Lateral phage transfer in obligate intracellular bacteria (Wolbachia): Verification from natural populations Letter Meghan E. Chafee 1 , Daniel J. Funk 1 , Richard G. Harrison 2 , and Seth R. Bordenstein 1, † 1 Department of Biological Sciences, Vanderbilt University, Nashville, TN; and 2 Department of Ecology and Evolutionary Biology, Cornell University, Ithaca, NY † Corresponding author: Seth R. Bordenstein, [email protected], (ph) 615.322.9087, (fax) 615.343.6707 Keywords: Lateral gene transfer, Wolbachia, bacteriophage, obligate intracellular bacteria, recombination, coinfection Running head: Lateral phage transfer Title length: 69 characters (with spaces) Abstract length: 192 words Total characters, including all legends and methods, but not Abstract: 11,132 Number of references 30
Transcript
Lateral phage transfer in obligate intracellular bacteria (Wolbachia):
Verification from natural populations
Letter
Meghan E. Chafee1, Daniel J. Funk1, Richard G. Harrison 2, and Seth R. Bordenstein1, †
1 Department of Biological Sciences, Vanderbilt University, Nashville, TN; and 2 Department of
Ecology and Evolutionary Biology, Cornell University, Ithaca, NY
† Corresponding author: Seth R. Bordenstein, [email protected], (ph) 615.322.9087,
and ORF7 haplotype. Pairs of haplotypes in shaded boxes illustrate putative lateral transfer
events in field specimens collected in this study. Bold taxa represent previously published
examples of lateral transfer through laboratory segregation experiments. The three group labels
(I-III) were assigned according to the nomenclature in Bordenstein and Wernegreen (2004).
Acknowledgements
This work was supported by grants NSF IOS-0852344 and NIH R01 GM085163-01 to S.R.B.
This paper’s contents are solely the responsibility of the authors and do not necessarily represent
the official view of the NSF or NIH.
References. Baldo, L., J. C. Dunning Hotopp, K. A. Jolley, S. R. Bordenstein, S. A. Biber, R. R. Choudhury,
C. Hayashi, M. C. Maiden, H. Tettelin, and J. H. Werren. 2006. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl Environ Microbiol 72:7098-7110.
Bordenstein, S. R., M. L. Marshall, A. J. Fry, U. Kim, and J. J. Wernegreen. 2006. The tripartite associations between bacteriophage, Wolbachia, and arthropods. PLoS Pathog 2:e43.
Bordenstein, S. R., and W. S. Reznikoff. 2005. Mobile DNA in obligate intracellular bacteria. Nat Rev Microbiol 3:688-699.
Bordenstein, S. R., and J. J. Wernegreen. 2004. Bacteriophage flux in endosymbionts (Wolbachia): infection frequency, lateral transfer, and recombination rates. Mol Biol Evol 21:1981-1991.
Brower, A. V. 1994. Rapid morphological radiation and convergence among races of the butterfly Heliconius erato inferred from patterns of mitochondrial DNA evolution. Proc Natl Acad Sci U S A 91:6491-6495.
Campbell, B. C., J. D. Steffen-Campbell, and J. H. Werren. 1993. Phylogeny of the Nasonia species complex (Hymenoptera: Pteromalidae) inferred from an internal transcribed spacer (ITS2) and 28S rDNA sequences. Insect Mol Biol 2:225-237.
Cordaux, R. 2008. ISWpi1 from Wolbachia pipientis defines a novel group of insertion sequences within the IS5 family. Gene 409:20-27.
Drummond, A., B. Ashton, M. Cheung, J. Heled, M. Kearse, R. Moir, S. Stones-Havas, T. Thierer, and A. Wilson. 2009. Geneious v.4.7, Available from http://www.geneious.com/.
Felsenstein, J. 1993. PHYLIP (Phylogeny Inference Package) version 3.567. Distributed by the author. Department of Genetics, University of Washington, Seattle.
Funk, D. J. 1999. Molecular systematics of cytochrome oxidase I and 16S from Neochlamisus leaf beetles and the importance of sampling. Mol Biol Evol 16:67-82.
Gavotte, L., H. Henri, R. Stouthamer, D. Charif, S. Charlat, M. Bouletreau, and F. Vavre. 2007. A survey of the bacteriophage WO in the endosymbiotic bacteria Wolbachia. Mol Biol Evol 24:427-435.
Gavotte, L., F. Vavre, H. Henri, M. Ravallec, R. Stouthamer, and M. Bouletreau. 2004. Diversity, distribution and specificity of WO phage infection in Wolbachia of four insect species. Insect Mol Biol 13:147-153.
