154 heart hindgut stomach hepatopancreas oesophagus anus abdominal segment eye stalk antenna pereiopods pleopods Internal and External Anatomy of a Penaeid Shrimp Internal and external anatomy of a penaeid shrimp.
Transcript
154
hear
t
hind
gut
stom
ach
hepa
topa
ncre
as
oeso
phag
us
anus
abdo
min
al�
segm
ent
eye
stal
k
ante
nna
pere
iopo
ds
pleo
pods
Internal and External Anatomy ofa Penaeid Shrimp
Inte
rnal
and
ext
erna
l ana
tom
y of
a p
enae
id s
hrim
p.
155
SECTION 4 -CRUSTACEAN DISEASES
Internal and External Anatomy of a Penaeid ShrimpSECTION 4 - CRUSTACEAN DISEASESC.1 GENERAL TECHNIQUESC.1.1 Gross ObservationsC.1.1.1 BehaviourC.1.1.1.1 GeneralC.1.1.1.2 MortalitiesC.1.1.1.3 FeedingC.1.1.2 Surface ObservationsC.1.1.2.1 Colonisation and ErosionC.1.1.2.2 Cuticle Softening, Spots and DamageC.1.1.2.3 ColourC.1.1.2.4 Environmental ObservationsC.1.1.3 Soft-Tissue SurfacesC.1.2 Environmental ParametersC.1.3 General ProceduresC.1.3.1 Pre-collection PreparationC.1.3.2 Background InformationC.1.3.3 Sample Collection for Health SurveillanceC.1.3.4 Sample Collection for Disease DiagnosisC.1.3.5 Live Specimen Collection for ShippingC.1.3.6 Preservation of Tissue SamplesC.1.3.7 Shipping Preserved SamplesC.1.4 Record-KeepingC.1.4.1 Gross ObservationsC.1.4.2 Environmental ObservationsC.1.4.3 Stocking RecordsC.1.5 References
VIRAL DISEASES OF SHRIMPC.2 Yellowhead Disease (YHD)C.3 Infectious Hepatopancreas and Haematopoietic
C.AII List of Regional Resource Experts for CrustaceanDiseases in the Asia-Pacific
C.AIII List of Useful Manuals/Guide to CrustaceanDiseases in Asia-Pacific
List of National Coordinators(NCs)Members of Regional Working Group (RWG) and
Technical Support Services (TSS)List of Figures
Section 4 - Crustacean Diseases
216
219
221225
230
157
C.1 GENERAL TECHNIQUES
General crustacean health advice and othervaluable information are available from the OIEReference Laboratories, Regional ResourceExperts in the Asia-Pacific, FAO and NACA.A list is provided in Annexes F.AI and AII, andup-to-date contact information may be ob-tained from the NACA Secretariat in Bangkok(E-mail:[email protected]) . Other useful guidesto diagnostic procedures which provide valu-able references for crustacean diseases arelisted in Annex F.AIII.
C.1.1 Gross Observations
Gross observations of clinical signs in shrimpcan be easily made at the farm or pond sideusing little, if any, equipment. Although, in mostcases, such observations are insufficient for adefinite diagnosis, such information is essen-tial for preliminary compilation of a strong “casedescription” (or case history). Accurate anddetailed gross observations also help with ini-tiation of an action plan which can effectivelyreduce losses or spread of the disease, e.g.,destruction or isolation of affected stocks, treat-ments or alterations to husbandry practices (i.e.,feeding regimes, stocking densities, pondfertilisation, etc.). These can all be started whilewaiting for more conclusive diagnostic results.
C.1.1.1 Behaviour (Level 1)
C.1.1.1.1 General
Abnormal shrimp behaviour is often the first signof a stress or disease problem. Farmers or farmworkers, through daily contact with their stocks,rapidly develop a subconscious sense of when“something is wrong”. This may be subtlechanges in feeding behaviour, swimming move-ment or unusual aggregations. Even predatoractivity can provide clues to more “hidden”changes such as when fish- or shrimp-eatingbirds congregate round affected ponds.Record-keeping (see C.1.4) can provide valu-able additional evidence that reinforces suchobservations and can indicate earlier dateswhen problems started to appear. It is impor-tant that farmers and workers on the farm, aswell as field support staff, get to know the “nor-mal” (healthy) behaviour of their stocks. Sincesome species and growing environments maydemonstrate or evoke subtle differences inbehaviour, these should be taken into account,especially if changing or adding species, orwhen information gathered from a different
growing environment is used. Where anychange from normal behaviour affects morethan small numbers of random individuals, thisshould be considered cause for concern andwarrants investigation.
Some clues to look out for in shrimp stocks in-clude:• unusual activity during the daytime - shrimps
tend to be more active at night and stick todeeper water during the day
• swimming at or near pond surface or edges- often associated with lethargy (shrimpswimming near the surface may attractpredatory birds)
• increased feed consumption followed bygoing off-feed
• reduction or cessation of feeding• abnormal feed conversion ratios, length/
weight ratios• general weakening - lethargy (note: lethargy
is also characteristic in crustaceans when thewater temperature or dissolved oxygen lev-els are low, so these possibilities should beeliminated as potential causes before diseaseinvestigations are started)
C.1.1.1.2 Mortalities
Mortalities that reach levels of concern to a pro-ducer should be examined for any patterns inlosses, such as:
• relatively uniform mortalities throughout asystem should be examined immediately andenvironmental factors determined (ideallywith pre-mortality records - see C.1.4)
• apparently random, or sporadic mortalitiesmay indicate a within-system or stock prob-lems. If the following conditions exist - (a)no history of stock-related mortalities, (b) allstock originate from the same source, and(c) there have been no changes to the rear-ing system prior to mortality problems -samples of affected and unaffected shrimpshould be submitted for laboratory exami-nation (Level II or III), as appropriate, andsupported by gross observations and stockhistory (see C.1.4)
• mortalities that spread suggest an infec-tious cause and should be sampled immedi-ately. Affected shrimp should be kept as faraway as possible from unaffected shrimp untilthe cause of the mortalities can be estab-lished.
158
C.1.1.1.3 Feeding
Absence of feeding behaviour and lack of feedin the gut are good indicators of potential prob-lems. Daily gut content checks can be madeon shrimp caught in feeding trays or bowls(where used) or, less frequently, from samplestaken to determine growth. Ideally examinationof feeding behaviour should be made every 1-2 weeks, even in extensive farming systems.Feeding behaviour is most easily checked byplacing feed in a tray or bowl (Fig.C.1.1.1.3a)and seeing how quickly the shrimp respond,ideally after the shrimp have not been fed for atleast a few hours. It is important that the feedused is attractive to the shrimp as poorly for-mulated, old or badly stored feeds may not beattractive to the shrimp. Gut contents can bechecked by holding the shrimp against a lightto show the gut in the tail segments(Fig.C.1.1.1.3b). If these are empty, especiallyjust after providing feed, it may indicate eitherof the following conditions: i) underfeeding, orii) onset of cessation of feeding (anorexia).
Where possible, feed records (see C.1.4) shouldbe maintained to determine normal feed con-sumption patterns (i.e., feeding activity byhealthy shrimp), which can be compared with“suspect” feeding activity. In many cases ofchronic loss, daily feed consumption patternsmay remain stable or oscillate over periods ofseveral weeks. These can be detected by mak-ing a graph of daily feed consumption or bycomparing daily feed consumption in the recordbook over an extended period (e.g. 3-4 weeks).
C.1.1.2 Surface Observations (Level 1)
C.1.1.2.1 Colonisation and Erosion
Colonisation of the shell (cuticle) and gills of acrustacean is an on-going process that is usu-ally controlled by grooming. The presence ofnumerous surface organisms (e.g. “parasites”- which damage their host; or “commensals” -that do not adversely impact their host) sug-gests sub-optimal holding conditions or a pos-sible disease problem. Apparent wearing away(erosion) of the cuticle or appendages (legs, tail,antennae, rostrum) (Fig.C.1.1.2.1a), or loss ofappendages, with or without blackening (mela-nization) are also highly indicative of a diseaseproblem. Breakage of the antennae is an earlywarning sign. In healthy penaeid shrimp, theseshould extend approximately 1/3 past thelength of the body (when bent back along thebody line). Likewise, erosion or swelling of the
tail (uropods and telson), with or without black-ening, is an early sign of disease(Fig.C.1.1.2.1b).
C.1.1.2.2 Cuticle Softening, Spots andDamage
Softening of the shell (Fig.C.1.1.2.2a andFig.C.1.1.2.2b), other than during a moult, mayalso indicate the presence of infection. Dam-age or wounds to the shell provide an opportu-nity for opportunistic infections (mainly bacte-rial and fungal) to invade the soft-tissues andproliferate, which can seriously impact thehealth of the shrimp.
Certain diseases, such as White Spot Disease,directly affect the appearance of the shell, how-ever, few changes are specific to a particularinfection. In the case of white spots on the cu-ticle, for example, recent work (Wang et al. 2000)has shown that a bacteria can produce signssimilar to those produced by White Spot Dis-ease (see C.4) and Bacterial White Spot Syn-drome (see C.4a).
C.1.1.2.3 Colour
Shrimp colour is another good indicator ofhealth problems. Many crustaceans becomemore reddish in color when infected by a widerange of organisms, or when exposed to toxicconditions (Fig.C.1.1.2.3a), especially those thataffect the hepatopancreas. This is thought tobe due to the release of yellow-orange (caro-tenoid) pigments that are normally stored in thehepatopancreas. This red colour is not specificfor any single condition (or groups of infections),however, so further diagnosis is needed.
Yellowish coloration of the cephalothorax isassociated with yellowhead disease (see C.2)and overall reddening can be associated withgill associated virus infections (see C.6), whitespot disease or bacteria, as described above,or bacterial septicemia (see C.10). In somecases, the colour changes are restricted to ex-tremities, such as the tail fan or appendages(Fig.C.1.1.2.3b), and these should be examinedclosely.
It should be noted that some shrimpbroodstock, particularly those from deeperwaters, can be red in colour (thought to be dueto a carotenoid-rich diet). This does not appearto be related to health and its normality can beestablished through familiarisation with the spe-cies being grown. Under certain conditions,some crustaceans may turn a distinct blue
C.1 General Techniques
159
(P Chanratchakool)
Fig.C.1.1.1.3a. Behaviour observation ofshrimp PL in a bowl.
(P Chanratchakool)
Fig.C.1.1.1.3b. Light coloured shrimp with fullguts from a pond with healthy phytoplankton.
(P Chanratchakool)
Fig.C.1.1.2.1a. Black discoloration of damagedappendages.
(P Chanratchakool)
(P Chanratchakool/MG Bondad-Reantaso)
Fig.C.1.1.2.2a,b. Shrimp with persistent softshell.
(P Chanratchakool)
Fig.C.1.1.2.3a. Abnormal blue and red discol-oration.
Fig.C.1.1.2.1b. Swollen tail due to localizedbacterial infection.
>
a
b
160
disease (see C.4) or the effect of salinity on theexpression of necrotising hepatopancreatitis(see C.10). This is especially important for spe-cies grown under conditions that bear littleresemblance to the wild situation. Water tem-perature, salinity, turbidity, fouling and plank-ton blooms (Fig. C.1.2 a,b,c and d) are all im-portant factors. Rapid changes in conditions,rather than gradual changes, are particularly im-portant as potential triggers for disease. There-fore, the farm manager and workers, should at-tempt to keep pond rearing conditions withinthe optimum range for the species and as con-stant as possible within that range. High stock-ing rates are common in aquaculture but pre-dispose individuals to stress so that even mi-nor changes in environmental conditions mayprecipitate disease. In addition, many smallchanges do not, on their own, affect shrimphealth. However, when several of these smallchanges occur simultaneously, results can befar more severe.
C.1.3 General Procedures
C.1.3.1 Pre-collection Preparation(Level I)
The diagnostic laboratory which will be receiv-ing the sample should be consulted to ascer-tain the best method of transportation (e.g., onice, preserved in fixative, whole or tissuesamples). The laboratory will also indicate ifboth clinically affected, as well as apparentlyhealthy individuals, are required for compara-tive purposes. As noted under C.1.3.3 andC.1.3.4, screening and disease diagnosis of-ten have different sample-size requirements.
The laboratory should be informed of exactlywhat is going to be sent (i.e., numbers, size-classes or tissues) and the intended date of col-lection and delivery, as far in advance as pos-sible. For screening healthy animals, samplesizes are usually larger so more lead time is re-quired by the laboratory. Screening can be alsobe planned ahead of time, based on predicteddates of shipping post-larvae (PL) orbroodstock, which means the shipper has moretime to notify the laboratory well in advance. Incases of disease outbreaks and significantmortalities, there may be less opportunity foradvance warning for the laboratory. However,the laboratory should still be contacted prior toshipment or hand-delivery of any diseasedsamples (for the reasons given under C.1.3.4).Some samples may require secured packag-ing or collection by designated personnel, ifthere are national or international certification
colour. This has been shown to be due to lowlevels of a carotenoid pigment in the hepato-pancreas (and other tissues), which may be in-duced by environmental or toxic conditions.Normal differences in colouration (light to dark)within a species may be due to other environ-mental variables. For example, Penaeusmonodon grown in low salinities, are often muchpaler than P. monodon grown in brackish-wa-ter or marine conditions. These variations donot appear to be related to general health.
C.1.1.2.4 Environmental Observations
Shrimp with brown gills or soft shells (or a rep-resentative sub-sample), should be transferredto a well aerated aquarium with clean sea wa-ter at the same salinity as the pond from whichthey came. They should be observed every 1-2hrs over 1 day. If the shrimp return to normalactivity within a few hours, check environmen-tal parameters in the rearing pond(s).
C.1.1.3 Soft-Tissue Surfaces (Level 1)
A readily observable change to soft tissues isfouling of the gill area (Fig. C.1.1.3a), sometimesaccompanied by brown discoloration (Fig.C.1.1.3b) (see C.1.1.2.4). This can be due todisease and should trigger action since it re-duces the shrimp’s ability to take up oxygenand survive.
Removal of the shell in the head region ofshrimp allows gross examination of the organsin this region, particularly the hepatopancreas(Fig.C.1.1.3c). In some conditions, the hepato-pancreas may appear discoloured (i.e., yellow-ish, pale, red), swollen or shrunken, comparedwith healthy shrimp. If the hepatopancreas isgently teased out of the shell, the mid-gut willbecome visible and permit direct examinationof colour (dark - feeding; light/white/yellow -mucoid, empty or not feeding - see C.1.1.1.3).This information is useful for determining thehealth of the shrimp and if infectious diseaseagents are present.
C.1.2 Environmental Parameters(Level 1)
Environmental conditions can have a significanteffect on crustacean health, both directly (withinthe ranges of physiological tolerances) and in-directly (enhancing susceptibility to infectionsor their expression). Examples include changesto dissolved oxygen levels and/or pH which maypromote clinical expression of previously latentyellowhead disease (see C.2) and white spot
C.1 General Techniques
161
C.1 General Techniques
(P Chanratchakool)
Fig.C.1.1.3a. Severe fouling on the gills.
(P Chanratchakool)
Fig.C.1.1.3b. Brown discolouration of the gills.
(P Chanratchakool)
Fig.C.1.1.3c. Shrimp on left side with smallhepatopancreas.
(P Chanratchakool)
Fig.C.1.2a, b, c. Examples of different kinds ofplankton blooms (a- yellow/green colouredbloom; b- brown coloured bloom; c- blue-greencoloured bloom.
(P Chanratchakool)
Fig.C.1.2d. Dead phytoplankton.
(V Alday de Graindorge and TW Flegel)
>
Fig. C.1.3.6. Points of injection of fixative.
a
b
c
162
requirements or risk of disease spread via trans-port of the sample to an area non-endemic fora suspected disease.Pre-collection discussions with the diagnosticlaboratory can significantly speed up process-ing and diagnosis of a sample (days to weeks)since it allows preparation of the required di-agnostic materials in advance of arrival of thesample(s) and ensures that emergency samplesare scheduled in for rapid diagnosis.
C.1.3.2 Background Information (Level 1)
All samples submitted for diagnosis should in-clude as much supporting information as pos-sible including:
• Gross observations and a history of environ-mental parameters (as described under C.1.1and C.1.2)
• Approximate prevalence and pattern of mor-tality (acute or chronic/sporadic cumulativelosses)
• History and origin of affected population• If the stock is not local, their origin(s) and
date(s) of transfer should be included• Details of feed, consumption rates and any
chemical treatments used
The above information provides valuable back-ground details which can help focus attentionon possible handling stress, changes in envi-ronment or infectious agents as the primarycause of any health problems.
C.1.3.3 Sample Collection for Health Sur-veillance
The most important factors associated withcollection of specimens for surveillance are:
• sample numbers that are high enough toensure adequate pathogen detection (seeC.1.3.1 and Table C.1.3.3). Check the num-ber of specimens required by the laboratorybefore collecting the sample(s) and ensurethat each specimen is intact. Larger numbersare generally needed for screening purposes,compared to numbers required for diseasediagnosis;
• susceptible species are sampled;• samples include age- or size-groups that are
most likely to manifest detectable infections.Such information is given under the specificdisease sections; and
• samples are collected during the seasonwhen infections are known to occur. Suchinformation is also given under the specificdisease sections.
As mentioned under C.1.3.1, check whether ornot designated personnel are required to do thecollection, or if secured packaging is necessary,or whether samples are being collected to meetnational or international certification require-ments.
C.1.3.4 Sample Collection for DiseaseDiagnosis
All samples submitted for disease diagnosisshould include as much supporting informationas possible, as described under C.1.3.2, withparticular emphasis on:
• rates and levels of mortality compared with“normal” levels for the time of year;
• patterns of mortality (random/sporadic,localised, spreading, widespread);
• history and origin(s) of the affectedpopulation(s); and
• details of feed used, consumption rates andany chemical treatments.
As in C.1.3.2, the above information will helpclarify whether or not an infectious agent is in-volved and will enable to focus the investiga-tive procedures required for an accurate diag-nosis. This information is also critical for labo-ratories outside the region or areas where thesuspected disease is endemic. Under such cir-cumstances, the laboratory may have to pre-pare for strict containment and sterile disposalof all specimen shipping materials and wasteproducts, in order to prevent escape from thelaboratory.
Wherever possible, check the number of speci-mens required by the laboratory for diagnosticexamination before collecting the sample(s).Also check with the laboratory to see whetheror not they require specimens showing clinicalsigns of disease only, or sub-samples of bothapparently healthy individuals and clinically af-fected specimens from the same pond/site. Thelatter option is usually used where a disease-outbreak or other problem is detected for thefirst time. Comparative samples can help pin-point abnormalities in the diseased specimens.
C.1.3.5 Live Specimen Collection forShipping (Level 1)
Once the required sample size is determined,the crustaceans should be collected from thewater. This should take place as close to ship-ping as possible to reduce possible mortalitiesduring transportation (especially important formoribund or diseased samples). Wherever pos-
C.1 General Techniques
163
Table C.1.3.31 . Sample sizes needed to detect at least one infected host in a population of agiven size, at a given prevalence of infection. Assumptions of 2% and 5% prevalences are mostcommonly used for surveillance of presumed exotic pathogens, with a 95% confidence limit.
Prevalence (%)
Population Size 0.5 1.0 2.0 3.0 4.0 5.0 10.0
50 46 46 46 37 37 29 20
100 93 93 76 61 50 43 23
250 192 156 110 75 62 49 25
500 314 223 127 88 67 54 26
1000 448 256 136 92 69 55 27
2500 512 279 142 95 71 56 27
5000 562 288 145 96 71 57 27
10000 579 292 146 96 72 29 27
100000 594 296 147 97 72 57 27
1000000 596 297 147 97 72 57 27
>1000000 600 300 150 100 75 60 30
1 Ossiander, F.J. and G. Wedermeyer. 1973. Journal Fisheries Research Board of Canada 30:1383-1384.
sible, ensure that each specimen is intact.
As noted under C.1.3.1, inform the laboratoryof the estimated time of arrival of the sampleso they can have the materials required to pro-cess prepared before the samples arrive. Thisshortens the time between removal from thepond and preparation of the specimens for ex-amination.
The crustaceans should be packed in seawa-ter in double plastic bags with the airspace inthe bag filled with oxygen. The bags should besealed tightly with rubber bands or rubber ringsand packed inside a foam box. A small amountof ice may be added to keep the water cool,especially if a long transport time is expected.This box is then taped securely and may bepackaged inside a cardboard carton. Checkwith the diagnostic laboratory about packingrequirements. Some laboratories have specificpackaging requirements for diseased organ-isms. Samples submitted for certification pur-poses may have additional shipping and col-lection requirements (see C.1.3.3).
Label containers clearly:
“LIVE SPECIMENS, STORE AT ___ to ___˚C, DONOT FREEZE”(insert temperature tolerance range of shrimpbeing shipped)
If being shipped by air also indicate:
“HOLD AT AIRPORT AND CALL FOR PICK-UP”
• Clearly indicate the name and telephonenumber of the contact person responsible forpicking up the package at the airport or re-ceiving it at the laboratory.
• Where possible, ship early in the week toavoid arrival during the weekend which maylead to loss through improper storage ofsamples.
• Inform the contact person as soon as theshipment has been sent and, where appro-priate, give them the name of the carrier, theflight number, the waybill number and theestimated time of arrival.
(Note: Some airlines have restrictions on ship-ping of seawater or preserved samples. It is agood idea to check with local airlines if they dohave any special requirements)
C.1 General Techniques
164
C.1.3.6 Preservation of Tissue Samples(Level 2)
In some cases, such as locations remote froma diagnostic laboratory or where transport con-nections are slow, it may not be possible to pro-vide a live shrimp sample. Since freezing is usu-ally inadequate for most diagnostic techniques(histology, bacteriology, mycology, etc.), speci-mens should be fixed (chemical preservationto prevent tissue breakdown and decay) on site.This makes the sample suitable for subsequenthistological examination, in situ hybridization,PCR or electron microscopy, but will preventroutine bacteriology, mycology, virology or othertechniques requiring live micro-organisms. Di-agnostic needs should therefore be dis-cussed with the laboratory prior to collect-ing the sample.
The best general fixative for penaeid shrimp isDavidson’s fixative.
330 ml 95% ethanol220 ml 100% formalin (37% w/v formalde-hyde in aqueous solution)115 ml glacial acetic acid335 ml distilled water.Mix and store at room temperature.
(It should be noted, however, that formalin resi-dues can interfere with the PCR process.Samples for PCR analysis should be fixed in70% ethanol.)
For any preservation procedure, it is essentialto remember that the main digestive organ ofthe shrimp (the hepatopancreas) is very impor-tant for disease diagnosis, but undergoes rapidautolysis (tissue digestion by digestive juicesreleased from the dying hepatopancreatic cells)immediately after death. This means that thepre-death structure of the hepatopancreas israpidly lost (turns to mush). Delays of even afew seconds in fixative penetration into this or-gan can result in the whole specimen beinguseless for diagnosis, thus, specimens must beimmersed or injected with fixative while stillalive. Dead shrimp, even when preserved onice (or frozen) are of no use for subsequent fixa-tion. In tropical areas, it is best to use cold fixa-tive that has been stored in the freezer or kepton ice, as this helps arrest autolysis and sec-ondary microbial proliferation, as the tissues arepreserved.
Larvae and early post larvae (PL) should beimmersed directly in a minimum of 10 volumesof fixative to one volume of shrimp tissue. This
10:1 ratio is critical for effective preservation.Attempts to cut costs by using lower ratios offixative to tissue can result in inadequate pres-ervation of tissues for processing.
