Involvement of protein kinase C and intracellular Ca 2+ in goldfish brain somatostatin-28 inhibitory action on growth hormone release in goldfish Y. Yu, J.P. Chang * Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada article info Article history: Received 7 January 2010 Revised 17 March 2010 Accepted 14 April 2010 Available online 18 April 2010 Keywords: Goldfish somatostatin signal transduction Identified somatotrope Ca 2+ -imaging Dispersed pituitary cell hormone release PKC abstract Goldfish brain somatostatin-28 (gbSS-28) is present in brain and pituitary tissues of goldfish. We assessed whether gbSS-28 targets Ca 2+ and/or protein kinase C (PKC)-dependent signaling cascades in inhibiting growth hormone (GH) release. gbSS-28 decreased basal GH release from primary cultures of dispersed goldfish pituitary cells and intracellular free calcium levels ([Ca 2+ ] i ) in goldfish somatotropes. gbSS-28 partially reduced [Ca 2+ ] i and GH responses induced by two endogeneous gonadotropin-releasing hormones (GnRHs), salmon (s)GnRH and chicken (c)GnRH-II. Furthermore, gbSS-28 reduced GH increases and abolished [Ca 2+ ] i elevations elicited by two PKC activators, tetradecanoyl 4b-phorbol-13-acetate and dioctanyl glycerol. The PKC inhibitors Gö6976 and Bis II abolished [Ca 2+ ] i responses to PKC activators, but only attenuated GnRH-induced increases in [Ca 2+ ] i and did not alter basal [Ca 2+ ] i . In cells pretreated with Bis II, gbSS-28 further reduced basal [Ca 2+ ] i . Our results suggest that gbSS-28 inhibits GnRH-induced GH release in part by attenuating PKC-mediated GnRH [Ca 2+ ] i signals. gbSS-28 reduces basal GH release also via reduction in [Ca 2+ ] i but PKC is not involved in this regard. Ó 2010 Elsevier Inc. All rights reserved. 1. Introduction The 14 amino acid somatostatin (SS-14) is a highly conserved neuropeptide that serves as the major inhibitory regulator of growth hormone (GH) release in vertebrates (Gahete et al., 2009). In mammals, an N-terminal extended form of SS-14, mSS- 28, is also derived from the same precursor gene as SS-14 (Spiess and Vale, 1980). Immunoreactivity for both SS-14 and mSS-28 are found in the rat and ovine preoptic-anterior hypothalamic re- gion, suggesting that mSS-28 is a hypophysiotropic hormone (Mes- guich et al., 1988; Scanlan et al., 2003). In fact, some earlier studies show that mSS-28 is more potent than SS-14 in inhibiting basal GH secretion from the rat pituitary in vitro and in vivo (Brazeau et al., 1981; Jacovidou and Patel, 1987). However, the possible roles of multiple SSs at the level of the mammalian pituitary and how mul- tiple SSs act on pituitary targets in the regulation of GH levels are not well understood. Neuroendocrine regulation of GH release by SS in fish is more complex. Multiple SS genes encode for multiple SS isoforms in a single species (Trabucchi et al., 1999; Sheridan and Kittilson, 2004; Li et al., 2009). To date, the goldfish is one of the fish species known to possess three SS genes in brain tissues, each encoding a distinct SS prohormone, PSS-I, PSS-II and PSS-III (Lin et al., 1999; Xing et al., 2005). These prohormones give rise to SS-14, goldfish brain (gb)SS-28 and [Pro 2 ]SS-14, respectively (Canosa et al., 2004). All three PSSs are expressed in the hypophysiotropic hypo- thalamus; in addition, PSS-I and PSS-II mRNAs are found in pitui- tary extracts (Yunker et al., 2003). All three endogenous hypothalamic SSs reduce basal GH levels and attenuate the GH secretion responses to neuroendocrine stimulators, including those to the two gonadotropin-releasing hormones native to goldfish (namely salmon (s)GnRH and chicken (c)GnRH-II) (Yunker et al., 2003). These observations suggest that goldfish pituitary cells are exposed to multiple endogenous SSs and they participate in the multifactorial neuroendocrine regulation of GH release in this spe- cies (Canosa et al., 2007). As in mammals, multiple SS receptors (Ssts) have been cloned and identified in the goldfish pituitary, where Sst2 and Sst5 mRNA levels are significantly higher than those in the brain (Lin et al., 2000, 2002). SS-14 and [Pro 2 ]SS-14 bind goldfish Sst2s expressed in COS-7 cells with similar affinity and exhibit similar potency in inhibiting aden- ylate cyclase. On the other hand, gbSS-28 has no effect on forskolin- stimulated cAMP production in COS-7 cells expressing Sst2s but the gbSS-28-selective Sst5s expressed in CCL39 cells are coupled to Gi/o- protein (Lin et al., 2000; Nunn et al., 2002). Interestingly, gbSS-28 is the most potent among the three goldfish SSs in suppressing basal GH secretion, but it is less effective than SS-14 in inhibiting GnRH-in- duced GH release in static incubation of dispersed goldfish pituitary cells (Yunker et al., 2003). These data indicate that goldfish pituitary 0016-6480/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2010.04.008 * Corresponding author. Address: CW405 Biol. Sci. Centre, Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada T6G 2E9. Fax: +1 780 492 9234. E-mail address: [email protected](J.P. Chang). General and Comparative Endocrinology 168 (2010) 71–81 Contents lists available at ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen
Transcript
General and Comparative Endocrinology 168 (2010) 71–81
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Involvement of protein kinase C and intracellular Ca2+ in goldfish brainsomatostatin-28 inhibitory action on growth hormone release in goldfish
Y. Yu, J.P. Chang *
Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada
a r t i c l e i n f o a b s t r a c t
Article history:Received 7 January 2010Revised 17 March 2010Accepted 14 April 2010Available online 18 April 2010
Keywords:Goldfish somatostatin signal transductionIdentified somatotropeCa2+-imagingDispersed pituitary cell hormone releasePKC
0016-6480/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.ygcen.2010.04.008
* Corresponding author. Address: CW405 Biol.Biological Sciences, University of Alberta, EdmontonFax: +1 780 492 9234.
Goldfish brain somatostatin-28 (gbSS-28) is present in brain and pituitary tissues of goldfish. Weassessed whether gbSS-28 targets Ca2+ and/or protein kinase C (PKC)-dependent signaling cascades ininhibiting growth hormone (GH) release. gbSS-28 decreased basal GH release from primary cultures ofdispersed goldfish pituitary cells and intracellular free calcium levels ([Ca2+]i) in goldfish somatotropes.gbSS-28 partially reduced [Ca2+]i and GH responses induced by two endogeneous gonadotropin-releasinghormones (GnRHs), salmon (s)GnRH and chicken (c)GnRH-II. Furthermore, gbSS-28 reduced GH increasesand abolished [Ca2+]i elevations elicited by two PKC activators, tetradecanoyl 4b-phorbol-13-acetate anddioctanyl glycerol. The PKC inhibitors Gö6976 and Bis II abolished [Ca2+]i responses to PKC activators, butonly attenuated GnRH-induced increases in [Ca2+]i and did not alter basal [Ca2+]i. In cells pretreated withBis II, gbSS-28 further reduced basal [Ca2+]i. Our results suggest that gbSS-28 inhibits GnRH-induced GHrelease in part by attenuating PKC-mediated GnRH [Ca2+]i signals. gbSS-28 reduces basal GH release alsovia reduction in [Ca2+]i but PKC is not involved in this regard.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
The 14 amino acid somatostatin (SS-14) is a highly conservedneuropeptide that serves as the major inhibitory regulator ofgrowth hormone (GH) release in vertebrates (Gahete et al.,2009). In mammals, an N-terminal extended form of SS-14, mSS-28, is also derived from the same precursor gene as SS-14 (Spiessand Vale, 1980). Immunoreactivity for both SS-14 and mSS-28are found in the rat and ovine preoptic-anterior hypothalamic re-gion, suggesting that mSS-28 is a hypophysiotropic hormone (Mes-guich et al., 1988; Scanlan et al., 2003). In fact, some earlier studiesshow that mSS-28 is more potent than SS-14 in inhibiting basal GHsecretion from the rat pituitary in vitro and in vivo (Brazeau et al.,1981; Jacovidou and Patel, 1987). However, the possible roles ofmultiple SSs at the level of the mammalian pituitary and how mul-tiple SSs act on pituitary targets in the regulation of GH levels arenot well understood.
Neuroendocrine regulation of GH release by SS in fish is morecomplex. Multiple SS genes encode for multiple SS isoforms in asingle species (Trabucchi et al., 1999; Sheridan and Kittilson,2004; Li et al., 2009). To date, the goldfish is one of the fish speciesknown to possess three SS genes in brain tissues, each encoding a
ll rights reserved.
Sci. Centre, Department of, Alberta, Canada T6G 2E9.
).
distinct SS prohormone, PSS-I, PSS-II and PSS-III (Lin et al., 1999;Xing et al., 2005). These prohormones give rise to SS-14, goldfishbrain (gb)SS-28 and [Pro2]SS-14, respectively (Canosa et al.,2004). All three PSSs are expressed in the hypophysiotropic hypo-thalamus; in addition, PSS-I and PSS-II mRNAs are found in pitui-tary extracts (Yunker et al., 2003). All three endogenoushypothalamic SSs reduce basal GH levels and attenuate the GHsecretion responses to neuroendocrine stimulators, including thoseto the two gonadotropin-releasing hormones native to goldfish(namely salmon (s)GnRH and chicken (c)GnRH-II) (Yunker et al.,2003). These observations suggest that goldfish pituitary cells areexposed to multiple endogenous SSs and they participate in themultifactorial neuroendocrine regulation of GH release in this spe-cies (Canosa et al., 2007).
