BSP-UP Professorial Chair Lectures 19-20 October 2015 Bangko Sentral ng Pilipinas Malate, Manila Lecture No. 7 Philippine Fungal Diversity: Benefits and Threats to Food Security by Dr. Christian Joseph R. Cumagun BSP UP Centennial Professor of Agriculture
Transcript
BSP-UP Professorial Chair Lectures 19-20 October 2015
Bangko Sentral ng Pilipinas Malate, Manila
Lecture No. 7
Philippine Fungal Diversity: Benefits and Threats to
Food Security
by
Dr. Christian Joseph R. Cumagun BSP UP Centennial Professor of Agriculture
Page 1 of 13
Philippine Fungal Diversity: Benefits and Threats to Food Security
Christian Joseph R. Cumagun1
Abstract
Fungal diversity plays a vital role in sustaining human life, either benefiting or threatening food security. Benefits derive from fungi range from food, medicine, to decomposition of organic matter, recycling of nutrients and plant growth promoter, including biological control of fungal plant pathogens. Fungal threats as pathogens of agricultural crops and mycotoxin producers may be generally less as compared with the benefits they provide to mankind. The paper briefly describes the beginnings and development of mycology as a science in the Philippines and presents the main repositories of fungal cultures in the Philippines. The four case studies generated from my research are highlighted, namely: biological control of plant pathogens; microbial gifts from the rainforests for agriculture; breeding for resistance against cereal blast and sheath blight pathogen including population genetic studies; and Fusarium diseases and mycotoxins. In view of habitat loss and climate change threatening fungi, there is an urgent need to promote fungal conservation for a sustainable agriculture and healthy planet.
1 Professor of Plant Pathology, Crop Protection Cluster, College of Agriculture, University of the Philippines Los Baños (UPLB), College, Laguna, 4031 Philippines E-mail: [email protected]
Page 2 of 13
Philippine Fungal Diversity: Benefits and Threats to Food Security
Christian Joseph R. Cumagun Introduction
Microorganisms particularly fungi are responsible for the outstanding breakthroughs in medicine and agriculture, including the threat it imposes on food security and food safety and yet very little is known about the need to conserve microbial diversity (Doyle et al., 2005; Colwell 1997). Mycology for most people is unheard of unless one mentions the word “mushroom” or “fungi” and more often than not the study of fungi is considered under botany because they are classified as lower plants. Mycology is first encountered in basic biology courses but very few students would be interested to pursue this study as a major in college and as a career in life. The reasons are varied and mostly stem from ones interest in other fields in biology whose organisms are more visible and that can be easily appreciated for its beauty like the animals and plants (Moore et al. 2008; Minter, 2011). Emphasis has focused on the importance of conservation of these life forms, but conservation of fungi is lagging behind due to lack of appreciation of the role of fungi in the environment by other biologists and conservationists. After all, threats to the survival of fungi such as climate change, loss of habitat and pollution are the same for the rest of living organisms. For each plant that gets extinct, a fungal species associated with it also disappears (Strobel, 2002).
In general, mankind benefits from fungi more than it suffers. Although some fungi are also major threats to food security, humanity has been beneficiary of the products derived from them including breakthroughs in modern medicine and the vital role they play in cycling of carbon and nutrients in the ecosystem, yet little do we know about conserving thousands of these fungal species (Hawkworth, 1991). It is estimated that there are 1.5 million species of fungi but only 10% of these have been discovered (Hawksworth, 2001). This estimate is rather conservative as there could be 5.1 million of fungal species based on high-thoroughput sequencing methods according to Blackwell (2011). Despite of declining research support, proper education and training of the next generation of specialists in the field of mycology is crucial if we want to ensure the viability and continuity of this neglected discipline. It is important to tell anyone regardless of age, background or occupation the nuts and bolts of mycology. The discipline of mycology should be a “megascience” (Hawksworth, 2009). In this chapter, the historical development of mycology as a science and the output of my research team in light of the importance of fungal diversity on food security in the Philippines are discussed. The genesis and development of Philippine mycology
Mycological expeditions in the Philippines have been documented as early as 1820 (Graff, 1916) but teaching and research on fungi formally began during the American colonial occupation through the founding of the University of the Philippines College of Agriculture in Los Baños, Laguna (Teodoro, 1937). During the Spanish period, there had been also reports of fungal collection mostly conducted by European and American mycologists (Teodoro, 1937). Graff in 1916 compiled a bibliography and new species of Philippine fungi from 1820 to 1915 comprising hundreds of newly described and first reports of fungal species. The most comprehensive fungal record in the Philippines was the classic book entitled, “An Enumeration of Philippine Fungi published” in 1937 by Dr. Nicanor G. Teodoro, a noted Filipino pioneer plant pathologist. For further readings on the growth and development of Philippine mycology, the reader is referred to the paper review of Tadiosa (2012). In this review, the author stated that the fungal diversity in the Philippines is estimated at 3956 species and 818 genera.
