International Journal of Innovative Research in Engineering & Management (IJIREM) ISSN: 2350-0557, Volume-2, Issue-6, November- 2015 40 Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and Characterization of Xylanases Dr. Pascoal José Gaspar Júnior Departamento de Bioquímica, Universidade de Campinas, e Departamento de Biologia, Centro Universitário de Formiga, Formiga, MG, Brazil; Dra. Fernanda Silva Torres Dr. Daniel Moreira dos Santos Departamento de Bioquímica e Imunologia, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil; Ana Paula Resende Pinto Departamento de Química, Universidade Federal de Ouro Preto, Ouro Preto, MG, Brazil; MsC. Camila de Melo Silva Departamento de Química, Centro Universitário de Formiga, Formiga, MG, Brazil; Nilo Sobreira Silva Departamento de Biologia, Centro Universitário de Formiga, Formiga, MG, Brazil; MsC. Lucas Vieira de Faria Departamento Microbiologia Agrícola, Universidade Federal de Lavras, Lavras, MG, Brazil; Juraci Lourenço Teixeira Departamento de Química, Instituto Federal do Triângulo Mineiro, Ituiutaba, MG, Brazil; Dr. Félix Gonçalves de Siqueira Laboratório de Bioquímica de Processos - Embrapa Agro-Energia, Brasília, DF, Brazil; MsC. Géssyca Paula de Alvarenga Soares Departamento Microbiologia Agrícola, Universidade Federal de Lavras, Lavras, MG, Brazil; Dra. Maria Elena de Lima Departamento de Bioquímica e Imunologia, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil; Dr. Sérgio Marangoni Departamento de Bioquímica, Universidade de Campinas, Campinas, SP, Brazil; ABSTRACT Screening strategies for the production of fungal holocellulases aims at obtaining enzymes that hydrolyze the lignocellulosic material from plant biomass, increasing the viability of cellulosic ethanol production. The present study evaluates the potential of Penicillium simplicissimum and Penicillium corylophilum to produce holocellulases from commercial and lignocellulosic substrates. These fungi were grown in a supplementary medium with 1.0% lignocellulosic substrate, at pH 7.0, at 10 8 spores.mL -1 . The supernatant was filtered and the enzyme activities were evaluated. P. simplicissimum xylanase activity on linseed substrate, after 72 and 120 h (3.8 and 3.9 IU.mL -1 , respectively), showed the highest values among all the enzymes tested. The lyophilized crude extract, after 72 h of incubation, was submitted to two chromatographic steps (gel filtration and anion exchange). A third chromatographic step using a reversed phase column, followed by mass spectrometry analyses, showed a pool of low- molecular-weight xylanases (18831 Da). The specific xylanase activity increased considerably after anion- exchange chromatography purification, from 3x10 -3 atv.mg -1 to 19.2 atv.mg -1 . Maximum enzyme activity was observed with 0.5% xylan and 0.04% xylose as carbon sources. Optimum pH and temperature for xylanase activity were 4.0 and 50°C, respectively. In conclusion, the fungal strains Penicillium simplicissimum and Penicillium corylophilum are potential sources for the production of different enzymes in several lignocellulosic substrates derived from oil plants. Moreover, we purified and partially characterized a pool of xylanase enzymes from P. simplicissimum that show promising industrial application. Keywords Agro-industrial residues; Hemicellulose; Enzyme purification; Xylanase; Hydrolysis, Penicillium simplicissimum. 1. INTRODUCTION Lignocellulosic residues have been widely explored as a strategy to solve the energy problem worldwide. These residues consist primarily of cellulose, hemicellulose and lignin, and they enable the growth of a variety of microorganisms that are known for producing a set of enzymes capable of degrading these insoluble polymers to soluble sugars, such as cellobiose and glucose (Monte et al. 2010). One type of hemicellulose is xylan, the second most abundant natural polysaccharide (Collins et al. 2005). Xylan is hydrolyzed by xylanases, which cleave their glycosidic bonds in the xylan backbone, reducing its polymerization (Reilly, 1981; Puls and Poutanen 1989; Li et al. 2000). There are several industrial applications for xylanases (Subramaniyan and Prema 2002). The major applications of these enzymes are in pulp, paper, food and baking industries (Beg et al. 2001). In the food industry, the action of xylanases, glucanases, proteinases and amylases reduces viscosity and increases the adsorption of nutrients (Bhat and Hazlewood 2001). Xylanases have also been used in combination with pectinases and other enzymes for processes such as clarification of juices, extraction of coffee, and extraction of plant oils and starch. Other applications include the production of bioethanol fuel from agricultural waste (Damaso et al. 2003).
Transcript
International Journal of Innovative Research in Engineering & Management (IJIREM) ISSN: 2350-0557, Volume-2, Issue-6, November- 2015
40
Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and
Characterization of Xylanases
Dr. Pascoal José Gaspar Júnior
Departamento de Bioquímica, Universidade de Campinas,
e Departamento de Biologia, Centro Universitário de Formiga, Formiga,
MG, Brazil; Dra. Fernanda Silva Torres
Dr. Daniel Moreira dos Santos Departamento de Bioquímica e
Imunologia, Universidade Federal de Minas Gerais,
Belo Horizonte, MG, Brazil; Ana Paula Resende Pinto
Departamento de Química, Universidade Federal de Ouro Preto,
Ouro Preto, MG, Brazil;
MsC. Camila de Melo Silva Departamento de Química, Centro Universitário de Formiga, Formiga,
MG, Brazil; Nilo Sobreira Silva
Departamento de Biologia, Centro Universitário de Formiga, Formiga,
MG, Brazil; MsC. Lucas Vieira de Faria
Departamento Microbiologia Agrícola, Universidade Federal de
Departamento de Química, Instituto Federal do Triângulo Mineiro,
Ituiutaba, MG, Brazil;
Dr. Félix Gonçalves de Siqueira
Laboratório de Bioquímica de Processos - Embrapa Agro-Energia,
Brasília, DF, Brazil; MsC. Géssyca Paula de
Alvarenga Soares Departamento Microbiologia Agrícola,
Universidade Federal de Lavras, Lavras, MG, Brazil;
Dra. Maria Elena de Lima Departamento de Bioquímica e
Imunologia, Universidade Federal de Minas Gerais, Belo Horizonte, MG,
Brazil; Dr. Sérgio Marangoni
Departamento de Bioquímica, Universidade de Campinas,
Campinas, SP, Brazil;
ABSTRACT Screening strategies for the production of fungal holocellulases aims at obtaining enzymes that hydrolyze the lignocellulosic material from plant biomass, increasing the viability of cellulosic ethanol production. The present study evaluates the potential of Penicillium simplicissimum and Penicillium corylophilum to produce holocellulases from commercial and lignocellulosic substrates. These fungi were grown in a supplementary medium with 1.0% lignocellulosic substrate, at pH 7.0, at 108 spores.mL-1. The supernatant was filtered and the enzyme activities were evaluated. P. simplicissimum xylanase activity on linseed substrate, after 72 and 120 h (3.8 and 3.9 IU.mL-1, respectively), showed the highest values among all the enzymes tested. The lyophilized crude extract, after 72 h of incubation, was submitted to two chromatographic steps (gel filtration and anion exchange). A third chromatographic step using a reversed phase column, followed by mass spectrometry analyses, showed a pool of low-molecular-weight xylanases (18831 Da). The specific xylanase activity increased considerably after anion-exchange chromatography purification, from 3x10-3 atv.mg-1 to 19.2 atv.mg-1. Maximum enzyme activity was observed with 0.5% xylan and 0.04% xylose as carbon sources. Optimum pH and temperature for xylanase activity were 4.0 and 50°C, respectively. In conclusion, the fungal strains Penicillium simplicissimum and Penicillium corylophilum are potential sources for the production of different enzymes in several lignocellulosic substrates derived from oil plants. Moreover, we purified and partially characterized a pool of xylanase enzymes from P. simplicissimum that show promising industrial application.
