lecture 10. Genetic Interactions lecture 10. Genetic Interactions synthetic lethality synthetic lethality high copy suppression suppression and enhancement high copy suppression suppression and enhancement mechanisms of suppression mechanisms of suppression
synthetic lethalitysynthetic lethalityhigh copy suppression suppression and enhancementhigh copy suppression suppression and enhancementmechanisms of suppressionmechanisms of suppression
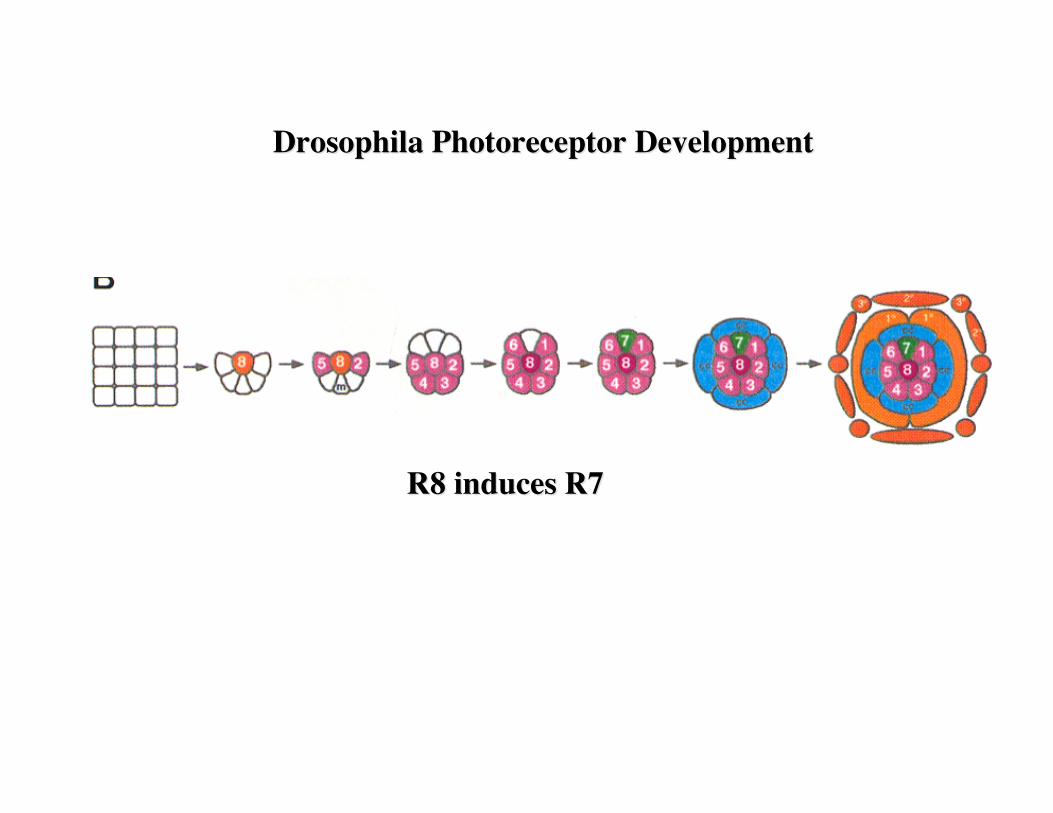
Drosophila Photoreceptor DevelopmentDrosophila Photoreceptor Development
R8 induces R7R8 induces R7
