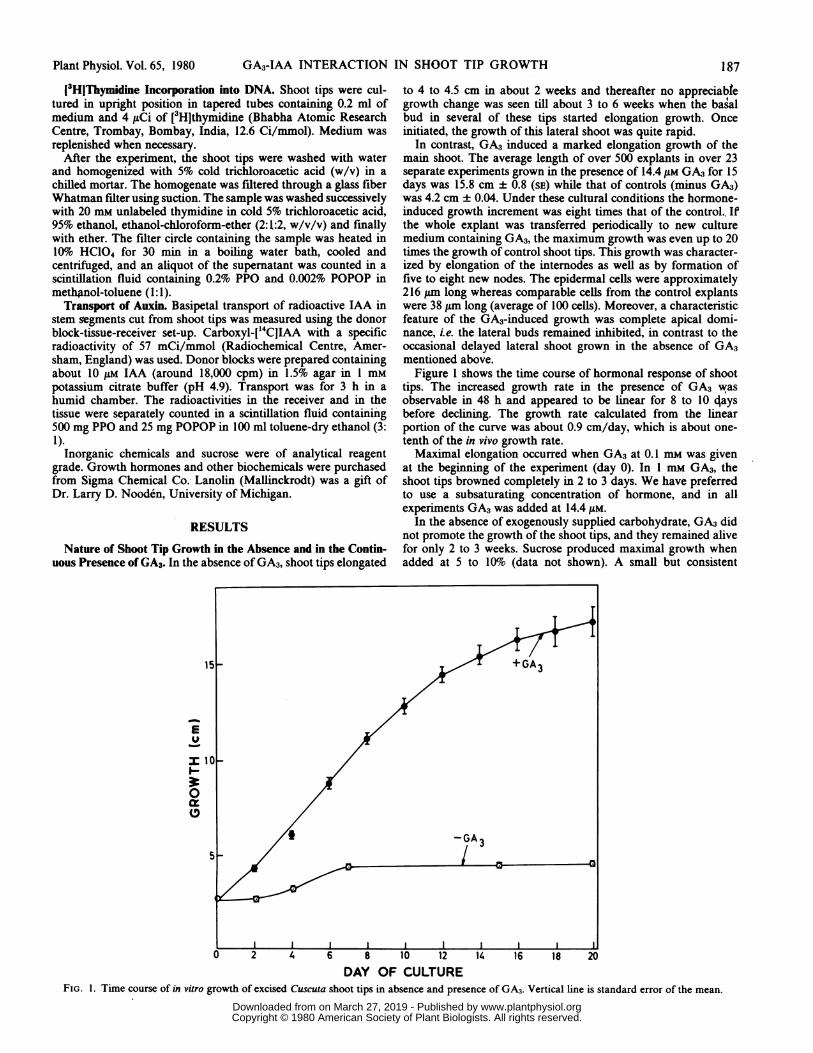
In contrast, GA3 induced a marked elongation growth of themain shoot. The average length of over 500 explants in over 23separate experiments grown in the presence of 14.4 ,UM GA3 for 15days was 15.8 cm ± 0.8 (SE) while that of controls (minus GA3)was 4.2 cm ± 0.04. Under these cultural conditions the hormone-induced growth increment was eight times that of the control.. Ifthe whole explant was transferred periodically to new culturemedium containing GA3, the maximum growth was even up to 20times the growth of control shoot tips. This growth was character-ized by elongation of the internodes as well as by formation offive to eight new nodes. The epidermal cells were approximately216 p,m long whereas comparable cells from the control explantswere 38 ,um long (average of 100 cells). Moreover, a characteristicfeature of the GA3-induced growth was complete apical domi-nance, i.e. the lateral buds remained inhibited, in contrast to theoccasional delayed lateral shoot grown in the absence of GA3mentioned above.