As in mammals, multiple SS receptors (Ssts) have been cloned andidentified in the goldfish pituitary, where Sst2 and Sst5 mRNA levelsare significantly higher than those in the brain (Lin et al., 2000, 2002).SS-14 and [Pro2]SS-14 bind goldfish Sst2s expressed in COS-7 cellswith similar affinity and exhibit similar potency in inhibiting aden-ylate cyclase. On the other hand, gbSS-28 has no effect on forskolin-stimulated cAMP production in COS-7 cells expressing Sst2s but thegbSS-28-selective Sst5s expressed in CCL39 cells are coupled to Gi/o-protein (Lin et al., 2000; Nunn et al., 2002). Interestingly, gbSS-28 isthe most potent among the three goldfish SSs in suppressing basalGH secretion, but it is less effective than SS-14 in inhibiting GnRH-in-duced GH release in static incubation of dispersed goldfish pituitarycells (Yunker et al., 2003). These data indicate that goldfish pituitary
72 Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81
cells have the ability to differentiate between the different endoge-nous SSs at the receptor and post-receptor levels.
While the above data do not provide insight into the functionalsignificance of the ability of the three endogenous SSs to suppressGH release, they indicate that goldfish pituitary GH release is aphysiological model to examine the differential actions of SS iso-forms on a single cell-type and to study how they interact withother neuroendocrine factors. Elucidating how the different SS iso-forms exert their effects is an important part of understanding themultifactorial neuroendocrine regulation of goldfish GH release. Asin many endocrine cells, elevation of intracellular free Ca2+ levels([Ca2+]i) and activation of protein kinase C (PKC) are important sig-naling pathways in goldfish somatotropes and both mechanismsparticipate in the actions of sGnRH and cGnRH-II on GH release.Both GnRHs are thought to increase extracellular Ca2+ entrythrough voltage-sensitive Ca2+ channels in part by PKC activation(Jobin and Chang, 1992a; Chang et al., 1996). In addition, sGnRHand cGnRH-II mobilize Ca2+ from non-identical suites of pharmaco-logically distinct intracellular Ca2+ stores (Johnson et al., 2000).Previous studies have shown that SS-14 decreases basal, as wellas cGnRH-II- and PKC activator-elicited GH responses withoutinducing parallel reductions in [Ca2+]i, suggesting that SS-14 actsat levels distal to Ca2+ signal generation (Yunker and Chang,2001). Whether [Pro2]SS-14 and gbSS-28 inhibit GH release viamodulation of Ca2+ signaling is unknown.
Given that gbSS-28 (PSS-II) mRNA is expressed in goldfish pitui-tary cells and in the hypophysiotropic hypothalamus, we examinedthe intracellular mechanisms targeted by gbSS-28 in its inhibition ofGH release. We hypothesized that gbSS-28 acts on PKC and Ca2+ sig-naling pathways to elicit its inhibitory action in goldfish pituitarycells in a manner different from that of SS-14. We used a column per-ifusion approach to examine the effects of gbSS-28 on the GH re-sponses to sGnRH, cGnRH-II and PKC activators. We alsomonitored, in parallel, [Ca2+]i changes under basal, GnRH- andPKC-stimulated conditions in single, identified somatotropes loadedwith Fura-2, a ratiometric indicator dye for measuring [Ca2+]i, ineither the absence or presence of gbSS-28 and/or PKC inhibitors. Re-sults demonstrate that gbSS-28 has moderate abilities to reduce ba-sal and PKC-dependent Ca2+ signals, in contrast to the generalinability of SS-14 to attenuate these parameters. Our findings alsosuggest that inhibition of GnRH-induced GH release by gbSS-28 in-volves modulation of PKC-dependent [Ca2+]i changes.
2. Materials and methods
2.1. Experimental animals and cell preparation
Common goldfish (Carassius auratus, 4–5 in. in length) werepurchased from Aquatic Imports (Calgary, Alberta, Canada) andmaintained in 1800-L tanks with flow-through, dechlorinated,charcoal filtered Edmonton municipal water. Fish were kept at18 �C with a simulated photoperiod (times of graded lights onand lights off controlled by an astronomical clock according to sun-rise and sunset of Edmonton, Alberta, Canada) for at least 2 weeksbefore all experiments. Both male and female goldfish were used.Goldfish were killed by cervical transection after deep anaesthesiawith 0.05% tricaine methanesulfonate (Syndel Lab., Vancouver, BC).Pituitaries were then excised and placed in dispersion medium(M199 with Hank’s salts, Gibco, Grand Island, NY, supplementedwith 2.2 g/l NaHCO3, 25 mM HEPES, 100,000 U/l penicillin,100 mg/l streptomycin, and 0.3% BSA, pH adjusted to 7.2 withNaOH). Dispersed pituitary cells were prepared by a trypsin/DNaseprotocol as previously described (Chang et al., 1990). All animalmaintenance and experimental protocols used in the present studyhave been approved by the Biological Sciences Animal Care Com-
mittee, University of Alberta in accordance with nationalguidelines.
2.2. Reagents
Synthetic gbSS-28 was a gift from Dr. J. Rivier of the Salk Insti-tute (La Jolla, CA). Commercial sGnRH ([Trp7, Leu8]GnRH) andcGnRH-II ([His5, Trp7, Tyr8]GnRH) were purchased from PeninsulaLaboratories (Belmont, CA). The PKC activators tetradecanoyl 4b-phorbol-13-acetate (TPA) and dioctanyl glycerol (DiC8), and twoPKC inhibitors, Bisindolylmaleimide II (Bis II) and Gö6976, werepurchased from Sigma Chemical Co (St. Louis, MO). The Ca2+-imag-ing dye Fura-2/AM and its solvent, Pluronic F-127, were purchasedfrom Invitrogen (Carlsbad, CA). Distilled, deionised water was usedto prepare stock solutions of gbSS-28, sGnRH and cGnRH-II. TPA,DiC8, Bis II and Gö6976 were dissolved in dimethyl sulphoxide(DMSO). Aliquots of stock solutions were stored at �20 �C. Finalconcentrations of DMSO were always less than 0.1% and had no ef-fect on basal GH release and [Ca2+]i in identified goldfish somato-tropes. Stock solutions of 10 mM Fura-2/AM was made up freshin DMSO with 20% Pluronic F-127 and sonicated for 10 min priorto use.
2.3. Column perifusion GH release assay
Perifusion experiments were carried out using an Accusyst sys-tem as described in Chang et al. (1990). Briefly, dispersed pituitarycells were cultured overnight with preswollen Cytodex beads(Cytodex I, Sigma) in plating medium (Medium M199 with Earles’salts, Gibco, Grand Island, NY, 26 mM NaHCO3, 25 mM HEPES,100 mg/l streptomycin, 100,000 units/l penicillin and 1% horse ser-um; pH adjusted to 7.2 with NaOH) at 28 �C, under 5% CO2 and sat-urated humidity. Cells on Cytodex beads were then transferred intotemperature controlled columns (1.5 million cells/column, 500 llchamber volume, 18 �C) and perifused with testing medium (Med-ium 199 with Hank’s salts, Gibco, Grand Island, NY, 26 mM NaH-CO3, 25 mM HEPES, 100 mg/l streptomycin, 100,000 units/lpenicillin and 0.1% BSA; pH adjusted to 7.2 with NaOH) at a flowrate of 13 ml/h (time to clear total dead-space �6.5 min). Fractionsof perifusate were collected every 1 min and stored at �26 �C untilGH levels were assayed by a previously validated radioimmunoas-say (RIA, Marchant et al., 1987). Treatments were performed induplicate in each experiment and all experiments were repeateda minimum of three times using different cell preparations.
2.4. Measurements of [Ca2+]i in single, identified somatotropes
Protocols described in (Johnson and Chang, 2000) were fol-lowed. Briefly, dispersed pituitary cells were cultured overnighton poly-L-lysine-coated coverslips (0.4 million cells/coverslip in2 ml plating medium). Cells were then loaded with Fura-2/AM(10 lM) by incubation for 35–40 min in clear testing medium(phenol red was omitted for imaging purposes) at 28 �C under 5%CO2 and saturated humidity. Using a small-volume, closed bathimaging chamber (260 ll bath volume, Series 20 closed bath imag-ing chamber; Warner Instruments, Hamden, CT), the cells wereperifused at a flow rate of 1 ml/min at 18–20 �C. Fura-2 was excitedwith a Hg–Xe arc lamp (Hammamatsu, Japan) at 340 and 380 nmusing a computer controlled filter wheel (Empix Imaging, Mississa-uga, ON). Emission fluorescence at 510 nm was recorded through aZeiss Axiovert 135 inverted microscope (Carl Zeiss Canada, DonMills, ON) and a Paultek Imaging ICCD camera (Grass Valley, CA)linked to a computer. Exposure time and camera gain were care-fully adjusted to optimize the signal-to-noise ratio, while neutraldensity filters (Omega Optical, Brattleboro, VT) were used to re-duce photobleaching. Pairs of images at different excitation
Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81 73
wavelengths were captured every 15 s. F340 and F380 ratios wereconverted to [Ca2+]i using constants determined by an in vitro Ca2+
calibration kit (Molecular Probes, Eugene, OR) according to the for-mula as described in Tsien (1989).
