J. Exp. Biol. (1970), sa, 95-107 Q^ With 5 text-figures Printed in Great Britain THE DIRECTION OF THE THRUST PRODUCED BY THE HETEROCERCAL TAILS OF TWO DISSIMILAR ELASMOBRANCHS: THE PORT JACKSON SHARK, HETERODONTUS PORTUSJACKSONI (MEYER), AND THE PIKED DOGFISH, SQUALUS MEGALOPS (MACLEAY) BY J. R. SIMONS School of Biological Sciences, The University of Sydney {Received 17 June 1969) It is generally held that the heterocercal tail of elasmobranchs (as well as certain primitive osteichthyans) generates a thrust which is angled upwards. The thrust is thought of as resolved into a horizontal component which drives the animal forward, and a vertical component which aids in the levitation of the creature in the water. To date, all attempts to demonstrate the 'lift' or 'upthrust' of such a tail have, either in qualitative (Grove & Newell, 1936; Affleck, 1950) or in quantitative approaches (Alexander, 1965), succeeded only in showing that the heterocercal tail produces a torque or moment of force about the animal's centre of gravity. A torque, being the product of a force and a distance, yields no information about the force producing it unless information is also known about the distance involved. It is quite conceivable, for instance, that the moment of negative pitch (Young, 1955) by which the tail is raised and the head depressed in elasmobranchs may, in fact, be produced by the tail generating a single horizontal force, provided the line of action of such a force passed dorsal to the animal's centre of gravity. It is thus possible that the action of the heterocercal tail may be to displace a horizontal thrust some distance above the general longitudinal axis of the body and yet still produce negative pitch. This possibility has never been excluded in any previous discussion of the problem of heterocercy and therefore the terms 'lift' and 'upthrust' remain obscure or meaning- less. Resolution of the doubt requires the demonstration of the line of action of the force rather than the torque it produces. It occurred to the present author that the direction and location of the thrust generated by any tail could be established empirically if one were to modify and extend the method used by Alexander (1965) in his study of the 'lift' produced by the tails of Scyliorhinus canicula and Galeorhinus galeus. Consequently, the work reported on below was undertaken. In it the position and direction of the thrust produced by the heterocercal tails of two dissimilar elasmobranchs were determined. The part played by the ventral lobe in modifying the direction of the thrust was also determined.
Transcript
J. Exp. Biol. (1970), sa, 95-107 Q^
With 5 text-figures
Printed in Great Britain
THE DIRECTION OFTHE THRUST PRODUCED BY THE HETEROCERCAL TAILS
OF TWO DISSIMILAR ELASMOBRANCHS: THE PORTJACKSON SHARK, HETERODONTUS PORTUSJACKSONI
(MEYER), AND THE PIKED DOGFISH, SQUALUSMEGALOPS (MACLEAY)
BY J. R. SIMONSSchool of Biological Sciences, The University of Sydney
{Received 17 June 1969)
It is generally held that the heterocercal tail of elasmobranchs (as well as certainprimitive osteichthyans) generates a thrust which is angled upwards. The thrust isthought of as resolved into a horizontal component which drives the animal forward,and a vertical component which aids in the levitation of the creature in the water.
To date, all attempts to demonstrate the 'lift' or 'upthrust' of such a tail have,either in qualitative (Grove & Newell, 1936; Affleck, 1950) or in quantitativeapproaches (Alexander, 1965), succeeded only in showing that the heterocercal tailproduces a torque or moment of force about the animal's centre of gravity.
A torque, being the product of a force and a distance, yields no information aboutthe force producing it unless information is also known about the distance involved.It is quite conceivable, for instance, that the moment of negative pitch (Young, 1955)by which the tail is raised and the head depressed in elasmobranchs may, in fact, beproduced by the tail generating a single horizontal force, provided the line of actionof such a force passed dorsal to the animal's centre of gravity. It is thus possible thatthe action of the heterocercal tail may be to displace a horizontal thrust some distanceabove the general longitudinal axis of the body and yet still produce negative pitch.This possibility has never been excluded in any previous discussion of the problem ofheterocercy and therefore the terms 'lift' and 'upthrust' remain obscure or meaning-less. Resolution of the doubt requires the demonstration of the line of action of theforce rather than the torque it produces.
