THE JOURNAL OF BKILOGICAL CHEMISTRY Vol. 245, No. 23, Issue of December 10, pp. 6357-6365, 1970 Printed in U.S.A. Isolation and Characterization of Plasma Membranes from Human Blood Platelets* (Received for publicat,ion, July 9, 1970) A. J. BARBER~ AND G. A. JAMIESON From The American National Red CrossBlood Research Laboratory, Bethesda,Maryland 2001/, and the Depart- ment of Biochemistry, Georgetown University Schoolsof Medicine and Dentistry, Washington, D. C. 20007 SUMMARY Human platelets were subjected to hypoto?ic lysis after being loaded intracellularly with glycerol (4.3 M) by centrifu- gation in a glycerol gradient under controlled conditions. A homogeneous membrane fraction, free of intracellular or- ganelles, was isolated after a single density step centrifuga- tion, while continuous sucrose density gradient centrifuga- tion gave two subfractions with densities of 1.090 and 1.120, and with identical isoelectric points (p1 3.9) as determined by isoelectric focusing. Compositional studies indicated that the platelet membrane was a lipoglycoprotein with a carbohydrate content of ap- proximately 7 %. The sugar components were identified as glucose, galactose, mannose, hexosamines (GlcN: GalN = 6:1), sialic acid, and fucose. The chemical composition of the platelet membranes isolated at d, 1.090 was protein, 31.9%; lipid, 55.9%; carbohydrate, 7.3%; RNA, 0.3 %; DNA, 0.0%; and for those isolated at d, 1.120, it was protein, 40.1%; lipid, 48.2%; carbohydrate, 6.9%; RNA, 0.4%; DNA, 0.0%. The membranes were characterized by a high molar ratio of cholesterol to phospholipid (0.49 and 0.45) that was similar to that found in liver plasma membranes. Phosphodiesterase, acid phosphatase and ATPase were purified 8-, 4-, and 2-fold, respectively, in the membrane fractions, whereas succinic dehydrogenase, esterase, and a variety of P-glycosidases were present only at very low levels. These results demonstrate the absence of intracellular mem- branes and, together with the chemical analyses, they suggest that the outer membrane of the platelet is similar to plasma membranes of other cells, and they provide biochemical con- firmation of its origin from the plasma membrane of the megakaryocyte. The vesicles of the two membrane bands differed in ultra- structure, the lighter (d, 1.090) having an average diameter of 1750 A with numerous concentric double membrane struc- * This work was supported in part by United States Public Health Service Grants GM-13057 and AI-09017. Platelets used in this study were provided by the Washington Regional Red Cross Blood Center under Contract PH 43-64-102 of the National Cancer Institute. This is Contribution 202 from the Blood Re- search Laboratory of the American National Red Cross, and it is the sixth paper in a series on studies on glycoproteins. $ This study is in partial fulfillment of the requirements for the degree of doctor of philosophy in the Department of Biochemistry, Georgetown University Schools of Medicine and Dentistry. tures, while the heavier (d, 1.120) consisted of single mem- branes of diameter 700 A. Recent studies in this laboratory have been directed toward the elucidation of certain structural (1, 2) and functional (3) features of the component glycoproteins of human platelet membranes. Two main functions can generally be attributed to cell surface membranes, one concerned with transport and permeability and the other with cell interactions and contact (for reviews, see References 4 and 5). The relative importance of these membrane functions differs according to the cell type. In the case of platelets, the cell contact function is increased, and the membranes play an important role in blood coagulation, hemostasis, and thrombosis (6-8), in the attachment of viruses (3), in serotonin uptake (9), in platelet aggregation (lo), and in adhesion to collagen (11). In addition, the surface membrane of the platelet contains several of the recognized HL-A antigenic determinants (12, 13) found on lymphocytes and other cells, and these may be involved in tissue histocompatibility and atherosclerosis (14). A major problem in biochemical studies of the platelet mem- brane has been the lack of a suitable method for their isolation. Such a method should be simple in operation and should give a high and reproducible yield of the platelet plasma membrane uncontaminated with intracellular components. As a result of an exhaustive study of methods applicable to the isolation of platelet membranes,’ we have developed a hypotonic lysis technique based on the extensive investigations of Meryman and associates in this laboratory concerning the unique differential properties of the platelet membrane with regard to the physio- logical transport of glycerol and water (15, 16). This method meets the above criteria for platelet membrane isolation. Application of this method has led to the isolation of two types of membrane vesicles in high yield, and extensive enzymatic, chemical, and ultrastructural analysis has shown that the plate- let membrane, despite its unique cytological origin, is similar to the plasma membrane of other cells. 1 A. J. Barber, D. S. Pepper, and G. A. Jamieson, unpublished studies. 6357 by guest on July 26, 2018 http://www.jbc.org/ Downloaded from
Transcript
THE JOURNAL OF BKILOGICAL CHEMISTRY Vol. 245, No. 23, Issue of December 10, pp. 6357-6365, 1970
Printed in U.S.A.
Isolation and Characterization of Plasma Membranes
from Human Blood Platelets*
(Received for publicat,ion, July 9, 1970)
A. J. BARBER~ AND G. A. JAMIESON
From The American National Red Cross Blood Research Laboratory, Bethesda, Maryland 2001/, and the Depart- ment of Biochemistry, Georgetown University Schools of Medicine and Dentistry, Washington, D. C. 20007
SUMMARY
Human platelets were subjected to hypoto?ic lysis after being loaded intracellularly with glycerol (4.3 M) by centrifu- gation in a glycerol gradient under controlled conditions. A homogeneous membrane fraction, free of intracellular or- ganelles, was isolated after a single density step centrifuga- tion, while continuous sucrose density gradient centrifuga- tion gave two subfractions with densities of 1.090 and 1.120, and with identical isoelectric points (p1 3.9) as determined by isoelectric focusing.