1. INTRODUCTION Lignocellulosic residues have been widely explored as a
strategy to solve the energy problem worldwide. These residues consist primarily of cellulose, hemicellulose and lignin, and they enable the growth of a variety of microorganisms that are known for producing a set of enzymes capable of degrading these insoluble polymers to soluble sugars, such as cellobiose and glucose (Monte et al. 2010). One type of hemicellulose is xylan, the second most abundant natural polysaccharide (Collins et al. 2005). Xylan is hydrolyzed by xylanases, which cleave their glycosidic bonds in the xylan backbone, reducing its polymerization (Reilly, 1981; Puls and Poutanen 1989; Li et al. 2000).
There are several industrial applications for xylanases (Subramaniyan and Prema 2002). The major applications of these enzymes are in pulp, paper, food and baking industries (Beg et al. 2001). In the food industry, the action of xylanases, glucanases, proteinases and amylases reduces viscosity and increases the adsorption of nutrients (Bhat and Hazlewood 2001). Xylanases have also been used in combination with pectinases and other enzymes for processes such as clarification of juices, extraction of coffee, and extraction of plant oils and starch. Other applications include the production of bioethanol fuel from agricultural waste (Damaso et al. 2003).
Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and Characterization of Xylanases
41
Bioethanol is considered a sustainable transportation fuel. The main motivation for investments in research and process development concerning bioethanol production is environmental concern, related to global warming. Based on this knowledge, this study focuses on the process of cellulose enzymatic hydrolysis. In general, the production of bioethanol from lignocellulose involves degradation of the polymeric compounds, primarily cellulose and hemicellulose, to sugars, which are then fermented by microorganisms to ethanol. This process can be performed with various procedures. We evaluated the enzyme production potential of Penicillium simplicissimum and Penicillium corylophilum grown in lignocellulosic substrates from oil plants used in biodiesel production. In addition, we purified and biochemically characterized some xylanases produced by Penicillium simplicissimum grown in linseed.
2. MATERIAL AND METHODS
2.1 Organisms and growth conditions P. simplicissimum was isolated in the Laboratory of
Applied Microbiology from Universidade de Formiga (UNIFOR), Formiga, Minas Gerais state, Brazil. P. corylophilum was isolated in the Laboratory of Microbiology from Universidade Estadual de São Paulo (UNESP), Ribeirão Preto, São Paulo state, Brazil. The fungi were maintained and replicated according to Siqueira et al. (2010). The lignocellulosic residues (1%) lupin, wild radish, crambe, sunflower, niger, linseed, castor, soybean, safflower, cottonseed, jatropha, were standardized by physical pretreatment (850 mm/micrometer) and incubated at 28°C with agitation (120 rpm). One percent microcrystalline cellulose (MC) was used as control. The crude extracts were filtered using common filter paper after 12, 24, 72 and 120 h of incubation. The filtrates were used in enzyme assays.
2.2 Physicochemical analysis of lignocellulosic residues
The content of neutral and acid detergent fiber (NDF and ADF, respectively), lignin, cellulose and hemicellulose was evaluated according to Van Soest (1967). Measurements of total sugar, pectin and soluble pectin followed the protocols of Antrona, method of Dische (1962) and Mccready and Mccomb (1952), and the solubility was calculated as the ratio of soluble pectin and total pectin X 100.
2.3 Enzymatic assays The release of reducing sugar was measured using the
3,5-dinitrosalicylic acid (DNS) method (Miller, 1959). For this, 2 mg/mL of each reducing sugar were added to 150 μL of distilled water and 300 μL of DNS and these were incubated at 100 °C for 10 min. Absorbance was evaluated at 540nm. All activities were expressed as IU.mL−1.
Total cellulase activity (FPase) was determined using Filter paper assay (Whatman No. 1 - 1 x 6 cm-50 mg) incubated with 150 μL of different lignocellulosic residues culture filtrate at 50 °C for 1 h, pH 7.0. Exoglucanase activity was determined using 100 μL of 1% MC in 0.05 M sodium acetate, pH 5.0
incubated for 2 h with 50 μL culture filtrate, at 50°C, 120 rpm. Endoglucanase (CMCase), xylanase, pectinase and mannanase activities were evaluated using specific substrates (0.5% carboximetilcellulose, CMC), 1% beechwood xylan, citrus peel pectin and locust bean gum galactomannan). In these assays, 50 μL of culture filtrate were incubated with 100 μL of specific substrate at 50ºC, pH 7.0 for 30 min. The release of reducing sugar in these assays was measured using the DNS method. β-glucosidase activity was determined using ρ-nitrophenyl β-D-glucopyranoside (pNPG) susbtrate according to Iembo et al. (2006), with modifications. The reaction was stopped by the addition of 1.0 M Na2CO3 (1mL) and absorbance was evaluated at 410 nm. The calibration curve was prepared using 10 mol/mL pNPG in 900 L of distilled water.
The effect of temperature on xylanase activity and enzyme-substrate complexation was determined in the temperature range of 30 to 80 °C in 50 mmol.L−1 sodium acetate buffer, pH 4.0. To determine the effect of pH on this activity, at 50 °C, a pH range of 3.0 to 9.0 was used. The following buffers were used in these experiments (mmol. L−1): 50 sodium acetate (pH 3.0–6.0), 50 sodium phosphate (pH 7.0), and 50 Tris-HCl (pH 8.0 and 9.0). Appropriate controls were included for all experiments.
2.4 Effect of carbon source on xylanase production
The effect of carbon sources on xylanase production was investigated by culturing the isolate in basic medium (pH 7.0) at room temperature (28 ±2 °C) for 168 h. Lactose, glucose, xylan, starch, MC, sucrose, cellobiose, pectin or xylose were used as a carbon source (0.5 and 2.0 %), individually, in supplementary medium. The crude extracts were filtered using common filter paper and the culture filtrate was used for enzyme assay.
2.5 Purification and partial characterization of xylanases
Lyophilized P. simplicissimum crude extract (300 mg) containing xylanases dissolved in 0.05 M sodium acetate, pH 7.0, was centrifuged at 4°C, 10,000 g for 10 min (Mickro 200R Hettich rotor 2424) and the supernatant was submitted to gel filtration chromatography (GFC) on HiPrep 16/60 Sephacryl S-200 HR column. This column was equilibrated and eluted with the same buffer used to dissolve the crude extract at a flow rate of 0.3 mL.min-1. Fractions of 1 mL were collected. The fractions containing xylanase proteins (0.293 mg) were pooled and further applied into the anion exchange chromatography (AEC) TSKgel DEAE-2SW column, previously equilibrated with 0.05 M sodium phosphate buffer, pH 7.0. The enzyme sample was eluted with a gradient of 0.05 M sodium phosphate buffer (B), containing 1 M NaCl, pH 7.0, according to the following program: 0% of B for 0.95 mL, 0 45% of B for 11.4 mL; 45 to 100% of B for 0.95 mL and 100% for 1.9 mL. Fractions of 0.150 mL were collected at a flow rate of 0.25 mL.min-1.
