This work highlights crucial features of a novel and still not completely understood population of mesenchymal progenitors isolated form endometrial decidual tissue.
Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 901821 14 pageshttpdxdoiorg1011552013901821
Research ArticleIsolation Characterization and Transduction ofEndometrial Decidual Tissue Multipotent MesenchymalStromalStem Cells from Menstrual Blood
Filippo Rossignoli1 Anna Caselli12 Giulia Grisendi1 Serena Piccinno1
Jorge S Burns1 Alba Murgia1 Elena Veronesi1 Pietro Loschi3 Cristina Masini1
Pierfranco Conte1 Paolo Paolucci4 Edwin M Horwiz5 and Massimo Dominici1
1 Division of Oncology Department of Medical and Surgical Sciences of Children amp AdultsUniversity Hospital of Modena and Reggio Emilia Via del Pozzo 71 41100 Modena Italy
2 Department of Internal Medicine and Oncology University of Bari Aldo Moro Bari Italy3 Unit of Plastic Surgery Department of Medical and Surgical Sciences of Children amp AdultsUniversity Hospital of Modena and Reggio Emilia Via del Pozzo 71 41100 Modena Italy
4Division of Pediatric Oncology Hematology and Marrow Transplantation Department of Medical and Surgical Sciences ofChildren amp Adults University Hospital of Modena and Reggio Emilia Via del Pozzo 71 41100 Modena Italy
5 Division of Oncology Childrenrsquos Hospital of Philadelphia Philadelphia PA 19104 USA
Correspondence should be addressed to Massimo Dominici massimodominiciunimoreit
Received 2 November 2012 Revised 22 January 2013 Accepted 28 January 2013
Academic Editor Thomas Skutella
Copyright copy 2013 Filippo Rossignoli et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Mesenchymal stromalstem cells (MSCs) reveal progenitor cells-like features including proliferation and differentiation capacitiesOne of the most historically recognized sources of MSC has been the bone marrow while other sources recently include adiposetissue teeth bone muscle placenta liver pancreas umbilical cord and cord blood Frequently progenitor isolation requirestraumatic procedures that are poorly feasible and associated with patient discomfort In the attempt to identify amore approachableMSC source we focused on endometrial decidual tissue (EDT) found within menstrual blood Based also on recent literaturefindings we hypothesized that EDT may contain heterogeneous populations including some having MSC-like features Thuswe here sought to isolate EDT-MSC processing menstrual samples from multiple donors Cytofluorimetric analyses revealedthat resulting adherent cells were expressing mesenchymal surface markers including CD56 CD73 CD90 CD105 and CD146and pluripotency markers such as SSEA-4 Moreover EDT-MSC showed a robust clonogenic potential and could be largelyexpanded in vitro as fibroblastoid elements In addition differentiation assays drove these cells towards osteogenic adipogenicand chondrogenic lineages Finally for the first time we were able to gene modify these progenitors by a retroviral vector carryingthe green fluorescent protein From these data we suggest that EDT-MSC could represent a new promising tool having potentialwithin cell and gene therapy applications
1 Introduction
Mesenchymal stromalstem cells (MSCs) are adult progenitorcells isolated from several human adult and perinatal tissues[1 2] While the acronymMSC seems to equate the biologicalproperties of these useful progenitors the possibility to isolatethese cells from different tissue has been outlining common
features combined with source-specific peculiarities that arestill under investigation
MSC demonstrated a positive impact in several patholog-ical conditions [1 3 4] exerting their therapeutic functionson damaged tissues through different mechanisms includingdifferentiation intomature cells and largely obscure paracrineeffects [5] Moreover MSCs have also been investigated for
2 BioMed Research International
their possible use to deliver wild-type or gene modification-induced bioactive molecules with promising but still unde-fined influence in cancer models [6ndash9] In the light ofthese findings it appears reasonable to propose that a tissuesource of MSC could be deemed relevant and useful bydetermining if it provides cells with varied differentiationpotential and distinct cytokine profiles that may indicatean advantageous role in the interaction with tumors [110] Therefore understanding these features from distinctMSC tissue sources shall be a primary objective to effi-ciently translate these cells into different clinical applica-tions
One most recognized source of MSC has been the bonemarrow obtained from iliac crest and more recently MSCprogenitors have been isolated from lipoaspirates and othertissue sources including teeth bone muscle placenta liverpancreas umbilical cord and cord blood [11] In the majorityof these cases especially the autologous sources tissue iso-lation requires traumatic procedures sometimes linked withpatientrsquos discomfort
In the attempt to identify a different and more approach-able MSC source for cell and gene therapies we focused onendometrial decidual tissue (EDT) obtained from menstrualblood Previous cloning studies of isolated human endome-trial cells provided early evidence of rare clonogenic mes-enchymal cells representing approximately 1 of endome-trial cells suspension obtained by uterine tissue digestionafter hysterectomy [12] These endometrial stromal elementsdemonstrated properties similar to bonemarrow and adiposetissue MSC including substantial self-renewal ability in vitrohigh proliferative potential and multilineage differentiation[13 14] However obtaining these cells directly from endo-metrium either after hysterectomy or by biopsy still impliesinvasive procedures
In order to evaluate the possibility of obtaining multi-potent cells from the uterus by a non-invasive and repro-ducible manner researchers started to analyze shed men-strual blood and tissue in the attempt to identify menstrualblood-derived MSC [15 16] These pilot studies confirmedwhat was already known for endometrial cells obtained fromhysterectomy suggesting that endometrial stemprogenitorcells may be shed in menstrual blood
Following these early findings and by different isolationsteps this study characterizes EDT as a novel tissue sourceof MSC with regards to possible applications in regenerativemedicine and gene therapyMSCs fromdifferent sources havebeen investigated for their possible uses as tumor-specificdelivery vehicles for suicide genes oncolytic viruses or sec-reted therapeutic proteins [8] The mechanisms throughwhich MSCs are considered to home to tumors are not com-pletely clear but seem to be dependent on biological proper-ties of tumor microenvironment as well as the native tropismof selected MSC and also on the experimental proceduresused [10 17]Thanks to genetic manipulation techniques thisnatural tropism has been recently and convincingly exploitedto transform MSC in ldquobulletsrdquo capable to deliver oncolyticviruses or various anticancer agents directly into tumor sites[8] The possibility of having noninvasive procedures toobtain autologous progenitor cells although with a gender
limitation paves the way to a more intense research activityaimed to deeply characterize MSC from a different source forselected biomedical applications
2 Materials and Methods
21 Cell Procurement and Processing Menstrual blood wascollected from healthy female volunteer donors (119899 = 3) dur-ing the first few days of the cycle Written consent wasobtained from each donor and the Local Ethical Committeeapproved cells donation for research purposes Each donorhas been endowed with a menstrual cup (DivaCup DivaInternational San Francisco CA USA) to collect bloodwhich was transferred in phosphate buffered saline (PBSPAA Laboratories Pasching Austria) with 1 penicillinstreptomycin (10000UmL Penicillin 10mgmL Strepto-mycin in 09 NaCl solution PAA Laboratories) 35mgmLfluconazole (Diflucan Pfizer New York NY USA) and hep-arin (500UmL Sigma St Louis MO USA) Samples weremaintained at 4∘C for 24ndash48 h after procurement until reach-ing the processing laboratory
The endometrial tissue if present was discarded and theremaining blood was homogenized by 20 passages through a19G needle using a 10mL syringe Cell suspension was thencultured as below reported In addition two alternative iso-lation protocols involving separation of the corpuscular frac-tion of blood through normal or density-gradient centrifu-gations were initially introduced However these latter twoapproaches were discontinued because of poor isolation effi-ciency (data not shown)
22 Cell Culture Isolated cells were seeded into culture flasksto obtain an adherent fraction by adding 120572MEM (Gibco)1 L-glutamine (200mM in 085 NaCl solution LonzaVerviers Belgium) 1penicillinstreptomycin (PAALabora-tories) and 10 fetal bovine serum (FBS PAA Laboratories)In addition a serum-deprived medium (Quantum 333) with1 penicillinstreptomycin was also introduced (all fromPAA Laboratories) Cells were cultured for 7 days washedwith PBS (PAA Laboratories) to remove the nonadherentfraction and fresh medium was added At confluence cellswere detached by trypsin (trypsin 005 EDTA 002 inPBS EuroClone Milan Italy) and subcultured at a densityof 6000 cellscm2 until functional assays All flasks were incu-bated at 37∘C 5CO
2 andmediumwas changed every other
day
23 Clonogenic Assay Adherent cells out of passage 1 (P1) orpassage 2 (P2) were seeded at clonal density of 100 cellscm2in 120572MEM (Gibco) 1 L-glutamine (Lonza) 1 penicillinstreptomycin (PAA Laboratories) and 10 FBS (PAA Labo-ratories) Colony formation was monitored daily On day 10cells were fixed with cooled absolute methanol for 2 minutesand stained for 5 minutes with 1 crystal violet aqueoussolution (Sigma) Colonies with more than 50 cells were thencounted as originally described for marrow MSC [18] Eachassaywas repeated in triplicate and cloning efficiency (119864) wascalculated as 119864 = (n clonescells seeded) sdot 100
BioMed Research International 3
24 Proliferation Assay Adherent cells were seeded at adensity of 6000 cellscm2 in 120572MEM (Gibco) 1 L-glutamine(Lonza) 1 penicillinstreptomycin (PAA Laboratories) and10 FBS (PAA Laboratories) in different flasks for thefollowing time points 24 48 72 and 96 hours At each timepoint cells were trypsinized and counted Data obtainedwereplotted as number of harvested cells (119910) against hours ofculture (119909) and the exponential growth curve was generatedusing GraphPad Prism software (GraphPad Software Incversion 500) The doubling time (119879) was obtained from thegrowth constant (119896) of the exponential equation 119910 = 119886 sdot 119890119896119909where 119896 = ln 2119879
25 Senescence-Associated 120573-Galactosidase Staining To as-sess cell senescence the activity of 120573-galactosidase (120573-gal) atpH 6 was evaluated 120573-gal expression is a feature of senescentcells [19] According to Senescence 120573-galactosidase StainingKit (Cell Signaling Technology Beverly MA USA) growthmedium was removed and cells were washed with PBS (PAALaboratories) Cells were then incubated for 15 minutes witha fixative solution (20 formaldehyde 2 glutaraldehyde in10x PBS) and washed twice with PBS (PAA Laboratories)Color development was obtained by incubation overnightat 37∘C with the provided staining solution (40mM citricacidsodium phosphate pH 60 150mMNaCl 2mMMgCl
2
5 nM potassium ferricyanide and 1mgmL X-gal in DMSO)Plateswere then observed bymicroscopy for the developmentof blue color
26 FACSAnalyses Adherent cells were harvested for surfaceantigen analysis Briefly cells were detached from plastic sup-port by trypsin (EuroClone) counted and aliquoted in FACSanalyses polypropylene tubes (05ndash1 sdot 106 cellstube) (VWRMilan Italy) EDT-MSCs were subsequently incubated inblocking buffer (100 120583L each 05ndash1 sdot 106 cells) containingDulbeccorsquos Modified Eaglersquos Medium (DMEM Gibco) 10FBS (PAA Laboratories) and 01M sodium azide and humanimmunoglobulin G (both from Sigma) and incubated for201015840 on ice After a PBS (PAA Laboratories) washing stepcells were resuspended in PBS (PAA Laboratories) with 05bovine serum albumin (BSA Sigma) and stained on ice andin the dark for 301015840 with the following monoclonal antibod-ies APC-anti-CD45 FITC-anti-HLADR PE-anti-CD34 PE-anti-CD14 and FITC-anti-CD56 (all from Becton Dickin-son Franklin Lakes NJ USA) PE-anti-CD31 (BioLegend)APC-anti-CD90 PE-anti-SSEA-4 (both from eBioscienceSan Diego CA USA) FITC-anti-CD105 PE-anti-CD73 (allfrom BD Pharmingen) APC-anti-CD146 (Miltenyi Biotec)In all the experiments the corresponding isotype-matchedantibodies were used as negative controls (BD Pharmingenand Becton Dickinson) Data were collected using a FACSAria III flow cytometer (BD Biosciences) and analyzed onFACS Diva software (BD Biosciences)
27 Multilineage Differentiation Assays To assess in vitrodifferentiation capacities adherent cells after P2 were cul-tured in specific induction media Media were changed everyother day and undifferentiated controls were concurrentlycultured in 120572MEM (Gibco) 1 L-glutamine (Lonza) 1
penicillinstreptomycin (PAA Laboratories) and 10 FBS(PAA Laboratories) for the same incubation time Each assaywas performed in triplicate
Adipogenic osteogenic and chondrogenic differentia-tions were performed as previously reported [20] Briefly forosteogenic differentiation cells were seeded at the density of10000 cellscm2 and maintained in the growth medium untilconfluence Culture medium was then substituted with theinduction one composed by 120572MEM (Gibco) supplementedwith 10 nM dexamethasone (Sigma) 10mM 120573-glycerol phos-phate (Sigma) 01mM L-ascorbic acid-2-phosphate (Sigma)10 defined FBS (Hyclone Logan UTUSA) 1 L-glutamine(Lonza) and 1 penicillinstreptomycin (PAA Laboratories)Induction was maintained for 14 days and from the seventhday 100 ngmL BMP-2 (Peprotech Rocky Hill NJ USA)was added Confirmation of osteogenic differentiation wasperformed through Alizarin Red staining combined by real-time qPCR analysis (described below) At the end of theinduction period culture wells were washed briefly witha buffer solution containing 20mM Tris-HCl and 150mMNaCl in water Cells were then fixed and dried with cooledabsolute methanol for 2 minutes After a washing step withddH2O cells were stained by 15 Alizarin Red aqueous
solution pH 40ndash42 (Sigma) for 5 minutes and washed againfirstwith ddH
2Oand thenwith PBS (PAALaboratories) for 15
minutes The last step consisted of dehydration using cooledabsolute ethanol for 2 minutes followed by microscopicobservation
For adipogenic differentiation EDT-MSCswere seeded atthe density of 10000 cellscm2 and maintained in the growthmedium until confluence The medium was then substi-tuted with the induction one composed of 120572MEM (Gibco)supplemented with 1120583M dexamethasone (Sigma) 60 120583Mindomethacin (Sigma) 10120583M insulin (Sigma) 05mM 3-isobutyl-1-methylxanthine (IBMX Sigma) 10 rabbit serum(EuroClone) 5 horse serum (EuroClone) 1 L-glutamine(Lonza) and 1 penicillinstreptomycin (PAA Laboratories)Induction lasted 10 days and confirmation of differentiationwas achieved through Oil Red O staining as follows Culturewells were washed briefly with PBS (PAA Laboratories) Cellswere fixed with vapors of 37 formaldehyde (Sigma) for 10minutes and then washed with water for 2 minutes Stainingwas obtained adding an Oil Red O solution (10mgmL OilRed O in ethanol 70 and acetone all from Sigma) into thewells for 3 minutes Excessive stain was removed by washingwith water and cells were then counterstained by Harrishematoxylin (Bio-Optica Milan Italy) for 30 seconds
Finally for chondrogenic differentiation 2ndash5 sdot 105 EDT-MSCs were aliquoted in a 2mL tube and then centrifugedat 1200 rpm for 10 minutes Cells were maintained pelletedin the growth medium at 37∘C with the plug opened After2 days of incubation tubes were centrifuged again and themedium was substituted with the induction one composedby 120572MEM (Gibco) supplemented with 100 nM dexametha-sone (Sigma) 200120583M L-ascorbic acid-2-phosphate (Sigma)100 120583gmL sodium pyruvate (Biochrom AG Berlin Ger-many) 40 120583gmL proline (Sigma) 1x ITS+ premix (BD Bio-sciences San Jose CA USA) 10 ngmL TGF-120573 (Peprotech)05 120583gmL BMP-6 (Peprotech) 1 L-glutamine (Lonza)