For PL that are more than approximately 20 mmin length, use a fine needle to make a small,shallow incision that breaks and slightly lifts thecuticle in the midline of the back, at the cuticu-lar junction between the cephalothorax and firstabdominal segment. This allows the fixative topenetrate the hepatopancreas quickly.
For larger PL’s, juveniles and adults, the fixa-tive should be injected directly into the shrimp,as follows:
• Place the shrimp briefly in ice water to se-date them
• Using surgical rubber gloves and protectiveeyeglasses, immediately inject the fixative(approximately 10% of the shrimp’s bodyweight) at the following sites (Fig. C.1.3.6):
º hepatopancreasº region anterior to the hepatopancreasº anterior abdominal region, andº posterior abdominal region.
Be careful to hold the shrimp so the angle ofinjection is pointed away from your body, sincefixative can sometimes spurt back out of aninjection site when the needle is removed andmay injure the eyes. It is also best to brace theinjection hand against the forearm of the handholding the shrimp, to avoid over penetrationof the needle into that hand. The hepatopan-creas should receive a larger proportion of theinjected fixative than the abdominal region. Inlarger shrimp it is better to inject the hepato-pancreas at several points. All signs of lifeshould cease and the colour should change atthe injection sites.
Immediately following injection, slit the cuticlewith dissecting scissors along the side of thebody from the sixth abdominal segment to thecuticle overlying the “head region” (cephalotho-rax). From there, angle the cut forward and up-ward until it reaches the base of the rostrum.Avoid cutting too deeply into the underlying tis-sue. Shrimp over 12 g should be transverselydissected, at least once, posterior of the abdo-men/cephalothorax junction and again mid-abdominally. The tissues should then be im-mersed in a 10:1 volume ratio of fixative to tis-sue, at room temperature. The fixative can bechanged after 24-72 hr to 70% ethanol, for long-term storage.
C.1 General Techniques
165
C.1.3.7 Shipping Preserved Samples(Level 1)
For shipping, remove specimens from ethanolstorage, wrap in paper towel saturated with50% ethanol and place in a sealed plastic bag.There should be no free liquid in the bag. Sealand place within a second sealed bag. In mostcountries, small numbers of such specimenscan be sent to diagnostic laboratories by air-mail. However, some countries or transportcompanies (especially air couriers) have strictregulations regarding shipping any chemicals,including fixed samples for diagnostic exami-nation. Check with the post office or carrierbefore collecting the samples to ensure theyare processed and packed in an appropriateand acceptable manner. All sample bags shouldbe packed in a durable, leak-proof container.
Label containers clearly with the name and tele-phone number of the contact person respon-sible for picking up the package at the airportor receiving it at the laboratory.
If being shipped by air indicate - “HOLD AT AIR-PORT AND CALL FOR PICK-UP”.
Where possible, ship early in the week to avoidarrival during the weekend which may lead toloss through improper storage of samples. In-form the contact person as soon as the ship-ment has been sent and, where appropriate,give them the name of the carrier, the flight num-ber, the waybill number and the estimated timeof arrival.
C.1.4 Record-Keeping (Level 1)
Record-keeping is essential for effective dis-ease management. For crustaceans, many ofthe factors that should be recorded on a regu-lar basis are outlined in sections C.1.1 andC.1.2. It is critical to establish and record nor-mal behaviour and appearance to compare withobservations during disease events.
C.1.4.1 Gross Observations (Level 1)
These could be included in routine logs of crus-tacean growth which, ideally, would be moni-tored on a regular basis either by sub-samplingfrom tanks or ponds, or by “best-guess” esti-mates from surface observations.
For hatchery operations, the minimum essen-tial information which should be recorded/logged include:
These observations should be recorded on adaily basis for all stages, and include date, time,tank, broodstock (where there are more thanone) and food-source (e.g., brine shrimp cul-ture batch or other food-source). Dates andtimes for tank and water changes should alsobe noted, along with dates and times for pipeflushing and/or disinfection. Ideally, these logsshould be checked regularly by the person re-sponsible for the site/animals.
Where possible, hatcheries should invest in amicroscope and conduct daily microscopicexaminations of the larvae. This will allow themto quickly identify problems developing withtheir stocks, often before they become evidentin the majority of the population.
For pond sites, the minimum essential obser-vations which need to be recorded/logged in-clude:
• growth• feed consumption• fouling• mortality
These should be recorded with date, site loca-tion and any action taken (e.g., sample collec-tion for laboratory examination). It is importantto understand that rates of change for theseparameters are essential for assessing thecause of any disease outbreak. This means lev-els have to be logged on a regular and consis-tent basis in order to detect patterns over time.Ideally, these logs should be checked regularlyby the person responsible for the site/animals.
C.1.4.2 Environmental Observations(Level 1)
This is most applicable to open ponds. Theminimum essential data that should be recordedare:
• temperature• salinity• pH• turbidity (qualitative evaluation or secchi disc)• algal bloom(s)• human activity (treatments, sorting, pond
changes, etc.)• predator activity
C.1 General Techniques
166
As with C.1.4.1, types and rates of changes inthese parameters prior to any disease out-breaks are extremely important in assessing thecause of the outbreak. Although helpful, datarecorded on the day of specimen collection aremuch less useful than continuous records.Thus, the importance of keeping careful, regu-lar and continuous records, regardless of the“expected” results, cannot be overstressed.
Frequency of record-keeping will vary with siteand, possibly, season. For example, more fre-quent monitoring may be required during un-stable weather, compared to seasons with ex-tended, stable, conditions.
Human and predator activity should be loggedon an “as it happens” basis.
C.1.4.3 Stocking Records (Level 1)
All movements of crustaceans into and out of ahatchery and pond/site should be recorded.These should include:
• the exact source of the broodstock or larvaeand any health certification history (e.g., re-sults of any tests carried out prior to/on ar-rival)
• condition on arrival• date, time and person responsible for receiv-
ing delivery of the stock• date, time and destination of stock shipped
out of the hatchery
In addition, all movements of stocks within ahatchery, nursery or grow out site should belogged with the date for tracking purposes if adisease situation arises.Where possible, animals from different sourcesshould not be mixed. If mixing is unavoidable,keep strict records of when mixing occurred.
C.1.5 References
Alday de Graindorge, V. and T.W. Flegel. 1999.Diagnosis of shrimp diseases with emphasison black tiger prawn, Penaeus monodon.Food and Agriculture Organization of theUnited Nations (FAO), Multimedia Asia Co.,Ltd, BIOTEC, Network of Aquaculture Cen-tres in Asia Pacific (NACA) and SoutheastAsian Chapter of the World Aquaculture So-ciety (WAS). Bangkok, Thailand. (InteractiveCD-ROM format).
Chanratchakool, P., J.F. Turnbull, S.J. Funge-Smith, I.H. MacRae and C. Limsuan.1998.Health Management in Shrimp Ponds. Third
Edition. Aquatic Animal Health Research In-stitute. Department of Fisheries. Bangkok,Thailand. 152p.
Chanratchakool, P., J.F. Turnbull, S. Funge-Smith and C. Limsuan. 1995. Health Man-agement in Shrimp Ponds. Second Edition.Aquatic Animal Health Research Institute.Department of Fisheries. Bangkok, Thailand.111p.
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDiseases of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Ossiander, F.J. and G. Wedermeyer. 1973. Com-puter program for sample size required todetermine disease incidence in fish popula-tions. J. Fish. Res. Bd. Can. 30: 1383-1384.
Wang,Y.G., K. L. Lee, M. Najiah, M. Shariff andM. D. Hassan. 2000. A new bacterial whitespot syndrome (BWSS) in cultured tigershrimp Penaeus monodon and its compari-son with white spot syndrome (WSS) causedby virus. Dis. Aquat.Org. 41:9-18.
C.1 General Techniques
167
C.2.1 Background Information
C.2.1.1 Causative Agent
Yellowhead disease (YHD) is caused byYellowhead Virus (YHV) (also reported in olderliterature as Yellowhead Baculovirus - YBV andYellowhead Disease Baculovirus - YHDBV). Itis now known not to be a member of theBaculoviridae. YHV is a single stranded RNA,rod shaped (44 ± 6 X 173 ±13 nm), envelopedcytoplasmic virus, likely related to viruses in theFamily Coronaviridae. Agarose gel electrophore-sis indicates a genome size of approximately 22Kilobases. Lymphoid organ virus (LOV) and gillassociated virus (GAV) (see C.6) of Penaeusmonodon in Australia are related to the YHV com-plex viruses, although, of the two, only GAV isknown to cause mortality. More detailed infor-mation about the disease can be found in the OIEDiagnostic Manual for Aquatic Animal Diseases(OIE 2000a) and Lightner (1996).
C.2.1.2 Host Range
Natural infections occur in Penaeus monodon,but experimental infections have been shown inP. japonicus, P. vannamei, P. setiferus, P. aztecus,P. duorarum and P. stylirostris. Penaeusmerguiensis, appear to be resistant to disease(but not necessarily infection). Palaemonstyliferus has been shown to be a carrier of vi-able virus. Euphausia spp. (krill), Acetes spp. andother small shrimp are also reported to carry YHDviruses.
C.2.1.3 Geographic Distribution
YHD affects cultivated shrimp in Asia includingChina PR, India, Philippines and Thailand.YHD has been reported from cultured shrimp inTexas and one sample has been reported to bepositive for YHV by antibody assay (Loh et al.1998).
C.2.1.4 Asia-Pacific Quarterly AquaticAnimal Disease Reporting System (1999-2000)
YHD was reported in Malaysia in June, in thePhilippines in January to March and July; in SriLanka in January and suspected for the wholeyear of 1999 in Thailand. For the reporting pe-riod for the year 2000, India reported it in Octo-ber and it was suspected for the whole year inThailand and Sri Lanka (OIE 1999, OIE 2000b).
VIRAL DISEASES OF SHRIMPC.2 YELLOWHEAD DISEASE (YHD)1
C.2.2 Clinical Aspects
Gross signs of disease (Fig.C.2.2) and mortal-ity occur within 2 to 4 days following an inter-val of exceptionally high feeding activity thatends in abrupt cessation of feeding. Mortali-ties can reach 100% within 3-5 days. Diseasedshrimp aggregate at the edges of the ponds ornear the surface. The hepatopancreas becomesdiscoloured which gives the cephalothorax ayellowish appearance, hence the name of thedisease. The overall appearance of the shrimpis abnormally pale. Post-larvae (PL) at 20-25days and older shrimp appear particularly sus-ceptible, while PL<15 appear resistant.
Care must be taken in gross diagnosis as mor-talities caused by YHD have been reported inthe absence of the classic yellowish appearanceof the cephalothorax. Clinical signs are not al-ways present, and their absence does not ruleout the possibility of YHD infection. Further con-firmatory diagnosis including a minimum of whole,stained gill mounts and haemolymph smearsshould be made in any cases of rapid unexplainedmortality in which YHV involvement cannot beruled out.
YHD virions are found generally in tissues ofectodermal and mesodemal embryonic origin,including: interstitial tissues of the hepatopan-creas, systemic blood cells and developing bloodcells in the haematopoietic tissues and fixed ph-agocytes in the heart, the lymphoid (Oka) organ,gill epithelial and pilar cells, connective andspongioform tissues, sub-cuticular epidermis,striated and cardiac muscles, ovary capsules,nervous tissue, neurosecretory and ganglial cells,stomach, mid-gut and midgut caecal walls. Theepithelial cells of hepatopancreatic tubules, mid-gut and midgut caecae (endodermal origin) arenot infected with YHV although underlying muscleand connective tissues are. The Oka organ, gill,heart and subcuticular tissues, including thoseof the stomach epithelium, contain the highestlevels of YHV. Infected cells show nuclear py-knosis and karyorrhexis which are apparentlysigns of viral triggered apoptosis (Khanobdee etal. 2001).
C.2.3 Screening Methods
More detailed information on methods forscreening YHD can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000a), at http://www.oie.int, or at selected ref-erences.
1 Yellowhead disease (YHD) is now classified as an OIE Notifiable Disease (OIE 2000a).
168
C.2.3.1 Presumptive
There are no gross observations (Level I) or his-topathological (Level II) diagnostic techniqueswhich can provide presumptive detection ofYHD in sub-clinical shrimp.
(TW Flegel)
Fig.C.2.2. Gross sign of yellow head disease(YHD) are displayed by the three Penaeusmonodon on the left.
(DV Lightner)
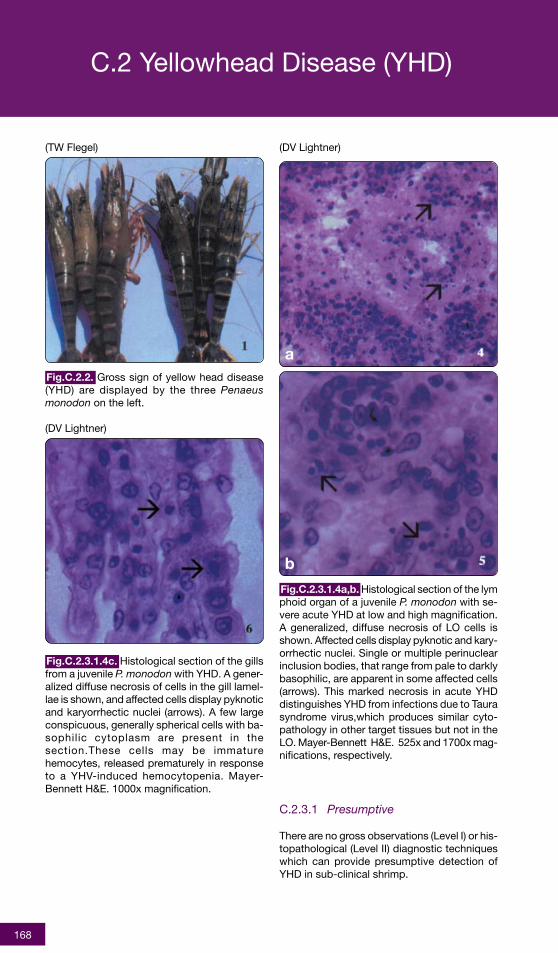
Fig.C.2.3.1.4a,b. Histological section of the lymphoid organ of a juvenile P. monodon with se-vere acute YHD at low and high magnification.A generalized, diffuse necrosis of LO cells isshown. Affected cells display pyknotic and kary-orrhectic nuclei. Single or multiple perinuclearinclusion bodies, that range from pale to darklybasophilic, are apparent in some affected cells(arrows). This marked necrosis in acute YHDdistinguishes YHD from infections due to Taurasyndrome virus,which produces similar cyto-pathology in other target tissues but not in theLO. Mayer-Bennett H&E. 525x and 1700x mag-nifications, respectively.
C.2 Yellowhead Disease (YHD)
(DV Lightner)
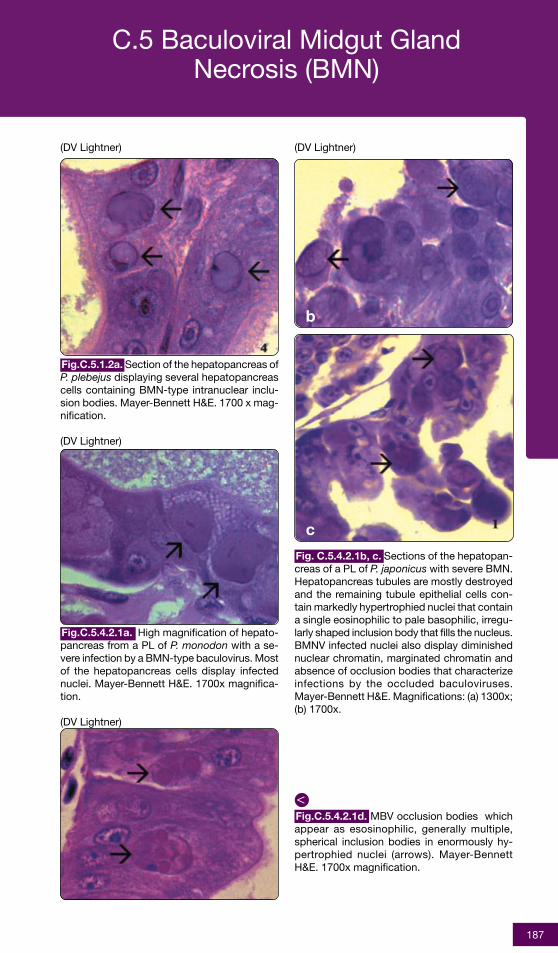
Fig.C.2.3.1.4c. Histological section of the gillsfrom a juvenile P. monodon with YHD. A gener-alized diffuse necrosis of cells in the gill lamel-lae is shown, and affected cells display pyknoticand karyorrhectic nuclei (arrows). A few largeconspicuous, generally spherical cells with ba-sophilic cytoplasm are present in thesection.These cells may be immaturehemocytes, released prematurely in responseto a YHV-induced hemocytopenia. Mayer-Bennett H&E. 1000x magnification.
For certification of YHV infection status ofbroodstock and fry, reverse transcriptase-poly-merase chain reaction (RT-PCR) technology isrecommended.
There are several commercially available RT-PCR kits now available to screen haemolymphfrom broodstock shrimp and PL tissues for evi-dence of YHV RNA.
C.2.4 Diagnostic Methods
More detailed information on methods for diag-nosis can be found in the OIE Diagnostic Manualfor Aquatic Animal Diseases (OIE 2000a), at http://www.oie.int, or at selected references.
C.2.4.1 Presumptive
C.2.4.1.1 Gross Observations (Level 1)
YHD can be suspected when an abnormal in-crease in feeding rates is followed by a sharpcessation in feeding. Moribund shrimp may ap-pear near the surface or edges of grow out pondsand show slow swimming behaviour in responseto stimuli. These may also show pale overall bodycolouration, a yellowish cephalothorax, pale gillsand hepatopancreas. YHD should be suspectedunder such circumstances, especially for P.monodon, and samples collected for confirma-tory diagnosis.
C.2.4.1.2 Gill Squash (Level II)
Fix whole shrimp, or gill filaments, in Davidson’sfixative overnight2 . Wash gill filament in tap wa-ter to remove the fixative and stain with Mayer-Bennett’s H&E. Clear in xylene and, using a finepair of needles (a stereo microscope is helpful),break off several secondary filaments and re-place the main filament in xylene for perma-nent reference storage in a sealed vial. Mountsecondary filaments, coverslip and use lightpressure to flatten the filaments as much aspossible, making them easy to see through. Thissame procedure can be used on thin layers ofsubcuticular tissue.
Moderate to large numbers of deeply baso-philic, evenly stained, spherical, cytoplasmic in-clusions approximately 2 mm in diameter orsmaller are presumptive for YHD, along withsimilar observations from haemolymph smears.As with tissue sections and wet-fixed gill fila-ments, these slides can be kept as a perma-nent record.
C.2.4.1.3 Haemolymph Smears (Level II)
Smears that show moderate to high numbers ofblood cells with pycnotic and karyorrhexic nu-clei, with no evidence of bacteria, can be indica-tive of early YHD. It is important that no bacteriaare present, since these can produce similarhaemocyte nucleus changes. Such changes aredifficult to see in moribund shrimp because ofthe loss of blood cells so grossly normal shrimpshould be sampled for these signs from the samepond where the moribund shrimp were obtained.The haemolymph is collected in a syringe con-taining twice the haemolymph volume of 25%formalin or modified Davidson’s fixative (i.e., withthe acetic acid component replaced by water orformalin). The blood cell suspension is mixedthoroughly in the syringe, the needle removedand a drop placed onto a microscope slide.Smear and air dry the preparation before stain-ing with H&E and eosin or other standard bloodstains. Dehydrate, mount and coverslip. The re-sults should be consistent with the gill wholemounts (above) or histopathology of tissue sec-tions, in order to make a presumptive YHD diag-nosis.
C.2.4.1.4 Histopathology (Level II)
Fix moribund shrimp from a suspected YHD out-break in Davidson’s fixative and process for stan-dard H&E stain. Most tissues where haemolympis present may be infected, however, principalsites include the lymphoid organ (Oka organ)(Fig.C.2.3.1.4a,b), hepatopancreatic interstitialcells (not tubule epithelial cells), heart, midgutmuscle and connective tissue (but not epithelialcells), stomach sub-cuticulum and gill tissues(Fig.C.2.3.1.4c). Light microscopy should revealmoderate to large numbers of deeply baso-philic, evenly stained, spherical, cytoplasmic in-clusions, approximately 2 mm in diameter(smaller in ectodermal and mesodermal tis-sues). Moribund shrimp show systemic necro-sis of gill and stomach sub-cuticular cells, with
2 If more rapid results are required, fixation can be shortened to 2 hours by substituting the acetic acid component of Davidson’sfixative with 50% concentrated HCl (this should be stored no more than a few days before use). After fixation, wash thoroughlyand check that the pH has returned to near neutral before staining. Do not fix for longer periods or above 25oC as this may resultin excessive tissue damage that will make interpretation difficult or impossible.
C.2 Yellowhead Disease (YHD)
170
intense basophilic cytoplasmic inclusions (H&Estaining) due to phagocytosed nuclei and viralinclusions. In the lymphoid organ, high num-bers of karyorrhexic and pyknotic basophilicinclusions are found in matrix cells of the nor-mal tubules. On the other hand, similar inclu-sions– are found only in lymphoid organ sphe-roids with Rhabdovirus of Penaeid Shrimp (RPS)described from Hawaii and Lymphoidal Parvo-like Virus (LPV, LOV) described from Australia;Lymphoid Organ Vacuolisation Virus (LOVV) inP. vannamei in Hawaii and the Americas; andTaura Syndrome Virus (TSV) in P. vannamei, P.stylirostris and P. setiferus from central andsouth America. Gill Associated Virus (GAV) inAustralian P. monodon; a Yellow-Head-Disease-Like Virus (YHDLV) in P. japonicus from TaiwanProvince of China produce similar histopathol-ogy to YHV.
C.2.4.2 Confirmatory
In cases where results from presumptive screen-ing indicate possible YHD infection, but confir-mation of the infectious agent is required (e.g.,first time finding or presence of other pathogenicfactors), bioassay (see C.2.4.2.1), electron mi-croscopy (see C.2.4.2.2) and molecular tech-niques (see C.2.4.2.3-5) are required.
C.2.4.2.1 Bioassay (Levels I-II)
The simplest bioassay method is to allow na?veshrimp (± 10 g wet weight) to feed on carcassesof suspect shrimp. Alternatively, preparehomogenates of gill tissues from suspect shrimp.Centrifuge solids into a loose pellet, decant andfilter (0.45 - 0.22 mm) the supernatant. Exposena?ve juvenile Penaeus monodon (± 10 g wetweight) to the supernatant Infected shrimp shouldevoke clinical signs in the na?ve shrimp within24-72 hours and 100% mortality will generallyoccur within 3-5 days. Infections should be con-firmed by histology of gills and haemolymph.
C.2.4.2.2 Transmission Electron Micros-copy (TEM) (Level III)
For TEM, the most suitable tissues of moribundshrimp suspected to be infected by YHD arethe lymphoid organ and gills. Fix tissues in 2.5%glutaraldehyde, 2% paraformaldehyde in ca-codylate buffer and post-fix in 1% osmiumtetroxide, prior to dehydration and embeddingin Spurr’s resin. 50nm sections are mounted onCu-200 grids and should be stained with ura-nyl acetate/70% methanol and Reynold’s leadcitrate. Diagnosis of YHV is confirmed by the
presence of non-occluded, enveloped, rod-shaped particles, 150-200 x 40-50 nm in sizein the perinuclear or cytoplasmic area of thetarget tissues or within cytoplasmic vesicles.Non-enveloped, filamentous forms measuring<800 nm may also be found in the cytoplasm.The cytoplasm of infected cells becomes frag-mented and breaks down within 32 hr of infec-tion.