Figure 1 shows the time course of hormonal response of shoottips. The increased growth rate in the presence of GA3 wasobservable in 48 h and appeared to be linear for 8 to 10 4aysbefore declining. The growth rate calculated from the linearportion of the curve was about 0.9 cm/day, which is about one-tenth of the in vivo growth rate.Maximal elongation occurred when GA3 at 0.1 mm was given
at the beginning of the experiment (day 0). In I mm GA3, theshoot tips browned completely in 2 to 3 days. We have preferredto use a subsaturating concentration of hormone, and in allexperiments GA3 was added at 14.4 ,UM.
In the absence of exogenously supplied carbohydrate, GA3 didnot promote the growth of the shoot tips, and they remained alivefor only 2 to 3 weeks. Sucrose produced maximal growth whenadded at 5 to 10%o (data not shown). A small but consistent
DAY OF CULTUREFIG. 1. Time course of in vitro growth of excised Cuscuta shoot tips in absence and presence of GA3. Vertical line is standard error of the mean.
improvement in growth was observed with 100 mg/l of myo-inositol (data not shown).During the first 15 days of culture, there was no significant
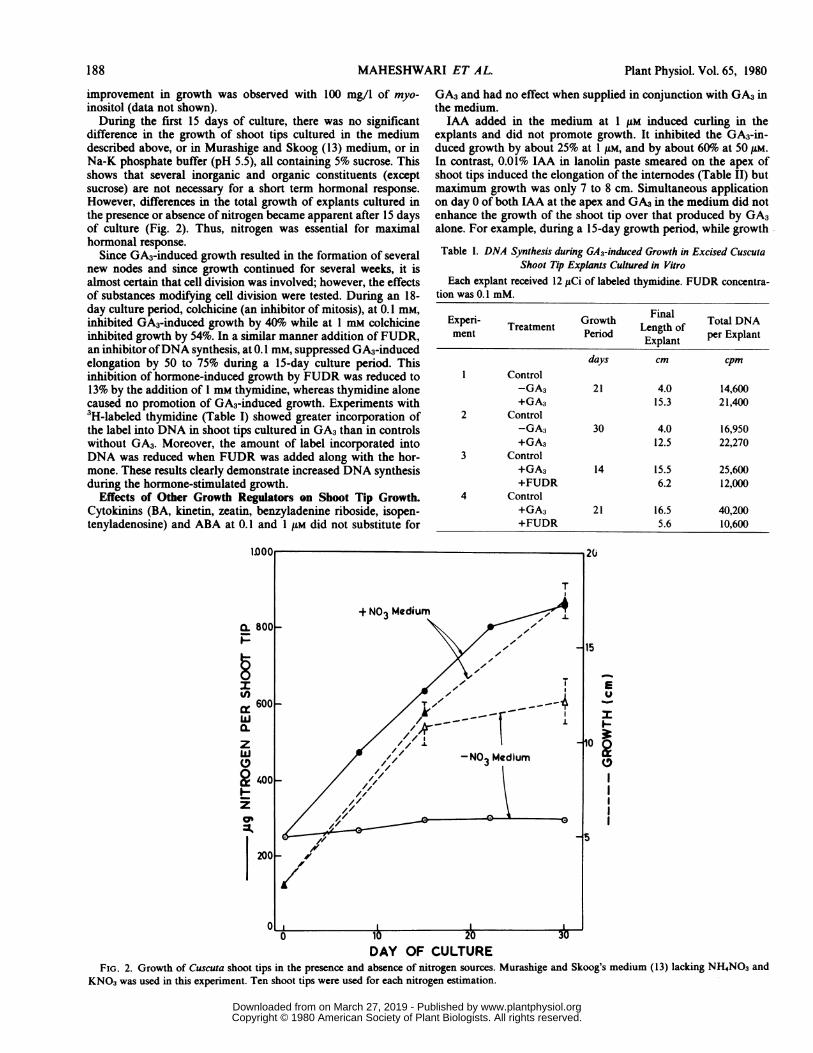
difference in the growth of shoot tips cultured in the mediumdescribed above, or in Murashige and Skoog (13) medium, or inNa-K phosphate buffer (pH 5.5), all containing 5% sucrose. Thisshows that several inorganic and organic constituents (exceptsucrose) are not necessary for a short term hormonal response.However, differences in the total growth of explants cultured inthe presence or absence of nitrogen became apparent after 15 daysof culture (Fig. 2). Thus, nitrogen was essential for maximalhormonal response.