Multiple Multiple OmmatidaOmmatidain each eye:in each eye:a population assaya population assay
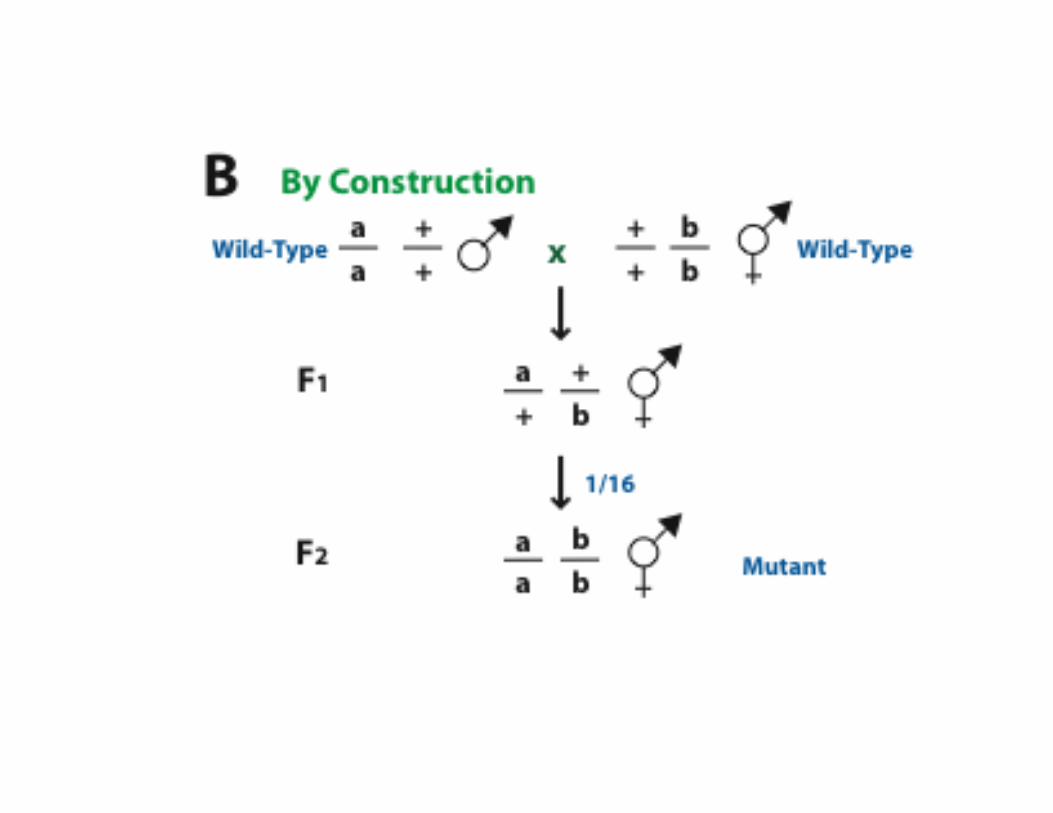
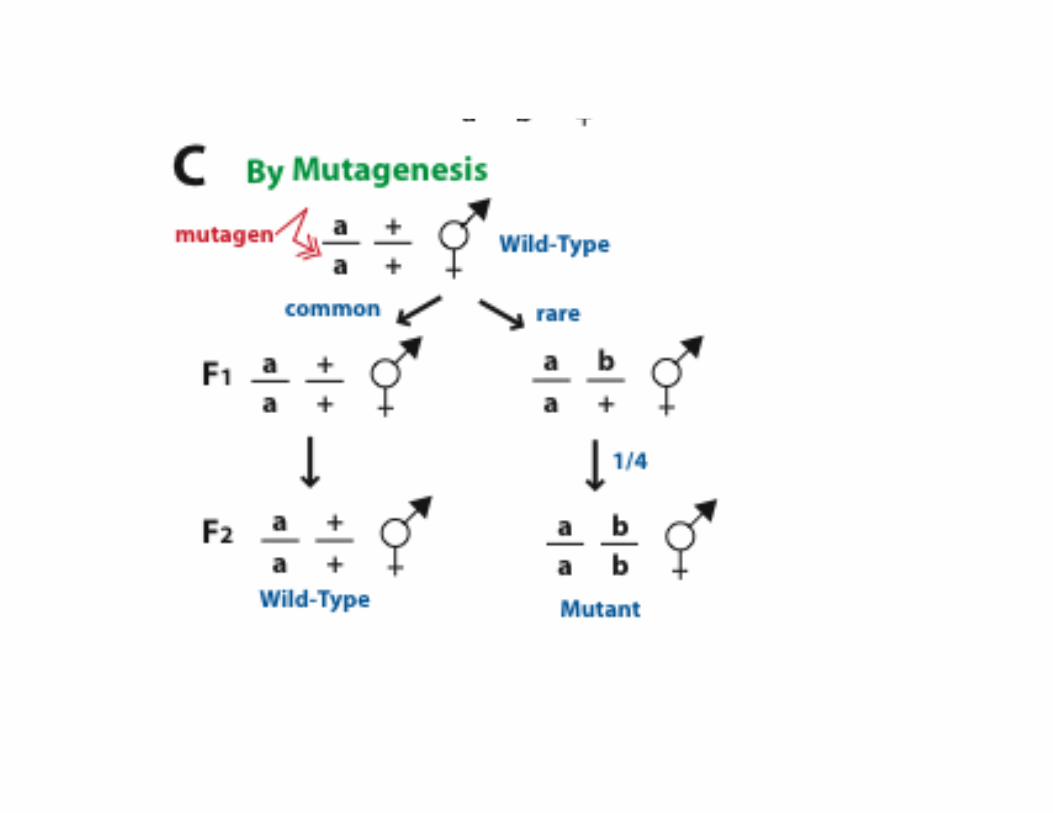
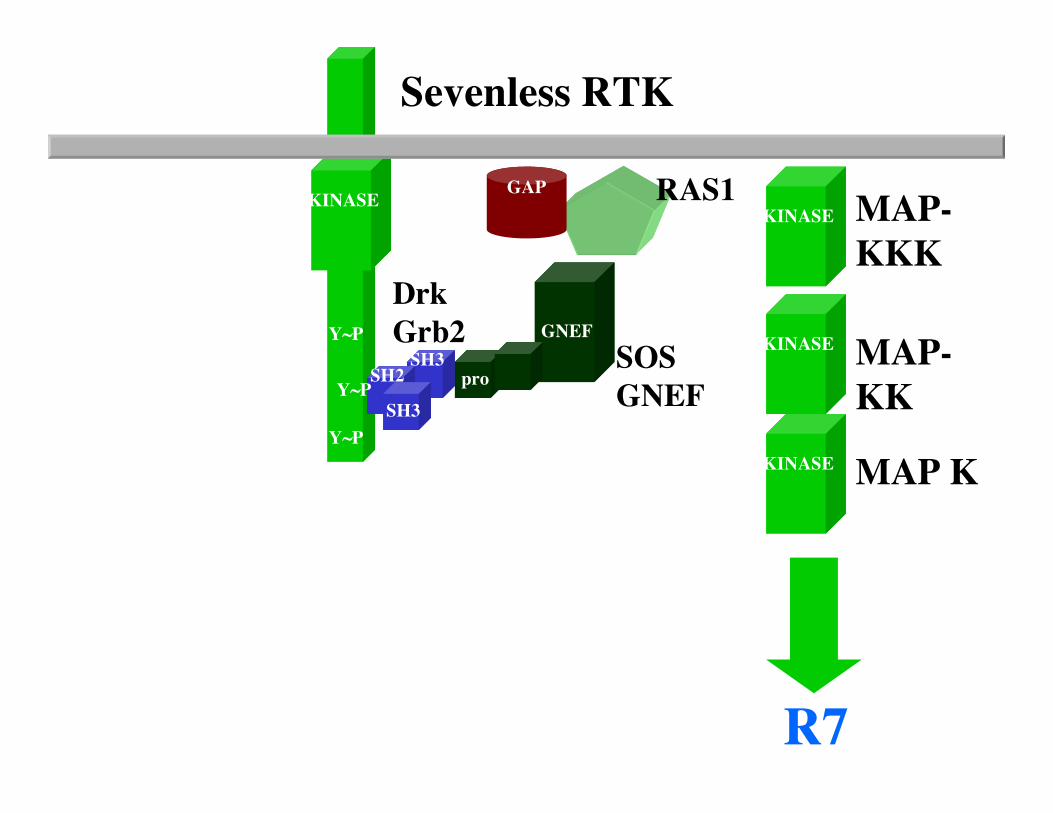
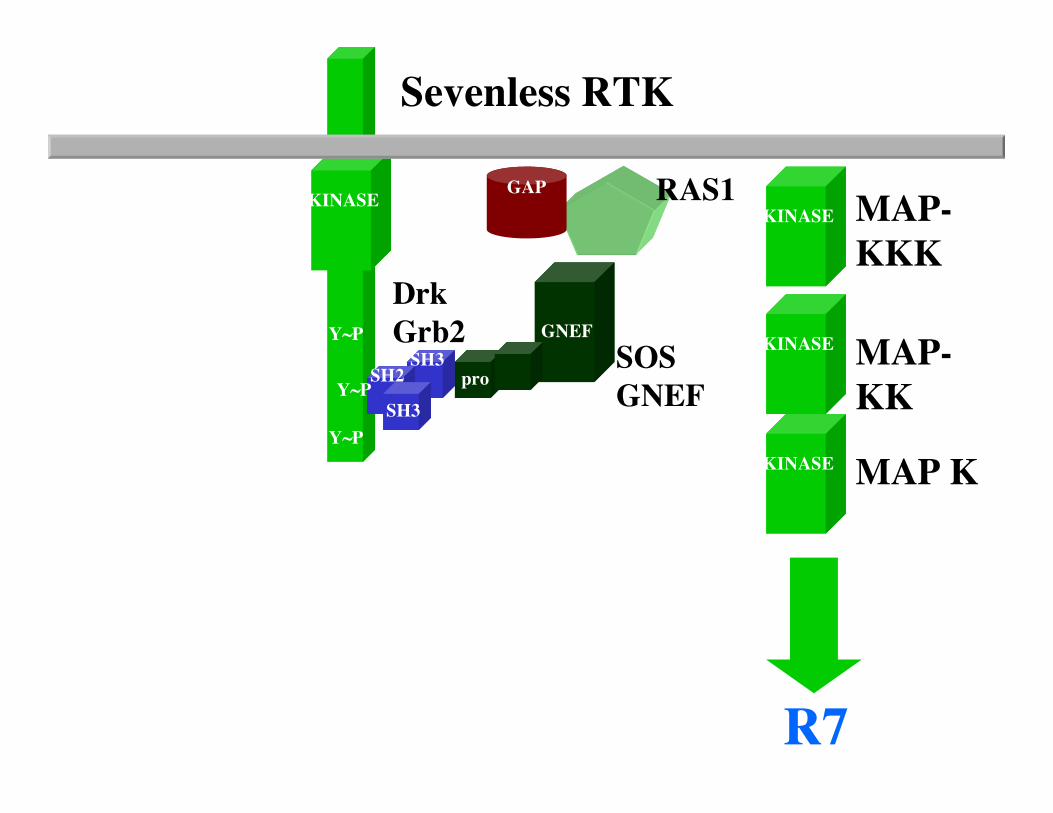
The fly eye consists of approximately 800 20-cell repeating units known as ommatidia. Each ommatidium consists of eight photoreceptor neurons (R1-R8), four lens secreting cone cells and eight additional accessory cells. The ommatidia arise from an undifferentiated epithelium by a series of cell