ALP alkaline phosphatase COL1A2 collagen 1A2 OC osteocalcin
and 1 penicillinstreptomycin (PAA Laboratories) Beforeand after each medium change tubes were centrifuged at1200 rpm for 10 minutes During the incubation period cellsremained as pellet with the tube plugs opened Inductionlasted 21 days and specimens were fixed for 1 hour in10 formaldehyde (Sigma) and then dehydrated by serialpassages into ethanol at increasing concentrations from 70to 100 Samples were then included into paraffin blocks andcut in slices on microscope slides for Alcian Blue stainingSlides were deparaffinized with the Histo-C cleaning agent(Celltech Turin Italy) and rehydrated through passagesinto a decreasing concentration alcoholic ladder (from 100ethanol to 70ethanol) Sample sectionswere then incubatedwith a 05mgmLHyaluronidase (Sigma) in buffer phosphatesolution (8 gL NaCl 2 gL NaH
2PO4 and 03 gL Na
2HPO4)
with 10mgmL BSA (Sigma) Slides were washed in water for5 minutes and then immersed in a 3 acetic acid solutionfor few seconds Staining with 10mgmLAlcian Blue solutionin 3 acetic acid (pH 25 Sigma) lasted 30 minutes andafter a washing step in water samples were counterstainedfor 5 minutes with nuclear fast red solution (Sigma) and thenwashed in water
28 Real-Time qPCR Total cellular RNA was isolated fromosteogenic committed and uncommitted cells using Trizolreagent method (Invitrogen) according to manufacturerrsquosinstructions Samples were concentrated by ethanol pre-cipitation and suspended in RNase-free water RNA quan-tity was assessed by spectrophotometry (DU730 UVVISSpectrophotometer Beckman Coulter Milan Italy) A 2120583galiquot was reverse transcribed into cDNA using Rever-tAid First Strand cDNA Synthesis Kit (Fermentas) by oligo(dT)18
primers in a final volume of 20 120583L Reactions wereperformed at 42∘C for 1 h with a final step at 70∘C for 5minutes cDNA was then used to determine the osteoblast-associated gene expression by quantitative real-time PCRtechnique using Step One Real-Time PCR System ThermalCycling Block (Applied Biosystems Foster City CA USA)Each sample was compared with noninduced control forthe expression of alkaline phosphatase collagen 1A2 andosteocalcin 120573-actin was used as a reference gene All primerswere purchased from Integrated DNA Technologies andsequences are reported in Table 1 PCR was performed withFast SYBR Green Master Mix (Applied Biosystems) whichuses AmpliTaq Fast DNA Polymerase SYBR Green I dye
to detect double-stranded DNA and ROX dye as a passiveinternal reference Reaction proceeded through an initial stepat 95∘C for 2010158401015840 followed by 40 cycles of denaturation (310158401015840 at95∘C) annealing and extending (3010158401015840 at 60∘C)The final stagecomprises the analysis of themelt curve through a denaturingstep (1510158401015840 at 95∘C) followed by annealing (11015840 at 60∘C) andramping to 95∘C with 03∘C incrementstep Levels of mRNAfor tested genes were quantified using ΔΔCT method andnormalized against human 120573-actin as a housekeeping geneData have been analyzed by StepOne software (version 21Life Technologies Corporation Carlsbad CA USA)
29 Gene Modification of EDT-MSC Cultured cells wereinfected by a bicistronicmurine stem cell virus-derived retro-viral vector (pMIGR1) encoding for green fluorescent Protein(GFP) Retrovirus production was performed by the FLYRDpackaging cell lines as published by Marx et al [21] FLYRDcells were seeded in a T175 flask with 10mL of medium com-posed by DMEM (Gibco) supplemented with 10 definedFBS (Hyclone) Cell supernatant was collected and filtered(PES) with a 045 120583m filter and EDT-MSC were incubatedfor 6 hrs with 5mL culture medium composed by 120572MEM(Gibco) 1 L-glutamine (Lonza) 1 penicillinstreptomycin(PAA Laboratories) and 10 FBS (PAA Laboratories) withthe addition of 5mL viral particles-containing supernatantand 6 120583gmL polybrene (Sigma) Cells were then washed byPBS (PAA Laboratories) and culture medium was changedThe infection step was repeated for three consecutive daysat which time cells were evaluated at FACS Aria III flowcytometer (BD Biosciences) for GFP protein expression 7-amino-actinomycin D (7AAD) staining was also performedto evaluate mortality after transduction Cells were evaluatedby FACS Aria III (BD Biosciences) and data were analyzedusing FACS Diva software (BD Biosciences)
210 Statistics Data are expressed as average values andanalyses were performed by GraphPad Prism software t-testwas considered as significant with 119875 value lt005
3 Results
31 EDT-Derived Cells Are In Vitro Heterogeneous but RetainPredominant MSC Features Adherent cells isolated frommenstrual blood initially displayed in vitro fibroblast shapemorphology in both 120572MEMwith 10 FBS and Quantum 333
BioMed Research International 5
(Figures 1(a)-1(b)) However prolonged cultures in 120572MEMwith 10 FBS showed a significant better growth perfor-mance and this medium was then preferentially used forall subsequent functional analyses Adherent elements hadtypical mesenchymal aspect being fibroblastoid-like spindle-shaped cells with an elongated cytoplasm Within this pop-ulation it was also possible to identify more infrequentcell clusters with a distinct morphology These appearedas endothelial-like sometimes binucleated cells forming amonolayer with polygonal shape (Figure 1(c)) However wedid not confirm the nature of this population because theirpresence disappeared after very early passages and becausethey were not the subjects of this study
32 EDT-Derived Cells Show Robust Clonogenic and Prolif-erative Potential Having observed the fibroblast shape ofEDT isolated cells we then focused on their clonogenic andproliferative potential We observed a high clonal efficiencywith an average of 141 (108ndash179) of the seeded cellsable to generating colonies This result indicates that themenstrual-derived cell population contains a large fractionof actively cycling cells with signficant clonogenic potential(Figure 2(a)) Of interests colonies did not appear homoge-nous and we were able to identify at least two kinds ofmorphologies On the one hand densely populated clonesconstituted by small size cells (Figures 2(b)-2(c)) and onthe other smaller cell clusters with elements having a largecytoplasm with an evident cytoskeleton (Figures 2(d)-2(e))
Isolated and expanded cells also demonstrated surpris-ingly low doubling time with an average value of 276 hours(219ndash330) (Figure 3) The high number of passages thatthese cells were able to reach further supported this remark-able growth property The 120573-galactosidase staining per-formed to evaluate senescent cells showed that cells cul-tured in Quantum 333 underwent senescence at passage 17while those cultured in 120572MEM supplemented with 10 FBSreached passage 26 before growth arrest (Figures 4(a)ndash4(d))Together these data suggest how plastic-adherent fibroblastshaped cells from EDT retain a strong proliferative potentialcomparable or even superior to MSC from other sour-ces
33 EDT-Derived Cells Display MSC Phenotypic FeaturesHaving evaluated the proliferation potential and to morecarefully define the MSC nature of isolated and expandedcells we assessed their antigen expression profile EDT-derived cells express typical MSC markers (Figures 5 and 6)and in particular more than 90 of tested cells were posi-tive for CD90 and CD73 constituting main features of MSCfrom other sources [22] We also observed that more than80 of cells expressed CD146 an adhesion molecule relatedbut not restricted to MSC and also expressed by endothe-lial cells [1] The levels of CD45 HLADR CD31 and CD14assessed early in culture and commonly used to distinguishMSC from hematopoietic and endothelial cells were below2 in most cases In sample 1 we observed a slightly increasein the CD45+ fraction suggesting the presence of hematopoi-etic elements which might have been isolated together withEDT-MSC Other markers were also considered such as
CD56 whose positivity was extremely variable by up to 45Collectively EDT-MSC from different donors showed thesame overall trend in markers expression (Figure 6) despitefor some markers such as for CD56 CD105 and CD146the variability has been considerably high The phenotypicheterogeneity was also confirmed by analysis of physicalparameters by FACS Forward scatter versus side scatter plotwas very dispersed and it was impossible to identify consist-ent cell groupswith similar physical parameters Interestinglywe have to report a consistent small fraction (gt10) of SSEA-4 positive cells that reached 194 suggesting the expressionof a pluripotency marker in this MSC type
34 EDT-MSCs Are Precursors of Three Mesenchymal TissuesIn Vitro FACS analyses were then followed by assays aim-ing to assess EDT-MSC multipotency We first focused onadipose commitment and after 10 days of adipogenic induc-ing cocktail EDT-MSCs were able to differentiate into vacu-ole-producing elements The Oil Red staining confirmedthe lipid content of those cells indicating the fat-producingability of isolated and induced cells (Figure 7(a)) Osteogenicmedium was then applied for 14 days and cells underwent toosteoblastic commitment forming a compact calcifiedmatrixwith calcium deposits confirmed by specific Alizarin Redstaining (Figure 7(b)) Finally after 21 days of chondrogenicinduction pellets of EDT-MSCs were included into paraffinblocks and Alcian Blue highlighted the sulfated proteogly-cans expression typical of the cartilaginousmatrix while FastRed staining revealed the nuclei of resident chondrocytesderived from EDT-MSC (Figure 7(c)) Since a main focus ofour research group is bone regeneration we then coupledthe cytochemical assay of osteogenic induced EDT-MSCwithqPCR to further confirm bone commitment As seen inFigure 8 all tested genes demonstrated an increased expres-sion in induced samples versus noninduced controls In part-icular in osteoblast-induced cells the expression of alkalinephosphatase resulted 45 times higher (119875 lt 00001) whilemore modest increases were observed for collagen 1A2 (29times 119875 lt 005) and osteocalcin (21 times 119875 lt 005) Thesefindings together with positive Alizarin staining reinforcedthe evidence for commitment of induced EDT-MSC to func-tional osteoblasts
35 EDT-MSC Can Be Efficiently Gene Modified To explorewhether ex vivo expanded EDT-MSC could be geneticallymanipulated for future gene delivery approaches cells wereincubated with supernatants containing retroviral particlescarrying the GFP gene As shown in Figure 9 RD114 pseu-dotyped retroviral particles were able to efficiently and stablytransduce EDT-MSCwith levels greater than 80 suggestinghow this MSC type could be suitable for gene deliveryapproaches
4 Discussion
SeveralMSC types have been obtained starting from differentsources here we have isolated a population of mesenchymalprogenitors from menstrual EDT and characterized themboth at morphological and molecular levels investigating
6 BioMed Research International
(a) (b)
(c)
Figure 1 In vitro adherent cells from decidual tissues (a) Representative photomicrograph of spindle-shaped adherent cells isolated by120572MEM and 10 FBS (b) Cells isolated in vitro by serum-deprived medium (Quantum 333) at early passages (c) Another population of cellswas in vitro isolated contextually with EDT-MSCThese elements share similarities with endothelial cells forming a monolayer of polygonalsometimes binucleated cells Original magnification 100x
their ability to proliferate to differentiate and to be genemodified EDT-MSCs appear as a heterogeneous cells pop-ulation that can be safely and easily isolated by noninvasivemanner providing an expandable source of cells In culturethey show mesenchymal morphology and although somestudies suggest they could derive from bone marrow [23]EDT-MSCs are generally considered to originate from theshedding of endometrial stemprogenitor cells which aremainly resident both in the basalis layer and partially in thefunctionalis area of endometrium [24]
The presence of a uterine population of progenitor cellswould be consistent with the high tissue turn-over after eachmenstrual cycle However it is still uncertain howmany sub-populations of progenitors are present in the endometriumand what are their specific properties Therefore in this pilotstudy we began by addressing the existence of mesenchymalprogenitors in the decidual endometrial tissue
The initial in vitro approach was performed to evaluatethe clonogenic potential of these cells EDT-MSC seeded atclonal density revealed the presence of two distinct colonytypes which closely resembled the stromal cell clones isolatedby Chan et al from endometrium after hysterectomy [12]One was composed of smaller and densely packed cells andthe other was composed of larger and sparser elements Thisfinding seems peculiar for EDT-MSC and differs from col-onies established from BM-MSC that appear larger and
characterized by a central nucleus of cells surrounded by acrown of more sparse ones [18] Next to qualitative evalua-tions of the colonies our data suggest an average EDT-MSCclonogenic efficiency of 14 This value is higher than whatis described for BM-MSC and more similar or even superiorto what reported for AT-MSC that retained around 10 ofclonogenic precursors [25] (Table 3) In addition our EDT-MSC population seemed to have a higher clonogenic effi-ciency compared to those described by other groups dealingwith endometrial stromal cells whose values ranged from 1to 11 [12 24 26] While this may be due to donor-relatedissues such as age in vitro technical aspects including harvest-ing and preservation procedures that may also drive thesedifferences Dimitrov et al observed a tendency for these cellsto decrease in clonal efficiency with the increase of seedingdensity [26] Similarly other culture conditions may generatebetter performing EDT-MSC In this pilot study we testedtwo different growthmedia such as120572MEMwith 10FBS anda serum deprived medium Although these cells apparentlydid not shown particular nutritional requirements serum-containing medium proved to be the best choice ensuring abetter morphological appearance and an improved longevityPrevious studies reported the use of different media such asDMEMF-12 and Chang complete media often supplementedwith calf serum hormones and growth factors [12 14 26 27]This wide range of culture conditions is likely to impact the
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
2 BioMed Research International
their possible use to deliver wild-type or gene modification-induced bioactive molecules with promising but still unde-fined influence in cancer models [6ndash9] In the light ofthese findings it appears reasonable to propose that a tissuesource of MSC could be deemed relevant and useful bydetermining if it provides cells with varied differentiationpotential and distinct cytokine profiles that may indicatean advantageous role in the interaction with tumors [110] Therefore understanding these features from distinctMSC tissue sources shall be a primary objective to effi-ciently translate these cells into different clinical applica-tions
One most recognized source of MSC has been the bonemarrow obtained from iliac crest and more recently MSCprogenitors have been isolated from lipoaspirates and othertissue sources including teeth bone muscle placenta liverpancreas umbilical cord and cord blood [11] In the majorityof these cases especially the autologous sources tissue iso-lation requires traumatic procedures sometimes linked withpatientrsquos discomfort
In the attempt to identify a different and more approach-able MSC source for cell and gene therapies we focused onendometrial decidual tissue (EDT) obtained from menstrualblood Previous cloning studies of isolated human endome-trial cells provided early evidence of rare clonogenic mes-enchymal cells representing approximately 1 of endome-trial cells suspension obtained by uterine tissue digestionafter hysterectomy [12] These endometrial stromal elementsdemonstrated properties similar to bonemarrow and adiposetissue MSC including substantial self-renewal ability in vitrohigh proliferative potential and multilineage differentiation[13 14] However obtaining these cells directly from endo-metrium either after hysterectomy or by biopsy still impliesinvasive procedures