Page 3 of 13
A Philippine society solely devoted on fungi - Mycological Society of the Philippines (MSP) was organized on October 19, 1998. At the University of the Philippines Visayas in Iloilo, Dr. Tricita H. Quimio, professor of mycology at the University of the Philippines Los Baños (UPLB) was appointed chairman of the organizing committee by a small group of mycologists attending a conference on tropical microbial diversity. She was then the Philippine representative to the International Mycological Association Committee for Asia (IMACA). The first scientific meeting was held on April 8, 1999 at the University of the Philippines Los Baños (UPLB) with 167 charter members. Dr. Kevin B. Hyde of the University of Hongkong talked on “Where are the missing fungi” and Dr. T. K. Tan of the National University of Singapore on “Mycology in ASEAN: Where are we.”
Despite the strong historical foundation of fungal research in the Philippines, there are only a few recent developments in terms of fungal survey to advance the field probably because of dearth of mycologists in the country. Some examples include work on wood decaying fungi in Philippine Dipterocarps (Tadiosa and Militante, 2006); macrofungi of Bazal-Baubo watersheds in Aurora province (Tadiosa et al., 2011) and Mt. Makulot, Cuenca, Batangas (Tadiosa et al., 2007). Tadiosa and Briones (2013) reported 75 species belonging to 36 genera and 23 families of fungi of Taal volcano protected landscape in Southern Luzon. Entomopathogenic fungi consisted of 53 species under 22 genera were reported on selected areas in the Philippines from 1998 to 2001 (Villacarlos and Meija, 2004). Fungi from bamboos were intensively studied by Cai et al (2003) on fresh water and river ecosystems and Hyde et al. (2002) on a vertical stratification. Su et al. (2014) studied the distribution and diversity of marine fungi in Manila Bay. The prospects of using DNA based methods such as the LSU and ITS, for identifying the forest fungi causing root and wood rot in the Philippines has just begun (Case et al., 2013). Case study 1: Biological control of fungal plant pathogens
Our work on biocontrol started with Trichoderma as a means to manage sheath blight
of rice caused by Rhizoctonia solani (Cumagun and Lapis, 1993; Cumagun and Ilag 1993a and 1993b). Our group is also the first to molecularly characterize Philippine Trichoderma strains (Cumagun et al. 2000). T. harzianum has been found to be the dominant strain in rice and rice based cropping systems whereas T. viride is present occasionally. The latter species has been found to possess a coconut smell indicative of volatile organic compounds that are inhibitory to many fungal pathogens (Cumagun et al. 2000). Of the many Trichoderma strains that have been isolated and screened for biocontrol of plant diseases in the Philippines, only a single strain has reached product commercialization (Cumagun, 2012).
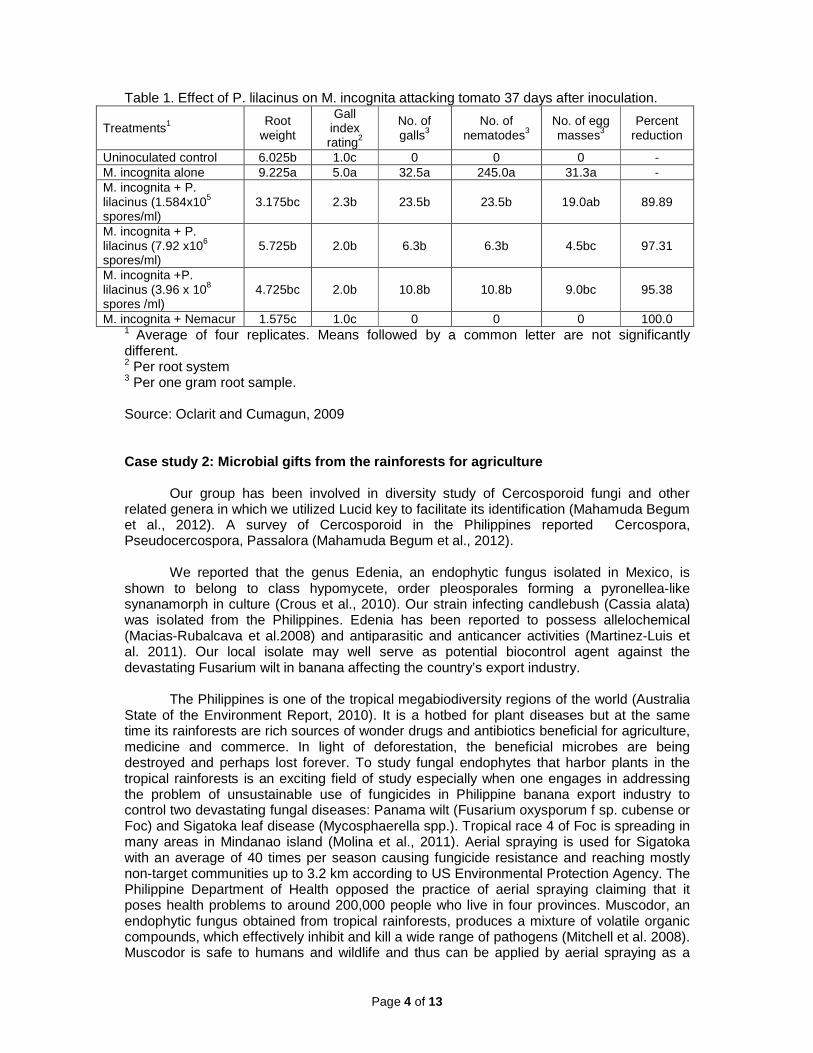
We also evaluated another fungus Paecilomyces lilacinus strain UP1 as biological control agent of the root knot nematode Meloidogyne incognita attacking tomato under screenhouse condition (Oclarit and Cumagun, 2009). Meloidogyne infects numerous hosts in agriculture and has a worldwide distribution. P. lilacinus was formulated on rice substrate in powder form. Root weight, gall index rating, number of galls, egg masses and nematodes per one gram root sample were determined and percent reduction in gall number was computed. Root weight and gall index ratings were significantly higher in untreated plants than those with P. lilacinus and with the commercial fungicide Nemacur. Number of galls, nematodes and egg masses per one gram root sample were significantly reduced by the application of P. lilacinus at all levels and this was comparable with Nemacur. However, egg mass count in plants treated with the lowest concentration of the biocontrol agent was not significantly different from the noninoculated control. Percent reduction in gall number was the highest at treatment with 7.92 x 106 spores per ml of P. lilacinus (Table 1). For a thorough review of biocontrol of plant nematodes, the reader is referred to Cumagun and Moosavchap (2015).