Huelsenbeck, J. P., and F. Ronquist. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17:754-755.
Ishmael, N., J. C. Hotopp, P. Ioannidis, S. Biber, J. Sakamoto, S. Siozios, V. Nene, J. Werren, K. Bourtzis, S. R. Bordenstein, and H. Tettelin. 2009. Extensive genomic diversity of closely related Wolbachia strains. Microbiology 155:2211-2222.
Iturbe-Ormaetxe, I., G. R. Burke, M. Riegler, and S. L. O'Neill. 2005. Distribution, expression, and motif variability of ankyrin domain genes in Wolbachia pipientis. J Bacteriol 187:5136-5145.
Kimura, M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16:111-120.
Klasson, L., J. Westberg, P. Sapountzis, K. Naslund, Y. Lutnaes, A. C. Darby, Z. Veneti, L. Chen, H. R. Braig, R. Garrett, K. Bourtzis, and S. G. Andersson. 2009. The mosaic genome structure of the Wolbachia wRi strain infecting Drosophila simulans. Proc Natl Acad Sci U S A 106:5725-5730.
Maddison, D., and W. Maddison. 2005. Sinauer Associates, Sunderland, MA. Mandel, M. J., C. L. Ross, and R. G. Harrison. 2001. Do Wolbachia infections play a role in
unidirectional incompatibilities in a field cricket hybrid zone? Mol Ecol 10:703-709. Masui, S., S. Kamoda, T. Sasaki, and H. Ishikawa. 2000. Distribution and evolution of
bacteriophage WO in Wolbachia, the endosymbiont causing sexual alterations in arthropods. J Mol Evol 51:491-497.
Moran, N. A., and G. R. Plague. 2004. Genomic changes following host restriction in bacteria. Curr Opin Genet Dev 14:627-633.
Ochman, H., S. Elwyn, and N. A. Moran. 1999. Calibrating bacterial evolution. Proc Natl Acad Sci USA 96:12638-12643.
Oliver, K. M., P. H. Degnan, G. R. Burke, and N. A. Moran. 2009a. Facultative symbionts of aphids and the horizontal transfer of ecologically important traits. Annu Rev Entomol.
Oliver, K. M., P. H. Degnan, M. S. Hunter, and N. A. Moran. 2009b. Bacteriophages encode factors required for protection in a symbiotic mutualism. Science 325:992-994.
Rozas, J., J. Sanchez-DelBarrio, X. Messeguer, and R. Rozas. 2003. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 19:2496-2497.
Sanogo, Y. O., and S. L. Dobson. 2006. WO bacteriophage transcription in Wolbachia-infected Culex pipiens. Insect Biochem Mol Biol 36:80-85.
Sinkins, S. P., T. Walker, A. R. Lynd, A. R. Steven, B. L. Makepeace, H. C. Godfray, and J. Parkhill. 2005. Wolbachia variability and host effects on crossing type in Culex mosquitoes. Nature 436:257-260.
Tamura, K., S. Subramanian, and S. Kumar. 2004. Temporal patterns of fruit fly (Drosophila) evolution revealed by mutation clocks. Mol Biol Evol 21:36-44.
Tanaka, K., S. Furukawa, N. Nikoh, T. Sasaki, and T. Fukatsu. 2009. Complete WO phage sequences revealed their dynamic evolutionary trajectories and putative functional elements required for integration into Wolbachia genome. Appl Environ Microbiol.
Wu, M., L. V. Sun, J. Vamathevan, M. Riegler, R. Deboy, J. C. Brownlie, E. A. McGraw, W. Martin, C. Esser, N. Ahmadinejad, C. Wiegand, R. Madupu, M. J. Beanan, L. M. Brinkac, S. C. Daugherty, A. S. Durkin, J. F. Kolonay, W. C. Nelson, Y. Mohamoud, P. Lee, K. Berry, M. B. Young, T. Utterback, J. Weidman, W. C. Nierman, I. T. Paulsen, K. E. Nelson, H. Tettelin, S. L. O'Neill, and J. A. Eisen. 2004. Phylogenomics of the reproductive parasite Wolbachia pipientis wMel: a streamlined genome overrun by mobile genetic elements. PLoS Biol 2:E69.
