Investigating the effects of musical training on functional brain development with a novel Melodic MMN paradigm Vesa Putkinen a,b,⇑ , Mari Tervaniemi a,b , Katri Saarikivi a,b , Nathalie de Vent c , Minna Huotilainen a,b,d a Cognitive Brain Research Unit, Cognitive Science, Institute of Behavioural Sciences, University of Helsinki, Helsinki, Finland b Finnish Centre of Excellence for Interdisciplinary Music Research, University of Jyväskylä, Jyväskylä, Finland c Brain and Cognition, Faculty of Social and Behavioural Sciences, University of Amsterdam, Amsterdam, Netherlands d Finnish Institute of Occupational Health, Helsinki, Finland article info Article history: Received 29 August 2013 Revised 15 December 2013 Accepted 8 January 2014 Available online 23 January 2014 Keywords: Neuroplasticity Event-relater potentials (ERP) Mismatch negativity (MMN) Musical training Brain development Auditory skills abstract Sensitivity to changes in various musical features was investigated by recording the mismatch negativity (MMN) auditory event-related potential (ERP) in musically trained and nontrained children semi-longi- tudinally at the ages of 9, 11, and 13 years. The responses were recorded using a novel Melodic multi-fea- ture paradigm which allows fast (<15 min) recording of an MMN profile for changes in melody, rhythm, musical key, timbre, tuning and timing. When compared to the nontrained children, the musically trained children displayed enlarged MMNs for the melody modulations by the age 13 and for the rhythm mod- ulations, timbre deviants and slightly mistuned tones already at the age of 11. Also, a positive mismatch response elicited by delayed tones was larger in amplitude in the musically trained than in the non- trained children at age 13. No group differences were found at the age 9 suggesting that the later enhancement of the MMN in the musically trained children resulted from training and not pre-existing difference between the groups. The current study demonstrates the applicability of the Melodic multi- feature paradigm in school-aged children and indicates that musical training enhances auditory discrim- ination for musically central sound dimensions in pre-adolescence. Ó 2014 Elsevier Inc. All rights reserved. 1. Introduction In various domains, expertise and long-term experience have been linked to specific neuroplastic changes in the brain (e.g., Cheour et al., 1998; Gauthier, Skudlarski, Gore, & Anderson, 2000; Lazar et al., 2005; Maguire et al., 2000). Musicianship is a prime example: In individuals who have mastered a musical instrument, brain function and structure show evidence of adapta- tion to the perceptual, motor, and cognitive demands of long-term musical training (Jäncke, 2009; Pantev & Herholz, 2011). The auditory event-related potential (ERP) response mismatch negativity (MMN) has been extensively used to compare the accuracy of auditory change detection between adult musicians and non-musicians (Tervaniemi, 2009). The MMN is elicited by sounds that deviate from a context established by the preceding sounds. Therefore, the elicitation of the MMN is thought to require rapid learning of invariant aspects of auditory input and cortical detection of auditory events that violate these regularities (Näätänen, Paavilainen, Rinne, & Alho, 2007; Winkler, Denham, & Nelken, 2009). Since the elicitation of the MMN does not require focused attention towards the stimuli the MMN appears to result from fairly automatic neural processes (Näätänen et al., 2007). Relative to non-musicians, musicians display larger MMNs especially to changes in musical sounds such as chords (Brattico et al., 2009; Koelsch, Schröger, & Tervaniemi, 1999), melodies (Brattico, Tervaniemi, Näätänen, & Peretz, 2006; Fujioka, Trainor, Ross, Kakigi, & Pantev, 2004, 2005; Tervaniemi, Rytkönen, Schröger, Ilmoniemi, & Näätänen, 2001) and rhythms (Vuust et al., 2005) as well as in other complex auditory stimuli (Herholz, Boh, & Pantev, 2011; van Zuijen, Sussman, Winkler, Näätänen, & Tervaniemi, 2005). Recent studies have shown that the MMN is enlarged in musically trained individuals already in childhood (Virtala, Huotilainen, Putkinen, Makkonen, & Tervaniemi, 2012; Meyer et al., 2011; Chobert, Marie, François, Schön, & Besson, http://dx.doi.org/10.1016/j.nlm.2014.01.007 1074-7427/Ó 2014 Elsevier Inc. All rights reserved. Abbreviations: EEG, electroencephalography; EOG, electro-oculogram; ERP, even-related potential; LDN, late discriminative negativity; MMN, mismatch negativity; SES, socioeconomic status. ⇑ Corresponding author at: Cognitive Brain Research Unit, Institute of Behav- ioural Sciences, University of Helsinki, P.O. Box 9, Helsinki FIN-00014, Finland. Fax: +358 9 191 29450. E-mail address: vesa.putkinen@helsinki.fi (V. Putkinen). Neurobiology of Learning and Memory 110 (2014) 8–15 Contents lists available at ScienceDirect Neurobiology of Learning and Memory journal homepage: www.elsevier.com/locate/ynlme
Transcript
Neurobiology of Learning and Memory 110 (2014) 8–15
Vesa Putkinen a,b,⇑, Mari Tervaniemi a,b, Katri Saarikivi a,b, Nathalie de Vent c, Minna Huotilainen a,b,d
a Cognitive Brain Research Unit, Cognitive Science, Institute of Behavioural Sciences, University of Helsinki, Helsinki, Finlandb Finnish Centre of Excellence for Interdisciplinary Music Research, University of Jyväskylä, Jyväskylä, Finlandc Brain and Cognition, Faculty of Social and Behavioural Sciences, University of Amsterdam, Amsterdam, Netherlandsd Finnish Institute of Occupational Health, Helsinki, Finland
a r t i c l e i n f o a b s t r a c t
Article history:Received 29 August 2013Revised 15 December 2013Accepted 8 January 2014Available online 23 January 2014
Sensitivity to changes in various musical features was investigated by recording the mismatch negativity(MMN) auditory event-related potential (ERP) in musically trained and nontrained children semi-longi-tudinally at the ages of 9, 11, and 13 years. The responses were recorded using a novel Melodic multi-fea-ture paradigm which allows fast (<15 min) recording of an MMN profile for changes in melody, rhythm,musical key, timbre, tuning and timing. When compared to the nontrained children, the musically trainedchildren displayed enlarged MMNs for the melody modulations by the age 13 and for the rhythm mod-ulations, timbre deviants and slightly mistuned tones already at the age of 11. Also, a positive mismatchresponse elicited by delayed tones was larger in amplitude in the musically trained than in the non-trained children at age 13. No group differences were found at the age 9 suggesting that the laterenhancement of the MMN in the musically trained children resulted from training and not pre-existingdifference between the groups. The current study demonstrates the applicability of the Melodic multi-feature paradigm in school-aged children and indicates that musical training enhances auditory discrim-ination for musically central sound dimensions in pre-adolescence.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
In various domains, expertise and long-term experience havebeen linked to specific neuroplastic changes in the brain (e.g.,Cheour et al., 1998; Gauthier, Skudlarski, Gore, & Anderson,2000; Lazar et al., 2005; Maguire et al., 2000). Musicianship is aprime example: In individuals who have mastered a musicalinstrument, brain function and structure show evidence of adapta-tion to the perceptual, motor, and cognitive demands of long-termmusical training (Jäncke, 2009; Pantev & Herholz, 2011).