Page 4 of 13
Table 1. Effect of P. lilacinus on M. incognita attacking tomato 37 days after inoculation.
Treatments1 Root
weight
Gall index rating2
No. of galls3
No. of nematodes3
No. of egg masses3
Percent reduction
Uninoculated control 6.025b 1.0c 0 0 0 - M. incognita alone 9.225a 5.0a 32.5a 245.0a 31.3a - M. incognita + P. lilacinus (1.584x105 spores/ml)
3.175bc 2.3b 23.5b 23.5b 19.0ab 89.89
M. incognita + P. lilacinus (7.92 x106 spores/ml)
5.725b 2.0b 6.3b 6.3b 4.5bc 97.31
M. incognita +P. lilacinus (3.96 x 108 spores /ml)
4.725bc 2.0b 10.8b 10.8b 9.0bc 95.38
M. incognita + Nemacur 1.575c 1.0c 0 0 0 100.0 1 Average of four replicates. Means followed by a common letter are not significantly different. 2 Per root system 3 Per one gram root sample. Source: Oclarit and Cumagun, 2009 Case study 2: Microbial gifts from the rainforests for agriculture
Our group has been involved in diversity study of Cercosporoid fungi and other related genera in which we utilized Lucid key to facilitate its identification (Mahamuda Begum et al., 2012). A survey of Cercosporoid in the Philippines reported Cercospora, Pseudocercospora, Passalora (Mahamuda Begum et al., 2012).
We reported that the genus Edenia, an endophytic fungus isolated in Mexico, is
shown to belong to class hypomycete, order pleosporales forming a pyronellea-like synanamorph in culture (Crous et al., 2010). Our strain infecting candlebush (Cassia alata) was isolated from the Philippines. Edenia has been reported to possess allelochemical (Macias-Rubalcava et al.2008) and antiparasitic and anticancer activities (Martinez-Luis et al. 2011). Our local isolate may well serve as potential biocontrol agent against the devastating Fusarium wilt in banana affecting the country’s export industry.
The Philippines is one of the tropical megabiodiversity regions of the world (Australia State of the Environment Report, 2010). It is a hotbed for plant diseases but at the same time its rainforests are rich sources of wonder drugs and antibiotics beneficial for agriculture, medicine and commerce. In light of deforestation, the beneficial microbes are being destroyed and perhaps lost forever. To study fungal endophytes that harbor plants in the tropical rainforests is an exciting field of study especially when one engages in addressing the problem of unsustainable use of fungicides in Philippine banana export industry to control two devastating fungal diseases: Panama wilt (Fusarium oxysporum f sp. cubense or Foc) and Sigatoka leaf disease (Mycosphaerella spp.). Tropical race 4 of Foc is spreading in many areas in Mindanao island (Molina et al., 2011). Aerial spraying is used for Sigatoka with an average of 40 times per season causing fungicide resistance and reaching mostly non-target communities up to 3.2 km according to US Environmental Protection Agency. The Philippine Department of Health opposed the practice of aerial spraying claiming that it poses health problems to around 200,000 people who live in four provinces. Muscodor, an endophytic fungus obtained from tropical rainforests, produces a mixture of volatile organic compounds, which effectively inhibit and kill a wide range of pathogens (Mitchell et al. 2008). Muscodor is safe to humans and wildlife and thus can be applied by aerial spraying as a
Page 5 of 13
viable alternative to fungicides and the banned soil fumigant - methyl bromide. This technology is expected to ensure food security and safety, promote biodiversity conservation, protect the health of the citizens and the environment, and provide sustainable pest management solutions (Figure 1).