Since GA3-induced growth resulted in the formation of severalnew nodes and since growth continued for several weeks, it isalmost certain that cell division was involved; however, the effectsof substances modifying cell division were tested. During an 18-day culture period, colchicine (an inhibitor of mitosis), at 0.1 mm,inhibited GA3-induced growth by 40%o while at I mm colchicineinhibited growth by 54%. In a similar manner addition of FUDR,an inhibitor ofDNA synthesis, at 0.1 nmm, suppressed GA3-inducedelongation by 50 to 75% during a 15-day culture period. Thisinhibition of hormone-induced growth by FUDR was reduced to13% by the addition of 1 mm thymidine, whereas thymidine alonecaused no promotion of GA3-induced growth. Experiments with3H-labeled thymidine (Table I) showed greater incorporation ofthe label into DNA in shoot tips cultured in GA3 than in controlswithout GA3. Moreover, the amount of label incorporated intoDNA was reduced when FUDR was added along with the hor-mone. These results clearly demonstrate increased DNA synthesisduring the hormone-stimulated growth.
Effects of Other Growth Regulators on Shoot Tip Growth.Cytokinins (BA, kinetin, zeatin, benzyladenine riboside, isopen-tenyladenosine) and ABA at 0.1 and 1 ,UM did not substitute for
GA3 and had no effect when supplied in conjunction with GA3 inthe medium.IAA added in the medium at I ,iM induced curling in the
explants and did not promote growth. It inhibited the GA3-in-duced growth by about 25% at 1 ,UM, and by about 60o at 50 UM.In contrast, 0.01% IAA in lanolin paste smeared on the apex ofshoot tips induced the elongation of the internodes (Table II) butmaximum growth was only 7 to 8 cm. Simultaneous applicationon day 0 of both IAA at the apex and GA3 in the medium did notenhance the growth of the shoot tip over that produced by GA3alone. For example, during a 15-day growth period, while growth
Table I. DNA Synthesis during GA3-induced Growth in Excised CuscutaShoot Tip Explants Cultured in Vitro
Each explant received 12 ,tCi of labeled thymidine. FUDR concentra-tion was 0.1 mM.
Experi- Growth Final Total DNAment PerioTreatmentd Length of
Explant per Explant
days cm cpmI Control
-GA3 21 4.0 14,600+GA3 15.3 21,400
2 Control-GA3 30 4.0 16,950+GA3 12.5 22,270
3 Control+GA3 14 15.5 25,600+FUDR 6.2 12,000
4 Control+GA3 21 16.5 40,200+FUDR 5.6 10,600
WuUr 726
a-In
w0lz
w
1uz
8oo0
6001-
4001-
2001
0 10 20
DAY OF CULTURE"U
15
Eu
Iio-IO:
5
10 flll
FIG. 2. Growth of Cuscuta shoot tips in the presence and absence of nitrogen sources. Murashige and Skoog's medium (13) lacking NH4NO3 andKNO3 was used in this experiment. Ten shoot tips were used for each nitrogen estimation.
in presence of GA3 was 13.06 cm ± 0.46 (SE), that in the presenceof GA3 + 0.01% IAA (applied at the apex) was 12.57 cm ± 0.58(SE). There was also no increase in growth when GA3 was appliedon day 8 to explants treated with IAA earlier (Table II).