In order to evaluate the possibility of obtaining multi-potent cells from the uterus by a non-invasive and repro-ducible manner researchers started to analyze shed men-strual blood and tissue in the attempt to identify menstrualblood-derived MSC [15 16] These pilot studies confirmedwhat was already known for endometrial cells obtained fromhysterectomy suggesting that endometrial stemprogenitorcells may be shed in menstrual blood
Following these early findings and by different isolationsteps this study characterizes EDT as a novel tissue sourceof MSC with regards to possible applications in regenerativemedicine and gene therapyMSCs fromdifferent sources havebeen investigated for their possible uses as tumor-specificdelivery vehicles for suicide genes oncolytic viruses or sec-reted therapeutic proteins [8] The mechanisms throughwhich MSCs are considered to home to tumors are not com-pletely clear but seem to be dependent on biological proper-ties of tumor microenvironment as well as the native tropismof selected MSC and also on the experimental proceduresused [10 17]Thanks to genetic manipulation techniques thisnatural tropism has been recently and convincingly exploitedto transform MSC in ldquobulletsrdquo capable to deliver oncolyticviruses or various anticancer agents directly into tumor sites[8] The possibility of having noninvasive procedures toobtain autologous progenitor cells although with a gender
limitation paves the way to a more intense research activityaimed to deeply characterize MSC from a different source forselected biomedical applications
2 Materials and Methods
21 Cell Procurement and Processing Menstrual blood wascollected from healthy female volunteer donors (119899 = 3) dur-ing the first few days of the cycle Written consent wasobtained from each donor and the Local Ethical Committeeapproved cells donation for research purposes Each donorhas been endowed with a menstrual cup (DivaCup DivaInternational San Francisco CA USA) to collect bloodwhich was transferred in phosphate buffered saline (PBSPAA Laboratories Pasching Austria) with 1 penicillinstreptomycin (10000UmL Penicillin 10mgmL Strepto-mycin in 09 NaCl solution PAA Laboratories) 35mgmLfluconazole (Diflucan Pfizer New York NY USA) and hep-arin (500UmL Sigma St Louis MO USA) Samples weremaintained at 4∘C for 24ndash48 h after procurement until reach-ing the processing laboratory
The endometrial tissue if present was discarded and theremaining blood was homogenized by 20 passages through a19G needle using a 10mL syringe Cell suspension was thencultured as below reported In addition two alternative iso-lation protocols involving separation of the corpuscular frac-tion of blood through normal or density-gradient centrifu-gations were initially introduced However these latter twoapproaches were discontinued because of poor isolation effi-ciency (data not shown)
22 Cell Culture Isolated cells were seeded into culture flasksto obtain an adherent fraction by adding 120572MEM (Gibco)1 L-glutamine (200mM in 085 NaCl solution LonzaVerviers Belgium) 1penicillinstreptomycin (PAALabora-tories) and 10 fetal bovine serum (FBS PAA Laboratories)In addition a serum-deprived medium (Quantum 333) with1 penicillinstreptomycin was also introduced (all fromPAA Laboratories) Cells were cultured for 7 days washedwith PBS (PAA Laboratories) to remove the nonadherentfraction and fresh medium was added At confluence cellswere detached by trypsin (trypsin 005 EDTA 002 inPBS EuroClone Milan Italy) and subcultured at a densityof 6000 cellscm2 until functional assays All flasks were incu-bated at 37∘C 5CO
2 andmediumwas changed every other
day
23 Clonogenic Assay Adherent cells out of passage 1 (P1) orpassage 2 (P2) were seeded at clonal density of 100 cellscm2in 120572MEM (Gibco) 1 L-glutamine (Lonza) 1 penicillinstreptomycin (PAA Laboratories) and 10 FBS (PAA Labo-ratories) Colony formation was monitored daily On day 10cells were fixed with cooled absolute methanol for 2 minutesand stained for 5 minutes with 1 crystal violet aqueoussolution (Sigma) Colonies with more than 50 cells were thencounted as originally described for marrow MSC [18] Eachassaywas repeated in triplicate and cloning efficiency (119864) wascalculated as 119864 = (n clonescells seeded) sdot 100
BioMed Research International 3
24 Proliferation Assay Adherent cells were seeded at adensity of 6000 cellscm2 in 120572MEM (Gibco) 1 L-glutamine(Lonza) 1 penicillinstreptomycin (PAA Laboratories) and10 FBS (PAA Laboratories) in different flasks for thefollowing time points 24 48 72 and 96 hours At each timepoint cells were trypsinized and counted Data obtainedwereplotted as number of harvested cells (119910) against hours ofculture (119909) and the exponential growth curve was generatedusing GraphPad Prism software (GraphPad Software Incversion 500) The doubling time (119879) was obtained from thegrowth constant (119896) of the exponential equation 119910 = 119886 sdot 119890119896119909where 119896 = ln 2119879
25 Senescence-Associated 120573-Galactosidase Staining To as-sess cell senescence the activity of 120573-galactosidase (120573-gal) atpH 6 was evaluated 120573-gal expression is a feature of senescentcells [19] According to Senescence 120573-galactosidase StainingKit (Cell Signaling Technology Beverly MA USA) growthmedium was removed and cells were washed with PBS (PAALaboratories) Cells were then incubated for 15 minutes witha fixative solution (20 formaldehyde 2 glutaraldehyde in10x PBS) and washed twice with PBS (PAA Laboratories)Color development was obtained by incubation overnightat 37∘C with the provided staining solution (40mM citricacidsodium phosphate pH 60 150mMNaCl 2mMMgCl
2
5 nM potassium ferricyanide and 1mgmL X-gal in DMSO)Plateswere then observed bymicroscopy for the developmentof blue color
26 FACSAnalyses Adherent cells were harvested for surfaceantigen analysis Briefly cells were detached from plastic sup-port by trypsin (EuroClone) counted and aliquoted in FACSanalyses polypropylene tubes (05ndash1 sdot 106 cellstube) (VWRMilan Italy) EDT-MSCs were subsequently incubated inblocking buffer (100 120583L each 05ndash1 sdot 106 cells) containingDulbeccorsquos Modified Eaglersquos Medium (DMEM Gibco) 10FBS (PAA Laboratories) and 01M sodium azide and humanimmunoglobulin G (both from Sigma) and incubated for201015840 on ice After a PBS (PAA Laboratories) washing stepcells were resuspended in PBS (PAA Laboratories) with 05bovine serum albumin (BSA Sigma) and stained on ice andin the dark for 301015840 with the following monoclonal antibod-ies APC-anti-CD45 FITC-anti-HLADR PE-anti-CD34 PE-anti-CD14 and FITC-anti-CD56 (all from Becton Dickin-son Franklin Lakes NJ USA) PE-anti-CD31 (BioLegend)APC-anti-CD90 PE-anti-SSEA-4 (both from eBioscienceSan Diego CA USA) FITC-anti-CD105 PE-anti-CD73 (allfrom BD Pharmingen) APC-anti-CD146 (Miltenyi Biotec)In all the experiments the corresponding isotype-matchedantibodies were used as negative controls (BD Pharmingenand Becton Dickinson) Data were collected using a FACSAria III flow cytometer (BD Biosciences) and analyzed onFACS Diva software (BD Biosciences)
27 Multilineage Differentiation Assays To assess in vitrodifferentiation capacities adherent cells after P2 were cul-tured in specific induction media Media were changed everyother day and undifferentiated controls were concurrentlycultured in 120572MEM (Gibco) 1 L-glutamine (Lonza) 1
penicillinstreptomycin (PAA Laboratories) and 10 FBS(PAA Laboratories) for the same incubation time Each assaywas performed in triplicate
Adipogenic osteogenic and chondrogenic differentia-tions were performed as previously reported [20] Briefly forosteogenic differentiation cells were seeded at the density of10000 cellscm2 and maintained in the growth medium untilconfluence Culture medium was then substituted with theinduction one composed by 120572MEM (Gibco) supplementedwith 10 nM dexamethasone (Sigma) 10mM 120573-glycerol phos-phate (Sigma) 01mM L-ascorbic acid-2-phosphate (Sigma)10 defined FBS (Hyclone Logan UTUSA) 1 L-glutamine(Lonza) and 1 penicillinstreptomycin (PAA Laboratories)Induction was maintained for 14 days and from the seventhday 100 ngmL BMP-2 (Peprotech Rocky Hill NJ USA)was added Confirmation of osteogenic differentiation wasperformed through Alizarin Red staining combined by real-time qPCR analysis (described below) At the end of theinduction period culture wells were washed briefly witha buffer solution containing 20mM Tris-HCl and 150mMNaCl in water Cells were then fixed and dried with cooledabsolute methanol for 2 minutes After a washing step withddH2O cells were stained by 15 Alizarin Red aqueous
solution pH 40ndash42 (Sigma) for 5 minutes and washed againfirstwith ddH
2Oand thenwith PBS (PAALaboratories) for 15
minutes The last step consisted of dehydration using cooledabsolute ethanol for 2 minutes followed by microscopicobservation
For adipogenic differentiation EDT-MSCswere seeded atthe density of 10000 cellscm2 and maintained in the growthmedium until confluence The medium was then substi-tuted with the induction one composed of 120572MEM (Gibco)supplemented with 1120583M dexamethasone (Sigma) 60 120583Mindomethacin (Sigma) 10120583M insulin (Sigma) 05mM 3-isobutyl-1-methylxanthine (IBMX Sigma) 10 rabbit serum(EuroClone) 5 horse serum (EuroClone) 1 L-glutamine(Lonza) and 1 penicillinstreptomycin (PAA Laboratories)Induction lasted 10 days and confirmation of differentiationwas achieved through Oil Red O staining as follows Culturewells were washed briefly with PBS (PAA Laboratories) Cellswere fixed with vapors of 37 formaldehyde (Sigma) for 10minutes and then washed with water for 2 minutes Stainingwas obtained adding an Oil Red O solution (10mgmL OilRed O in ethanol 70 and acetone all from Sigma) into thewells for 3 minutes Excessive stain was removed by washingwith water and cells were then counterstained by Harrishematoxylin (Bio-Optica Milan Italy) for 30 seconds
Finally for chondrogenic differentiation 2ndash5 sdot 105 EDT-MSCs were aliquoted in a 2mL tube and then centrifugedat 1200 rpm for 10 minutes Cells were maintained pelletedin the growth medium at 37∘C with the plug opened After2 days of incubation tubes were centrifuged again and themedium was substituted with the induction one composedby 120572MEM (Gibco) supplemented with 100 nM dexametha-sone (Sigma) 200120583M L-ascorbic acid-2-phosphate (Sigma)100 120583gmL sodium pyruvate (Biochrom AG Berlin Ger-many) 40 120583gmL proline (Sigma) 1x ITS+ premix (BD Bio-sciences San Jose CA USA) 10 ngmL TGF-120573 (Peprotech)05 120583gmL BMP-6 (Peprotech) 1 L-glutamine (Lonza)
ALP alkaline phosphatase COL1A2 collagen 1A2 OC osteocalcin
and 1 penicillinstreptomycin (PAA Laboratories) Beforeand after each medium change tubes were centrifuged at1200 rpm for 10 minutes During the incubation period cellsremained as pellet with the tube plugs opened Inductionlasted 21 days and specimens were fixed for 1 hour in10 formaldehyde (Sigma) and then dehydrated by serialpassages into ethanol at increasing concentrations from 70to 100 Samples were then included into paraffin blocks andcut in slices on microscope slides for Alcian Blue stainingSlides were deparaffinized with the Histo-C cleaning agent(Celltech Turin Italy) and rehydrated through passagesinto a decreasing concentration alcoholic ladder (from 100ethanol to 70ethanol) Sample sectionswere then incubatedwith a 05mgmLHyaluronidase (Sigma) in buffer phosphatesolution (8 gL NaCl 2 gL NaH
2PO4 and 03 gL Na
2HPO4)
with 10mgmL BSA (Sigma) Slides were washed in water for5 minutes and then immersed in a 3 acetic acid solutionfor few seconds Staining with 10mgmLAlcian Blue solutionin 3 acetic acid (pH 25 Sigma) lasted 30 minutes andafter a washing step in water samples were counterstainedfor 5 minutes with nuclear fast red solution (Sigma) and thenwashed in water
28 Real-Time qPCR Total cellular RNA was isolated fromosteogenic committed and uncommitted cells using Trizolreagent method (Invitrogen) according to manufacturerrsquosinstructions Samples were concentrated by ethanol pre-cipitation and suspended in RNase-free water RNA quan-tity was assessed by spectrophotometry (DU730 UVVISSpectrophotometer Beckman Coulter Milan Italy) A 2120583galiquot was reverse transcribed into cDNA using Rever-tAid First Strand cDNA Synthesis Kit (Fermentas) by oligo(dT)18
primers in a final volume of 20 120583L Reactions wereperformed at 42∘C for 1 h with a final step at 70∘C for 5minutes cDNA was then used to determine the osteoblast-associated gene expression by quantitative real-time PCRtechnique using Step One Real-Time PCR System ThermalCycling Block (Applied Biosystems Foster City CA USA)Each sample was compared with noninduced control forthe expression of alkaline phosphatase collagen 1A2 andosteocalcin 120573-actin was used as a reference gene All primerswere purchased from Integrated DNA Technologies andsequences are reported in Table 1 PCR was performed withFast SYBR Green Master Mix (Applied Biosystems) whichuses AmpliTaq Fast DNA Polymerase SYBR Green I dye
to detect double-stranded DNA and ROX dye as a passiveinternal reference Reaction proceeded through an initial stepat 95∘C for 2010158401015840 followed by 40 cycles of denaturation (310158401015840 at95∘C) annealing and extending (3010158401015840 at 60∘C)The final stagecomprises the analysis of themelt curve through a denaturingstep (1510158401015840 at 95∘C) followed by annealing (11015840 at 60∘C) andramping to 95∘C with 03∘C incrementstep Levels of mRNAfor tested genes were quantified using ΔΔCT method andnormalized against human 120573-actin as a housekeeping geneData have been analyzed by StepOne software (version 21Life Technologies Corporation Carlsbad CA USA)
29 Gene Modification of EDT-MSC Cultured cells wereinfected by a bicistronicmurine stem cell virus-derived retro-viral vector (pMIGR1) encoding for green fluorescent Protein(GFP) Retrovirus production was performed by the FLYRDpackaging cell lines as published by Marx et al [21] FLYRDcells were seeded in a T175 flask with 10mL of medium com-posed by DMEM (Gibco) supplemented with 10 definedFBS (Hyclone) Cell supernatant was collected and filtered(PES) with a 045 120583m filter and EDT-MSC were incubatedfor 6 hrs with 5mL culture medium composed by 120572MEM(Gibco) 1 L-glutamine (Lonza) 1 penicillinstreptomycin(PAA Laboratories) and 10 FBS (PAA Laboratories) withthe addition of 5mL viral particles-containing supernatantand 6 120583gmL polybrene (Sigma) Cells were then washed byPBS (PAA Laboratories) and culture medium was changedThe infection step was repeated for three consecutive daysat which time cells were evaluated at FACS Aria III flowcytometer (BD Biosciences) for GFP protein expression 7-amino-actinomycin D (7AAD) staining was also performedto evaluate mortality after transduction Cells were evaluatedby FACS Aria III (BD Biosciences) and data were analyzedusing FACS Diva software (BD Biosciences)
210 Statistics Data are expressed as average values andanalyses were performed by GraphPad Prism software t-testwas considered as significant with 119875 value lt005
3 Results
31 EDT-Derived Cells Are In Vitro Heterogeneous but RetainPredominant MSC Features Adherent cells isolated frommenstrual blood initially displayed in vitro fibroblast shapemorphology in both 120572MEMwith 10 FBS and Quantum 333
BioMed Research International 5
(Figures 1(a)-1(b)) However prolonged cultures in 120572MEMwith 10 FBS showed a significant better growth perfor-mance and this medium was then preferentially used forall subsequent functional analyses Adherent elements hadtypical mesenchymal aspect being fibroblastoid-like spindle-shaped cells with an elongated cytoplasm Within this pop-ulation it was also possible to identify more infrequentcell clusters with a distinct morphology These appearedas endothelial-like sometimes binucleated cells forming amonolayer with polygonal shape (Figure 1(c)) However wedid not confirm the nature of this population because theirpresence disappeared after very early passages and becausethey were not the subjects of this study