2.6 Reverse-phase chromatography (RPC) The fraction from GFC (544 mg) containing xylanase
proteins was submitted to RPC on Discovery® BIO Wide Pore
International Journal of Innovative Research in Engineering & Management (IJIREM) ISSN: 2350-0557, Volume-2, Issue-6, November- 2015
42
C18 column. This column was equilibrated with 0.1% TFA in MilliQ water (A) and the xylanases were eluted with a linear gradient of 0.1% TFA in acetonitrile. Fractions of 1 mL were collected at a flow rate of 4.0 mL.min-1. The xylanases were detected by mass spectrometry analysis.
2.7 Determination of Protein Concentration Protein concentration was determined according to
Bradford (1976) using bovine serum albumin as reference.
2.8 Mass spectrometry (MS) MS analyzes were carried out using a MALDI-TOF MS
AutoFlex III instrument in positive/linear mode controlled by FlexControl 3.3 software. Instrument calibration was achieved by using Calibration Standard Protein I and Dihydroxybenzoic acid and 3,5-dimethoxy-4-hydroxycinnamic acid were used as matrices. 0.5 microliter samples, were spotted to MTPAnchorChip-400/384 targets using standard protocols for the dried droplet method. MS data analysis was performed using the FlexAnalysis 3.3 software.
2.9 Graphics and Statistical analyses Results were expressed as mean ± SEM of three
experiments. Graphics were constructed in GraphPad Prisma 5 and statistical analyses were performed using two-way ANOVA followed by Bonferroni post hoc test for multiple comparisons or Dunnets post test. A value of p<0.05 was considered statistically significant.
3. RESULTS 3.1 Physicochemical analysis
The physicochemical analysis of the carbon sources is shown in table 1. Among all lignocellulosic substrates shown, the highest percentages of acid detergent fiber (ADF) and neutral detergent fiber (NDF) were found with castor beans (47.4 and 50.5%) and linseed (31.3 and 77.6%) substrates, respectively. These substrates also presented the highest values of holocellulose (hemicellulose + cellulose), 45% for castor beans and 63% for linseed, followed by cotton (37%). Lignin was found in high percentages in crambe and linseed substrates. In contrast, soybean had the lowest values of lignin. The ratios of total sugars, total and soluble pectin were obtained for all substrates, but the highest values were found for wild radish (21.7, 1,243.5 and 1,144.8 mg /100 g, respectively). Wild radish, linseed and cotton presented the highest solubility, 92.0, 51.7 and 45.1%, respectively.
Table 1 Determination of some physicochemical parameters of lignocellulosic residues, by using different carbon sources.
3.2 Enzymatic production of P. corylophilum and P. simplicissimum in different lignocellulosic substrates
Several enzymatic activities of P. corylophilum and P. simplicissimum were determined after 12, 24, 72 and 120 h of incubation using as substrates: lupin, wild radish, crambe, sunflower, nyger, linseed, castor, soybean, safflower, cottonseed and jatropha. Endoglucanase activity for P. corylophilum was significantly higher in lupin, wild radish, nyger, linseed and jatropha substrates compared with microcrystalline cellulose control (0.075 IU.mL-1). The highest activities at 72 h were found in sunflower and wild radish, 0.185 and 0.184 IU.mL-1, respectively, and at 120 h, in nyger and jatropha substrates (0.187 and 0.203 IU.mL-1). The castor bean substrate presented the smallest activities at all times (data not shown). Results obtained for P. simplicissimum showed higher activities than control for all substrates. The linseed substrate, at 72 and 120 h, showed the highest activities, 0.137 and 0.177 IU.mL-1, respectively
The exoglucanase activities were smaller when compared with endoglucanase. The highest values were obtained for P. corylophilum at 24 h cultivated in soybean and safflower (0.034 IU.mL-1 for both); at 72 h and 120 h in lupine (0.033 and 0.034 IU.mL-1) and at 120 h in linseed residues (0.031 IU.mL-1), compared with 0.019 IU.mL-1 for the control. For P. simplicissimum the activities were significantly higher than the control (0.006 IU/mL) at 12 h in soybean and cottonseed (0.043 and 0.044 IU.mL-1), at 24 h in crambe and soybean (0.042 and 0.048 IU.mL-1), at 72 and 120 h in crambe (0.043 and 0.046 IU.mL-1) and in cottonseed substrates, 0.043 IU.mL-1, respectively.
We determined total cellulase activities (FPases), and the crambe substrate, at 72 and 120 h, showed the highest activity. For P. corylophilum, at 120 h, the nyger and jatropha substrates showed the best results, and for P. simplicissimum, at 24, 72 and 120 h, linseed and castor bean substrates showed the most significant results. Other activities investigated were β-glucosidase and pectinase. The first was more active for P. corylophilum at 72 and 120 h with all substrates, with the highest values achieved with soybean (0.47 and 0.92 IU.mL-1) and safflower (0.61 and 1.0 IU.mL-1). For P. simplicissimum, soybean, safflower and cotton residues showed the biggest values. The pectinase activity for the two fungi were higher than the control in all times and substrates used (data not shown).
Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and Characterization of Xylanases
43
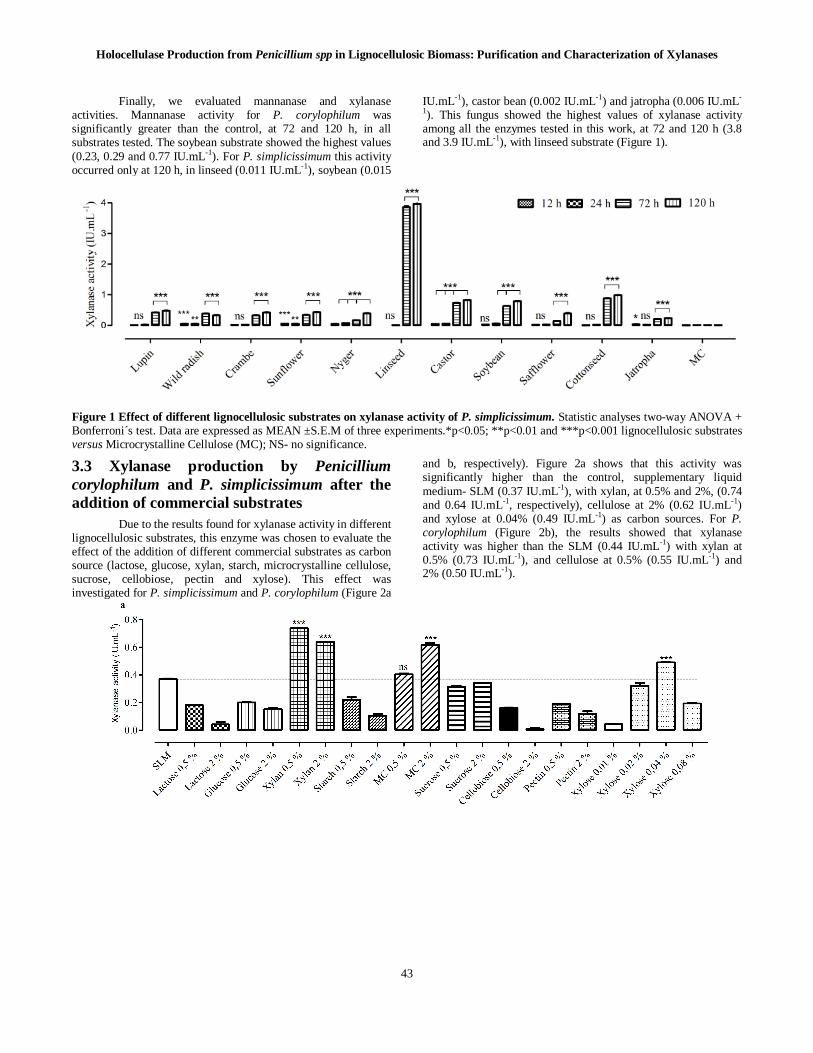
Finally, we evaluated mannanase and xylanase activities. Mannanase activity for P. corylophilum was significantly greater than the control, at 72 and 120 h, in all substrates tested. The soybean substrate showed the highest values (0.23, 0.29 and 0.77 IU.mL-1). For P. simplicissimum this activity occurred only at 120 h, in linseed (0.011 IU.mL-1), soybean (0.015
IU.mL-1), castor bean (0.002 IU.mL-1) and jatropha (0.006 IU.mL-
1). This fungus showed the highest values of xylanase activity among all the enzymes tested in this work, at 72 and 120 h (3.8 and 3.9 IU.mL-1), with linseed substrate (Figure 1).