The auditory event-related potential (ERP) response mismatchnegativity (MMN) has been extensively used to compare theaccuracy of auditory change detection between adult musicians
and non-musicians (Tervaniemi, 2009). The MMN is elicited bysounds that deviate from a context established by the precedingsounds. Therefore, the elicitation of the MMN is thought to requirerapid learning of invariant aspects of auditory input and corticaldetection of auditory events that violate these regularities(Näätänen, Paavilainen, Rinne, & Alho, 2007; Winkler, Denham, &Nelken, 2009). Since the elicitation of the MMN does not requirefocused attention towards the stimuli the MMN appears to resultfrom fairly automatic neural processes (Näätänen et al., 2007).Relative to non-musicians, musicians display larger MMNsespecially to changes in musical sounds such as chords (Bratticoet al., 2009; Koelsch, Schröger, & Tervaniemi, 1999), melodies(Brattico, Tervaniemi, Näätänen, & Peretz, 2006; Fujioka, Trainor,Ross, Kakigi, & Pantev, 2004, 2005; Tervaniemi, Rytkönen,Schröger, Ilmoniemi, & Näätänen, 2001) and rhythms (Vuustet al., 2005) as well as in other complex auditory stimuli (Herholz,Boh, & Pantev, 2011; van Zuijen, Sussman, Winkler, Näätänen, &Tervaniemi, 2005). Recent studies have shown that the MMN isenlarged in musically trained individuals already in childhood(Virtala, Huotilainen, Putkinen, Makkonen, & Tervaniemi, 2012;Meyer et al., 2011; Chobert, Marie, François, Schön, & Besson,
V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15 9
2011). Although the aforementioned cross-sectional studies withinone age group cannot tease apart the influence of training andpre-existing factors, the common interpretation drawn from thesefindings is that musical training enhances automatic memory-based cortical discrimination of sound changes.
Importantly, recent longitudinal studies provide stronger evi-dence in favor of a causal role of training in such effects. Chobertet al. (2012) showed that already within 12 months 8–10-yearold children who had been randomly assigned to musical grouplessons showed a larger increase in MMN amplitude than controlchildren assigned to painting lessons. Putkinen et al. (2013), inturn, found that between the ages of 7 and 13 years the MMN elic-ited by minor chord deviants among major chord standards grewmore in amplitude in musically trained children than in childrenwithout musical training. No group difference in the MMN ampli-tude was found at the baseline measurement at the age 7 indicat-ing that the enhancement of the MMN in the musically trainedchildren emerged as a result of the training.
In recent years, the so called multi-feature paradigm (Näätänen,Pakarinen, Rinne, & Takegata, 2004) has been increasingly used torecord the MMN. In contrast to the oddball paradigm that has beentraditionally employed in MMN studies, the multi-feature para-digm enables fast recording of responses to several deviant typesin one stimulus sequence and is therefore well suited for studiesin children (Lovio et al., 2009; Putkinen, Niinikuru, Lipsanen, Ter-vaniemi, & Huotilainen, 2011; Sambeth et al., 2009). The originalmulti-feature paradigm was designed to probe fairly basic, low-le-vel auditory discrimination skills, such as detecting changes in fre-quency or duration of a repeating tone. Since such skills appear todevelop irrespective of musical training the traditional multi-fea-ture paradigm has not been very effective in differentiating musi-cally trained and nontrained individuals (Putkinen et al., 2013;Tervaniemi et al., 2006). Specifically, with the exception of a loca-tion deviant, neither Putkinen et al. (2013) nor Tervaniemi et al.(2006) found evidence of clear MMN amplitude differences be-tween musically trained and nontrained individuals. In contrast,multi-feature paradigms with subtle and musically relevant devi-ants such as the recently introduced Melodic multi-feature paradigm(Huotilainen, Putkinen, & Tervaniemi, 2009) might be more sensi-tive to the effects of musical training. Unlike the traditional multi-feature paradigm, the Melodic multi-feature paradigm is com-posed of short melodies and includes violations of complex spec-tral and temporal regularities, namely, melody, rhythm, key,timbre, tuning and timing. Furthermore, the stimuli are presentedso that frequent updating of the memory representation for themelody, rhythm, and key is needed for detecting the changes inthese features (as described below). A study employing theMelodic multi-feature paradigm in adult folk musicians and non-musicians found enlarged MMNs for the mistuned sounds as wellas group differences in the scalp topographies for the melody,rhythm, timbre, and timing MMNs (Tervaniemi, submitted forpublication). The paradigm has also been used successfully toobtain mismatch response profiles from 2 to 3-year old children(Huotilainen et al., 2009). Therefore, the Melodic multi-featureparadigm shows promise as a method for studying how musicaltraining influences the development of musically central auditoryskills in children.
Here, we employ the Melodic multi-feature paradigm semi-longitudinally to examine changes over the ages of 9–13 in theencoding of melody, rhythm, key, timbre, tuning, and timing inchildren who play a musical instrument and in children with nomusical training. Based on previous studies, it was expected thatwith age and accumulation of training the musically trainedchildren would show progressively larger MMNs relative to thecontrol children. In contrast, no group differences were expectedat the early stages of training (cf. Putkinen et al., 2013).
2. Methods
2.1. Subjects
Data from 185 recordings from 117 children are reported here.The Music group consisted of 55 children who had started takingmusic lessons approximately at the age of seven and attended amusic-oriented elementary school. The music education integratedin the daily curriculum had an emphasis on classical music and in-cluded weekly music theory studies as well as solo instrument les-sons and orchestra and choir practice. The instruments played bythe children included, among others, violin, viola, cello, doublebass, guitar, and flute. The Control group consisted of 62 childrenwithout formal musical training or music-related hobbies. A greatmajority of the children in the Control group frequently partici-pated in an adult-guided extra-curricular activity (mostly teamsports). The number of children in the two groups at each ageand their gender distribution are given in Table 1.
The current study is a part of a larger ongoing longitudinalstudy in which the same children are invited to participate in sev-eral EEG experiments and a new group of children is recruitedevery two years. Therefore, the children who were recruited earlierhad already participated in several measurements while those re-cruited last had been measured only once. There were also 12recordings from which the data were discarded because of toofew accepted trials. Subject attrition also occurred due to schedul-ing issues or because some children were not reached at the timewhen the recordings were conducted. The number of childrenwho participated in one, two or three recordings was 24, 26, and5 for the Music group and 36, 20, and 6 for the Control group.
The children in both groups were mainly from middle class orupper middle class families in the Helsinki area. The socioeconomicstatus (SES) of the children at age 13 was indexed by parental in-come and education both measured on 6-step scales (Income scalein steps of 1000 euros from 1 = under a 1000 Euros/month to6 = over 5000/month; Education scale: 1 = comprehensive school,2 = upper secondary school or vocational school, 3 = a higher de-gree than upper secondary school or vocational school which isnot a bachelor’s, master’s, licenciate, or doctoral degree, 4 = Bache-lor’s degree or equivalent, 5 = Master’s degree or equivalent,6 = licenciate or doctoral level degree). A composite SES score foreach child was formed by averaging together the answers of bothparents to these questions. There was no statistically significantdifference between the groups in SES at age 13 when the group dif-ferences in response amplitudes were expected to be largest(t(53) = 1.175, p = .245, SES data was not obtained from 3 childrenin the Music group and 4 children in the Control group). Accordingto the answers given by the parents in a questionnaire regardingtheir child’s health status, none of the children had hearing deficitsor neurological impairments.