C.2.4.2.3 Western Blot Assay (Level III)
Remove 0.1 ml of haemolymph from live YHD-suspected shrimp and dilute with 0.1 ml of cit-rate buffer for immediate use or store at -80oCuntil examination. A purified viral preparation isrequired as a positive control, and confirmationis made on the presence of 4 major protein bandscharacteristic of YHV at 135 and 175 kDa. Thesensitivity of the Western blot assay is 0.4 ngof YHV protein.
RT-PCR can be conducted on the haemolymphof suspect shrimp or on post-larvae (seeC.2.3.2.1). There are several commercially avail-able RT-PCR kits now available to screenhaemolymph from broodstock shrimp and PL tis-sues for evidence of YHV RNA.
C.2.4.2.5 In situ Nucleic Acid Hybridiza-tion (Level III)
Commercial in situ hybridization kits for YHD arenow available.
C.2.5 Modes of Transmission
Infections are generally believed to be horizon-tally transmitted. Survivors of YHD infection, how-ever, maintain chronic sub-clinical infections andvertical transmission is suspected with such in-dividuals. There are a number of known or sus-pected carrier crustaceans including the brack-ish water shrimp, Palaemon styliferus and Acetessp., which can potentially transmit YHD to farmedshrimp.
C.2.6 Control Measures
There are no known treatments for shrimp in-fected with YHV. However, a number of pre-ventative measures are recommended to re-duce spread. These include the following:
• broodstock specimens be screened for YHV
C.2 Yellowhead Disease (YHD)
171
• infected individuals and their offspring bedestroyed in a sanitary manner
• associated equipment and rearing water aredisinfected
• exclude potential carriers of YHD by screen-ing PL pre-stocking in ponds
• prevention of exposure to potential carriers,post-stocking, can be achieved by filtrationor prior treatment in storage ponds of waterused for water exchanges.
• avoidance of rapid changes in pH or pro-longed periods of low (<2ppm) dissolvedoxygen. These can trigger sub-lethal out-breaks of YHD. Alkalinity should not varymore than 0.5 pH units daily and water pHlevels > 9 should be avoided. Changes in sa-linity apparently do not trigger outbreaks.
• avoid fresh aquatic feeds in grow-out ponds,maturation units and hatchery facilities, un-less the feed is subjected to prior steriliza-tion (gamma radiation) or pasteurization (i.e.,holding at 7˚C for 10 min).
If an outbreak occurs, it is recommended that theaffected pond be treated with 30 ppm chlorine tokill the shrimp and potential carriers. The deadshrimp and other animals should be removed andburied or burned. If they cannot be removed, thepond should be thoroughly dried before re-stocking.
If the outbreak pond can be emergency har-vested, the discharge water should be pumpedinto an adjacent pond for disinfection with chlo-rine and holding for a minimum of 4 days beforedischarge. All other waste materials should beburied or burned. Harvesting personnel shouldchange clothing and shower at the site with wa-ter that will be discharged into the treatment pond.Clothing used during harvesting should be placedin a specific container to be sent for chlorine treat-ment and laundering. Equipment, vehicles andrubber boots and the outside of shrimp contain-ers should be disinfected with chlorine and thedischarge water run into the treatment pond.Neighbours should be notified of any YHD out-break and control efforts, and advised not carryout any water exchange for at least 4 days fol-lowing discharge from the pond used for disin-fection. Processing plants receiving emergencyharvested shrimp should be notified that thespecific lot of shrimp is YHV infected and ap-propriate measures should be taken at the plantto avoid transfer of the disease via transportcontainers and processing wastes. Prohibitionof introduction of living shrimp from YHV andGAV enzootic areas into historically uninfectedareas is recommended.
C.2.6 Selected References
Khanobdee, K., C. Soowannayan, T.W. Flegel,S. Ubol, and B. Withyachumnarnkul. 2001.Evidence for apoptosis correlated with mor-tality in the giant black tiger shrimp Penaeusmonodon infected with yellow head virus.Dis. Aquat. Org. (in press).
Lightner, D.V. 1996. A Handbook of Shrimp Pa-thology and Diagnostic Procedures for Dis-ease of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Loh, P.C., E.C.B. Nadala, Jr., L.M. Tapay, andY. Lu. 1998. Recent developments in im-munologically-based and cell culture proto-cols for the specific detection of shrimp viralpathogens, pp. 255-259. In: Flegel T.W. (ed)Advances in Shrimp Biotechnology. NationalCenter for Genetic Engineering and Biotech-nology, Bangkok, Thailand.
Lu, Y., L.M. Tapay, and P.C. Loh. 1996. Devel-opment of a nitrocellullose-enzyme immu-noassay for the detection of yellow-head vi-rus from penaeid shrimp. J. Fish Dis. 19(1):9-13.
Nadala, E.C.B. Jr., L.M. Tapay, S. Cao, and P.C.Loh. 1997. Detection of yellowhead virus andChinese baculovirus in penaeid shrimp by thewestern blot technique. J. Virol. Meth.69(1-2): 39-44.
OIE. 1999. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 35p.
OIE. 2000a. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
OIE. 2000b. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 40p.
Spann, K.M., J.E. Vickers, and R.J.G. Lester.1995. Lymphoid organ virus of Penaeusmonodon from Australia. Dis. Aquat. Org.23(2): 127-134.
Spann, K.M., J.A. Cowley, P.J. Walker, andR.J.G. Lester. 1997. A yellow-head-like virusfrom Penaeus monodon cultured in Austra-lia. Dis.Aquat. Org. 31(3): 169-179.
C.2 Yellowhead Disease (YHD)
172
Wang, C.S., K.F.J.Tang, G.H. Kou, S.N. Chen.1996. Yellow head disease like virus infec-tion in the Kuruma shrimp Peneaus japonicuscultured in Taiwan. Fish Pathol. 31(4): 177-182.
Wongteerasupaya, C., V. Boonsaeng, S.P a n y i m , A . T a s s a n a k a j o n ,B.Withyachumnarnkul, and T.W. Flegel.1997. Detection of yellow-head virus (YHV)of Penaeus monodon by RT-PCR amplifica-tion. Dis. Aquat. Org. 31(3): 181-186.
C.2 Yellowhead Disease (YHD)
173
C.3.1 Background Information
C.3.1.1 Causative Agent
Infectious Hypodermal and Hematopoietic Necro-sis (IHHN) is caused by a non-enveloped icosa-hedral virus, Infectious Hypodermal and Hemato-poietic Necrosis Virus (IHHNV), averaging 22 nmin diameter, with a density of 1.40 g/ml in CsCl,containing linear ssDNA with an estimated sizeof 4.1 kb, and a capsid that has four polypep-tides with molecular weights of 74, 47, 39, and37.5 kD. Because of these characteristics,IHHNV has been classified as a member of thefamily Parvoviridae. More detailed informationabout the disease can be found at OIE Diagnos-tic Manual for Aquatic Animal Diseases (OIE2000a) and Lightner (1996).
C.3.1.2 Host Range
IHHNV infects a wide range of penaeid shrimps,but does not appear to infect other decapod crus-taceans. Natural infections have been reportedin Penaeus vannamei, P. stylirostris, P.occidentalis, P. monodon, P. semisulcatus, P.californiensis and P. japonicus. Experimental in-fections have also been reported for P. setiferus,P. aztecus and P. duorarum. Penaeus indicusand P. merguiensis appear to be refractory toIHHNV infection.
C.3.1.3 Geographic Distribution
IHHN occurs in wild and cultured penaeid shrimpsin Central America, Ecuador, India, Indonesia,Malaysia, Philippines, Peru, Taiwan Province ofChina, and Thailand. Although IHHNV has beenreported from cultured penaeid shrimp from mostregions of the western hemisphere and in wildpenaeids throughout their geographic range alongthe Pacific coast of the Americas (Peru to north-ern Mexico), it has not been found in penaeidson the Atlantic side of the Americas. IHHNV hasbeen reported in cultured penaeid shrimp fromGuam, French Polynesia, Hawaii, Israel and NewCaledonia. An IHHN-like virus has also been re-ported from Australia.
C.3.1.4 Asia-Pacific Quarterly Aquatic Ani-mal Disease reporting System (1999-2000)
The disease was suspected in India during the2nd quarter reporting period for 1999 and 1stquarter reporting period for 2000 (OIE 1999, OIE2000b).
Penaeus stylirostris. Infection by IHHNV causesacute epizootics and mass mortality (> 90%) inP. stylirostris. Although vertically infected lar-vae and early postlarvae do not become dis-eased, juveniles >35 days old appear suscep-tible showing gross signs followed by massmortalities. In horizontally infected juveniles, theincubation period and severity of the diseaseappears size and/or age dependent, with youngjuveniles always being the most severely af-fected (Fig.C.3.2a). Infected adults seldomshow signs of the disease or mortalities.
Penaeus vannamei. The chronic disease, “runtdeformity syndrome” (RDS) (Fig.C.3.2b,c), iscaused by IHHNV infection of P. vannamei. Ju-veniles with RDS show wide ranges of sizes,with many smaller than average (“runted”)shrimp. Size variations typically exceed 30%from the mean size and may reach 90%.Uninfected populations of juvenile P. vannameiusually show size variations of < 30% of themean. Similar RDS signs have been observedin cultured P. stylirostris.
C.3.3 Screening Methods
More detailed information on methods forscreening IHHN can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000a), at http://www.oie.int, or at selected ref-erences.
C.3.3.1 Presumptive
There are no gross signs (Level I) or histologi-cal features (Level II) that can be used to indi-cate presumptive infection by IHHNV in sub-clinical carriers.
C.3.3.2 Confirmatory
Molecular methods are required to detectIHHNV in sub-clinical carriers.
C.3.3.2.1 Dot Blot Hybridization (Level III)
Haemolymph samples or a small appendage(pleiopod) can be used for dot blot testing.Commercial dot blot hybridization kits for IHHNare now available.
174
(DV Lightner)
Fig.C.3.2a. A small juvenile Penaeus stylirostrisshowing gross signs of acute IHHN disease.Visible through the cuticle, especially on the ab-domen, are multifocal white to buff colored le-sions in the cuticular epithelium or subcutis (ar-rows). While such lesions are common in P.stylirostris with acute terminal IHHN disease,they are not pathognomonic for IHHN disease.
(DV Lightner)
Fig.C.3.2b. Dorsal view of juvenile P. vannamei(preserved in Davidson’s AFA) showing grosssigns of IHHNV-caused RDS. Cuticular abnor-malities of the sixth abdominal segment andtail fan are illustrated.
Fig.C.3.4.1.2a. A high magnification of gillsshowing eosinophilic intranuclear inclusions(Cowdry type A inclusions or CAIs) that arepathognomonic for IHHNV infections. Mayer-Bennett H&E. 1800x magnification.
Fig.C.3.2c.Lateral viewof juvenile P.v a n n a m e i(preserved inDav idson ’sAFA) showinggross signs ofI H H N V -caused RDS.Cuticular ab-normalities ofthe sixth ab-dominal seg-ment and tailfan are illus-trated.
(DV Lightner)
(DV Lightner)
Fig.C.3.4.1.2b. A low magnification photomi-crograph (LM) of an H&E stained section of ajuvenile P. stylirostris with severe acute IHHNdisease. This section is through the cuticularepithelium and subcuticular connective tissuesjust dorsal and posterior to the heart. Numer-ous necrotic cells with pyknotic nuclei or withpathognomonic eosinophilic intranuclear inclu-sion bodies (Cowdry type A) are present (ar-rows). Mayer-Bennett H&E. 830x magnification.
H&E stain) intranuclear, Cowdry type A inclu-sion bodies (CAIs) provide a presumptive diag-nosis of IHHNV infection. Infected nuclei areenlarged with a central eosinophilic inclusionsometimes separated from the marginatedchromatin by an unstained ringwhen tissues arepreserved with acetic acid containing fixatives.Since IHHNV intranuclear inclusion bodies canbe confused with developing intranuclear in-clusion bodies due to White Spot Disease, elec-tron microscopy (C.3.4.2.2) or in situ hybridiza-tion assays of suspect sections with IHHNV-specific DNA probes (C.3.4.2.3-5) may be re-quired for definitive diagnosis. Basophilicstrands may be visible within the CAIs and cy-toplasmic inclusion bodies may also be present.
C.3.4.2 Confirmatory
C.3.4.2.1 Bioassay (Levels I/II)
Prevalence and severity of IHHNV infectionsmay be “enhanced” in a quarantined popula-tion by holding the suspect shrimp in crowdedor other stressful conditions (low dissolved oxy-gen, elevated water temperature, or elevatedammonia or nitrite). These conditions may en-courage expression of low grade IHHNV infec-tions and transmission from sub-clinical carri-ers to uninfected shrimp. This increase in preva-lence and severity can enhance detection us-ing screening methods.
Indicator shrimp (0.1-4.0 gm juvenile P.stylirostris) can also be used to assess the pres-ence of IHHNV by cohabitation, feeding ofminced carcasses or injection with cell-freehomogenates from suspect shrimp.
C.3.4.2.2 Transmission Electron Micros-copy (TEM) (Level III)
Negative stain preparations of purified virusshow non-enveloped, icosahedral virions, 20-22 nm in diameter. Transmission electron mi-croscopic preparations show intranuclear inclu-sions containing virions 17-26 nm in diameter.Viral particles are also present in the cytoplasmwhere they assemble and replicate. Chromatinstrands (that may be visible as basophilic in-clusions under light microscopy) are a promi-nent feature of IHHNV intranuclear inclusionbodies. Paracrystalline arrays of virions corre-spond to cytoplasmic inclusion bodies that maybe detected under light microscopy.
The same tissue samples described in C.3.3.2.1can be used for non-lethal screening of non-clinical broodstock and juveniles of susceptiblespecies, using PCR.
C.3.4 Diagnostic Methods
More detailed information on methods for di-agnosis of IHHN can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000b), at http://www.oie.int, or at selected ref-erences.
C.3.4.1 Presumptive
C.3.4.1.1 Gross Observations (Level I)
Gross signs are not IHHN specific. Acute in-fections of juvenile P. stylirostris may result in amarked reduction in food consumption, fol-lowed by changes in behaviour and appear-ance. The shrimp may rise slowly to the watersurface, become motionless and then roll-over,and slowly sink (ventral side up) to the bottom.This behavior may continue for several hoursuntil the shrimp become too weak to continue,or are cannibalised by healthier siblings. By thisstage of infection white or buff-coloured spots(which differ from the white spots that occur inWSD - C.4) in the cuticular epidermis, espe-cially at the junction of the abdominal tergalplates, resulting in a mottled appearance. Thismottling may later fade in P. stylirostris. Mori-bund P. stylirostris may further develop a dis-tinctly bluish colour and opaque abdominalmusculature. Although P. monodon is frequentlyfound to be infected with IHHNV, it does notgenerally appear to cause any major clinicaldisease in the species. Juvenile shrimp (P.vannamei and P. stylirostris) with RDS displaybent or deformed rostrums, wrinkled antennalflagella, cuticular roughness, and other cuticu-lar deformities. They also show a high percent-age (30-90%) of stunted growth (“runt shrimp”)compared with less than 30% below averagesize in uninfected populations.
C.3.4.1.2 Histopathology (Level II)
Infected cells occur in the gills (Fig.C.3.4.1.2a),epidermal (Fig.C.3.4.1.2b) and hypodermal epi-thelia of fore and hindgut, nerve cord and nerveganglia, as well as mesodermal haematopoieticorgans, antennal gland, gonads, lymphoid or-gan, and connective tissue. Eosinophilic (with
IHHNV-specific DNA probes are now availablefor in situ hybridization confirmation of histo-logical and/or electron microscopic observa-tion.
C.3.5 Modes of Transmission
Some members of populations of P. stylirostrisand P. vannamei, which survive IHHNV infec-tions and/or epizootics, may carry sub-clinicalinfections for life which may be passed hori-zontally to other stocks, or vertically, if used asbroodstock.
C.3.6 Control Measures
Eradication methods for IHHNV can be appliedto certain aquaculture situations. These meth-ods are dependent upon eradication of infectedstocks, disinfection of the culture facility, avoid-ance of re-introduction of the virus (from othernearby culture facilities, wild shrimp, etc.), andre-stocking with IHHNV-free post-larvae thathave been produced from IHHNV-freebroodstock.
C.3.7 Selected References
Bell, T.A. and D.V. Lightner, D.V. 1984. IHHN vi-rus: Infectivity and pathogenicity studies inPenaeus stylirostris and Penaeus vannamei.Aquac. 38: 185-194.
Bray, W.A., A.L. Lawrence, and J.R. Leung-Trujillo. 1994. The effect of salinity on growthand survival of Penaeus vannamei, with ob-servations on the interaction of IHHN virusand salinity. Aquac. 122(2-3): 133-146.
Browdy, C.L., J.D. Holloway, Jr., C.O. King, A.D.Stokes, J.S. Hopkins, and P.A. Sandifer. 1993.IHHN virus and intensive culture of Penaeusvannamei: Effects of stocking density andwater exchange rates. Crus. Biol.13(1): 87-94.
Carr, W.H., J.N. Sweeney, L. Nunan, D.V.Lightner, H.H. Hirsch, and J.J. Reddington.1996. The use of an infectious hypodermaland hematopoietic necrosis virus gene probeserodiagnostic field kit for screening of can-didate specific pathogen-free Penaeusvannamei broodstock. Aquac.147(1-2): 1-8.
Castille, F.L., T.M. Samocha, A.L. Lawrence, H.He, P. Frelier, and F. Jaenike. 1993. Variabil-ity in growth and survival of early postlarvalshrimp (Penaeus vannamei Boone 1931).Aquac. 113(1-2): 65-81.
Karunasagar, I. and I. Karunasagar. 1996.Shrimp diseases and control. AquacultureFoundation of India, Madras, India 1996: 63-67
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDisease of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Lu, Y., P.C. Loh, and J.A. Brock. 1989. Isola-tion, purification and characterisation of
infectious hypodermal and hematopoietic ne-crosis virus (IHHNV) from penaeid shrimp. J.
Virol. Meth. 26: 339-344.
Mari, J., J.R. Bonami, and D.V. Lightner. 1993.Partial cloning of the genome of infectioushypodermal and hematopoietic necrosis vi-rus, an unusual parvovirus pathogenic forpenaeid shrimps - diagnosis of the diseaseusing a specific probe. J. Gen. Vir.74(12):2637-2643.
Nunan, L.M., B. Poulos, and D.V. Lightner. 1994.Detection of the infectious hypodermal andhematopoietic necrosis virus (IHHNV) inPenaeus shrimp tissue homogenate andhemolymph using polymerase chain reaction(PCR). International Symposium on AquaticAnimal Health: Program and Abstracts. Uni-versity of California, School of VeterinaryMedicine, Davis, CA, USA. 1994: P-62.
OIE. 1999. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 35p.
OIE. 2000a. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
OIE. 2000b. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 40p.
Owens, L., I.G. Anderson, M. Kenway, L. Trott,and J.A.H. Benzie. 1992. Infectious hypoder-mal and haematopoietic necrosis virus(IHHNV) in a hybrid penaeid prawn from tropi-cal Australia. Dis. Aquat. Org. 14: 219-228.
Poulos, B.T., D.V. Lightner, B. Trumper, and J.R.Bonami. 1994. Monoclonal antibodies to apenaeid shrimp parvovirus, infectious hypo-dermal and hematopoeitic necrosis virus(IHHNV). J. Aquat. Anim. Health 6(2): 149-154.
The causative agent of white spot disease(WSD) is the white spot syndrome virus (WSSV)or white spot virus (WSV), a double strandedDNA (dsDNA) virus. In initial reports, WSV wasdescribed as a non-occluded baculovirus butsubsequent analysis of WSV-DNA sequencesdoes not support this contention. The virusesin this complex have recently been shown tocomprise a new group with the proposed nameof Nimaviridae (Van Hulten et al. 2001). In theliterature, however, several names have beenused to describe the virus, including baculoviralhypodermal and haematopoietic necrosis(HHNBV), Shrimp Explosive Epidemic Disease(SEED), China virus disease, rod-shapednuclear virus of Penaeus japonicus (RV-PJ);systemic ectodermal and mesodermalbaculovirus (SEMBV), white spot baculovirus(WSBV) and white spot syndrome virus (WSSV).More detailed information about the diseasecan be found in the OIE Manual for AquaticAnimal Diseases (OIE 2000a) and Lightner(1996).
C.4.1.2 Host Range
White spot disease has a wide spectrum of hosts.Outbreaks were first reported from farmedPenaeus japonicus in Japan and natural infec-tions have subsequently been observed in P.chinensis, P. indicus, P. merguiensis, P. monodon,P. setiferus, P. stylirostris, and P. vannamei. Inexperimental studies, WSD is also lethal to P.aztecus, P. duodarum and P. setiferus.
C.4.1.3 Geographic Distribution
WSD was first reported in Taiwan Province ofChina and China mainland between 1991-1992,and in Japan in 1993 from shrimp imported fromChina PR. Later outbreaks have been reportedfrom elsewhere in Asia including China PR, In-dia, Indonesia, Korea RO, Malaysia, TaiwanProvince of China, Thailand, and Vietnam. Inaddition to the Asian countries listed above,farmed shrimp exhibiting the gross signs and his-tology of WSD have been reported in the USAand Latin America.
As of 1999, WSD has been reported in at leastnine countries in the Americas: Columbia, Ec-uador, Guatemala, Honduras, Mexico, Nicara-
C.4 WHITE SPOT DISEASE (WSD)3
3 White spot disease (WSD) is now classified as an OIE Notifiable Disease (OIE 2000a).
gua, Panama, Peru and USA (Subasinghe etal. 2001).
C.4.1.4 Asia-Pacific Quarterly AquaticAnimal Disease Reporting System (1999-2000)
WSD was reported by Bangladesh, China PR,India, Indonesia, Japan, Korea RO, Malaysia,Philippines, Taiwan Province of China, SriLanka, and Thailand; and suspected in Paki-stan during the reporting period for the year1999. In year 2000, Bangladesh, India, Japan,Korea RO, Malaysia, Philippines, Sri Lanka,Thailand and Vietnam reported positive occur-rence of the disease (NACA/FAO 2000a,b,c; OIE1999, OIE 2000a,b).
C.4.2 Clinical Aspects
WSD outbeaks are often characterised by highand rapid mortality of infected populations,usually shortly after the first appearance of theclinical signs. Acutely affected shrimp demon-strate anorexia and lethargy, have a loose cu-ticle with numerous white spots (about 0.5 to2.0 mm in diameter) on the inside surface ofthe carapace (Fig.C.4.2a,b). These spots arewithin the cuticle structure and cannot be re-moved by scraping. Moribund shrimp may alsoshow a pink to red discolouration. Susceptibleshrimp species displaying these clinical signsare likely to undergo high levels of mortality.Pathology is associated with systemic destruc-tion of the ectodermal and mesodermal tissuesof the gills and sub-cuticular tissues.
C.4.3 Screening Methods
More detailed information on methods forscreening for WSD can be found in the OIE Di-agnostic Manual for Aquatic Animal Diseases(OIE 2000a), at http://www.oie.int, or in selectedreferences.
C.4.3.1 Presumptive
There are no gross observations (Level I) or his-topathological (Level II) diagnostic techniqueswhich can provide presumptive detection ofWSD in sub-clinical shrimp.