Compositional studies indicated that the platelet membrane was a lipoglycoprotein with a carbohydrate content of ap- proximately 7 %. The sugar components were identified as glucose, galactose, mannose, hexosamines (GlcN: GalN = 6:1), sialic acid, and fucose. The chemical composition of the platelet membranes isolated at d, 1.090 was protein, 31.9%; lipid, 55.9%; carbohydrate, 7.3%; RNA, 0.3 %; DNA, 0.0%; and for those isolated at d, 1.120, it was protein, 40.1%; lipid, 48.2%; carbohydrate, 6.9%; RNA, 0.4%; DNA, 0.0%. The membranes were characterized by a high molar ratio of cholesterol to phospholipid (0.49 and 0.45) that was similar to that found in liver plasma membranes.
Phosphodiesterase, acid phosphatase and ATPase were purified 8-, 4-, and 2-fold, respectively, in the membrane fractions, whereas succinic dehydrogenase, esterase, and a variety of P-glycosidases were present only at very low levels. These results demonstrate the absence of intracellular mem- branes and, together with the chemical analyses, they suggest that the outer membrane of the platelet is similar to plasma membranes of other cells, and they provide biochemical con- firmation of its origin from the plasma membrane of the megakaryocyte.
The vesicles of the two membrane bands differed in ultra- structure, the lighter (d, 1.090) having an average diameter of 1750 A with numerous concentric double membrane struc-
* This work was supported in part by United States Public Health Service Grants GM-13057 and AI-09017. Platelets used in this study were provided by the Washington Regional Red Cross Blood Center under Contract PH 43-64-102 of the National Cancer Institute. This is Contribution 202 from the Blood Re- search Laboratory of the American National Red Cross, and it is the sixth paper in a series on studies on glycoproteins.
$ This study is in partial fulfillment of the requirements for the degree of doctor of philosophy in the Department of Biochemistry, Georgetown University Schools of Medicine and Dentistry.
tures, while the heavier (d, 1.120) consisted of single mem-
branes of diameter 700 A.
Recent studies in this laboratory have been directed toward the elucidation of certain structural (1, 2) and functional (3) features of the component glycoproteins of human platelet membranes. Two main functions can generally be attributed to cell surface membranes, one concerned with transport and permeability and the other with cell interactions and contact (for reviews, see References 4 and 5). The relative importance of these membrane functions differs according to the cell type. In the case of platelets, the cell contact function is increased, and the membranes play an important role in blood coagulation, hemostasis, and thrombosis (6-8), in the attachment of viruses (3), in serotonin uptake (9), in platelet aggregation (lo), and in adhesion to collagen (11). In addition, the surface membrane of the platelet contains several of the recognized HL-A antigenic determinants (12, 13) found on lymphocytes and other cells, and these may be involved in tissue histocompatibility and atherosclerosis (14).
A major problem in biochemical studies of the platelet mem- brane has been the lack of a suitable method for their isolation. Such a method should be simple in operation and should give a high and reproducible yield of the platelet plasma membrane uncontaminated with intracellular components. As a result of an exhaustive study of methods applicable to the isolation of platelet membranes,’ we have developed a hypotonic lysis technique based on the extensive investigations of Meryman and associates in this laboratory concerning the unique differential properties of the platelet membrane with regard to the physio- logical transport of glycerol and water (15, 16). This method meets the above criteria for platelet membrane isolation.
Application of this method has led to the isolation of two types
of membrane vesicles in high yield, and extensive enzymatic, chemical, and ultrastructural analysis has shown that the plate- let membrane, despite its unique cytological origin, is similar to the plasma membrane of other cells.
1 A. J. Barber, D. S. Pepper, and G. A. Jamieson, unpublished studies.
Characterization of Plasma Membranes from Human Platelets Vol. 245, No. 23
FIG. 1. A, density step (10 ml of sucrose, 27% (w/v), d, 1.106) cent,rifugation of platelet lysate. B, continuous density gradient (28 ml of sucrose, 15 t.o 40y0 (w/v), d, 1.060 to 1.150) centrifugation of membrane fraction from A.
PLATELET CONCENTRATES
I Erythrocyte Pellet
1.500 xg, I min. 4O t
Supernotant-Platelet Rich Plasma
(i 1 4300 xg, IOmin, 4O (ii! Suspend in 0.001 M EDTA, 0.01 M
Tris (pH 7.5), 0.15 M NoCl (iii) Wash two times
I Supernotont
I I
Washed Platelets
I I) waver on O-40% lsotonlc qlycerol gradlent
I I ii) 1465 xg, 30 min, 4O (iii) 5860 xg, IO min. 4’
Supernotant
t Pellet-Glycerol Loaded Platelets
(i ) Lyse by addition of 0.01 M Tris (pH 7.5)- 0.25 M Sucrose
(ii) Layer on d, I.106 sucrose (iii) 63500 xg, 3-5 hours, 4O
(I) Dilute wth Tris-sucrose (II) 105,000 xg, I hour, 4O
t Membrane Pellet
( I 1 Susoend I” Trwsucrose
I (ii) Lay& on 15-40% linear sucrose’grodient cd, 1.060-1.1501
(iii) 63,500 xg, I8 hours, 4O t t t
Upper Bond d. 1.090 Lower Band d, I.120 Platelet Membrane Platelet Membrane
FIG. 2. Isolatiou of platelet membranes
MATERIBLS .4X1) METHODS
Prepxation of Iluman Platelet Membranes-Single units (-450 ml) of blood were drawn from normal volunteer blood donors, with acid-citrate dextrose (NIH Solution A) as anti- coagulant. Within 4 hours of blood collection, the platelets were isolated from individual units by two cycles of differential centrifugation (17) and were finally freed of residual erythrocytes by repeated differential centrifugation at 3300 rpm (1500 X g) for 1 min in an International swing-out rotor (International
Equipment Company, Needham Heights, Massachusetts). The platelet preparation was then sedimented at 6000 rpm (4300 X g) in a Sorvall RC-2 centrifuge (Ivan Sorvall, Inc., Norwalk, Connecticut) and washed three times in cold 0.001 M EDTA, 0.01 M Tris-HCl (pH 7.5), 0.15 M NaCl. The methods described in this section were routinely applied to a pool of platelets ob- tained from 16 individual donor units (m-8 liters of blood) but were found to be equally satisfactory when applied at a level of 1 unit or 30 units in a single run.