Figure 1. Benefits derived from Muscodor technology
A list of commercialized fungal based products developed by National Institute of Microbiology and Biotechnology (BIOTECH) at UPLB is shown in Table 2 whereas Table 3 shows the repositories of fungal cultures and specimens in the Philippines. All microbial collections in Table 2 are members of the Philippine National Collection of Microorganisms (PNCM). PNCM serves as the national repository of microbial strains in the country. It offers microbiological analyses, identification of isolates and preservation of cultures. PNCM is an accredited laboratory of the Food and Drug Administration -Department of Health (FDA-DOH) and identified as a National Reference Laboratory (NRL) for microbiology by the Bureau of Products Standard of the Department of Trade and Industry (DTI). It is also a member of the World Federation for Culture Collection. At the international level, the Asian Consortium for the Conservation and Sustainable Use of Microbial Resources (ACM) was founded in 2004 by 12 Asian countries (Cambodia, China, Indonesia, Japan, Korea, Laos, Malaysia, Mongolia, Myanmar, Philippines, Thailand and Vietnam) on the occasion of the 10th International Congress on Culture Collections in Tsukuba, Japan to strengthen the relationships among Asian countries, as well as encourage and facilitate international cooperation for biotechnology using microbiological resources in Asia. Currently, 22 organizations from 13 countries including India are members of ACM.
Page 6 of 13
Table 2. BIOTECH commercialized products derived from Philippine fungi. Product Name Fungi Uses
Brown Magic Endomycorrhiza Growth promoter of orchid seedlings and protects them from diseases
Bio Quick Trichoderma sp. Bio-organic fertilizer Bio Green Trichoderma sp. Bio-organic fertilizer Mycogroe Ecto Mycorrhizal Fungi Bio fertilizer
Mycovam Vesicular Arbuscular Fungi Soil-based biofertilizer for crops except crucifers and lowland rice
VAM Root Inoculant Vesicular Arbuscular Fungi Growth promoting subtances and disease control
Lipase Rhizopus sp. Hydrolyses coconut oil to produce high value β- monoglyceride
Pectinase Aspergillus sp. Juice and wine clarification oil extraction from freshly grated coconut and essential oil extraction
Microbial Rennet Rhizopus chinensis Milk coagulation for cheese production
Table 3. Major repositories of fungal cultures and specimens in the Philippines. Name of Culture Collection Description
UPLB Mycological Herbarium
A collection of about 11,000 ascomycetes, basidiomycetes and myxomycetes. It provides identification services of macrofungal specimens to students, university faculty and researchers.
UPLB Microbial Culture Collection
A repository of microorganisms and provides cultures upon request for instruction, research and industrial purposes. It offers other services such as microbiological analysis of samples which includes microbial cultural count, isolation and identification, water quality analysis, short-term training in basic microbiological techniques and other microbiologically-related analyses
Philippine National Herbarium Collection (PNHC)
Fungi is just one part of the vast botanical collections of 180,000 specimens of the PNHC.
Microbial Research and Service Laboratory (MRSL), University of the Philippines, Natural Sciences Research Institute (UPCC) in Diliman
MRSL is one of the four technical laboratories of the Natural Sciences Research Institute. MRSL houses the UPCC which maintains about 500 cultures of bacteria, fungi and yeasts in viable and pure form.
University of Santo Tomas Collection of Microbial Strains (USTCMS)
The USTCMS offers a unique collection of indigenous, clinical and biotechnological microbial strains such filamentous fungi and yeasts of terrestrial and marine origin, microalgae and Streptomyces along with bacteria.
Page 7 of 13
Case study 3. Breeding for resistance and population genetic structure: cereal blast and sheath blight pathogen
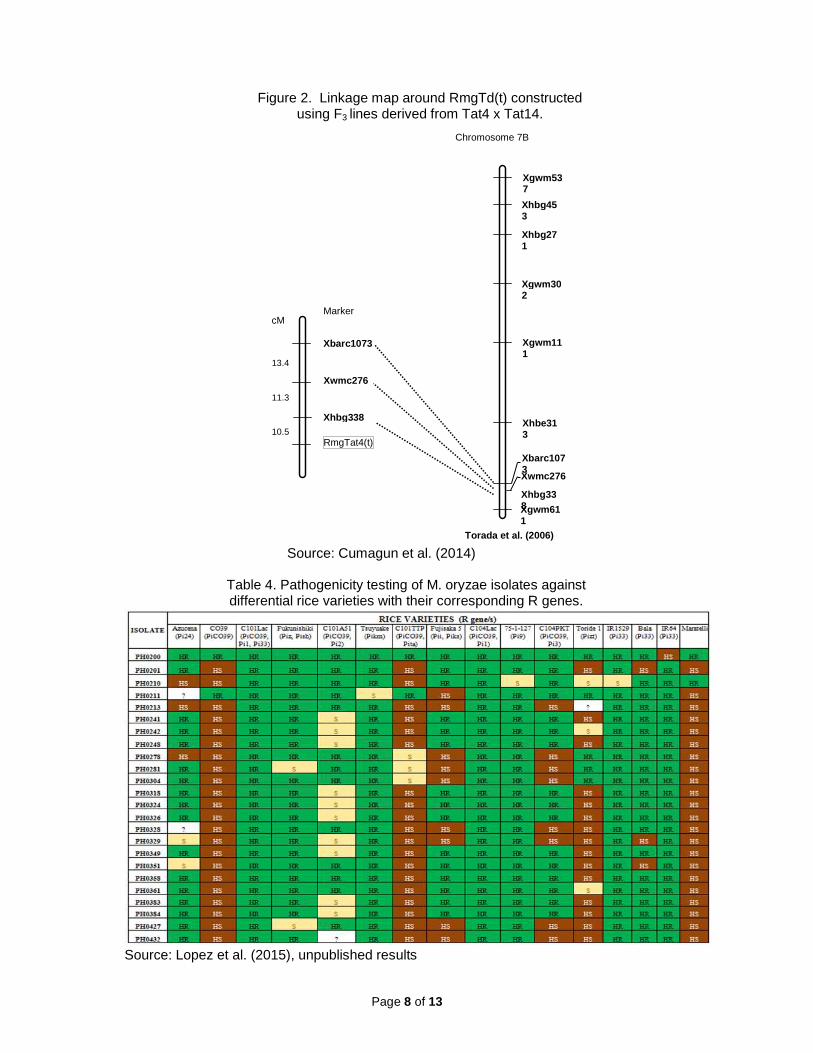