179
C.4.3.2 Confirmatory
C.4.3.2.1 Nested PCR of Tissues andHaemolymph (Level III)
The protocol described by Lo et al (1996, 1998)is the recommended procedure for nested PCRof tissues and haemolymph. There are alsocommercially available kits for detection of WSDin sub-clinical carriers using PCR-based tech-niques.
C.4.3.2.2 Polymerase Chain Reaction(PCR) of Postlarvae (Level III)
From a nursery or hatchery tank containing 100000 postlarvae (PL) or more, sample approxi-mately 1000 PL from each of 5 different points.
C.4 White Spot Disease (WSD)
(DV Lightner)
Fig.C.4.2a. A juvenile P. monodon with distinc-tive white spots of WSD.
(DV Lightner/P. Saibaba)
Fig.C.4.2b. Carapace from a juvenile P.monodon with WSD. Calcareous deposits onthe underside of the shell account for the whitespots.
(DV Lightner)
Fig.C.4.3.3.1.2a. Histological section from thestomach of a juvenile P.chinensis infected withWSD. Prominent intranuclear inclusion bodiesare abundant in the cuticular epithelium andsubcuticular connective tissue of the organ (ar-rows).
(DV Lightner)
Fig.C.4.3.3.1.2b. Section of the gills from a ju-venile P. chinensis with WSBV. Infected cellsshow developing and fully developed intra-nuclear inclusion bodies of WSBV (arrows).Mayer-Bennett H&E. 900x magnification.
Pool the samples in a basin, gently swirl thewater and select an assay sample from livingPL collected at the center of the basin. A sampleof 150 PL is required to give a 95% confidenceof detecting an infection at 2% prevalence inthe population (see Table C.1.3.3 of C.1 Gen-eral Techniques).
For PL 11 and older, exclude shrimp eyes fromany tissue samples, since these inhibit the PCRprocess. Follow the procedures from the rec-ommended source for nested PCR given un-der C.4.3.2.1.
180
to 2 hrs by changing the acetic acid in theDavidson’s fixative to 50% concentrated HCl(this should not be stored longer than a few daysbefore use). After fixation, wash the tissues thor-oughly and ensure pH is near neutral beforestaining. Do not fix for longer periods, or above25oC, as this can cause tissue damage that willmake interpretation difficult or impossible. Stainwith Meyer’s H&E and dehydrate to xylene (orequivalent clearing solution). Place a gill fila-ment on a microscope slide tease off severalsecondary filaments. Replace the main filamentin a sealed vial filled with xylene as a perma-nent back-up reference. Being careful not tolet the secondary gill filaments dry, tease apartand remove any large fragments or particlesfrom the slide. Add a drop of mounting fluidand a cover glass, using light pressure to flat-ten the tissue as much as possible. The sameprocedure can be used for thin layers of sub-cuticular tissue.
Examine under a compound microscope at 40xmagnification for moderate to large numbersof hypertrophied nuclei with basophilic, cen-trally-positioned, inclusions surrounded bymarginated chromatin. The whole mount slidescan also be kept as permanent records.
C.4.4.1.3 Histopathology (Level II)
Moribund shrimp from a suspected WSD out-break should be fixed in Davidson’s fixative andstained with haematoxylin and eosin (H&E). Thehistopathology of WSD is distinctive, and canprovide a conclusive diagnosis. However, firsttime detection or detection in species not pre-viously reported to be susceptible, requiremolecular assay or electron microscopy dem-onstration of a viral aetiology.
Moribund shrimp with WSV show systemicdestruction of ectodermal and mesodermal tis-sues. Nuclei of infected cells are hypertrophiedand when stained with haematoxylin and eosinshow lightly to deeply basophilic central inclu-sions surrounded by marginated chromatin.These intranuclear inclusions can also be seenin squash mounts of gills or sub-cuticular tis-sue (see C.4.4.1.2), or in tissue sections. Thebest tissues for examination are the subcuticu-lar tissue of the stomach (Fig.C.4.3.3.1.2a),cephalothorax or gill tissues (Fig.C.4.3.3.1.2b).
C.4.4.2 Confirmatory
A definitive diagnosis can be accomplished bypolymerase chain reaction (PCR) technology
C.4.3.2.3 Dot Blot Hybridization (Level III)
Details on dot blot hybridisation techniques anddetection kit availability are provided in the OIEDiagnostic Manual (OIE 2000a).
C.4.3.2.4 In situ Hybridization (Level III)
Details on in situ hybridization techniques anddetection kit availability are provided in the OIEDiagnostic Manual (OIE 2000a).
C.4.4 Diagnostic Methods
More detailed information on methods for di-agnosis of WSD can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000a), at http://www.oie.int, or in selected ref-erences.
C.4.4.1 Presumptive
C.4.4.1.1 Gross Observations (Level I)
WSD outbreaks are generally preceded by ces-sation of feeding followed, within a few days,by the appearance of moribund shrimp swim-ming near the surface at the edge of rearingponds. These shrimp exhibit white inclusionsembedded in the cuticle and often show red-dish discolouration of the body. The cuticularinclusions range from minute spots to discsseveral mm in diameter that may coalesce intolarger plaques. They are most easily observedby removing the cuticle from the cephalotho-rax, scraping away any attached tissue andholding the cuticle up to the light. The appear-ance of white spots in the cuticle can be causedby other conditions. In particular, Wang et al.,2000, report a condition called bacterial whitespot syndrome (BWSS) which can easily bemistaken for WSD (see C.4a). Therefore, histo-pathological examination is required for confir-matory diagnosis.
C.4.4.1.2 Rapid Squash Mount Prepara-tions (Level II)
Two types of rapid squash mount preparationsthat can be used for presumptive diagnosis ofWSD: i) fresh, unstained wet mounts fixed in10% formalin solution and viewed by dark fieldmicroscopy with a wet-type condenser, and ii)fixed tissues stained with H&E.
For method ii) fix whole shrimp or gill filamentsin Davidson’s fixative overnight. If more rapidresults are required, fixation can be shortened
C.4 White Spot Disease (WSD)
181
(single-step or nested), in situ hybridization,Western blot analysis (detailed protocols canbe found in OIE (2000a) or electron microscopy(TEM).
C.4.4.2.5 Transmission Electron Micros-copy (TEM) (Level III)
The most suitable tissues for TEM examinationare subcuticular tissues, gills and pereiopodsthat have been pre-screened by histology(C.4.4.1.3) or rapid-stain tissue squashes(C.4.4.1.2) which show signs of hypertrophiednuclei with Cowdry A-type inclusions or mar-ginated chromatin surrounding a basophilic in-clusion body. Fix tissues for at least 24h in a10:1 fixative to tissue volume ration of 6%gluteraldehyde at 4°C and buffered with sodiumcacodylate or phosphate solution to pH7. Forlonger term storage, reduce gluteraldehyde to0.5-1.0% concentration. Post-fix in 1% osmiumtetroxide, and stain with uranyl acetate and leadcitrate (or equivalent TEM stain). WSD virionsare rod-shaped to elliptical with a trilaminarenvelope and measure 80-120 x 250-380 nm.
C.4.4.2.6 Negative Stain Electron Micros-copy (Level III)
Negative stain preparations from shrimphaemolymph may show virions with unique, tail-like appendages within the hypertrophied nu-clei of infected cells, but no evidence of occlu-sion bodies.
C.4.5 Modes of Transmission
Wild broodstock and fry used to stock rearingponds are known to carry WSV, as are numer-ous other crustaceans and even aquatic insectlarvae. Molecular techniques have been usedto confirm infection of non-penaeid carriers ofWSV and transmission studies show that thesecan transmit WSV to shrimp.
C.4.6 Control Measures
There are no known treatments for shrimp in-fected with WSV, however, a number of pre-ventative measures are recommended to re-duce spread.
At facilities used for the production of PL, it isrecommended that wild broodstock bescreened for WSD by nested PCR. Any infectedindividuals, and their offspring, should be de-stroyed in a sanitary manner and all contami-nated equipment and rearing water be disin-
fected. It is also recommended that broodstockP. monodon be tested for WSD after spawningto increase the probability of viral detection.
At grow-out, PL should be screened for free-dom from WSV by nested PCR using sufficientlylarge numbers of PL to ensure detection of sig-nificant infections. A biased sampling regime,which selects weaker animals for testing, canfurther increase the probability of detecting in-fected batches.
During cultivation, it is suspected that rapidchanges in water temperature, hardness andsalinity, or reduced oxygen levels (<2 ppm) forextended periods, can trigger outbreaks ofWSD in shrimp with sub-clinical infections. It isnot yet known whether large diurnal pH changescan trigger outbreaks but stable pond-water pHis known to reduce general stress levels inshrimp. Fresh or fresh-frozen feeds of aquaticanimal origin should not be used in the grow-out ponds, maturation units and hatchery fa-cilities unless subjected to prior sterilization(gamma radiation) or pasteurization (i.e., hold-ing at 70°C for 10 min).
Any affected ponds should be treated immedi-ately with 30 ppm chlorine to kill the infectedshrimp and any potential carriers. The deadshrimp and other animals should be removedand buried or burned. The water should thenbe held for a minimum of 4 days before dis-charge. Neighbouring pond owners should beimmediately informed and should not carry outwater exchange for a minimum of 4 days afterwater is discharged from an outbreak pond if itis likely to come into contact with their ownsupply water.
If the outbreak pond is emergency harvested,the discharge water should be pumped into anadjacent pond or reservoir for disinfection withchlorine and holding for a minimum of 4 daysbefore discharge. All water from the harvestedpond should be discharged into the treatmentpond and any waste materials should be bur-ied or burned. Harvesting personnel shouldchange clothing and shower at the site withwater that will be discharged into the treatmentpond. Clothing used during harvesting shouldbe placed in a specific container to be sent fordisinfection and laundering. Equipment, ve-hicles, footwear and the outside of shrimp con-tainers should be disinfected and the wastewater discarded into the treatment pond. Theprocessing plant should be notified that thespecific lot of shrimp is WSD infected and ap-propriate measures should be taken at the plant
C.4 White Spot Disease (WSD)
182
to avoid transfer of the disease via transportcontainers and processing wastes. Preventionof introduction of live shrimp from WSV enzooticareas into historically uninfected areas or ar-eas defined as free from the disease is recom-mended.
C.4.7 Selected References
Chou, H.Y., C.Y. Huang, C.H. Wang, H.C.Chiang and C.F. Lo. 1995. Pathogenicity of abaculovirus infection causing white spot syn-drome in cultured penaeid shrimp in Taiwan.Dis. Aquat. Org. 23: 165-173.
Inouye, K, S. Miwa, N. Oseko, H. Nakano, T.Kimura, K. Momoyama and M. Hiraoka.1994. Mass mortalities of cultured Kurumashrimp Penaeus japonicus in Japan in 1993:electron microscopic evidence of the caus-ative virus. Fish Pathol. 29:149-158.
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDisease of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Lo, C.F., Y.S. Chang, C.T. Cheng, and G.H.Kou 1998. PCR monitoring of cultured shrimpfor white spot syndrome virus (WSSV) infec-tion in growout ponds. In: Flegel T.W. (ed)Advances in shrimp biotechnology, pp. 281-286. National Center for Genetic Engineer-ing and Biotechnology. Bangkok, Thailand.
Lo, C.F., J.H. Leu , C.H. Ho, C.H. Chen, S.E.Peng, Y.T. Chen, C.M. Chou, , P.Y. Yeh , C.J.Huang, H.Y. Chou, C.H. Wang, and G.K. Kou.1996. Detection of baculovirus associatedwith white spot syndrome (WSBV) in penaeidshrimps using polymerase chain reaction.Dis. Aquat. Org. 25: 133-141.
Network of Aquaculture Centres in Asia-Pacificand Food and Agriculture Organization of theUnited Nations. 2000a. Quarterly Aquatic Ani-mal Disease Report (Asia and Pacific Region),2000/1, January-March 2000. FAO ProjectTCP/RAS/6714. Bangkok, Thailand. 57p.
Network of Aquaculture Centres in Asia-Pacificand Food and Agriculture Organization of theUnited Nations. 2000b. Quarterly AquaticAnimal Disease Report (Asia and Pacific Re-gion), 2000/2, April-June 2000. FAO ProjectTCP/RAS/6714. Bangkok, Thailand. 59p.
Network of Aquaculture Centres in Asia-Pacificand Food and Agriculture Organization of theUnited Nations. 2000c. Quarterly Aquatic Ani-mal Disease Report (Asia and Pacific Region),2000/3, July-September 2000. FAO ProjectTCP/RAS/6714. Bangkok, Thailand. 57p.
OIE. 1999. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 35p.
OIE. 2000a. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
OIE. 2000b. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 40p.
Subasinghe, R.P., M.G. Bondad-Reantaso, andS.E. McGladdery. 2001. Aquaculture devel-opment, health and wealth. In: R.P.Subasinghe, P. Bueno, M.J. Phillips, C.Hough, S.E. McGladdery & J.R. Arthur, eds.Aquaculture in the Third Millennium. Techni-cal Proceedings of the Conference onAquaculture in the Third Millennium,Bangkok, Thailand, 20-25 February 2000.NACA, Bangkok and FAO, Rome. (in press)
Van Hulten, M.C. , J. Witteveldt, S. Peters, N.Kloosterboer, R. Tarchini, M. Fiers, H.Sandbrink, R.K. Lankhorst, and J.M. Vlak.2001 The white spot syndrome virus DNAgenome sequence. Virol. 286 (1):7-22.
Wang, C.H., C.F. Lo, J.H. Leu, C.M. Chou, M.C.Tung, C.F. Chang, M.S. Su and G.H. Kou.1995. Purification and genomic analysis ofbaculoviruses associated with white spotsyndrome (WSBV) of Penaeus monodon. Dis.Aquat. Org. 23:239-242.
Wongteerasupaya, C., J.E. Vickers, S.Sriurairatana, G.L. Nash, A. Akarajamorn, V.Boonsaeng, S. Panyim, A. Tassanakajon, B.Withyachumnarnkul and T.W. Flegel. 1995.A non-occluded, systemic baculovirus thatoccurs in cells of ectodermal and mesoder-mal origin and causes high mortality in theblack tiger prawn, Penaeus monodon. Dis.Aquat. Org. 21:69-77.
C.4 White Spot Disease (WSD)
183
Bacterial White Spot Syndrome (BWSS) is arecently described condition which affectsPenaeus monodon. It is, as yet, poorly under-stood condition and is included in the AsiaDiagnostic Guide due to the possibility of di-agnostic confusion with viral White Spot Dis-ease (WSD).
C.4a.1 Background Information
Since 1993, white spot disease virus (WSDV)has caused massive losses to the shrimp indus-try in Asia and Latin America. Recently, anotherdisease syndrome showing similar gross clinicalsigns of white spots, has been detected and re-ported as “bacterial white spot syndrome’’(BWSS) (Wang et al., 1999, 2000). The similargross clinical signs have also caused confusionduring PCR-based screening for WSD since,shrimp with apparent WSDV clinical signs, givenegative results. The clinical effects of BWSS,appear far less significant than those of WSDinfection, although it has been suggested thatsevere infections may reduce moulting andgrowth.
C.4a.1.1 Causative Agent(s)
The bacterium Bacillus subtilis has been sug-gested as the possible causative agent due toits association with the white spots (Wang et al.,2000) but no causal relationship has been dem-onstrated, nor have infectivity studies been con-ducted. Vibrio cholerae is also often isolated insignificant numbers and similar white spots havebeen described in farmed shrimp in Thailand asa result of exposure to high pH and alkalinity inponds in the absence of the White Spot virus orbacterial colonisation of the spots, indicating thatthe bacterial involvement may be secondary. Thelack of certainty as to the causative agent andthe possibility of secondary involvement of bac-teria needs to be addressed through further re-search. Until the bacterial etiology is clearly dem-onstrated, bacteria cannot be definitively re-garded as the causative agent.
C.4a.1.2 Host Range
To date, the syndrome has only been reported incultured Penaeus monodon.
C.4a.1.3 Geographic Distribution
BWSS was first detected from a shrimp (Penaeusmonodon) farm in Malaysia in 1998 (Wang et al.
C.4a BACTERIAL WHITE SPOTSYNDROME (BWSS)
1999, 2000). This remains the only confirmedreport of the condition.
C.4a.2 Clinical Aspects
Dull white spots are seen on the carapace andall over the body but are more noticeable whenthe cuticle is peeled away from the body. Thewhite spots are rounded and not as dense asthose seen in WSD (Fig.C.4a.2). Wet mountmicroscopy reveals the spots as opaque brown-ish lichen-like lesions with a crenellated margin(although this is also the case with spots in theearly stages of WSD and cannot be used as adistinctive diagnostic feature). The spot centeris often eroded and even perforated. During theearly stage of infection, shrimp are still active,feeding and able to moult – at which point thewhite spots may be lost. However, delayedmoulting, reduced growth and low mortalitieshave been reported in severely infected shrimp(Wang et al., 2000).
C.4a.3 Screening Methods
There are no reported methodologies availableto screen for sub-clinical infections, sinceBWSS appears to be an opportunistic infec-tion.
C.4a.4 Diagnostic Methods
C.4a.4.1 Presumptive
C.4a.4.1.1 Gross Observations (Level I)
The presence of white spots on shrimp cuticleswithout significant mortality.
C.4a.4.1.2 Wet Mounts (Level I)
If cuticular spots are detected in P. monodon,which show an opaque brownish lichen-likeappearance with a crenallated margin and thecenter shows signs of erosion and/or perfora-tion, along with extensive bacterial involvement,such infections could be attributable to BWSS.Such infections should be confirmed as beingnegative for WSD.
Negative WSDV-PCR results from samplesshowing gross clinical signs attributed to WSD,may be suggestive of the alternate aetiology ofBWSS.
184
C.4a Bacterial White SpotSyndrome (BWSS)
(M. Shariff)
Fig. C.4a.2. Penaeus monodon dense whitespots on the carapace induced by WSD.
(M. Shariff/ Wang et al. 2000 (DAO 41:9-18))
Fig. C.4a.4.2.2a, b. Bacterial white spots(BWS), which are less dense than virus-inducedwhite spots. Note some BWS have a distinctwhitish marginal ring and maybe with or with-out a pinpoint whitish dot in the center
(M. Shariff/ Wang et al. 2000 (DAO 41:9-18))
C.4a.4.2 Confirmatory
C.4a.4.2.1 Histopathology (Level II)
Histological examinations should be conductedto ensure that the soft-tissues associated withthe cuticular lesions do not show signs of theWSDV characteristic endodermal and mesoder-mal intranuclear inclusian bodies. In the caseof BWSS, bacteria will be the primary micro-bial foreign particle and this should be in pri-mary association with the cuticular lesionsthemselves.
C.4a.4.2.2 Scanning Electron Microscopy(TEM) (Level III)
The presence of spot lesions (Fig. C.4a.4.2.1a,b)together with numerous bacteria (Fig.C.4a.4.2.2c) under scanning electron micros-copy will confirm BWSS.
C.4a.5 Modes of Transmission
Since bacteria are only localized on the bodysurface, the mode of transmission is thoughtto be through the rearing water. However, thishas yet to be demonstrated using transmissionstudies.
C.4a.6 Control Measures
Although the exact aetiology is unknown, somemeasures may help to reduce the risk of BWSS.Build up of high bacterial density in rearingwater should be avoided. Changing water fre-quently is recommended . Indiscriminate useof probiotics containing Bacillus subtilis shouldalso be avoided until the relationship betweenthis bacteria and the BWSS syndrome is betterunderstood. It has been claimed that BWSS inshrimp ponds can be treated with quick lime(CaO) at 25 ppm, however, this is still under in-vestigation and the use of quicklime may itselfcause problems due to rapid increases in pond-water pH (see C.4.6).
Fig. C.4a.4.2.2c. Presence of large number ofbacteria attached to exposed fibrillar laminaeof the endocuticle.
>
b
a
185
C.4a.7 Selected References
Wang, Y.G., M. Shariff, K.L. Lee and M.D.Hassan. 1999. A review on diseases ofcultured shrimp in Malaysia. Paper waspresented at Workshop on Thematic Reviewon Management Strategies for MajorDiseases in Shrimp Aquaculture, 28-30November 1999, Cebu, Philippines. WB,NACA, WWF and FAO.
Wang, Y. G., K.L. Lee, M. Najiah, M. Shariffand M.D. Hassan. 2000. A new bacterialwhite spot syndrome (BWSS) in cultured tigershrimp Penaeus monodon and its compari-son with white spot syndrome (WSS) causedby virus. Dis. Aquat. Org. 41: 9-18.
C.4a Bacterial White SpotSyndrome (BWSS)
186
C.5.1 Background Information
C.5.1.1 Causative Agent
The pathogen responsible for Baculoviral Mid-gut Gland Necrosis (BMN) disease is Baculoviralmidgut gland necrosis virus (BMNV), a non-oc-cluded gut-infecting baculovirus, whose non-en-veloped nucleocapsid measures 36 by 250 nm;enveloped virions measures ~ 72 by ~ 310 nm.More detailed information about the disease canbe found in the OIE Diagnostic Manual for AquaticAnimal Diseases (2000a), and Lighter (1996).
C.5.1.2 Host Range
BMN was observed as natural infections inPenaeus japonicus, P. monodon and P. plebejus(Fig.C.5.1.2a); and as experimental infections inP. chinensis and P. semisulcatus.
C.5.1.3 Geographic Distribution
BMN has occurred in the Kyushu and Chugokuarea of Japan since 1971. BMN-like virus (non-occluded, type C baculovirus) has also been re-ported in P. japonicus in Korea RO and fromP. monodon in the Philippines and possibly inAustralia and Indonesia.
C.5.1.4 Asia-Pacific Quarterly AquaticAnimal Diseases Reporting System (1999-2000)
For the reporting year 1999, no positive reportfrom Japan (1992 was last year of occurrence).The disease was suspected in Korea RO fromJanuary to September 1999 and whole year of2000 (OIE 1999, OIE 2000a).
C.5.2 Clinical Aspects
In Japan, BMN is considered to be one of themajor problems in hatcheries where it infects lar-vae and early postlarval stages causing highmortalities. The apparent white turbidity of thehepatopancreas is caused by necrosis of hepato-pancreas tubule epithelium and possibly also themucosal epithelium. Larvae float inactively butlater stages (late PL) tend to show resistance tothe disease.
C.5.3 Screening Methods
More detailed information on methods for screen-ing BMN can be found in the OIE DiagnosticManual for Aquatic Animal Diseases (OIE 2000a),at http://www.oie.int, or at selected references.
C.5 BACULOVIRAL MIDGUT GLANDNECROSIS (BMN)
C.5.3.1 Presumptive
Techniques suitable for presumptive screeningof asymptomatic animals at Levels I or II arenot available.
C.5.3.2 Confirmatory
C.5.3.2.1 Histopathology (Level II)
Histopathology as described for C.5.4.2.1 is thestandard screening method recommended byOIE (2000a).
C.5.4 Diagnostic Methods
More detailed information on methods for di-agnosis can be found in the OIE DiagnosticManual for Aquatic Animal Diseases (OIE2000a), at http://www.oie.int, or at selected ref-erences.
C.5.4.1 Presumptive
C.5.4.1.1 Gross Observations (Level 1)
Morbid or larvae heavily infected with BMNVshows a cloudy midgut gland, easily observ-able by the naked eye.
C.5.4.1.2 Wet-Mount Technique (Level II)
Hypertrophied nuclei in fresh squashes (viewedunder dark-field microscopy) or in stainedsmears of hepatopancreas (using light micros-copy) are demonstrated in BMNV infectedsamples. When viewed under dark-field illumi-nation equipped with a wet-type condenser, theinfected nuclei appear white against the darkbackground. This is due to the increased re-flected and diffracted rays produced by numer-ous virus particles in the nucleus. Samples fixedin 10% formalin also give same results.