It occurred to the present author that the direction and location of the thrustgenerated by any tail could be established empirically if one were to modify and extendthe method used by Alexander (1965) in his study of the 'lift' produced by the tailsof Scyliorhinus canicula and Galeorhinus galeus. Consequently, the work reported onbelow was undertaken. In it the position and direction of the thrust produced by theheterocercal tails of two dissimilar elasmobranchs were determined. The part playedby the ventral lobe in modifying the direction of the thrust was also determined.
96 J. R. SIMONS
MATERIAL
The species of elasmobranchs used in this study were the Port Jackson shark(Heterodontus portusjacksoni) and the piked dogfish (Squalus megalops).
The tails, together with the final 6 to 8 cm. of the trunk region, were cut from freshspecimens as they became available. These were immediately mounted on theapparatus (see below) and the experiment was carried out. After all the relevantinformation on the whole tail had been gathered, the ventral lobe of the tail wasremoved and the experiment was repeated.
The tails of six specimens of each species were used.
APPARATUS
The apparatus employed was a modified version of one (originally built for classuse) replicating that designed and used by Alexander (1965) to whose paper thereader is referred for full details (Fig. 1).
The modifications made were:(a) Raising the fulcrum, about which the crossbar holding the tail moved, some
distance (32 cm.) above the horizontal midline of the crossbar.
Fig. 1. Diagram of the apparatus for determining the location and direction of the thrustdeveloped by a tail, as modified from the original design of Alexander (1965). (a) Fulcrum con-sisting of two roller bearings. (6) Graduated arc indicating any deflexion of the lower assemblyfrom its equilibrium position, (c) Cross arm of lower assembly along which weights may be sus-pended. Used in conjunction with the lower counterpoise to determine moments about thefulcrum more precisely, (d) Screw and slip-ring allowing inversion of mounted tail, (e) Remov-able element of horizontal arm.
The thrust of heterocercal tails 97
(b) Making provision for the tail to be mounted initially upright and subsequentlyupside down without the necessity for removing the tail from the assembly.
(c) Making provision for the tail to be displaced a known distance (5 cm.) from itsinitial position with respect to the vertical driving shaft of the apparatus.
These modifications allowed the tail to be mounted on the apparatus in fourpositions: (1) Upright (sagittal plane vertical) at a distance 6 cm. from the centrelineof the vertical driving shaft. (2) Upside down at the same distance. (3) Upright at adistance 11 cm. from the centreline of the vertical shaft. (4) Upside down at this samedistance.
Any tendency of the tail and crossbar, as it was rotated, to move away from ahorizontal equilibrium position was indicated by a pointer moving over a graduatedarc held on the upper part of the vertical shaft.
With these differences in construction the apparatus was used basically as describedby Alexander.