After the nature of the experiment and the rights of the partic-ipants were explained to the parents and children, a written con-sent was obtained from the parents and a verbal one from thechildren. The children were rewarded with movie tickets and atoy, game or a book for their participation. The experiment proto-
10 V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15
col was approved by the Ethical Committee of the former Depart-ment of Psychology, University of Helsinki, Finland.
2.2. Procedure
In addition to the experiment reported here, the session in-cluded several other experiments (reported elsewhere) and tookaltogether 2 h including the attachment and detachment of elec-trodes and one break halfway through session. During the experi-ment, the participants sat in an armchair watching a muted,captioned, self-chosen movie in an electrically shielded and sound-proofed room. They were asked to ignore the sounds and to avoidunnecessary movements. The stimuli were presented though head-phones (Sony Dynamic Stereo Headphones, MDR-7506) binaurallyat 65 dB.
2.3. Stimuli
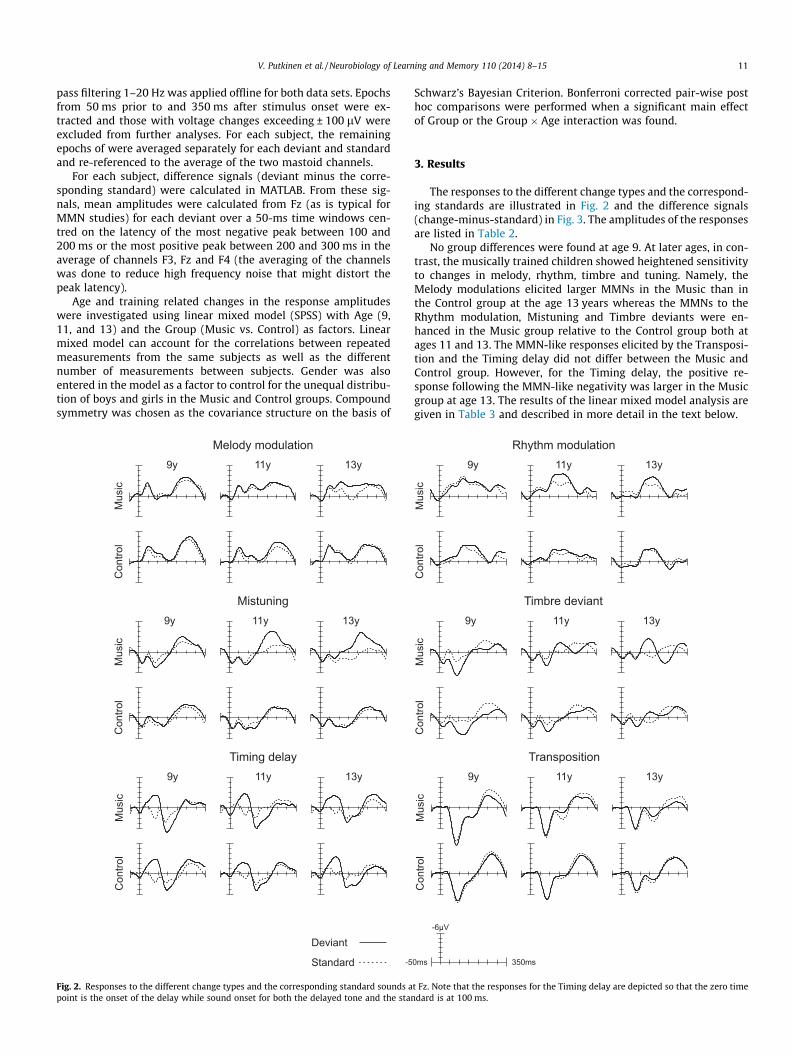
Fig. 1 gives an illustration of the experimental paradigm. Thestimuli consisted of 360 melodies played with a synthesized pianotimbre (expect for the Timbre deviants, see below). The melodiesconformed to the tonal conventions of Western music. They werecomposed of a major triad chord (i.e. a chord composed of threesimultaneously sounded tones: the so called root, third and fifthwhich in the case of C-major triad are C, E and G, respectively) fol-lowed by five tones. The chord at the beginning of each melodywas either a B[, B, C, C], D or a E[ major triad. First inversions ofthe triads were used so that the root was the highest in pitch(the third being the lowest and fifth the second highest) and there-by the most audible of the tones contained in the chords. Each ofthe four following tones was either the second, third, fourth orthe fifth tone of the major scale with a tonic corresponding tothe root of the chord at the beginning of the melody. The last toneof the melody was always the tonic of the scale. The fundamentalfrequencies of the tones used to construct the stimuli ranged from233.1 to 466.2 Hz.
The duration of the chord at the beginning of each melody was300 ms. The duration of two out of the four following tones was125 ms (short inter-tones), and the duration of the other twowas 300 ms (long inter-tones). The final tone of each melody wasalways the tonic and was 575 ms in duration. The chord and thetones within a melody were separated by 50-ms silent intervalsand the last tone of the melody was followed by a 125-ms silentperiod lasting until the beginning of the next melody. Therefore,the interval from the beginning of each melody to the beginningof the next one was 2.1 s in total. The duration of the whole stim-ulus sequence was approximately 13 min.
Occasionally one of six changes took place in the melodies. In 80melodies, one of the long inter-tones at the fourth or fifth positionof the melody was replaced with another in-key tone (Melody mod-ulation, see Fig. 1). In 72 melodies, the duration of two successivetones were switched, i.e., the rhythmical pattern was changed(Rhythm modulation). Ninety-six of the melodies were transposedby a semitone up or down (Transposition). In 96 melodies, one of
Transposition
Rhythm modulation
Melody mod
Transposition standard
Tim
Freq
uenc
y
2100 4200
Timbre deviantTiming standard
Fig. 1. An illustration of the Melo
the long inter-tones or the final tones was played with a flute tim-bre instead of the piano timbre (Timbre deviant). In 72 melodies,one of the long inter-tones was mistuned by a ½ semitone, i.e., lessthan 3% (Mistuning). Finally, in 100 melodies, one of the short orlong inter-tones or the final tones was presented 100 ms too late(Timing delay).
After a Melody or Rhythm modulation or a Transposition, themelody was repeated with the new rhythmic or pitch pattern orin the new key until the next corresponding change at least onceand on average 3–4 times in the so called roving standard manner(cf. Cowan, Winkler, Teder, & Näätänen, 1993).
