When cells can coordinate these signals, they begin to live within a community either singly or communal and begin to establish a division of labour and therefore specialization of function
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
might have created multicell orgs through endocrine sys
Theories of Prebiotic Synthesis: Energy Sources
There were, in theory, several sources of energy that could have been used for the prebiotic synthesis of organic chemicals
Sunlight
Lightning
Geothermal vents
Cosmic Radiation
Volcanic activity
CSB$325$Lecture$4:$Hormone$Evolu7on$
Figure 2-1: Examples of organic compounds present in the prebiotic Earth. Such
compounds were used as substrates and building blocks for more complex molecules.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
NH4
A
B
C
D
E
Evolution of Protocells
Association of organic material on inorganic substrate
Concentration of organic material
Partitioning of Hydrophilic and Hydrophobic material
Formation of Hydrophobic membrane
Separation of protocell From substrate
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
there was an E gradient
CSB$325$Lecture$4:$Hormone$Evolu7on$
Protocells and First Signalling Networks
What is a protocell?
Partitioning of biotic and non-biotic environments The need to signal
Theoretical evolution and structure of a protocell
Partitioning of hydrophilic and hydrophobic molecules Development of lipid membrane Ion gradients across membrane and energy production Signalling capacity Membrane diffusion Pores in membrane Protein channels Early adhesion proteins
CSB$325$Lecture$4:$Hormone$Evolu7on$
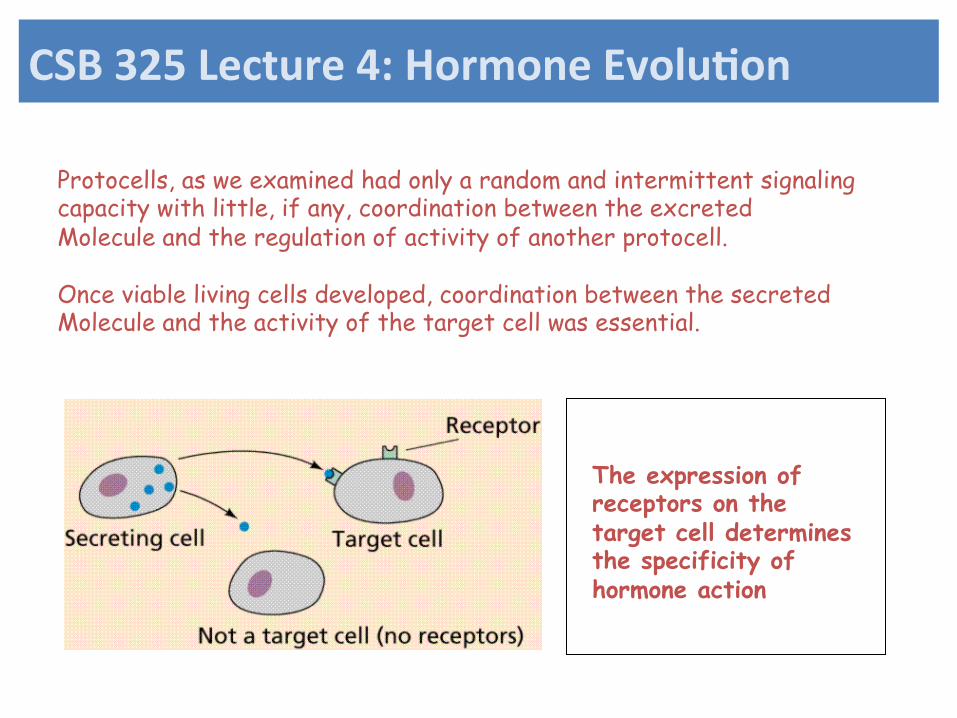
Protocells, as we examined had only a random and intermittent signaling capacity with little, if any, coordination between the excreted Molecule and the regulation of activity of another protocell. Once viable living cells developed, coordination between the secreted Molecule and the activity of the target cell was essential.
The expression of receptors on the target cell determines the specificity of hormone action
CSB$325$Lecture$4:$Hormone$Evolu7on$
First Cells and Signaling Ability First true cell
Metabolic and genetic replication ability Required Attributes
Stable membrane Cytoskeleton adhesion signaling system endocytosis, exocytosis Reasonably accurate replication
Evolution
2 billion years from first cell to first multicellular animal Elaboration of membranes act as a substrate for biosynthetic enzymes required for exocytosis and endocytosis Symbiosis with other organisms Transformation from anaerobic to aerobic formation of organelles
CSB$325$Lecture$4:$Hormone$Evolu7on$
Figure 2-11: Increase in cellular complexity by symbiosis. The formation of organelles
occurs by the ingestion or infection of other cells. Overtime, a symbiotic relationship is
formed. Intracellular or intracrine signalling pathways are required to coordinate the
actions of the new organelle with the actions of the rest of the cell.