C.5.4.2 Confirmatory
C.5.4.2.1 Histopathology (Level II)
Samples are fixed in Davidson’s fixative, stainedwith standard H&E and examined under brightfield microscopy. Infected shrimps show greatlyhypertropied nuclei (Fig.C.5.4.2.1a) inhepatopancreatic epithelial cells undergoingnecrosis. Infected nuclei show diminishednuclear chromatin, marginated chromatin (Fig.C.5.4.2.1b, c) and absence of occlusion bod-ies characteristic of Baculovirus penaei (BP) (seealso Fig. C.9.3.2.3a,b – section C.9) and
187
C.5 Baculoviral Midgut GlandNecrosis (BMN)
(DV Lightner)
Fig.C.5.1.2a. Section of the hepatopancreas ofP. plebejus displaying several hepatopancreascells containing BMN-type intranuclear inclu-sion bodies. Mayer-Bennett H&E. 1700 x mag-nification.
(DV Lightner)
Fig.C.5.4.2.1a. High magnification of hepato-pancreas from a PL of P. monodon with a se-vere infection by a BMN-type baculovirus. Mostof the hepatopancreas cells display infectednuclei. Mayer-Bennett H&E. 1700x magnifica-tion.
(DV Lightner)
(DV Lightner)
Fig. C.5.4.2.1b, c. Sections of the hepatopan-creas of a PL of P. japonicus with severe BMN.Hepatopancreas tubules are mostly destroyedand the remaining tubule epithelial cells con-tain markedly hypertrophied nuclei that containa single eosinophilic to pale basophilic, irregu-larly shaped inclusion body that fills the nucleus.BMNV infected nuclei also display diminishednuclear chromatin, marginated chromatin andabsence of occlusion bodies that characterizeinfections by the occluded baculoviruses.Mayer-Bennett H&E. Magnifications: (a) 1300x;(b) 1700x.
Fig.C.5.4.2.1d. MBV occlusion bodies whichappear as esosinophilic, generally multiple,spherical inclusion bodies in enormously hy-pertrophied nuclei (arrows). Mayer-BennettH&E. 1700x magnification.
>
c
b
188
Dis. 8:585-589.
Natividad, J.M. and D.V. Lightner. 1992. Preva-lence and geographic distribution of MBVand other diseases in cultured giant tigerprawns (Penaeus monodon) in the Philip-pines, pp.139-160. In: Diseases of CulturedPenaeid Shrimp in Asia and the UnitedStates, Fulks, W. and Main, K.L (eds.). TheOceanic Institute, Honolulu, Hawaii, USA.
OIE. 1999. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 35p.
OIE. 2000a. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
OIE. 2000b. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 40p.
Park, M.A. 1992. The status of culture and dis-eases of penaeid shrimp in Korea, pp. 161-167. In: Diseases of Cultured Penaeid Shrimpin Asia and the United States, Fulks, W. andMain, K.L (eds.). The Oceanic Institute, Ho-nolulu, Hawaii, USA.
Sano, T. and K. Momoyama. 1992. Baculovirusinfection of penaeid shrimp in Japan, pp. 169-174. In: Diseases of Cultured Penaeid Shrimpin Asia and the United States, Fulks, W. andMain, K.L (eds.). The Oceanic Institute, Ho-nolulu, Hawaii, USA.
Sano, T. T. Nishimura, K. Oguma, K. Momoyamaand N. Takeno. 1981. Baculovirus infectionof cultured Kuruma shrimp Penaeusjaponicus in Japan. Fish Pathol. 15:185-191.
C.5.4.2.2 Transmission Electron Micros-copy (TEM) (Level III)
Tranmission electron microscopy can be usedconfirm diagnosis of BMN through demonstra-tion of the rod-shaped enveloped virions asdescribed in C.5.1.1.
C.5.4 Modes of Transmission
The oral route has been demonstrated to bethe main infection pathway for BMNV infection.Viruses released with faeces into the environ-mental water of intensive culture systems ofP. japonicus play an important role in diseasespread.
C.5.5 Control Measures
The concentrations of various disinfectants re-quired to kill BMNV are toxic to shrimp larvae.Complete or partial eradication of viral infec-tion may be accomplished by thorough wash-ing of fertile eggs or nauplii using clean seawater to remove the adhering excreta. Disin-fection of the culture facility and the avoidanceof re-introduction of the virus are critical fac-tors to control BMN disease.
The suggested procedure for eradication ofBMN infection involves collection of fertile eggsfrom broodstock and passing them through asoft gauze with pore size of 800 mm to removedigested excrement or faeces of the shrimp.The eggs are then washed with running seawater at salinity level of 28-30% for 3-5 min tomake sure all the faecal debris has been re-moved. The eggs are then collected by pass-ing the suspension through a soft gauze withpore size of 100 mm. The eggs are then furtherwashed with running sea water at salinity levelof 28-30% for 3-5 min to remove the adhesiveviral particles.
C.5.6 Selected References
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDisease of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Momoyama, K. and T. Sano. 1989. Develop-mental stages of kuruma shrimp larvae,Penaeus japonicus Bate, with baculoviralmid-gut gland necrosis (BMN) virus. J. Fish
C.5 Baculoviral Midgut GlandNecrosis (BMN)
189
C.6.1 Background
C.6.1.1 Causative Agent
Gill-associated virus (GAV) is a single-strandedRNA virus related to viruses of the familyCoronaviridae. It is closely related to yellow headvirus and is regarded as a member of the yellowhead complex. GAV can occur in healthy ordiseased shrimp and was previously called lym-phoid organ virus (LOV) when observed inhealthy shrimp.
C.6.1.2 Host Range
Natural infection with GAV has only been reportedin Penaeus monodon but experimental infectionhas caused mortalities in P. esculentus, P.merguiensis and P. japonicus. An age or sizerelated resistance to disease was observed in P.japonicus.
C.6.1.3 Geographic Distribution
GAV has only been recorded from Queenslandon the north-east coast of Australia and is en-demic to P. monodon in this region.
Australia reported widespread occurrence of LOVamong healthy farmed and wild P. monodon inQueensland. Other countries reported “no infor-mation available” for GAV for the reporting pe-riod for 1999 and 2000 (OIE 1999, OIE 2000).
C.6.2 Clinical Aspects
GAV is endemic in healthy P. monodon in north-ern Queensland. It is unclear whether the onsetof disease results from environmental stress lead-ing to clinical expression of the pre-existing vi-rus as can occur with YHD and WSD or whetherthe disease arises from a new infection with apathogenic strain of GAV. GAV is predominantlyfound in the gill and lymphoid organ but has alsobeen observed in haemocytes. During acute in-fections, there is a rapid loss of haemocytes, thelymphoid organs appear disorganised and devoidof normal tubule structure, and the virus is de-tected in the connective tissues of all major or-gans.
The PCR primers below are designed to am-plify a 618 bp region of GAV:
GAV-5 5’-AAC TTT GCC ATC CTC GTCAC-3’
GAV-6 5’-TGG ATG TTG TGT GTT CTCAAC-3’
The PCR primers below are designed to am-plify a 317 bp region internal to the region am-plified by GAV5 and GAV6:
GAV-1 5’-ATC CAT ACT ACT CTA AAC TTCC-3’
GAV-2 5’-GAA TTT CTC GAA CAA CAGACG-3’
Total RNA (100 ng) is denatured in the pres-ence of 35 pmol of each primer (GAV-5 andGAV-6) by heating at 98°C for 8 min in 6 mlDEPC-water containing 0.5 ml deionisedformamide and quenched on dry ice. cDNA issynthesised by the addition of 2 ml SuperscriptII buffer x 5, 1 ml 100 mM DTT, 0.5 ml 10 mMdNTPs, 20 U rRNasinTM (Promega) and 100 USuperscript II Reverse Transcriptase (Life Tech-nologies) and DEPC-water to 10 ml and the re-action is incubated at 42oC for 1 hr followed byheating at 99oC for 5 min before quenching onice. One tenth of the cDNA reaction (1 ml = 10ng RNA) is amplified in 50 ml using Taq buffer(10 mM Tris-HCl pH 9.0, 50 mM KCl, 0.1% Tri-ton X-100), 1.5 mM MgCl2, 35 pmol each primerGAV-5 and GAV-6 and 200 mM dNTPs overlaidwith 50 ml liquid paraffin. PCRs are initiatedusing a “hot-start” protocol in which the reac-tion was heated at 85oC for 5 min prior to theaddition of 2.5 U Taq polymerase (Promega).DNA is amplified by 30 cycles of 95oC/1 min,58oC/1 min, 72oC/40 sec followed by 72oC/10min final extension and 20oC hold using eithera Corbett Research or Omnigene (Hybaid) ther-mal cycler. PCR products (10 ml) are resolvedin 2% agarose-TAE gels containing 0.5 mg/mlethidium bromide.
When the result of the primary RT-PCR is nega-tive or inconclusive, 0.5 ml of the primary PCRis amplified by nested PCR as above in a 50 mlreaction volume using primers GAV-1 and GAV-2. In some cases, 5 ml of the RT-PCR is used.Nested PCR conditions are as for the primaryPCR except that the extension time is reducedto 30 sec and number of cycles is reduced to20. Nested PCR aliquots (10 ml) are analysedin 2% agarose-TAE gels.
190
C.6.4 Diagnostic Methods
C.6.4.1 Presumptive
C.6.4.1.1 Gross Observations (Level I)
Shrimp with an acute GAV infection demon-strate lethargy, lack of appetite and swim onthe surface or around the edge of ponds. Thebody may develop a dark red colour particu-larly on the appendages, tail fan and mouthparts; gills tend to be yellow to pink in colour.Barnacle and tube worm attachment togetherwith gill fouling have also been observed. Thegross signs of acute GAV infection are variableand not always seen and thus, they are not re-liable, even for preliminary diagnosis.
C.6.4.1.2 Cytology/Histopathology(Level II)
The cephalothorax of infected prawns is sepa-rated from the abdomen and split longtudinally.The sample is then fixed in Davidson’s fixativeand processed for histology. Sections arestained with H&E. Lymphoid organs from dis-eased shrimp display loss of the normal tubulestructure. Where tubule structure is disrupted,there is no obvious cellular or nuclear hyper-trophy, pyknotic nuclei or vacuolization. Foci ofabnormal cells are observed within the lym-phoid organ and these may be darkly eosino-philic. The gills of diseased shrimp displaystructural damage including fusion of gill fila-ment tips, general necrosis and loss of cuticlefrom primary and secondary lamellae. The cy-tology of the gills appears normal apart fromsmall basophilic foci of necrotic cells.
C.6.4.2 Confirmatory
C.6.4.2.1 Transmission Electron Micros-copy (TEM) (Level III)
Tissue samples are fixed in 2.5% glutaralde-hyde/2% paraformaldehyde in cacodylatebuffer and post-fixed in 1% osmium tetroxide.Fixed samples are then dehydrated through agraded series of ethanol concentrations andmounted in Spurr’s resin. 50 nm sections aremounted on Cu-200 grids, stained with uranylacetate/70% methanol and Reynold’s lead cit-rate. The cytoplasm of lymphoid organ cellsfrom diseased shrimps contains both rod-shaped enveloped virus particles and viralnucleocapsids. The nucleocapsids are from166-435 nm in length 16-18 nm in width.
C.6 Gill-Associated Virus (GAV)
(P Walker)
Fig. C.6.4.2.1. Transmission electron micros-copy of GAV.
Nucleocapsids have striations with a periodic-ity of 7 nm and are often found associated withthe endoplasmic reticulum. Enveloped virionsare less common, occurring in about 20% ofcells within the disrupted areas of the lymphoidorgan. The enveloped virions (Fig. C.6.4.2.1) are183-200 nm long and 34-42 nm wide againassociated with the endoplasmic reticulum.Both enveloped virions and nucleocapsids arepresent in gill tissue but the nucleocpsids aremore commonly occurring in 40-70% of cellswhereas enveloped virions are present in lessthan 10% of cells.
The most effective form of horizontal transmis-sion is direct cannibalism but transmission canalso be water-borne. GAV is also transmittedvertically from healthy broodstock. The virusmay be transmitted from either or both parentsbut it is not clear if infection is within the egg.
C.6.6 Control Measures
There are no known control measures for GAV.Prevention of the movement of GAV infectedstock into historically uninfected areas is rec-ommended. Drying out of infected ponds ap-pears effective in preventing persistence of thevirus.
191
C.6.7 Selected References
Cowley, J.A., C.M. Dimmock, C.Wongteerasupaya, V. Boonsaeng, S. Panyamand P.J. Walker. 1999.Yellow head virus fromThailand and gill-associated virus from Aus-tralian are closely related but distinct viruses.Dis. Aquat. Org. 36:153-157.
Cowley, J.A., C.M. Dimmock, K.M. Spann andP.J. Walker. 2000b. Gill-associated virus ofPenaeus monodon prawns: an invertebratevirus with ORF1a and ORF 1b genes relatedto arteri- and coronaviruses. J. Gen. Virol. 81:1473 – 1484.
Spann, K.M., J.E. Vickers and R.J.G.Lester.1995. Lymphoid organ virus ofPenaeus monodon from Australia. Dis.Aquat. Org. 23: 127-134
Spann, K.M., J.A. Cowley, P.J. Walker andR.J.G. Lester.1997. A yellow-head-like virusfrom Penaeus monodon cultured in Austra-lia. Dis. Aquat. Org. 31: 169-179.
Spann, K.M., A.R. Donaldson, I.J. East, J.A.Cowley and P.J. Walker. 2000. Differencesin the susceptibility of four penaeid prawnspecies to gill-associated virus (GAV). Dis.Aquat. Org. 42: 221-225.
Walker, P.J., J.A. Cowley, K.M. Spann, R.A.J.Hodgson, M.A. Hall and B.Withyachumnernkul. 2001. Yellow head com-plex viruses: transmission cycles and topo-graphical distribution in the Asia-Pacific re-gion, pp. 227-237. In: C.L. Browdy and D.E.Jory (eds).The New Wave: Proceedings of theSpecial Session on Sustainable Shrimp Cul-ture, Aquaculture 2001. The World Aquacul-ture Society, Baton Rouge, LA.
C.6 Gill-Associated Virus (GAV)
192
C.7.1 Background Information
C.7.1.1 Causative Agent
Spawner-isolated Mortality Virus Disease(SMVD) is caused by a single-stranded icosa-hedral DNA virus measuring 20-25 nm. Thesecharacteristics are most closely associated withthose of the Family Parvoviridae. The virus hasbeen named Spawner-isolated Mortality Virus(SMV) and other disease names include SpawnerMortality Syndrome (SMS) and Midcrop Mortal-ity Syndrome (MCMS). More detailed informa-tion about the disease can be found in OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000a).
C.7.1.2 Host Range
SMVD affects Penaeus monodon. Experimentalinfections have also resulted in mortalities of P.esculentus, P. japonicus, P. merguiensis andMetapenaeus ensis. Moribund, farmed freshwa-ter crayfish (Cherax quadricarinatus) have alsobeen associated with putative SMV infection us-ing DNA-probe analyses.
C.7.1.3 Geographic Distribution
SMVD has been reported from Queensland, aswell as the Philippines and Sri Lanka.
C.7.1.3.4 Asia-Pacific Quarterly AquaticAnimal Disease Reporting System (1999-2000)
Most countries reported “no information available”or “never reported” for the 2 year reporting pe-riod (1999 and 2000) except for Sri Lanka whichsuspected the disease in August 1999 and re-ported positive occurrence in September 1999(OIE 1999, OIE 2000b). Philippines reported posi-tive occurrence of SMV in October to December1998 where samples of P. monodon sent to Aus-tralia for insitu hybridization using SMV probeproduced positive results (NACA/FAO 1999).
C.7.2 Clinical Aspects
There are no specific clinical signs known forSMV. It is one of several viruses associated withmid-crop mortality syndrome (MCMS) which re-sulted in significant mortalities of juvenile andsub-adult P. monodon cultured in Australia from1994 to 1996. Similarly affected P. monodon
from the Philippines were also infected with lu-minous vibriosis (Vibrio harveyi).
C.7.3 Screening Methods
More detailed information on methods forscreening SMVD can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000a), at http://www.oie.int, or at selected ref-erences.
There are no standard screening methods avail-able for asymptomatic animals.
C.7.4 Diagnostic Methods
More detailed information on methods for di-agnosis of SMVD can be found in the OIE Di-agnostic Manual for Aquatic Animal Diseases(OIE 2000a), at http://www.oie.int, or at selectedreferences.
C.7.4.1 Presumptive
C.7.4.1.1 Gross Observations (Level 1)
There are no specific clinical signs for SMVD.Juvenile P. monodon in grow-out ponds mayshow discolouration, lethargy, fouling and an-orexia. Since this may be caused by severalviral or bacterial infections, however, other di-agnostic methods are required.
C.7.4.1.2 Cytology/Histopathology(Level II)
The histopathology associated with SMVD isnot disease specific. In naturally infected juve-nile P. monodon, haemocyte infiltration andcytolysis is focussed around the enteric epithe-lial surfaces. Experimental infections, using tis-sue extracts from shrimp with SMVD developsystemic infections manifest by systemichaemocytic infiltration, necrosis and sloughingof epithelial cells of the midgut and hepatopan-creas.
C.7.4.2 Confirmatory
C.7.4.2.1 Transmission Electron Micros-copy (TEM) (Level III)
SMV virions are found in the gut epithelial tis-sues. The viral particles measure approximately20-25 nm in diameter and have hexagonal
C.7 SPAWNER-ISOLATEDMORTALITY VIRUS DISEASE
(SMVD)4
4 This disease is listed in the current FAO/NACA/OIE Quarterly Aquatic Animal Disease Reporting System as Spawner mortalitysyndrome (‘Midcrop mortality syndrome’).
193
(icosahedral) symmetry.
C.7.5 Modes of Transmission
Moribund and dead individuals are cannibalisedby surviving animals, which is assumed to fa-cilitate horizontal transmission.
C.7.6 Control Measures
Prevention of introduction of shrimp from SMVinfected stock into historically uninfected areasis recommended. Daily removal of moribundanimals from ponds, particularly early in pro-duction, has also been recommended. Stock-ing of ponds with progeny of spawners withSMV-negative faecal testing using PCR-probeshas been shown to reduce mortality by 23%.
C.7.7 Selected References
Albaladejo, J.D., L.M. Tapay, V.P. Migo, C.G.Alfafara, J.R. Somga, S.L. Mayo, R.C.Miranda, K. Natividad, F.O. Magbanua, T.Itami, M. Matsumura, E.C.B. Nadala, Jr. andP.C. Loh. 1998. Screening for shrimp virusesin the Philippines, pp. 251-254. In: Advancesin shrimp Biotechnology, Flegel, T.W. (ed).National Center for Genetic Engineering andBiotechnology. Bangkok, Thailand.
Fraser, C.A. and L. Owens. 1996. Spawner-iso-lated mortality virus from Australian Penaeusmonodon. Dis. Aquat. Org. 27: 141-148.
NACA/FAO. 1999. Quarterly Aquatic AnimalDisease Report (Asia-Pacific Region), 98/2,October to December 1998. FAO ProjectTCP/RAS/6714. Bangkok, Thailand. 41p.
OIE. 1999. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 35p.
OIE. 2000a. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
OIE. 2000b. Regional Aquatic Animal DiseaseYearbook 1999 (Asian and Pacific Region).OIE Representation for Asia and the Pacific.Tokyo, Japan. 40p.
Owens, L. and C. McElnea. 2000. Natural in-fection of the redclaw crayfish Cheraxquadricarinatus with presumptive spawner-isolated mortality virus. Dis. Aquat. Org. 40:219-233.
Owens, L., G. Haqshenas, C. McElnea and R.Coelen. 1998. Putative spawner-isolatedmortality virus associated with mid-crop mor-tality syndrome in farmed Penaeus monodonfrom northern Australia. Dis. Aquat. Org. 34:177-185.
Taura Syndrome (TS) is caused by a virus, TauraSyndrome Virus (TSV) tentatively classified asa member of the Picornaviridae based on itsmorphology (31- 32 nm non-enveloped icosa-hedron), cytoplasmic replication, buoyant den-sity of 1.338 g/ml, genome consisting of a linear,positive-sense ssRNA of approximately 10.2 kbin length, and a capsid comprised of three ma-jor (55, 40, and 24 kD) and one minor (58 kD)polypeptides. More detailed information aboutthe pathogen can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases(2000) and Lightner (1996).
C.8.1.2 Host Range
TSV infects a number of American penaeid spe-cies. The most susceptible species is the Pa-cific white shrimp Penaeus vannamei, althoughP. stylirostris, and P. setiferus can also be infected.Post-larvae and juvenile P. schmittii, P. aztecus,P. duorarum, P. chinensis, P. monodon, andMarsupenaeus (Penaeus) japonicus have beeninfected experimentally.
C.8.1.3 Geographic Distribution
Taura Syndrome was first detected in shrimpfarms near the Taura River, Ecuador (hence thename of the disease) in 1992. It then spreadthroughout most shrimp growing regions of LatinAmerica including Hawaii (infections successfullyeradicated ) and the Pacific coasts of Colombia,Costa Rica, Ecuador, El Salvador, Guatemala,Honduras, Mexico, Nicaragua, Panama, andPeru.
TSV has also been reported from cultured shrimpalong the Atlantic coasts of Belize, Brazil, Co-lumbia, Mexico, and Venezuela and the south-eastern U.S. states of Florida, South Carolina andTexas. TSV has, however, been successfullyeradicated from cultured stocks in Florida andBelize. TSV is found in wild penaeids in Ecua-dor, El Salvador, Honduras, and Mexico. The onlyrecord of TSV in the eastern hemisphere is fromTaiwan, Province of China, where the diseasewas likely introduced with P. vannamei from Cen-tral America.
C.8 TAURA SYNDROME (TS)5
C.8.2 Clinical Aspects
Taura Syndrome is particularly devastating topost-larval P. vannamei within approximately 14to 40 days of stocking into grow-out ponds ortanks, however, larger stages may also be se-verely affected. Three distinct phases charac-terize TS disease progression: i) the acute stage,during which most mortalities occur; ii) a brieftransition phase, and iii) a chronic ‘carrier’ stage.In the acute phase, the cuticular epithelium isthe most severely affected tissue. In the chronicphase, the lymphoid organ becomes the pre-dominant site of infection. In P. vannamei, theacute phase of infection may result in highmortalities (40-90%), while most strains of P.stylirostris appear resistant to fatal levels ofinfection. Survivors of acute TSV infection passthrough a brief transition phase and enter thechronic phase which may persist for the rest oftheir lives. This sub-clinical phase of infectionis believed to have contributed to the spreadof the disease via carriage of viable TSV.
C.8.3 Screening Methods
Detailed information on methods for screeningTSV can be found in the OIE Diagnostic Manualfor Aquatic Animal Diseases (OIE 2000), at http://www.oie.int, or at selected references.
C.8.3.1 Presumptive
C.8.3.1.1 Gross Observation (Level I)
Any Penaeus vannamei, or other susceptiblepenaeid survivors of a TS outbreak, should beconsidered suspect carriers of TSV. However,there are no gross observation or Level I signsthat can be used to screen sub-clinical carri-ers.