Somatotropes were identified using unique morphological fea-tures under Nomarski Differential Interference Contrast (DIC)microscope as previously described (Van Goor et al., 1994). Briefly,somatotropes possess a large ovoid nucleus containing a centrallylocated nucleolus, numerous granules within the cytoplasm andsmall cytoplasmic extensions (identification accuracy >95% veri-fied by immunostaining with GH anti-serum).
2.5. Data expression and statistics
GH release data from perifusion studies are expressed as per-centages of pretreatment values (average of the values in the firstfive fractions of perifusate collected). The GH responses to treat-ments were quantified by calculating the net response of GH levelsas area under the curve (baseline value subtracted, baseline de-fined as the average value of the five fractions prior to theparticular drug treatment), and the response separated into twotime-dependent phases. Peak responses were determined as thenet response observed within a pulse of stimulator treatment. Pla-teau responses were defined as the net response observed after thepeak to end of recording. Total response was defined as the sum ofthe peak and plateau responses.
Ca2+ traces from individual somatotropes were normalized as apercentage of resting [Ca2+]i (average of the values in the first 10observations; % pretreatment). Data from individual cells aregrouped and kinetic analysis of the Ca2+ traces were performed.A response was deemed to have occurred with an increase in[Ca2+]i of >20% pretreatment (Yunker and Chang, 2001). The totalresponse and duration were calculated as the net change of
0
20
40
60
80
100
*
gbSS-28Basal
Mea
n [C
a2+] i
(% p
retr
eatm
ent)
Bas
al C
alci
um(%
pre
trea
tmen
t)
0 4000
50
100
150
200Basal [
10nM g
Time
g
A
B
Fig. 1. Effects of 10 nM gbSS-28 on resting [Ca2+]i in identified goldfish somatotropes. (AgbSS-28 (horizontal bar, n = 12) from cells prepared from fish at times of gonadal regr(mean ± SEM, average pretreatment [Ca2+]i = 125.6 ± 8.3 nM, n = 18) are presented. Comobservations before and during gbSS-28 treatment. Asterisk indicates a significant differtest, p < 0.05).
[Ca2+]i and the amount of time over the response period. The max-imal amplitude of the Ca2+ responses was the maximal value re-corded during the response period. The time to maximalamplitude was calculated as the amount of time between the onsetof the Ca2+ response and maximal amplitude. Experiments were re-peated with cells from at least four separate cell preparations.
All values are expressed as mean ± SEM. Results were comparedusing either Student’s t-test or analysis of variance (ANOVA) fol-lowed by protected least significant difference (LSD) multiple com-parisons. Differences were considered significant when p < 0.05.
3. Results
3.1. gbSS-28 reduced basal [Ca2+]i in identified somatotropes andsuppressed basal GH release from mixed populations of dispersedgoldfish pituitary cells
In Fura-2-loaded goldfish somatotropes, application of a 5-minpulse of 10 nM gbSS-28, a dose previously shown to be maximallyeffective in suppressing basal GH release in 2-h static incubationstudies (Yunker et al., 2003), significantly decreased basal [Ca2+]i
by approximately 29% and this attenuating effect of gbSS-28 on[Ca2+]i was observed in all the cells tested (n = 12; Fig. 1). Account-ing for dead-space and flow rate of the system, the [Ca2+]i decreaseoccurred within 15 s of the gbSS-28 treatment reaching the cells(i.e., almost instantaneously and within the first observation periodfollowing exposure of cell to gbSS-28). Maximal level of suppres-sion of about 32% was observed by 2–2.5 min of exposure afterwhich an average reduction of �25–30% was maintained. The re-sponse duration lasted 306 ± 11 s, indicating that [Ca2+]i values re-turned almost immediately to basal levels following thewithdrawal of gbSS-28 treatment (Fig. 1); however, a rebound in-crease in [Ca2+]i characteristic of the termination of SS-14 actions
0
5
10
15
20
*
gbSS-28Basal
CV
of
[Ca2+
] i(%)
800 1200
Ca2+]i(n=6)
bSS-28 (n=12)
(S)
bSS-28
C
) Pooled [Ca2+]i traces in the absence (basal, n = 6) and presence of a 5-min pulse ofession (May to June). Results are expressed as a percentage of pretreatment valueparisons of (B) mean [Ca2+]i values and (C) coefficient of variation (CV) of [Ca2+]i
ence between parameter observed before and during gbSS-28 treatment (paired t-
0 500 1000 1500 2000
60
80
100
120
140 Basal
10nM gbSS-28
gbSS-28GH
(%
pre
trea
tmen
t)
Time(s)
Fig. 2. Effects of 10 nM gbSS-28 on resting GH secretion from goldfish pituitarycells. Pooled GH release traces in the absence (basal, n = 8) and presence of a 5-minpulse of gbSS-28 (horizontal bar, n = 10) from cells prepared from fish at late stagesof gonadal maturation (March to April) are presented. Results are expressed as apercentage of pretreatment value (mean ± SEM, average pretreatment GH values forcontrols and gbSS-28-treated groups = 45.2 ± 7.7 and 36.3 ± 5.8 ng/ml,respectively).
0
100
200
300 100nM cGnRH-II
100nM cGnRH-II+10nM gbSS-28
cGnRH-II+gbSS-28cGnRH-II
[Ca2+
] i (
% p
retr
eatm
ent)
0 200 400 600 800 1000 1200 1400
0 200 400 600 800 1000 1200 14000
100
200
300 100nM sGnRH
100nM sGnRH+10nM gbSS-28
sGnRH+gbSS-28sGnRH[C
a2+] i (
% p
retr
eatm
ent)
Time(s)
Time(s)
A
B
Fig. 3. Effects of 10 nM gbSS-28 on the [Ca2+]i responses to 100 nM sGnRH (A) andcGnRH-II. (B) 2-min pulses of GnRH (open horizontal bar) were applied either alone(n = 5 and 6 for sGnRH and cGnRH-II, respectively) or 2 min into a 5-min applicationof gbSS-28 (hatched bar; n = 12 and 12, respectively, for combination treatmentwith either sGnRH or cGnRH-II). Results are expressed as a percentage ofpretreatment value (mean ± SEM, pretreatment [Ca2+]i = 100.9 ± 0.5 and101.5 ± 0.9 nM for experiments with sGnRH and cGnRH-II, respectively) and pooled[Ca2+]i traces are presented. Pituitary cells were prepared from fish at times ofgonadal regression (June to July).
74 Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81
in mammals was not observed (McFarlane et al., 1993). Accordingto previously established criteria (Yunker and Chang, 2001), morethan 90% of the cells recorded can be judged as quiet cells havingrelatively stable basal [Ca2+]i (average coefficient of variation, CV,of fluctuation in [Ca2+]i < 20%). Interestingly, gbSS-28 not only re-duced the mean basal [Ca2+]i, but also turned cells into even ‘‘qui-eter” somatotropes (CV of spontaneous oscillations in [Ca2+]i
following gbSS-28 treatment was �42% of that prior to gbSS-28application, p < 0.05, Fig. 1C).
Whether the acute inhibitory actions of gbSS-28 on basal [Ca2+]i
was reflected in rapid suppression of basal GH release was exam-ined in parallel in a column perfusion system with fast (1 min)fraction collections (Fig. 2). A 5-min application of 10 nM gbSS-28 rapidly suppressed basal GH secretion. Reduction in secretionwas observed starting generally within 1 min following the esti-mated arrival of the gbSS-28 pulse (i.e., within the time resolutionof the system) and a maximal suppression of about 25% wasreached by 3 min into gbSS-28 exposure. This magnitude of maxi-mal suppression was in a similar range as that observed with[Ca2+]i measurement (about 28%). This significant reduction ofGH release was maintained for 781 ± 48 s, three times longer thanduration of gbSS-28 application. A transient rebound increase inGH levels characteristic upon termination of SS-14 treatment inmammals (Fig. 2; McFarlane et al., 1993) was not observed.
3.2. gbSS-28 inhibited GnRH-stimulated [Ca2+]i increases and GHrelease
In goldfish somatotropes, application of sGnRH and cGnRH-IIelicited increases in [Ca2+]i and GH release (Johnson and Chang,2000). In the present study, we tested whether gbSS-28 could af-fect GnRH-evoked Ca2+ signals and GH release in a manner similarto what was seen in the basal conditions. GnRH was used at a doseof 100 nM, a concentration that has previously been shown to bemaximally effective in stimulating GH secretion (Chang et al.,1994).
As shown in Fig. 3A, treatment of single, identified somato-tropes with sGnRH (100 nM) for 2 min induced acute elevationsin [Ca2+]i that lasted for approximately 5 min and with an averagemaximal increase of about 2.2-fold above basal. Addition of 10 nMgbSS-28 significantly reduced the magnitude of the total sGnRH-induced [Ca2+]i response (Table 1). Similarly, a 2-min challenge of100 nM cGnRH-II induced an increase in [Ca2+]i (Fig. 3B). This re-sponse had a similar maximal magnitude as that with sGnRH but
the kinetics was different. Relative to sGnRH, the cGnRH-II-elicitedCa2+ response had a delayed peak response but the duration of theresponse was roughly doubled (Table 1). gbSS-28 pretreatment sig-nificantly reduced the total response induced by cGnRH-II, as wellas the maximal amplitude, time to maximal response and durationof response (Table 1).