THEORY OF THE METHOD
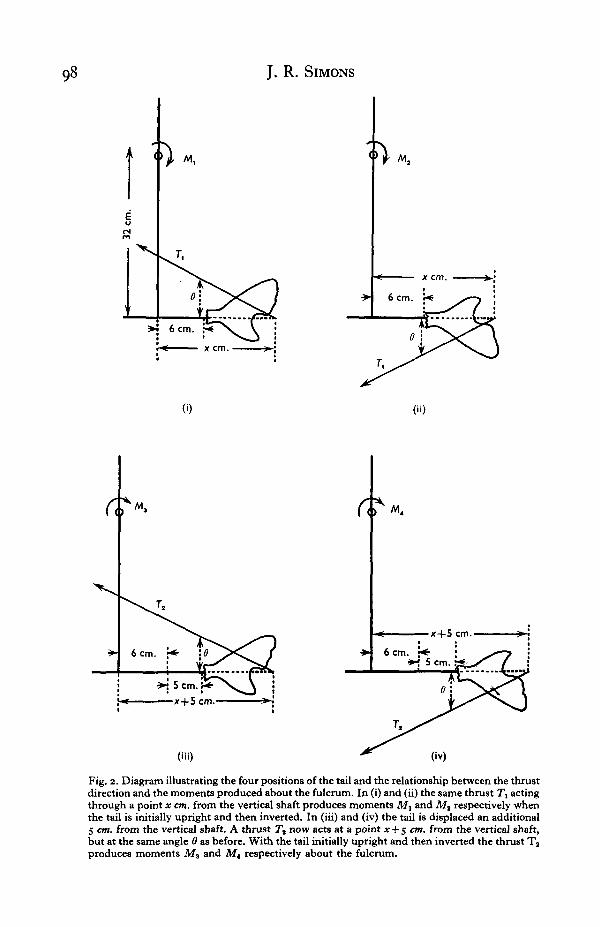
The arrangement of the tail, the thrusts and moments generated in the fourdifferent positions of mounting are shown in diagrammatic form in Fig. 2(i), (ii),(iii) and (iv).
In (i) and (ii) the tail is mounted at the same distance from the centre line of thevertical shaft but in the upright and inverted position respectively. If, in thesepositions, the vertical shaft is rotated at a given speed, the tail will develop a particularthrust represented by a single resultant force 7\. Let the thrust be inclined at theangle 6 to the horizontal and intersect the horizontal (i.e. the midline of the crossbarand hence the line of the vertebral column as it passes into the tail) at a distance x cm.from the centreline of the vertical shaft.
The thrust generates moments about the fulcrum A. Let the moments generatedwhen the tail is upright be Mt and those generated when the tail is upside down beM8.
From the geometry of the arrangement it can be shown that
Mx = ( a - # t a n d) Tx cos 6and
M2 = (a + x tan 6) 7\ cos 6
where a is the distance (32 cm.) of the fulcrum A above the midline of the crossbar.The magnitudes of 7\ and hence M1 and M2 depend upon the speed of rotation of
the vertical shaft, but the ratio Af1/M2 is a constant, provided always that both mem-bers of any pair of Mx and M2 are determined at the same speed of rotation.
Let kx denote the ratio M1/Ma and rearrange the relationship
, _ a — x tan d1 ~ a + a t a n 6
to the form
x tan 6 = -±—T~ (1)1 +«i
7 EXB 52
J. R. SIMONS
0) (ii)
(iii)
Fig. 2. Diagram illustrating the four positions of the tail and the relationship between the thrustdirection and the moments produced about the fulcrum. In (i) and (ii) the same thrust 7\ actingthrough a point x cm. from the vertical shaft produces moments M, and Mt respectively whenthe tail is initially upright and then inverted. In (iii) and (iv) the tail is displaced an additional5 cm. from the vertical shaft. A thrust Tt now acts at a point * + 5 cm. from the vertical shaft,but at the same angle 6 as before. With the tail initially upright and then inverted the thrust Taproduces moments M3 and M4 respectively about the fulcrum.
The thrust of heterocercal tails 99
Consider the situation (iii) and (iv). Here the line of thrust is still angled at 6°, butit now intersects the horizontal at a distance (*+5) cm. from the vertical shaft's centre.If the thrust developed in the new situation is Ta and the respective moments M3 andMt, and if the ratio MzjMi is denoted by k2, it can be shown in steps paralleling thoseexplained above that
, _ a-(x+5) tan 62 ~ a + (* + 5)tan 6
Rearranging this relationship and substituting a(i—k^)\(\+h^) for* tan 6 from(1) above yields first the solution
tan 0 = 7 ^ ^
and subsequentlya i—k.
tan 0i+&x
The solution of these two values enables one to locate the line of thrust with respectto the position of the tail on the apparatus.
THE METHOD AS PRACTISED
The various moments Mlt M2, M3 and Mt were determined as means of themoments generated by alternately rotating the vertical shaft in clockwise and anti-clockwise directions. Each moment was also determined at five different speeds ofrotation. Thus, five values of Mx were obtained and each of these was paired by avalue M2 determined at the same speeds. A similar series of paired values of M3 andMt was determined.