Figure 1 Effect of different lignocellulosic substrates on xylanase activity of P. simplicissimum. Statistic analyses two-way ANOVA + Bonferroni´s test. Data are expressed as MEAN ±S.E.M of three experiments.*p<0.05; **p<0.01 and ***p<0.001 lignocellulosic substrates versus Microcrystalline Cellulose (MC); NS- no significance.
3.3 Xylanase production by Penicillium corylophilum and P. simplicissimum after the addition of commercial substrates
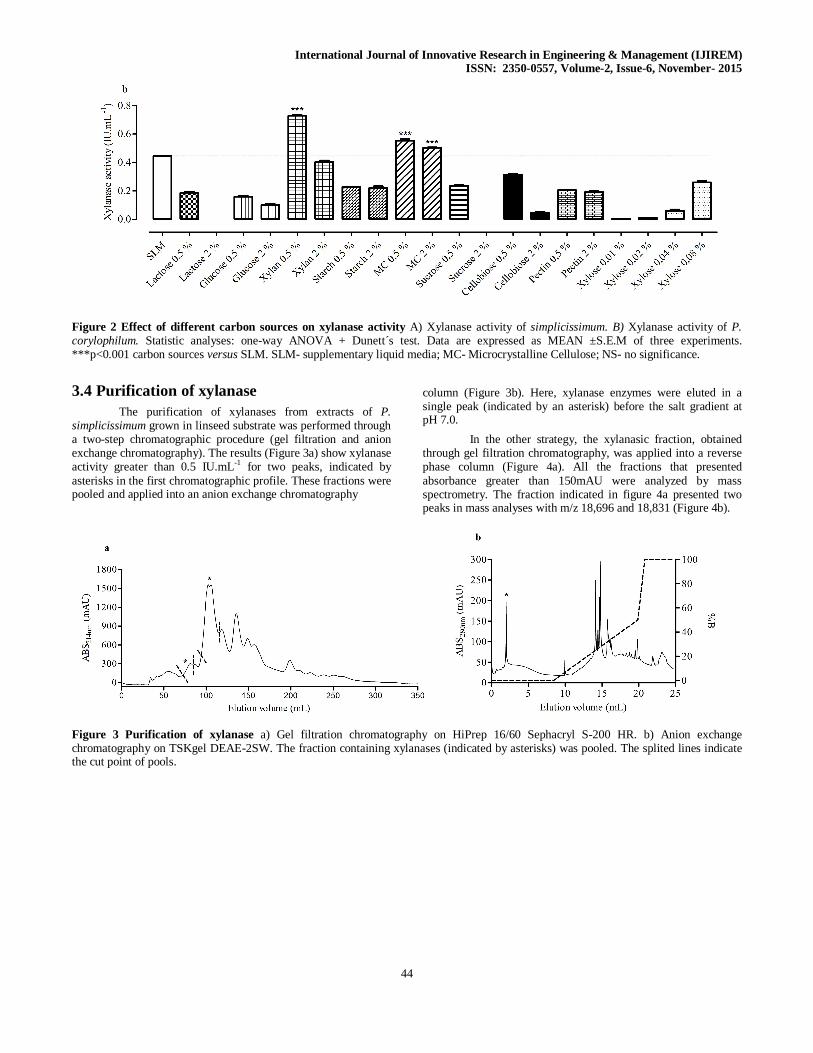
Due to the results found for xylanase activity in different lignocellulosic substrates, this enzyme was chosen to evaluate the effect of the addition of different commercial substrates as carbon source (lactose, glucose, xylan, starch, microcrystalline cellulose, sucrose, cellobiose, pectin and xylose). This effect was investigated for P. simplicissimum and P. corylophilum (Figure 2a
and b, respectively). Figure 2a shows that this activity was significantly higher than the control, supplementary liquid medium- SLM (0.37 IU.mL-1), with xylan, at 0.5% and 2%, (0.74 and 0.64 IU.mL-1, respectively), cellulose at 2% (0.62 IU.mL-1) and xylose at 0.04% (0.49 IU.mL-1) as carbon sources. For P. corylophilum (Figure 2b), the results showed that xylanase activity was higher than the SLM (0.44 IU.mL-1) with xylan at 0.5% (0.73 IU.mL-1), and cellulose at 0.5% (0.55 IU.mL-1) and 2% (0.50 IU.mL-1).
International Journal of Innovative Research in Engineering & Management (IJIREM) ISSN: 2350-0557, Volume-2, Issue-6, November- 2015
44
Figure 2 Effect of different carbon sources on xylanase activity A) Xylanase activity of simplicissimum. B) Xylanase activity of P. corylophilum. Statistic analyses: one-way ANOVA + Dunett´s test. Data are expressed as MEAN ±S.E.M of three experiments. ***p<0.001 carbon sources versus SLM. SLM- supplementary liquid media; MC- Microcrystalline Cellulose; NS- no significance.
3.4 Purification of xylanase The purification of xylanases from extracts of P.
simplicissimum grown in linseed substrate was performed through a two-step chromatographic procedure (gel filtration and anion exchange chromatography). The results (Figure 3a) show xylanase activity greater than 0.5 IU.mL-1 for two peaks, indicated by asterisks in the first chromatographic profile. These fractions were pooled and applied into an anion exchange chromatography
column (Figure 3b). Here, xylanase enzymes were eluted in a single peak (indicated by an asterisk) before the salt gradient at pH 7.0.