32 EDT-Derived Cells Show Robust Clonogenic and Prolif-erative Potential Having observed the fibroblast shape ofEDT isolated cells we then focused on their clonogenic andproliferative potential We observed a high clonal efficiencywith an average of 141 (108ndash179) of the seeded cellsable to generating colonies This result indicates that themenstrual-derived cell population contains a large fractionof actively cycling cells with signficant clonogenic potential(Figure 2(a)) Of interests colonies did not appear homoge-nous and we were able to identify at least two kinds ofmorphologies On the one hand densely populated clonesconstituted by small size cells (Figures 2(b)-2(c)) and onthe other smaller cell clusters with elements having a largecytoplasm with an evident cytoskeleton (Figures 2(d)-2(e))
Isolated and expanded cells also demonstrated surpris-ingly low doubling time with an average value of 276 hours(219ndash330) (Figure 3) The high number of passages thatthese cells were able to reach further supported this remark-able growth property The 120573-galactosidase staining per-formed to evaluate senescent cells showed that cells cul-tured in Quantum 333 underwent senescence at passage 17while those cultured in 120572MEM supplemented with 10 FBSreached passage 26 before growth arrest (Figures 4(a)ndash4(d))Together these data suggest how plastic-adherent fibroblastshaped cells from EDT retain a strong proliferative potentialcomparable or even superior to MSC from other sour-ces
33 EDT-Derived Cells Display MSC Phenotypic FeaturesHaving evaluated the proliferation potential and to morecarefully define the MSC nature of isolated and expandedcells we assessed their antigen expression profile EDT-derived cells express typical MSC markers (Figures 5 and 6)and in particular more than 90 of tested cells were posi-tive for CD90 and CD73 constituting main features of MSCfrom other sources [22] We also observed that more than80 of cells expressed CD146 an adhesion molecule relatedbut not restricted to MSC and also expressed by endothe-lial cells [1] The levels of CD45 HLADR CD31 and CD14assessed early in culture and commonly used to distinguishMSC from hematopoietic and endothelial cells were below2 in most cases In sample 1 we observed a slightly increasein the CD45+ fraction suggesting the presence of hematopoi-etic elements which might have been isolated together withEDT-MSC Other markers were also considered such as
CD56 whose positivity was extremely variable by up to 45Collectively EDT-MSC from different donors showed thesame overall trend in markers expression (Figure 6) despitefor some markers such as for CD56 CD105 and CD146the variability has been considerably high The phenotypicheterogeneity was also confirmed by analysis of physicalparameters by FACS Forward scatter versus side scatter plotwas very dispersed and it was impossible to identify consist-ent cell groupswith similar physical parameters Interestinglywe have to report a consistent small fraction (gt10) of SSEA-4 positive cells that reached 194 suggesting the expressionof a pluripotency marker in this MSC type
34 EDT-MSCs Are Precursors of Three Mesenchymal TissuesIn Vitro FACS analyses were then followed by assays aim-ing to assess EDT-MSC multipotency We first focused onadipose commitment and after 10 days of adipogenic induc-ing cocktail EDT-MSCs were able to differentiate into vacu-ole-producing elements The Oil Red staining confirmedthe lipid content of those cells indicating the fat-producingability of isolated and induced cells (Figure 7(a)) Osteogenicmedium was then applied for 14 days and cells underwent toosteoblastic commitment forming a compact calcifiedmatrixwith calcium deposits confirmed by specific Alizarin Redstaining (Figure 7(b)) Finally after 21 days of chondrogenicinduction pellets of EDT-MSCs were included into paraffinblocks and Alcian Blue highlighted the sulfated proteogly-cans expression typical of the cartilaginousmatrix while FastRed staining revealed the nuclei of resident chondrocytesderived from EDT-MSC (Figure 7(c)) Since a main focus ofour research group is bone regeneration we then coupledthe cytochemical assay of osteogenic induced EDT-MSCwithqPCR to further confirm bone commitment As seen inFigure 8 all tested genes demonstrated an increased expres-sion in induced samples versus noninduced controls In part-icular in osteoblast-induced cells the expression of alkalinephosphatase resulted 45 times higher (119875 lt 00001) whilemore modest increases were observed for collagen 1A2 (29times 119875 lt 005) and osteocalcin (21 times 119875 lt 005) Thesefindings together with positive Alizarin staining reinforcedthe evidence for commitment of induced EDT-MSC to func-tional osteoblasts
35 EDT-MSC Can Be Efficiently Gene Modified To explorewhether ex vivo expanded EDT-MSC could be geneticallymanipulated for future gene delivery approaches cells wereincubated with supernatants containing retroviral particlescarrying the GFP gene As shown in Figure 9 RD114 pseu-dotyped retroviral particles were able to efficiently and stablytransduce EDT-MSCwith levels greater than 80 suggestinghow this MSC type could be suitable for gene deliveryapproaches
4 Discussion
SeveralMSC types have been obtained starting from differentsources here we have isolated a population of mesenchymalprogenitors from menstrual EDT and characterized themboth at morphological and molecular levels investigating
6 BioMed Research International
(a) (b)
(c)
Figure 1 In vitro adherent cells from decidual tissues (a) Representative photomicrograph of spindle-shaped adherent cells isolated by120572MEM and 10 FBS (b) Cells isolated in vitro by serum-deprived medium (Quantum 333) at early passages (c) Another population of cellswas in vitro isolated contextually with EDT-MSCThese elements share similarities with endothelial cells forming a monolayer of polygonalsometimes binucleated cells Original magnification 100x
their ability to proliferate to differentiate and to be genemodified EDT-MSCs appear as a heterogeneous cells pop-ulation that can be safely and easily isolated by noninvasivemanner providing an expandable source of cells In culturethey show mesenchymal morphology and although somestudies suggest they could derive from bone marrow [23]EDT-MSCs are generally considered to originate from theshedding of endometrial stemprogenitor cells which aremainly resident both in the basalis layer and partially in thefunctionalis area of endometrium [24]
The presence of a uterine population of progenitor cellswould be consistent with the high tissue turn-over after eachmenstrual cycle However it is still uncertain howmany sub-populations of progenitors are present in the endometriumand what are their specific properties Therefore in this pilotstudy we began by addressing the existence of mesenchymalprogenitors in the decidual endometrial tissue
The initial in vitro approach was performed to evaluatethe clonogenic potential of these cells EDT-MSC seeded atclonal density revealed the presence of two distinct colonytypes which closely resembled the stromal cell clones isolatedby Chan et al from endometrium after hysterectomy [12]One was composed of smaller and densely packed cells andthe other was composed of larger and sparser elements Thisfinding seems peculiar for EDT-MSC and differs from col-onies established from BM-MSC that appear larger and
characterized by a central nucleus of cells surrounded by acrown of more sparse ones [18] Next to qualitative evalua-tions of the colonies our data suggest an average EDT-MSCclonogenic efficiency of 14 This value is higher than whatis described for BM-MSC and more similar or even superiorto what reported for AT-MSC that retained around 10 ofclonogenic precursors [25] (Table 3) In addition our EDT-MSC population seemed to have a higher clonogenic effi-ciency compared to those described by other groups dealingwith endometrial stromal cells whose values ranged from 1to 11 [12 24 26] While this may be due to donor-relatedissues such as age in vitro technical aspects including harvest-ing and preservation procedures that may also drive thesedifferences Dimitrov et al observed a tendency for these cellsto decrease in clonal efficiency with the increase of seedingdensity [26] Similarly other culture conditions may generatebetter performing EDT-MSC In this pilot study we testedtwo different growthmedia such as120572MEMwith 10FBS anda serum deprived medium Although these cells apparentlydid not shown particular nutritional requirements serum-containing medium proved to be the best choice ensuring abetter morphological appearance and an improved longevityPrevious studies reported the use of different media such asDMEMF-12 and Chang complete media often supplementedwith calf serum hormones and growth factors [12 14 26 27]This wide range of culture conditions is likely to impact the
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
BioMed Research International 3
24 Proliferation Assay Adherent cells were seeded at adensity of 6000 cellscm2 in 120572MEM (Gibco) 1 L-glutamine(Lonza) 1 penicillinstreptomycin (PAA Laboratories) and10 FBS (PAA Laboratories) in different flasks for thefollowing time points 24 48 72 and 96 hours At each timepoint cells were trypsinized and counted Data obtainedwereplotted as number of harvested cells (119910) against hours ofculture (119909) and the exponential growth curve was generatedusing GraphPad Prism software (GraphPad Software Incversion 500) The doubling time (119879) was obtained from thegrowth constant (119896) of the exponential equation 119910 = 119886 sdot 119890119896119909where 119896 = ln 2119879
25 Senescence-Associated 120573-Galactosidase Staining To as-sess cell senescence the activity of 120573-galactosidase (120573-gal) atpH 6 was evaluated 120573-gal expression is a feature of senescentcells [19] According to Senescence 120573-galactosidase StainingKit (Cell Signaling Technology Beverly MA USA) growthmedium was removed and cells were washed with PBS (PAALaboratories) Cells were then incubated for 15 minutes witha fixative solution (20 formaldehyde 2 glutaraldehyde in10x PBS) and washed twice with PBS (PAA Laboratories)Color development was obtained by incubation overnightat 37∘C with the provided staining solution (40mM citricacidsodium phosphate pH 60 150mMNaCl 2mMMgCl
2
5 nM potassium ferricyanide and 1mgmL X-gal in DMSO)Plateswere then observed bymicroscopy for the developmentof blue color
26 FACSAnalyses Adherent cells were harvested for surfaceantigen analysis Briefly cells were detached from plastic sup-port by trypsin (EuroClone) counted and aliquoted in FACSanalyses polypropylene tubes (05ndash1 sdot 106 cellstube) (VWRMilan Italy) EDT-MSCs were subsequently incubated inblocking buffer (100 120583L each 05ndash1 sdot 106 cells) containingDulbeccorsquos Modified Eaglersquos Medium (DMEM Gibco) 10FBS (PAA Laboratories) and 01M sodium azide and humanimmunoglobulin G (both from Sigma) and incubated for201015840 on ice After a PBS (PAA Laboratories) washing stepcells were resuspended in PBS (PAA Laboratories) with 05bovine serum albumin (BSA Sigma) and stained on ice andin the dark for 301015840 with the following monoclonal antibod-ies APC-anti-CD45 FITC-anti-HLADR PE-anti-CD34 PE-anti-CD14 and FITC-anti-CD56 (all from Becton Dickin-son Franklin Lakes NJ USA) PE-anti-CD31 (BioLegend)APC-anti-CD90 PE-anti-SSEA-4 (both from eBioscienceSan Diego CA USA) FITC-anti-CD105 PE-anti-CD73 (allfrom BD Pharmingen) APC-anti-CD146 (Miltenyi Biotec)In all the experiments the corresponding isotype-matchedantibodies were used as negative controls (BD Pharmingenand Becton Dickinson) Data were collected using a FACSAria III flow cytometer (BD Biosciences) and analyzed onFACS Diva software (BD Biosciences)
27 Multilineage Differentiation Assays To assess in vitrodifferentiation capacities adherent cells after P2 were cul-tured in specific induction media Media were changed everyother day and undifferentiated controls were concurrentlycultured in 120572MEM (Gibco) 1 L-glutamine (Lonza) 1
penicillinstreptomycin (PAA Laboratories) and 10 FBS(PAA Laboratories) for the same incubation time Each assaywas performed in triplicate
Adipogenic osteogenic and chondrogenic differentia-tions were performed as previously reported [20] Briefly forosteogenic differentiation cells were seeded at the density of10000 cellscm2 and maintained in the growth medium untilconfluence Culture medium was then substituted with theinduction one composed by 120572MEM (Gibco) supplementedwith 10 nM dexamethasone (Sigma) 10mM 120573-glycerol phos-phate (Sigma) 01mM L-ascorbic acid-2-phosphate (Sigma)10 defined FBS (Hyclone Logan UTUSA) 1 L-glutamine(Lonza) and 1 penicillinstreptomycin (PAA Laboratories)Induction was maintained for 14 days and from the seventhday 100 ngmL BMP-2 (Peprotech Rocky Hill NJ USA)was added Confirmation of osteogenic differentiation wasperformed through Alizarin Red staining combined by real-time qPCR analysis (described below) At the end of theinduction period culture wells were washed briefly witha buffer solution containing 20mM Tris-HCl and 150mMNaCl in water Cells were then fixed and dried with cooledabsolute methanol for 2 minutes After a washing step withddH2O cells were stained by 15 Alizarin Red aqueous
solution pH 40ndash42 (Sigma) for 5 minutes and washed againfirstwith ddH
2Oand thenwith PBS (PAALaboratories) for 15
minutes The last step consisted of dehydration using cooledabsolute ethanol for 2 minutes followed by microscopicobservation
For adipogenic differentiation EDT-MSCswere seeded atthe density of 10000 cellscm2 and maintained in the growthmedium until confluence The medium was then substi-tuted with the induction one composed of 120572MEM (Gibco)supplemented with 1120583M dexamethasone (Sigma) 60 120583Mindomethacin (Sigma) 10120583M insulin (Sigma) 05mM 3-isobutyl-1-methylxanthine (IBMX Sigma) 10 rabbit serum(EuroClone) 5 horse serum (EuroClone) 1 L-glutamine(Lonza) and 1 penicillinstreptomycin (PAA Laboratories)Induction lasted 10 days and confirmation of differentiationwas achieved through Oil Red O staining as follows Culturewells were washed briefly with PBS (PAA Laboratories) Cellswere fixed with vapors of 37 formaldehyde (Sigma) for 10minutes and then washed with water for 2 minutes Stainingwas obtained adding an Oil Red O solution (10mgmL OilRed O in ethanol 70 and acetone all from Sigma) into thewells for 3 minutes Excessive stain was removed by washingwith water and cells were then counterstained by Harrishematoxylin (Bio-Optica Milan Italy) for 30 seconds
Finally for chondrogenic differentiation 2ndash5 sdot 105 EDT-MSCs were aliquoted in a 2mL tube and then centrifugedat 1200 rpm for 10 minutes Cells were maintained pelletedin the growth medium at 37∘C with the plug opened After2 days of incubation tubes were centrifuged again and themedium was substituted with the induction one composedby 120572MEM (Gibco) supplemented with 100 nM dexametha-sone (Sigma) 200120583M L-ascorbic acid-2-phosphate (Sigma)100 120583gmL sodium pyruvate (Biochrom AG Berlin Ger-many) 40 120583gmL proline (Sigma) 1x ITS+ premix (BD Bio-sciences San Jose CA USA) 10 ngmL TGF-120573 (Peprotech)05 120583gmL BMP-6 (Peprotech) 1 L-glutamine (Lonza)
ALP alkaline phosphatase COL1A2 collagen 1A2 OC osteocalcin
and 1 penicillinstreptomycin (PAA Laboratories) Beforeand after each medium change tubes were centrifuged at1200 rpm for 10 minutes During the incubation period cellsremained as pellet with the tube plugs opened Inductionlasted 21 days and specimens were fixed for 1 hour in10 formaldehyde (Sigma) and then dehydrated by serialpassages into ethanol at increasing concentrations from 70to 100 Samples were then included into paraffin blocks andcut in slices on microscope slides for Alcian Blue stainingSlides were deparaffinized with the Histo-C cleaning agent(Celltech Turin Italy) and rehydrated through passagesinto a decreasing concentration alcoholic ladder (from 100ethanol to 70ethanol) Sample sectionswere then incubatedwith a 05mgmLHyaluronidase (Sigma) in buffer phosphatesolution (8 gL NaCl 2 gL NaH