The values of the ratios MJM2 and M3/M4 were computed from each pair ofmoments and thus the average values of £x and k2 were obtained. These values werethen substituted in the equations given above to arrive at the solutions for tan 6 and x.
Since the tails used were of different sizes, the value x was expressed as a ratio (r)of the overall length of the tail as measured along the horizontal extension of thevertebral column. By this simple device it becomes possible to speak in averageterms about the location of the line of thrust in the ' average' tail.
As has been mentioned, after the tail had been subjected to the procedure describedabove, the ventral lobe was cut away and the procedure was repeated in order todetermine what difference resulted from the removal of the lobe.
RESULTS
(a) Whole tails
The values of the angle of inclination (6) and the relative position (r) of the line ofthrust determined for the tails of six Port Jackson sharks and six piked dogfish aregiven in Table 1.
As may be seen, the values of the angles of inclination found for the dogfish wereconsistently higher than those found for Heterodontus. In average terms the line ofthrust developed by the tail of Heterodontus is inclined upwards at about 120 to the
7-2
ioo J. R. SIMONS
horizontal, whereas that of Squalus approximates 26*5°. Using the mean values of 6and r, the results may be expressed in diagrams of 'average' tails as shown in Fig.3A and B.
Table 1. The values of the angles of inclination 6 to the horizontal and theproportional distances (r) of tail length intersected along the horizontal by theline of thrust found for six whole tails each of Heterodontus portusjacksoni
and Squalus megalops
Tail I
Tail II
Tail III
Tail IV
Tail V
Tail VI
Mean 6
Mean r
Heterodontus portus-
er0r
er
er
er
0r
= i:
= i
jacksoni
H°57'1-02
- 2 ° 3 6 'o-S4
I9°S6'°535°24'2-73
I3°37'0 9 2
23°37'0 7 4
J O I6 '
[•08
Squalusmegalops
38V0-56
2 3 V0 8 6
2O°3O'0 7 7
3 2 V°-33
24°42'0-47
2O°23'0-67
26°27'0 6 1
Table 2. The values of the angles of inclination (9) to the horizontal and theproportional distances (r) of tail length intersected along the horizontal by theline of thrust after removal of the ventral lobes of the same tails detailed in
Table 1
Heterodontus portus- Squalusjacksoni megalops
Tail I 6 38°i4' 5 0 V 'r 076 0-49
Tail II 6 4i°56' 40°i2'r 077 0-78
Tail III 0 48°i9' 47°37'r 0-54 0-82
Tail IV 0 4 i°57' 5 4°29 'r o-86 0-55
TailV 6 47°i2' 42°27'r 0-72 o-6o
Tail VI 0 34°59' 47°28'r 077 072
Mean 0 = 42°6' 47°9'Mean r = 0-74 O-66
(b) Tails with ventral lobe removed
The values of 6 and r for the same tails detailed above, after removal of their ventrallobes, are given in Table 2.
Comparing this set of results with the previous set indicates that in each case
The thrust of heterocercal tails 101
removal of the ventral lobe of the tail had the effect of increasing the angle of inclina-tion of the thrust developed by the tail in both species of fish. In both species the newelevations of the thrust line reached values of some 400, enabling one to conclude that,minus their ventral lobes, the tails of Heterodontus and Squalus have closely com-parable actions (Fig. 3 C and D).
It is obviously the case that in both species the action of the ventral lobe of the tail
Fig. 3. The location and direction of the thrust (T) of the whole tail of A. Heterodontusportusjacksoni and B. Squalus megalops. C. and D. illustrate the change in direction andlocation of the thrust after removal of the ventral lobes from the same tails.
102 J. R. SIMONS
is such that the line of thrust of the whole tail is brought nearer to the horizontal. Oneconcludes, in effect, that the action of the ventral lobe reduces lift.