Magnaporthe is considered to be the world’s most important pathogen of cereals and a causal agent of blast disease of approximately 100 grass species. These isolates are divided into host-specific subgroups such as Oryza isolates pathogenic to rice, Setaria isolates pathogenic to foxtail millet, Setaria isolates pathogenic on Setaria spp. including foxtail millet (S. italica), Eleusine isolates pathogenic on Eleusine spp. including finger millet (E. coracana)), etc. (Kato et al., 2000). In the 1980s in Brazil, blast disease was reported for the first time in wheat, the most widely cultivated crop in the world. Its causal agent proved to be a new subgroup (Triticum isolates) of M. oryzae which had spread to other countries in South America and became a major threat to wheat production. North America is currently under alert regarding the spread of this disease. Conquest of the blast disease in rice alone will be enough to feed 60 million people yearly. Fungicides by foliar and seed treatment provides insufficient control of the disease, thus development of resistant cultivars remains to be an effective, economical and environmentally friendly way to manage the disease. An isolate from oat in the Kobe University collection was isolated in 1980s in Brazil. An infection assay revealed that this isolate (Avena isolate) was an Avena-specific strain. Although this strain has not yet caused a serious epidemic of oat blast, there is a possibility that the next blast outbreak may occur on oat in South America. To develop a strategy to combat against oat blast, genetic analyses of its avirulence/virulence and identification of resistance genes are needed. In the present study, we mapped PWT3 by molecular approach and demonstrated that it is an avirulence gene under the control of gene-for-gene interactions by successfully identifying its corresponding resistance gene RmgTd(t) in wheat and barley cultivars which we mapped on chromosome 7b (Cumagun et al., 2014; see Figure 2).
To keep abreast of the current genetic diversity of the populations of Magnaporthe
oryzae, the rice blast disease pathogen, we characterized the genetic diversity of 453 monoconidial isolates recently collected from different geographic areas in the Philippines in experimental plots and in farmer fields (Lopez et al., 2013). With 13 microsatellite markers, a total of 93 new genotypes were determined when compared to the reference isolates representing the genotypes previously published. Analysis revealed a weak geographic structuring of pathogen genotypes, with some of those collected from Luzon clustering with those from Visayas or Mindanao. Nevertheless, none of the strains from Mindanao clustered with those from the Visayas or vice versa. Significant differences were noted on isolates collected from distinct agroecosystems and isolates collected from the upland tended to be more diverse when compared to isolates from lowland. This structure likely results from limited natural migration and from some events of long distance migration, probably through the transport of infected seeds. A subset of 30 isolates representative of the genotypic diversity were tested for fertility status. Of these, eight induced the production of perithecia, of which six also were only male-fertile and two were also female-fertile. These fertile isolates were mostly found in the upland agroecosystem. In addition, the same isolates and 23 additional isolates were subjected to PCR test for mating type. MAT1.1 isolates represented 36% of our sample whereas it represented only 5% in the collection of reference strains. This result confirms that this new sampling improves our knowledge of the blast population in the Philippines. Pathogenicity test of a subset of 24 isolates inoculated to 16 differential rice varieties demonstrated that most isolates were avirulent to varieties C101Lac and C104Lac which carry the Pi1 resistance gene (Table 4). The very popular Pi33 gene, integrated into the varieties IR 64, Bala, and IR1529 also conferred resistance to most of the isolates. In addition to Pi1 and Pi33, other possible candidate resistance gene for plant breeding effort to manage the rice blast disease in the Philippines are Pi9, Pikm, and the combination of Piz and Pish (Lopez et al, 2015, unpublished results).
Page 8 of 13
Figure 2. Linkage map around RmgTd(t) constructed using F3 lines derived from Tat4 x Tat14.
Marker
Xwmc276
Xhbg338
Xbarc1073
RmgTat4(t)
cM
11.3
13.4
10.5
Torada et al. (2006)
Chromosome 7B
Xgwm611
Xhbg338
Xbarc1073
Xhbe313
Xwmc276
Xgwm111
Xgwm302
Xhbg271
Xgwm537
Xhbg453
Source: Cumagun et al. (2014)
Table 4. Pathogenicity testing of M. oryzae isolates against differential rice varieties with their corresponding R genes.