The responses to the changes were compared to responses elic-ited by standard tones that were matched in duration and positionwithin the melody for each change type. The responses to the Mel-ody modulation and Mistuning were compared to the responseselicited by the long inter-tones. For the Transpositions, the stan-dards were the chords at the beginning of non-transposed melo-dies. For the Rhythm modulations and Timing delays, in turn,both the long inter-tones and short inter-tones served as the stan-dards. Finally, for the Timbre deviants, the standards included thelong inter-tones and the final tones at the end of the melodies. Fur-thermore, since our preliminary analyses of the data indicated thatthe early part of the response to the changes and the long latencyresponses evoked by the preceding sounds overlapped, the stan-dards were also matched separately for each change type by theduration and type of the preceding sound (i.e., inter-tone vs. chord;change vs. repeated tone) to avoid differences in the responses tothe standards and changes arising from the differential responsesto the preceding sounds.
2.4. EEG recording
The EEG was recorded either with a Neuroscan or a BioSemi Ac-tive-Two system. The respective percentages of the recordings con-ducted with the Biosemi system at the age of 9, 11, and 13 were 10%,24%, and 100%. For the recordings conducted with the Neuroscansystem, the EEG (filter bandwith: 0.10–70 Hz ; sampling rate:500 Hz) was registered from the channels F3, Fz, F4, C3, Cz, C4, P3,Pz, P4 (according to the International 10–20 system), and the leftand right mastoids using Ag/AgCl electrodes with a common refer-ence electrode placed at the nose. For the recordings conductedusing the BioSemi systen, the EEG (DC-102 Hz, sampling rate:512 Hz) was registered with 64 active Ag-AgCl electrodes from thescalp (according to the International 10–20 system) and additionalactive electrodes placed at the right and left mastoid and nose. Theelectro-oculogram (EOG) was registered with electrodes situatedbelow and at the outer canthus of the right eye in all recordings.
2.5. Data analysis
The data were analyzed using the BESA 5.1 software. For the 64-channel Biosemi data, noisy channels were interpolated or ex-cluded, the automatic ocular artifact correction incorporated inBESA was used, and the data were down-sampled to 500 Hz. Band
ulation
Melody/Timbre standard
Rhythm standard
Transposition standard
e (ms)6300
100 ms
Timing standardMistuning
Mistuning standard
Timing delay
Transposition
8400
dic multi-feature paradigm.
V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15 11
pass filtering 1–20 Hz was applied offline for both data sets. Epochsfrom 50 ms prior to and 350 ms after stimulus onset were ex-tracted and those with voltage changes exceeding ± 100 lV wereexcluded from further analyses. For each subject, the remainingepochs of were averaged separately for each deviant and standardand re-referenced to the average of the two mastoid channels.
For each subject, difference signals (deviant minus the corre-sponding standard) were calculated in MATLAB. From these sig-nals, mean amplitudes were calculated from Fz (as is typical forMMN studies) for each deviant over a 50-ms time windows cen-tred on the latency of the most negative peak between 100 and200 ms or the most positive peak between 200 and 300 ms in theaverage of channels F3, Fz and F4 (the averaging of the channelswas done to reduce high frequency noise that might distort thepeak latency).
Age and training related changes in the response amplitudeswere investigated using linear mixed model (SPSS) with Age (9,11, and 13) and the Group (Music vs. Control) as factors. Linearmixed model can account for the correlations between repeatedmeasurements from the same subjects as well as the differentnumber of measurements between subjects. Gender was alsoentered in the model as a factor to control for the unequal distribu-tion of boys and girls in the Music and Control groups. Compoundsymmetry was chosen as the covariance structure on the basis of
Melody modulation
Mistuning
Mus
ic
9y 11y 13y
9y 11y 13y
Mus
icC
ontro
l
Timing delay9y 11y 13y
Mus
icC
ontro
lC
ontro
l
Standard
Deviant
-5
Fig. 2. Responses to the different change types and the corresponding standard sounds apoint is the onset of the delay while sound onset for both the delayed tone and the stan
Schwarz’s Bayesian Criterion. Bonferroni corrected pair-wise posthoc comparisons were performed when a significant main effectof Group or the Group � Age interaction was found.
3. Results
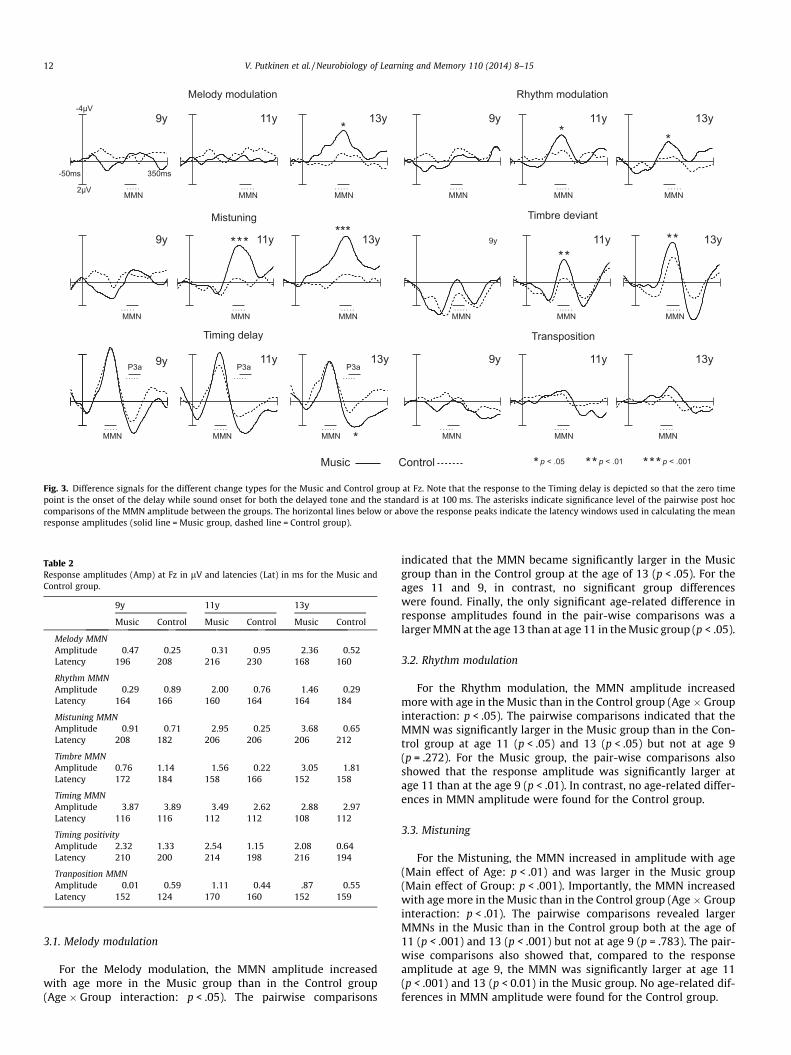
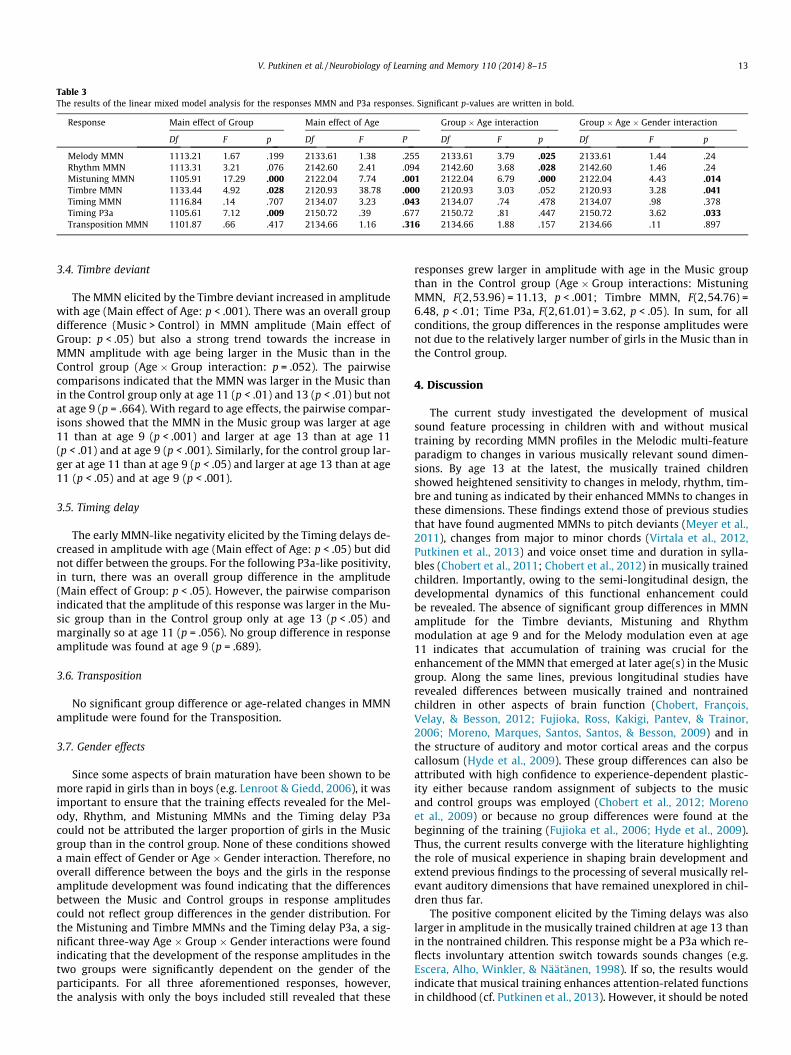
The responses to the different change types and the correspond-ing standards are illustrated in Fig. 2 and the difference signals(change-minus-standard) in Fig. 3. The amplitudes of the responsesare listed in Table 2.