C.8.3.1.2 Histopathology (Level II)
Post-larvae, juveniles and adults can bescreened using routine histological techniquesand stains. Chronic stages of infection arecharacterised by the presence of spherical ac-cumulations of cells in the lymphoid organ, re-ferred to as ‘lymphoid organ spheroids’ (LOS).These masses are composed of presumed ph-agocytic hemocytes, which have sequesteredTSV and aggregate within intertubular spacesof the lymphoid organs.
5 Taura Syndrome (TS) is now classified as an OIE Notifiable Disease (OIE 2000).
195
C.8.3.1.3 Immunoassays (Level III)
A commercial dot blot detection kit is availablefor TSV from DiagXotics (Wilton, CT, USA).ELISA kits using a TSV MAb have also beenproduced. These can be used to screen pos-sible TSV carriers, but any positive resultsshould be cross-checked with another confir-matory technique, or by bioassay, sincevisualisation of clinical signs or the virus is notpossible with molecular screening techniques(this also applies to screening with PCR probes- C.8.3.1.5)
C.8.3.1.4 In situ Hybridization (Level III)
A commercial in situ hybridization detection kitis available for TSV from DiagXotics (Wilton, CT,USA). This technique is usually reserved forconfirmation of observations made using rou-tine histology (C.8.3.1.2), rather than as a stand-alone technique for screening.
C.8.3.1.5 PCR Probes (Level III)
An RT-PCR based assay uses shrimphaemolymph for screening purposes, giving theadvantage of being able to screen livebroodstock and assist selection of TSV-nega-tive shrimp for spawning. Positive results fromsurvivors of previous TSV outbreaks can beconsidered confirmatory, however, first timepositive results from non-susceptible speciesor non-enzootic sources should be analyzedusing another, confirmatory, technique for thesame reasons given for dot-bot hybridization(C.8.3.1.3).
C.8.4 Diagnostic Methods
Detailed information on methods for diagnosisof TSV can be found in the OIE DiagnosticManual for Aquatic Animal Diseases (OIE 2000),at http://www.oie.int, or at selected references.
C.8.4.1 Presumptive
C.8.4.1.1 Gross Observations (Level I)
Penaeus vannamei post-larvae or older shrimpmay show a pale reddish discolouration, espe-cially of the tail fan (Fig.C.8.4.1.1a,b) andpleiopods (hence the name “red tail” disease,applied by farmers when the disease first ap-peared in Ecuador). This colour change is dueto expansion of the red chromatophores withinthe cuticular epithelium. Magnification of the
C.8 Taura Syndrome (TS)
edges of the pleiopods or uropods may revealevidence of focal necrosis. Shrimp showingthese signs typically have soft shells, an emptygut and often die during moulting. During se-vere epizootics, sea birds (gulls, terns, cormo-rants, etc.) may be attracted to ponds contain-ing shrimp over 1 gm in size.
Although the transition stage of TS only lasts afew days, some shrimp may show signs of ran-dom, multi-focal, irregularly shaped melanizedcuticular lesions (Fig.C.8.4.1.1c,d,e). Thesecorrespond to blood cell repair activity aroundthe necrotic lesions induced by TSV infectionof the cuticular epithelium. Such shrimp may,or may not, have soft cuticles and reddiscolouration, and may be behaving and feed-ing normally.
C.8.4.1.2 Histopathology (Level II)
Diagnosis of TS in acute stages of the diseaserequires histological (H&E stain preparations)demonstration of multi-focal areas of necrosisin the cuticular epithelium of the general bodysurface, appendages, gills (Fig.C.8.4.1.2a),hind-gut, esophagus and stomach(Fig.C.8.4.1.2b). Sub-cuticular connective tis-sue and striated muscle fibers basal or adja-cent to affected cuticular epithelium may alsoshow signs of necrosis. Rarely, the antennalgland tubule epithelium is affected. Cuticularlesions may contain foci of cells with abnormallyeosinophilic (pink-staining) cytoplasm and py-knotic (condensed nucleoplasm) or karyorrhe-ctic (fragmented nucleoplasm) nuclei. Rem-nants of necrotic cells are often abundant withinacute phase lesions and appear as roughlyspherical bodies (1-20 µm diameter) that rangein stain uptake from eosinophilic to lightly ba-sophilic (blue-staining). Another feature of acuteTS is the absence of haemocyte infiltration, orother signs of a host defense response. Thesefeatures combine to give acute phase TS le-sions a “peppered” appearance(Fig.C.8.4.1.2c), that is considered to be diag-nostic for the disease, and can be consideredconfirmatory (C.8.4.2.2) in susceptible speciesin enzootic waters. Confirmation by anothertechnique is recommended for first time obser-vations of these histopathological features, ortheir appearance in abnormal penaeid speciesor locations.
In the transitional phase of TS, the number andseverity of the cuticular lesions that charac-terize acute phase infections decrease andaffected tissues become infiltrated byhaemocytes. These may become melanized
196
(DV Lightner)
(C.8.4.1.1). If the acute cuticular lesions perfo-rate the epicuticle, the affected surfaces mayshow evidence of colonization and invasion byVibrio spp, or other secondary infections.
In the chronic phase of TS, the only sign of in-fection is the presence of prominent lymphoidorgan spheres (LOS) (Fig.C.8.4.1.2d), whichcorrespond to aggregations of presumedhemocytes within the intertubular spaces of thelymphoid organ.
Fig. C.8.4.1.1a,b. a. Moribund, juvenile, pond-reared Penaeus vannamei from Ecuador in theperacute phase of Taura Syndrome (TS). Shrimpare lethargic, have soft shells and a distinct redtail fan; b. Higher magnification of tail fan show-ing reddish discoloration and rough edges ofthe cuticular epithelium in the uropods sugges-tive of focal necrosis at the epithelium of thosesites (arrows).
(DV Lightner/F Jimenez)
Fig. C.8.4.1.1c,d,e. Juvenile, pond-reared P.vannamei (c – from Ecuador; d – from Texas; e– from Mexico) showing melanized foci marksites of resolving cuticular epithelium necrosisdue to TSV infection.
C.8 Taura Syndrome (TS)
a
b
d
c
e
197
(DV Lightner)
Fig. C.8.4.1.2c. Higher magnification of Fig.C.8.4.1.2b showing the cytoplasmic inclusionswith pyknotic and karyorrhectic nuclei giving a‘peppered’ appearance. Mayer-Bennett H&E.900x magnification.
(DV Lightner)
Fig. C.8.4.1.2d. Mid-sagittal section of the lym-phoid organ (LO) of an experimentally infectedjuvenile P. vannamei. Interspersed among nor-mal appearing lymphoid organ (LO) cords or tis-sue, which is characterized by multiple layersof sheath cells around a central hemolymphvessel (small arrow), are accumulations of dis-organized LO cells that form LO ‘spheroids”.Lymphoid organs spheres (LOS) lack a centralvessel and consists of cells which showkaryomegaly and large prominent cytoplasmicvacuoles and other cytoplasmic inclusions(large arrow). Mayer-Bennett H&E. 300x mag-nification.
(DV Lightner)
Fig. C.8.4.1.2a. Focal TSV lesions in the gills(arrow). Nuclear pykinosis and karyorrhexis, in-creased cytoplasmic eosinophilia, and an abun-dance of variably staining generally sphericalcytoplasmic inclusions are distinguishing char-acteristics of the lesions. 900x magnification.
(DV Lightner)
Fig. C.8.4.1.2b. Histological section throughstomach of juvenile P. vannamei showing promi-nent areas of necrosis in the cuticular epithe-lium (large arrow). Adjacent to focal lesions arenormal appearing epithelial cells (small arrows).Mayer-Bennett H&E. 300x magnification.
C.8 Taura Syndrome (TS)
198
C.8.4.2 Confirmatory
C.8.4.2.1 Bioassay (Levels I/II)
Specific Pathogen Free (SPF) juvenile Penaeusvannamei can be used to test suspect TSV-in-fected shrimp. Three exposure methods can beused:i) Suspect shrimp can be chopped up and fed
to SPF juvenile P. vannamei held in smalltanks. Another tank should hold SPF shrimpfrom the same source, but fed regular feedonly (controls). If the suspect shrimp werepositive for TSV, gross signs and histopatho-logical lesions should become evident within3-4 days of initial exposure. Significant mor-talities usually occur by 3-8 days post-ex-posure. The control shrimp should stayhealthy and show no gross or histologicalsigns of TS.
ii) Whole shrimp collected from a presumptiveTSV epizootic can be homogenized for in-oculation challenge. Alternatively, heads maybe used where presumptive TS signs appearto be at the transitional phase of develop-ment (melanized lesions) or where there areno clinical signs of infection (presumptivechronic phase) since this contains the lym-phoid organ.
iii) Haemolymph samples may be taken frombroodstock and used to expose SPF indica-tor shrimp, as for method ii) above.
C.8.4.2.2 Histopathology (Level II)
Observation of the lesions described underC.8.4.1.2 can be considered confirmatory forsusceptible species from sources known to beenzootic for TSV.
C.8.4.2.3 Transmission Electron Micros-copy (TEM) (Level III)
Transmission electron microscopy of acutephase epithelial lesions or lymphoid organspheroids that demonstrate the presence ofnon-enveloped icosahedral viral particles, 31-32 nm in diameter, in the cytoplasm of affectedcells, can be considered confirmatory whereconsistent with gross and histological clinicalsigns in a susceptible penaeid species. Furtherconfirmation using molecular techniques(C.8.4.2.4-6) are recommended, however, forfirst-time diagnoses or detection in speciesother than those listed as being naturally orexperimentally susceptible.
C.8.4.2.4 Dot Blot (Level III)
As described under C.8.3.1.3.
C.8.4.2.5 In situ Hybridization (Level III)
As described under C.8.3.1.4.
C.8.4.2.6 PCR Probes (Level III)
As described under C.8.3.1.5.
C.8.5 Modes of Transmission
Shrimps that have survived the acute and tran-sitional phases of TS can maintain chronic sub-clinical infections within the lymphoid organ, forthe remainder of their lives. These shrimp maytransmit the virus horizontally to other suscep-tible shrimp. Vertical transmission is suspected,but this has yet to be conclusively demon-strated.
In addition to movement of sub-clinical carri-ers of TSV, aquatic insects and sea birds havebeen implicated in transmission of the disease.The water boatman, Trichocorixa reticulata(Corixidae), feeds on dead shrimp and is be-lieved to spread TSV by flying from pond topond. Laughing gull, Larus atricilla, faeces col-lected from around TSV-infected ponds in Texasduring the 1995 epizootic, were also found tocontain viable TSV. Viable TSV has also beenfound in frozen shrimp products.
C.8.6 Control Measures
In much of Central America where TS is en-zootic, shrimp farm management has shiftedtowards increased use of wild caught P.vannamei PL, rather than hatchery-reared PL.This has improved survival to harvest. It is sus-pected that wild PL may have increased toler-ance of TS due to natural exposure and selec-tion of survivors. Another management strat-egy has been doubling post-larval stockingdensities in semi-intensive pond culture. Heavylosses due to TS early in the production cycleare compensated for by the survivors (5-40%of the original number stocked) being TS toler-ant. Selective breeding is showing promise fordevelopment of TSV resistant stocks of P.vannamei and P. stylirostris (which are resistantto both IHHNV and TSV). Initial results show a20-40% improvement in survival.
Eradication depends on total removal of in-fected stocks, disinfection of the culture facil-ity, avoidance of re-introduction of the virus
C.8 Taura Syndrome (TS)
199
(from nearby culture facilities, wild shrimp, orsub-clinical carriers etc.), and re-stocking withTSV-free PL produced from TSV-freebroodstock.
C.8.7 Selected References
Aragon-Noriega, E.A., J.H. Cordova-Murueta,and H.L. Trias-Hernandez. 1998. Effect ofTaura-like viral disease on survival of thewestern white shrimp (Penaeus vannamei)cultured at two densities in NorthwesternMexico. World Aquac. 29(3):66-72.
Bonami, J.R., K.W. Hasson, J. Mari, B.T. Poulos,and D.V. Lightner. 1997. Taura syndrome ofmarine penaeid shrimp: Characterisation ofthe viral agent. J. Gen. Virol. 78(2):313-319.
Brock, J.A., R. Gose, D.V. Lightner, and K.W.Hasson . 1995. An overview of Taura Syn-drome, an important disease of farmedPenaeus vannamei, pp. 84-94. In: Swimmingthrough troubled water. Proceedings of theSpecial Session on Shrimp Farming, WorldAquaculture Society, Baton Rouge, LA.
Dixon, H. and J. Dorado. 1997. Managing Taurasyndrome virus in Belize: A case study.Aquac. Mag. 23(2): 30-42.
Garza, J.R., K.W. Hasson, B.T. Poulos, R.M.Redman, B.L. White, and D.V. Lightner. 1997.Demonstration of infectious Taura syndromevirus in the feces of seagulls collected dur-ing an epizootic in Texas. J. Aquat. Anim.Health 9(2):156-159.
Hasson, K.W., D.V. Lightner, B.T. Poulos, R.M.Redman, B.L. White, J.A. Brock, and J.R.
Bonami. 1995. Taura syndrome in Penaeusvannamei: Demonstration of a viral etiology.Dis. Aquat. Org. 23(2):115-126.
Hasson, K.W., J. Hasson, H. Aubert, R.M.Redman, and D.V. Lightner. 1997. A new RNA-friendly fixative for the preservation ofpenaeid shrimp samples for virological de-tection using cDNA genomic probes. J. Virol.Meth. 66:227-236.
Hasson, K.W., D.V. Lightner, J. Mari, J.R.Bonami, B.T. Poulos, L.L. Mohney, R.M.Redman, and J.A Brock. 1999a. The geo-graphic distribution of Taura Syndrome Vi-rus (TSV) in the Americas: determination byhistopathology and in situ hybridisation us-ing TSV-specific cDNA probes. Aquac. 171(1-2):13-26.
Hasson, K.W., Lightner, D.V., Mohney, L.L.,Redman, R.M., Poulos, B.T. and B.M. White.1999b. Taura syndrome virus (TSV) lesion de-velopment and the disease cycle in the Pa-cific white shrimp Penaeus vannamei. Dis.Aquat. Org. 36(2):81-93.
Hasson, K.W., D.V. Lightner, L.L. Mohney, R.M.Redman, and B.M. White. 1999c. Role of
lymphoid organ spheroids in chronic Taurasyndrome virus (TSV) infections in Penaeusvannamei. Dis. Aquat. Org. 38(2):93-105.
Jimenez, R., R. Barniol, L. Barniol and M.Machuca. 2000. Periodic occurrence of epi-thelial viral necrosis outbreaks in Penaeusvannamei in Ecuador. Dis. Aquat.Org.42(2):91-99.
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDiseases of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Lightner, D.V. 1999. The penaeid shrimp virusesTSV, IHHNV, WSSV and YHV: Current Statusin the Americas, available diagnostic meth-ods and management strategies. J. AppliedAquac. 9(2):27-52.
Lightner, D.V. and R.M. Redman. 1998. Strate-gies for the control of viral diseases of shrimpin the Americas. Fish Pathol. 33:165-180.
Lotz, J.M. 1997a. Effect of host size on viru-lence of Taura virus to the marine shrimpPenaeus vannamei (Crustacea: Penaeidae).Dis. Aquat. Org. 30(1):45-51.
Lotz, J.M. 1997b. Disease control and patho-gen status assurance in an SPF-basedshrimp aquaculture industry, with particularreference to the United States, pp. 243-254.In: Diseases in Asian Aquaculture III. Flegel,T.W. and I.H. MacRae (eds.). Fish Health Sec-tion, Asian Fisheries Society, Manila, The Phil-ippines.
Morales-Covarrubias, M.S. and C. Chavez-Sanchez. 1999. Histopathological studies onwild broodstock of white shrimp Penaeusvannamei in the Platanitos Area, adjacent toSan Blas, Nayarit, Mexico. J. World Aquac..
C.8 Taura Syndrome (TS)
200
Soc. 30(2):192-200.
Nunan, L.M., B.T. Poulos, and D.V. Lightner.1998. Reverse transcriptase polymerasechain reaction (RT-PCR) used for the detec-tion of Taura Syndrome virus (TSV) in experi-mentally infected shrimp. Dis. Aquat. Org.34(2):87-91.
OIE. 2000. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
Overstreet, R.M., D.V. Lightner, K.W. Hasson,S. McIlwain, and J.M. Lotz. 1997. Suscepti-bility to Taura syndrome virus of somepenaeid shrimp species native to the Gulf ofMexico and the southeastern United States.J. Invert. Pathol. 69(2):165-176.
Poulos, B.T., R. Kibler, D. Bradley-Dunlop, L.L.Mohney, and D.V. Lightner. 1999. Productionand use of antibodies for the detection of theTaura syndrome virus in penaeid shrimp. Dis.Aquat. Org. 37(2):99-106.
Yu, C.-I. and Y.-L. Song. 2000. Outbreaks ofTaura syndrome in Pacific white shrimpPenaeus vannamei cultured in Taiwan. FishPathol. 35(1):21-24.
Zarain-Herzberg, M. and F. Ascencio-Valle.2001. Taura syndrome in Mexico: follow-upstudy in shrimp farms of Sinaloa. Aquac.193(1-2):1-9.
C.8 Taura Syndrome (TS)
201
C.9.1 Background Information
C.9.1.1 Causative Agent
Nuclear Polyhedrosis Baculoviroses (NPB) in-fections are caused by the Baculoviridae,Baculovirus penaei (BP - PvSNPV) and Mondonbaculovirus (MBV – PmSNPV). The diseasesassociated with these viruses are Baculovirusdisease, Nuclear polyhedrosis disease, polyhe-dral inclusion body virus disease (PIB), polyhe-dral occlusion body virus disease (POB) andBaculovirus penaei (BP) virus disease. Moredetailed information about the disease can befound at OIE Diagnostic Manual for Aquatic Ani-mal Diseases (OIE 2000).
C.9.1.2 Host Range
BP infects in a wide range of penaeid shrimp in-cluding Penaeus duorarum, P. aztecus, P.setiferus, P. vannamei, P. stylirostris and P.marginatus. BP has also been reported from P.penicillatus, P. schmitti, P. paulensis and P. subtilis.
MBV-type baculoviruses are, by definition, pri-marily found in cultured P. monodon. Other co-cultured species may also acquire MBV-type vi-rus infections, but these have not been associ-ated with severe pathology, or developed non-monodon reservoirs.
C.9.1.3 Geographical Distribution
BP is found throughout the Americas from theGulf of Mexico to Central Brazil on the East Coastand from Peru to Mexico on the Pacific Coast.BP has also been found in wild shrimp in Hawaii.Multiple strains of BP are recorded within thisgeographic range.
MBV has been reported from Australia, East Af-rica, the Middle East, many Indo-Pacific coun-tries and from south and eastern Asia. MBV-typeviruses have also been found in sites associatedwith P. monodon culture in the Mediterranean andWest Africa, Tahiti and Hawaii, as well as sev-eral locations in North and South America andthe Caribbean.
C.9.2 Clinical Aspects
The impact of BP varies from species to spe-cies. Penaeus aztecus and P. vannamei arehighly susceptible. Penaeus stylirostris is mod-erately susceptible and P. monodon and P.setiferus appear to be resistant/tolerant. In sus-ceptible species, BP infection is characterised
by a sudden onset of high morbidity and mor-tality in larval and post larval stages. Growthrates decrease, the shrimp stop feeding, ap-pear lethargic and show signs of epibiont foul-ing (due to reduced grooming activity). The vi-rus attacks the nuclei of hepatopancreas epi-thelia but can also infect mid-gut epithelia. Al-though infections may be chronic to acute, withhigh cumulative mortality, presence of the BPvirus is not always associated with disease andpost-larvae older than 63 days show no clini-cal signs of infection (see C.9.6).
MBV causes similar clinical signs to BP, due tosimilar infection of the hepatopancreatic andmid-gut epithelial nuclei. Infections of MBV mayalso occur in the lymphoid organ. Larval stagesof P. monodon are particularly susceptible, how-ever, prevalences of >45% may be present injuvenile and adult developmental stages withno overt clinical effects.
C.9.3 Screening Methods
More detailed information on methods forscreening NPB can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000), at http://www.oie.int, or at selected ref-erences.
C.9.3.1 Presumptive
There are no presumptive screening methodsfor asymptomatic carriers of BP and MBV, sincedirect microscopic methods (C.9.3.2) demon-strating the characteristic occlusion bodies (tet-rahedral for BP and spherical-ovoid for MBV)are considered to be confirmatory.
C.9.3.2 Confirmatory
C.9.3.2.1 Wet Mount of Fresh Tissue(Level I/II)
BP infections can be confirmed by bright-fieldor phase contrast microscopic observation ofsingle or multiple tetrahedral (polyhedral) inclu-sion (occlusion) bodies (Fig. C.9.3.2.1a) withinenlarged nuclei of hepatopancreas or midgutepithelia. These bodies can range in size from0.1-20.0 µm (modal range = 8-10 µm) along theperpendicular axis from the base of thepyrimidal shape to the opposite point.
MBV infections observed using the same mi-croscope apparatus appear as single or mul-tiple spherical or sub-spherical inclusion bod-ies within enlarged nuclei of hepatopancreas
Fig. C.9.3.2.1a. Wet mount of feces from a P.vannamei infected with BP showing tetrahedralocclusion bodies (arrows) which are diagnos-tic for infection of shrimp’s hepatopancreas ormidgut epithelial cells. Phase contrast, no stain.700x magnification.
(DV Lightner)
Fig. C.9.3.2.1b,c. Mid and high magnificationviews of tissue squash preparations of thehepatopancreas (HP) from PL of P. monodonwith MBV infections. Most HP cells in both PLsusually display multiple, generally spherical, in-tranuclear occlusion bodies (arrow) that are di-agnostic for MBV. 0.1% malachite green. 700x(b) and 1 700x (c) magnifications.
(DV Lightner)
Fig. C.9.3.2.3a,b. a. Mid-magnification view ofmid-sagittal sections of PL of P. vannamei withsevere BP infections of the hepatopancreasshowing multiple eosinophilic BP tetrahedralocclusion bodies within markedly hypertrophiedhepatopnacreas (HP) cell nuclei (arrows).Mayer-Bennett H&E. 700x magnification; b.High magnification of an HP tubule showingseveral BP-infected cells that illustrate well theintranuclear, eosinophilic, tetrahedral occlusionbodies of BP (arrows). Mayer-Bennett H&E.1800x magnification.
>
b
a
c
b
203
C.9.4.1 Presumptive
C.9.4.1.1 Gross Observations (Level I)
Gross signs of BP vary between susceptiblespecies but include decreased growth, cessa-tion of feeding and preening, lethargy and in-creased epibiont fouling. Some shrimp mayexhibit a white mid-gut line through the ventralabdominal cuticle. None of these symptoms arespecific to BP, but can be considered suspectin susceptible species and at early developmen-tal/post-larval stages which have a history ofbeing affected by BP.
MBV causes similar clinical signs to BP, butprincipally affects larval of P. monodon with aninverse correlation between larval age andpathogenic effects. Adults can be infected withno overt signs (see C.9.3). As with BP, thesesigns are not specific to MBV.
C.9.4.2 Confirmatory
C.9.4.2.1 Wet Mount of Fresh Tissue(Level I/II)
As described for C.9.3.2.1.
C.9.4.2.2 Faecal Examination (Level I/II)
As described for C.9.3.2.2.
C.9.4.2.3 Histopathology (Level II)
As described for C.9.3.2.3.
C.9.4.2.4 Autofluorescence with phloxinestain (Level II)
An aqueous solution of 0.001% phloxine usedon tissue squash preparations or faeces, willcause occlusion bodies of both BP and MBVto fluoresce yellow-green when examined us-ing a fluorescent microscope (barrier filter 0-515 nm and exciter filter of 490 nm) (Thurmanet al. 1990). The same effect is achieved using0.005% phloxine in routine haematoxylin andeosin stain of histological tissue preparations.