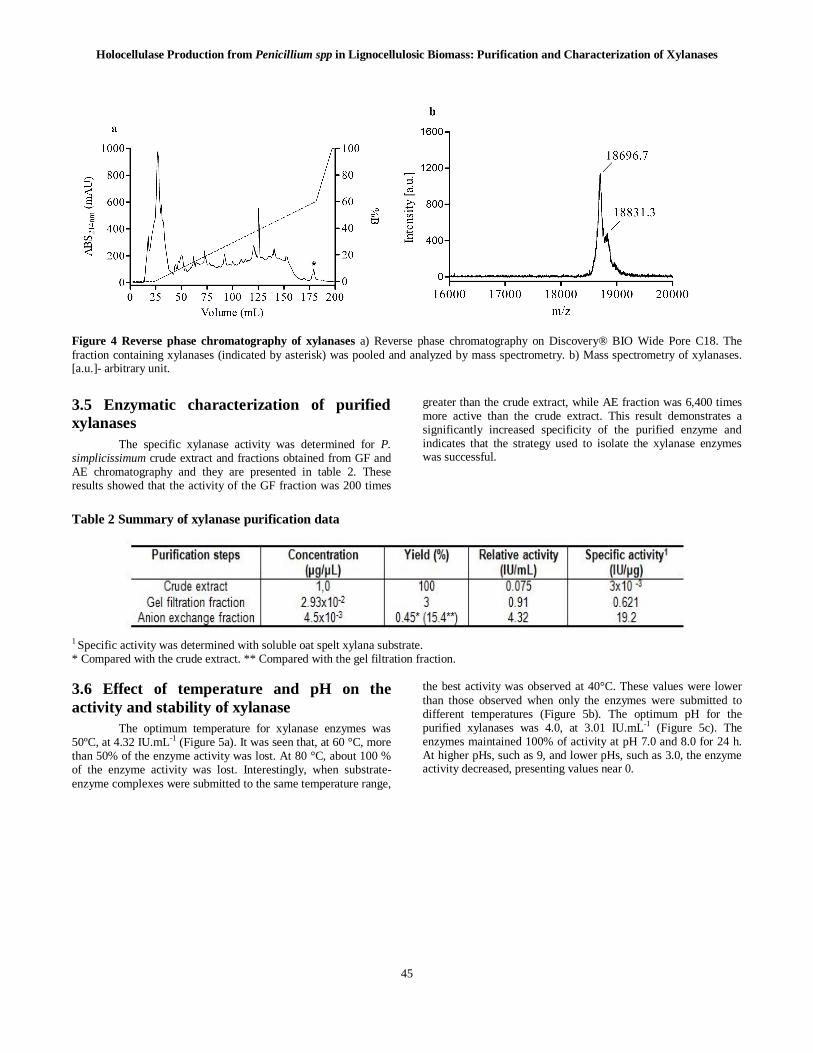
In the other strategy, the xylanasic fraction, obtained through gel filtration chromatography, was applied into a reverse phase column (Figure 4a). All the fractions that presented absorbance greater than 150mAU were analyzed by mass spectrometry. The fraction indicated in figure 4a presented two peaks in mass analyses with m/z 18,696 and 18,831 (Figure 4b).
Figure 3 Purification of xylanase a) Gel filtration chromatography on HiPrep 16/60 Sephacryl S-200 HR. b) Anion exchange chromatography on TSKgel DEAE-2SW. The fraction containing xylanases (indicated by asterisks) was pooled. The splited lines indicate the cut point of pools.
Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and Characterization of Xylanases
45
Figure 4 Reverse phase chromatography of xylanases a) Reverse phase chromatography on Discovery® BIO Wide Pore C18. The fraction containing xylanases (indicated by asterisk) was pooled and analyzed by mass spectrometry. b) Mass spectrometry of xylanases. [a.u.]- arbitrary unit.
3.5 Enzymatic characterization of purified xylanases
The specific xylanase activity was determined for P. simplicissimum crude extract and fractions obtained from GF and AE chromatography and they are presented in table 2. These results showed that the activity of the GF fraction was 200 times
greater than the crude extract, while AE fraction was 6,400 times more active than the crude extract. This result demonstrates a significantly increased specificity of the purified enzyme and indicates that the strategy used to isolate the xylanase enzymes was successful.
Table 2 Summary of xylanase purification data
1 Specific activity was determined with soluble oat spelt xylana substrate. * Compared with the crude extract. ** Compared with the gel filtration fraction.
3.6 Effect of temperature and pH on the activity and stability of xylanase
The optimum temperature for xylanase enzymes was 50ºC, at 4.32 IU.mL-1 (Figure 5a). It was seen that, at 60 °C, more than 50% of the enzyme activity was lost. At 80 °C, about 100 % of the enzyme activity was lost. Interestingly, when substrate-enzyme complexes were submitted to the same temperature range,
the best activity was observed at 40°C. These values were lower than those observed when only the enzymes were submitted to different temperatures (Figure 5b). The optimum pH for the purified xylanases was 4.0, at 3.01 IU.mL-1 (Figure 5c). The enzymes maintained 100% of activity at pH 7.0 and 8.0 for 24 h. At higher pHs, such as 9, and lower pHs, such as 3.0, the enzyme activity decreased, presenting values near 0.
International Journal of Innovative Research in Engineering & Management (IJIREM) ISSN: 2350-0557, Volume-2, Issue-6, November- 2015
46
Figure 5 Effect of different temperatures and pH on xylanase activity A) Effect of different temperatures on xylanase activity. This enzyme was incubated at 30, 40, 50, 60, 70 and 80°C and the enzymatic activity was determined as described in Methods B) Effect of different temperatures on xylanase-xylan complex. This complex was incubated at 30, 40, 50, 60, 70 and 80°C and the xylanase activity was determined as described in Methods. C) Effect of different pH on xylanase activity. This enzyme was incubated at pH 3, 4, 5, 6, 7, 8 and 9 and the enzymatic activity was determined.
4. DISCUSSION Bioethanol is predominantly produced from biomass,
and can be categorized into three generations (1st, 2nd and 3rd), according to the origin of the biomasses used (Dragone et al. 2010). Lignocellulosic substrates are the non-edible part of the plants and (Nigam and Singh 2010) can potentially provide novel “second generation” bioethanol, due to the fact that they do not compete with food agriculture (Ohman et al. 2006; Kleinert and Barth 2008).
The composition of lignocellulosic biomasses (mainly hemicellulose, cellulose and lignin) varies quantitatively and qualitatively according to the plant type, varieties, part and maturity (Aman 1993; Mosier et al. 2005; Vanholme et al. 2010). Some compositional and structural features, such as the presence of lignin, the crystalline structure of cellulose and its accessible surface area, can limit their enzymatic degradation (Tong et al. 1990; Chang and Holtzapple 2000; Monlau et al. 2012). In this context, to achieve adequate enzymatic hydrolysis for bioethanol production from some lignocellulosic substrates that are more difficult to degrade, a pretreatment step is necessary (Chang and Holtzapple 2000; Taherzadeh and Karimi 2008). Due to this pretreatment, the production of 2nd generation fuels is not cost effective, because there are many technical barriers to be overcome before their potential can be achieved (IEA 2008).
In this work, the content of lignocellulosic residues from different carbon sources was determined (Table 1). Hemicellulose, which connects lignin and cellulose fibers and confers rigidity to the whole matrix (Salmen and Olsson 1998; Watanabe et al. 2003), was more abundant in linseed (46.3%) and jatropha (22.0%). The other component studied, lignin, whose distribution and composition are very important for enzyme accessibility and the digestibility of biomass (Adler 1977; Yuan et al. 2008; Clark et al. 2009; Guo et al. 2010; Ntaikou et al. 2010), was found in high percentages in linseed (14.8%) and crambe (10.8%) substrates. In contrast, the smallest quantities were observed in soybean (0.075%). In our study, even with high concentrations of lignin in linseed, the xylanase activity was the highest among all substrates analyzed after 72 h of cultivation.