Symbiotic cells – increase of complexity
CSB$325$Lecture$4:$Hormone$Evolu7on$
Selection of Types of Signaling Molecules
Amino acids among first amino acids synthesized acid/base ability form elongated chains Peptides and Proteins structural and signalling ability several types of structural organization can carry and transmit information Lipids pass through membrane early prebiotic synthesis Gases and ions present in prebiotic Earth Nucleic acids common as signal molecules but evolutionary origins are not clear Sugars not common as signalling molecule ! !!
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
impart a lot of E and are hydrophillic might not be greatsog mols because they dissolve
Figure 2-15: Evolution of signalling systems in the Metazoa.
A possible timeline for the evolution of cells and their signalling systems
CSB$325$Lecture$4:$Hormone$Evolu7on$
Problems associated with the development Of multicellularity: 1. Adhesion;cells need to be physically connected or attached 2. Nutrient, ion, water, oxygen transport to inner cells 3. Coordination of function: signalling systems 4. Procurement of nutrient sources
CSB$325$Lecture$4:$Hormone$Evolu7on$
Signaling Processes Earliest Cells:
Intracrine and Exocrine abilities More evolved systems
Simple and facilitated diffusion Channel and transporter mediated diffusion
CSB$325$Lecture$4:$Hormone$Evolu7on$
Basic signaling systems: Growth and Differentiation
feeding, growth maturation cell cycle differentiation cell death
Sensory systems
locomotion toward food source away from danger chemical (olfactory input) visual (input) geotaxis mechanical
CSB$325$Lecture$4:$Hormone$Evolu7on$
Figure 3-8: Phylogeny of the phylogenetically oldest lineages of metazoans.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
know how the signalling systems evolved
Shabd A
know the taxonomy
Shabd A
distinct organismsevolved from single celled orgs
Shabd A
basal orgs = something that's close to the original
Shabd A
Shabd A
no symetry between sponges and placozoans
Shabd A
us
Shabd A
mollusks and insects
Shabd A
second mouth = opening
Simplest$Known$Metazoan$
Trichoplax adherens Phylum Placozoa No endocrine or nervous system asymmetrical
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
the only signalling they have might beparacrine signalling. ionic transmission like gap junctions. simple signalling systemdoesn't require much sophistication to communicate and stuff
Signaling in primitive Extant Metazoans
Placozoa and Poriferans as models of early metazoans role of extracellular matrix
Trichoplax adherens as a model
asymmetrical, two tissue types autocrine paracrine signalling only
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Figure 3-10 Photoreception in a sponge larvae. Cilia, associated with a ring of
photoreceptive cells at the base of the larve act to propel the organism away from
sunlight. Reprinted with permission from Leys and Degnan (2001) Biol. Bull.
Epithelial cells of the Poriferans (Sponges) are capable of transmitting
signals
No true nervous system with axons and action potentials
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
works the same way as placozoans. so don't know if they're sister types or how they're related. they're negatively phototaxic, take the larva are symetrical, their photoreceptors can stimulate the surrounding cells by gap junctions which can stimulate the flagella moving it away from light. it's capable of behavior despite the lack of NS
Formation of the Nerve net with Neurons and neurosecretory cells
Placozoans and poriferans do not have nervous systems, however the Radiate phyla, Ctenophora and Cnidaria do. There are no known metazoans that display transitional forms of the nervous System. Phylogenetically older species of deuterostomes and protostomes also show The presence of a nerve net, We assume that the first functional neural/neurosecretory system was a nerve net.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
have a reasonably elaborate NS, there's no transitional forms doe, don't know how it got to that point. but they go have a nerve net, which is fundamentaly different from how we do it. looks like the next slide
A model of a nerve net
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
blue = somatic cells, the other cells are neurosecretory cells, which are connected by axons and are in communication with each other, they get stimulate and interact with each other. can efficiently regulate the somatic cells
Figure 3-13 Nerve net in Hydra. A network of nerve cells is seen in A) hypostome B)
peduncle and C) basal disc after visualization with an anti vasopressin antiserum. See text
for additional details. The scale bars represent 100 µm. Reprinted with permission from
Morashita et al (2003) Elsevier Ltd.