2PO4 and 03 gL Na
2HPO4)
with 10mgmL BSA (Sigma) Slides were washed in water for5 minutes and then immersed in a 3 acetic acid solutionfor few seconds Staining with 10mgmLAlcian Blue solutionin 3 acetic acid (pH 25 Sigma) lasted 30 minutes andafter a washing step in water samples were counterstainedfor 5 minutes with nuclear fast red solution (Sigma) and thenwashed in water
28 Real-Time qPCR Total cellular RNA was isolated fromosteogenic committed and uncommitted cells using Trizolreagent method (Invitrogen) according to manufacturerrsquosinstructions Samples were concentrated by ethanol pre-cipitation and suspended in RNase-free water RNA quan-tity was assessed by spectrophotometry (DU730 UVVISSpectrophotometer Beckman Coulter Milan Italy) A 2120583galiquot was reverse transcribed into cDNA using Rever-tAid First Strand cDNA Synthesis Kit (Fermentas) by oligo(dT)18
primers in a final volume of 20 120583L Reactions wereperformed at 42∘C for 1 h with a final step at 70∘C for 5minutes cDNA was then used to determine the osteoblast-associated gene expression by quantitative real-time PCRtechnique using Step One Real-Time PCR System ThermalCycling Block (Applied Biosystems Foster City CA USA)Each sample was compared with noninduced control forthe expression of alkaline phosphatase collagen 1A2 andosteocalcin 120573-actin was used as a reference gene All primerswere purchased from Integrated DNA Technologies andsequences are reported in Table 1 PCR was performed withFast SYBR Green Master Mix (Applied Biosystems) whichuses AmpliTaq Fast DNA Polymerase SYBR Green I dye
to detect double-stranded DNA and ROX dye as a passiveinternal reference Reaction proceeded through an initial stepat 95∘C for 2010158401015840 followed by 40 cycles of denaturation (310158401015840 at95∘C) annealing and extending (3010158401015840 at 60∘C)The final stagecomprises the analysis of themelt curve through a denaturingstep (1510158401015840 at 95∘C) followed by annealing (11015840 at 60∘C) andramping to 95∘C with 03∘C incrementstep Levels of mRNAfor tested genes were quantified using ΔΔCT method andnormalized against human 120573-actin as a housekeeping geneData have been analyzed by StepOne software (version 21Life Technologies Corporation Carlsbad CA USA)
29 Gene Modification of EDT-MSC Cultured cells wereinfected by a bicistronicmurine stem cell virus-derived retro-viral vector (pMIGR1) encoding for green fluorescent Protein(GFP) Retrovirus production was performed by the FLYRDpackaging cell lines as published by Marx et al [21] FLYRDcells were seeded in a T175 flask with 10mL of medium com-posed by DMEM (Gibco) supplemented with 10 definedFBS (Hyclone) Cell supernatant was collected and filtered(PES) with a 045 120583m filter and EDT-MSC were incubatedfor 6 hrs with 5mL culture medium composed by 120572MEM(Gibco) 1 L-glutamine (Lonza) 1 penicillinstreptomycin(PAA Laboratories) and 10 FBS (PAA Laboratories) withthe addition of 5mL viral particles-containing supernatantand 6 120583gmL polybrene (Sigma) Cells were then washed byPBS (PAA Laboratories) and culture medium was changedThe infection step was repeated for three consecutive daysat which time cells were evaluated at FACS Aria III flowcytometer (BD Biosciences) for GFP protein expression 7-amino-actinomycin D (7AAD) staining was also performedto evaluate mortality after transduction Cells were evaluatedby FACS Aria III (BD Biosciences) and data were analyzedusing FACS Diva software (BD Biosciences)
210 Statistics Data are expressed as average values andanalyses were performed by GraphPad Prism software t-testwas considered as significant with 119875 value lt005
3 Results
31 EDT-Derived Cells Are In Vitro Heterogeneous but RetainPredominant MSC Features Adherent cells isolated frommenstrual blood initially displayed in vitro fibroblast shapemorphology in both 120572MEMwith 10 FBS and Quantum 333
BioMed Research International 5
(Figures 1(a)-1(b)) However prolonged cultures in 120572MEMwith 10 FBS showed a significant better growth perfor-mance and this medium was then preferentially used forall subsequent functional analyses Adherent elements hadtypical mesenchymal aspect being fibroblastoid-like spindle-shaped cells with an elongated cytoplasm Within this pop-ulation it was also possible to identify more infrequentcell clusters with a distinct morphology These appearedas endothelial-like sometimes binucleated cells forming amonolayer with polygonal shape (Figure 1(c)) However wedid not confirm the nature of this population because theirpresence disappeared after very early passages and becausethey were not the subjects of this study
32 EDT-Derived Cells Show Robust Clonogenic and Prolif-erative Potential Having observed the fibroblast shape ofEDT isolated cells we then focused on their clonogenic andproliferative potential We observed a high clonal efficiencywith an average of 141 (108ndash179) of the seeded cellsable to generating colonies This result indicates that themenstrual-derived cell population contains a large fractionof actively cycling cells with signficant clonogenic potential(Figure 2(a)) Of interests colonies did not appear homoge-nous and we were able to identify at least two kinds ofmorphologies On the one hand densely populated clonesconstituted by small size cells (Figures 2(b)-2(c)) and onthe other smaller cell clusters with elements having a largecytoplasm with an evident cytoskeleton (Figures 2(d)-2(e))
Isolated and expanded cells also demonstrated surpris-ingly low doubling time with an average value of 276 hours(219ndash330) (Figure 3) The high number of passages thatthese cells were able to reach further supported this remark-able growth property The 120573-galactosidase staining per-formed to evaluate senescent cells showed that cells cul-tured in Quantum 333 underwent senescence at passage 17while those cultured in 120572MEM supplemented with 10 FBSreached passage 26 before growth arrest (Figures 4(a)ndash4(d))Together these data suggest how plastic-adherent fibroblastshaped cells from EDT retain a strong proliferative potentialcomparable or even superior to MSC from other sour-ces
33 EDT-Derived Cells Display MSC Phenotypic FeaturesHaving evaluated the proliferation potential and to morecarefully define the MSC nature of isolated and expandedcells we assessed their antigen expression profile EDT-derived cells express typical MSC markers (Figures 5 and 6)and in particular more than 90 of tested cells were posi-tive for CD90 and CD73 constituting main features of MSCfrom other sources [22] We also observed that more than80 of cells expressed CD146 an adhesion molecule relatedbut not restricted to MSC and also expressed by endothe-lial cells [1] The levels of CD45 HLADR CD31 and CD14assessed early in culture and commonly used to distinguishMSC from hematopoietic and endothelial cells were below2 in most cases In sample 1 we observed a slightly increasein the CD45+ fraction suggesting the presence of hematopoi-etic elements which might have been isolated together withEDT-MSC Other markers were also considered such as
CD56 whose positivity was extremely variable by up to 45Collectively EDT-MSC from different donors showed thesame overall trend in markers expression (Figure 6) despitefor some markers such as for CD56 CD105 and CD146the variability has been considerably high The phenotypicheterogeneity was also confirmed by analysis of physicalparameters by FACS Forward scatter versus side scatter plotwas very dispersed and it was impossible to identify consist-ent cell groupswith similar physical parameters Interestinglywe have to report a consistent small fraction (gt10) of SSEA-4 positive cells that reached 194 suggesting the expressionof a pluripotency marker in this MSC type
34 EDT-MSCs Are Precursors of Three Mesenchymal TissuesIn Vitro FACS analyses were then followed by assays aim-ing to assess EDT-MSC multipotency We first focused onadipose commitment and after 10 days of adipogenic induc-ing cocktail EDT-MSCs were able to differentiate into vacu-ole-producing elements The Oil Red staining confirmedthe lipid content of those cells indicating the fat-producingability of isolated and induced cells (Figure 7(a)) Osteogenicmedium was then applied for 14 days and cells underwent toosteoblastic commitment forming a compact calcifiedmatrixwith calcium deposits confirmed by specific Alizarin Redstaining (Figure 7(b)) Finally after 21 days of chondrogenicinduction pellets of EDT-MSCs were included into paraffinblocks and Alcian Blue highlighted the sulfated proteogly-cans expression typical of the cartilaginousmatrix while FastRed staining revealed the nuclei of resident chondrocytesderived from EDT-MSC (Figure 7(c)) Since a main focus ofour research group is bone regeneration we then coupledthe cytochemical assay of osteogenic induced EDT-MSCwithqPCR to further confirm bone commitment As seen inFigure 8 all tested genes demonstrated an increased expres-sion in induced samples versus noninduced controls In part-icular in osteoblast-induced cells the expression of alkalinephosphatase resulted 45 times higher (119875 lt 00001) whilemore modest increases were observed for collagen 1A2 (29times 119875 lt 005) and osteocalcin (21 times 119875 lt 005) Thesefindings together with positive Alizarin staining reinforcedthe evidence for commitment of induced EDT-MSC to func-tional osteoblasts
35 EDT-MSC Can Be Efficiently Gene Modified To explorewhether ex vivo expanded EDT-MSC could be geneticallymanipulated for future gene delivery approaches cells wereincubated with supernatants containing retroviral particlescarrying the GFP gene As shown in Figure 9 RD114 pseu-dotyped retroviral particles were able to efficiently and stablytransduce EDT-MSCwith levels greater than 80 suggestinghow this MSC type could be suitable for gene deliveryapproaches
4 Discussion
SeveralMSC types have been obtained starting from differentsources here we have isolated a population of mesenchymalprogenitors from menstrual EDT and characterized themboth at morphological and molecular levels investigating
6 BioMed Research International
(a) (b)
(c)
Figure 1 In vitro adherent cells from decidual tissues (a) Representative photomicrograph of spindle-shaped adherent cells isolated by120572MEM and 10 FBS (b) Cells isolated in vitro by serum-deprived medium (Quantum 333) at early passages (c) Another population of cellswas in vitro isolated contextually with EDT-MSCThese elements share similarities with endothelial cells forming a monolayer of polygonalsometimes binucleated cells Original magnification 100x
their ability to proliferate to differentiate and to be genemodified EDT-MSCs appear as a heterogeneous cells pop-ulation that can be safely and easily isolated by noninvasivemanner providing an expandable source of cells In culturethey show mesenchymal morphology and although somestudies suggest they could derive from bone marrow [23]EDT-MSCs are generally considered to originate from theshedding of endometrial stemprogenitor cells which aremainly resident both in the basalis layer and partially in thefunctionalis area of endometrium [24]
The presence of a uterine population of progenitor cellswould be consistent with the high tissue turn-over after eachmenstrual cycle However it is still uncertain howmany sub-populations of progenitors are present in the endometriumand what are their specific properties Therefore in this pilotstudy we began by addressing the existence of mesenchymalprogenitors in the decidual endometrial tissue
The initial in vitro approach was performed to evaluatethe clonogenic potential of these cells EDT-MSC seeded atclonal density revealed the presence of two distinct colonytypes which closely resembled the stromal cell clones isolatedby Chan et al from endometrium after hysterectomy [12]One was composed of smaller and densely packed cells andthe other was composed of larger and sparser elements Thisfinding seems peculiar for EDT-MSC and differs from col-onies established from BM-MSC that appear larger and
characterized by a central nucleus of cells surrounded by acrown of more sparse ones [18] Next to qualitative evalua-tions of the colonies our data suggest an average EDT-MSCclonogenic efficiency of 14 This value is higher than whatis described for BM-MSC and more similar or even superiorto what reported for AT-MSC that retained around 10 ofclonogenic precursors [25] (Table 3) In addition our EDT-MSC population seemed to have a higher clonogenic effi-ciency compared to those described by other groups dealingwith endometrial stromal cells whose values ranged from 1to 11 [12 24 26] While this may be due to donor-relatedissues such as age in vitro technical aspects including harvest-ing and preservation procedures that may also drive thesedifferences Dimitrov et al observed a tendency for these cellsto decrease in clonal efficiency with the increase of seedingdensity [26] Similarly other culture conditions may generatebetter performing EDT-MSC In this pilot study we testedtwo different growthmedia such as120572MEMwith 10FBS anda serum deprived medium Although these cells apparentlydid not shown particular nutritional requirements serum-containing medium proved to be the best choice ensuring abetter morphological appearance and an improved longevityPrevious studies reported the use of different media such asDMEMF-12 and Chang complete media often supplementedwith calf serum hormones and growth factors [12 14 26 27]This wide range of culture conditions is likely to impact the
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
ALP alkaline phosphatase COL1A2 collagen 1A2 OC osteocalcin
and 1 penicillinstreptomycin (PAA Laboratories) Beforeand after each medium change tubes were centrifuged at1200 rpm for 10 minutes During the incubation period cellsremained as pellet with the tube plugs opened Inductionlasted 21 days and specimens were fixed for 1 hour in10 formaldehyde (Sigma) and then dehydrated by serialpassages into ethanol at increasing concentrations from 70to 100 Samples were then included into paraffin blocks andcut in slices on microscope slides for Alcian Blue stainingSlides were deparaffinized with the Histo-C cleaning agent(Celltech Turin Italy) and rehydrated through passagesinto a decreasing concentration alcoholic ladder (from 100ethanol to 70ethanol) Sample sectionswere then incubatedwith a 05mgmLHyaluronidase (Sigma) in buffer phosphatesolution (8 gL NaCl 2 gL NaH
2PO4 and 03 gL Na
2HPO4)
with 10mgmL BSA (Sigma) Slides were washed in water for5 minutes and then immersed in a 3 acetic acid solutionfor few seconds Staining with 10mgmLAlcian Blue solutionin 3 acetic acid (pH 25 Sigma) lasted 30 minutes andafter a washing step in water samples were counterstainedfor 5 minutes with nuclear fast red solution (Sigma) and thenwashed in water
28 Real-Time qPCR Total cellular RNA was isolated fromosteogenic committed and uncommitted cells using Trizolreagent method (Invitrogen) according to manufacturerrsquosinstructions Samples were concentrated by ethanol pre-cipitation and suspended in RNase-free water RNA quan-tity was assessed by spectrophotometry (DU730 UVVISSpectrophotometer Beckman Coulter Milan Italy) A 2120583galiquot was reverse transcribed into cDNA using Rever-tAid First Strand cDNA Synthesis Kit (Fermentas) by oligo(dT)18
primers in a final volume of 20 120583L Reactions wereperformed at 42∘C for 1 h with a final step at 70∘C for 5minutes cDNA was then used to determine the osteoblast-associated gene expression by quantitative real-time PCRtechnique using Step One Real-Time PCR System ThermalCycling Block (Applied Biosystems Foster City CA USA)Each sample was compared with noninduced control forthe expression of alkaline phosphatase collagen 1A2 andosteocalcin 120573-actin was used as a reference gene All primerswere purchased from Integrated DNA Technologies andsequences are reported in Table 1 PCR was performed withFast SYBR Green Master Mix (Applied Biosystems) whichuses AmpliTaq Fast DNA Polymerase SYBR Green I dye