No group differences were found at age 9. At later ages, in con-trast, the musically trained children showed heightened sensitivityto changes in melody, rhythm, timbre and tuning. Namely, theMelody modulations elicited larger MMNs in the Music than inthe Control group at the age 13 years whereas the MMNs to theRhythm modulation, Mistuning and Timbre deviants were en-hanced in the Music group relative to the Control group both atages 11 and 13. The MMN-like responses elicited by the Transposi-tion and the Timing delay did not differ between the Music andControl group. However, for the Timing delay, the positive re-sponse following the MMN-like negativity was larger in the Musicgroup at age 13. The results of the linear mixed model analysis aregiven in Table 3 and described in more detail in the text below.
Rhythm modulation
Timbre deviant
Transposition
9y 11y 13y
Mus
icC
ontro
l
9y 11y 13y
Mus
icC
ontro
l
9y 11y 13y
Mus
icC
ontro
l
-6µV
0ms 350ms
t Fz. Note that the responses for the Timing delay are depicted so that the zero timedard is at 100 ms.
Melody modulation Rhythm modulation
Timbre deviantMistuning
Timing delay Transposition
-4µV
2µV
-50ms 350ms
*
*
* *
****
******
Music Control **** **p < .05 p < .01 p < .001
9y 11y 13y 9y 11y 13y
9y 11y 13y9y 11y 13y
9y 11y 13y 9y 11y 13y
MMNMMN MMN MMNMMNMMN
MMN MMNMMNMMN MMNMMN
MMN MMNMMNMMN MMNMMN
P3a P3a P3a
Fig. 3. Difference signals for the different change types for the Music and Control group at Fz. Note that the response to the Timing delay is depicted so that the zero timepoint is the onset of the delay while sound onset for both the delayed tone and the standard is at 100 ms. The asterisks indicate significance level of the pairwise post hoccomparisons of the MMN amplitude between the groups. The horizontal lines below or above the response peaks indicate the latency windows used in calculating the meanresponse amplitudes (solid line = Music group, dashed line = Control group).
Table 2Response amplitudes (Amp) at Fz in lV and latencies (Lat) in ms for the Music andControl group.
12 V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15
3.1. Melody modulation
For the Melody modulation, the MMN amplitude increasedwith age more in the Music group than in the Control group(Age � Group interaction: p < .05). The pairwise comparisons
indicated that the MMN became significantly larger in the Musicgroup than in the Control group at the age of 13 (p < .05). For theages 11 and 9, in contrast, no significant group differenceswere found. Finally, the only significant age-related difference inresponse amplitudes found in the pair-wise comparisons was alarger MMN at the age 13 than at age 11 in the Music group (p < .05).
3.2. Rhythm modulation
For the Rhythm modulation, the MMN amplitude increasedmore with age in the Music than in the Control group (Age � Groupinteraction: p < .05). The pairwise comparisons indicated that theMMN was significantly larger in the Music group than in the Con-trol group at age 11 (p < .05) and 13 (p < .05) but not at age 9(p = .272). For the Music group, the pair-wise comparisons alsoshowed that the response amplitude was significantly larger atage 11 than at the age 9 (p < .01). In contrast, no age-related differ-ences in MMN amplitude were found for the Control group.
3.3. Mistuning
For the Mistuning, the MMN increased in amplitude with age(Main effect of Age: p < .01) and was larger in the Music group(Main effect of Group: p < .001). Importantly, the MMN increasedwith age more in the Music than in the Control group (Age � Groupinteraction: p < .01). The pairwise comparisons revealed largerMMNs in the Music than in the Control group both at the age of11 (p < .001) and 13 (p < .001) but not at age 9 (p = .783). The pair-wise comparisons also showed that, compared to the responseamplitude at age 9, the MMN was significantly larger at age 11(p < .001) and 13 (p < 0.01) in the Music group. No age-related dif-ferences in MMN amplitude were found for the Control group.
Table 3The results of the linear mixed model analysis for the responses MMN and P3a responses. Significant p-values are written in bold.
Response Main effect of Group Main effect of Age Group � Age interaction Group � Age � Gender interaction
V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15 13
3.4. Timbre deviant
The MMN elicited by the Timbre deviant increased in amplitudewith age (Main effect of Age: p < .001). There was an overall groupdifference (Music > Control) in MMN amplitude (Main effect ofGroup: p < .05) but also a strong trend towards the increase inMMN amplitude with age being larger in the Music than in theControl group (Age � Group interaction: p = .052). The pairwisecomparisons indicated that the MMN was larger in the Music thanin the Control group only at age 11 (p < .01) and 13 (p < .01) but notat age 9 (p = .664). With regard to age effects, the pairwise compar-isons showed that the MMN in the Music group was larger at age11 than at age 9 (p < .001) and larger at age 13 than at age 11(p < .01) and at age 9 (p < .001). Similarly, for the control group lar-ger at age 11 than at age 9 (p < .05) and larger at age 13 than at age11 (p < .05) and at age 9 (p < .001).