The intracellular glycerol concentration was raised to 4.3 M
by centrifuging the platelets very slowly through isotonic glycerol gradients; 30 ml (0 to 40%, 3,000 rpm (1,465 X g) for 30 min, followed by 6,000 rpm (5,860 x g) for 10 mm) at 4” in equal portions in four tubes in the Sorvall HB-4 swing-out rotor. The supernatant glycerol solutions were carefully and completely removed by aspiration. A volume of cold Tris-sucrose (0.01 M Tris-HCl (pH 7.5)-0.25 M sucrose) was added equal to approximately 4 to 5 times the volume of the platelet buttons, and the platelets were suspended and simultaneously lysed by rapid mixing of each tube on a Vibromixer. All operations were carried out at ice-bath temperatures.
For isolation of the membrane fraction, the lgsed platelet suspension was divided in three portions and layered on top of a solution of 27% (w/v) sucrose (d, 1.106) in the three tubes of the SW 25.1 rotor of a Reckman model L ultracentrifuge and centrifuged at 4” at 25,000 rpm (63,500 x g) for 3 to 5 hours. T;nbroken platelets, granules, and other debris sedimented to the bottom of the tubes, while the membrane fraction remained as a narrow band above the sucrose interface (Fig. IA). The membrane bands were isolated, diluted 1 :lO with cold Tris- sucrose buffer, and pelleted by centrifugation in the No. 30 rotor for 1 hour at 4” and 30,000 rpm (105,000 x g).
The membrane pellet was then suspended in cold Tris-sucrose buffer, layered on a linear sucrose density gradient (15 to 40% (w/v), d, 1.060 to 1.150), and centrifuged for 18 hours in the SW 25.1 rotor of the ultracentrifuge at 63,500 x g. The two resulting membrane bands (Fig. 1B) were removed by careful aspiration, diluted 1: 10 with cold Tris-sucrose buffer, and cen- trifuged at 4” at 30,000 rpm for 1 hour in the No. 30 rotor. The pellets were suspended in Tris-sucrose and stored at - 40”. The various steps of the isolation procedure are summarized in Fig. 2.
Electron Microscopy-Pellets of the various fractions obtained during the isolation procedure were fixed for 2 to 3 hours in a freshly prepared solution of glutaraldehyde (1.25%) in 0.05 M
phosphate buffer, pH 7.3, containing 0.2 M sucrose. After washing, the pellets were fixed in 1% OsO*, dehydrated in ethanol-water solutions of increasing concentrations of ethanol, stained with many1 acetate, and embedded in Araldite (Ciba pharmaceuticals, Inc., Summit, Kew Jersey). Thin sections, tnounted on carbon-covered grids, were stained with lead citrate (pH 12). Specimens were examined with an .1EI-6 13 electron microscope (Biodynamics Research Corporation, Rockville, Maryland).
Chemical Analyses-The various fractions obtained from the isolation procedure were dialyzed exhaustively for 2 days against distilled water, lyophilized, and weighed.
Protein--Protein was determined by the procedure of Lowry et al. (18), the biuret method (19), or the Kjeldahl procedure as reported by Koch and McKeekin (20). Human serum albumin was used as standard.
Carbohydrate-Sialic acid was determined by thiobarbituric
ksue of December 10, 1970 A. J. Barber and G. A. Jamieson 6359
300 nm 200 nm -- -
FIG. 3. Electron micrographs of human platelet fractions. A, platelet membranes isolated after stepwise centrifugation. No contaminating subcellular structures are present. X 20,000. 13, upper band (d, 1.090) of platelet membranes isolated after continuous gradient centrifugation. No contaminat.ing struc- tures are present. The average diameter of the vesicles is 1750 A. Note t,he appearance of many double membrane structures. X 40,000. c’, lower band (d, 1.120) of platelet membranes iso- lated after continuous gradient centrifugation. No contaminat- ing structures are present. The average diameter of the vesicles is 700 A. Note the absence of any double membrrtne structures.
acid assay (21) after hydrolysis with 0.1 N H2S04 at 85” for 30 min. N-Acetylneuraminic acid (synthetic) was used as stand- ard. Optical density readings were made routinely at 532 and 549 nm, and a correction was made for interfering substances (22). Total hexosamine was determined by the Elson-Morgan procedure as modified by Gatt and Berman (23) after hydrolysis in 2 N HCl at 100” for 12 hours in sealed tubes. The long column (60 cm) of the Beckman amino acid analyzer, model 120 C, was employed for the individual estimation of glucosamine and galactosamine. Glucosamine and galactosamine were eluted at. 51 and 77 ml, respectively, after phenylalanine. Crystalline glucosamine and galactosamine hydrochloride from Mann were used as st.andards.
Total hexose was determined by the phenol-sulfuric acid method of Dubois et al. (24), with an equimolar mixture of galactose and glucose as standard. Glucose, galactose, mannose, and fucose were determined by quantitative gas chromatography, with the use of a modified alditol acetate procedure (I), following hydrolysis in 0.25 N HzS04 at 100” for 24 hours in a nitrogen atmosphere. n-hrabinose (96 pg) was added prior to hydrolysis as an internal standard. Chromatography was carried out with a 4-foot column of 3% ECNSS-M on Gaschrom Q (Applied Science Laboratories, Inc., State College, Pennsylvania) (100 to 200 mesh) at 190”, at a nitrogen flow rate of 80 ml per min, with a flame ionization detector.
Nucleic A&%--RNA determinations were performed by the Schmidt-Thannhauser method as described by Mum-o and Fleck (25), assuming an optical density at 260 nm of 1.000 for an RNA concentration of 32 pg per ml. DNA was measured by the diphenylamine reaction (26) after hydrolysis in 0.5 N perchloric acid for 15 min at 90". Calf thymus DKA (Calbiochem) was used as standard.
Lipid-Total lipids were determined gravimetrically after extraction three times with chloroform-methanol (2:l w/v) at room temperature for 24 hours and partitioning against NaCl by the method of Folch, Lees, and Sloane Stanley (27). Total phosphorus and organic-extractable phosphorus, determined after perchloric acid combustion (28), were analyzed by the method of Bartlett (29). Phospholipid was calculated by assuming 25 pg of phospholipid per pg of lipid phosphorus (30). Cholesterol was determined by the method of Watson (31).