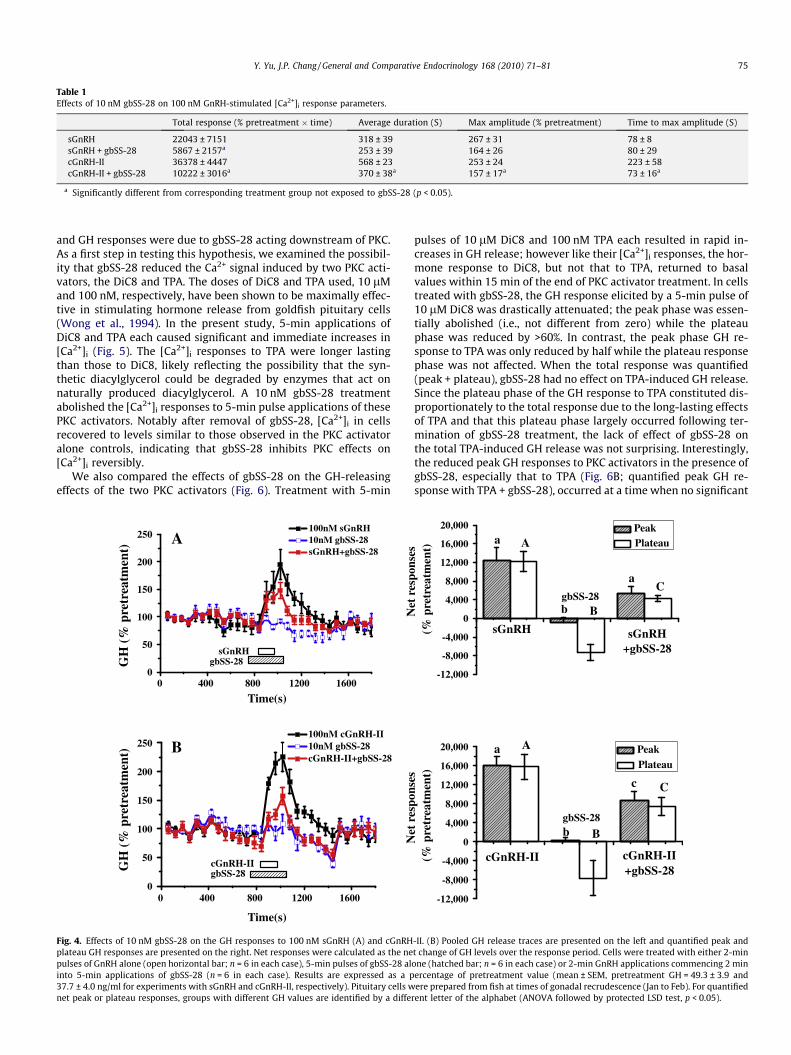
Next, we examined the effects of gbSS-28 in inhibiting the GnRH-induced GH responses. The 2-min pulses of 100 nM sGnRH andcGnRH-II each induced a rapid increase in GH secretion and withthe maximal elevation in GH release observed within 3 min of theestimated exposure of dispersed goldfish pituitary cells to the GnRHs(Fig. 4). Pretreatment with 10 nM gbSS-28 significantly reduced theGH responses to sGnRH and cGnRH-II. Interestingly, significantreduction was only observed in the plateau phase of the sGnRH-in-duced GH response whereas both the peak and plateau phases ofthe cGnRH-II-elicited GH increase were significantly attenuated(Fig. 4). The greater effectiveness of gbSS-28 to inhibit cGnRH-II- thansGnRH-induced GH secretion was similar to data obtained from[Ca2+]i imaging, suggesting that gbSS-28 inhibited GnRH-stimulatedGH secretion by interfering with Ca2+-dependent mechanisms.
3.3. gbSS-28 abolished PKC-mediated elevations in [Ca2+]i
and increases in GH release
In goldfish, PKC in part mediates GnRH-induced hormone re-lease. GnRH activation of PKC is thought to cause an increase inCa2+ entry through VSCC (Wong et al., 1994). Thus, we hypothe-sized that the ability of gbSS-28 to reduce GnRH-induced [Ca2+]i
Table 1Effects of 10 nM gbSS-28 on 100 nM GnRH-stimulated [Ca2+]i response parameters.
Total response (% pretreatment � time) Average duration (S) Max amplitude (% pretreatment) Time to max amplitude (S)
a Significantly different from corresponding treatment group not exposed to gbSS-28 (p < 0.05).
Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81 75
and GH responses were due to gbSS-28 acting downstream of PKC.As a first step in testing this hypothesis, we examined the possibil-ity that gbSS-28 reduced the Ca2+ signal induced by two PKC acti-vators, the DiC8 and TPA. The doses of DiC8 and TPA used, 10 lMand 100 nM, respectively, have been shown to be maximally effec-tive in stimulating hormone release from goldfish pituitary cells(Wong et al., 1994). In the present study, 5-min applications ofDiC8 and TPA each caused significant and immediate increases in[Ca2+]i (Fig. 5). The [Ca2+]i responses to TPA were longer lastingthan those to DiC8, likely reflecting the possibility that the syn-thetic diacylglycerol could be degraded by enzymes that act onnaturally produced diacylglycerol. A 10 nM gbSS-28 treatmentabolished the [Ca2+]i responses to 5-min pulse applications of thesePKC activators. Notably after removal of gbSS-28, [Ca2+]i in cellsrecovered to levels similar to those observed in the PKC activatoralone controls, indicating that gbSS-28 inhibits PKC effects on[Ca2+]i reversibly.
We also compared the effects of gbSS-28 on the GH-releasingeffects of the two PKC activators (Fig. 6). Treatment with 5-min
0
50
100
150
200
250100nM sGnRH10nM gbSS-28sGnRH+gbSS-28
gbSS-28sGnRH
GH
(%
pre
trea
tmen
t)
0 400 800 1200 1600
0 400 800 1200 16000
50
100
150
200
250100nM cGnRH-II10nM gbSS-28cGnRH-II+gbSS-28
gbSS-28cGnRH-IIG
H (
% p
retr
eatm
ent)
Time(s)
Time(s)
A
B
Fig. 4. Effects of 10 nM gbSS-28 on the GH responses to 100 nM sGnRH (A) and cGnRHplateau GH responses are presented on the right. Net responses were calculated as the nepulses of GnRH alone (open horizontal bar; n = 6 in each case), 5-min pulses of gbSS-28 alinto 5-min applications of gbSS-28 (n = 6 in each case). Results are expressed as a p37.7 ± 4.0 ng/ml for experiments with sGnRH and cGnRH-II, respectively). Pituitary cells wnet peak or plateau responses, groups with different GH values are identified by a diffe
pulses of 10 lM DiC8 and 100 nM TPA each resulted in rapid in-creases in GH release; however like their [Ca2+]i responses, the hor-mone response to DiC8, but not that to TPA, returned to basalvalues within 15 min of the end of PKC activator treatment. In cellstreated with gbSS-28, the GH response elicited by a 5-min pulse of10 lM DiC8 was drastically attenuated; the peak phase was essen-tially abolished (i.e., not different from zero) while the plateauphase was reduced by >60%. In contrast, the peak phase GH re-sponse to TPA was only reduced by half while the plateau responsephase was not affected. When the total response was quantified(peak + plateau), gbSS-28 had no effect on TPA-induced GH release.Since the plateau phase of the GH response to TPA constituted dis-proportionately to the total response due to the long-lasting effectsof TPA and that this plateau phase largely occurred following ter-mination of gbSS-28 treatment, the lack of effect of gbSS-28 onthe total TPA-induced GH release was not surprising. Interestingly,the reduced peak GH responses to PKC activators in the presence ofgbSS-28, especially that to TPA (Fig. 6B; quantified peak GH re-sponse with TPA + gbSS-28), occurred at a time when no significant
-12,000
-8,000
-4,000
0
4,000
8,000
12,000
16,000
20,000
C
B
A
c
bgbSS-28
cGnRH-II+gbSS-28
PeakPlateau
Net
res
pons
es (
% p
retr
eatm
ent)
cGnRH-II
a
-12,000
-8,000
-4,000
0
4,000
8,000
12,000
16,000
20,000
C
B
A
a
bgbSS-28
sGnRH+gbSS-28
PeakPlateau
Net
res
pons
es (
% p
retr
eatm
ent)
sGnRH
a
-II. (B) Pooled GH release traces are presented on the left and quantified peak andt change of GH levels over the response period. Cells were treated with either 2-minone (hatched bar; n = 6 in each case) or 2-min GnRH applications commencing 2 minercentage of pretreatment value (mean ± SEM, pretreatment GH = 49.3 ± 3.9 andere prepared from fish at times of gonadal recrudescence (Jan to Feb). For quantified
rent letter of the alphabet (ANOVA followed by protected LSD test, p < 0.05).
0
100
200
300
400 100nM TPA100nM TPA+10nM gbSS-28
gbSS-28TPA[C
a2+] i (
% p
retr
eatm
ent)
0 500 1000 1500
0 500 1000 1500
0
100
200
300
400
DiC8gbSS-28
10µM DiC810µM DiC8+10nM gbSS-28
[Ca2+
] i (%
pre
trea
tmen
t)
Time(s)
Time(s)
A
B
Fig. 5. Effects of 10 nM gbSS-28 on the the [Ca2+]i responses to 2 PKC activators,10 mM DiC8 (A) and 100 nM TPA (B). Five-min pulses of PKC activator (openhorizontal bar) were applied either alone (n = 6 and 5 for DiC8 and TPA,respectively) or 2 min into a 8-min application of gbSS-28 (hatched bar; n = 12and 12, respectively for combination treatment with either DiC8 or TPA). Results areexpressed as a percentage of pretreatment value (mean ± SEM, pretreatment[Ca2+]i = 117.6 ± 4.9 and 102.3 ± 6.6 nM for experiments with DiC8 and TPA,respectively) and pooled [Ca2+]i traces are presented. Pituitary cells were preparedfrom fish at times of gonadal recrudescence (December to January).