interactions. We will only consider an interaction between theR8 and presumptive R7 cells that determines the fate of R7. The R7 photoreceptor detects light in the UV range. Screens for mutants with ommatidia that lack R7 cells identified three genes: sevenless (sev), bride of sevenless (Boss) and seven-in-abstentia (sina). Adult flies homozygous for mutations in any of these genes have ommatidia that lack an R7 cell and contain an additional cone cell. In the absence of R7 differentiation, the presumptive R7 cell becomes a cone cell. sev and sina are a receptor tyrosine kinase and a nuclear protein, respectively, and both genes act in R7 to specifyR7's fate. boss appears to encode the ligand for the Sev receptor tyrosine kinase, and in contrast to sev and sina, acts in R8 cell to specify R7's fate.
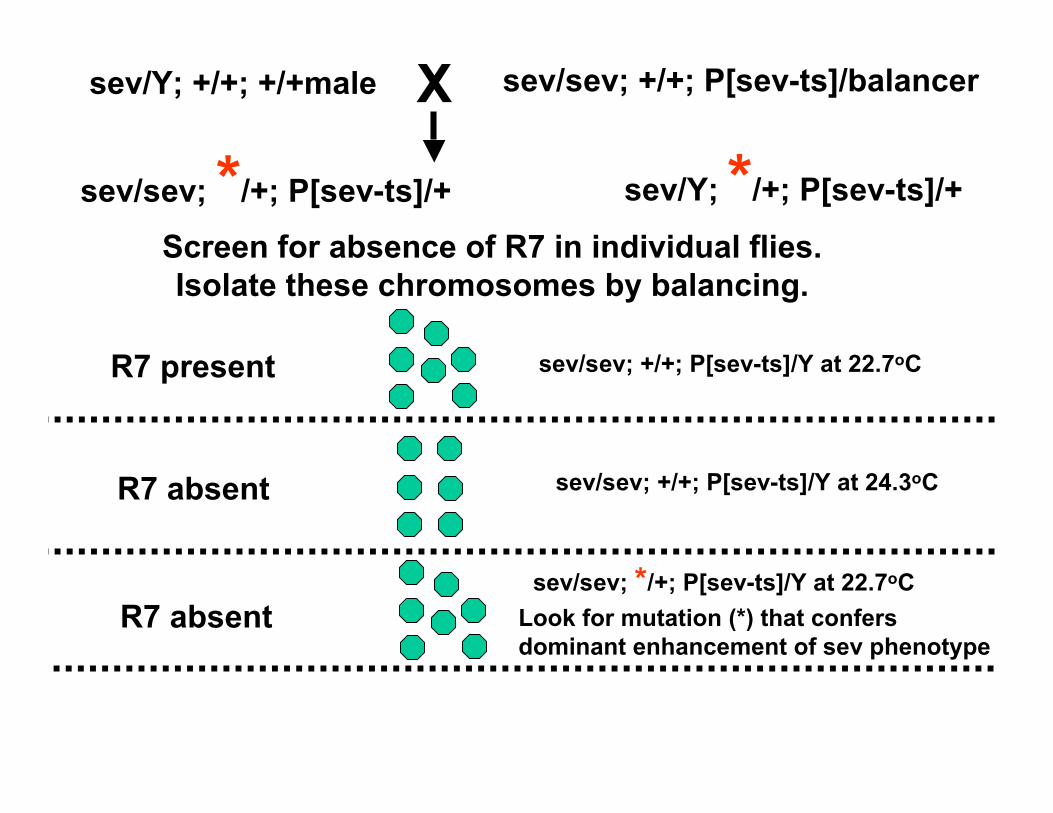
Now consider the problem that many genes functioning downstream of receptor tyrosine kinse receptor activation arelikely to be required for other tyrosine kinase signaling pathways that are required for the viability of the organism. How can one use the fly eye to identify such mutations in such genes.