Immunohistochemical visualization of a Nerve net in Hydra
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Signaling Systems in the Radiata Growth and Differentiation pathways comparatively well developed Wnt/frizzled pathway:
wnt (wingless) ligands: a family of polypeptide ligands frizzled receptor: a family of proteins consisting of membrane bound receptors with 7 alpha helices Similar to G-protein coupled receptors but without the G-protein component.
Nuclear receptors: similar to steroid-thyroid hormone receptors although
ligands are not well understood Serine/threonine and Tyrosine kinase based systems Sensory type pathways present
G-protein coupled receptors are present but in low numbers and diversity
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
frizzled = GPCR and is a very early version of it.
Formation of the Nerve net with Neurons and neurosecretory cells
The first neuron/neurosecretory cells may have been derived from Early sensory cells These cells released a chemical signal via a robust depolarizing current resulting from the appropriate sensory input. Later these cells evolved into a new morphology with extended Process that could interact with structures associated with movement And feeding (ie. Cells with cilia). As movement became more complex, interneurons evolved and bridged The sensory cells with the locomotory cells
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Evolution of Signaling pathways
Sensory cell
Locomotor cell with cilia
Sensory input
Ionic coupling
Nervous
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
pre nervous type of system.
Sensory cell
Locomotor cell with cilia
Sensory input
Ionic coupling
Nervous
Sensory cell
Sensory input
Chemical secretion and paracrine diffusion
Neuroendocrine
Evolution of Signaling pathways
CSB$325$Lecture$4:$Hormone$Evolu7on$
Sensory cell
Locomotor cell with cilia
Sensory input
Ionic coupling
Nervous
Sensory cell
Sensory input
Chemical secretion and paracrine diffusion
Neuroendocrine
Sensory cell
Sensory input
Chemical secretion and paracrine diffusion
Endocrine
Non-nervous (depolarizing) Secretory cell
Evolution of Signaling pathways
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
primitive NS
Shabd A
primitive
Shabd A
primitive
Shabd A
these two splitand specialized
Genetic and Morphological Complexity and Signalling Systems: Key points
Development of Nervous and Vascular Systems
Integration of sensory systems and locomotor systems Formation of nervous system and integration with vascular system
Triploblastic organism
Formation of all known bilateral metazoans Increase in complexity of signalling systems
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
endocrine and NS developed complementary systems b/c NS = fast but can only reach few cells and endo = slow but reaches all cells .
Relationship among symmetry, endocrine system, and the nervous system
How an organism moves has a bearing On where sensory organs are located. Once bilateral symmetry evolved, then Sense organs could be concentrated In the direction of movement (i.e. head)
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
not the most efficent way of moving
Shabd A
more efficient, looks like a pnacake and move in two dimensions, sensory sys can be located on the edge, the NS linked all fo these things they have radial NS.
Shabd A
most efficient. they move along one direction. the sensory organs become concentrated on one secion. the amount of E to create a small but efficent it less than the first one. the amount of E utlized for movement is also lesscompared to the other ones.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Relationship among symmetry, endocrine system, and the nervous system
Symmetry allowed for structural similarity among individuals of a population A systemic signaling system could therefore develop in same way in all
individuals. This set the foundation for co-ordination among sensory systems, feeding,
and locomotory structures. This could not be consistently achieved in a non symmetrical animal.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
more effective. it also creates a behaviour which allows them to interact with each other even more and they come together as a population
Basal (phylogenetically older) Lineages of bilateral animals
Deuterostomes Protostomes Acoela
Echinoderms Ecdysozoa Lophotrochozoa
Molluscs Arthropods Chordates
Radiata
Placozoa Porifera
Precambrian explosion
Triploblastic animals
Moving forward toward the Chordates
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
pangenomic theory that somehow the genomesweere similar (?) find more info
Figure$4B2$Evolu'on!of!the!nervous!system!
Formation of the Chordate Nervous System Plan
Segmentation Encephalization Dorsal nerve chord
Radiates: Nerve ring Nerve net
Ventral nerve chord
Segmented nervous System with Ganglia and Ventral nerve chord
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Shabd A
bilateral animals. we see the beginning of a NS
Shabd A
protestomes
A few words about Endocrine systems…
Any cell that evolved the capability for robust paracrine secretion had the potential to become and endocrine cell. These cells were present in all the basic tissue types of the earliest Metazoans, therefore, all tissues had the capability of becoming Endocrine organs. In fact, this is the case today: all tissues and organs have a variety of Substances that are secreted into the blood stream. Therefore, all organs, by definition, may be considered endocrine organs.