Effect of GA3 and IAA on Growth of Decapitated Shoot Tips.Decapitation of shoot tips (removal of about 0.5 cm from apex)reduced the growth response to GA3, presumably by the removalof tissue capable of cell division and cell elongation. Growth insuch explants was about two times that of controls (Table II).As in intact shoot tips, IAA had no significant effect when
Table II. Effect of GA3 and IAA on Growth of Intact and DecapitatedShoot Tip Explants of Cuscuta
Initial length of explants was 2.5 cm. GA3 was added in the mediumand IAA in lanolin was smeared on the apex.
TreatmentFinal Explant
Lengthcm ± SE
A. Apex intact (growth period 25 days)(a) Control 4.1 0.1(b) +GA3 18.0 ± 0.9(c) +GA3 (applied on day 8) 10.7 ± 0.9(d) +IAA (applied on day 0) 8.0 ± 0.5(e) +IAA (applied on day 0) 7.9 ± 0.4
+GA3 (applied on day 8)B. Decapitated (growth period 30 days)
(f) Control 4.2 ± 0.1(g) +GA38 5.7 ± 0.3(h) +IAA 6.7 ± 0.3(i) +IAA + GA3 7.8 ± 0.3a In these explants, the uppermost lateral bud grew out. Significant
difference at 1% level between treatments (c) and (e), (f) and (g), and (g)and (i). Significant difference at 5% level between treatments (g) and (h),and (h) and (i).
provided in the medium. However, when IAA was applied at thecut apex, it induced a growth increment about 2.5 times that ofthe control. The growth of decapitated explants was significantlygreater in presence of IAA plus GA3 than when either one wasapplied alone but the growth was much less than that of the intacttips.
Discontinuous Application of GA3. To determine whether GA3acts as a trigger or is continuously required for growth, shoot tipswere placed upright in presence ofGA3 for different time intervals,rinsed in sterile water, and then allowed to grow in the BM. Thedata in Figure 3 show that GA3 must be supplied for the first 6days of culture in order to produce maximum growth of the shoottips.
In pulse experiments, GA3 of different concentrations wasapplied for 1 h. With shorter hormone application time, a higherGA3 concentration was required to produce a given maximalgrowth. One-h pulse applications of up to 50 ,LM GA3 wereineffective in promoting growth (data not shown) while maximalgrowth was obtained with 1 mm GA3. This concentration ofhormone was, however, toxic if given for 6 h, the explants brown-ing within 2 days after hormone application. These results suggestthat the elongation response depends upon the amount ofhormonetaken up by the explant.Delayed Application of GA3. Maximal elongation of shoot tips
occurred when GA3 was given at the beginning of the cultureperiod. The decline in growth with progressively delayed appli-cation of GA3 appeared to be more or less linear with time (Fig.4). It was also variable from experiment to experiment and thisappeared to depend on the season when the Cuscuta vines wereobtained. When GA3 was given 1 to 3 weeks after the beginningof culture, the growth response of shoot tips to hormone applica-tion was either small or totally absent. In many such explants, thebasal bud activated by this time (i.e. protruding through the scale),grew into a shoot. GA3 application did not hasten the growth rateor augment the maximum growth of the lateral shoot. As alreadymentioned, lateral shoot growth also occurred in several control
DAY OF CULTUREFIG. 3. Response of excised Cuscuta shoot tips to discontinuous application of GA3.
DAYS ELAPSED BEFORE APPLICATION OF GA3FIG. 4. Response of excised Cuscuta shoot tips to delayed application of GA3. Shoot tips were cultured in BM for different periods before transferring
to BM + GA3. Measurements made 20 days after hormone treatment. Vertical lines are standard error. Experiment 1 was done in January; experiment2 was done in April.