Amino Acids-Amino acid analyses were performed on the membrane and lysate fractions. Hydrolysis was carried out with glass-distilled, constant boiling HCl at 108” in an atmos- phere of nitrogen for periods of 24, 48, 72, and 96 hours at a weight concentration of 1 mg of membrane or lysate per ml. After hydrolysis, the HCl was evaporated to dryness, and the dried samples were dissolved in 0.2 N citrate buffer, pH 2.2. The amino acid analyses were performed on aliquots of the hydrolysate on a Beckman model 120 C amino acid analyzer.
Enzyme Assays-All enzyme assays were performed at 37” unless otherwise indicated. Under the conditions of each assay, the rate of the reaction was shown to be linear with time and proportional to the amount of enzyme preparation used. One unit of activity was defined as the amount causing the removal of 1 pmole of substrate or yielding 1 pmole of product per hour. When Triton X-100 was used, the final concentration in the incubation mixture was 0.15% v/v.
Mg2+- (or Ca2+-)ATPase (EC 3.6.1 .S)-The Mg2+- or Ca2f- dependent adenosine triphosphatase was measured according to the method of Chambers, Salzman, and Neri (32).
Na+-K+-Mg2+-A TPase (EC S .6.1.4)-The Na+, Kf-stimu- lated, Mgzf-dependent ATPase was measured by the method of Wallach and Kamat (33).
Phosphodiesterase (EC S 1.4. I)-This enzyme was assayed at 420 nm by the liberation of p-nitrophenol from bis(p-nitro- phenyl)phosphate (34).
Acid Phosphatase (EC S.1 .S.d)-This enzyme was measured by the release of p-nitrophenol from p-nitrophenyl phosphate (35).
Esterme (EC 3.1 .l .I)-This enzyme was estimated with p-nitrophenyl acetate as substrate (36).
N-Acetylglucosaminidase (EC S. 2.1. SO)-This enzyme was determined by the method described by Langley and Jevons (37), with p-nitrophenyl-N-acetyl-P-D-glucosaminide as sub- strate. The reaction was terminated with 0.2 M carbonate- bicarbonate buffer (pH 10.8).
/3-Glucuronidase (EC S .d. 1. .%-The glucuronidase activity was assayed with phenolphthalein glucuronide as substrate (38).
&Galactosidase (EC S. 2.1.2,!?-This enzyme was determined with o-nitrophenyl-P-n-galactopyranoside as substrate (39).
Succinic Dehydrogenase (EC 1. S. 99. I)-This enzyme was
Issue of December 10, 1970 A. J. Barber and G. A. Jamieson 6361
measured at 30” with 2,6-dichlorophenolindophenol as the electron acceptor (40).
Lactic Dehydrogenase (EC 1.1.1. %‘)-This enzyme was deter- mined spectrophotometrically by the method of Wroblewski and LaDue (41).
Alkaline Phosphatase (EC S. 1. S. 1)-The alkaline phosphatase activity was measured by the liberation of p-nitrophenol from p-nitrophenyl phosphate (42).
Leucine Aminopeptidase (EC S .4.1. I)-This activity was estimated by the hydrolysis of L-leucyl-P-naphthylamide (43).
Isoelectric Focusing-The method used was identical with that developed for the study of red cell and lymphocyte membranes.2 An LKB model 8101 electrofocusing column (110 ml) was used with a 2% (w/v) Ampholine carrier of pH range 3.0 to 5.0 (LKB Instruments, Inc., Rockville, Maryland). Membrane fractions (4.0 mg of protein) were added to the heavy solution in lieu of water. After running for 80 hours at 4” (300 volts; 2.5 ma, decreasing to 1.0 ma) (44), the sucrose gradients were pumped out, and 3-ml fractions were collected, dialyzed exhaustively for 3 days against distilled water, concentrated, and assayed for protein and sialic acid content.
RESULTS
Isolation Procedure-The method described permits the isola- tion of a homogeneous membrane fraction in about 5 hours.
Examination of the initial platelet lysate by phase contrast microscopy showed that 80 to 90% of the platelets were ruptured by the present method of glycerol-loaded, hypotonic lysis. The figure was confirmed by the fact that over 75% of the lactic dehydrogenase activity, a known soluble cytoplasmic enzyme, was found in the soluble fraction of the platelet lysate.
Examination of the lysate in the electron microscope indicated the presence of membranous structures, numerous granules, and small dense bodies, demonstrating that extensive damage to intracellular organelles does not occur by the glycerol-lysis technique.
The sucrose step centrifugation was most critical. When a preliminary differential centrifugation was performed, the mem- brane fraction, mitochondria, granules, and other cellular debris sedimented together. Consequently, the isopycnic density of the membranes was determined, and a sucrose step (d, 1.106) was employed. The material isolated from the interface of the sucrose step showed only membranous vesicles of different size and shape, with no evidence of contamination by large granules, mitochondria, or other intracellular organelles (Fig. 3A). The intracellular debris found at the bottom of the tube was devoid of large membrane fragments but rich in osmophilic granules, in empty vacuoles, and in a wide variety of other, undefined struc- tures. Soluble protein occurred above the interface, and all traces of it could be removed by washing the membranes.
Two distinct membrane fractions were then isolated by means of isopycnic gradient centrifugation. These two fractions, which had peak densities of 1.090 and 1.120, were obtained in equal amounts, each corresponding to about 3% of the dry weight of the platelet (Table I).
Morphology of Isolated Platelet Membranes-Examination of the platelet membrane fractions under the electron microscope revealed that each fraction was morphologically homogeneous and consisted of large membranous structures completely devoid
z N. Groh and G. A. Jamieson, unpublished procedure.