CSB$325$Lecture$4:$Hormone$Evolu7on$
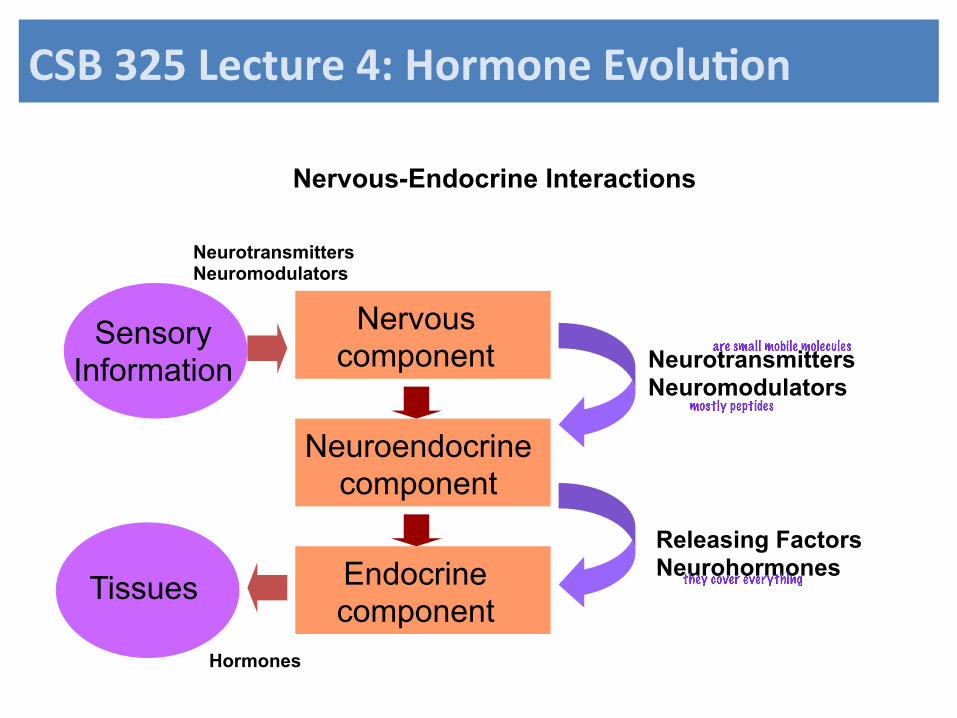
Nervous-Endocrine Interactions
Nervous component
Neuroendocrine component
Releasing Factors Neurohormones Endocrine
component
Sensory Information
Tissues
Neurotransmitters Neuromodulators
Neurotransmitters Neuromodulators
Hormones
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
are small mobile molecules
Shabd A
mostly peptides
Shabd A
Shabd A
they cover everything
General Organization of the Vertebrate Brain
Nerve cord/spinal cord
Hindbrain rhombencephalon
Midbrain mesencephalon
Forebrain prosencephalon
The brain is bilateral consisting of both left and right halves. One side is basically a mirror image of the other. This means also that the neuroendocrine cells are similarly divided.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
shark brain
Human brain development
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
midbrain
Figure 4-13 Organization of the vertebrate nervous system A) Central Nervous
System B) Spinal Cord and ganglia
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
most is on diencephalon main component
Ventricles of the Brain
I
II III IV
The brain and nerve cord are hollow and consists of four ventricles filled with cerebral spinal fluid
Forebrain Midbrain Hindbrain
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
fishes, sharks, amphibians
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Shabd A
evolution, folded back on its self
Cerebrospinal fluid CSF
Resembles ultrafiltrate of plasma There are no substances in CSF that are not found in plasma Proportions differ
I
II III IV
No diffusion barrier
Brain-blood barier There is a barrier between CSF and blood.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
protects brain from shockand stabilizing itand also clears out shit form there
Figure 4-15 Basic components of the vertebrate blood brain barrier
Blood-brain barrier
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
can expand and contract to allow openings
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
windows or holes
Shabd A
ones that are fat soluble they passthrough BBB very easily which thewater soluable can't
Transported: glucose, neutral amino acids used for neurotransmiter synthesis Peptides such as met-enkephalin, IL1, NPY, MCH, CRF use saturatable Transporters Peptides such as calcitonin, amylin, adrenomedulin enter via specific receptors In circumventricular organs
CSB$325$Lecture$4:$Hormone$Evolu7on$
Figure 4-16. Position of circumventricular organs in a human brain
CSB$325$Lecture$4:$Hormone$Evolu7on$Pineal gland – melatonin, rhythms Subfornical organ – regulates fluids, osmoreceptor cells Organum vasculosum – peptides, osmoregulation, fever Area postrema – vomiting centre, osmoregulation Median eminence associated with anterior pituitary Neurohypophysis: neurosecretion AVP, oxytocin
The posterior lobe consists of neurosecretory terminal originating from cells in the hypothalamus. In mammals, there are two main hormones that are released: Vasopressin and Oxytocin. In non-mammals variants of vasotocin are released.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Chordate Evolution and Phylogeny
CSB$325$Lecture$4:$Hormone$Evolu7on$