Table 1 Average Pairwise Nucleotide Diversity Per Site (π) in Five Genes Taxa
Gene
Length of
Alignment (bp) φ
N
π
Wolbachia pipientis
ftsZ
435
38
0.058
gatB
369 38 0.066
hcpA
444 38 0.073
coxA
402 38 0.080
Bacteriophage WO-B ORF7
240 59 0.146
φ Length of the alignment is based on removal of indels
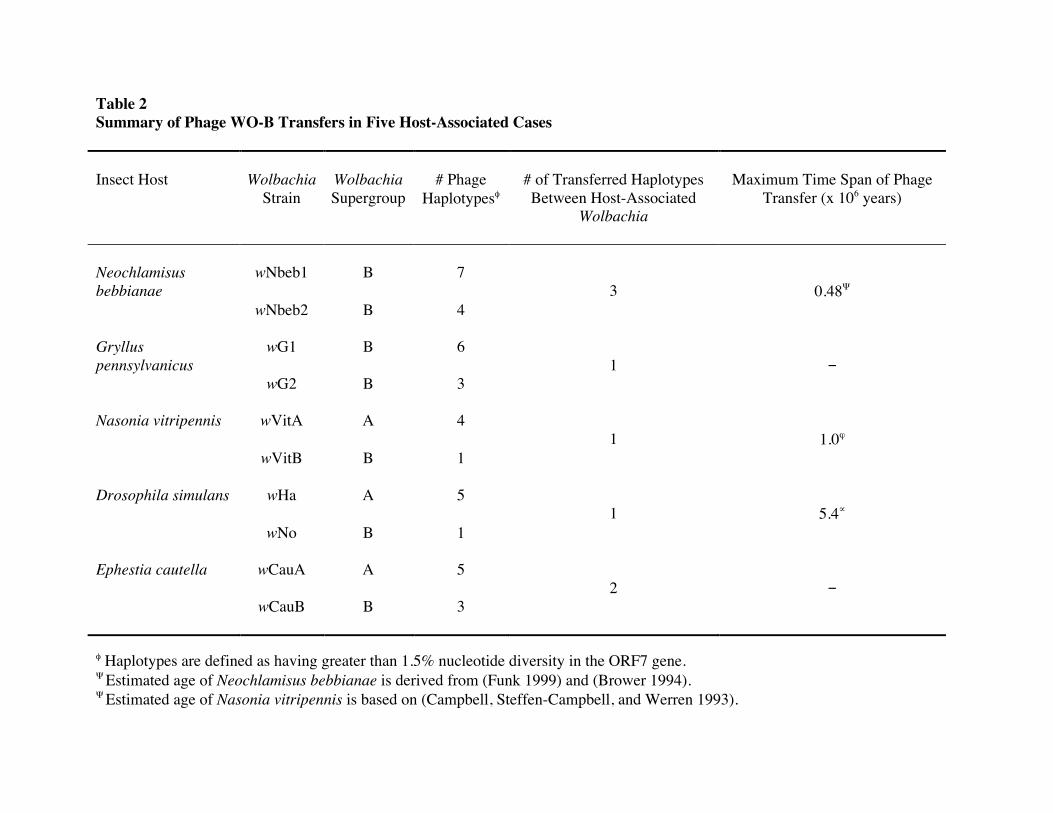
Table 2 Summary of Phage WO-B Transfers in Five Host-Associated Cases Insect Host
Wolbachia
Strain
Wolbachia Supergroup
# Phage
Haplotypesφ
# of Transferred Haplotypes Between Host-Associated
Wolbachia
Maximum Time Span of Phage
Transfer (x 106 years)
Neochlamisus bebbianae
wNbeb1
B
7
3
0.48Ψ wNbeb2
B 4
Gryllus pennsylvanicus
wG1
B
6
1
−
wG2
B 3
Nasonia vitripennis wVitA
A 4 1
1.0ϕ
wVitB
B 1
Drosophila simulans wHa
A 5 1
5.4∝
wNo
B 1
Ephestia cautella wCauA
A
5 2
−
wCauB
B 3
φ Haplotypes are defined as having greater than 1.5% nucleotide diversity in the ORF7 gene. Ψ Estimated age of Neochlamisus bebbianae is derived from (Funk 1999) and (Brower 1994). Ψ Estimated age of Nasonia vitripennis is based on (Campbell, Steffen-Campbell, and Werren 1993).
∝ Estimated age of Drosophila simulans is based on (Tamura, Subramanian, and Kumar 2004).
























![Wolbachia Seminar Master [Compatibility Mode]](https://static.documents.pub/doc/80x56/54679b73b4af9f623f8b588c/wolbachia-seminar-master-compatibility-mode.jpg)