TABLE II
Analysis of human platelet membranes and total lysate
a Residues per 1000 residues of amino acid, to the nearest whole number.
b Unless otherwise indicated, the values for the amino acids represent an average of the analyses obtained at 24, 48, and 72 hours.
c Corrected for destruction during hydrolysis. d Determined on platelets less than 2 hours after phlebotomy;
if expressed as nondialyeable glucose, the value would be 0.038. B As the N-acetyl derivatives.
of contamination with subcellular elements. Several charac- teristic differences could be noted. The average diameter of the vesicles in the lighter band (d, 1.090) was 1750 A (Fig. 3B), while that of those in the heavier band (d, 1.120) was 700 A (Fig. 3C). That is, the vesicles of the upper band were, on the average, 2.5-fold greater in size than those in the lower band. Another characteristic difference was that the upper band corn tained what appeared to be double membrane structures arranged one inside the other, whereas no such structures were present in the lower band.
Chemical Composition of Platelet Membranes-The individual components recovered in the total platelet lysate, the soluble
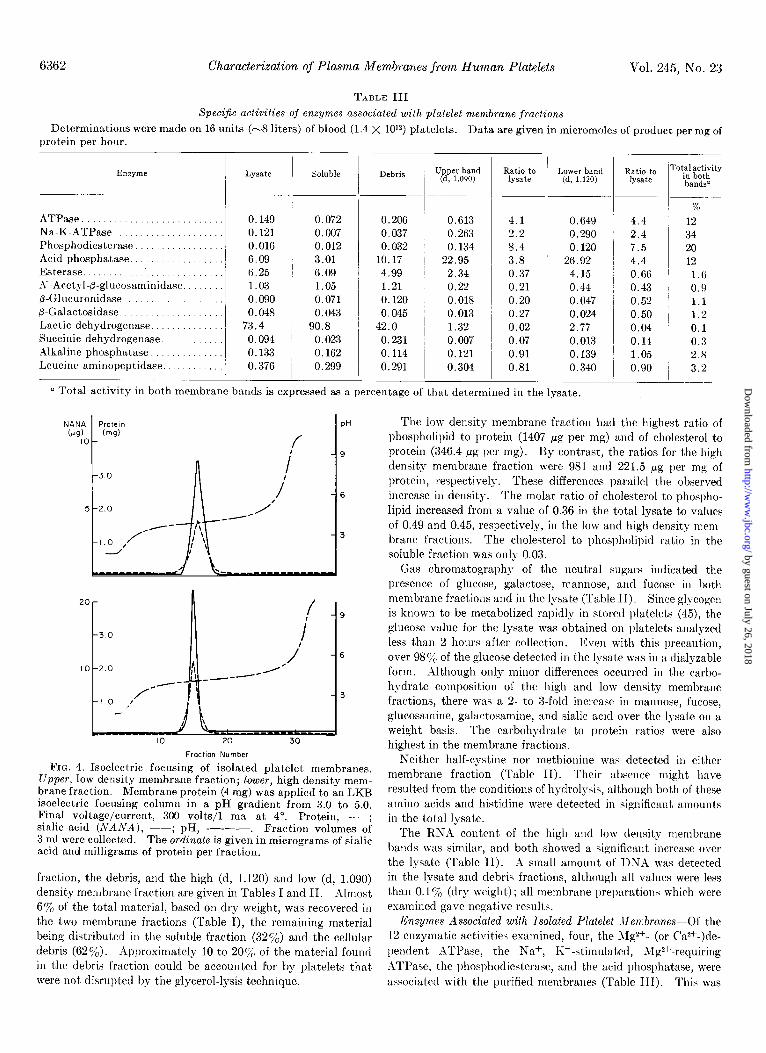
a Total activity in both membrane bands is expressed as a percentage of that determined in the lysat,e.
IO 20 30
Fraction Number
FIG. 4. Isoelectric focusing of isolated platelet membranes. Upper, low density membrane fraction; Zourer, high density mem- brane fraction. Membrane protein (4 mg) was applied to an LKB isoelectric focusing column in a pH gradient from 3.0 to 5.0. Final voltage/current, 300 volts/l ma at 4”. Protein, -----; sialic acid (AJAVA), -; pH, ------. Fraction volumes of 3 ml were collected. The ordinate is given in micrograms of sialic acid and milligrams of protein per fraction.
fraction, the debris, and the high (d, 1.120) and low (d, 1.090) density membrane fraction are given in Tables I and II. Almost 6% of the total material, based on dry weight, was recovered in the two membrane fractions (Table I), the remaining material being distributed in the soluble fraction (32%) and the cellular debris (6270). Approximately 10 to 20% of the material found in the debris fraction could be accounted for by platelets that were not disrupted by the glycerol-lysis technique.
The low density membrane fraction had the highest ratio of phospholipid to protein (1407 pg per mg) and of cholesterol to protein (346.4 pg per mg). I{y contrast, the ratios for the high density membrane fract.ion were 981 and 221.5 pg I)er mg of protein, respectively. These differences parallel the observed increase in density. The molar ratio of cholesterol to phospho- lipid increased from a value of 0.36 in the total lysate to values of 0.49 and 0.45, respectively, in the low and high delrsity n7em brane fractions. The cholesterol to phospholipid ratio in the soluble fraction was only 0.03.
Gas chromatography of the neutral sugars indicated the presence of glucose, galactose, Irannose, and fucose ill both membrane fractions and in the lysate (Table II). Since glycogen is known to be metabolized rapidly in stored platelets (45), the glucose value for the lysate was obtained on platelets analyzed less than 2 hours after collection. Even with this precaution, over 98% of the glucose detected in the lysate was in a dialyzable form. Although only minor differences occurred in the carbo- hydrate composition of the high and low density membrane fractions, there was a 2- to a-fold increase in mannose, fucose, glucosamine, galactosamine, and sialic acid over the lysate on a weight, basis. The carbohydrate to protein ratios were also highest in the membrane fractions.
Neither half-cystine nor methionine was detected in either membrane fraction (Table II). Their absence might have
resulted from the conditions of hydrolysis, alt.hough both of these amino acids and histidine mere detected in significant amouuts
in the total lysate. The RNA content of the high and low density membrane
bands was similar, and both showed a significant increase over the lysate (Table II). -1 small amount of l)N,Z was detected in the lysate and debris fract,ions, although all values were less than O.lGiO (dry weight); all membrane preparations which were examined gave negative results.