A further demonstration of increasing negative pitch (and therefore, since we nowknow that the line of thrust is angled upwards, of lift) with removal of the ventral lobeis provided in Fig. 4. This figure shows traces taken by a pen recorder harnessed tothe crossbar of a replica of Alexander's original apparatus. As the tail was rotated ittended to move the crossbar out of the horizontal plane. A pressure transducer wasconnected to the crossbar. The moment of negative pitch produced by the tail was thustransformed and 'read' by the pen recorder as a rise in pressure. Readings weretaken first of the whole tail and subsequently after the ventral lobe was cut away. Theexperiment was performed mainly for the purpose of providing a simple and effectiveclass demonstration of the 'action' of the ventral lobe of the tail.
B
Fig. 4. Traces of the moment of negative pitch developed by the tail of Heterodontus portus-jacksoni. A. Whole tail. B. Ventral lobe cut away. The greater moment is generated in B.The overall time taken in B is shorter than in A since the removal of the ventral lobe decreasesthe resistance of the tail to lateral motion. Hence, for the same driving torque the tail ismoved more rapidly. The increased moment generated is due in part to the faster motion as wellas to the increase in the angle of inclination of the thrust resulting from the removal of theventral lobe.
DISCUSSION
The results detailed above show that it is possible to speak of the lift generated bythe heterocercal tails of Heterodontus portusjacksoni and Squalus megalops in terms ofa vertical component of the thrust rather than in terms of a torque or moment of force;that there is, in reality, a force generated which aids in the levitation of the elasmo-branchs as they swim.
It has also been shown that the ventral lobe of the tail in these fish acts to reducelift. Since previous authors have explained 'lift' as being due to the presence of theventral lobe, the result of the present work is seen to be in conflict with the usuallyaccepted ideas about the action and method of working of the heterocercal tail. Theventral lobe, it should be noted, is referred to variously as ' hypochordal' (Grove &Newell, 1936; Affleck ,1950) or 'hypocaudal' (Young, 1955; Alexander, 1965) butthe present author prefers to follow a translation of the Latin terminology used byMarinelli & Strenger (1959).
According to Grove & Newell (1936) and Affleck (1950), who used models ratherthan actual tails, when the tail moves laterally through the water the ventral lobe isdeflected in such a way as to form an inclined plane of which the ' edge' fixed to thevertebral column precedes the free ventral edge in the direction of the lateral displace-ment. The inclined plane is thus oriented to produce a component of force directedvertically upwards. At the same time the dorsal lobe of the tail produces a similarinclined plane which, opposed in sense to the ventral one, produces a downwardly
The thrust of heterocercal tails 103
directed force. The resulting' lift' is the algebraic sum of these two oppositely directedforces.
It will be noticed, from the various illustrations given, that in the tail of Hetero-dontus there are two distinct areas which constitute the hypochordal region of the tail.
Fig. 5. Diagram illustrating the position and movement of the leading and trailing edges ofthe tail of Heterodontus portusjacksoni when the tail is moved laterally in water. A. The outlineof the tail as seen from the left. The leading edges of the lobes are indicated by the thicklined margins; the trailing edges, by thinner lines. The position of the vertebral column isalso indicated. B. The tail viewed from above as it moves laterally to the left. Note that theleading edge of the ventral lobe advances before the general position of the vertebral column.The posterior third of the leading edge of the dorsal lobe behaves similarly. The trailing edgeof the inferior lobe (i./.) trails behind the vertebral column. C. The configuration of verticalsections a-a and b-b as viewed from the rear when the tail moves laterally to the left. D. Theconfiguration of horizontal sections c-c and d-d viewed from above as the tail moves laterally tothe left. Note that each horizontal section is a symmetrical hydrofoil outline which becomesbent at each oscillation of the tail so that the thinner region trails the direction of oscillation.Consideration of the diagrams leads to the appreciation that in lateral movement the tailassumes a configuration approximating part of the wall of a hollow cone in which the narrowend is directed anteriorly and the expanded end posteriorly. The concavity of the wall of thecone is directed towards the direction of lateral motion of the tail. i./. = inferior lobe ofepichordal lobe. v.l. = ventral lobe. The displacement and thickness of the sections has beenexaggerated.