The highest contents of pectin were found for wild radish, 1,243.5%, and this substrate presented the highest
solubility (92.04%). This content has been shown to limit the enzymatic accessibility to cellulose (Berlin et al. 2007; Frigon et al. 2012; Pakarinen et al. 2012). Indeed, Berlin et al. (2007) and Pakarinen et al. (2012) suggested that the use of pectinase enzymes could increase the hydrolysis of cellulose.
The main enzymatic activity found in this work was xylanase activity. Xylanases (endo-1, 4-β-D-xylan xylanohydrolase, EC 3.2.1.8) are endo-glycosyl hydrolases that catalyze the hydrolysis of β-1,4 glycosidic bonds to xylan, a linear polymer of β-D-xylopyranosyl units (Polizeli et al. 2005). The xylanases derived from microorganisms are used in many biotechnological strategies, such as the clarification of fruit juices and wines, food processing, in combination with cellulases, grains and poultry food and bio-pulping (Biely et al 1985; Siqueira et al. 2010, Murthy and Naidu 2010). However, the main problem faced by various industries while using enzymatic treatments is the availability and cost of these enzymes (Hinnman 1994). The use of low cost substrates for the production of industrial enzymes is one of the ways to greatly reduce production costs.
A great number of substrates can induce xylanase production. In this context, we investigated the xylanase production of P. corylophilum and P. simplicissimum fungi, in various lignocellulosic and commercial substrates. Our results showed that the second fungus present the highest activities after 72 and 120 h of incubation, using linseed as a substrate (Figure 1). Probably, this finding can be explained by the largest percentage of hemicellulose (46.3%) found in this substrate (Table 1). Our result corroborates with the findings of Badhan et al. (2007), who observed that the wheat straw produced the highest xylanase amount and that this lignocellulosic material possessed a high content of hemicellulose (50%).
The xylanase production after addition of commercial substrates (Figure 2) showed that the use of xylan, at 0.5% and 2%, cellulose at 2% and xylose at 0.04% as carbon sources significantly increased this activity when compared to SLM for both fungi. Bakri et al. (2003), similarly, found that lower concentrations of xylan (0.5%) developed the highest activity. However, an increase in the concentration of xylan (2.0%) inhibited the enzymatic activity. The presence of 0.04% of xylose was enough to increase P. simplicissimum xylanase activity.
Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and Characterization of Xylanases
47
Pandya and Gupte (2012) found that the optimum xylose concentration was 0.5% w/w, suggesting that this sugar acts as an inducer rather than a carbon source. Moreover, the xylanase production decreased with lower and higher xylose concentrations, suggesting a repression phenomena. Another study from Bakri et al. (2003) tested the effect of increasing concentrations of xylose on P. canescens xylanase production. The results showed that the xylanase production was higher with 2.0% xylose. However, 4 to 10% xylose reduced xylanase production.
The results described above show that P. simplicissimum fungus is a good producer of xylanases. These enzymes, present on crude extract grown in linseed substrate, were chosen for a partial purification procedure through liquid chromatography (Figure 3). The fractionation by GF chromatography (Figure 3a) showed xylanase activities in two regions of the chromatogram, suggesting enzyme multiplicity. The first protein peak, detected after fractionation on AEC, corresponding to the void volume fractionation range, presented xylanase activity. This result demonstrated the cationic property of the purified enzymes. In the other hand, to determine the mass of xylanases, another purification step on an RPC was performed with the fractions obtained from GF chromatography (Figure 4a). All fractions (absorbance > 150mAU) were analyzed by MS. These analyses indicated that fraction 1 (Figure 4a) presented two peaks with masses of 18,696 and 18,831 m/z, respectively (Figure 4b). In others works, Duarte et al. (2012) isolated a low molecular weight xylanase from A. oryzae culture with a molecular mass of 21.5 kDa, determined through gel electrophoresis and Liao et al. (2014) isolated a xylanse from Penicillium oxalicum GZ-2 with a predicted molecular weight of 21.3 kDa. These results corroborate with the findings in this work, emphasizing the presence of this xylanases in different crude extracts.
We determined the specific xylanase activity in P. simplicissimum lyophilized crude extract and fractions obtained from GF and AE chromatography (table 2). The results showed an increase in the specific activity of this enzyme, indicating that the purification strategy was successful. In contrast to our findings, Duarte et al. (2012) showed that the xylanase activity decreased 18 times after the lyophilization of the crude extract.
The level of production and the activity of xylanase are affected by many environmental factors (substrate, incubation time, temperature and pH) that determine the growth rate of microorganisms (Prakasham et al. 2005). In our work, the effect of temperature and pH on the activity and stability of xylanase was investigated. The optimum temperature for xylanase activity was 50 ºC (Figure 5a) and for substrate-enzyme complexes, 40°C. The optimum pH was 4.0 (Figure 5c) and this activity was reduced in acidic (3.0) and in highly alkaline (8.0 and 9.0) ranges. This may be due to the unfavorable pH, which may denature the enzymes and limit their activity. This influence was studied in others works. Liao et al. 2012 showed that the optimum temperature and pH for xylanase activity, produced from Penicillium oxalicum GZ-2, were 50°C and 4.0, respectively. In another study, Duarte et al. (2012) showed that the most efficient enzyme catalysis was at 50 °C and pH 6.0. Roy et al. (2013) showed that the optimum temperature and pH for the partially purified xylanase from Simplicillium obclavatum MTCC 9604 were 50°C and pH 5.0. These results corroborate with our findings.
The use of abundant and low cost agricultural residues as substrate for xylanase production substantially reduces the production costs of this enzyme (Murthy and Naidu 2010; Duarte et al. 2012). The cost of the substrate is crucial in the xylanase production process, because pure xylan, due to its high cost, is not suitable for large-scale production processes. The results obtained during our study suggest that the use of lignocellulosic biomass from agro-industrial residues increases the fungal production of xylanase. This could represent a good strategy to recycle agro-industrial wastes, considering their potential to produce fuels.
5. ACKNOWLEDGMENTS This study was funded by the Brazilian agencies:
Instituto Nacional de Ciência e Tecnologia em Toxinas/Fundação de Amparo à Pesquisa do Estado de São Paulo (INCTTOX/FAPESP), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo a Pesquiso do Estado de Minas Gerais (FAPEMIG) and Financiadora de Estudos e Projetos/ Ministério da Ciência, Tecnologia e Inovação (FINEP/MCTI) The authors greatly appreciated the assistance of Mrs. Flávia De Marco in reviewing this manuscript.
6. CONFLICT OF INTEREST The authors declare that there are no conflicts of
interest.