C.9.4.2.5 Transmission Electron Micros-copy (TEM) (Level III)
BP virions are rod-shaped with an envelopednucleocapsid measuring 286-337 nm x 56-79nm. The virions are found either free or occludedwithin a crystalline protein matrix (the occlu-sion body). In early infections, virions are found
or midgut epithelia. MBV occlusion bodiesmeasure 0.1 –20.0 µm in diameter (Fig.C.9.3.2.1b,c). The occlusion bodies can bestained using a 0.05% aqueous solution ofmalachite green, which stains them moredensely than surrounding, similarly sized spheri-cal bodies (cell nuclei, secretory granules, lipiddroplets, etc.).
C.9.3.2.2 Faecal Examination (Level I/II)
Make wet mounts of faecal strands and exam-ine for occlusion bodies, as described for freshtissue mounts (C.9.3.2.1).
C.9.3.2.3 Histopathology (Level II)
Tissues from live or moribund (but not dead,due to rapid liquefaction of the target organ –the hepatopancreas) shrimp should be fixed inDavidson’s fixative to ensure optimum fixationof the hepatopancreas (10% buffered formalinprovides sub-optimal hepatopancreas preser-vation). The fixative should be administered bydirect injection into the hepatopancreas. Thecuticle should be cut along the dorsal line ofthe cephalothorax to enhance fixative penetra-tion of the underlying tissues and the tissuesshould be fixed for 24-48 hr before transfer to70% ethanol for storage. The tissues can thenbe processed for routine paraffin embedding,sectioning at 5-7 µm thickness and staining withHarris’ haematoxylin and eosin or other Giemsaor Gram tissue-staining methods. Brown andBrenn’s histological Gram stain provides intensered or purple colouration of both MBV (see alsoFig. C.5.4.2.1d – C.5) and BP occlusion bodies(Fig. C.9.3.2.3a,b), aiding in their differentiationfrom surrounding tissues.
Two primers sequences are available for theMBV polyhedrin gene (Lu et al 1993) and an-other pair are available for a 1017bp fragmentof the viral genome (Mari et al 1993). Details onthe PCR procedures for screening tissue or fae-cal samples are provided in the OIE DiagnosticManual (OIE 2000) or selected references(C.9.7).
C.9.4 Diagnostic Methods
More detailed information on methods for di-agnosis of NPB can be found in the OIE Diag-nostic Manual for Aquatic Animal Diseases (OIE2000), at http://www.oie.int, or at selected ref-erences.
in association with nuclear enlargements, ab-errant stromatic patterns of the nucleoplasm,degenerate nucleoli, and nuclear membraneproliferation into labyrinths. Occlusion bodiesoccur during later stages of infection.
MBV has been shown to have two types ofocclusion bodies using electron microscopicexaminations (Ramasamy et al. 2000). Type 1has a paracrystalline array of polyhedrin unitswithin a lattice work spacing of 5-7 nm, whichcontains occluded virions (along with a fewperipheral non-occluded virions) that have adouble envelope and measure 267 ± 2 x 78 ± 3nm. Type 2 occlusion bodies consist of non-crystalline, granulin-like sub-units 12 nm in di-ameter, containing mostly non-occluded virionsmeasuring 326 ± 4 x 73 ± 1 nm. In addition, anon-enveloped stage has recently been de-tected (Vickers et al. 2000) in the cytoplasm ofinfected cells and close association with thenuclear membrane.
C.9.4.2.6 In situ Hybridization (Level III)
Details of the preparation and analytical proce-dures requried for in situ hybridisation for con-firming BP and MBV infections are provided inthe OIE Diagnostic Manual (OIE 2000a) underboth the Nuclear Polyhedrosis Baculoviroseschapter (Chapter 4.2.2) as well as the InfectiousHypodermal and Haematopoietic Necrosischapter (Chapter 4.2.3).
C.9.5 Modes of Transmission
BP and MBV are both transmitted orally viauptake of virus shed with the faeces of infectedshrimp (C.9.3.2.2), or cannibalism on dead anddying shrimp. Infected adults have also beenshown to infect their offspring via faecal con-tamination of the spawned egg masses.
C.9.6 Control Measures
Overcrowding, chemical and environmentallyinduced stress, have all been shown to increasethe virulence of MBV and BP infections in sus-ceptible shrimp species under culture condi-tions.
Exposure of stocks to infection can be avoidedby pre-screening the faeces of potentialbroodstock and selecting adults shown to befree of faecal contamination by occlusion bod-ies of either baculovirus. Prevention of infec-tions may also be achieved by surface disin-fection of nauplii larvae or fertilised eggs with
formalin, iodophore and filtered clean seawa-ter as follows:• Collect nauplii and wash in gently running sea
water for 1-2 minutes.• Immerse the nauplii in a 400 ppm solution of
formalin for 1 minute followed by a solutionof 0.1 ppm iodine for an additional minute.The same procedure can be used on fertilisedeggs except the formalin concentration is re-duced to 100ppm.
• Rinse the treated nauplii in running sea wa-ter for 3-5 min and introduce to the hatchery.
Eradication of clinical outbreaks of BP and MBVmay be possible in certain aquaculture situa-tions by removal and sterile disposal of infectedstocks, disinfection of the culture facility, theavoidance of re-introduction of the virus (fromother nearby culture facilities, wild shrimp, etc.).
C.9.7 Selected References
Alcivar-Warren,A., R.M. Overstreet, A.K. Dhar,K. Astrofsky, W.H. Carr, J. Sweeny and J.M.Lotz. 1997. Genetic susceptibility of culturedshrimp (Penaeus vannamei) to infectioushypodermal and hematopoietic necrosis vi-rus and Baculovirus penaei: Possible relation-ship with growth status and metabolic geneexpression. J. Invertebr. Pathol. 70(3): 190-197.
Belcher, C.R. and P.R. Young. 1998.Colourimetric PCR-based detection ofmonodon baculovirus in whole Penaeusmonodon postlarvae. J. Virol. Methods 74(1):21-29.
Brock, J.A., D.V. Lightner and T.A. Bell. 1983. Areview of four virus (BP, MBV, BMN, andIHHNV) diseases of penaeid shrimp with par-ticular reference to clinical significance, di-agnosis and control in shrimp aquaculture.Proc. 71st Intl. Council for the Exploration ofthe Sea, C.M. 1983/Gen: 10/1-18.
Brock, J.A., L.K. Nakagawa, H. Van Campen,T. Hayashi, S. Teruya. 1986. A record ofBaculovirus penaei from Penaeus marginatusRandall in Hawaii. J. Fish Dis. 9: 353-355.
Bruce, L.D., B.B. Trumper, and D.V. Lightner.1991. Methods of viral isolation and DNAextraction for a penaeid shrimp baculovirus.J. Virol. Meth. 34:245-254.
Bruce, L.D., R.M. Redman and D.V. Lightner.1994. Application of gene probes to deter-mine target organs of a penaeid shrimp
baculovirus using in situ hybridisation.Aquaculture 120(1-2): 45-51.
Bruce, L.D., D.V. Lightner, R.M. Redman andK.C. Stuck. 1994. Comparison of traditionaland molecular tools for Baculovirus penaeiinfections in larval Penaeus vannamei. J.Aquatic Anim. Health 6(4): 355-359.
Bueno, S.L., R.M. Nascimento and I.Nascimento. 1990. Baculovirus penaei infec-tion in Penaeus subtilis: A new host and anew geographic range of the disease. J.World Aquacult. Soc. 21(3): 235-237.
Chen, S.N., P.S. Chang and G.S. Kou. 1993.Diseases and treatment strategies onPenaeus monodon in Taiwan. pp. 43-57 In:Proceedings of the Symposium on Aquacul-ture held in Beijing, 21-23 December 1992,Taiwan Fisheries Research Institute, Keelung,TRFI Conf. Proc. #3.
Chen, S.N., P.S. Chang, C.C. Chen and G.H.Kou. 1993. Studies on infection pathway ofMonodon Baculovirus (MBV). COA Fish. Ser.40: 81-85.
Chen, X., D. Wu, H. Huang, X. Chi and P. Chen.1995. Ultrastructure on Penaeus monodonbaculovirus. J. Fish. China (Shuichan Xuebao)19(3): 203-209.
Fegan, D.F., T.W. Flegel, S. Sriurairatana andM. Waiyakruttha. 1991. The occurrence, de-velopment and histopathology of monodonbaculovirus in Penaeus monodon in Thailand.Aquac. 96(3-4): 205-217.
Flegel, T.W., V. Thamavit, T. Pasharawipas andV. Alday-Sanz. 1999. Statistical correlationbetween severity of hepatopancreaticparvovirus infection and stunting of farmedblack tiger shrimp (Penaeus monodon).Aquac.174(3-4): 197-206.
Hammer, H.S., K.C. Stuck and R.M. Overstreet.1998. Infectivity and pathogenicity ofBaculovirus penaei (BP) in cultured larval andpostlarval Pacific white shrimp, Penaeusvannamei, related to the stage of viral devel-opment. J. Invertebr. Pathol. 72(1): 38-43.
Hao, N.V., D.T. Thuy, L.T. Loan, L.T.T. Phi, L.H.Phuoc, H.H.T. Corsin and P. Chanratchakool.
Presence of the two viral pathogens WSSVand MBV in three wild shrimp species(Penaeus indicus, Metapenaeus ensis andMetapenaeus lysianassa). Asian Fish. Sci.
12(4): 309-325.
Hsu, Y.L., K.H. Wang, Y.H. Yang, M.C. Tung,C.H. Hu, C.F. Lo, C.H. Wang and T. Hsu. 2000.Diagnosis of Penaeus monodon-typebaculovirus by PCR and by ELISA. Dis.Aquatic Org. 40(2): 93-99.
Karunasagar, I., S.K. Otta and I. Karunasagar.1998. Monodon baculovirus (MBV) and bac-terial septicaemia associated with massmortality of cultivated shrimp (Penaeusmonodon) from the east coast of India. In-dian J. Virol. 14(1): 27-30.
LeBlanc, B.D. and R.M. Overstreet. 1990.Prevalence of Baculovirus penaei in experi-mentally infected white shrimp (Penaeusvannamei) relative to age. Aquac. 87(3-4):237-242.
LeBlanc, B.D. and R.M. Overstreet. 1991. Ef-fect of dessication, pH, heat and ultravioletirradiation on viability of Baculovirus penaei.J. Invertebr. Pathol. 57(2): 277-286.
LeBlanc, B.D. and R.M. Overstreet. 1991. Effi-cacy of calcium hypochlorite as a disinfec-tant against the shrimp virus Baculoviruspenaei. J. Aquatic Anim. Health 3(2): 141-145.
LeBlanc, B.D., R.M. Overstreet and J.M. Lotz.1991. Relative susceptibility of Penaeusaztecus to Baculovirus penaei. J. WorldAquacult. Soc. 22(3): 173-177.
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDisease of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Lightner, D.V. and R.M. Redman. 1989.Baculovirus penaei in Penaeus stylirostris(Crustacea: Decapoda) cultured in Mexico:Unique cytopathology and a new geographicrecord. J. Invertebr. Pathol. 53(1): 137-139.
Lightner, D.V., R.M. Redman, and E.A. AlmadaRuiz. 1989. Baculovirus penaei in Penaeusstylirostris (Crustacea: Decapoda) cultured inMexico: unique cytopathology and a newgeographic record. J. Inverteb. Pathol.53:137-139.
Lu, C.C., K.F.J. Tang, G.H. Kou and S.N. Chen.1995. Detection of Penaeus monodon-type
baculovirus (MBV) infection in Penaeusmonodon Fabricius by in situ hybridisation.J. Fish Dis. 18(4): 337-345.
Lu, C.C., K.F.J. Tang and S.N. Chen. 1996.Morphogenesis of the membrane labyrinthin penaeid shrimp cells infected with Penaeusmonodon-baculovirus (MBV). J. Fish Dis.19(5): 357-364.
OIE. 2000. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
Poulos, B.T., J. Mari, J-R. Bonami, R. Redmanand D.V. Lightner. 1994. Use of non -radio-actively labeled DNA probes for the detec-tion of a baculovirus from Penaeus monodonby in situ hybridisation on fixed tissues. J.Virol. Methods 49(2): 187-194.
Ramasamy, P., P.R. Rajan, V. Purushothamanand G.P. Brennan. 2000. Ultrastructure and
pathogenesis of Monodon baculovirus (PmSNPV) in cultuerd larvae and natural brood-ers of Penaeus monodon. Aquac.184(1-2):45-66.
Shariff, M., R.P. Subasinghe and J.R. Arthur(eds) (1992) Diseases in Asian Aqaculture.Proceedings of the First Symposium on Dis-eases in Asian Aquaculture, Bali 1990. FishHealth Section, Asian Fisheries Society, Ma-nila, Philippines, 585pp.
Spann, K.M., R.J.G. Lester and J.L. Paynter.1993. Efficiency of chlorine as a disinfectantagainst Monodon baculovirus. Asian Fish.Sci. 6(3): 295-301.
Stuck, K.C. and R.M. Overstreet. 1994. Effectof Baculovirus penaei on growth and survivalof experimentally infected postlarvae of thePacific white shrimp, Penaeus vannamei. J.Invertebr. Pathol. 64(1): 18-25.
Stuck, K.C. and S.Y. Wang. 1996. Establishmentand persistence of Baculovirus penaei infec-tions in cultured Pacific white shrimpPenaeus vannamei. J. Invertebr. Pathol. 68(1):59-64.
Stuck, K.C., L.M. Stuck, R.M. Overstreet andS.Y. Wang. 1996. Relationship between BP
(Baculovirus penaei) and energy reserves inlarval and postlarval Pacific white shrimpPenaeus vannamei. Dis. Aquat. Org. 24(3):191-198.
Thurman, R.B., T.A. Bell, D.V. Lightner and S.Hazanow. 1990. Unique physicochemicalproperties of the occluded penaeid shrimp
baculoviruses and their use in diagnosis ofinfections. J. Aquat. Anim. Health 2(2): 128-131.
Vickers, J.E., J.L. Paynter, P.B. Spradbrow andR.J.G. Lester. 1993. An impression smearmethod for rapid detection of Penaeusmonodon-type baculovirus (MBV) in Austra-lian prawns. J. Fish Dis. 16(5): 507-511.
Vickers, J.E., R. Webb and P.R. Young. 2000.Monodon baculovirus from Australia: ultra-structural observations. Dis. Aquat. Org.39(3): 169-176.
Wang, S.Y., C. Hong and J.M. Lotz. 1996. De-velopment of a PCR procedure for the de-tection of Baculovirus penaei in shrimp. Dis.Aquat. Org. 25(1-2): 123-131.
Necrotising Hepatopancreatitis (NHP) is causedby a bacterium that is relatively small, highly pleo-morphic, Gram negative, and an apparent obli-gate intracellular pathogen. The NHP bacteriumhas two morphologically different forms: one is asmall pleomorphic rod and lacks flagella; whilethe other is a longer helical rod possessing eightflagella on the basal apex of the bacterrium, andan additional flagellum (or possibly two) on thecrest of the helix.. The NHP bacterium occupiesa new genus in the alpha Proteobacteria, and isclosely related to other bacterial endosymbiontsof protozoans. NHP is also known as Texasnecrotizing hepatopancreatitis (TNHP), TexasPond Mortality Syndrome (TPMS) and Peru ne-crotizing hepatopancreatitis (PNHP). More infor-mation about the disease is found in Lightner(1996).
C.10.1.2 Host Range
NHP can infect both Penaeus vannamei and P.stylirostris but causes higher mortalities in theformer species. NHP has also been reported inP. aztecus, P. californiensis and P. setiferus.
C.10.1.3 Geographic Distribution
NHP was first described in Texas in 1985. Otheroutbreaks have been reported in most LatinAmerican countries on both the Pacific and At-lantic Ocean coasts, including Brazil, Costa Rica,Ecuador, Mexico, Panama, Peru and Venezu-ela.
C.10.2 Clinical Aspects
The NHP bacterium apparently infects only theepithelial cells lining the hepatopancreatic tu-bules, and, to date, no other cell type has beenshown to become infected. The hepatopancreasin shrimp is a critical organ involved in food di-gestion, nutrient absorption and storage, and anyinfection has obvious and serious consequencesfor the affected animal, from reduced growth todeath. Various environmental factors appear tobe important for the onset of NHP clinical signs;the most prominent ones are water salinity over16 ppt (parts per thousand) and water tempera-ture of 26˚C or higher.
BACTERIAL DISEASE OF SHRIMPC.10 NECROTISING
HEPATOPANCREATITIS (NHP)
C.10.3 Screening Methods
C.10.3.1 Confirmatory
C.10.3.1.1 Dot Blot for AsymptomaticAnimals (Level III)
A commercial dot blot detection kit is availablefor NHP from DiagXotics (Wilton, CT, USA).
C.10.3.1.2 In situ Hybridization (Level III)
A commercial in situ hybridization detection kitis available for NHP from DiagXotics (Wilton,CT, USA).
Samples of hepatopancreas are fixed in 70%ethanol and triturated prior to processing. DNAis isolated as follows: 25 mg of the trituratedhepatopancreas is suspended in 250 µl of di-gestion buffer (50 mM Tris, 20 mM EDTA, 0.5%SDS, pH 8.5) in 0.5 ml eppendorf tubes. Pro-teinase K (7.5 µl of a 20 mg ml-1 stock solution)is added and the tube incubated at 60°C for2 h with periodic vortexing. The tube is thenincubated at 95°C for 10 min to inactivate theproteinase K. The tube is then centrifuged for 3min at 13,000 rpm (16,000 x g) and 75 µl of thesupernatant applied to a CHROMA SPIN TE-100 (Clontech Labs) column and centrifuged ina horizontal rotor according to themanufacturer’s protocol. The solution collectedby centrifugation is diluted 1:100 and 1:1000 indistilled water prior to use in the PCR assay.
Below is the sequence of oligonucleotide prim-ers used for amplifying variable regions of the16S rRNA sequence:
Forward: 5'-ACG TTG GAG GTT CGT CCT TCA G-3'
Reverse1 5'-TCA CCC CCT TGC TTC TCA TTG T-3'
Reverse2 5'-CCA GTC ATC ACC TTT TCT GTG GTC-3'
The forward primer and reverse primer 1 am-plify a 441 bp fragment, the forward primer andreverse primer 2 amplify a 660 bp fragent. PCRis performed in 50 µl reactions containing 10mM Tris-HCl (pH 8.3), 50mM KCl, 1.5 mM MgCl,200 mM deoxynucleotides, 0.5 m?M of the for-ward and the paired reverse primers and 0.03to 0.3 µg of template DNA. The reactants areheated to 94°C in a programmable thermocycler
208
(DV Lightner)
Fig. C.10.4.1.1. Juvenile P. vannamei with NHPshowing markedly atrophied hepatopancreas,reduced to about 50% of its normal volume.
(DV Lightner)
Fig. C.10.4.1.2. Wet- mount of the HP of in-fected shrimp with inflamed hemocyte, mela-nized HP tubules and absence of lipid drop-lets. No stain. 150x magnification.
(DV Lightner)
Fig. C.10.4.1.3a,b. Low and mid-magnificationof photographs of the HP of a severely NHPinfected juvenile P. vannamei. Severehemocytic inflammation of the intratubularspaces (small arrow) in response to necrosis,cytolysis and sloughing of HP tubule epithelialcells (large arrow), are among the principal his-topathological changes due to NHP. Mayer-Bennett H&E. 150x (a) and 300x (b) magnifica-tions.
C.10 Necrotising Hepatopancreatitis(NHP)
(DV Lightner)
Fig. C.10.4.1.3c. Low magnification view of theHP of a juvenile P.vannamei with severe, chronicNHP. The HP tubule epithelium is markedly at-rophied, resulting in the formation of largeedematous (fluid filled or “watery areas in theHP. Mayer-Bannett H & E. 100x magnification.
>
b
a
209
C.10 Necrotising Hepatopancreatitis(NHP)
prior to adding 1.25 U of Amplitaq DNA poly-merase. The final solution is then overlayed withmineral oil. The amplification profile consists of35 cycles of 30 s at 94°C, 30 s at 58°C and 1min at 72°C with an additional 5 min at 72°Cfollowing the final cycle. PCR products is ex-amined by electrophoresis in 1% agarose in TAEbuffer containing 0.5 m?g ml-1 ethidium bro-mide.
C.10.4 Diagnostic Methods
More detailed information on methods for di-agnosis of NHP can be found in Lightner (1996)or in selected references.
C.10.4.1 Presumptive
C.10.4.1.1 Gross Observations (Level 1)
A wide range of gross signs can be used toindicate the possible presence of NHP. Theseinclude: lethargy, reduced food intake, higherfood conversion ratios, anorexia and emptyguts, noticeable reduced growth and poorlength weight ratios (“thin tails”); soft shells andflaccid bodies; black or darkened gills; heavysurface fouling by epicommensal organisms;bacterial shell disease, including ulcerative cu-ticle lesions or melanized appendage erosion;and expanded chromatophores resulting in theappearance of darkened edges in uropods andpleopods. The hepatopancreas may be atro-phied (Fig.C.10.4.1.1) and have any of the fol-lowing characteristics: soft and watery; fluidfilled center; paled with black stripes (melanizedtubules); pale center instead of the normal tanto orange coloration. Elevated mortality ratesreaching over 90% can occur within 30 days ofonset of clinical signs if not treated.
C.10.4.1.2 Wet Mounts (Level II)
Wet mounts of the hepatopancreas of shrimpwith NHP may show reduced or absent lipiddroplets and/or melanized hepatopancreas tu-bules (Fig.C.10.4.1.2).
C.10.4.1.3 Histopathology (Level II)
NHP is characterised by an atrophied hepato-pancreas showing moderate to extreme atro-phy of the tubule mucosa and the presence ofthe bacterial forms through histological prepa-rations. Principal histopathological changes dueto NHP include hemocytic inflammation of theintertubular spaces in response to necrosis,cytolysis, and sloughing of hepatopancreas
tubule epithelial cells (Fig. C.10.4.1.3a,b). Thehepatopancreas tubule epithelium is markedlyatropied, resulting in the formation of largeedematous (fluid filled or “watery”) areas in thehepatopancreas (Fig.C.10.4.1.3c).Tubule epi-thelial cells within granulomatous lesions aretypically atrophied and reduced from simplecolumnar to cuboidal in morphology. They con-tain little or no stored lipid vacuoles(Fig.C.10.4.1.3d) and markedly reduced or nosecretory vacuoles.
C.10.4.2 Confirmatory
C.10.4.2.1 Transmission Electron Micros-copy (TEM) (Level III)
Two distinct versions of the NHP bacteriumoccur in infected hepatopancreatic cells. Thefirst is a rod-shaped rickettsial-like form mea-suring 0.3 µm x 9 µm which lacks flagella. Thesecond is a helical form (Fig.C.10.4.2.1) mea-suring 0.2 µm x 2.6-2.9 µm which has eightperiplasmic flagella at the basal apex of thebacterium and an additional 1-2 flagella on thecrest of the helix.
C.10.4.2.2 Dot Blot for AsymptomaticAnimals (Level III)
A commercial dot blot detection kit is availablefor NHP from DiagXotics (Wilton, CT, USA).
C.10.4.2.3 In situ Hybridization (Level III)
A commercial in situ hybridization detection kitis available for NHP from DiagXotics (Wilton,CT, USA).