Enzymes Associated with Isolated Platelet Mexbranes-Of the 12 enzymatic activitiei examined, four, the Jig*+- (or CW-)de-
pendent ATPase, the Na+, K+-stimulated, Mp2+-requiring ATl’ase, the phosphodiesterase, and the acid phosphatase, were associated with the purified membranes (Table III). This was
Issue of December 10, 1970 A. J. Barber and G. A. Jamieson 6363
indicated by the high percentage of the total activity recovered in the membrane fraction (12 to 34’%) and by the increase in specific activity of these enzymes as compared with the lysate fraction (purification ratio, 4.4 to 7.5).
The extremely low recovery of lactic dehydrogenase in the membrane fractions indicated that they were free from soluble, cytoplasmic contamination. Similarly, the low recoveries of esterase, succinic dehydrogenase, and P-glycosidase activities indicated the absence of contamination with membranes derived from the endoplasmic reticulum, from mitochondria, and from granules, respectively. Selected enzyme assays were carried out in the presence of Triton X-100. The activity of the membrane- bound enzymes was increased approximately 2-fold in all cases, except for @-galactosidase.
Isoelectric Focusing of Isolated Platelet &fembranes-Isoelectric focusing can be used to determine the isoelectric points of cell membranes as well as soluble proteins.2 Estimation of the separated high and low density membrane fractions individually by the isoelectric focusing technique gave single bands, with identical pH values of 3.95 f 0.05 (Fig. 4). In both cases, the sialic acid peak coincided with the protein peak and was localized at the expected p1. It is interesting to note that the low density membrane fraction gave a more diffuse band under identical experimental conditions. This p1 value of 3.95 is in excellent agreement with the values of 4.0 (46) and 3.9 (47) determined by cytoelectrophoresis of intact platelets.
DISCUSSION
The isolation of human platelet membranes presents particular difficulties because of the small volume of these cells, 5 ~3 (48) and their resistance to mechanical shearing forces, although a number of physical techniques have been applied for this purpose (49-54). We have carried out a systematic comparison of various physical methods for platelet disruption, including sonic disruption, nitrogen cavitation in a high pressure bomb, and the use of a “no clearance” tissue grinder, both in the presence and absence of surface-stabilizing agents (55), and have found all such techniques unsuitable for the large scale isolation of platelet membranes on considerations of yield, reproducibility, and simplicity in application. Similarly, simple osmotic lysis was not an effective method for the isolation of platelet mem- branes.
In an attempt to develop a more effective method for platelet homogenization, we have made use of the extensive studies of Meryman and associates (15, 16) in this laboratory on the effects of ionic stress on membrane permeability in erythrocytes, plate- lets, and a wide variety of other cells. These studies have shown that the physiological integrity of the membrane in these cells is unaffected by transient osmotic stress induced by glycerol or other solutes, provided that these solutes do not exceed 4.5 times the isotonic value (56). In the particular case of the platelet membrane, however, the rate of passage of glycerol is exceedingly slow, being on the order of minutes,3 whereas water transport is relatively rapid. From this, it follows that platelets may be loaded with high intracellular concentrations of glycerol, under carefully controlled conditions, without impairment of membrane integrity, and that the cell may then be lysed by exposure to hypotonic conditions when there is a rapid influx of water but a relatively slow efflux of glycerol.
8 H. T. Meryman, personal communication.
We have utilized these observations to develop a rapid method for the lysis of intact platelets. The use of the glycerol gradient technique allows the very slow accumulation of glycerol into the platelet to a final concentration of 140~ isotonic without introducing transient concentration differences in excess of 4.5~ isotonic. This technique seems to effect a minimum of damage to the platelet that could initiate a platelet release reaction, platelet clumping, and viscous metamorphosis. More- over, since the glycerol-lysis technique depends on the integrity of the factors mediating membrane transport of glycerol, it is possible that platelets that are not lysed by this technique have an impaired passive transport system since the technique does not appear to be as effective on aged platelets.
The single sucrose density step centrifugation allows the isolation of a homogeneous membrane fraction. The membranes are freed of intracellular debris in this step by a combination of factors that depend on the density of the sedimenting particles and the g forces applied. The step density of d, 1.106 was chosen because of maximum removal of debris in a short period of time (3 to 5 hours) which does not allow significant sedimenta- tion of the high density (d, 1.120) membrane fraction. A higher step density would require a longer period of ultracentrifugation to remove debris, whereas a lower step density would result in a decreased yield of the high density membranes. The further continuous sucrose gradient centrifugation allows a subdivision into two membrane fractions that differ in phospholipid content and in ultrastructure and that have buoyant densities of 1.090 and 1.120. The origin of these two types of membrane vesicles is unknown and, although two membrane fractions have been isolated from platelets by other techniques (57, 58), they were not extensively characterized.
Human platelets are formed from the fragmentation in the bone marrow of the precursor cell, the megakaryocyte, and the demarcation membranes may arise from intracellular structures (59), although further studies by electron microscopy suggest that they are derived from its plasma membrane (60). Thus, it is not clear whether the outer membrane of the platelet can be considered analogous to the plasma membrane of other cells.
The enzyme distribution in both membrane bands isolated in the present work was similar to those of the plasma membranes of other cells. Phosphodiesterase, acid phosphatase, and ATPase showed enrichments of 8-, 4-, and 2-fold, respectively, in platelet membranes, compared with values in liver cells of 15-, 4- (61), and 6-fold (62), respectively, for plasma membranes as compared with microsomes. The low recovery and negligible increase in specific activity of alkaline phosphatase (Table III) were similar to the low activity of this enzyme in liver cell plasma membranes (63), and the low recoveries of fl-glycosidase, esterase, and succinic dehydrogenase, which usually occur in intracellular membranes, were also consistent with these two membrane bands having arisen from the plasma membrane of the platelet. Finally, a high molar ratio of cholesterol to phospholipid is characteristic of plasma membranes relative to other cellular components of the same tissue (63). For platelets, the molar ratios of 0.49 and 0.45, which were highest in the low and high density membrane fractions, respectively, fell within the range of 0.38 to 0.74 determined for liver cells (62, 64) and are com- parable with the values reported for membrane preparations from L cells (0.69) (65), HeLa cells (1.05) (42), and erythrocytes (1.15) (66).