7. REFERENCES [1] Adler. E. 1977. Lignin chemistry: Past, Present and Future. Wood Science Technol. 11(Jun. 1976), 169-218. DOI= http://dx.doi.org/10.1007/BF00365615. [2] Aman, P. 1993. Composition and structure of cell wall polysaccharides in forages. In: J. Ralph (ed), Forage Cell Wall Structure and Digestibility, Madison, pp 183-200 (Oct, 1991). [3] Badhan, A.K., Chadha, B.S., Kaur, J., Sonia, K.G., Saini, H.S., Bhat, M.K. 2007. Role of transglycosylation products in the expression of multiple Xylanases in Myceliophthora sp. IMI 387099. Current Microbiol. 54, (Oct, 2006) 405-409. DOI= http://dx.doi.org/0.1007/s00284-006-0204-5 [4] Bakri, Y., Jacques, P. and Thonart, P. 2003. Xylanase production by Penicillium canescens 10-10c in solid-state fermentation. Appl. Biochem. and Biotechnol., 105, (Mar. 2003) 108. DOI= http://dx.doi.org/10.1007/978-1-4612-0057-4_61. [5] Beg, Q.K., Kappor, M., Mahajan, L. 2001. Microbial xylanases and their industrial applications: a review. Appl. Microbiol. and Biotechnol., 56, (Jun.2001) 326-338. DOI= http://dx.doi.org/10.1007/s002530100704. [6] Berlin, A., Maximenko, V., Gilkes, N., Saddler, J. 2007. Optimization of enzyme complexes for lignocellulose hydrolysis. Biotechnol. Bioeneng., 97, (Oct.2006) 287-296. DOI= http://dx.doi.org/10.1002/bit.21238. [7] Bhat, M.K. and Hazlewood, G.P. 2001. Enzymology and Other Characteristics of Cellulases and Xylanases. In: Enzymes in Farm Animal Nutrition. Bedford MR and Partridge CC (eds.), ISBN 0 85199 393 1. CAB International, Wallingford, UK. [8] Biely, P., Markovic, O. and Mislovicova, D. 1985. Sensitive detection of endo-1,4-betaglucanases and endo-1,4-beta-xylanases
International Journal of Innovative Research in Engineering & Management (IJIREM) ISSN: 2350-0557, Volume-2, Issue-6, November- 2015
48
in gels. Anal. Biochem., 144, (Jul. 1984) 147-151. DOI= http://dx.doi.org/10.1016/0003-2697(85)90096-X. [9] Bradford, M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, (Jan.1976) 248-254. DOI= http://dx.doi.org/10.1016/0003-2697(76)90527-3. [10] Chang, V.S. and Holtzapple, M.T. 2000. Fundamental factors affecting biomass enzymatic reactivity. Appl. Biochem. Biotechnol., 84, (Mar. 2000) 35-37. DOI= http://dx.doi.org/10.1385/ABAB:84-86:1-9:5. [11] Clark, J.H., Deswarte, F.E.I. and Farmer, T.J. 2009. The integration of green chemistry into future biorefineries. Biofpr., 3, (Oct. 2008) 72-90. DOI= http://dx.doi.org/10.1002/bbb.119. [12] Collins, T., Gerday, C. and Feller, G. 2005. Xylanases, xylanases families and extremophilic xylanases. FEMS Microbiol. Rev. 29, (Jun. 2004) 3–23. DOI= http://dx.doi.org/10.1016/j.femsre.2004.06.005. [13] Damaso, M.C.T., Almeida, M.S. and Kurtenbach, E. 2003. Optimized expression of a thermostable xylanase from Thermomyces lanuginosus in Pichia pastoris. Appl. Environ. Microbiol., 69, (Jul. 2003) 6064-6072. DOI= http://dx.doi.org/10.1128/AEM.69.10.6064-6072.2003. [14] Dische, Z.G. 1962. General color reactions. In: Whistler, R.L., Wolfram, M.L. Carbohydrate Chemistry. New York, Academic Press, p.477-512. [15] Dragone, D., Fernandes, B., Vicente, A. A., Teixeira, J.A. 2010. Third generation biofuels from microalgae. Curr Res, Technol. Educ. Top. in Appl. Microbiol. and Microbial Biotechnol., A. Mendez-Vilas (Ed). [16] Duarte, G.C., Moreira, L.R.S., Gómez-Mendoza, D.P., Siqueira, F.G., Batista, L.R., Amaral, L.I.V., Ricart, C.A.O., Ferreira Filho, E.X. 2012. Use of residual biomass from the Textile Industry as carbon source for production of a low-molecular-weight Xylanase from Aspergillus oryzae. Appl. Sci., 2, (Oct. 2012) 754-772. DOI= http://dx.doi.org/10.3390/app2040754. [17] Frigon, J.C., Mehta, P. and Guiot, S.R. 2012. Impact of mechanical, chemical and enzymatic pre-treatments on the methane yield from the anaerobic digestion of switchgrass. Biomass and Bioenergy, 36, (Mar. 2011) 1-11. DOI= http://dx.doi.org/10.1016/j.biombioe.2011.02.013. [18] Guo, Y., Wang, S.Z., Xu, D.H., Gong, Y.M., Ma, H.H., Tang, X.Y. 2010. Review of catalytic supercritical water gasification for hydrogen production from biomass. Renewable and Sustain Energy Rev., 14, (Aug. 2009) 334-343. DOI= http://dx.doi.org/10.1016/j.rser.2009.08.012. [19] Hinnman, R.L. 1994. The changing face of the fermentation industry. Chem. Technol., 24, 45-48. [20] IEA. 2008. Commercial investments in 2nd generation plants. In: Sims R. (ed), From 1st to second generation Biofuel Technologies: An Overview of Current Industry and Rand D activities, Italy, (Nov. 2008) pp 62-82. [21] Iembo, I., Azevedo, M.O., Bloch Junior, C., Ferreira Filho, E.X. 2006. Purification and partial characterization οf a new β-xylosidase from Humicola grisea var. thermoidea. World J. Microbiol. Biotechnol., 22, (Set. 2005) 475-479. DOI= http://dx.doi.org/10.1007/s11274-005-9059-3 [22] Kleinert, M. and Barth, T. 2008. Towards a Lignincellulosic Biorefinery: direct one-step conversion of lignin to Hydrogen-Enriched biofuel. Energy and Fuels, 22, (Jan. 2008) 1371-1379. DOI= http://dx.doi.org/10.1021/ef700631w [23] Li, K., Azadi, P., Collins, R., Tolan, J., Kim, J.S., Karl-Erik, E.L. 2000. Relationships between activities of xylanases and