Make a partially active mutant version of sev and introduce it into a sev mutant background. These flies have atemperature-sensitive phenotype. A fly carrying one copy of this transgene is wildtype at 22.7oC (R7 is present).However, at 24.3oC R7 is absent
An enhancer screen for essential genes required for R7 development
Screen for absence of R7 in individual flies.Isolate these chromosomes by balancing.
sev/sev; +/+; P[sev-ts]/Y at 22.7oC
sev/sev; +/+; P[sev-ts]/Y at 24.3oC
sev/sev; */+; P[sev-ts]/Y at 22.7oCLook for mutation (*) that confersdominant enhancement of sev phenotype
R7 present
R7 absent
R7 absent
SH3
Sevenless RTK
DrkGrb2
SOSGNEF
RAS1
R7
KINASE
SH2
SH3
pro
GNEF
Y~P
Y~P
GAP
Y~P
KINASE
KINASE
KINASE
MAP-KKK
MAP-KK
MAP K
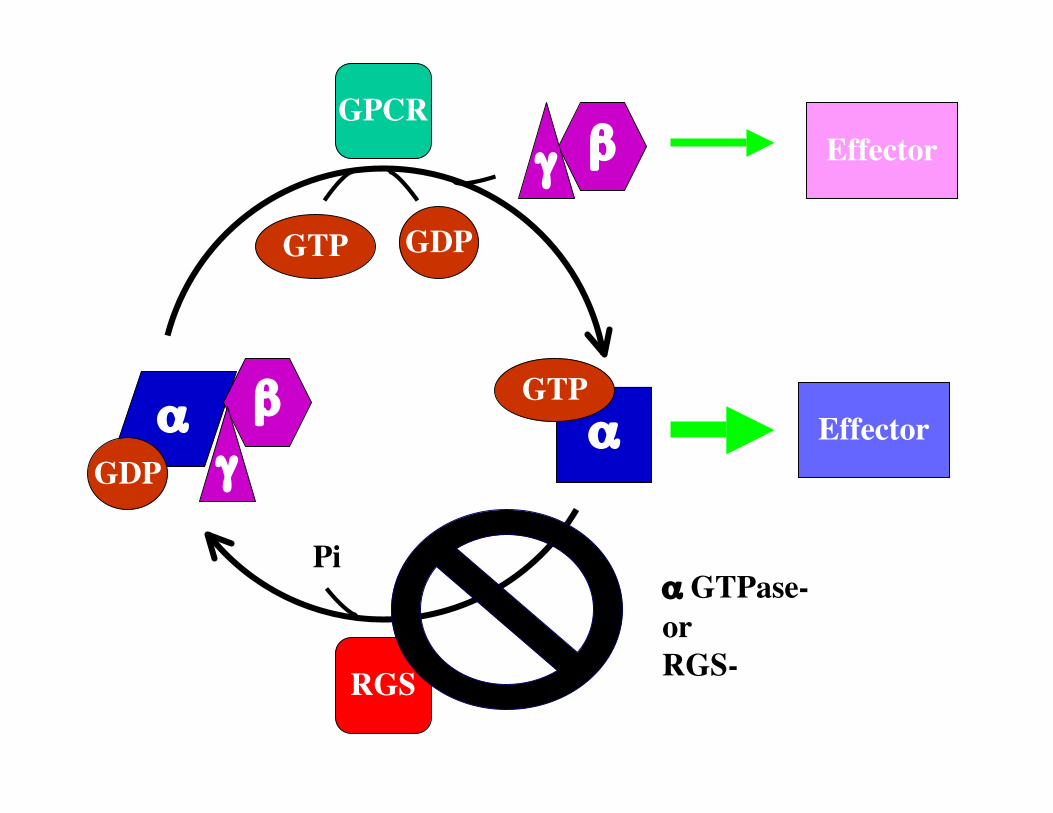
αα Effector
RGS
GPCR
GDP
GTP
GDPGTP
βγ
βγ Effector
Pi
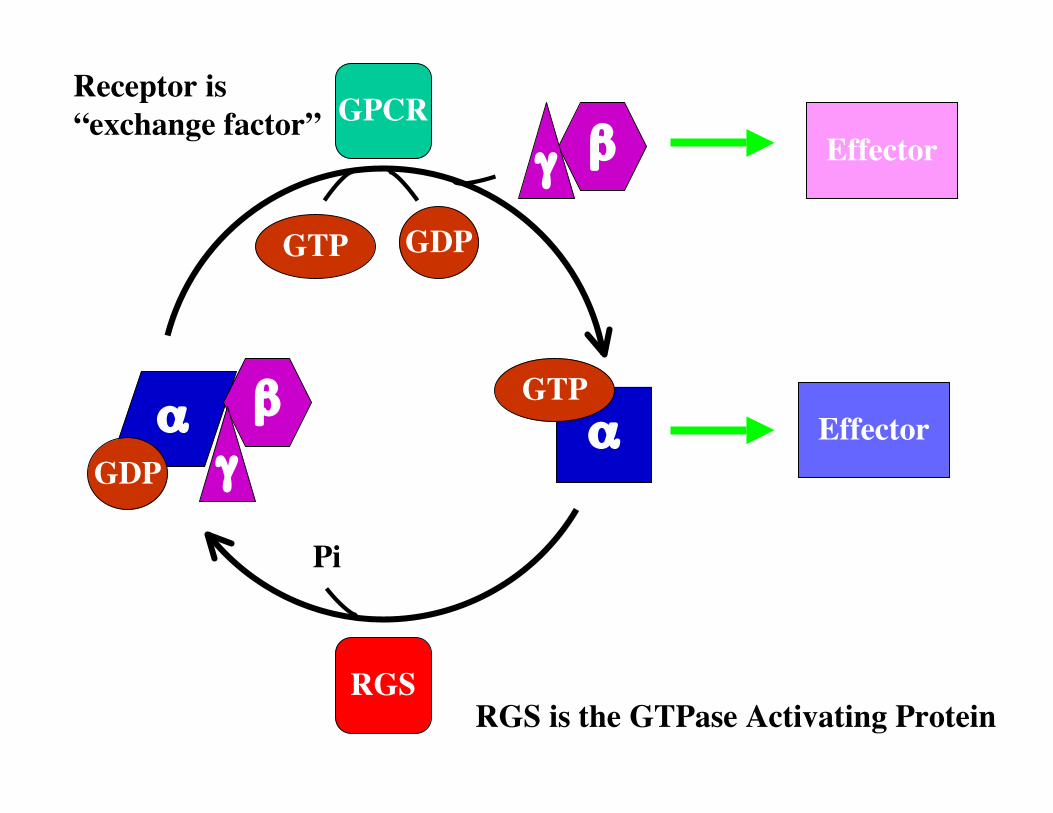
Receptor is“exchange factor”
RGS is the GTPase Activating Protein
αα Effector