Figure 4-18. Neural Complex in a Tunicate. A number of hypothalamic and pituitary
hormones and processing enzymes have been found in the neural complex suggesting a
relationship with the chordate hypothalamo-pituitary system. See text for further
details.CS, cloacal siphon; OS, oral siphon. Reprinted with permission from Kawamura
et al, (2002) Elsevier Science (USA).
Neuroendocrine secretion in a Tunicate
CSB$325$Lecture$4:$Hormone$Evolu7on$
Structure of Agnathan Pituitary Glands The jawless fishes, or agnathans, are the most primitive known chordates. These fish which include the hagfishes (Myxini) and the lampreys (Cephalospidomorphi) are separate classes. The hagfishes lack a vertebrae and therefore are not classified As vertebrates. The Cephalospidomorphi do possess a vertebrae and are considered The phylogenetically oldest vertebrates. The Cephalospidomorphi likely evolved At least 50 million years after the Myxini. Both groups of fishes are highly degenerate. Despite this, there are common aspects to their structure of pituitary and hypothalamus That provide us with clues as to how this system evolved in the vertebrates
Hagfish Class: Myxini
Lamprey Class: Cephalospidomorphi
100
200
300
400
500
600
CSB$325$Lecture$4:$Hormone$Evolu7on$
Myxini Cephalospidomorphi
Agnathan Pituitary Structure
For both the Myxini and the Cephalospidomorphi there appears to be a complete separation of the pars distalis from the rest of the hypothalamus. Diffusion of the hypothalamic releasing factors appears to be the main route of communication between the hypothalamus and pars distalis. However, a simple portal system may be present although this is unclear. In hagfishes, (Myxini), the pars nervosa is a relatively undifferentiated part of the hypothalamus where the axonal termini of neurosecretory cells impinge on a vascularized region at the hypothalamus wall. In lampreys (Cephalospidomorphi) the pars nervosa becomes more differentiated into a discrete lobular structure.
Pars distalis Pars intermedia Pars nervosa
Structure of Agnathan Pituitary Glands CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
communicate throughstemic bloodsnot efficentbut it works
Shabd A
much closer association with distalisn and nervosa
Shabd A
both don't have cortical sys
Hagfish and lampreys may also utilize the systemic blood as a route for releasing factors to regulate the pars distalis Ectopic transplantation of the hagfish pars distalis in other regions of the body, for example do not appear to compromise the normal physiology of the these animals. The various cell types in the pars distalis are relatively intermingled.
Structure of Agnathan Pituitary Glands
CSB$325$Lecture$4:$Hormone$Evolu7on$
The Class Chondrichthyes or cartilaginous fish are the phylogenetically oldest Group of gnathostomes or jawed fish. The cartilaginous fish include the Subclass Elasmobranchii (sharks, skates and rays) and the Subclass Holocephali (chimaeras). The cartilaginous fish evolved 50-100 million years After the lineage leading to the lampreys and about 400 million years before Present. The Elasmobranchs and Holocephali diverged early in Chondrichthyan evolution, but still possess a number of related features.