76 Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81
PKC-induced increases in [Ca2+]i was observed in Fig. 5A (abolish-ment of [Ca2+]i response during the presence of gbSS-28). Whetherthis observation implied that a Ca2+-independent PKC action of GHrelease was present could not be determined since [Ca2+]i changeswere measured across the whole focal plane of the cell and smallregional increases in [Ca2+]i remaining might not be detected.None-the-less, the GH results, coupled with those from [Ca2+]i
studies, revealed that gbSS-28 was a more effective inhibitoragainst DiC8 than TPA in terms of GH release and the relative effi-cacy of gbSS-28 was associated with its ability to modulate theCa2+ signal induced by these PKC activators.
3.4. The effects of TPA and DiC8 on [Ca2+]i are expressed via activationof PKC
In the literature, it is known that phorbol esters can affect Ca2+-independent PKC activation (McFerran and Guild, 1994) and thatdiacylglycerol may also have PKC-independent activities (Chakrab-arti and Kumar, 2000). Thus, we needed to confirm that DiC8 andTPA were indeed acting through PKC to induce increases in[Ca2+]i in our study system. To address this question, two PKCinhibitors, Bis II and Gö6976, were used. Bis II, the broader spec-trum PKC inhibitor, was used at 100 nM, a concentration that is10 times that of the Ki (10 nM) reported for PKC (Toullec et al.,1991). The PKCa/bI isoenzyme-selective inhibitor Gö6976 wastested at 100 nM, a concentration that is about 20-fold of the Kifor blocking PKCa (2.3 nM) and PKCb1 (6.2 nM) isozymes (Marti-
ny-Baron et al., 1993). These two inhibitors were chosen becausegoldfish somatotropes have been shown to contain PKCa and thatgoldfish pituitary extracts also contain PKCd and PKCh (Klausenet al., 2005). PKCa belongs to the subfamily of Ca2+- and diacylglyc-erol-regulated (conventional) PKCs, whereas PKCd and PKCh areCa2+-independent isoforms and are activated by diacylglyceroland phosphatidylserine (novel PKCs; (Steinberg, 2008)). As shownin Fig. 7A and C, pretreatment of cells with Bis II or Gö6976 com-mencing 1 h prior to the beginning of recording abolished the[Ca2+]i response induced by DiC8. There was no difference in the ef-fects of the two PKC inhibitors, suggesting that involvement of con-ventional PKC could totally account for DiC8 actions in goldfishsomatotropes. TPA-stimulated [Ca2+]i increase was significantlyattenuated in the presence of either 100 nM Bis II or Gö6976(Fig. 7B and D), and a higher concentration of Bis II (500 nM) com-pletely blocked the TPA effect (data not shown). These results sug-gest that TPA effects on goldfish somatotropes are also mediatedthrough PKC activation.
3.5. gbSS-28 inhibits resting and GnRH-triggered [Ca2+]i signalsin the presence of PKC inhibitors
Based on the fact that GnRH stimulation of GH release requiresPKC and that PKC activators can lead to increases in [Ca2+]i just likeGnRH (Wong et al., 1994; Yunker and Chang, 2001), it is logical toassume that GnRH-induced Ca2+ signaling involves PKC; however,this requires direct testing. Validating that GnRH-induced [Ca2+]i
increases involved PKC is crucial in evaluating whether gbSS-28may attenuate GnRH-induced increase in [Ca2+]i by targetingPKC. To test the involvement of PKC in GnRH-induced [Ca2+]i re-sponses, we examined the ability of Bis II and Gö6976 to affectGnRH-elicited increase in [Ca2+]i in goldfish somatotropes inCa2+-imaging studies. When cells were pretreated for 1 h with100 nM Bis II or Gö6976, a protocol which was able to abolishthe [Ca2+]i increases in response to DiC8 (Fig. 7), [Ca2+]i increasesinduced by 2-min pulses of sGnRH and cGnRH-II were reducedby 35–55% (Fig. 8A,B,F and G). Although Bis II and Gö6976 had sim-ilar effects on sGnRH-induced [Ca2+]i response, the peak and pla-teau phases of the response to cGnRH-II were selectivelymodulated by Gö6976 and Bis II, respectively. These latter observa-tions were not at variance with the idea that different PKC isoformsmediate the rapid and sustained effects of cGnRH-II. Regardless,these results demonstrate that GnRH-elicited increases in [Ca2+]i
were partially mediated by PKC.To evaluate the extent to which inhibition of PKC-dependent in-
crease in [Ca2+]i contributes to the total ability of gbSS-28 to reducethe [Ca2+]i response under basal and/or GnRH-stimulated condi-tions, we tested the effects of combined 10 nM gbSS-28 and100 nM Bis II treatment. In cells pretreated with Bis II, applicationof gbSS-28 caused an immediate further reduction in [Ca2+]i, sug-gesting that reduction in PKC activity did not totally account forthe inhibitory action of gbSS-28 on basal [Ca2+]i level (Fig. 8 Eand J). The magnitudes of the peak [Ca2+]i responses to the twoGnRHs in the presence of both Bis II and gbSS-28 were generallylower than those in the presence of either Bis II or gbSS-28 alonebut the differences were not significant (Fig. 8E and J).
Since we hypothesize that gbSS-28 inhibition of GnRH action ingoldfish somatotrope involved attenuation of PKC-dependent Ca2+
entry, we also examined the effects of treatment with nominallyCa2+-free media (testing media prepared without the addition ofCa2+ salts) on GnRH-induced [Ca2+]i responses. Exposure to nomi-nally Ca2+-free media caused a rapid decrease in resting [Ca2+]i
and impaired the ability of sGnRH and cGnRH-II to elicit [Ca2+]i in-creases (Fig. 8D and I). The effect of Ca2+-free media and gbSS-28on [Ca2+]i are similar (average [Ca2+]i values: 78.1 ± 3.5% pretreat-ment for Ca2+-free media, Fig. 8; 71.8 ± 3.4% pretreatment for
-200
0
200
400
600
800
1000
1200
1400
1600
gbSS-28TPA
100nM TPA10nM gbSS-28100nM TPA+10nM gbSS-28
GH
(%
pre
trea
tmen
t)
0 1000 2000 3000 4000 5000 6000
0 500 1000 1500 2000 2500 3000 350040
60
80
100
120
140
160
180
200
220
gbSS-28DiC8
10µM DiC8 10nM gbSS-28 10µM DiC8+10nM gbSS-28
GH
(%
pre
trea
tmen
t)
Time (s)-20,000
0
20,000
40,000
60,000
C
B
A
cb
gbSS-28
Dic8
+gbSS-28
Net
res
pons
es
(%
pre
trea
tmen
t)
Dic8
a
Time (s)
0
2 x 105
4 x 105
6 x 105
2.4 x 106
2.6 x 106
2.2 x 106
a
A
bB c
C
Net
res
pons
es
(% p
retr
eatm
ent)
TPA gbSS-28 TPA+gbSS-28
PeakPlateau
PeakPlateauA
B
Fig. 6. Effects of 10 nM gbSS-28 on the GH responses to two PKC activators, 10 mM DiC8 (A) and 100 nM TPA. (B) Pooled GH release traces are presented on the left andquantified peak and plateau GH responses are presented on the right. Net responses were calculated as the net change of GH levels over the response period. Cells weretreated with either 5-min pulses of PKC activator alone (open horizontal bar, n = 8 and 6 for DiC8 and TPA, respectively), 8-min pulses of gbSS-28 alone (n = 8 and 6 for DiC8and TPA experiments, respectively) or 5-min pulses of PKC activators commencing 2 min into gbSS-28 application (n = 8 and 6, respectively for combination treatment witheither DiC8 or TPA). Results are expressed as a percentage of pretreatment value (mean ± SEM, pretreatment GH = 49.3 ± 4.8 and 46.4 ± 1.7 ng/ml for experiments with DiC8and TPA, respectively). Pituitary cells were prepared from fish at times of gonadal recrudescence (Jan to Feb). For quantified net peak or plateau responses, groups withdifferent GH values are identified by a different letter of the alphabet (ANOVA followed by protected LSD test, p < 0.05).
Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81 77
gbSS-28, Fig. 1), and gbSS-28 did not further reduce [Ca2+]i whencells were already exposed to Ca2+-free media (average [Ca2+]i val-ues: 72.9 ± 1.2% pretreatment, n = 7; p > 0.05 vs. Ca2+-free mediaalone or gbSS-28 alone). Interestingly, the combined inhibitory ef-fects of gbSS-28 and Bis II on sGnRH-induced [Ca2+]i response canbe mimicked by incubation with nominally Ca2+-free media(Fig. 8D), indicating that the residual Ca2+ in both instances werelikely due to mobilization of Ca2+ from intracellular sources. In con-trast, cGnRH-II failed to elicit a significant increase in [Ca2+]i in thepresent study when cells were incubated with nominally Ca2+-freemedia while a small cGnRH-II-induced [Ca2+]i remained in thepresence of both gbSS-28 and Bis II (Fig. 8I and J).
4. Discussions
Results from this study provide insight into signal transductionelement(s) targeted by gbSS-28 to effect its inhibition on basal andGnRH-stimulated GH release. Whereas the reduction of basal[Ca2+]i by gbSS-28 is largely PKC-independent, inhibition of GnRHsignaling by gbSS-28 affects [Ca2+]i through a pathway that de-pends on PKC. Thus, results support the hypothesis that gbSS-28inhibitory mechanisms are different from those of SS-14.