RGS
GPCR
GDP
GTP
GDPGTP
βγ
βγ Effector
Piα GTPase-orRGS-
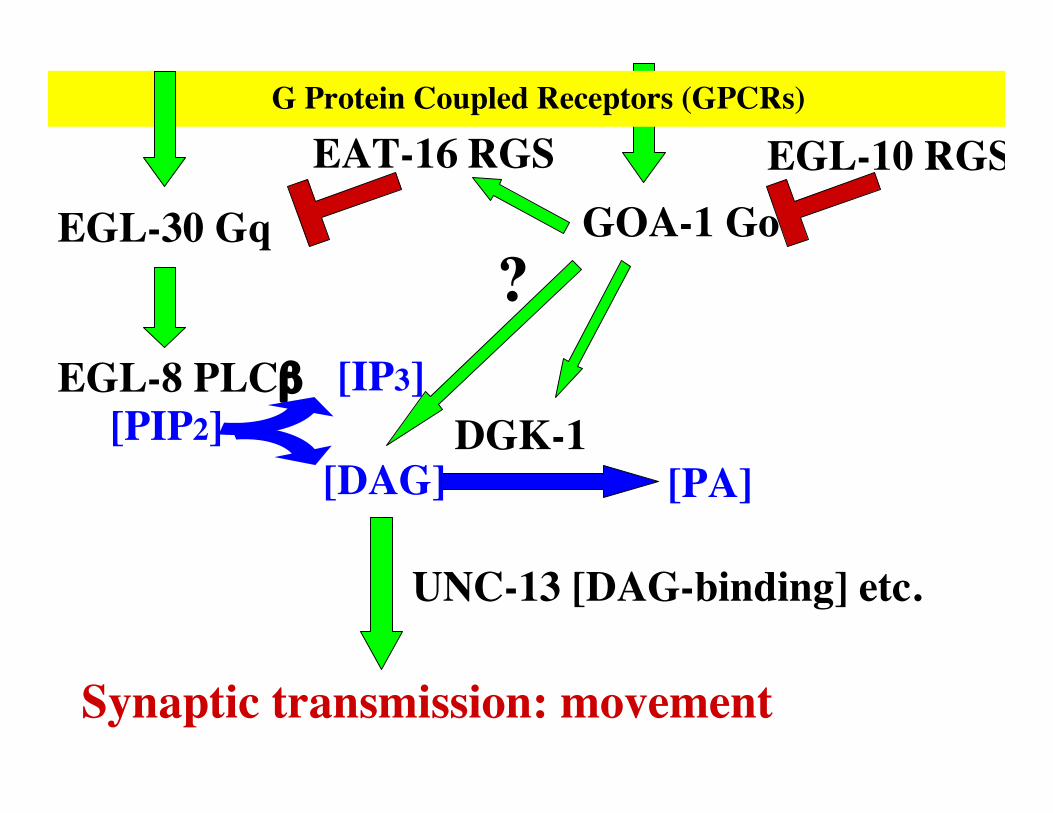
G proteins Gq and Go control movement
Genotype Phenotype
Wild type wild-typeegl-30(lf) paralyzedegl-30(gf) hyperactivegoa-1(lf) hyperactivegoa-1(gf) paralyzedegl-30(lf) goa-1(lf) paralyzed
lf, loss-of-function; gf, gain-of-function
C. C. eleganselegans
Mutations that Suppress activated Goα
syIs17 syIs17; sag-4(sy433)
Before Heat Shock
AfterHeat Shock
Jane Mendel, Yvonne Jane Mendel, Yvonne HajduHajdu-Cronin, -Cronin, Wen Wen ChenChen
Suppressors of Activated Goα (Sag)
• sag-4, 8 sag-4 encodes cyclin L homologueCLASS II wild type
CLASS I hyperactive• dgk-1/sag-1 (14 alleles) encodes diacylgycerol kinase• eat-16(sy348) (p.k.a. sag-2) encodes RGS7 homologue
CLASS III Egg-laying defective• sag-3, 5 sag-3 encodes Heat Shock Factor
CLASS IV wild type• sag-6
CLASS V Egg-laying defective• sag-7
Yvonne Hajdu-Cronin & Wen Chen
EGL-30 Gq
EGL-8 PLCβ [IP3]
[DAG]
Synaptic transmission: movement
[PIP2] DGK-1[PA]
GOA-1 GoEGL-10 RGSEAT-16 RGS
?
UNC-13 [DAG-binding] etc.
G Protein Coupled Receptors (GPCRs)
Extragenic suppression• many mechanisms--key issue is the genetic specificity of
the suppressor
‘informational’suppression
gene-nonspecificallele-specific
direct interaction?gene-specificallele-specific
epistasis (bypasssuppression)
gene-specificallele-nonspecific
suppression by compensatorychange in direct interactor?
• ‘Lock and Key’ model: binding site is restored• in general a very rare event as target size is 1(or a few) bp-
-need screens of >106 genomes• RNA-RNA interactions:
– restoration of base pairing (nonsense suppression)– splice site suppression e.g. Lesser + Guthrie 1993 Science 262:
1982• protein-DNA interactions
– lac operon: oC mutations suppressed by mutations in repressor thatbind more tightly to operator (Pfahl 1981, J. Mol. Biol. 147: 1-10)
• protein-protein interactions?