Source: Lopez et al. (2015), unpublished results
Page 9 of 13
The rice sheath blight disease, caused by the fungal pathogen Rhizoctonia solani
AG1-IA is one of the most economically important diseases worldwide. Our group conducted a study to determine the genetic structure and reproductive mode of 18 rice-infecting populations from China, Japan and the Philippines. This study is important because no population genetic data of R. solani has been reported among three of the most important rice production countries in Asia. A total of 423 unique multilocus genotypes were found among 717 fungal isolates genotyped using nine microsatellite loci. The populations of R. solani AG1-IA from the three countries exhibited a mixed reproductive mode consisting of sexual recombination and clonal reproduction. However, a moderate to high clonal fraction (0.38-0.51) indicated that asexual reproduction by sclerotia represent an important source of inoculum in field populations. The current levels of population subdivision varied among and within countries. While Фst values in some populations from China (Фst = 0.11- 0.36; P ≤ 0.05) and almost all populations from the Philippines (Фst = 0.5-0.6; P ≤ 0.05) indicated a moderate level of subdivision among populations while no significant subdivisions in all populations in Japan were found. There was evidence of isolation by distance with a genetic distance RST (R2 = 0.45; P ≤ 0.001). Migration rate from Japan to China was the highest while migrants from the Philippines to China and Japan were rather small. Migration rate supports the number of immigrant genotypes detected across populations (Cumagun et al 2015, unpublished results; see Figure 3).
Figure 3. Structure inferred membership coefficient for multilocus microsatellite genotypes of Rhizoctonia solani AG- 1 IA
from rice-infecting population samples from China, Japan and the Philippines.
Source: Cumagun et al. (2015, unpublished results
Page 10 of 13
Case study 4. Fusarium diseases and mycotoxins
Panama wilt, caused by Fusarium oxysporum f. sp. cubense (Foc) is considered one of the most devastating banana diseases in recorded history. The disease is threatening the banana industry due to Tropical Race 4 (TR4) infecting Cavendish cultivar. Using race-specific primers, FocTr4-F/FocTr4-R and Foc-1/Foc-2, the absence of TR4 in Luzon has been confirmed, however, the occurrence of subtropical race 4 (ST4) has been reported, which should also be taken in to consideration as the latter can also cause severe damage under favorable condition. Overall, high genetic variability was recorded in Foc Philippine population using M13, ERIC and REP primers, which might render host resistance vulnerable. This is the first study on genetic diversity of Foc in the Philippines. (Aguilar-Hawod, 2014). Recently, Solpot (2015) found that out of 75 Foc isolates collected from South Central Mindanao, 25 were confirmed as TR4 from Lakatan (12), Cavendish (11) and Latundan (2).
Fusarium verticillioides is a major pathogen of corn worldwide and is the most
frequently reported Fusarium species in corn causing seedling, stalk, and ear rots. The fungus is also common in farm and storage samples, reduce kernel quality and nutritional value and is a potential threat to human and animal health as a result of a mycotoxin called fumonisins that it produces. Pre- and post-harvest grain losses could ranged from 30 to 80% reduction. Our group evaluated Fusarium contamination and mycotoxin production in maize seeds in the main island of Luzon, Philippines. Fusarium species producing toxins are always associated with cereals from warm tropical countries. This work suggests that mycotoxins pose a serious food problem in the Philippines but that there has been extremely lack of research. For example, the FAO website for the Philippines data relate only to two corn fumonisins (FB-1, FB-2) which were found in >50% maize samples (D’Mello and Macdonald, 1998; Placinta, D’Mello and MacDonald, 1999). High diversity in the population of F. verticillioides exists in Philippine corn fields and that almost all the isolates tested were fumonisin producers using molecular tools (Magculia and Cumagun, 2011). Similarly, high variation in aggressiveness was found in the population, suggesting the ability of the fungus to evolve in response to chemicals and resistant cultivars (Cumagun et al., 2011). The project provided important information relevant to human or animal consumption of corn and generated useful results such as the report on the high fumonisin ability of the isolates from the Northern part of Luzon which is a considered a major corn growing area in the Philippines (Cumagun et al, 2009). Improved farming and post handling storage and processing practices for grains should be a priority concern so that post harvest grain losses and the risk of mycotoxin contamination could be significantly reduced. Conclusion and future outlook
In 1998, International Congress of Plant Pathology President David Ingram gave a thought provoking paper on why there is a need to conserve biodiversity not only of beneficial microbes but plant pathogens as well. The reasons are: 1) they serve a role in revealing genetic diversity in potential breeding material and provide vital screens for the development of new cultivars, 2) they are the raw material for much of the basic scientific research on life cycles and genetics that generate an understanding of pathogen variation, evolution and population dynamics, and 3) they constitute a potentially significant biotechnological resource of particular importance to the genetic engineer (Ingram, 1998).
Research on fungal diversity in the Philippines is limited as there is a lack of trained
personnel in the field of mycology. Hence, there is a need to train the next generation of mycologists who will promote the viability of this neglected field of biological sciences. My research on the four areas I have presented in this paper provides an overview on how several possibilities a person with training on fungi can do to safeguard food security and promote diversity and conservation for a sustainable and healthy planet.
Page 11 of 13
References 1. Aguilar-Hawod, K. 2014. Genetic diversity of Fusarium oxysporum f. sp. cubense (E.