4.1. Mechanisms of gbSS-28 inhibition of basal GH release
Previous hormone release results have shown that basal GH re-lease is highly sensitive to the manipulations of extracellular Ca2+
availability and inhibition of voltage-sensitive Ca2+ channel func-
tions (Jobin and Chang, 1992b). Furthermore in this cell system,general inhibition of intracellular Ca2+ release channels is notaccompanied by acute decreases in [Ca2+]i and GH release (Johnsonand Chang, 2000). In the present study, we confirmed that incuba-tion of goldfish somatotropes with nominally Ca2+-free media de-creased [Ca2+]i and further demonstrated that gbSS-28 inhibitionof basal GH release is accompanied by decreases in [Ca2+]i in soma-totropes. Furthermore, gbSS-28 failed to suppress [Ca2+]i whenthese [Ca2+]i levels were already reduced by treatment with Ca2+-free media. Taken together, the available information suggests thatattenuation of extracellular Ca2+ entry into somatotropes is amechanism by which gbSS-28 reduces basal GH release. Interest-ingly, although the somatotropes examined in the present studyfits into previous criteria defining quiet cells in terms of [Ca2+]i
fluctuations under resting conditions (Yunker and Chang, 2001),gbSS-28 further reduced the variability in [Ca2+]i. In normal mam-malian somatotropes, the [Ca2+]i oscillations have been proposedto be the result of Ca2+ influx into the cell, possibly as the resultof spontaneous electrical activity of the membrane (Kwiecienet al., 1997). This observation lends further support to the idea thatmanipulation of [Ca2+]i from an extracellular source is a criticalcomponent of gbSS-28 inhibition of basal GH release in these cells.
Although the effects of gbSS-28 on basal [Ca2+]i and GH releaseare both rapid and reversible, [Ca2+]i returns to basal values verysoon after termination of gbSS-28 treatment while the suppressionof GH release lasted for a longer time, with the duration of suppres-sion of GH release approximately doubled that of the reduction in[Ca2+]i. A delay in recovery of GH release following the return of
50
100
150
200
250
300
350 100nM G o 6976 Without G o 6976
DiC8
100nM G o 6976 Without G o 6976
[Ca2+
] i (%
pre
trea
tmen
t)
TPA
050
010
0015
00
50
100
150
200
250
300
350 100nM Bis II
Without Bis II
DiC8
100nM Bis II
Without Bis II
[Ca2+
] i (%
pre
trea
tmen
t)
050
010
0015
00
050
010
0015
00 050
010
0015
00
TPA
A B
C D
Time (s)
Time (s)
Fig. 7. Effects of 100 nM PKC inhibitors Bis II (A and B) and Gö6976 (C and D) on the [Ca2+]i responses in identified goldfish somatotropes to 8-min treatments (hatched bar) ofeither 10 mM DiC8 (A and C) or 100 nM TPA (B and D). The PKC inhibitors were applied commencing 1 h prior to beginning of experiments and continued to the end of datacollection. Pooled data are presented (mean ± SEM; n = 6, 9, 10, 5, 9 and 10 for DiC8, DiC8 + Bis II, DiC8 + Gö6976, TPA, TPA + Bis II and TPA + Gö6976, respectively).Pretreatment [Ca2+]i values for cells exposed to Bis II and Gö6976 are 114.5 ± 11.9 nM and 136.1 ± 21.2 nM, respectively and 115.2 ± 6.6 nM for cells not pretreated with PKCinhibitors. Cells used were prepared from fish at times of gonadal regression (August to September).
78 Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81
[Ca2+]i to normal levels is not totally unexpected because of the fol-lowing two reasons. First, the total dead-volume between the Ca2+-imaging and GH release experiments are different, and thus theirwashout kinetics are possibly different. Second, the prolonged ef-fects of gbSS-28 on GH release relative to the [Ca2+]i changesmay also be a reflection of the fact that Ca2+ is only one of manysignal transduction components involved in the regulation of exo-cytosis. This time delay may be explained by additional primingsteps required by vesicles to enter the readily releasable pool andbecome release competent (Morgan et al., 2005).
How the hypothesized gbSS-28-effected reduction of extracel-lular Ca2+ entry is manifested and whether gbSS-28 also targetsother intracellular components leading to suppression of basalGH secretion are unclear, but PKC is unlikely to be a target forgbSS-28 action in this regard. PKC would have been a candidatebased on a number of previous and current findings, includingthe following: (1) PKC activation is known to inhibit K+ channelsand thus enhance membrane depolarization and activation ofCa2+ entry through voltage-sensitive Ca2+ channels in rat GH4C1cell (Shipston and Armstrong, 1996); (2) PKC activators elevates[Ca2+]i in goldfish somatotropes (Yunker and Chang, 2001 andFig. 5); and (3) gbSS-28 inhibits the [Ca2+]i response to two PKCactivators (Fig. 5). However, applications of PKC inhibitors neitheraffected basal [Ca2+]i in goldfish somatotropes in the present study(basal [Ca2+]i in controls, Gö6976- and Bis II-treated cells were115.2 ± 6.6, 114.5 ± 11.9 and 136.1 ± 21.2 nM, respectively) norsuppressed basal GH secretion in previous experiments (Wonget al., 1994). Furthermore, in the presence of Bis II, gbSS-28 further
reduced basal [Ca2+]i. On the other hand, SS-14 has been shown toincrease K+ conductance leading to hyperpolarization of the cellmembrane and inhibition of voltage-sensitive Ca2+ influx in severalsystems including mammalian pituitary and pancreatic beta cells(Gromada et al., 2001; Tsaneva-Atanasova et al., 2007). Direct eval-uation of gbSS-28 effects on membrane ion channels in goldfishsomatotropes will be an important future experimental directionin understanding the mechanisms of gbSS-28 reduction of [Ca2+]i,which seems to be closely tied to its ability to inhibit basal GH re-lease from these cells.
4.2. Mechanisms of gbSS-28 inhibition of sGnRH- and cGnRH-II-induced GH release
It has been shown that GnRH-stimulated GH release in goldfish ismediated by PKC activation and mobilization of Ca2+ from bothextracellular and intracellular sources (Chang et al., 1994; Wonget al., 1994). Results in the present study demonstrate directly, forthe first time, that sGnRH and cGnRH-II actions on [Ca2+]i increasesin goldfish somatotropes are mediated in part through PKC and thatgbSS-28 attenuates the [Ca2+]i and GH responses to both GnRHs.gbSS-28 differentially affects the peak and plateau GH release phasesof sGnRH relative to those of cGnRH-II and these effects mirror therelative efficacy of gbSS-28 to attenuate the peak and plateau[Ca2+]i responses to these two GnRHs. These results lend further cre-dence to the idea that gbSS-28 inhibition of GnRH-induced GH re-lease is dependent on its ability to affect GnRH-induced [Ca2+]i
increase. In addition, the co-application of Bis II and gbSS-28 fails
050
010
00
60
80
100
120
140
160
180
200
220
sG
[Ca2+
] i (%
pre
trea
tmen
t)
050
010
00
Med.
Med.
With Bis II
sG
050
010
00
With G ö 6976
sG
050
010
00
0Ca 2+
sG
050
010
00
SS28sG
With Bis II
A B C D E
F G H I J
Time(s)
Time(s)
050
010
0015
0050
75
100
125
150
175
200
cG
[Ca2+
] i (%
pre
trea
tmen
t)
050
010
00
With Bis II
cG
050
010
00
Med.
Med.With
G ö6976
cG
050
010
0015
00
0Ca 2+cG
050
010
0015
00
cGSS28
With Bis-II
sG
SS28+sG
Bis II+
sG
Go697
6+sG
0Ca
+sG
SS28+Bis
II+sG
0
40
80
120
160
200
bb
b a,bb
Net
max
imal
am
plit
udes
a
..
2+
cG
SS28+cG
BisII+
cG
Go697
6+cG
0Ca
+cG
SS28+Bis
II+cG
0
40
80
120
160
200
b
b
ba,ba,b
Net
max
imal
am
plit
udes a
..
2+
Fig. 8. [Ca2+]i responses to 2-min pulses (open bar) of 100 nM sGnRH (sG) (A–E) and 100 nM cGnRH-II (cG) (F–J) either alone (in media, med; A and F) and in the presence ofeither 100 nM Bis II (B and G), 100 nM Gö6976 (C and H), norminally Ca2+-free media (0 Ca2+) (D and I), or 10 nM gbSS-28 in the presence of 100 nM Bis II (E and J). PKCinhibitors were applied commencing 1 h prior to the beginning of experiments and continued to the end of data collection. Average [Ca2+]i traces (n = 4–6 in each case) frompooled results expressed as a percentage of pretreatment value (pretreatment = 104.4 ± 11.9 and 125.8 ± 7.0 nM for sGnRH and cGnRH-II experiments, respectively;mean ± SEM) are presented in the left panels, standard error bars are excluded for the sake of clarity. Duration of exposure to nominally Ca2+-free media or gbSRIF-28 areidentified by the hatched bar. Quantified results of the net maximal amplitudes (where the basal levels of [Ca2+]i were subtracted) in [Ca2+]i in response to GnRH are presentedin the right panel and the corresponding effects of gbSS-28 on GnRH obtained from experiments presented in Figure 4 are included for comparison. Groups with different netmaximal amplitudes of [Ca2+]i increases are identified by a different letter of the alphabet (ANOVA followed by protected LSD test, p < 0.05). Cells used were prepared fromfish at times of gonadal regression (June to July).
Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81 79
to produce a larger inhibitory effect than that of gbSS-28 alone. Theseobservations, when viewed together with the ability of gbSS-28 toinhibit the PKC-dependent increase in [Ca2+]i, further suggest thatgbSS-28 inhibits GnRH-stimulated GH release in part by blockingPKC-dependent [Ca2+]i increase. On the other hand, the inability ofthe two PKC inhibitors to totally inhibit GnRH effects on [Ca2+]i whenused at concentrations that virtually abolished DiC8-induced [Ca2+]i
increases indicates that GnRH-activated Ca2+ signaling also has aPKC-independent component. Whether gbSS-28 inhibition ofGnRH-induced rise in [Ca2+]i also acts though this PKC-independentpathway is unresolved, although in some cases, gbSS-28 effects onthe magnitude of peak [Ca2+]i in response to cGnRH-II are more pro-nounced than those of the PKC inhibitors, with PKC inhibitors reduc-ing the cGnRH-II-induced response by 30–40% and gbSS-28attenuating the cGnRH-II effects by �60%. The identity of thesePKC-independent Ca2+ mobilization mechanism(s), if present, re-mains to be determined in future studies.
Previous studies indicate that PKC activators do not affect mobi-lization of Ca2+ from intracellular stores in goldfish somatotropes(Johnson and Chang, 2000). Whether gbSS-28 affects GnRH-inducedmobilization of Ca2+ from intracellular sources cannot be addressedbased on results in the present study. However, the similaritybetween the sGnRH-induced Ca2+ signal remaining following pre-
exposure to nominally Ca2+-free media and that observed in thecombined presence of gbSS-28 and Bis II suggest that the residualCa2+ signal following this combined treatment is derived from intra-cellular sources. In goldfish somatotropes, sGnRH is known to mobi-lize Ca2+ from intracellular stores that are sensitive to IP3 andcaffeine, but not ryanodine (Johnson et al., 2000). Thus, gbSS-28 ap-pears not to abolish the ability of sGnRH to mobilize Ca2+ from one ormore of these stores. On the other hand, cGnRH-II is known to utilizeryanodine-sensitive and caffeine-sensitive Ca2+ stores (Johnsonet al., 2000), and the residual cGnRH-II induced Ca2+ signal observedin the presence of gbSS-28 and Bis II is likely to be from one or both ofthese sources. Surprisingly, unlike the situation with sGnRH, cGnRH-II failed to elicit a significant Ca2+ response in somatotropes pre-treated with nominally Ca2+-free media. This observation indicatesthat the intracellular Ca2+ stores utilized by cGnRH-II and sGnRHmay have different sensitivities to and/or requirements for extracel-lular Ca2+ in their refilling.
4.3. Differences in the actions of gbSS-28 and SS-14 in goldfishsomatotropes
Results from the present study indicate that gbSS-28 and SS-14mechanisms of action on goldfish somatotropes are not identical.
Fig. 9. Working model of gbSS-28 inhibition of basal and GnRH-stimulated GHrelease in goldfish. gbSS-28 attenuates basal GH release by decreasing [Ca2+]i
manifested in part as a consequent of inhibitory actions on non-PKC-sensitivemembrane Ca2+ channels to extracellular Ca2+ entry (solid blunt-ended arrow).Attenuation of GnRH-induced GH release is also effected through reduction ofGnRH-elicited increases in [Ca2+]i. In this case, gbSS-28 action includes inhibition ofPKC-mediated extracellular Ca2+ entry (open blunt-ended arrow). Whether PKC-insensitive but GnRH-sensitive intracellular Ca2+ release channels are targeted bygbSS-28 is unknown. If this occurs, IP3- and ryanodine-receptor Ca2+ channels maybe candidate targets regulated by gbSS-28 (stippled blunt-ended arrow). Incontrast, attenuation of Ca2+ signals is not a primary mechanism through whichSS-14 lowers basal, or abolish GnRH-stimulated hormone release. Instead, previousresults indicate that SS-14 acts downstream of Ca2+ mobilization to reduce GHsecretion (dotted blunt-ended arrow).
80 Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81
This study shows that the inhibitory effects of gbSS-28 on basal, aswell as GnRH- and PKC-stimulated, GH release are strongly associ-ated with the ability of gbSS-28 to affect [Ca2+]i. In contrast,whereas SS-14 also reduced basal GH release from populations ofgoldfish pituitary cells, parallel reductions in [Ca2+]i are not ob-served within single, identified somatotropes (Yunker and Chang,2001). Similarly, SS-14 abolishes the GH responses to the two na-tive GnRHs and the PKC activators DiC8 and TPA but these effectsare not accompanied by pronounced reductions in [Ca2+]i signals.In addition, the [Ca2+]i response induced by DiC8 is larger in thepresence than in the absence of SS-14 (Yunker and Chang, 2001).gbSS-28 is also less effective than SS-14 in inhibiting GH responsesto Ca2+ ionophores (Yunker et al., 2003). These data, when takentogether, suggest that, unlike gbSS-28, SS-14 action is mainly tar-geting sites distal to [Ca2+]i elevations along the GnRH/PKC signal-ing cascade in goldfish somatotropes to affect GH secretion. Inaddition, when tested at concentrations that maximally suppressbasal GH secretion, SS-14 abolishes the GH responses to the twoendogenous GnRHs (Yunker et al., 2003) while gbSS-28 is only par-tially effective in this regard (Fig. 4).
The reasons behind the different effects of these two nativegoldfish SS isoforms on GH regulation are not completely under-stood, however, a possible explanation exists. Previous studiesindicate that Sst2 and Sst5 are the major Sst isoforms expressedin the goldfish pituitary (Lin et al., 2000, 2002). gbSS-28 binds gold-fish Sst5 with 4-fold higher affinity than SS-14 (Lin et al., 2002).Paralleling the effects of gbSS-28 to inhibit basal and stimulated[Ca2+]i responses, Sst5 activation in mammals is linked to suppres-sion of [Ca2+]i in addition to inhibition of adenylate cyclase (Liapa-kis et al., 1996; Siehler and Hoyer, 1999; Peverelli et al., 2009). Inaddition, unlike SS-14, gbSS-28 is not effective in inhibiting aden-ylate cyclase via Sst2 receptors (Lin et al., 2000). Thus, one canspeculate that inhibitory action of SS isoforms may be mediatedby multiple subsets of receptors which are differentially coupleto [Ca2+]i and other signaling pathways.
4.4. Further evidence on differential signaling in sGnRH- and cGnRH-II-induced GH secretion
In addition to demonstrating, for the first time, a possible differ-ence in sensitivity of intracellular Ca2+ stores utilized by sGnRHand cGnRH-II to manipulation of extracellular Ca2+ availability, re-sults from the present study also provide evidence for the presenceof other differences in sGnRH and cGnRH-II signaling in goldfishsomatotropes. The [Ca2+]i responses to 2-min applications ofcGnRH-II are more prolonged relative to responses to sGnRH, withthe former having a more prominent plateau phase response. gbSS-28 affects both the peak and plateau phases of the GH and [Ca2+]i
responses to cGnRH-II while preferentially only attenuating theplateau phase responses to sGnRH. Since a single goldfish somato-trope can respond to both sGnRH and cGnRH-II (Johnson andChang, 2000), these observations are consistent with the idea thatthe peak and plateau responses elicited by the two GnRHs aremediated by non-identical signaling cascades within the samecells. Whether and how signaling cascade(s) participate in the dif-ferential actions of sGnRH and cGnRH-II on the peak and plateauresponses is not known but PKC appears to play a role. ThePKCa/b selective inhibitor Gö6976 preferentially blunts the peakphase [Ca2+]i response to cGnRH-II, indicating that conventionalPKC isoform(s), such as the PKCa that is know to be present ingoldfish somatotropes (Klausen et al., 2005), participates in medi-ating this early stage of the cGnRH-II induced Ca2+ signal. Con-versely, the general PKC inhibitor Bis II, but not Gö6976, has astrong attenuating effect on the plateau phase [Ca2+]i response tocGnRH-II suggesting that non-conventional (perhaps novel) PKCisoforms are involved in the prolonged action of cGnRH-II on[Ca2+]i. In contrast, such differences in the involvement of PKC iso-forms are not evident with sGnRH although it must be said that theplateau phase of the sGnRH Ca2+ response is not very prominentmaking such comparisons of phasic involvement of PKC isoformsin sGnRH action more difficult in our present short-pulse challengestudies.
4.5. Summary and model of gbSS-28 action
Results from the present study are consistent with the hypoth-esis that gbSS-28 and SS-14 differ in their intracellular signalingleading to suppression of basal and stimulated GH secretion ingoldfish (Fig. 9). While SS-14 reduction of basal and GnRH-inducedGH release is independent of changes in [Ca2+]i, gbSS-28 decreasesbasal GH release by reducing resting [Ca2+]i. Reduction in a PKC-dependent increases in [Ca2+]i is one of the mechanisms by whichgbSS-28 inhibitory action on GnRH-induced GH release is ex-pressed. Given the importance of extracellular Ca2+ in controllingGH release and the known effects of SS on voltage-sensitive ionchannels in other animal/tissue models (Yang and Chen, 2007),gbSS-28 may similarly act on membrane voltage-sensitive ionchannels in goldfish somatotropes, resulting in this reduction of[Ca2+]i. Whether gbSS-28 attenuates GnRH-elicited, but PKC-inde-pendent, [Ca2+]i increases via interference with mobilization ofCa2+ from intracellular sources remain to be investigated. None-the-less, results from the present study provided insights contrib-uting to the understanding of how interactions between inhibitoryand stimulatory neuroendocrine regulators might occur in the reg-ulation of GH release in goldfish.