Early detection of clinical NHP is important forsuccessful treatment because of the potentialfor cannibalism to amplify and transmit the dis-ease. Molecular testing of PL from infectedbroodstock indicates that vertical transmissiondoes not occur.
C.10.6 Control Measures
Periodic population sampling and examination(through histopathology, TEM or commercialgene probe) are highly recommended in farms
210
(DV Lightner)
Fig. C.10.4.1.3d. The HP tubule epithelial cellsshow no cytoplasmic lipid droplets, but insteadcontain masses of the tiny, non-membranebound, intracytoplasmic NHP bacteria (arrow).Mayer-Bennett H&E. 1700x magnification.
(DV Lightner)
Fig. C.10.4.2.1. Low magnification TEM of ahepatopancreatocyte from a juvenile P.vannamei with NHP. Profiles of intracellular rod-shaped forms (large arrow) and helical forms(small arrow) of the NHP bacterium are abun-dant in the cytoplasm. 10 000x magnification.
with a history of NHP occurrence and whereenvironmental conditions favor outbreaks. Theuse of the antibiotic oxytetracycline (OTC) inmedicated feeds is probably the best NHP treat-ment currently available, particularly if diseasepresence is detected early.
There is also some evidence that deeper pro-duction ponds (2 m) and the use of hydratedlime (Ca(OH)2) to treat pond bottoms duringpond preparation before stocking can help re-duce NHP incidence. Preventive measures caninclude raking, tilling and removing pond bot-tom sediments, prolonged sun drying of pondsand water distribution canals for several weeks,disinfection of fishing gear and other farmequipment using calcium hypochlorite and dry-ing and extensive liming of ponds.
C.10.7 Selected References
Brock, J.A. and K. Main. 1994. A Guide to theCommon Problems and Diseases of Cul-tured Penaeus vannamei. Oceanic Institute,Makapuu Point, Honolulu, Hawaii. 241p.
Frelier,P.F., R.F. Sis,T.A. Bell and D.H. Lewis.1992. Microscopic and ultrastructural stud-ies of necrotizing hepatopancreatitis in Texascultured shrimp (Penaeus vannamei). Vet.Pathol. 29:269-277.
Lightner, D.V. 1996. A Handbook of ShrimpPathology and Diagnostic Procedures forDiseases of Cultured Penaeid Shrimp. WorldAquaculture Society, Baton Rouge, LA. 304p.
Lightner, D.V., R.M. Redman and J.R. Bonami.1992. Morphological evidence for a singlebacterial aetiology in Texas necrotizinghepatopancreatitis in Penaeus vannamei(Crustacea:Decapoda). Dis. Aquat. Org.13:235-239.
C.10 Necrotising Hepatopancreatitis(NHP)
211
C.11.1 Background Information
C.11.1.1 Causative Agent
Crayfish Plague (also known as Krebspest,Kraftpest, ‘la peste’ or ‘crayfishaphanomyciasis’) is caused by the Oomycetefungus, Aphanomyces astaci. This is a closerelative of species associated with serious fin-fish diseases, such as A. invadans, in EpizooticUlcerative Syndrome (EUS) of South-East Asia(see section F.11).
C.11.1.2 Host Range
Crayfish plague affects the Noble crayfishAstacus astacus of north-west Europe, thestone crayfish Austropotamobius pallipes ofsouth-west and west Europe, the mountaincrayfish Austropotamobius torrentium of south-west Europe, and the slender clawed or Turk-ish crayfish Astacus leptodactylus of easternEurope and Asia Minor. The Chinese mitten crab(Eriocheir sinensis) can be infected experimen-tally. North American crayfish (Pacifasticusleniusculus, the signal crayfish, andProcambarus clarkii, the Louisiana swamp cray-fish) can also be infected by A. astaci, but arerelatively tolerant of the disease, only exhibit-ing clinical signs under intensive culture condi-tions.
C.11.1.3 Geographical Distribution
Aphanomyces astaci is widespread in Europe,as well as in North America. The disease firstappeared in northern Italy in the mid 19th cen-tury, and then spread down to the Balkans andBlack Sea, as well as into Russia, Finland andSweden. In the 1960’s the disease appeared inSpain with further spread to the British Isles, Tur-key, Greece and Norway in the 1980’s.
C.11.2 Clinical Aspects
The hyphae of A. astaci grow throughout the non-calcified parts of the cuticle and may extend alongthe nerve cord. The more disease tolerant spe-cies of crayfish (North American) encapsulate thefungal hyphae within melanised nodules, arrest-ing the hyphal proliferation. Susceptible speciesappear incapable of producing such a defensereaction, and the fungus proliferates throughoutthe epicuticle and exocuticular layers of the ex-oskeleton. The cuticle and related soft-tissuedamage leads to death which, under warm wa-ter conditions, can be rapid and result in 100%mortality. Resistant North American species that
FUNGAL DISEASE OF CRAYFISHC.11 CRAYFISH PLAGUE
survive initial infection can become sub-clini-cal carriers of the fungus. Under adverse hold-ing conditions, however, such infections maybecome pathogenic.
C.11.3 Screening Methods
More detailed information on methods forscreening crayfish plague can be found in theOIE Diagnostic Manual for Aquatic Animal Dis-eases (OIE 2000), at http://www.oie.int or se-lected references.
C.11.3.1 Presumptive
C.11.3.1.1 Gross Observations (Level I)
Melanized spots in the cuticle of any crayfishspecies may be indicative of crayfish plaguesurvival. Such crayfish should be consideredto be potential carriers of the disease andscreened for Aphanomyces astaci using con-firmatory diagnostic techniques (C.11.3.2 andC.11.4.2).
C.11.3.1.2 Microscopy (Level I/II)
Foci of infection as described under C.12.3.1.1,may not be readily visible. Examination using adissecting microscope may reveal small whit-ened patches in the muscle tissues underlyingthin spots in the cuticle. There may also bebrownish discolouration of the cuticle. Finebrown lines through the cuticle should also beconsidered as suspect fungal hyphae. The ar-eas that should be examined closely are theintersternal soft-ventral cuticle of the abdomenand tail; the cuticle between the carapace andtail, the joints of the periopods (especially theproximal joints), the perianal cuticle and the gills.
C.11.3.2 Confirmatory
C.11.3.2.1 Culture (Level II)
The fungus can be isolated from suspect cu-ticle and tissues using an agar medium thatcontains yeast extract, glucose and antibiotics(penicillin G and oxolinic acide) made up withnatural (not demineralised) river water. Identifi-cation to species requires morphologicalcharacterisation of the sexual reproductiveparts of the fungus, however, these stages areabsent in A. astaci, thus, confirmation of infec-tion is usually based on isolation of fungalcolonies with the following characteristics (sinceno other closely-related Oomycetes are knownto infect crayfish):
212
• growth within the agar medium (unless cul-tured at < 7°C, which promotes superficialgrowth);
phae, 7-9 µm in diameter (min-max 5-10 µm);• young hyphae are densely packed with
coarse, granular cytoplasm and containhighly refractile globules;
• older hyphae are highly vacuolated and theoldest hyphae appear to be empty
When thalli are transferred from the culturemedium to sterile distilled water, they developsporangia within 12-15 h (20°C) or 20-30 h(16°C). Elongate, irregularly amoeboid shapedspores are released and rapidly encyst as amass around the sporangial tip(Fig.C.11.3.2.1a). Encysted primary sporesmeasure 9-11 µm in diameter (min-max 8-15µm). Release of the secondary zoospores oc-curs from papillae that develop on the surfaceof the primary spore cyst. This occurs at tem-peratures as low as 4°C, peaking at 20°C andstopping at temperatures >24°C. Thezoospores have lateral flagella and measure 8x 12 µm. More details on culture media, tech-niques and developmental stage morphologyare provided in the OIE Manual (OIE 2000).
C.11.3.2.2 Bioassay (Level I/II)
Confirmation of crayfish plague can be doneusing zoospores cultured from fungal isolatesfrom suspect crayfish tissues. Rapid mortali-ties in the susceptible crayfish, along with re-isolation of the fungus as described above,should be considered conclusive for A. astaci.
C.11.4 Diagnostic Methods
More detailed information on methods for di-agnosis of crayfish plague can be found in theOIE Diagnostic Manual for Aquatic Animal Dis-eases (OIE 2000), at http://www.oie.int or se-lected references.
There is no other disease, or pollution effect,that can cause total mortality of crayfish butleave all other animals in the same water un-harmed. In such situations and with known sus-ceptible species, presumptive diagnosis can befairly conclusive. In first-time cases or in situa-tions with resistant species, however, confirma-tory isolation of the pathogen is recommended.
C.11.4.1 Presumptive
C.11.4.1.1 Gross observation (Level I)
Large numbers of crayfish showing activity dur-ing daylight should be considered suspect,since crayfish are normally nocturnal. Somemay show uncoordinated movement, easily tiponto their backs, and be unable to right them-selves.
Gross clinical signs of crayfish plague vary fromnone to a wide range of external lesions. White
(EAFP/DJ Alderman)
Fig. C.11.3.2.1a. Fresh microscopic mount ofa piece of infected exoskeleton showing fun-gal spores.
(EAFP/DJ Alderman)
Fig. C.11.4.1.1a,b. Clinical signs of infectedcrayfish showing whitened necrotic muscula-ture in the tail, and often accompanied inchronic infections by melanisation (blackening)of affected exoskeleton.
C.11 Crayfish Plague
b
a
213
avoided or undertaken with disinfection precau-tions. Sodium hypochlorite and iodophores canbe used to disinfect equipment and thoroughdrying (>24 hours) is also effective, sinceoomycetes cannot withstand desiccation.
C.11.7 Selected References
Alderman, D.J. 1996. Geographical spread ofbacterial and fungal diseases of crustaceans.OIE International Conference on the preven-tion of diseases of aquatic animals throughinternational trade. Office International desEpizooties, Paris, France, June 7-9 1995. Rev.Sci. Tech. Off. Int. Epiz. 15: 603-632.
Alderman, D.J. and J.L. Polglase. 1986.Aphanomyces astaci: isolation and culture.J. Fish Dis. 9: 367-379.
Alderman, D.J., J.L. Polglase, . Frayling and J.Hogger. 1984. Crayfish plague in Britain. J.Fish Dis. 7(5): 401-405.
Alderman, D.J., J.L. Polglase and M. Frayling.1987. Aphanomyces astaci pathoogenicityunder laboratory and field conditions. J. FishDis. 10: 385-393.
Alderman, D.J., D. Holdich and I. Reeve. 1990.Signal crayfish as vectors of crayfish plaguein Britain. Aquac. 86(1): 306.
Dieguez-Uribeondo, J., C. Temino and J.L.Muzquiz. 1997. The crayfish plagueAphanomyces astaci in Spain. Bull. Fr. PechePiscic. 1(347): 753-763.
Fuerst, M. 1995. On the recovery of Astacusastacus L. populations after an epizootic ofthe crayfish plague (Aphanomyces astaciShikora). Eighth Int. Symp. Astacol., Louisi-ana State Univ. Printing Office, Baton Rouge,LA, pp. 565-576.
Holdich, D.M. and I.D. Reeve. 1991. Distribu-tion of freshwater crayfish in the British Isles,with particular reference to crayfish plague,alien introductions and water quality. Aquat.Conserv. Mar. Freshwat. Ecosyst, 1(2): 139-158.
Lilley, J.H. and V. Inglis. 1997. Comparative ef-fects of various antibiotics, fungicides anddisinfectants on Aphanomyces invaderis andother saprolegniaceous fungi. Aquac. Res.28(6): 461-469.
patches of muscle tissue underlying transpar-ent areas of cuticle (especially the ventral ab-domen and periopod joints), and focal brownmelanised spots (Fig.C.11.4.1.1a,b), are themost consistent signs.
C.11.4.1.2 Microscopy (Level I/II)
As for C.11.3.1.2.
C.11.4.2 Confirmatory
C.11.4.2.1 Culture (Level II)
As for C.12.3.2.1, diagnosis of crayfish plaguerequires the isolation and characterisation of thepathogen, A. astaci, using mycological mediafortified with antibiotics to control bacterial con-tamination. Isolation is only likely to be success-ful before or within 12 hours of the death ofinfected crayfish.
C.11.4.2.2 Bioassay (Level I/II)
As for C.11.3.2.2.
C.11.5 Mode of Transmission
Transmission is horizontal and direct via themotile biflagellate zoospore stage of A. astaci,which posseses a positive chemotaxis towardscrayfish. The disease can spread downstreamat the speed of flow of the river, and has beendocumented to spread upstream at 2-4 km peryear. The upstream spread is suspected todriven by movements of crayfish between in-fection and the terminal stages of the disease.
Transmission has also been linked to the waterused to move fish between farms, as well as tocontaminated equipment (boots, fishing gear,crayfish traps, etc.). Introductions of NorthAmerican crayfish for crayfish farming are be-lieved to have been the source of the Europeanoutbreaks of crayfish plague.
C.11.6 Control Measures
There is no treatment for crayfish plague, andthe high levels of mortality have precluded natu-ral selection for disease resistance in the mostsusceptible species (some populations are nowendangered). Control of the disease is bestachieved by preventing introductions or escapeof crayfish into unaffected waters. In addition,movement of water or any equipment betweenaffected to unaffected watersheds should be
C.11 Crayfish Plague
214
Lilley, J.H., L. Cerenius and K. Soderhall. 1997.RAPD evidence for the origin of crayfishplague outbreaks in Britain. Aquac. 157(3-4): 181-185.
Nylund, V. and K. Westman. 1995. Fequency ofvisible symptoms of the crayfish plague fun-gus (Aphanomyces astaci) on the signal cray-fish (Pacifasticus leniusculus) in natural popu-lations in Finland in 1979-1988. Eighth Int.Symp. Astacol., Louisiana State Univ. Print-ing Office, Baton Rouge, LA.
Oidtmann, B., M. El-Matbouli, H. Fischer, R.Hoffmann, K. Klaerding, I. Schmidt and R.Schmidt. 1997. Light microscopy of Astacusastacus L. under normal and selected patho-logical conditions, with special emphasis toporcelain disease and crayfish plague. Fresh-water Crayfish 11. A Journal of Astacology,Int. Assoc. Astacology, pp. 465-480.
Oidtmann B., L. Cerenius, I. Schmid, R. Hoffmanand K. Soederhaell. 1999. Crayfish plagueepizootics in Germany – classification of twoGerman isolates of the crayfish plague fun-gus Apahnomyces astaci by random ampli-fication of polymorphic DNA. Dis. Aquat. Org.35(3): 235-238.
OIE. 2000. Diagnostic Manual for Aquatic Ani-mal Diseases, Third Edition, 2000. Office In-ternational des Epizooties, Paris, France.237p.
Reynolds, J.D. 1988. Crayfish extinctions andcrayfish plague in central Ireland. Biol.Conserv. 45(4): 279-285.
Vennerstroem, P., K. Soederhaell andL.Cerenius. 1998. The origin of two crayfishplague (Aphanomyces astaci) epizootics inFinland on noble crayfish, Astacus astacus.Ann. Zool. Fenn. 35(1): 43-46.
Crustacean pathogens Prof. D. LightnerAquaculture Pathology SectionDepartment of Veterinary ScienceUniversity of ArizonaBuilding 90, Room 202Tucson AZ 85721USATel: (1.520) 621.84.14Fax: (1.520) 621.48.99E-mail: [email protected]. S.N. ChenDepartment of ZoologyDirector, Institute of Fishery BiologyNational Taiwan UniversityNo. 1 Roosevelt RoadSection 4 , Taipei, Taiwan 10764TAIWAN PROVINCE of CHINATel: 886-2-368-71-01Fax: 886-2-368-71-22E-mail: [email protected]
216
ANNEX C.AII. LIST OF REGIONAL RESOURCEEXPERTS FOR CRUSTACEAN DISEASES
IN ASIA-PACIFIC1
Expert
Dr. Richard CallinanNSW Fisheries, Regional Veterinary LaboratoryWollongbar NSW 2477AUSTRALIATel (61) 2 6626 1294Mob 0427492027Fax (61) 2 6626 1276E-mail: [email protected]. Indrani KarunasagarDepartment of Fishery MicrobiologyUniversity of Agricultural SciencesMangalore – 575 002INDIATel: 91-824 436384Fax: 91-824 436384E-mail: [email protected]. C.V. MohanDepartment of AquacultureCollege of FisheriesUniversity of Agricultural SciencesMangalore-575002INDIATel: 91 824 439256 (College); 434356 (Dept), 439412 (Res)Fax: 91 824 438366E-mail: [email protected]. Mohammed ShariffFaculty of Veterinary MedicineUniversiti Putra Malaysia43400 Serdang, SelangorMALAYSIATel: 603-9431064; 9488246Fax: 603-9488246; 9430626E-mail: [email protected]. Jie HuangYellow Sea Fisheries Research InstituteChinese Academy of Fishery Sciences106 Nanjing RoadQingdao, Shandong 266071PEOPLE’S REPUBLIC of CHINATel: 86 (532) 582 3062Fax: 86 (532) 581 1514E-mail: [email protected]. Jian-Guo HeSchool of Life SciencesZhongshan UniversityGuangzhou 510275PEOPLE’S REPUBLIC of CHINATel: +86-20-84110976Fax: +86-20-84036215E-mail: [email protected]. Juan D. AlbaladejoFish Health SectionBureau of Fisheries and Aquatic ResourcesArcadia Building, 860 Quezon AvenueQuezon City, Metro ManilaPHILIPPINES
Disease
Shrimp diseases
217
Tel/Fax: 632-372-5055E-mail: [email protected]. Joselito R. SomgaFish Health SectionBureau of Fisheries and Aquatic ResourcesArcadia Building, 860 Quezon AvenueQuezon City, Metro ManilaPHILIPPINESTel/Fax: 632-372-5055E-mail: [email protected]. Leobert de la PenaFish Health SectionAquaculture DepartmentSoutheast Asian Fisheries Development CenterTigbauan, Iloilo 5021PHILIPPINESTel: 63 33 335 1009Fax: 63 33 335 1008E-mail: [email protected]; [email protected]. P.P.G.S.N. SiriwardenaHead, Inland Aquatic Resources and AquacultureNational Aquatic Resources Research and Develoment AgencyColombo 15,SRI LANKATel: 941-522005Fax: 941-522932E-mail: [email protected]. Yen-Ling SongDepartment of ZoologyCollege of ScienceNational Taiwan University1, Sec. 4, Roosevelt Rd.TAIWAN PROVINCE OF CHINAE-mail: [email protected]. Pornlerd ChanratchakoolAquatic Animal Health Research InstituteDepartment of FisheriesKasetsart University CampusJatujak, Ladyao, Bangkok 10900THAILANDTel: 662-5794122Fax: 662-5613993E-mail: [email protected]. Daniel F. FeganNational Center for Genetic Engineering and Biotechnology (BIOTEC)Shrimp Biotechnology Programme18th Fl. Gypsum BuidlingSri Ayuthya Road, BangkokTHAILANDTel: 662-261-7225Fax:662-261-7225E-mail: [email protected]. Chalor LimsuanChalor LimsuwanFaculty of Fisheries, Kasetsart UnviersityJatujak, Bangkok 10900THAILANDTel: 66-2-940-5695
Annex C.AII. List of Regional Resource Expertsfor Crustacean Diseases in Asia-Pacific
218
Dr. Gary NashCenter for Excellence for Shrimp Molecular Biology and BiotechnologyChalerm Prakiat BuildingFaculty of Science, Mahidol UniversityRama 6 RoadBangkok 10400THAILANDTel: 66-2-201-5870 to 5872Fax: 66-2-201-5873E-mail: [email protected] Nguyen Thanh PhuongAquaculture and Fisheries Sciences Institute (AFSI)College of AgricultureCantho University, CanthoVIETNAMTel.: 84-71-830-931/830246Fax: 84-71-830-247.E-mail: [email protected] Peter WalkerAssociate Professor and Principal Research ScientistCSIRO Livestock IndustriesPMB 3 Indooroopilly Q 4068AUSTRALIATel: 61 7 3214 3758Fax: 61 7 3214 2718E-mail : [email protected] P.K.M. WijegoonawardenaNational Aquatic Resources Research and Development AgencyColombo 15,SRI LANKATel: 941-522005Fax: 941-522932E-mail: [email protected]. Tim FlegelCentex Shrimp, Chalerm Prakiat BuildingFaculty of Science, Mahidol UniversityRama 6 Road, Bangkok 10400THAILANDPersonal Tel: (66-2) 201-5876Office Tel: (66-2) 201-5870 or 201-5871 or 201-5872Fax: (66-2) 201-5873Mobile Phone: (66-1) 403-5833E-mail: [email protected]. Celia Lavilla-TorresFish Health SectionAquaculture DepartmentSoutheast Asian Fisheries Development CenterTigbauan, Iloilo 5021PHILIPPINESTel: 63 33 335 1009Fax: 63 33 335 1008E-mail: [email protected]
Bacterial Diseases
Annex C.AII. List of Regional Resource Experts
for Crustacean Diseases in Asia-Pacific
Shrimp Viruses
219
Asian Fish Health Bibliography III Japan by Wakabayashi H (editor). Fish Health SpecialPublication No. 3. Japanese Society of Fish Pathology, Japan and Fish Health Section of AsianFisheries Society, Manila, PhilippinesInformation: Japanese Society of Fish Pathology
Manual for Fish Diseases Diagnosis: Marine Fish and Crustacean Diseases in Indonesia(1998) by Zafran, Des Roza, Isti Koesharyani, Fris Johnny and Kei YuasaInformation: Gondol Research Station for Coastal Fisheries
Health Management in Shrimp Ponds. Third Edition (1998) by P. Chanratchakool, J.F.Turnbull,S.J.Funge-Smith,I.H. MacRae and C. Limsuan.Information: Aquatic Animal Health Research Institute
Department of FisheriesKasetsart University CampusJatujak, Ladyao, Bangkok 10900THAILANDTel: (66.2) 579.41.22Fax: (66.2) 561.39.93E-mail: [email protected]
Fish Health for Fishfarmers (1999) by Tina ThorneInformation: Fisheries Western Australia
3rd Floor, SGIO Atrium186 St. Georges Terrace, Perth WA 6000Tel: (08) 9482 7333 Fax: (08) 9482 7389Web: http://www.gov.au.westfish
Australian Aquatic Animal Disease – Identification Field Guide (1999) by Alistair Herfort andGrant RawlinInformation: AFFA Shopfront – Agriculture, Fisheries and Forestry – Australia
Diseases in Penaeid Shrimps in the Philippines. Second Edition (2000). By CR Lavilla-Pitogo,G.D. Lio-Po, E.R. Cruz-Lacierda, E.V. Alapide-Tendencia and L.D. de la PenaInformation: Fish Health Section
Manual for Fish Disease Diagnosis - II: Marine Fish and Crustacean Diseases in Indonesia(2001) by Isti Koesharyani, Des Roza, Ketut Mahardika, Fris Johnny, Zafran and Kei Yuasa,edited by K. Sugama, K. Hatai, and T NakaiInformation: Gondol Research Station for Coastal Fisheries
ANNEX C.AIII. LIST OF USEFUL DIAGNOSTICMANUALS/GUIDES TO CRUSTACEAN
DISEASES IN ASIA-PACIFIC
220
Annex C.AIII.List of Useful Diagnostic Manuals/Guides to Crustacean Diseases in Asia-Pacific
Reference PCR Protocol for Detection of White Spot Syndrome Virus (WSSV) in Shrimp.Shrimp Biotechnology Service Laboratory. Vol. 1, No. 1, March 2001Information: Shrimp Biotechnology Service Laboratory