The larger more ventral region is referred to in this work as the ventral lobe. Sepa-rated from the ventral lobe by a large gap is a more posterior and smaller lobe whichmay be termed the inferior region of the dorsal lobe. In all the observations which thepresent author has made in the movements of the tail of Heterodontus the inferiorregion alone is the only part of the tail which behaves in the way expected from the
104 J- R- SIMONS
account given by Grove & Newell. The larger, ventral lobe behaves in quite a contraryfashion.
When the tail of Heterodontus moves laterally, any given point on the ventral edgeof the ventral lobe can be seen to move either with or in front of the correspondingpoint vertically above on the vertebral column. No point on the lower edge ever lagsbehind the vertebral column (see Fig. 5). That is to say, the ' inclined plane' formedby the lobe is oriented in the reverse sense to that expected from Grove & NewelPsaccount.
An explanation of the deflexion of the ventral lobe of Heterodontus may be givenin terms of its general structure. A horizontal section of the lobe taken at any leveldiscloses a symmetrical hydrofoil section (Fig. 5D). The greater bulk of the internalstructure necessarily lies in the anterior third to a quarter of the section whileposteriorly, the section thins to form a fine edge. The anterior part of the section ismuch less flexible than the posterior part.
When a flexible rectangular plate having such a section is moved laterally against aresisting medium it tends to be deflected, because of the difference in flexibilities, sothat the thicker region turns towards the direction of lateral motion and the thinneraway. The regions thus form the leading and trailing edges respectively.
In the tail of the elasmobranch the hydrofoil sectioned lobe is not a simple rec-tangular plate. Furthermore, the leading edge tapers as it nears its distal extremity.Consequently, the angle of deflexion in lateral motion of the tail is not uniform at allpoints on the surface of the lobe. It remains true, however, that stiffened edges leadand thinner, more flexible edges trail.
In the tail of Heterodontus the leading edge of the ventral lobe is not the vertebralcolumn but the ventral margin of the lobe. In a similar way, the leading edge of thedorsal lobe is its dorsal margin. All other margins—the posterior margin of the dorsallobe, the margin of the inferior region of the dorsal lobe and the posterior margin ofthe ventral lobe—are trailing edges (Fig. 5 A).
In the tail of Squalus the vertebral column is not extended to the tip of the dorsallobe as it is in Heterodontus. Consequently, the lobe is not so obviously marked offinto epichordal and inferior regions, but gives the appearance of a more or less unitfin. Again, the ventral lobe is not separated by a large gap from the dorsal area, butby a notch. Nevertheless, both ventral and dorsal lobes behave in the same way asthe corresponding lobes in Heterodontus. They both possess symmetrical hydrofoilsections and thus the leading edge of the dorsal lobe is the dorsal margin and that ofthe ventral lobe, the ventral margin.
The contradiction between the results of the present work and the contentions ofprevious authors lies in the different arrangement of leading and trailing edges foundin actual tails, compared with those found in the various models that Grove & Newelland Affleck used in their studies.
In all models previously used the central shaft simulating the vertebral column hasgenerally been some form of metal rod or strip whereas the vanes simulating the lobesof the tail have been flat, uniformly thin plates of some material (cardboard, celluloidetc.) which compared with the central shaft have been highly flexible. In these circum-stances the leading edge has been the metal shaft and the free margins of the vanes thetrailing edges. The result, as the authors have noted, is that the dorsal and ventral
The thrust of heterocercal tails 105
vanes respectively generated down and upthrusts—the reverse of the results foundin this present work. The positioning of the leading and trailing edges is a matter ofsome importance, and it is of interest to point out that Affleck (1950) did, in fact,observe that stiffening the outer edge of a vane altered or nullified its contribution tothe overall action of the model. It was not a point he pursued, except to note thatsome elasmobranchs possess tails in which both lobes were considerably strengthenedon their outer edges and therefore perhaps act as homocercal tails.