Elasmobranchii Holocephali
Chondricthyan Pituitary Gland
100
200
300
400
500
600
CSB$325$Lecture$4:$Hormone$Evolu7on$
General Structure of Elasmobranch Pituitary Gland
Pars nervosa
Pars distalis
Pars intermedia
Portal system The Elasmobranch pituitary consists of a fused pars nervosa and pars intermedia. In this situation where there is considerable Intermingling of pars nervosa and pars Intermedia, it is referred to as a pars Neurointermedia or Neurointermediate lobe. There is a well developed portal system that connects most of the pars distalis with the hypothalamus. However, the ventral lobe, has no vascular or nervous connection to the rest of the brain. Releasing factors are secreted into the systemic blood supply to communicate with the ventral lobe.
Neurointermediate lobe
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Shabd A
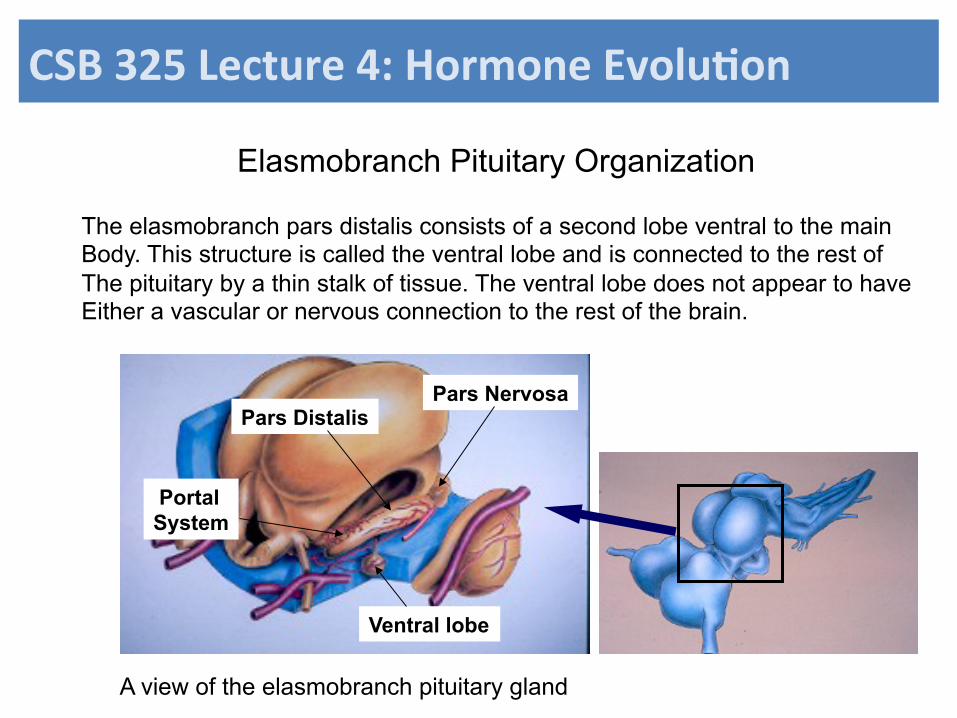
Elasmobranch Pituitary Organization
Ventral lobe
Pars Distalis Pars Nervosa
Portal System
The elasmobranch pars distalis consists of a second lobe ventral to the main Body. This structure is called the ventral lobe and is connected to the rest of The pituitary by a thin stalk of tissue. The ventral lobe does not appear to have Either a vascular or nervous connection to the rest of the brain.
A view of the elasmobranch pituitary gland
CSB$325$Lecture$4:$Hormone$Evolu7on$
Pituitary and Diencephalon Structure in the Holocephali
Buccal lobe
Brain
Position of Pituitary Gland
The structure of the pituitary in holocephalans is similar to that of the Elasmobranchii. The main difference lies in the structure of the ventral lobe. In holocephalans, the ventral lobe is called a buccal lobe and is found in the roof of the mouth. It is separated from the brain by a layer of cartilage. The hypothalamus communicates with the buccal lobe by secreting releasing factors into the systemic blood.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
Although there are few studies to draw upon, the location of cells in the pituitary gland in the Cartilaginous fish appears to have some organization. For example, the gonadotropes are located primarily in the ventral lobe of elasmobranches (or the buccal lobe of holocephalons). Although there is some interdigitation of the pars nervosa with aspects of the pars distalis, there is currently no evidence that there is any direct neural link between the hypothalamus and pars distalis.However, given the organization of the neurosecretory cells of the hypothalamus and the presence of interdigitation in cartilaginous fish, there is the distinct potential that some form of rudimentary neural connection may exist. If so, one might also expect that some of the basic pituitary cell types are organized to a limited degree in clusters of related cells.