xylan structures. Enzyme Microbiol. Technol., 27, (Feb. 2000) 89–94. DOI= http://dx.doi.org/10.1016/S0141-0229(00)00190-3. [24] Liao, H., Sun, S., Wang, P., Bi, W., Tan, S., Wei, Z., Mei, X., Liu, D., Raza, W., Shen, Q., Xu, Y. 2014. A new acidophilic endo-β-1,4-xylanase from Penicillium oxalicum: cloning, purification, and insights into the influence of metal ions on xylanase activity. J. of Industrial Microbiol. & Biotechnol., 41, (Ju. 2012) 7, pp 1071-1083. http://dx.doi.org/10.1007/s10295-014-1453-0. [25] Mccready, P., Mccomb, E.A. 1952. Extraction and determination of total pectic materials fruit. Anal. Chem., 24, (Dec. 1952) 1586-1588. DOI= http://dx.doi.org/10.1021/ac60072a033. [26] Miller, G.L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31, (Mar. 1959) 426-429. DOI= http://dx.doi.org/10.1021/ac60147a030. [27] Monlau, F., Barakat, A., Trably Dumas, C., Steyer, J.P., Carrere, H. 2013. Lignocellulosic materials into Biohydrogen and Biomethane: impact of structural features and pretreatment. Crit. Rev. in Environ. Science and Technol., 46, (Oct. 2011) 12217-12225. DOI= http://dx.doi.org/10.1080/10643389.2011.604258. [28] Monte, J. R., Carvalho, W., Milagres, A.M.F. 2010. Use of a mixture of thermophilic enzymes produced by the fungus Thermoascus aurantiacus to enhance the enzymatic hydrolysis of the sugarcane bagasse cellulose. Am. J. Agric. Biol. Science, 5, (Oct. 2010) 468-476. DOI= http://dx.doi.org/ 10.3844/ajabssp.2010.468.476. [29] Mosier, N., Wyman, C., Dale, B., Elander, R., Lee, Y.Y., Holtzapple, M., Ladisch, M. 2005. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresourc. Technol., 96, (Sep. 2004) 673-686. DOI= http://dx.doi.org/10.1016/j.biortech.2004.06.025. [30] Murthy, P. and Naidu, M. 2010. Production and application of xylanase from Penicillium sp. utilizing coffee by-products. Food Bioprocess. Technol. 5, (Feb. 2009) 657–664. DOI= http://dx.doi.org/10.1007/s11947-010-0331-7. [31] Nigam, P.S. and Singh, A. 2010. Production of liquid biofuels from renewable resources. Prog. in Energy and Combust. Sci. 37, (May.2010) 52-68. DOI= http://dx.doi.org/10.1016/j.pecs.2010.01.003. [32] Ntaikou, I., Gavala, H.N., Kornaros, M., Lyberatos, G. 2007. Hydrogen production from sugars and sweet sorghum biomass using Ruminococcus albus. Int. J. Hydrogen. Energy., 33, (May. 2010) 1153-1163. DOI= http://dx.doi.org/10.1016/j.ijhydene.2007.10.053. [33] Ohman, M., Boman, C., Hedman, H., Eklund, R. 2006. Residential combustion performance of pelletized hydrolysis residue from lignocellulosic ethanol production. Energy and Fuels, 20, (Jan. 2006) 1298-1304. DOI= http://dx.doi.org/10.1021/ef058030g. [34] Pakarinen, A., Zhang, J., Brock, T., Maijala, P., Viikari, L. 2012. Enzymatic accessibility of fiber hemp is enhanced by enzymatic or chemical removal of pectin. Bioresourc. Technol., 107, (Dec. 2011) 275-281. DOI= http://dx.doi.org/10.1016/j.biortech.2011.12.101. [35] Pandya, J.J. and Gupte, A. 2012. Production of xylanase under solid-state fermentation by Aspergillus tubingensis JP-1 and its application. Bioprocess Biosyst. Eng., 35, (Jan. 2012) 769-779. DOI= http://dx.doi.org/10.1007/s00449-011-0657-1. [36] Polizeli, M.L.T.M., Rizzatti, A.C.S., Monti, R., Terenzi, H.F., Jorge, J.A., Amorim, D.S. 2005. Xylanases from fungi: properties and industrial applications. Appl. Microbiol.
Holocellulase Production from Penicillium spp in Lignocellulosic Biomass: Purification and Characterization of Xylanases
49
Biotechnol., 67, (Jan. 2005) 577–591. DOI= http://dx.doi.org/10.1007/s00253-005-1904-7. [37] Prakasham, R.S., Subba Rao C.H., Sreenivas Rao, R., Rajesham, S., Sarma, P.N. 2005. Optimization of alkaline protease production by Bacillus sp. using Taguchi methodology. Appl. Biochem. Biotechnol., 120, (Aug. 2004) 133–144. DOI= http://dx.doi.org/10.1385/ABAB:120:2:133. [38] Puls, J. and Poutanen, K. 1989. Mechanisms of enzymatic hydrolysis of hemicelluloses xylans and procedures for determination of the enzyme activities involved. In: Ericksson KEE, Ander P (eds) Proceedings of the 3rd International Conference on Biotechnology in the Pulp and Paper Industry. STFI, Stockholm, pp 93–95, (Jun 1989). [39] Reilly, P.J. 1981. Xylanases, structure and function. In: Hollaender A (ed) Trends in the biology of fermentation for fuels and chemicals. Plenum, New York, pp 111–129 (Jul. 1982). [40] Roy, S., Dutta, T., Sarkar, T.S. and Ghosh, S. 2013. Novel xylanases fromSimplicillium obclavatum MTCC 9604: comparative analysis of production, purification and characterization of enzyme from submerged and solid state fermentation. SpringerPlus, 2, (Aug. 2013) 382. http://dx.doi.org/ 10.1186/2193-1801-2-382 [41] Salmen, L. and Olsson, M. 1998. Interaction between hemicelluloses, lignin and cellulose: Structure property relationships. J. Pulp and Paper Sci., 24, 99-103. DOI= http://dx.doi.org/35400007511992.0050. [42] Siqueira, F.G., Siqueira, E.G., Jaramillo, P.M.D., Silveira, M.H.L., Andreaus, J., Couto, F.A., Batista, L.R., Filho, E.X.F. 2010. The potential of agro-industrial residues for production of holocellulase from filamentous fungi. Int. Biodeterior. Biodegrad., 64, (Nov. 2009) 20–26. DOI= http://dx.doi.org/10.1016/j.ibiod.2009.10.002. [43] Subramaniyan, S. and Prema, P. 2002. Biotechnology of microbial xylanases: enzymology, molecular biology, and application. Crit. Rev. Biotechnol., 22, (Sep.2008) 33–64. DOI= http://dx.doi.org/10.1080/07388550290789450. [44] Taherzadeh, M.J. and Karimi, K. 2008. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production: A review. Intern. J. Mol. Sci., 9, (Sep. 2008) 1621-1651. DOI= http://dx.doi.org/10.3390/ijms9091621. [45] Tong, X., Smith, L.H. and McCarthy, P.L. 1990. Methane fermentation of selected lignocellulosic materials. Biomass, 21, (Jun. 1989) 239. DOI= http://dx.doi.org/10.1016/0144-4565(90)90075-U. [46] Van Soest, P.J. 1967. Development of a comprehensive system of feed analyses and its application to forages. J. Anim. Sci., 26,119-28. DOI= http://dx.doi.org/10.2134/jas1967.261119x. [47] Vanholme, R., Demedts, B., Morreel, K., Ralph, J., Boerjan, W. 2010. Lignin Biosynthesis and Structure. Plant Physiol., 153, (Jul. 2010) 895-905. DOI= http://dx.doi.org/10.1104. [48] Watanabe, M., Inomata, H., Osada, M., Sato, T., Adschiri, T., Arai, K. 2003. Catalytic effects of NaOH and ZrO2 for partial oxidative gasification of n-hexadecane and lignin in supercritical water. Fuel, 82, (Oct. 2002) 545-552. DOI= http://dx.doi.org/10.1016/S0016-2361(02)00320-4. [49] Yuan, J.S., Tiller, K.H., Al-Ahmad, H., Stewart, N.R., Stewart, Jr. C.N. 2008. Plants to power: bioenergy to fuel the future. Trends in Plant Sci., 13, (Jul.2008) 421-429. DOI= http://dx.doi.org/10.1016/j.tplants.2008.06.001.