to detect double-stranded DNA and ROX dye as a passiveinternal reference Reaction proceeded through an initial stepat 95∘C for 2010158401015840 followed by 40 cycles of denaturation (310158401015840 at95∘C) annealing and extending (3010158401015840 at 60∘C)The final stagecomprises the analysis of themelt curve through a denaturingstep (1510158401015840 at 95∘C) followed by annealing (11015840 at 60∘C) andramping to 95∘C with 03∘C incrementstep Levels of mRNAfor tested genes were quantified using ΔΔCT method andnormalized against human 120573-actin as a housekeeping geneData have been analyzed by StepOne software (version 21Life Technologies Corporation Carlsbad CA USA)
29 Gene Modification of EDT-MSC Cultured cells wereinfected by a bicistronicmurine stem cell virus-derived retro-viral vector (pMIGR1) encoding for green fluorescent Protein(GFP) Retrovirus production was performed by the FLYRDpackaging cell lines as published by Marx et al [21] FLYRDcells were seeded in a T175 flask with 10mL of medium com-posed by DMEM (Gibco) supplemented with 10 definedFBS (Hyclone) Cell supernatant was collected and filtered(PES) with a 045 120583m filter and EDT-MSC were incubatedfor 6 hrs with 5mL culture medium composed by 120572MEM(Gibco) 1 L-glutamine (Lonza) 1 penicillinstreptomycin(PAA Laboratories) and 10 FBS (PAA Laboratories) withthe addition of 5mL viral particles-containing supernatantand 6 120583gmL polybrene (Sigma) Cells were then washed byPBS (PAA Laboratories) and culture medium was changedThe infection step was repeated for three consecutive daysat which time cells were evaluated at FACS Aria III flowcytometer (BD Biosciences) for GFP protein expression 7-amino-actinomycin D (7AAD) staining was also performedto evaluate mortality after transduction Cells were evaluatedby FACS Aria III (BD Biosciences) and data were analyzedusing FACS Diva software (BD Biosciences)
210 Statistics Data are expressed as average values andanalyses were performed by GraphPad Prism software t-testwas considered as significant with 119875 value lt005
3 Results
31 EDT-Derived Cells Are In Vitro Heterogeneous but RetainPredominant MSC Features Adherent cells isolated frommenstrual blood initially displayed in vitro fibroblast shapemorphology in both 120572MEMwith 10 FBS and Quantum 333
BioMed Research International 5
(Figures 1(a)-1(b)) However prolonged cultures in 120572MEMwith 10 FBS showed a significant better growth perfor-mance and this medium was then preferentially used forall subsequent functional analyses Adherent elements hadtypical mesenchymal aspect being fibroblastoid-like spindle-shaped cells with an elongated cytoplasm Within this pop-ulation it was also possible to identify more infrequentcell clusters with a distinct morphology These appearedas endothelial-like sometimes binucleated cells forming amonolayer with polygonal shape (Figure 1(c)) However wedid not confirm the nature of this population because theirpresence disappeared after very early passages and becausethey were not the subjects of this study
32 EDT-Derived Cells Show Robust Clonogenic and Prolif-erative Potential Having observed the fibroblast shape ofEDT isolated cells we then focused on their clonogenic andproliferative potential We observed a high clonal efficiencywith an average of 141 (108ndash179) of the seeded cellsable to generating colonies This result indicates that themenstrual-derived cell population contains a large fractionof actively cycling cells with signficant clonogenic potential(Figure 2(a)) Of interests colonies did not appear homoge-nous and we were able to identify at least two kinds ofmorphologies On the one hand densely populated clonesconstituted by small size cells (Figures 2(b)-2(c)) and onthe other smaller cell clusters with elements having a largecytoplasm with an evident cytoskeleton (Figures 2(d)-2(e))
Isolated and expanded cells also demonstrated surpris-ingly low doubling time with an average value of 276 hours(219ndash330) (Figure 3) The high number of passages thatthese cells were able to reach further supported this remark-able growth property The 120573-galactosidase staining per-formed to evaluate senescent cells showed that cells cul-tured in Quantum 333 underwent senescence at passage 17while those cultured in 120572MEM supplemented with 10 FBSreached passage 26 before growth arrest (Figures 4(a)ndash4(d))Together these data suggest how plastic-adherent fibroblastshaped cells from EDT retain a strong proliferative potentialcomparable or even superior to MSC from other sour-ces
33 EDT-Derived Cells Display MSC Phenotypic FeaturesHaving evaluated the proliferation potential and to morecarefully define the MSC nature of isolated and expandedcells we assessed their antigen expression profile EDT-derived cells express typical MSC markers (Figures 5 and 6)and in particular more than 90 of tested cells were posi-tive for CD90 and CD73 constituting main features of MSCfrom other sources [22] We also observed that more than80 of cells expressed CD146 an adhesion molecule relatedbut not restricted to MSC and also expressed by endothe-lial cells [1] The levels of CD45 HLADR CD31 and CD14assessed early in culture and commonly used to distinguishMSC from hematopoietic and endothelial cells were below2 in most cases In sample 1 we observed a slightly increasein the CD45+ fraction suggesting the presence of hematopoi-etic elements which might have been isolated together withEDT-MSC Other markers were also considered such as
CD56 whose positivity was extremely variable by up to 45Collectively EDT-MSC from different donors showed thesame overall trend in markers expression (Figure 6) despitefor some markers such as for CD56 CD105 and CD146the variability has been considerably high The phenotypicheterogeneity was also confirmed by analysis of physicalparameters by FACS Forward scatter versus side scatter plotwas very dispersed and it was impossible to identify consist-ent cell groupswith similar physical parameters Interestinglywe have to report a consistent small fraction (gt10) of SSEA-4 positive cells that reached 194 suggesting the expressionof a pluripotency marker in this MSC type
34 EDT-MSCs Are Precursors of Three Mesenchymal TissuesIn Vitro FACS analyses were then followed by assays aim-ing to assess EDT-MSC multipotency We first focused onadipose commitment and after 10 days of adipogenic induc-ing cocktail EDT-MSCs were able to differentiate into vacu-ole-producing elements The Oil Red staining confirmedthe lipid content of those cells indicating the fat-producingability of isolated and induced cells (Figure 7(a)) Osteogenicmedium was then applied for 14 days and cells underwent toosteoblastic commitment forming a compact calcifiedmatrixwith calcium deposits confirmed by specific Alizarin Redstaining (Figure 7(b)) Finally after 21 days of chondrogenicinduction pellets of EDT-MSCs were included into paraffinblocks and Alcian Blue highlighted the sulfated proteogly-cans expression typical of the cartilaginousmatrix while FastRed staining revealed the nuclei of resident chondrocytesderived from EDT-MSC (Figure 7(c)) Since a main focus ofour research group is bone regeneration we then coupledthe cytochemical assay of osteogenic induced EDT-MSCwithqPCR to further confirm bone commitment As seen inFigure 8 all tested genes demonstrated an increased expres-sion in induced samples versus noninduced controls In part-icular in osteoblast-induced cells the expression of alkalinephosphatase resulted 45 times higher (119875 lt 00001) whilemore modest increases were observed for collagen 1A2 (29times 119875 lt 005) and osteocalcin (21 times 119875 lt 005) Thesefindings together with positive Alizarin staining reinforcedthe evidence for commitment of induced EDT-MSC to func-tional osteoblasts
35 EDT-MSC Can Be Efficiently Gene Modified To explorewhether ex vivo expanded EDT-MSC could be geneticallymanipulated for future gene delivery approaches cells wereincubated with supernatants containing retroviral particlescarrying the GFP gene As shown in Figure 9 RD114 pseu-dotyped retroviral particles were able to efficiently and stablytransduce EDT-MSCwith levels greater than 80 suggestinghow this MSC type could be suitable for gene deliveryapproaches
4 Discussion
SeveralMSC types have been obtained starting from differentsources here we have isolated a population of mesenchymalprogenitors from menstrual EDT and characterized themboth at morphological and molecular levels investigating
6 BioMed Research International
(a) (b)
(c)
Figure 1 In vitro adherent cells from decidual tissues (a) Representative photomicrograph of spindle-shaped adherent cells isolated by120572MEM and 10 FBS (b) Cells isolated in vitro by serum-deprived medium (Quantum 333) at early passages (c) Another population of cellswas in vitro isolated contextually with EDT-MSCThese elements share similarities with endothelial cells forming a monolayer of polygonalsometimes binucleated cells Original magnification 100x
their ability to proliferate to differentiate and to be genemodified EDT-MSCs appear as a heterogeneous cells pop-ulation that can be safely and easily isolated by noninvasivemanner providing an expandable source of cells In culturethey show mesenchymal morphology and although somestudies suggest they could derive from bone marrow [23]EDT-MSCs are generally considered to originate from theshedding of endometrial stemprogenitor cells which aremainly resident both in the basalis layer and partially in thefunctionalis area of endometrium [24]
The presence of a uterine population of progenitor cellswould be consistent with the high tissue turn-over after eachmenstrual cycle However it is still uncertain howmany sub-populations of progenitors are present in the endometriumand what are their specific properties Therefore in this pilotstudy we began by addressing the existence of mesenchymalprogenitors in the decidual endometrial tissue
The initial in vitro approach was performed to evaluatethe clonogenic potential of these cells EDT-MSC seeded atclonal density revealed the presence of two distinct colonytypes which closely resembled the stromal cell clones isolatedby Chan et al from endometrium after hysterectomy [12]One was composed of smaller and densely packed cells andthe other was composed of larger and sparser elements Thisfinding seems peculiar for EDT-MSC and differs from col-onies established from BM-MSC that appear larger and
characterized by a central nucleus of cells surrounded by acrown of more sparse ones [18] Next to qualitative evalua-tions of the colonies our data suggest an average EDT-MSCclonogenic efficiency of 14 This value is higher than whatis described for BM-MSC and more similar or even superiorto what reported for AT-MSC that retained around 10 ofclonogenic precursors [25] (Table 3) In addition our EDT-MSC population seemed to have a higher clonogenic effi-ciency compared to those described by other groups dealingwith endometrial stromal cells whose values ranged from 1to 11 [12 24 26] While this may be due to donor-relatedissues such as age in vitro technical aspects including harvest-ing and preservation procedures that may also drive thesedifferences Dimitrov et al observed a tendency for these cellsto decrease in clonal efficiency with the increase of seedingdensity [26] Similarly other culture conditions may generatebetter performing EDT-MSC In this pilot study we testedtwo different growthmedia such as120572MEMwith 10FBS anda serum deprived medium Although these cells apparentlydid not shown particular nutritional requirements serum-containing medium proved to be the best choice ensuring abetter morphological appearance and an improved longevityPrevious studies reported the use of different media such asDMEMF-12 and Chang complete media often supplementedwith calf serum hormones and growth factors [12 14 26 27]This wide range of culture conditions is likely to impact the
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
BioMed Research International 5
(Figures 1(a)-1(b)) However prolonged cultures in 120572MEMwith 10 FBS showed a significant better growth perfor-mance and this medium was then preferentially used forall subsequent functional analyses Adherent elements hadtypical mesenchymal aspect being fibroblastoid-like spindle-shaped cells with an elongated cytoplasm Within this pop-ulation it was also possible to identify more infrequentcell clusters with a distinct morphology These appearedas endothelial-like sometimes binucleated cells forming amonolayer with polygonal shape (Figure 1(c)) However wedid not confirm the nature of this population because theirpresence disappeared after very early passages and becausethey were not the subjects of this study
32 EDT-Derived Cells Show Robust Clonogenic and Prolif-erative Potential Having observed the fibroblast shape ofEDT isolated cells we then focused on their clonogenic andproliferative potential We observed a high clonal efficiencywith an average of 141 (108ndash179) of the seeded cellsable to generating colonies This result indicates that themenstrual-derived cell population contains a large fractionof actively cycling cells with signficant clonogenic potential(Figure 2(a)) Of interests colonies did not appear homoge-nous and we were able to identify at least two kinds ofmorphologies On the one hand densely populated clonesconstituted by small size cells (Figures 2(b)-2(c)) and onthe other smaller cell clusters with elements having a largecytoplasm with an evident cytoskeleton (Figures 2(d)-2(e))
Isolated and expanded cells also demonstrated surpris-ingly low doubling time with an average value of 276 hours(219ndash330) (Figure 3) The high number of passages thatthese cells were able to reach further supported this remark-able growth property The 120573-galactosidase staining per-formed to evaluate senescent cells showed that cells cul-tured in Quantum 333 underwent senescence at passage 17while those cultured in 120572MEM supplemented with 10 FBSreached passage 26 before growth arrest (Figures 4(a)ndash4(d))Together these data suggest how plastic-adherent fibroblastshaped cells from EDT retain a strong proliferative potentialcomparable or even superior to MSC from other sour-ces
33 EDT-Derived Cells Display MSC Phenotypic FeaturesHaving evaluated the proliferation potential and to morecarefully define the MSC nature of isolated and expandedcells we assessed their antigen expression profile EDT-derived cells express typical MSC markers (Figures 5 and 6)and in particular more than 90 of tested cells were posi-tive for CD90 and CD73 constituting main features of MSCfrom other sources [22] We also observed that more than80 of cells expressed CD146 an adhesion molecule relatedbut not restricted to MSC and also expressed by endothe-lial cells [1] The levels of CD45 HLADR CD31 and CD14assessed early in culture and commonly used to distinguishMSC from hematopoietic and endothelial cells were below2 in most cases In sample 1 we observed a slightly increasein the CD45+ fraction suggesting the presence of hematopoi-etic elements which might have been isolated together withEDT-MSC Other markers were also considered such as
CD56 whose positivity was extremely variable by up to 45Collectively EDT-MSC from different donors showed thesame overall trend in markers expression (Figure 6) despitefor some markers such as for CD56 CD105 and CD146the variability has been considerably high The phenotypicheterogeneity was also confirmed by analysis of physicalparameters by FACS Forward scatter versus side scatter plotwas very dispersed and it was impossible to identify consist-ent cell groupswith similar physical parameters Interestinglywe have to report a consistent small fraction (gt10) of SSEA-4 positive cells that reached 194 suggesting the expressionof a pluripotency marker in this MSC type