These results strongly support the hypothesis that the outer
6364 Characterixdim of Plasma Membranes from Human Platelets Vol. 245, No. 23
membrane of the platelet is analogous to the plasma membrane of other cells, and they provide biochemical confirmation of the suggestion of Behnke (60) that the demarcation membrane arises directly from the plasma membrane of the megakaryocyte.
The isolation of two types of membrane fraction is not unique, two subfractions having been isolated from liver plasma (67) and nuclear (68) membranes and from the plasma membranes of chick fibroblasts (69). Recently, Steck et al. (70) have shown that the erythrocyte membrane may be disrupted to give vesicles that are “inside out” or “outside out,” depending on the nature of the ionic environment. The vesicles have different densities which may arise from differences in the distribution of fixed charges on the inner membrane surface.
Although it has not been possible to determine the origin of the two types of platelet membranes, these observations offer one explanation, since the discontinuities in the outer membrane surface introduced by the canals of the surface-connected system (71) would make the platelet particularly susceptible to the inversions and eversions that are the source of the two types of membrane in the red cell. A second possibility arises from the observations of Nakao and Angrist (72) that the platelet mem- brane stains irregularly with ruthenium red, suggesting a hetero- geneity in composition, and that the interior cytoplasm of the platelet contains many inverted vesicular profiles otherwise identical in appearance with the outer membrane.
The membranes are rich in carbohydrate (7%), which accounts for the release of glycopeptides from the platelet membrane by trypsin, their viral receptor activity, and their reaction with the periodic acid-Schiff stain following solubilization in sodium dodecylsulfate.4 The ratio of glucosamine to galactosamine of 6: 1 differs widely from the value of 2: 1 in erythrocyte mem- branes (73) but is consistent with the high glucosamine content of glycopeptides derived from intact platelets (1, 2). No DNA is found in the membrane fractions, and the small amount found in the lysate and debris is probably of cytoplasmic or mito- chondrial origin (74). RNA is increased a-fold in both the low and high density membrane fractions, a datum that is compatible with protein synthesis occurring in the surface membranes of platelets (57) and L cells (75).
The present work provides a suitable method for the rapid isolation of two types of well characterized platelet membranes in good yield, and it indicates the similarity of these platelet membranes to the plasma membranes of other cells. Studies on the solubilization and characterization of the glycoprotein components of these membranes are in progress.
Acknowledgments-We are indebted to Dr. D. S. Pepper for his guidance in certain of the density gradient techniques and to Mr. Nelson Groh for his assistance in the experiments on isoelectric focusing and gas-liquid chromatography.
REFERENCES
1. PEPPER, D. S., AND JAMIESON, G. A., Biochemistry, 8, (1969).
2. PEPPER, D. S., AND JAMIESON, G. A., Biochemistry, 9, (1970).
3362
3706
3. PEPPER, D. S., AND JAMIESON, G. A., Nature, 219,1252 (1968). 4. DOWBEN. R. M.. Bioloaical membranes, Little, Brown and
Comp&y, Boston, 1967. 5. DALTON, A. J., AND HAQUENAU, F., The membranes, A
Press, New York, 1968. 6. &RCUS, A. J.. New Engl. J. Med., 280, 1213 (1969).
4 A. J. Barber and G. A. Jamieson, unpublished observations.
7. MICHAL, F., AND FIRKIN, B. G.. Annu. Rev. Pharmacol.. 99 95 (1969).
I-
8. HASLAM, R. J., in E. R. JAFFE (Editor), Proceedings of the Twelfth International Congress of the International Societu
” ”
of Hkmatology, Grune and Stratton, Inc., New York, 1968, p. 198.
9. MICHAL, F., Nature, 231, 1253 (1969). 10. MARQUIS, N. R., VIGDAHL, R. L., AND TAVORMINA, P. A.,
Biochem. Biophys. Res. Commun., 36,965 (1969). 11. WILNER, G. D., NOSSEL, H. L., AND LEROY, E. D., J. Clin.
LEGRAND. L.. in E. S. CURTONI. P. L. NL~TTIUZ. AND R. M. TOSI (Editors), Histocompatibikty testing, The Williams and Wilkins Company, Baltimore, 1968, p. 189.
13. SHULMAN, N. R., MARDER, V. J., HILLER, M. C., AND COLLIER, E. M., Progr. Hematol., 4, 222 (1964).
14. MANLEY, G., AND MULLINGER, R. N., Brit. J. Exp. Pathol., 48, 529 (1967).
15. LUNDBERG, A. S., ESTU’ICK, N., AND MERYMAN, H. T., Blood J. Hematol., 23, 545 (1969).
16. MERYMAN, H. T., in G. E. W. WOLSTENHOLME AND M. O’CON- nor (Editors), C&a Foundation Symposium on the Frozen Cell, J. and A: Churchill, Ltd., Loidoi, 1970, p. 51.
17. PERT, J. H., ZUCKER, M. B., LUNDBERG, A., YANKEE, R., AND HENDERSON, E., Voz Sang., 13,119 (1967).
18. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J., J. Biol. Chem., 193, 265 (1951).
19. GORNA~L, A. G., BAR~AWI~L, c‘. J.,’ AND DAVID, M. M., J. Biol. Chem., 177, 751 (1949).
20. KOCH, F. C., AND MCKEEKIN, T. L., J. Amer. Chem. Sot., 46, 2066 (1924).
21. AMINOFF, D., Biochem. J., 81, 384 (1961). 22. WARREN. L.. J. Biol. Chem.. 234. 1971 (1959). 23. GATT, R:, A&D BERMAN, E. k., Anal. B‘ioch&., 16,167 (1965). 24. DUBOIS, M., GILLES, K. A., HAMILTON, J. K., REBERS, P. A.,
AND SMITH, F., Anal. Chem., 28,350 (1956). 25. MUNRO, H. N., AND FLECK, A., Methods Biochem. Anal., 14,
113 (1966). 26. CROFT, D. N., AND LUBRAN, M., Biochem. J., 96,612 (1965). 27. FOLCH, J., LEES, M., AND SLOANE STANLEY, G. H., J. Biol.