Quite apart from the degree of extension of the vertebral column in the dorsal lobe,the tails of Heterodontus and Squalus differ significantly in two other respects. Theventral lobe of Heterodontus is larger relative to the whole tail area than that ofSqualus. In the latter animal the larger dorsal lobe also extends posteriorly furtherthan in Heterodontus. These differences in relative size and layout of the lobes explainwhy the tail of Squalus produces a thrust which is more inclined to the horizontal thanthat produced by Heterodontus.
The significance of the different angles of inclination is best understood in terms ofthe general habits and food-gathering behaviour of the two fish. Heterodontus, beinga slow-swimming, placid creature, feeding on molluscs and echinoderms, may betermed a quiet browsing animal. The present author has never yet seen a specimenthat developed any turn of speed, even when repeatedly prodded with a sharp spear.The creature habitually swims with the body horizontal but can climb slowly at ashallow angle. The impression given in this activity is one of labour.
In contrast, Squalus is fast-swimming, highly manoeuvrable and capable of climbingrapidly with the body sharply inclined to the horizontal. A. A. Racek (personalcommunication) has, while engaged in underwater investigations, watched theseelasmobranchs preying on schools of mullet. The dogfish follows the school fromslightly below. In attacking, it accelerates and dives further to a point which brings itsome four or five fathoms below the centre of the school. Pulling out of the dive itascends rapidly and, metaphorically, bursts into the school, striking about with quickchanges of direction as the prey scatters.
The high degree of manoeuvrability exhibited by Squalus is due in part to thepronounced elevation of the line of thrust of the tail. It passes dorsal to the centre ofgravity and at some distance (estimated at about one-third of the body length) away.The moment of negative pitch generated when the dogfish is exerting full power (aswhen it attacks) must be considerable. This torque together with the opposing largetorque generated by the pectoral fins constitutes a system by which rapid changes ofthe fore and aft inclination of the body are made possible.
The greater elevation of the line of thrust is also advantageous in producing itsupwardly directed component of force which, together with the vertical force com-ponent, produced by the pectoral fins, brings the dogfish up rapidly in the ascentphase of its attack.
If one might draw a parallel with aircraft, Heterodontus may be compared with somefreighter aeroplane, large winged and capable of attaining lift and stability at lowspeeds but slow climbing and sluggishly manoeuvrable; whereas Squalus resemblesmore some fighter aeroplane with small lift and control surfaces operating at speedto give high manoeuvrability with the power to climb swiftly.
The tail of Heterodontus, in conformity with the creature's general primitiveness, is
106 J. R. SIMONS
much more strongly heterocercal than that of Squalus. Since it is widely, if somewhatvaguely, held that heterocercal tails produce upthrust it is at first sight a contradictoryresult to find that the more primitive tail studied in this work produces, by virtue ofthe lower angle of elevation of the line of thrust, less upthrust than the other.
Affleck (1950) pointed out that an anatomically based classification of tails (asheterocercal, homocercal etc.) does not necessarily coincide with a classification basedon the action of the tail. He found, for instance, that while some heterocercal tails hadan epibatic action (i.e. they produced a negative pitch), others were isobatic (i.e.produced neither positive nor negative pitch) and therefore acted as homocercal tailsare said to act. Now, it has been pointed out in the introduction of this work that theeffective pitch produced by a tail must depend on where the line of thrust passes withrespect to the creature's centre of gravity. If the thrust line passes above, the action ofthe tail must be epibatic; if below, hypobatic, and if through the centre of gravity,isobatic. In respect of the average results obtained in the present work, the tails ofboth Heterodontus and Squalus are epibatic in action, but with the more pronouncedheterocercal one (Heterodontus) less so. With respect to the individual results thereare two instances which would seem to suggest the possibility that the action of thetail of Heterodontus may at times be isobatic or even hypobatic since in both cases theline of thrust is angled very close to the horizontal (see Results—whole-tail specimensII and IV). The datum of the horizontal, as has been explained, has been takenthroughout this study as the line of the vertebral column of the trunk region im-mediately preceding the tail. In Heterodontus this datum line, if extended forwards,passes either through or, more likely, just below the centre of gravity of the fish(J. R. Simons, in preparation). Hence, in the case where the thrust line is angled at— 2-5°, the action of the tail must be regarded as hypobatic, and in the case where theangle is 5-5°, possibly isobatic or weakly epibatic.