34 EDT-MSCs Are Precursors of Three Mesenchymal TissuesIn Vitro FACS analyses were then followed by assays aim-ing to assess EDT-MSC multipotency We first focused onadipose commitment and after 10 days of adipogenic induc-ing cocktail EDT-MSCs were able to differentiate into vacu-ole-producing elements The Oil Red staining confirmedthe lipid content of those cells indicating the fat-producingability of isolated and induced cells (Figure 7(a)) Osteogenicmedium was then applied for 14 days and cells underwent toosteoblastic commitment forming a compact calcifiedmatrixwith calcium deposits confirmed by specific Alizarin Redstaining (Figure 7(b)) Finally after 21 days of chondrogenicinduction pellets of EDT-MSCs were included into paraffinblocks and Alcian Blue highlighted the sulfated proteogly-cans expression typical of the cartilaginousmatrix while FastRed staining revealed the nuclei of resident chondrocytesderived from EDT-MSC (Figure 7(c)) Since a main focus ofour research group is bone regeneration we then coupledthe cytochemical assay of osteogenic induced EDT-MSCwithqPCR to further confirm bone commitment As seen inFigure 8 all tested genes demonstrated an increased expres-sion in induced samples versus noninduced controls In part-icular in osteoblast-induced cells the expression of alkalinephosphatase resulted 45 times higher (119875 lt 00001) whilemore modest increases were observed for collagen 1A2 (29times 119875 lt 005) and osteocalcin (21 times 119875 lt 005) Thesefindings together with positive Alizarin staining reinforcedthe evidence for commitment of induced EDT-MSC to func-tional osteoblasts
35 EDT-MSC Can Be Efficiently Gene Modified To explorewhether ex vivo expanded EDT-MSC could be geneticallymanipulated for future gene delivery approaches cells wereincubated with supernatants containing retroviral particlescarrying the GFP gene As shown in Figure 9 RD114 pseu-dotyped retroviral particles were able to efficiently and stablytransduce EDT-MSCwith levels greater than 80 suggestinghow this MSC type could be suitable for gene deliveryapproaches
4 Discussion
SeveralMSC types have been obtained starting from differentsources here we have isolated a population of mesenchymalprogenitors from menstrual EDT and characterized themboth at morphological and molecular levels investigating
6 BioMed Research International
(a) (b)
(c)
Figure 1 In vitro adherent cells from decidual tissues (a) Representative photomicrograph of spindle-shaped adherent cells isolated by120572MEM and 10 FBS (b) Cells isolated in vitro by serum-deprived medium (Quantum 333) at early passages (c) Another population of cellswas in vitro isolated contextually with EDT-MSCThese elements share similarities with endothelial cells forming a monolayer of polygonalsometimes binucleated cells Original magnification 100x
their ability to proliferate to differentiate and to be genemodified EDT-MSCs appear as a heterogeneous cells pop-ulation that can be safely and easily isolated by noninvasivemanner providing an expandable source of cells In culturethey show mesenchymal morphology and although somestudies suggest they could derive from bone marrow [23]EDT-MSCs are generally considered to originate from theshedding of endometrial stemprogenitor cells which aremainly resident both in the basalis layer and partially in thefunctionalis area of endometrium [24]
The presence of a uterine population of progenitor cellswould be consistent with the high tissue turn-over after eachmenstrual cycle However it is still uncertain howmany sub-populations of progenitors are present in the endometriumand what are their specific properties Therefore in this pilotstudy we began by addressing the existence of mesenchymalprogenitors in the decidual endometrial tissue
The initial in vitro approach was performed to evaluatethe clonogenic potential of these cells EDT-MSC seeded atclonal density revealed the presence of two distinct colonytypes which closely resembled the stromal cell clones isolatedby Chan et al from endometrium after hysterectomy [12]One was composed of smaller and densely packed cells andthe other was composed of larger and sparser elements Thisfinding seems peculiar for EDT-MSC and differs from col-onies established from BM-MSC that appear larger and
characterized by a central nucleus of cells surrounded by acrown of more sparse ones [18] Next to qualitative evalua-tions of the colonies our data suggest an average EDT-MSCclonogenic efficiency of 14 This value is higher than whatis described for BM-MSC and more similar or even superiorto what reported for AT-MSC that retained around 10 ofclonogenic precursors [25] (Table 3) In addition our EDT-MSC population seemed to have a higher clonogenic effi-ciency compared to those described by other groups dealingwith endometrial stromal cells whose values ranged from 1to 11 [12 24 26] While this may be due to donor-relatedissues such as age in vitro technical aspects including harvest-ing and preservation procedures that may also drive thesedifferences Dimitrov et al observed a tendency for these cellsto decrease in clonal efficiency with the increase of seedingdensity [26] Similarly other culture conditions may generatebetter performing EDT-MSC In this pilot study we testedtwo different growthmedia such as120572MEMwith 10FBS anda serum deprived medium Although these cells apparentlydid not shown particular nutritional requirements serum-containing medium proved to be the best choice ensuring abetter morphological appearance and an improved longevityPrevious studies reported the use of different media such asDMEMF-12 and Chang complete media often supplementedwith calf serum hormones and growth factors [12 14 26 27]This wide range of culture conditions is likely to impact the
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
6 BioMed Research International
(a) (b)
(c)
Figure 1 In vitro adherent cells from decidual tissues (a) Representative photomicrograph of spindle-shaped adherent cells isolated by120572MEM and 10 FBS (b) Cells isolated in vitro by serum-deprived medium (Quantum 333) at early passages (c) Another population of cellswas in vitro isolated contextually with EDT-MSCThese elements share similarities with endothelial cells forming a monolayer of polygonalsometimes binucleated cells Original magnification 100x
their ability to proliferate to differentiate and to be genemodified EDT-MSCs appear as a heterogeneous cells pop-ulation that can be safely and easily isolated by noninvasivemanner providing an expandable source of cells In culturethey show mesenchymal morphology and although somestudies suggest they could derive from bone marrow [23]EDT-MSCs are generally considered to originate from theshedding of endometrial stemprogenitor cells which aremainly resident both in the basalis layer and partially in thefunctionalis area of endometrium [24]
The presence of a uterine population of progenitor cellswould be consistent with the high tissue turn-over after eachmenstrual cycle However it is still uncertain howmany sub-populations of progenitors are present in the endometriumand what are their specific properties Therefore in this pilotstudy we began by addressing the existence of mesenchymalprogenitors in the decidual endometrial tissue
The initial in vitro approach was performed to evaluatethe clonogenic potential of these cells EDT-MSC seeded atclonal density revealed the presence of two distinct colonytypes which closely resembled the stromal cell clones isolatedby Chan et al from endometrium after hysterectomy [12]One was composed of smaller and densely packed cells andthe other was composed of larger and sparser elements Thisfinding seems peculiar for EDT-MSC and differs from col-onies established from BM-MSC that appear larger and
characterized by a central nucleus of cells surrounded by acrown of more sparse ones [18] Next to qualitative evalua-tions of the colonies our data suggest an average EDT-MSCclonogenic efficiency of 14 This value is higher than whatis described for BM-MSC and more similar or even superiorto what reported for AT-MSC that retained around 10 ofclonogenic precursors [25] (Table 3) In addition our EDT-MSC population seemed to have a higher clonogenic effi-ciency compared to those described by other groups dealingwith endometrial stromal cells whose values ranged from 1to 11 [12 24 26] While this may be due to donor-relatedissues such as age in vitro technical aspects including harvest-ing and preservation procedures that may also drive thesedifferences Dimitrov et al observed a tendency for these cellsto decrease in clonal efficiency with the increase of seedingdensity [26] Similarly other culture conditions may generatebetter performing EDT-MSC In this pilot study we testedtwo different growthmedia such as120572MEMwith 10FBS anda serum deprived medium Although these cells apparentlydid not shown particular nutritional requirements serum-containing medium proved to be the best choice ensuring abetter morphological appearance and an improved longevityPrevious studies reported the use of different media such asDMEMF-12 and Chang complete media often supplementedwith calf serum hormones and growth factors [12 14 26 27]This wide range of culture conditions is likely to impact the
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
BioMed Research International 7
2500 seeded EDT-MSC
(a) (b)
(c) (d) (e)
Figure 2 EDT-MSC clonogenic precursors (a) A representative T-25 culture flask with crystal violet-stained EDT-MSC colonies (b) and(c) EDT-MSC compact clones with small size cells (d) and (e) EDT-MSCs could also generate less populated colonies with larger cellularelements Original magnification 50x
times105
25
20
15
10
5
0
Cel
l yie
ld
0 24 48 72 96 120Time (h)
Sample 1Sample 2Sample 3
Figure 3 EDT-MSC proliferation Cell proliferation assays performed to assess EDT-MSC doubling time Doubling time for sample 1 was330 h for sample 2 was 219 h and for sample 3 was 279 h
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
8 BioMed Research International
120572MEM with 10 FBSPassage 3
100x
(a)
Quantum 333Passage 17
100x
(b)
Passage 26100x
120572MEM with 10 FBS
(c)
200xPassage 26
120572MEM with 10 FBS
(d)
Figure 4 120573-galactosidase staining (a) Activity of 120573-galactosidase at pH 6 was assessed on passage 3 EDT-MSC cultured in FBS resultingalmost negative for the presence of typical blue senescent cellsThe conspicuous number ofmitotic event is also visible highlighting an intenseproliferative activity (b) Cells grown in Quantum 333 underwent senescence at passage 17 and resulted in deep blue staining within most ofthe amplified EDT-MSC (c) Cells cultured in FBS were capable to reach passage 26 before showing senescence features such as blue staininglarge cytoplasm multinucleated elements and evident cytoskeleton (d) A detail of a senescent cell Original magnification 100x
in vitro behavior leading to different performance in ex vivopivotal parameters
Using a FBS-based medium EDT-MSC demonstrated aremarkable proliferation rate rapidly doubling their numberin less than 28 hrs This result agrees with other similarstudies which reported a doubling time between 18 and 36 hfor EDT-MSC [15 28] As seen in Table 3 similar attitude hasbeen also reported for dental pulp stem cells (DPSCs) andumbilical cord (UC) MSC that duplicate in approximately30 hrs considerably faster versus both BM-MSC and AT-MSC [29ndash31]
Beside this robust proliferative attitude we also observedthat plastic adherent fraction from shedded DT could beexpanded for more than 25 passages before reaching senes-cence a result once again higher than what reported forMSCfrom other sources As summarized in Table 3 BM-MSCand AT-MSC senescence has been reported from passages7 and 8 [32] while UC-MSC grow longer in culture untilpassage 16 [31] These findings reinforce the hypothesis thatEDT-MSC can duplicate for long time before senesce and are
consistent with reports asserting 25ndash30 population doublingsfor uterine-derived precursors [15 28 33]This unique featurefit well with the high rate of renewal retained in vivo by theendometrium leading to the conclusion that this new MSCsource could have a tremendous potential for large-scale clin-ical applications in regenerative medicine
In this work EDT-MSC showed other similarities withMSC from the other sources and in particular under theimmunophenotypical point of view satisfying recognizedphenotypic criteria for MSC [22] The biomarker profile ascompared in Table 2 shows a close resemblance of EDT-MSC with other sources MSC expressing high levels ofCD73 and CD90 and being negative for CD14 CD45 CD31and HLA-DR Other markers have been additionally eval-uated CD56 also known as neural cell adhesion molecule(NCAM) is a glycoprotein expressed by neuroectodermal-derived cells such as neurons and glia but also by skeletalmuscle NK cells and activated T cells [34] Among our EDT-MSC CD56 expression has been extremely variable (13ndash45) and this may support the observation made by others
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
BioMed Research International 9
SSC-
A
FSC-A
(a)CD
45
HLADR
(b)
CD45
350300250200150100
500
36
(c)
HLADR
200
150
100
50
0
09
(d)
CD14
CD31
(e)
CD31
350300250200150100
500
05
(f)
CD14
600
500
400
300
200
100
0
08
(g)
CD10
5
CD73
(h)
CD73
150
100
50
0
973
(i)
CD105
64250
200
150
100
50
0
(j)
SSEA
4
CD146
(k)
SSEA4
1300250200150100
500
(l)
CD146
823
75
100
25
50
0
(m)
CD31
CD56
(n)
CD56
150
100
50
0
174
(o)
CD90
CD105
(p)
CD90
200
150
100
50
0
974
(q)
Figure 5 EDT-MSCs express typicalMSCmarkers Representative FACS analyses of EDT-MSC immunophenotype Percentages are referringto the average positivity for each marker Cells were gated out of 7-AAD positivity to exclude nonviable elements
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
10 BioMed Research International
Table 2 Immunophenotype of MSC from different sources
Marker CD14 CD45 CD73 CD90 CD105 CD146 CD31 HLA-DR ReferencesBM-MSC minus minus +++ +++ +++ ++ minus minus [43 44]PB-MSC minus minus NA +++ ++ NA minus minus [45]DPSC minus minus +++ +++ +++ NA minus minus [30]UC-MSC minus minus +++ +++ +++ NA minus minus [2 43]AT-MSC minus minus +++ +++ ++ NA minus minus [2 43]EDT-MSC minus minus +++ +++ ++ +++ minus minus Dagger
Comparison between positive and negative MSC surface markers from different sources including EDT-MSC Variable expression is highlighted with plusmn BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulp stromal cells UC-MSC umbilical cord MSC AT-MSC adipose tissue MSCEDT-MSC decidual tissue MSC DaggerOriginal data from this study references
Posit
ivity
()
100
80
60
40
20
0
CD45
HLA
DR
CD31
CD56
CD14
CD90
CD73
CD10
5
CD14
6
SSEA
-4
Sample 1Sample 2Sample 3
Figure 6 Overall EDT-MSC antigens expression after passage 1 invitro Marker expressions are here reported as average percentage(plusmnstandard deviation) of each tested sample
exploiting CD56 expression to discriminate distinct marrowMSC subpopulations [35 36] Moreover we evaluated theexpression of SSEA-4 a glycolipid recently proposed to bea novel marker for BM-MSC isolation and that is expressedby teratocarcinoma cells and embryonic stem cells [37]EDT-MSC demonstrated a noticeable expression of SSEA-4 that while reaching 194 appeared far from the 90positivity reported by others on decidual-derived cells [15]However different laboratories reported discrepancies onEDT-MSC phenotype [15 28 38] and this could be likely dueto the heterogeneity of menstrual blood cell population todifferences in cell selection and as stated to culture condi-tions [13] In addition the relatively short time from the earlyevidence of these progenitors does not help in the pheno-typical standardization and certainly will require deeper andmulticentric studies
Multipotency is another key feature ofMSCs and all EDT-MSC lines showed the in vitro capacity to differentiate intothreemesodermal lineages such as osteoblast able to generatemineralized matrix lipid vacuole-containing adipocyte andmatrix-producing chondrocytes Additional experimentswillbe necessary to confirm this ability prompting their broadintroduction in regenerativemedicine applications Howeverwe have here begun to assess the capacity of these cells