3.5. Timing delay
The early MMN-like negativity elicited by the Timing delays de-creased in amplitude with age (Main effect of Age: p < .05) but didnot differ between the groups. For the following P3a-like positivity,in turn, there was an overall group difference in the amplitude(Main effect of Group: p < .05). However, the pairwise comparisonindicated that the amplitude of this response was larger in the Mu-sic group than in the Control group only at age 13 (p < .05) andmarginally so at age 11 (p = .056). No group difference in responseamplitude was found at age 9 (p = .689).
3.6. Transposition
No significant group difference or age-related changes in MMNamplitude were found for the Transposition.
3.7. Gender effects
Since some aspects of brain maturation have been shown to bemore rapid in girls than in boys (e.g. Lenroot & Giedd, 2006), it wasimportant to ensure that the training effects revealed for the Mel-ody, Rhythm, and Mistuning MMNs and the Timing delay P3acould not be attributed the larger proportion of girls in the Musicgroup than in the control group. None of these conditions showeda main effect of Gender or Age � Gender interaction. Therefore, nooverall difference between the boys and the girls in the responseamplitude development was found indicating that the differencesbetween the Music and Control groups in response amplitudescould not reflect group differences in the gender distribution. Forthe Mistuning and Timbre MMNs and the Timing delay P3a, a sig-nificant three-way Age � Group � Gender interactions were foundindicating that the development of the response amplitudes in thetwo groups were significantly dependent on the gender of theparticipants. For all three aforementioned responses, however,the analysis with only the boys included still revealed that these
responses grew larger in amplitude with age in the Music groupthan in the Control group (Age � Group interactions: MistuningMMN, F(2,53.96) = 11.13, p < .001; Timbre MMN, F(2,54.76) =6.48, p < .01; Time P3a, F(2,61.01) = 3.62, p < .05). In sum, for allconditions, the group differences in the response amplitudes werenot due to the relatively larger number of girls in the Music than inthe Control group.
4. Discussion
The current study investigated the development of musicalsound feature processing in children with and without musicaltraining by recording MMN profiles in the Melodic multi-featureparadigm to changes in various musically relevant sound dimen-sions. By age 13 at the latest, the musically trained childrenshowed heightened sensitivity to changes in melody, rhythm, tim-bre and tuning as indicated by their enhanced MMNs to changes inthese dimensions. These findings extend those of previous studiesthat have found augmented MMNs to pitch deviants (Meyer et al.,2011), changes from major to minor chords (Virtala et al., 2012,Putkinen et al., 2013) and voice onset time and duration in sylla-bles (Chobert et al., 2011; Chobert et al., 2012) in musically trainedchildren. Importantly, owing to the semi-longitudinal design, thedevelopmental dynamics of this functional enhancement couldbe revealed. The absence of significant group differences in MMNamplitude for the Timbre deviants, Mistuning and Rhythmmodulation at age 9 and for the Melody modulation even at age11 indicates that accumulation of training was crucial for theenhancement of the MMN that emerged at later age(s) in the Musicgroup. Along the same lines, previous longitudinal studies haverevealed differences between musically trained and nontrainedchildren in other aspects of brain function (Chobert, François,Velay, & Besson, 2012; Fujioka, Ross, Kakigi, Pantev, & Trainor,2006; Moreno, Marques, Santos, Santos, & Besson, 2009) and inthe structure of auditory and motor cortical areas and the corpuscallosum (Hyde et al., 2009). These group differences can also beattributed with high confidence to experience-dependent plastic-ity either because random assignment of subjects to the musicand control groups was employed (Chobert et al., 2012; Morenoet al., 2009) or because no group differences were found at thebeginning of the training (Fujioka et al., 2006; Hyde et al., 2009).Thus, the current results converge with the literature highlightingthe role of musical experience in shaping brain development andextend previous findings to the processing of several musically rel-evant auditory dimensions that have remained unexplored in chil-dren thus far.
The positive component elicited by the Timing delays was alsolarger in amplitude in the musically trained children at age 13 thanin the nontrained children. This response might be a P3a which re-flects involuntary attention switch towards sounds changes (e.g.Escera, Alho, Winkler, & Näätänen, 1998). If so, the results wouldindicate that musical training enhances attention-related functionsin childhood (cf. Putkinen et al., 2013). However, it should be noted
14 V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15
that even though this response was approximately in the expectedlatency range of the P3a with regard to the onset of the delay (i.e.the zero time point in Fig. 2), relative to the onset of the delayedtone (i.e. 100 ms in Fig. 2) it falls in the time range of the obligatoryP1 response. In school-aged children, the P1 is enlarged withprolonged inter-stimulus intervals (Sussman, Steinschneider,Gumenyuk, Grushko, & Lawson, 2008). Therefore, it cannot beruled out that a less refractory P1 elicited by the delayed tonecontributed to the positivity seen in the difference signal for theTiming delays. The P1 has also been previously reported to beenlarged in musically trained children (Shahin et al., 2004).
Not only is the Melodic multi-feature paradigm a time-efficientmethod for recording an extensive discrimination profile formusical sounds but was also found to be highly effective in differ-entiating musically trained and nontrained children in the currentstudy. In contrast, a previous study (Putkinen et al., 2013),conducted partly with the same subjects, found no evidence forMMN enhancement in the musically trained children for changesin the frequency, duration, intensity, and temporal structurerecorded in a simpler, non-musical multi-feature paradigm. Thesefindings suggest that musical training facilitates the processing ofsubtle sound changes that are especially relevant for music pro-cessing. Also in adult musicians, enhanced MMNs are most consis-tently found for changes in complex, rather than simple sounds.For instance, Koelsch et al. (1999) found that while a slight pitchchange embedded in a major chord elicited a larger MMN in musi-cians than in non-musicians, an equal change in the frequency of asimple repeating tone did not differentiate the two groups (for cor-responding results pertaining to melodies vs. tones, see Brattico,Näätänen, & Tervaniemi, 2001; Fujioka et al., 2004). The enhanceddiscrimination of fine grained sound changes can obviously beuseful in other, non-musical domains – a case in point being thefacilitated processing of speech sounds seen in musically trainedadults and children (Chobert et al., 2011; Moreno et al., 2009;Musacchia, Sams, Skoe, & Kraus, 2007).
Interestingly, group differences in MMN amplitude emergedrelatively slowly, i.e., they were not seen before the children inthe Music group had received at least 3 years of training. Some pre-vious studies, by contrast, have found neuroplastic changes in chil-dren already after considerably shorter training periods. Forinstance, Fujioka et al. (2006) found that after only six months ofSuzuki violin lessons 6-year-old children showed enhancement ofan early cortical evoked field elicited by violin sounds. A studyby Chobert et al. (2012) suggests that also the MMN can be en-hanced by musical training already within a year in children inschool-aged children. Although Chobert et al. (2012) used a fairlycomplex multi-feature paradigm with speech sounds, the para-digm in the current study was arguably even more demanding.Thus the difficulty of the paradigm might at least in part explainthe slow emergence of the MMN enhancement in the Music group.