shoot tips cultured in the absence of the hormone after 3 to 6weeks.The loss of apical dominance suggested the possibility that
either the production of auxin at the apex and/or its basipetaltransport might have been affected. Ifthe first possibility is correct,then application of auxin to the apex of such shoot tips mightpromote growth. When IAA was smeared on the apex of shoottips incubated 8 to 21 days in a medium lacking GA3, growth didtake place in response to this delayed GA3 application (Table III).Delayed IAA application in the absence ofGA3 was ineffective ininducing growth. Even in the presence of GA3, IAA promotedgrowth only if it was applied directly to the apex rather than whenadded in the medium, apparently due to lack of acropetal trans-port. Table III shows a significant difference in growth betweenGA3 (control) and explants treated with GA3 + IAA. Theseexperiments show that growth of shoot tips is regulated by bothan auxin and a gibberellin.The second possibility, namely that basipetal transport of auxin
from the apex of shoot tips might decline in the absence of GA3,appeared to be supported by the observation that a 15- to 20-daydelay in the application of IAA in lanolin paste to the shoot tipoften failed to inhibit the development of basal bud. Experimentswith labeled IAA (Table IV) confirmed that the extent ofbasipetaltransport of auxin declined significantly with time in the absenceof GA3. This decline in transport was apparently not due to a
reduction in the uptake of IAA from the donor blocks by thesetissue segments (experiment 1). In contrast, equivalent segmentsfrom explants cultured in a medium containing GA3 appeared toretain the capacity to transport auxin. We studied the effectproduced by the application of 0.01% IAA in lanolin to shoot tipsfrom which GA3 application had been withheld for 20 days(experiment 2). It was found that following a delayed treatmentwith GA3 alone, the rate of transport of auxin remained low. The
Table III. Induction of Growth in Cuscuta Shoot Tip by DelayedApplication ofIAA and GA3
Shoot tips were cultured in BM for 8 to 21 days. These were then either(A) transferred to BM + GA3 (control), or (B) treated with 0.01% IAA inlanolin applied to the apex of the shoot tip and then transferred to BM +GA3. Growth was measured 1 month after the application of hormones.
B. +IAA 18 12.01 ± 1.00a2 A. Control 15 21 5.8 ± 0.48
B. +IAA 20 11.7 ± 1.4a3 A. Control 21 18 5.8 ± 0.15
B. +IAA 15 7.6 0.42ba Significantly different from control at 1% level.b Significantly different from control at 5% level.
application of auxin together with gibberellin, therefore, had arestorative effect, not only on growth but on auxin transport as
well.Effect of TIBA. TIBA, an inhibitor of polar transport of auxin,
was used to test further the involvement of IAA in the GA3-stimulated growth of shoot tips. TIBA at 0.5% in lanolin pastewas applied to the apex of shoot tip explants cultured in GA3 atthe beginning ofthe experiment. After 4 days, TIBA was reappliedat the apex. In an 18-day culture period, TIBA inhibited the GA3-stimulated growth by 60%o. Moreover, in all explants treated withTIBA, the basal bud developed by 15 days, indicating the abolition
Table IV. Basipetal Transport of['4CJIAA in Segments of Cuscuta ShootTips Cultured in Presence and Absence of GA3
Experiments 1: 1.0-cm-long segments from 1.5 cm below the apex wereused. Each datum is the average of 10 replicates. Experiment 2: 0.6-cm-long segments from 1.2 cm below the apex were used. Each datum is theaverage of four replicates.
RadioactivityExpen- Treatment DayReceiver Tissue
cpm ±SE
I A. Control (-GA3) 0 253 ± 58 4,021 ± 4962 100±6 4,490±3164 12±5 5,740±535
B. +GA3 2 318 ± 55 5,346 ± 4014 211±23 6,512±660
2 A. Control (GA3 ap-plied on day20) 408 25 ± 13 Not deter-
minedB. + Cold IAA (ap-
plied on day20) 40b 128 ± 21 Not deter-
mined
aThe average length was 4.2 cm.b The average length was 7.2 cm.
of apical dominance. The lateral shoots varied in length from 1 to15 cm. Such an early development of lateral shoots in shoot tipshad not been encountered under any experimental condition.
DISCUSSION
The elongation response of isolated Cuscuta shoot tips to GA3appears to be the largest so far observed for an isolated plantsystem. The response to GA3 was specific insofar as basally appliedauxin or cytokinin either inhibited or had no effect on elongation.Our observations with Cuscuta parallel those of Montague et al.