F. Smith) Snyder & Hansen causing Panama wilt of banana (Musa spp.) in the Philippines. MS Thesis, UPLB, College, Laguna.
2. Australia State of the Environment Report. 2001 [http://www.conservation.org/documentaries/Pages/megadiversity.aspx Megadiversity: The 17 Biodiversity Superstars
3. Blackwell, M. 2011. The Fungi: 1, 2, 3 … 5.1 million species? American Journal of Botany. 98: 426-438.
4. Case, Anna E et al. 2013. DNA-based approaches to identify forest fungi in Pacific Islands: A pilot study. In: Browning, John; Palacios, Patsy, comps. Proceedings of the 60th Annual Western International Forest Disease Work Conference; October 8-12 October 2012; Tahoe City, CA. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Forest Health Technology and Enterprise Team. p. 149-151.
5. Collwell, R.R. 1997. Microbial diversity: the importance of exploration and conservation Journal of Industrial Microbiology and Biotechnology 18: 302-307
6. Crous, P.W., Braun, U., Wingfield, M.J., Wood, A.R., Shin, H.D., Summerell, B.A. Alfenas, A.C., Cumagun, C.J.R. and Groenewald, J.Z. 2009. Phylogeny and taxonomy of obscure genera of microfungi. Persoonia Molecular Phylogeny and Evolution 22:139-161.
7. Cumagun, C.J.R., Vargas, M. L.D.V., and Alviar, A.N. 2011. Evaluation of greenhouse and field aggressiveness of Fusarium verticillioides from corn in Laguna province, Philippines. African Journal of Agricultural Research 6: 6586-6591.
8. Cumagun, C. J. R.2012. Managing plant disease and promoting sustainability and productivity with Trichoderma: The Philippine Experience. Journal of Agricultural Science and Technology 14: 699-714.
9. Cumagun, C.J.R. and Ilag, L.L.1998a. Enhancing the efficacy of Trichoderma harzianum Rifai by chitin amendment against sheath blight of rice. Philippine Phytopathology 33: 72-86.
10. Cumagun, C.J.R. and Ilag, L.L.1998b. Parasitism of sclerotial bodies of Rhizoctonia solani Kuhn by Trichodema harzianum Rifai and Penicillium oxalicum Currie and Thom. Philippine Phytopathology 33: 17-26.
11. Cumagun, C.J.R. and Lapis, D.B. 1993. Note: Practical approach in mass production of Trichoderma spp. as a means of biological control against sheath blight of rice. Philippine Agriculturist 76: 251-257.
12. Cumagun, C.J.R., Hockenhull, J. and Lubeck, M.2000. Characterization of Trichoderma isolates from Philippines rice fields by UP-PCR and rDNA-ITS1 analysis: Identification of UP-PCR markers. Journal of Phytopathology 143: 109-115.
13. Cumagun, C.J.R. and Moosavchap. 2015. Significance of biocontrol agents of phytonematodes. In
14. Cumagun, C.J.R., Ramos, J.S., Dimaano, A.O., Munaut, F. and Van Hove, F. 2009. Genetic characteristics of Fusarium verticillioides from corn in the Philippines. Journal of General Plant Pathology 75: 405-412
15. Cumagun, C.J.R., Anh, V.L., Phuong, Vy, T.T., Asano, H., Chuma, I. and Tosa Y. 2014 Identification a hidden resistance gene in tetraploid wheat using laboratory strains of Pyricularia oryzae produced by backcrosses. Phytopathology 104:634-640.
Page 12 of 13
16. Cumagun, C.J.R. Zala, M., Oliva, R., Arakawa, M., McDonald, B.A., and Ceresini, P.C. 2015. Genetic structure of populations of Rhizoctonia solani AG1-IA from rice in China, Japan and the Philippines. unpublished results.
17. Doyle, M., Jaykus, L. and Metz, M. 2005. Research Opportunities in Food and Agriculture Microbiology. American Academy of Microbiology. March 12-14, 2005, Washington, DC.
18. Graff, P.W. 1916. Bibliography and new species of Philippine fungi. Mycologia 8: 253-288.
19. Hawksworth, D.L.1991. The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycological Research 95: 641-655.
20. Hawksworth, D.L. 2001. The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycological Research 105:1422-1432.
21. Hawksworth, D.L. 2009. Mycology as a neglected megascience. In : Rai, M. and Bridge, P.D. (eds) Applied Mycology. CAB International, London,UK. pp1-16
22. Hyde, K.D., Zhou, D., McKenzie, E.H.C., Ho, W.H. and Dalisay, T. Vertical distribution of saprobic fungi on bamboo culms. Fungal Diversity 11:109-118.
23. Ingram, D.S. 1998. Biodiversity and Plant Pathogens and Conservation. http://www.apsnet.org/publications/apsnetfeatures/Pages/ICPP98Ingram.aspx
24. Lopez, A.L.C., Adreit, H., Milazzo, J., Cumagun, C.J.R., and Thareau, D. 2015. Genetic structure of populations of Magnaporthe oryzae in farmer fields and experimental plots in the Philippines. unpublished results.