Pituitary Cell Organization in the Cartilaginous Fish
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
interdigitation
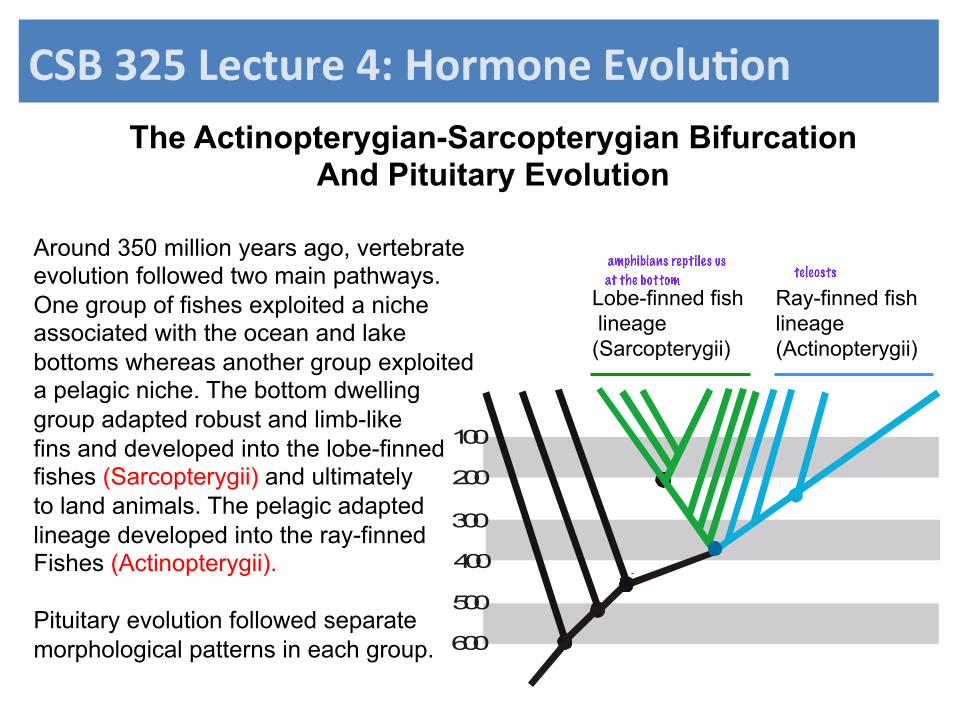
The Actinopterygian-Sarcopterygian Bifurcation And Pituitary Evolution
100
200
300
400
500
600
Ray-finned fish lineage (Actinopterygii)
Lobe-finned fish lineage (Sarcopterygii)
Around 350 million years ago, vertebrate evolution followed two main pathways. One group of fishes exploited a niche associated with the ocean and lake bottoms whereas another group exploited a pelagic niche. The bottom dwelling group adapted robust and limb-like fins and developed into the lobe-finned fishes (Sarcopterygii) and ultimately to land animals. The pelagic adapted lineage developed into the ray-finned Fishes (Actinopterygii). Pituitary evolution followed separate morphological patterns in each group.
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
at the bottom
Shabd A
teleosts
Shabd A
amphibians reptiles us
Evolu7on$of$the$Pituitary$Portal$System$
Early jawed vertebrate
Lobe-finned fishes/tetrapods Ray-finned fishes
oc
oc
oc
vIII vIII
vIII
Elaboration of portal system Direct neural connection
Jawed vertebrates began with a pituitary structure similar to chondrichtyan fishes where there was a portal system and a more or less distinct pars distalis and pars nervosa. In the lobe-finned fishes and tetrapods, the portal system became more developed, whereas in the ray-finned fishes the portal system was lost and a direct neural connection developed
Neurosecretory cells With projections to The median eminence And portal system
Neurosecretory cells With projections to the pars nervosa
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
cells are intermingled here
Shabd A
cells are clumped by groups
Amniotes The earliest amniote tetrapods appears about 30 million years after the evolution of the first amphibians. This group includes the mammals, reptiles and birds, and refers to the amniotic egg present in all groups. The early amniotes began to radiate extensively in the Carboniferous and early Permian times, displacing the non-amniotes (early amphibians) from those habitats and niches. Evidence suggested that virtually all of the early amniotes were carnivorous. The extensive radiation of insects at this time is thought to have been a food source that was exploited by the early amniotes thus acting to stimulate this radiation of species. Physiological changes that accompanied this adaptation included changes in respiratory mechanisms, diuresis and excretion, reproduction and integration of sensory systems.