As in other elasmobranchs, the tails of the animals studied here are equipped withradial muscles in the hypochordal regions. From their position and direction of con-traction it is probable, as Alexander (1965) remarks, that they act to alter the contribu-tion the ventral lobe makes to the action of the whole tail. The present author isstrongly convinced that by virtue of the radial muscles, the ventral lobe becomes adevice for altering the 'horizontal trim' of the fish, by causing a change principally inthe angle of elevation of the thrust and, to a lesser extent, in the magnitude of thethrust developed. Such a trim mechanism would be advantageous because in thesefish a large meal or, in the case of a gravid female Squalus, large embryos, alters thelocation of both centres of gravity and centres of buoyancy (J. R. Simons, in prepara-tion) and hence horizontal equilibrium tends to be upset. As in aircraft where ahorizontal trim device is used to restore the proper flight attitude as the load is varied,so in elasmobranch fish an analogous mechanism would be advantageous in mini-mising, particularly at cruising speeds, energy losses due to drag and inefficientangling of thrust forces which would otherwise occur.
SUMMARY
1. A method of determining the location and direction of the thrust developed bythe tail of a fish is described.
The thrust of heterocercal tails 107
2. The thrust produced by the heterocercal tails of Heterodontus portusjacksoni andSqualus megalops was found to be upwardly inclined to the horizontal at approximately120 and 260 respectively.
3. The angle of elevation of the line of thrust developed by the tails correlates withthe general habits of the two elasmobranchs; particularly in regard to manoeuvrabilityin the vertical plane.
4. The direction of the line of thrust depends upon the positions of the leading andtrailing edges of the lobes of the tail as well as the relative sizes, and general arrange-ment, of the lobes.
5. The ventral lobe of the tail acts primarily to reduce the angle of the elevation ofthe thrust. Minus the ventral lobe, both types of tail were found to be strongly epibaticwith lines of thrust considerably more inclined than those of the whole tail.
6. The ventral lobe of the tail, by virtue of the radial muscles, also probably func-tions as a variable trim device whereby the elasmobranch can correct its 'fore and afttrim' by adjusting the direction of the line of thrust appropriately.
I should like to express thanks to my colleagues Drs V. B. Morris and A. A. Racek,who have taken considerable interest, both critical and constructive, in the progressof this investigation.
REFERENCES
AFFLECK, R. J. (1950). Some points in the function, development and evolution of the tail in fishes.Proc. zool. Soc. Lond. iao, 349-68.
ALEXANDER, R. M C N . (1965). The lift produced by the heterocercal tails in Selachii. J. exp. Biol. 43,131-8.
GROVE, A. J. & NEWELL, G. E. (1936). A mechanical investigation into the effectual action of the caudalfin of some aquatic chordates. Arm. Mag. nat. Hist. 17, 280-90.
MARINELLI, W. & STRENGER, A. L. (1959). Vergleichende Anatomie und Morphologie der Wirbeltiere.Band 1. Wien: Franz Deuticke.
YOUNG, J. Z. (1955). The Life of Vertebrates. Oxford: Clarendon Press.

























![vixra.org · 2015. 9. 2. · G8,2 Geometric Algebra, DCGA byRobert Benjamin Easter 1Introduction This paper1introduces2an application of theG 8;2geometric algebra[3][4], tenta-tively](https://static.documents.pub/doc/80x56/60ac8e80c53dd316b87c5800/vixraorg-2015-9-2-g82-geometric-algebra-dcga-byrobert-benjamin-easter-1introduction.jpg)