(11) with Avena, and of Silk and Jones (17) with lettuce, in thatthe tissue segments showed a decreased growth if GA3 treatmentswere given at increasingly later times after excision from the plant.Montague et al. (1 1) showed that this loss of response of segmentswas not due to decrease in the capacity of the segments to take upthe hormone. The decreased responsiveness to GA3 was inhibitedby low temperature and accelerated by anaerobiosis, and it wassuggested that this may be taken as physiological senescence ofthe segment. On the other hand, Silk and Jones (12) explained thereduced growth resulting from delayed GA3 application as beingdue to loss of ability to grow rather than to loss in sensitivity tohormone. In Cuscuta, the concentration of GA3 required formaximal response remained unaltered whether it was given onday 0 or on day 15 (data not given). When GA3 application wasdelayed, the shoot tip remained sensitive to a toxic concentrationof GA3 (1 mM). Thus, it is unlikely that there was any significantloss in sensitivity to hormone in its absence. The fact that somegrowth did resume in response to application of both IAA andGA3 (Table III) shows that the potential to grow was also nottotally lost, at least up to 2 to 3 weeks of culture in the absence ofGA3. However, the magnitude of growth response to delayedapplication of GA3 (Table II) or of IAA and GA3 (Table III) wassmaller than when GA3 alone was given on day 0 (Fig. I andTable II), suggesting that the nature of the decreased growth thatresulted from delay in the GA3 application is complex.The observation of an auxin requirement for growth in response
to delayed application of gibberellin gives some insight into thenature of the decreased growth referred to above. Data in TableIII reveal that both an auxin and a gibberellin interact to controlshoot tip growth in Cuscuta. If so, how then can we explain that
an exogenous application of IAA was required for growth onlywhen GA3 application was delayed, but not when it was given atthe beginning of the culture period? We hypothesize that contin-ued production of auxin at the apex of the shoot is dependentupon gibberellin; in the absence of gibberellin the production ofauxin at the apex of the shoot becomes very low or ceasesaltogether. This hypothesis, although not yet substantiated exper-imentally, is in accord with the observations. For example, thistype of gibberellin action would explain apical dominance inhormone-stimulated growth. Thus, a continuous supply of auxinfrom the shoot apex would not only be necessary for the concertedaction of hormones in controlling mitotic activity in the subapicaltissue (16), but also the increased concentration ofauxin functionalat a distance from the site of its production (4, 15) would inhibitthe growth oflateral buds in the presence ofgibberellin. Moreover,the hypothesis is in accordance with the observed inhibitory effectof TIBA on GA3-induced growth and with the loss of apicaldominance in TIBA-treated shoot tip explants. Furthermore, thehypothesis does not vitiate the observation (Table II) that IAAalone produced a small growth response when applied to the tipof day 0 of culture.A simple test of the above hypothesis would be to see whether
the auxin content of shoot tips cultured in the presence and theabsence of gibberellin was different. Support for such a hypothesisis given by Kuraishi and Muir (9) who found an increase indiffusible auxin from the apical portion of plants that were treatedwith GA3. That gibberellin application stimulates IAA productionin many plants is well established (5, 14). However, as in the peainternode (2) and the cucumber hypocotyl (6-8) gibberellin cannotact by an auxin-sparing mechanism, for both are required forgrowth. The experiment with decapitated shoot tips (Table II) isalso in accord with the postulate that in Cuscuta gibberellin actswith auxin in promoting shoot tip growth and that gibberellinregulates the synthesis of auxin in shoot tips. The site of auxinbiosynthesis in plants is the terminal bud (18). Therefore, gibber-ellin applied to a decapitated shoot tip may not be expected toresult in increased auxin production. Thus, decapitated Cuscutashoot tips responded optimally to. application of both auxin andgibberellin.