100
200
300
400
500
600
Phylogenetic Position of Amniotes
CSB$325$Lecture$4:$Hormone$Evolu7on$
Structure of Reptile Pituitary Glands
100
200
300
400
500
600
rhynchocephalia
chelonia
crocodilia
squamata squamata
There are four subclasses of reptiles: Chelonia (tortoises, turtles and terrapins), Rhynchocephalia (tuataras), crocodilia (alligators, crocodiles, caimans, gharials) And squamata (lizards snakes).
CSB$325$Lecture$4:$Hormone$Evolu7on$
Structure of Reptile Pituitary Glands
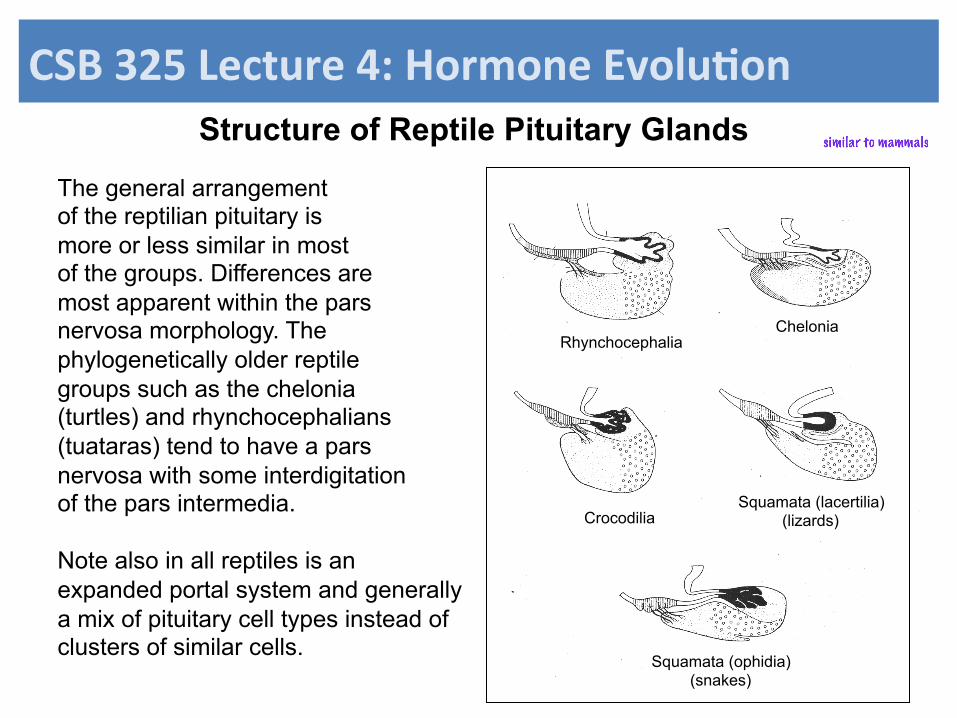
The general arrangement of the reptilian pituitary is more or less similar in most of the groups. Differences are most apparent within the pars nervosa morphology. The phylogenetically older reptile groups such as the chelonia (turtles) and rhynchocephalians (tuataras) tend to have a pars nervosa with some interdigitation of the pars intermedia. Note also in all reptiles is an expanded portal system and generally a mix of pituitary cell types instead of clusters of similar cells.
Rhynchocephalia Chelonia
Crocodilia Squamata (lacertilia)
(lizards)
Squamata (ophidia) (snakes)
CSB$325$Lecture$4:$Hormone$Evolu7on$
Shabd A
similar to mammals
Structure of Teleost Pituitary Gland
100
200
300
400
500
600
The Teleostei are a vast assemblage of fish that include about 25,000 species. The pituitary gland is unusual in that cells of the pars distalis are clustered in nuclei of similar cells. There is extensive interdigitation of the neural lobe with the rest of the pituitary, and direct neural connection between the hypothalamus And pars distalis
CSB$325$Lecture$4:$Hormone$Evolu7on$
Anguilla anguilla (eel) as a model for the teleost pituitary gland
The teleost pituitary gland morphology represents the most advanced structural features among the fishes. Two major changes have occurred: neurosecretory cells of the hypothalamus send their axons through the extensive interdigitations of the pars nervosa with the pars intermedia and pars distalis. Within the pars distalis cells have become grouped into discrete clusters. Thus, in this manner the releasing factor can stimulate the appropriate pituitary cell.