The demanding nature of the paradigm probably also explainswhy, except for the responses to the Timing delays, the MMNs toin the control group were fairly small and showed increase inamplitude with age only for the perceptually distinct Timbredeviants. In contrast, the study of Tervaniemi et al. (submittedfor publication) found prominent MMNs in the Melodic multi-feature paradigm not only in the musician group but also innon-musicians (see also Vuust et al., 2011). These results suggestthat the ability to neurally discriminate subtle changes in inter-leaved musical sound regularities develops with age even withoutmusical training but at a slower pace than in individuals with suchtraining. Previous studies suggest similar protracted developmentfor the processing of other types of musical sounds. Firstly, asalready mentioned, Putkinen et al. (2013) obtained a larger MMNto minor chord deviants in musically trained children than in non-trained children. Using highly similar stimuli, Brattico et al. (2009),
however, found no difference in the strength of the magnetic coun-terpart of the MMN between adult musicians and non-musiciansindicating that by adulthood non-musicians had acquired the samelevel of proficiency in detecting minor chord deviants among majorchord standards. Furthermore, a study by Virtala et al. (2012)found an MMN-like response to minor chord deviants in musicallytrained 13-year-old children but no evidence for an MMN inmusically nontrained children. In another study employing thesame stimuli, by contrast, Virtala et al. (2011) did obtain a signifi-cant MMN in adult non-musicians. Together these studies suggestthat without musical training the sensitivity for this particularstimulus contrast develops between age 13 and adulthood.
With regard to the gender effects, it was unequivocally shownthat the enhanced responses in the musically trained children werenot due to the larger proportion of girls in the Music group than inthe Control group. Firstly, the Melody and Rhythm MMNs showedno effects involving the Gender factor. Secondly, for the Mistuningand Timbre MMNs and the Timing P3a, which showed a significantinteraction between factors Age, Group and Gender, a significantlylarger age-related increase in response amplitudes in the Musicgroup was found even when the analysis was done only for themale subjects. Tentatively, it seems that the aforementionedthree-way interactions might in fact have resulted from slightlysteeper increase in response amplitudes in the musically trainedboys than the musically trained girls (see the Supplementary mate-rial). However, these effects should be interpreted with caution asthey were only seen for three out of the six conditions and werethereby not very robust. Furthermore, they were merely incidentalfindings that resulted from the necessity of controlling for the un-equal gender distribution in the statistical model. Therefore, weleave the issue of putative gender differences in the effects of mu-sical training on MMN and P3a maturation to be investigated byfuture studies that—unlike the current one—are specifically de-signed for this purpose.
Returning to the role of training in the differences betweenmusicians and non-musicians, it is evident that long-term practiceis a key determinant of musical ability. Still, some of the compo-nent skills of ‘‘musicality’’, i.e., the perceptual and motor abilitiesneeded for accurate perception and production of music, may verywell show individual differences that reflect inborn, genetic factors(cf. Drayna, Manichaikul, de Lange, Snieder, & Spector, 2001; Pulliet al., 2008; for a discussion, see Levitin, 2012). Even if this is thecase, the extent to which such skills are related to one’s ability tobenefit from musical training and thereby predict whether an indi-vidual can attain high level of musical skills is unclear. Less music-specific individual factors which might have a strong genetic com-ponent such as personality and ability to focus attention obviouslyplay a role in whether an individual is willing to invest time andeffort to master a musical instrument (cf. Corrigall, Schellenberg,& Misura, 2013). Efforts to disentangle the contribution of train-ing-related and other factors in musical skill development willundoubtedly continue to drive future research. For now, the cur-rent results lend support for the causal role of musical training inshaping the development of musically central auditory skills inchildhood.
5. Summary and conclusions
In the current semi-longitudinal study, MMN responses were re-corded from musically trained and nontrained school-aged childrento changes in various features of musical sounds in the Melodicmulti-feature paradigm. In the musically trained children, MMNsto changes in melody, rhythm, timbre and tuning increased inamplitude during the follow-up ultimately reaching higheramplitudes compared to the responses of the nontrained children.
V. Putkinen et al. / Neurobiology of Learning and Memory 110 (2014) 8–15 15
In contrast, very little age-related change was seen in the responsesin the children without musical training indicating that the abilityto automatically detect subtle changes in complex musical soundsas reflected by the MMN develops slowly if at all in school-age.Since no group differences were seen until the age of eleven forany of the changes, the results imply that the enhanced MMNs thatemerged in the Music group were due to the training and not pre-existing group differences. Therefore, the current results speak fora causal role of experience in the heightened sensitivity to changesin various musical features in musically trained children.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.nlm.2014.01.007.
References
Brattico, E., Näätänen, R., & Tervaniemi, M. (2001). Context effects on pitchperception in musicians and nonmusicians: Evidence from event-related-potential recordings. Music Perception, 19(2), 199–222.
Brattico, E., Pallesen, K. J., Varyagina, O., Bailey, C., Anourova, I., Järvenpää, M., et al.(2009). Neural discrimination of nonprototypical chords in music experts andlaymen: An MEG study. Journal of Cognitive Neuroscience, 21(11), 2230–2244.
Brattico, E., Tervaniemi, M., Näätänen, R., & Peretz, I. (2006). Musical scaleproperties are automatically processed in the human auditory cortex. BrainResearch, 1117(1), 162–174.
Cheour, M., Ceponiene, R., Lehtokoski, A., Luuk, A., Allik, J., Alho, K., et al. (1998).Development of language-specific phoneme representations in the infant brain.Nature Neuroscience, 1(5), 351–353.
Chobert, J., François, C., Velay, J., & Besson, M. (2012). Twelve months of activemusical training in 8-to 10-year-old children enhances the preattentiveprocessing of syllabic duration and voice onset time. Cerebral Cortex.
Chobert, J., Marie, C., François, C., Schön, D., & Besson, M. (2011). Enhanced passiveand active processing of syllables in musician children. Journal of CognitiveNeuroscience, 23(12), 3874–3887.
Corrigall, K. A., Schellenberg, E. G., & Misura, N. M. (2013). Music training, cognition,and personality. Frontiers in Psychology, 4.
Cowan, N., Winkler, I., Teder, W., & Näätänen, R. (1993). Memory prerequisites ofmismatch negativity in the auditory event-related potential (ERP). Journal ofExperimental Psychology. Learning, Memory, and Cognition, 19(4), 909.
Drayna, D., Manichaikul, A., de Lange, M., Snieder, H., & Spector, T. (2001). Geneticcorrelates of musical pitch recognition in humans. Science, 291(5510),1969–1972.
Escera, C., Alho, K., Winkler, I., & Näätänen, R. (1998). Neural mechanisms ofinvoluntary attention to acoustic novelty and change. Journal of cognitiveneuroscience, 10(5), 590–604.
Fujioka, T., Ross, B., Kakigi, R., Pantev, C., & Trainor, L. J. (2006). One year of musicaltraining affects development of auditory cortical-evoked fields in youngchildren. Brain, 129(10), 2593–2608.
Fujioka, T., Trainor, L. J., Ross, B., Kakigi, R., & Pantev, C. (2004). Musical trainingenhances automatic encoding of melodic contour and interval structure. Journalof Cognitive Neuroscience, 16(6), 1010–1021.