In shoot tips deprived of GA3, the capacity to transport IAAbecame greatly reduced (Table IV). This loss in transport capacitycould be due to the inhibition of growth, as shown by the fact thatdelayed application ofGA3 alone did not improve auxin transport.Transport was restored when growth commenced in response toapplication of both IAA and GA3. These observations are similarto those of Morris (12) who studied auxin transport in two-branched bean plants comprising a growing and an inhibitedshoot. The capacity of the inhibited shoot to transport IAA wasmuch less than that of the growing shoot. Following the release ofthe subordinate shoot from correlative inhibition, its capacity forauxin transport was restored. It is not known whether followingthe delayed application of IAA and GA3 to Cuscuta shoot tips,the capacity to synthesize endogenous auxin is restored or not.
Acknowledgments-The authors thank Prof. Kenneth V. Thimann, University ofCalifornia, for stimulating discussions during his visit to Bangalore in 1976 and forhis comments on the manuscript. Prof. Paul J. Vithayathil's help in assemblingresearch facilities and his constant encouragement are much appreciated.
LITERATURE CITED
1. BALDEV B 1959 In vitro culture of shoot apex of Cuscuta reflexa Roxb. MemoirsIndian Bot Soc 2: 40-42
2. BRIAN PW, HG HEMMING 1958 Complementary action of gibberellic acid andauxins in pea internode extension. Ann Bot 22: 1-77
3. HUMPHRIES EG 1956 Mineral components and ash analysis. In K Paech, MVTracey, eds, Moderne Methoden Der Pflanzenanalyse, Vol 1. Springer-Verlag,Berlin, p 481
4. JACOBS WP, DB CASE 1965 Auxin transport, gibberellin, and apical dominance.Science 148: 1729-1731
5. JONESRL 1973 The gibberellins, their physiological role. Annu Rev Plant Physiol
24: 571-5986. KATSUMI M, BO PHINNEY, WK PURVES 1965 The role of gibberellin and auxin
in cucumber hypocotyl growth. Physiol Plant 18: 462-4737. KAZAMA H, M KATSUMI 1973 Auxin-gibberellin relationships in their effects on
hypocotyl elongation of light-grown cucumber seedlings. Responses ofsectionsto auxin, gibberellin and sucrose. Plant Cell Physiol 14: 449-458
8. KAZAMA H, M KATSUMI 1974 Auxin-gibberellin relationships in their effects onhypocotyl elongation of light-grown cucumber seedlings. II. Effect of GA3-pretreatment of IAA-induced elongation. Plant Cell Physiol 15: 307-314
9. KURAISHI S, RM MUIR 1962 Increase in diffusible auxin after treatment withgibberellin. Science 137: 660-661
10. Loo SW 1946 Cultivation of excised stem tips of dodder in vitro. Am J Bot 33:295-300
1 1. MONTAGUE MJ, H IKUMA, PB KAUFMAN 1973 On the nature of the physiologicalresponses ofA vena stem segments to gibberellic acid treatment. Plant Physiol
PlaQt Physiol. Vol. 65, 1980
51: 1026-103212. MoRRs DA 1977 Transport of exogenous auxin in two branched dwarf pea
seedlings (Pisum sativum L.). Some implications for polarity and apical domi-nance. Planta 136: 91-96
13. MURASHIGE T, F SKOOG 1962 A revised medium for rapid growth and bioassayswith tobacco tissue cultures. Physiol Plant 15: 473497
14. PALEG LG 1965 Physiological effects of gibberellins. Annu Rev Plant Physiol 16:291-322
15. PILET PE 1965 Action of gibberellic acid on auxin transport. Nature 208: 134416. SACHs RM 1961 Gibberellin, auxin, and growth retardant effects upon cell
division and shoot histogenesis. Adv Chem 28: 49-5817. SiLK WK, RL JONES 1975 Gibberellin response in lettuce hypocotyl sections.
Plant Physiol 56: 267-27218. THIMANN KV, F SKOOG 1934 On the inhibition of bud development and other
functions of growth substance in Viciafaba. Proc R Soc Lond B 114: 317-339