Acknowledgments
This research was supported by an NSERC Individual DiscoveryGrant to J.P.C. Financial support for Y.Y. was provided by a F.S. ChiaPhD Recruitment Scholarship and Graduate Teaching Assistant-
Y. Yu, J.P. Chang / General and Comparative Endocrinology 168 (2010) 71–81 81
ships from the Department of Biological Sciences, University of Al-berta. We thank Dr. Jean Rivier for the generous supply of gbSS-28.
References
Brazeau, P., Ling, N., Esch, F., Bohlen, P., Benoit, R., Guillemin, R., 1981. Highbiological activity of the synthetic replicates of somatostatin-28 andsomatostatin-25. Regul. Pept. 1, 255–264.
Canosa, L.F., Cerda-Reverter, J.M., Peter, R.E., 2004. Brain mapping of threesomatostatin encoding genes in the goldfish. J. Comp. Neurol. 474, 43–57.
Canosa, L.F., Chang, J.P., Peter, R.E., 2007. Neuroendocrine control of growthhormone in fish. Gen. Comp. Endocrinol. 151, 1–26.
Chakrabarti, R., Kumar, S., 2000. Diacylglycerol mediates the T-cell receptor-drivenCa2+ influx in T cells by a novel mechanism independent of protein kinase Cactivation. J. Cell Biochem. 78, 222–230.
Chang, J.P., Cook, H., Freedman, G.L., Wiggs, A.J., Somoza, G.M., de Leeuw, R., Peter,R.E., 1990. Use of a pituitary cell dispersion method and primary culture systemfor the studies of gonadotropin-releasing hormone action in the goldfish,Carassius auratus I. Initial morphological, static, and cell column perifusionstudies. Gen. Comp. Endocrinol. 77, 256–273.
Chang, J.P., Van Goor, F., Jobin, R.M., Lo, A., 1996. GnRH signaling in goldfishpituitary cells. Biol. Signals 5, 70–80.
Chang, J.P., Van Goor, F., Wong, A.O., Jobin, R.M., Neumann, C.M., 1994. Signaltransduction pathways in GnRH- and dopamine D1-stimulated growthhormone secretion in the goldfish. Chin. J. Physiol. 37, 111–127.
Gahete, M.D., Duran-Prado, M., Luque, R.M., Martinez-Fuentes, A.J., Quintero, A.,Gutierrez-Pascual, E., Cordoba-Chacon, J., Malagon, M.M., Gracia-Navarro, F.,Castano, J.P., 2009. Understanding the multifactorial control of growth hormonerelease by somatotropes: lessons from comparative endocrinology. Ann. N. Y.Acad. Sci. 1163, 137–153.
Gromada, J., Hoy, M., Olsen, H.L., Gotfredsen, C.F., Buschard, K., Rorsman, P., Bokvist,K., 2001. Gi2 proteins couple somatostatin receptors to low-conductance K+
channels in rat pancreatic alpha-cells. Pflugers Arch. 442, 19–26.Jacovidou, N., Patel, Y.C., 1987. Antiserum to somatostatin-28 augments growth
hormone secretion in the rat. Endocrinology 121, 782–785.Jobin, R.M., Chang, J.P., 1992a. Actions of two native GnRHs and protein kinase C
modulators on goldfish pituitary cells studies on intracellular calcium levelsand gonadotropin release. Cell Calcium 13, 531–540.
Jobin, R.M., Chang, J.P., 1992b. Differences in extracellular calcium involvementmediating the secretion of gonadotropin and growth hormone stimulated bytwo closely related endogenous GnRH peptides in goldfish pituitary cells.Neuroendocrinology 55, 156–166.
differentially involved in gonadotropin-releasing hormone-induced growthhormone gene expression in the goldfish pituitary. Am. J. Physiol. Regul.Integr. Comp. Physiol. 289, R1625–1633.
Kwiecien, R., Tseeb, V., Kurchikov, A., Kordon, C., Hammond, C., 1997. Growthhormone-releasing hormone triggers pacemaker activity and persistent Ca2+
oscillations in rat somatotrophs. J. Physiol. 499, 613–623.Li, C.J., Wei, Q.W., Zhou, L., Cao, H., Zhang, Y., Gui, J.F., 2009. Molecular and
expression characterization of two somatostatin genes in the Chinese sturgeon,Acipenser sinensis. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 154, 127–134.
Liapakis, G., Tallent, M., Reisine, T., 1996. Molecular and functional properties ofsomatostain receptor subtypes. Metabolism 45, 12–13.
Lin, X., Janovick, J.A., Cardenas, R., Conn, P.M., Peter, R.E., 2000. Molecular cloningand expression of a type-two somatostatin receptor in goldfish brain andpituitary. Mol. Cell. Endocrinol. 166, 75–87.
Lin, X., Nunn, C., Hoyer, D., Rivier, J., Peter, R.E., 2002. Identification andcharacterization of a type five-like somatostatin receptor in goldfish pituitary.Mol. Cell. Endocrinol. 189, 105–116.
Lin, X., Otto, C.J., Peter, R.E., 1999. Expression of three distinct somatostatinmessenger ribonucleic acids (mRNAs) in goldfish brain: characterization of thecomplementary deoxyribonucleic acids, distribution and seasonal variation ofthe mRNAs, and action of a somatostatin-14 variant. Endocrinology 140, 2089–2099.
Marchant, T.A., Fraser, R.A., Andrews, P.C., Peter, R.E., 1987. The influence ofmammalian and teleost somatostatins on the secretion of growth hormone
from goldfish (Carassius auratus L.) pituitary fragments in vitro. Regul. Pept 17,41–52.
Martiny-Baron, G., Kazanietz, M.G., Mischak, H., Blumberg, P.M., Kochs, G., Hug, H.,Marme, D., Schachtele, C., 1993. Selective inhibition of protein kinase Cisozymes by the indolocarbazole Go 6976. J. Biol. Chem. 268, 9194–9197.
McFarlane, M.B., Bruhn, T.O., Jackson, I.M., 1993. Postsomatostatin hypersecretionof growth hormone from perifused rat anterior pituitary cells is dependent oncalcium influx. Neuroendocrinology 57, 496–502.
McFerran, B.W., Guild, S.B., 1994. Effects of protein kinase C activators upon the latestages of the ACTH secretory pathway of AtT-20 cells. Brit. J. Pharmacol. 113,171–178.
Mesguich, P., Benoit, R., Dubois, P.M., Morel, G., 1988. Somatostatin-28- andsomatostatin-14-like immunoreactivities in the rat pituitary gland. Cell TissueRes. 252, 419–427.
Morgan, A., Burgoyne, R.D., Barclay, J.W., Craig, T.J., Prescott, G.R., Ciufo, L.F., Evans,G.J., Graham, M.E., 2005. Regulation of exocytosis by protein kinase C. Biochem.Soc. Trans. 33, 1341–1344.
Nunn, C., Feuerbach, D., Lin, X., Peter, R., Hoyer, D., 2002. Pharmacologicalcharacterization of the goldfish somatostatin sst5 receptor. Eur. J. Pharmacol.436, 173–186.
Peverelli, E., Lania, A.G., Mantovani, G., Beck-Peccoz, P., Spada, A., 2009.Characterization of intracellular signaling mediated by human somatostatinreceptor 5: role of the DRY motif and the third intracellular loop. Endocrinology150, 3169–3176.
Scanlan, N., Dufourny, L., Skinner, D.C., 2003. Somatostatin-14 neurons in the ovinehypothalamus: colocalization with estrogen receptor alpha and somatostatin-28(1–12) immunoreactivity, and activation in response to estradiol. Biol.Reprod. 69, 1318–1324.
Sheridan, M.A., Kittilson, J.D., 2004. The role of somatostatins in the regulation ofmetabolism in fish. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 138, 323–330.
Shipston, M.J., Armstrong, D.L., 1996. Activation of protein kinase C inhibitscalcium-activated potassium channels in rat pituitary tumor cells. J. Physiol.493, 665–672.
Siehler, S., Hoyer, D., 1999. Characterization of human recombinant somatostatinreceptors. 3. Modulation of adenylate cyclase activity. Naunyn SchmiedebergsArch. Pharmacol. 360, 510–521.
Spiess, J., Vale, W., 1980. Multiple forms of somatostatin-like activity in rathypothalamus. Biochemistry 19, 2861–2866.
Steinberg, S.F., 2008. Structural basis of protein kinase C isoform function. Physiol.Rev. 88, 1341–1378.
Toullec, D., Pianetti, P., Coste, H., Bellevergue, P., Grand-Perret, T., Ajakane, M.,Baudet, V., Boissin, P., Boursier, E., Loriolle, F., 1991. The bisindolylmaleimide GF109203X is a potent and selective inhibitor of protein kinase C. J. Biol. Chem.266, 15771–15781.
Trabucchi, M., Tostivint, H., Lihrmann, I., Jegou, S., Vallarino, M., Vaudry, H., 1999.Molecular cloning of the cDNAs and distribution of the mRNAs encoding twosomatostatin precursors in the African lungfish Protopterus annectens. J. Comp.Neurol. 410, 643–652.
Tsaneva-Atanasova, K., Sherman, A., Van Goor, F., Stojilkovic, S.S., 2007. Mechanismof spontaneous and receptor-controlled electrical activity in pituitarysomatotrophs: experiments and theory. J. Neurophysiol. 98, 131–144.