allele-specific suppression
• null mutants are not suppressed, so notbypass suppressor
• stabilization or altered processing of mutantgene product
suppression by formation of newprotein-protein interactions
Adams + Botstein 1989.suppressors of ts actinmutants– get sac mutants. sac6 is
fimbrin, actin-binding– sac6 mutations are
missense in actin bindingdomain, increase affinityfor mutant actin
– But the affinity for wildtype actin is also increased
ACT SAC
act SAC
act sac
ACT sac
gene non-specific, allele specific
• suppression at level of gene expression:‘informational’
1. Nonsense suppression2. Frameshift suppression3. Splicing machinery4. stabilization of unstable mRNA or protein5. suppression of transposon insertion alleles
nonsense suppression• conditional ‘amber’ mutations in many T4 genes
(Epstein et al)– grow on one E coli strain (CR63) but not on B– cause premature termination– suppression due to mutant tRNA that can recognize
amber codon UAG and insert amino acid (usually Trp;codon is UGG)
– amber suppressor strains are a bit sick because ofreadthrough
frameshift suppression
• extragenic suppression of frameshifts bytwo mechanisms– limitation of Trp-tRNA– other tRNAs loosely bind to codon (mismatch)
and allow frameshifting
– also mutant tRNA with 4-base anticodon now‘reads’ frameshift as a 3-base codon…
suppression by stabilization ofmessage
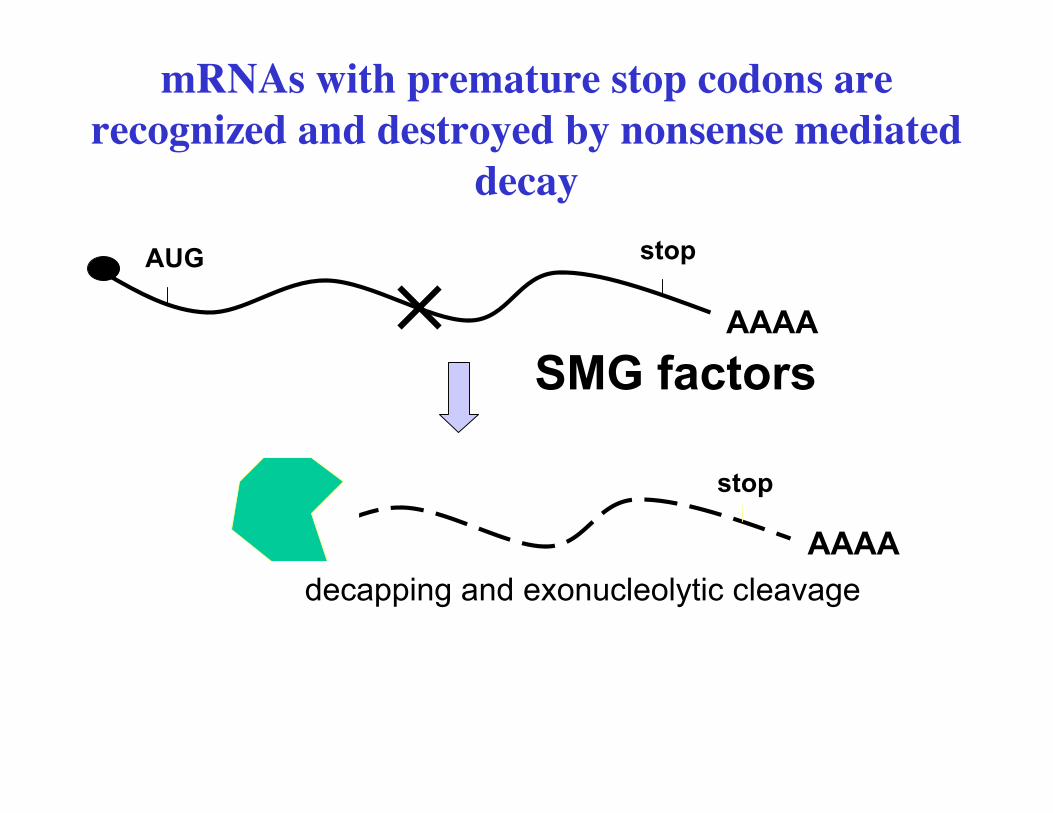
• mRNAs with ‘premature’ stop codons are recognized anddegraded– nonsense mediated decay/ ‘mRNA surveillance’– Upf pathway (yeast), SMG pathway (worms)– get rid of aberrant mRNAs before they get to ribosome
• some nonsense mutations can be suppressed if partiallyfunctional protein can be made
AAAA
stopAUG
mRNAs with premature stop codonsproduce truncated proteins.
Expression of these from many loci can be detrimental to the animal.Cells have mechanisms of removing aberrant mRNAs
mRNAs with premature stop codons arerecognized and destroyed by nonsense mediated
decay
AAAA
SMG factors
stopAUG
AAAAdecapping and exonucleolytic cleavage
stop
Screens for suppressors of nonsensemutations revealed smg genes
Hodgkin J, Papp A, Pulak R, Ambros V, Anderson P. A new kind of informational suppression in thenematode Caenorhabditis elegans. Genetics. 1989 Oct;123(2):301-13.
Mutations in the proteins required for nonsense mediated decaysuppress nonsense mutations by allowing stabilizing mRNAs withpremature stop codons.Functional proteins are made since low levels of readthrough makesome normal protein or because expression of the truncated proteincan suppress the phenotype
AAAA
stopAUG
In the absence of SMG proteins mRNAs withpremature stop codons will persist
Expression of these from many loci can be detrimental to the animal
mRNAs with premature stop codons have a low level of readthrough,these levels may be enough to rescue the mutant phenotype
Short protein fragmentis not functional orantimorphic
suppression by stabilization ofprotein
• E. coli lon protease degrades aberrantproteins
• mutations in lon suppress thermolabilemutations in many genes (RNA polymeraseetc)