for robust osteogenic commitment confirmed by the differ-entiation data obtained by staining for mineralized matrixwe observed a relevant increase in the expression of typicalosteoblast-associated genes such as alkaline phosphataseosteocalcin and collagen 1A2
Regenerative therapies based on stem cells and morespecifically on MSC infusion may counteract tissue-losingstates and may also be useful to substitute the congenitalabnormal stem cell compartment in order to regeneratehealthy functional cells In this perspective ease of supplywide window of harvesting and availability of EDT-MSCwould allow autologous or ultimately allogeneic EDT-MSCtransplantation These properties have already been explo-ited and EDT-MSC have demonstrated a therapeutic effectin a murine model of muscular dystrophy [38] and a neuro-protective effect in an experimental stroke model [27] More-over endometrial-derived cells seem to have robust anti-inflammatory and immunomodulatory properties similarlyto other MSCs [27 39] For this reason they were intraven-ously and intrathecally transplanted in 4 patients with mult-iple sclerosis notably without arising immunological reac-tions or other adverse effects and with a clinical benefit thatshall be carefully assessed [40]
Another original result we here report is the high trans-duction efficiency of EDT-MSC by a GFP-carrying retroviralvector The procedure generated exogenous gene expressionin more than 80 of the cells and to our knowledge this isone of the first reports of such highly efficient genetic mani-pulation of EDT-MSC by a single GFP-carrying vector pro-viding a basis for use of these cells in gene therapy approachesincluding targeted cancer treatment or their use as cell vec-tors for peptides or trophic factors Having extensively cha-racterized EDT-MSCwe presume that these cellsmay be use-ful for a more efficient delivery of proapoptotic moleculesthanks to the high transduction efficiency and the rapid pro-liferationThismay allow amore targeted approach andbetterpersistence of the cells in the tumor microenvironment
Starting from these findings future studies shall focuson the characterisation of the heterogeneous cell populationsidentifiable in menstrual decidual tissue investigating theexistence of different progenitor subtypes as well as under-standing the origin of these cells More reliable techniques toisolate and cultivate pure populations of EDT-MSC should bedeveloped and standardized in parallel to allow a rapid intro-duction of this promising therapeutic tool into the clinic
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
BioMed Research International 11
100x 100x 200xControl Induced
(a)
50x 50x 100x
(b)
100x 100x 200x
(c)
Figure 7 EDT-MSCs differentiate into committed mesenchymal tissues (a) Oil Red O staining in adipogenic induced and control EDT-MSC The red-brown vacuoles demonstrate differentiation into fat-producing cells Original magnification 100x (b) Alizarin Red stainingshowing calcium deposits in induced cells when compared with noninduced controls Original magnification 50x (c) Alcian Blue stainingoutlining sulfated proteoglycans expression suggesting differentiation into a chondrogenic tissue Original magnification 100x
Table 3 Growth characteristics of MSC from different sources
Cell type Source Isolation yield Doubling time Clonogenic efficiency Senescence passageBM-MSC Bone marrow aspirate 60ndash600 cells from 1mL of blood [41] 612 h [29] 39lowast 7 [32]
PB-MSC Peripheral blood 1 to 13 MSC from 1 million ofmononuclear cells [11] NA NA NA
DPSC Deciduous and adult teeth NA 30 h [30] NA NA
UC-MSC Umbilical cord of newborns 5 sdot 104ndash5 sdot 105 cells from 1 cm3 of
Comparison between growth parameters of MSC from different sources BM-MSC bone marrow MSC PB-MSC peripheral blood MSC DPSC dental pulpstromal cells UC-MSC umbilical cordMSC AT-MSC adipose tissueMSC EDT-MSC decidual tissueMSClowastUnpublished data Daggeroriginal data from this studyreferences
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
12 BioMed Research International
ALP
COLL
1A2
OC
50
45
40
35
30
25
20
15
10
5
0
Relat
ive e
xpre
ssio
n
Figure 8 qRT-PCR for osteoblast-associated genes The relative quantity of osteoblast-associated transcripts in induced EDT-MSC versusnoninduced controls revealed an increased expression of the considered markers All values were statistically significant (119875 lt 005) ALPalkaline phosphatase COLL1A2 collagen 1A2 and OC osteocalcin
SSC-
A
FSC-A
(a)
Control
SSC-
A
GFP-A
(b)
Transduced
839
SSC-
A
GFP-A
(c)
Figure 9 GFP expression of transduced andwild-type EDT-MSC Representative FACS analyses assessingGFP positivity of wild-type controland transduced EDT-MSC
5 Conclusions
This work highlights crucial features of a novel and still notcompletely understood population of mesenchymal progeni-tors isolated form endometrial decidual tissue Cells grownin vitro were characterized by a rapid proliferation a highclonogenic potential and a long-term survival Advantagesin comparison with MSC from other sources include agreater ease of supply and the protracted availability during awomanrsquos lifetime with the additional benefit of deriving stemcells from a waste tissue thus avoiding critical ethical issuesIn the light of their capacity to differentiate intomesenchymaltissues and their propensity to undergo geneticmanipulationEDT-MSCs hold promise for novel therapeutic approachesfor still incurable and highly disabling diseases
Conflict of Interests
The authors do not have direct financial conflicts of interest
Authorsrsquo Contribution
F Rossignoli and A Caselli are equally contributing authors
Acknowledgments
This work was supported in part by the Italian Ministryof Health ldquoBando Giovani Ricercatori 2008rdquo (MD) ldquoBandoCellule Staminali 2008rdquo (MD PP) Regione Emilia Romagna(PP MD) and the Associazione ASEOP (PP)
References
[1] D Campioni and F Lanza ldquoCellule mesenchimali stromali car-atterizzazione immunofenotipica e funzionalerdquoLettereGIC vol20 pp 13ndash19 2011
[2] W Wagner F Wein A Seckinger et al ldquoComparative charac-teristics of mesenchymal stem cells from human bone marrow
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
BioMed Research International 13
adipose tissue and umbilical cord bloodrdquo Experimental Hema-tology vol 33 no 11 pp 1402ndash1416 2005
[3] HM Lazarus S E Haynesworth S L Gerson N S Rosenthaland A I Caplan ldquoEx vivo expansion and subsequent infusionof human bone marrow-derived stromal progenitor cells (mes-enchymal progenitor cells) implications for therapeutic userdquoBone Marrow Transplantation vol 16 no 4 pp 557ndash564 1995
[4] L M Ball M E Bernardo H Roelofs et al ldquoCotransplanta-tion of ex vivo-expanded mesenchymal stem cells accelerateslymphocyte recovery and may reduce the risk of graft failure inhaploidentical hematopoietic stem-cell transplantationrdquo Bloodvol 110 no 7 pp 2764ndash2767 2007
[5] E M Horwitz and M Dominici ldquoHow do mesenchymalstromal cells exert their therapeutic benefitrdquo Cytotherapy vol10 no 8 pp 771ndash774 2008
[6] A Y Khakoo S Pati S A Anderson et al ldquoHuman mesenchy-mal stem cells exert potent antitumorigenic effects in a modelof Kaposirsquos sarcomardquoThe Journal of Experimental Medicine vol203 no 5 pp 1235ndash1247 2006
[7] G Grisendi R Bussolari L Cafarelli et al ldquoAdipose-derivedmesenchymal stem cells as stable source of tumor necrosisfactor-related apoptosis-inducing ligand delivery for cancertherapyrdquo Cancer Research vol 70 no 9 pp 3718ndash3729 2010
[8] R M Dwyer S Khan F P Barry T OrsquoBrien and M J KerinldquoAdvances inmesenchymal stem cell-mediated gene therapy forcancerrdquo Stem Cell Research amp Therapy vol 1 no 3 article 252010
[9] S Kidd L Caldwell M Dietrich et al ldquoMesenchymal stro-mal cells alone or expressing interferon-120573 suppress pancreatictumors in vivo an effect countered by anti-inflammatory treat-mentrdquo Cytotherapy vol 12 no 5 pp 615ndash625 2010
[10] G Chamberlain J Fox B Ashton and J Middleton ldquoConcisereview mesenchymal stem cells their phenotype differentia-tion capacity immunological features and potential for hom-ingrdquo Stem Cells vol 25 no 11 pp 2739ndash2749 2007
[11] R Hass C Kasper S Bohm and R Jacobs ldquoDifferent popula-tions and sources of human mesenchymal stem cells (MSC)a comparison of adult and neonatal tissue-derived MSCrdquo CellCommunication and Signaling vol 9 article 12 2011
[12] R W S Chan K E Schwab and C E Gargett ldquoClonogenicityof human endometrial epithelial and stromal cellsrdquo Biology ofReproduction vol 70 no 6 pp 1738ndash1750 2004
[13] C E Gargett and H Masuda ldquoAdult stem cells in the endomet-riumrdquoMolecular Human Reproduction vol 16 no 11 Article IDgaq061 pp 818ndash834 2010
[14] C E Gargett K E Schwab R M Zillwood H P T Nguyenand D Wu ldquoIsolation and culture of epithelial progenitors andmesenchymal stem cells from human endometriumrdquo Biology ofReproduction vol 80 no 6 pp 1136ndash1145 2009
[15] A N Patel E Park M Kuzman F Benetti F J Silva and JG Allickson ldquoMultipotent menstrual blood stromal stem cellsisolation characterization and differentiationrdquo Cell Transplan-tation vol 17 no 3 pp 303ndash311 2008
[16] R A Musina A V Belyavski O V Tarusova E V Solovyovaand G T Sukhikh ldquoEndometrial mesenchymal stem cells iso-lated from the menstrual bloodrdquo Bulletin of ExperimentalBiology and Medicine vol 145 no 4 pp 539ndash543 2008
[17] N DrsquoSouza J S Burns G Grisendi et al ldquoMSC and tumorshoming differentiation and secretion influence the therapeuticpotentialrdquo Advances in Biochemical EngineeringBiotechnology2012
[18] H Castro-Malaspina R E Gay G Resnick et al ldquoCharacteri-zation of human bone marrow fibroblast colony-forming cells(CFU-F) and their progenyrdquo Blood vol 56 no 2 pp 289ndash3011980
[19] G P Dimri X Lee G Basile et al ldquoA biomarker that identifiessenescent human cells in culture and in aging skin in vivordquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 92 no 20 pp 9363ndash9367 1995
[20] G Grisendi C Anneren L Cafarelli et al ldquoGMP-manufac-tured density gradient media for optimized mesenchymal stro-malstem cell isolation and expansionrdquo Cytotherapy vol 12 no4 pp 466ndash477 2010
[21] J C Marx J A Allay D A Persons et al ldquoHigh-efficiencytransduction and long-term gene expression with a murinestem cell retroviral vector encoding the green fluorescent pro-tein in humanmarrow stromal cellsrdquoHumanGeneTherapy vol10 no 7 pp 1163ndash1173 1999
[22] M Dominici K Le Blanc I Mueller et al ldquoMinimal crite-ria for defining multipotent mesenchymal stromal cells TheInternational Society for Cellular Therapy position statementrdquoCytotherapy vol 8 no 4 pp 315ndash317 2006
[23] H Du and H S Taylor ldquoContribution of bone marrow-derivedstem cells to endometrium and endometriosisrdquo Stem Cells vol25 no 8 pp 2082ndash2086 2007
[24] K E Schwab and C E Gargett ldquoCo-expression of two perivas-cular cell markers isolates mesenchymal stem-like cells fromhuman endometriumrdquoHuman Reproduction vol 22 no 11 pp2903ndash2911 2007
[25] K Bieback A Hecker T Schlechter et al ldquoReplicative agingand differentiation potential of human adipose tissue-derivedmesenchymal stromal cells expanded in pooled human or fetalbovine serumrdquo Cytotherapy vol 14 pp 570ndash583 2012
[26] R Dimitrov T Timeva D Kyurkchiev et al ldquoCharacterizationof clonogenic stromal cells isolated fromhuman endometriumrdquoReproduction vol 135 no 4 pp 551ndash558 2008
[27] C V Borlongan Y Kaneko M Maki et al ldquoMenstrual bloodcells display stem cell-like phenotypic markers and exert neu-roprotection following transplantation in experimental strokerdquoStem Cells and Development vol 19 no 4 pp 439ndash451 2010
[28] X Meng T E Ichim J Zhong et al ldquoEndometrial regenerativecells a novel stem cell populationrdquo Journal of TranslationalMedicine vol 5 article 57 2007
[29] L Peng Z Jia X Yin et al ldquoComparative analysis of mesenchy-mal stem cells from bonemarrow cartilage and adipose tissuerdquoStem Cells and Development vol 17 no 4 pp 761ndash773 2008
[30] B C Perry D Zhou XWu et al ldquoCollection cryopreservationand characterization of human dental pulp-derived mesenchy-mal stem cells for banking and clinical userdquo Tissue EngineeringC vol 14 no 2 pp 149ndash156 2008
[31] L L Lu Y J Liu S G Yang et al ldquoIsolation and characteri-zation of human umbilical cord mesenchymal stem cells withhematopoiesis-supportive function and other potentialsrdquo Hae-matologica vol 91 no 8 pp 1017ndash1028 2006
[32] S Kern H Eichler J Stoeve H Kluter and K BiebackldquoComparative analysis of mesenchymal stem cells from bonemarrow umbilical cord blood or adipose tissuerdquo StemCells vol24 no 5 pp 1294ndash1301 2006
[33] N Hida N Nishiyama S Miyoshi et al ldquoNovel cardiac pre-cursor-like cells from human menstrual blood-derived mes-enchymal cellsrdquo Stem Cells vol 26 no 7 pp 1695ndash1704 2008
[34] H Zola B Swart I Nicholson and E Voss Eds Leukocyte andStromal Cell MoleculesmdashThe CDMarkers Wiley-Liss 2007
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006
14 BioMed Research International
[35] V L Battula S Treml PM Bareiss et al ldquoIsolation of function-ally distinct mesenchymal stem cell subsets using antibodiesagainst CD56 CD271 and mesenchymal stem cell antigen-1rdquoHaematologica vol 94 no 2 pp 173ndash184 2009
[36] H J Buhring S Treml F Cerabona P De Zwart L Kanz andM Sobiesiak ldquoPhenotypic characterization of distinct humanbone marrow-derived MSC subsetsrdquo Annals of the New YorkAcademy of Sciences vol 1176 pp 124ndash134 2009
[37] E J Gang D Bosnakovski C A Figueiredo J W Visser andR C R Perlingeiro ldquoSSEA-4 identifies mesenchymal stem cellsfrom bone marrowrdquo Blood vol 109 no 4 pp 1743ndash1751 2007
[38] CHCui TUyama KMiyado et al ldquoMenstrual blood-derivedcells confer human dystrophin expression in the murine modelof duchenne muscular dystrophy via cell fusion and myogenictransdifferentiationrdquoMolecular Biology of the Cell vol 18 no 5pp 1586ndash1594 2007
[39] M P Murphy H Wang A N Patel et al ldquoAllogeneic endome-trial regenerative cells an rdquoOff the shelf solutionrdquo for criticallimb ischemiardquo Journal of Translational Medicine vol 6 article45 2008
[40] Z Zhong A N Patel T E Ichim et al ldquoFeasibility investigationof allogeneic endometrial regenerative cellsrdquo Journal of Transla-tional Medicine vol 7 article 15 2009
[41] M F Pittenger A M Mackay S C Beck et al ldquoMultilineagepotential of adult human mesenchymal stem cellsrdquo Science vol284 no 5411 pp 143ndash147 1999
[42] R C Schugar S M Chirieleison K E Wescoe et al ldquoHighharvest yield high expansion and phenotype stability of CD146mesenchymal stromal cells fromwhole primitive humanumbil-ical cord tissuerdquo Journal of Biomedicine and Biotechnology vol2009 Article ID 789526 11 pages 2009
[43] D T Covas R A Panepucci A M Fontes et al ldquoMultipotentmesenchymal stromal cells obtained from diverse human tis-sues share functional properties and gene-expression profilewith CD146+ perivascular cells and fibroblastsrdquo ExperimentalHematology vol 36 no 5 pp 642ndash654 2008
[44] D Campioni S Moretti L Ferrari M Punturieri G L Cas-toldi and F Lanza ldquoImmunophenotypic heterogeneity of bonemarrow-derived mesenchymal stromal cells from patients withhematologic disorders correlation with bonemarrowmicroen-vironmentrdquo Haematologica vol 91 no 3 pp 364ndash368 2006
[45] I Kassis L Zangi R Rivkin et al ldquoIsolation of mesenchymalstem cells from G-CSF-mobilized human peripheral bloodusing fibrin microbeadsrdquo Bone Marrow Transplantation vol 37no 10 pp 967ndash976 2006






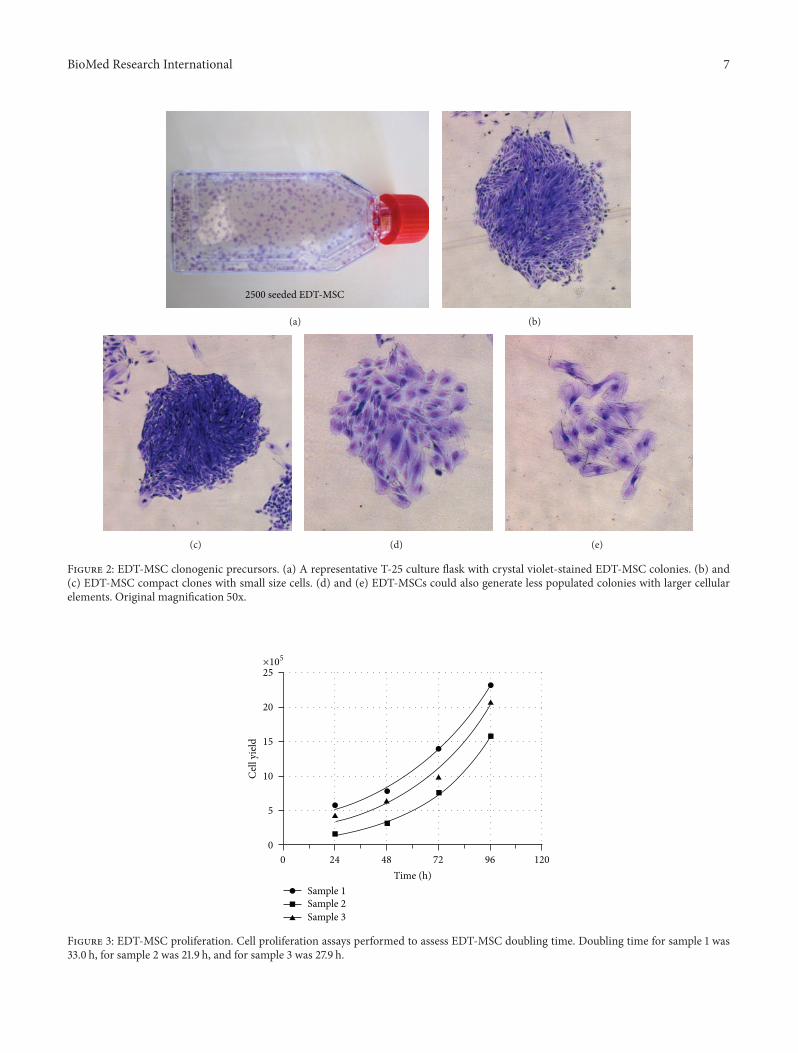
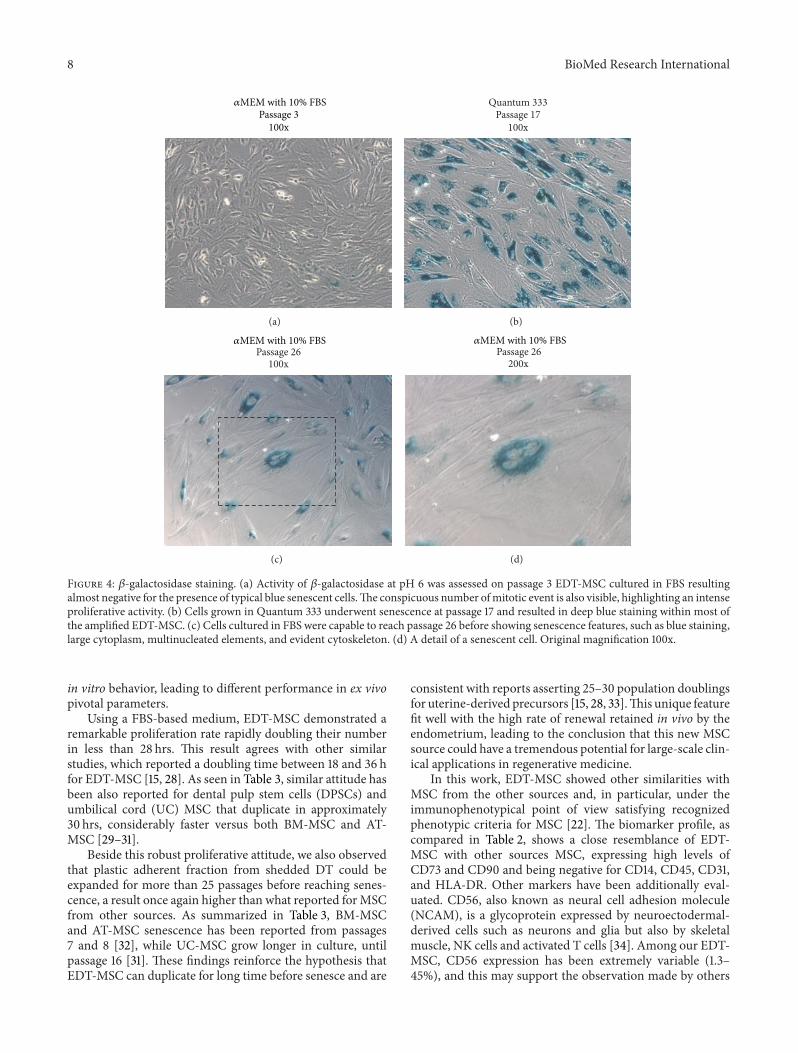
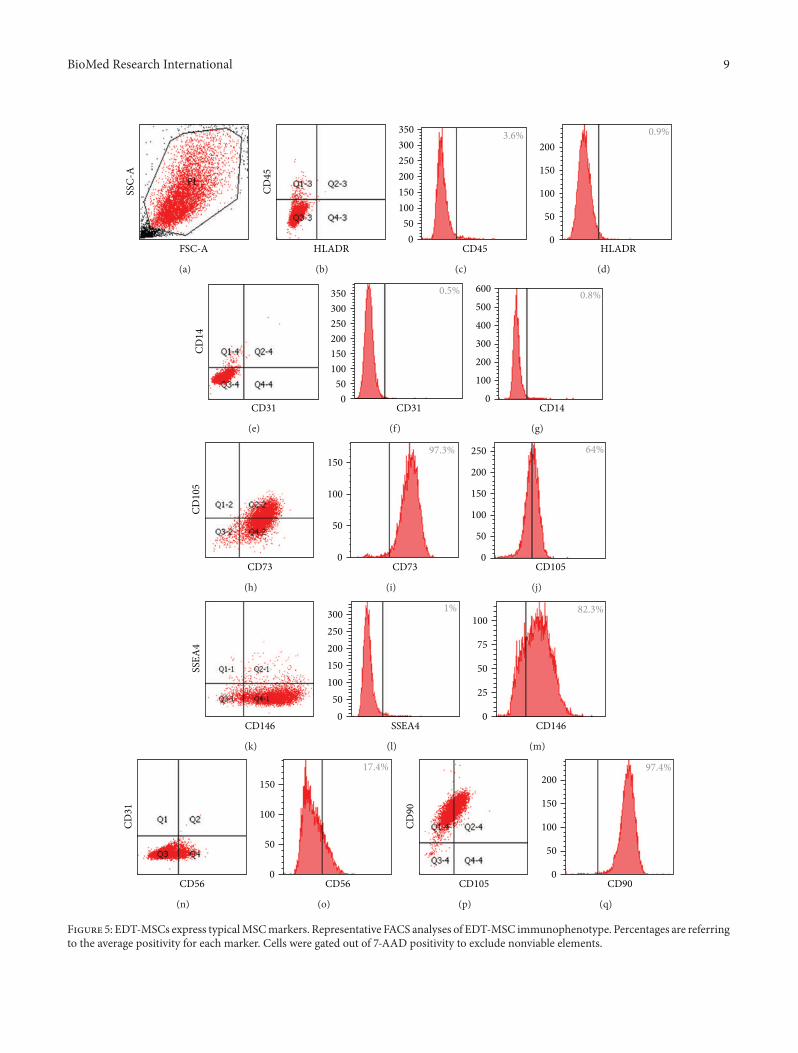
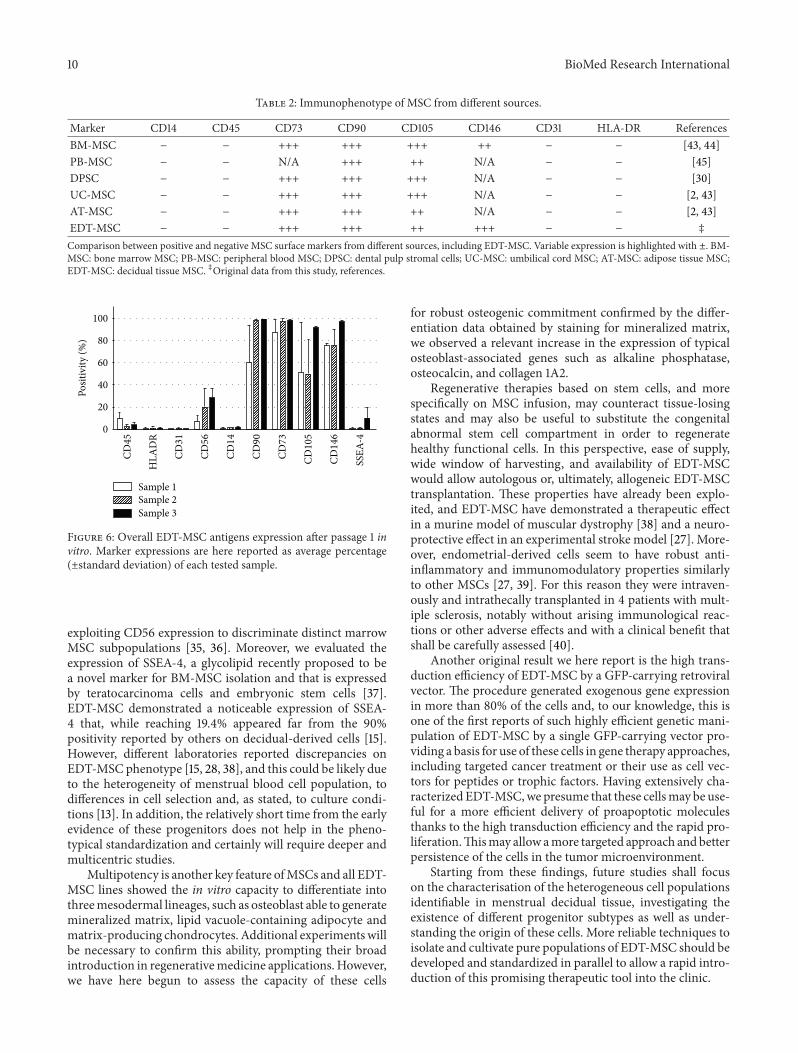
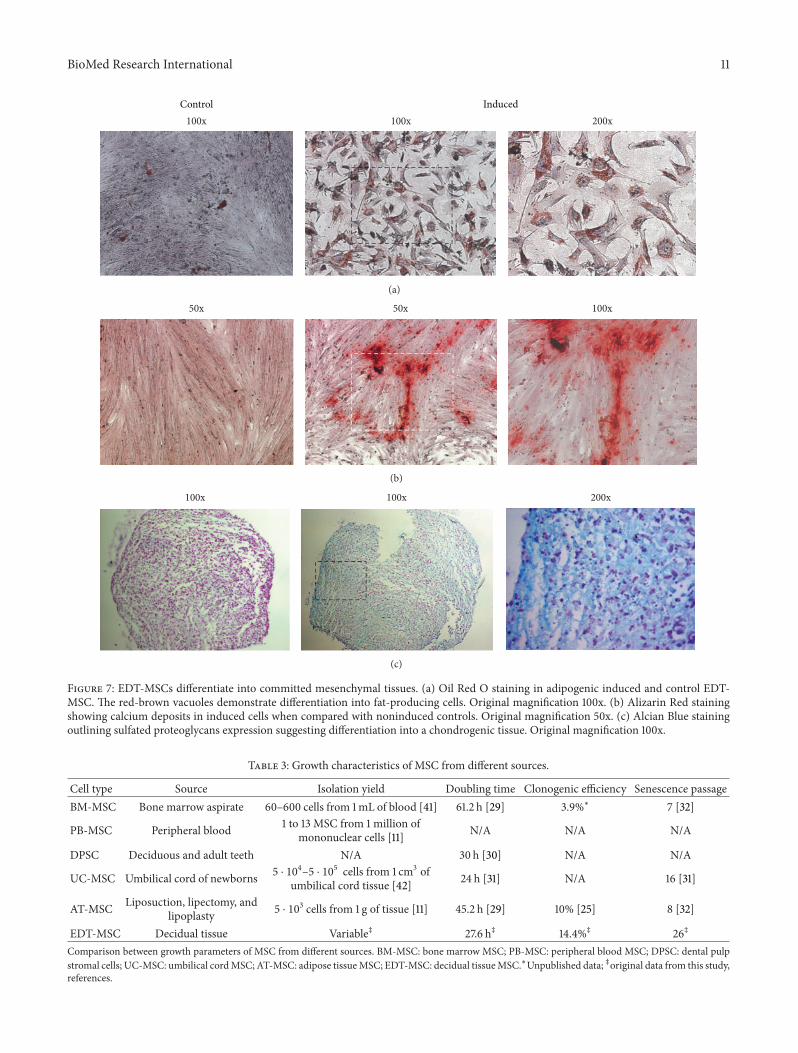







![RESEARCH Open Access Human multipotent stromal cells ...mesenchymal lineage cell types including bone, cartilage, adipose tissue, muscle and tendon [4]. MSCs have been isolated from](https://static.documents.pub/doc/80x56/5e6b65b920f9b208741edf9a/research-open-access-human-multipotent-stromal-cells-mesenchymal-lineage-cell.jpg)