25. Kato, H. Yamamoto, M., Yamaguchi-ozaki, T., Kadouchi, H. , Iwamoto, Y., Nakayashiki, H., Tosa, Y., Mayama, S. and Mori, N. 2000. Pathogenicity, mating ability and DNA restriction fragment length polymorphisms of Pyricularia populations isolated from Gramineae, Bambusideae and Zingiberaceae plants. Journal of General Plant Pathology 66: 30-47.
26. Macías-Rubalcava ML, Hernández-Bautista BE, Jiménez-Estrada M, González MC, Glenn AE, Hanlin RT, Hernández-Ortega S, Saucedo-García A, Muria-González JM, Anaya AL. 2008. Naphthoquinone spiroketal with allelochemical activity from the newly discovered endophytic fungus Edenia gomezpompae. Phytochemistry 69:1185-1196.
27. Magculia, N, Cumagun, C.J.R. 2011. Genetic diversity and PCR based identification of fumonisin producing isolates of Fusarium verticillioides in corn. Tropical Plant Pathology 36: 225-232.
28. Mahamuda Begum, M.M., Dalisay, T.U. and Cumagun, C.JR. 2012. Taxonomic review and development of a Lucid key for Philippime Cercosporoids and related fungi. In Cumagun, C.J.R. (ed) Plant Pathology, In Tech Press, Croatia. pp.1-40.
29. Martinez-Luis, S. Cherigo, L., Higginbotham, S., Arnold, E., Spadafora, C., Ibañez, Gerwick, W.H. and Cubilla-Rios, L. 2011. Screening and evaluation of antiparasitic and in vitro anticancer activities of Panamanian endophytic fungi. International Microbiology 14;95-102.
30. Minter, D. W. 2011. What every botanist and zoologist should know –and what every mycologist should be telling them. IMA Fungus 2:1-5.
31. Mitchell, A.M. et al. 2008. Muscodor crispans, a novel endophyte from Ananas ananassoides in the Bolivian Amazon Fungal Divers 31:37–43.
Page 13 of 13
32. Mitchell, A.M., Strobel, G.A., Hess, W.M., Vargas, P.N. and Ezra, D. 2008. Muscodor crispans, a novel endophyte from Ananas ananassoides in the Bolivian Amazon. Fungal Diversity 31: 37-43.
33. Molina, A. B. et al. 2011. Field resistance of selected banana cultivars against Tropical Race 4 of Fusarium oxysporum f. sp. cubense in the Philippines Phytopathology 101:S122.
34. Moore, D., Nauta, M.M., Evans, S.E. and Rotheroe, M. 2008. Fungal Conservation: Issues and Solutions. Cambridge University Press. 276 pp.
35. Oclarit, E.L. and Cumagun, C.J.R. 2009. Evaluation of efficacy of Paecilomyces lilacinus as biological control agent of Meloidogyne incognita attacking tomato. Journal of Plant Protection Research 49:459-462
36. Solpot, T.C. 2015. Phenotypic analysis, molecular detection and management of Fusarium oxysporum f sp. cubense Tropical Race 4 (FocTR4) causing Fusarium wilt of banana in South Central Mindanao. MS Thesis, UPLB, College, Laguna.
37. Strobel, G. A. Microbial gifts from rainforests. Can. J. Plant Pathol. 24:14-20.
38. Su G.S. et al. 2014. Distribution and Diversity of Marine Fungi in Manila Bay, Philippines. Annual Research and Review in Biology 4: 4166-4173.
39. Tadiosa, E.R. 2012. The growth and development of mycology in the Philippines. Fungal Conservation 2: 18-22.
40. Tadiosa, E.R, Agbayani, E.S. & Agustin, N.T. 2011. Preliminary study on the macrofungi of Bazal-Baubo Watersheds, Aurora Province, Central Luzon, Philippines. Asian Journal of Biodiversity 2: 149-171.
41. Tadiosa, E.R, Arsenio, J.J. & Marasigan, M.C. 2007. Macroscopic fungal diversity of Mt. Makulot, Cuenca, Batangas, Philippines. Journal of Nature Studies 6: nos 1& 2.
42. Tadiosa, E.R. and Briones, R.U. 2013. Fungi of Taal volcano protected landscape, Southern Luzon, Philippines. Asian Journal of Biodiversity 4: 46-64.
43. Tadiosa, E.R. & Militante, E.P. 2006. Identification of important wood-decaying fungi associated with some Philippine dipterocarps at the Makiling Forest. Sylvatrop 16: nos 1 & 2.
44. Takabayashi, N., Tosa, Y., Oh, H. S., and Mayama, S. 2002. A gene-for-gene relationship underlying the species-specific parasitism of Avena/Triticum isolates of Magnaporthe grisea on wheat cultivars. Phytopathology 92:1182-1188.
45. Teodoro, N.G. 1937. An Enumeration of Philippine Fungi. Commonwealth of the Philippines, Department of Agriculture and Commerce, Technical Bulletin 4. Bureau of Printing, Manila.
46. Torada A, Koike M, Mochida K, Ogihara Y. 2006. SSR-based linkage map with new markers using an intraspecific population of common wheat. Theor Appl Genet. 112:1042–1051.
47. Villacarlos, L.T. and Meija, B.S. 2004. Philippine entomopathogenic fungi I. Occurrence and diversity. Philippine Agricultural Scientist 87: 249-265.