Chem., 226, 497 (1957). 28. B~~TTCHER, C. J. F., VAN GENT, C. M., AND PRIES, C., Anal.
Chim. Acta, 24, 203 (1961). 29. BARTLETT, G: R., J. Biol. Chem., 234,466 (1959). 30. KOPACZYK. K.. PERDUE. J.. AND GREEN. D. E.. Arch. Bio-
them. Biophys., 116, 215 (1966). ’ ’ 31. WATSON, D., C&n. Chim. Acta, 6,637 (1960). 32. CHAMBERS, D. A., SALZMAN, E. W., AND NERI, L. L., Arch.
Biochem. Biophys., 119, 173 (1967). 33. WALLACH, D. F. H., AND KAMAT, V. B., in S. P. COLOWICK
AND N. 0. KAPLAN (Editors), Methods in enzymology, Vol. 8, Academic Press, New York, 1966, p. 164.
34. KOERNER, J. F., AND SINSHEIMER, R. L., J. Biol. Chem., 228, 1049 (1957).
35. LINHARDT, K., AND WALTER, K., in H. U. BERGMEYER (Edi- tor), Methods of enzymatic analysis, Academic Press, New York, 1963, p. 783.
36. BIER, M., in S. P. COLOWICK AND N. 0. KAPLAN (Editors), Methods in enzymology, Vol. 1, Academic Press, New York, 1955, p. 632.
37. LANGLEY, T. J., AND JEVONS, F. R., Arch. Biochem. Biophys., 128,312 (1968).
38. TALALAY, P., FISHMAN, W. H., AND HUGGINS, C., J. Biol. Chem., 166,757 (1946).
39. HUGHES, R. C., AND JEANLOZ, R. W., Biochemistry, 3, 1636 (1964).
40. GREEN, D. E., MII, S., AND KOHOUT, P. M., J. Biol. Chem., 217,551 (1956).
41. WROBLEWSKI, R., AND LADTJE, J. S., Proc. Sot. Exp. Biol. Med., 90, 210 (1955).
42. BOSMANN, H. B., HAGOPIAN, A., AND EYLAR, E. H., Arch. Biochem. Biophys., 128,51 (1968).
43. MARTINEK, R. G., BERGER, L., AND BROIDA, D., Clin. Chem., 10, 1087 (1964).
45. SCOTT, R. B., Blood J. Hematol., 30, 321 (1967). 46. Ross, S. W., AND EBERT, R. V., J. Clin. Invest., 38,155 (1959). 47. MADOFF, M. A., EBBE, S., BALDINI, M., J. Clin. Invest., 43,
870 (1964). 48. BULL, B. S., AND ZUCKER, M. G., Proc. Sot. Exp. Biol. Med.,
120, 296 (1965). 49. MARCUS, A. J., ZUCKER-FRANKLIN, D., SAFIER, L. B., AND
ULLMAN, H. L., J. Clin. Invest., 45,14 (1966). 50. BUCKINGHAM, S., AND MAYNERT, E. W., J. Pharmacol. Exp.
Ther., 143,332 (1964). 51. SOLATUNTURI, E., AND PAASONEN, M. K., Ann. Med. Exp.
Biol. Fenn.,.44,427 (1966). 52. SIEGEL. A.. AND L~SCHER. E. F.. Nature. 216,745 (1967). &. WEBER: E.: AND MONOT, k., K&z. Wochenschr., 45,165 (1967). 54. DAY, H. J., HOLMSEN, H., AND HOVIG, T., &and. J. Haematol.,
Suppl., 7,l (1969). 55. WARREN, L., GLICK, M. C., AND NASS, M. K., J. Cell. Physiol.,
68, 266 (1966). 56. MERYMAN. H. T.. Nature. 218, 333 (1968). .&‘. BOOYSE, 6’. M., AND RAF~LSO~, M.‘E., in S. A. JOHNSON AND
M. M. GUEST (Editors), Dynamics of thrombus formation and dissolution, J. B. Lippincott Company, Philadelphia, 1969, p. 149.
58. SABA, S. R., RODMAN, N. F., AND MASON, R. G., Amer. J. Pathol., 66,225 (1969).
61. LANSING, A. I., BELKHODE, M. L., LYNCH, W. E., AND LIE- BERMAN, I., J. Biol. Chem., 242,1772 (1967).
62. EMMELOT, P., Bos, C. J., BENEDETTI, E. L., AND R~MKE, P., Biochim. Biophys. Acta, 90, 126 (1964).
63. COLEMAN, R., AND FINEAN, J. B., Biochim. Biophys. Acta, 126, 197 (1966).
64. PFLEGER, R. C., ANDERSON, N. G., AND SNYDER, F., Biochem- istry, 7, 2826 (1968).
65. WEINSTEIN, D. B., in L. A. MANSON (Editor), Biologicat properties of the mammalian surface membrane, The Wistar Institute Press, Philadelphia, 1968, p. 17.
66. BAKERMAN, S., AND WASEMILLER, G., Biochemistry, 6, 1100 (1967).
67. EVANS, W. H., Biochem. J., 116,833 (1970). 68. KASHNIG, D. M., AND KASPER, C. B., J. Biol. Chem., 244,
3786 (1969). 69. PERDUE, J. F., AND SNEIDER, J., Biochim. Biophys. Acta, 196,
125 (1970’). 70. STECK,~ T. i., WEINSTEIN, R. S., STRAUS, J. H., AND WALLACH,
D. F. H.. Science. 168. 2.55 0970). 71. HOVIG, T.,‘Ser. Hakmatol., 1,‘3 (1968). 72. NAKAO, K., AND ANQRIST, A. A., Nature, 217,960 (1968). 73. ROSENBERQ, S. A., AND GUIDOTTI, G., J. Biol. Chem., 243,
1985 (1968). 74. NASS, M. M. K., NASS, S., AND AFZELIUS, B. A., Exp. Cell
Res., 37, 516 (1965). 75. GLICK, M. C., AND WARREN, L., Proc. Nat. Acad. Xci. U. S. A.,