Fujioka, T., Trainor, L. J., Ross, B., Kakigi, R., & Pantev, C. (2005). Automatic encodingof polyphonic melodies in musicians and nonmusicians. Journal of CognitiveNeuroscience, 17(10), 1578–1592.
Gauthier, I., Skudlarski, P., Gore, J. C., & Anderson, A. W. (2000). Expertise for carsand birds recruits brain areas involved in face recognition. Nature Neuroscience,3(2), 191–197.
Herholz, S. C., Boh, B., & Pantev, C. (2011). Musical training modulates encoding ofhigher-order regularities in the auditory cortex. European Journal ofNeuroscience, 34(3), 524–529.
Huotilainen, M., Putkinen, V., & Tervaniemi, M. (2009). Brain research revealsautomatic musical memory functions in children. Annals of the New YorkAcademy of Sciences, 1169(1), 178–181.
Hyde, K. L., Lerch, J., Norton, A., Forgeard, M., Winner, E., Evans, A. C., et al. (2009).Musical training shapes structural brain development. The Journal ofNeuroscience, 29(10), 3019–3025.
Jäncke, L. (2009). The plastic human brain. Restorative Neurology and Neuroscience,27(5), 521–538.
Koelsch, S., Schröger, E., & Tervaniemi, M. (1999). Superior pre-attentive auditoryprocessing in musicians. NeuroReport, 10(6), 1309–1313.
Lazar, S. W., Kerr, C. E., Wasserman, R. H., Gray, J. R., Greve, D. N., Treadway, M. T.,et al. (2005). Meditation experience is associated with increased corticalthickness. NeuroReport, 16(17), 1893.
Lenroot, R. K., & Giedd, J. N. (2006). Brain development in children and adolescents:Insights from anatomical magnetic resonance imaging. Neuroscience andBiobehavioral Reviews, 30(6), 718–729.
Levitin, D. J. (2012). What does it mean to be musical? Neuron, 73(4), 633–637.Lovio, R., Pakarinen, S., Huotilainen, M., Alku, P., Silvennoinen, S., Näätänen, R., et al.
(2009). Auditory discrimination profiles of speech sound changes in 6-year-oldchildren as determined with the multi-feature MMN paradigm. ClinicalNeurophysiology, 120(5), 916–921.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R.S., et al. (2000). Navigation-related structural change in the hippocampi of taxidrivers. Proceedings of the National Academy of Sciences, 97(8), 4398–4403.
Meyer, M., Elmer, S., Ringli, M., Oechslin, M. S., Baumann, S., & Jäncke, L. (2011).Long-term exposure to music enhances the sensitivity of the auditory system inchildren. European Journal of Neuroscience, 34(5), 755–765.
Moreno, S., Marques, C., Santos, A., Santos, M., & Besson, M. (2009). Musical traininginfluences linguistic abilities in 8-year-old children: More evidence for brainplasticity. Cerebral Cortex, 19(3), 712–723.
Musacchia, G., Sams, M., Skoe, E., & Kraus, N. (2007). Musicians have enhancedsubcortical auditory and audiovisual processing of speech and music.Proceedings of the National Academy of Sciences, 104(40), 15894–15898.
Näätänen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007). The mismatch negativity(MMN) in basic research of central auditory processing: A review. ClinicalNeurophysiology, 118(12), 2544–2590.
Näätänen, R., Pakarinen, S., Rinne, T., & Takegata, R. (2004). The mismatch negativity(MMN): Towards the optimal paradigm. Clinical Neurophysiology, 115(1),140–144.
Pantev, C., & Herholz, S. C. (2011). Plasticity of the human auditory cortex related tomusical training. Neuroscience and Biobehavioral Reviews, 35(10), 2140–2154.
Pulli, K., Karma, K., Norio, R., Sistonen, P., Göring, H. H. H., & Järvelä, I. (2008).Genome-wide linkage scan for loci of musical aptitude in Finnish families:Evidence for a major locus at 4q22. Journal of Medical Genetics, 45(7), 451–456.
Putkinen, V., Niinikuru, R., Lipsanen, J., Tervaniemi, M., & Huotilainen, M. (2012).Fast measurement of auditory event-related potential profiles in 2–3-year-olds.Developmental Neuropsychology, 37(1), 51–75.
Putkinen, V., Tervaniemi, M., Saarikivi, K., Ojala, P., & Huotilainen, M. (2013).Enhanced development of auditory change detection in musically trainedschool-aged children: a longitudinal event-related potential study.Developmental science.
Sambeth, A., Pakarinen, S., Ruohio, K., Fellman, V., van Zuijen, T. L., & Huotilainen, M.(2009). Change detection in newborns using a multiple deviant paradigm: Astudy using magnetoencephalography. Clinical Neurophysiology, 120(3),530–538.
Shahin, A., Roberts, L. E., & Trainor, L. J. (2004). Enhancement of auditory corticaldevelopment by musical experience in children. Neuroreport, 15(12),1917–1921.
Sussman, E., Steinschneider, M., Gumenyuk, V., Grushko, J., & Lawson, K. (2008). Thematuration of human evoked brain potentials to sounds presented at differentstimulus rates. Hearing Research, 236(1), 61–79.
Tervaniemi, M., Castaneda, A., Knoll, M., & Uther, M. (2006). Sound processing inamateur musicians and nonmusicians: event-related potential and behavioralindices. Neuroreport, 17(11), 1225–1228.
Tervaniemi M., Huotilainen, M. & Brattico, E. (submitted for publication). Melodicmulti-feature paradigm reveals auditory profiles in music-sound.
Tervaniemi, M. (2009). Musicians—Same or different? Annals of the New YorkAcademy of Sciences, 1169(1), 151–156.
Tervaniemi, M., Rytkönen, M., Schröger, E., Ilmoniemi, R. J., & Näätänen, R. (2001).Superior formation of cortical memory traces for melodic patterns in musicians.Learning & Memory, 8(5), 295–300.
van Zuijen, T. L., Sussman, E., Winkler, I., Näätänen, R., & Tervaniemi, M. (2005).Auditory organization of sound sequences by a temporal or numericalregularity—A mismatch negativity study comparing musicians and non-musicians. Cognitive Brain Research, 23(2), 270–276.
Virtala, P., Huotilainen, M., Putkinen, V., Makkonen, T., & Tervaniemi, M. (2012).Musical training facilitates the neural discrimination of major versus minorchords in 13-year-old children. Psychophysiology, 49(8), 1125–1132.
Vuust, P., Brattico, E., Glerean, E., Seppänen, M., Pakarinen, S., Tervaniemi, M., et al.(2011). New fast mismatch negativity paradigm for determining the neuralprerequisites for musical ability. Cortex, 47(9), 1091–1098.
Vuust, P., Pallesen, K. J., Bailey, C., van Zuijen, T. L., Gjedde, A., Roepstorff, A., et al.(2005). To musicians, the message is in the meter: Pre-attentive neuronalresponses to incongruent rhythm are left-lateralized in musicians. Neuroimage,24(2), 560–564.
Winkler, I., Denham, S. L., & Nelken, I. (2009). Modeling the auditory scene:Predictive regularity representations and perceptual objects. Trends in CognitiveSciences, 13(12), 532–540.