Research article Lipopolysaccharides elicit an oxidative burst as a component of the innate immune system in the seagrass Thalassia testudinum Kyle Loucks, David Waddell, Cliff Ross * Department of Biology, University of North Florida, 1 UNF Drive, Jacksonville, FL 32224, USA article info Article history: Received 17 April 2013 Accepted 20 May 2013 Available online 5 June 2013 Keywords: Elicitation Hydrogen peroxide Oxidative burst Pathogen defense Seagrass Signaling pathways abstract This study represents the first report characterizing the biological effects of a lipopolysaccharide (LPS) immune modulator on a marine vascular plant. LPS was shown to serve as a strong elicitor of the early defense response in the subtropical seagrass Thalassia testudinum Banks ex König and was capable of inducing an oxidative burst identified at the single cell level. The formation of reactive oxygen species (ROS), detected by a redox-sensitive fluorescent probe and luminol-based chemiluminescence, included a diphenyleneiodonium sensitive response, suggesting the involvement of an NADPH oxidase. A 900 bp cDNA fragment coding for this enzyme was sequenced and found to encode a NAD binding pocket domain with extensive homology to the Arabidopsis thaliana rbohF (respiratory burst oxidase homolog) gene. The triggered release of ROS occurred at 20 min post-elicitation and was dose-dependent, requiring a minimal threshold of 50 mg/mL LPS. Pharmacological dissection of the early events preceding ROS emission indicated that the signal transduction chain of events involved extracellular alkalinization, G-proteins, phospholipase A 2 , as well as K þ , Ca 2þ , and anion channels. Despite exclusively thriving in a marine environment, seagrasses contain ROS-generating machinery and signal transduction components that appear to be evolutionarily conserved with the well-characterized defense response systems found in terrestrial plants. Ó 2013 Elsevier Masson SAS. All rights reserved. 1. Introduction Seagrasses are widely distributed marine vascular plants that not only serve as critical sources of coastal primary production and habitat, but also contribute significantly toward biogeochemical cycling and near shore sediment stabilization [1]. Seagrass meadows have experienced significant declines in recent decades due to persistent threats from both local and global scale sources. One such threat is the incidence of disease in which pathogenic protists of the genus Labyrinthula spp. (Division Stramenopila) cause local and regional-scale seagrass die-off events; a phenom- enology termed wasting disease [2]. Wasting disease outbreaks have had long-lasting effects on dominant seagrass species (e.g. Thalassia testudinum Banks ex König, which is found in tropics and subtropics and Zostera marina Linnaeus, which is found in temperate regions) as some populations affected by the disease have not fully recovered, and other populations have been driven to local extinction [3]. While there is a recent increasing awareness of seagrass wasting disease in the marine environment [4,5] the biochemical basis of pathogen recognition and defense responses in seagrasses are virtually unexplored. Considering seagrasses are descendants of terrestrial plants that evolved into a distinct lineage w75 million years ago [6], it would be anticipated that there would be a selected degree of homology in the principles of activated innate immunity. A substantial amount of evidence has accumulated highlighting the importance of pathogen-associated molecular patterns (PAMPs) in plant defense systems [7]. PAMPs are expressed by a wide range of microorganisms and include well-characterized elicitors such as chitin, eubacterial flagellin, peptidoglycans, lip- oteichoic acid (LTA) of Gram-positive bacteria and the lipopoly- saccharides (LPS) of Gram-negative bacteria, which often serve in many experimental systems as the prototypical model PAMP [8]. LPS is a structurally conserved lipoglycan capable of inducing both systemic acquired resistance (SAR) and induced systemic resistance (ISR) in terrestrial plant studies [9]. In addition, LPS has been implicated in the elicitation of immune responses modulating extracellular alkalinization (K þ /H þ exchange), nitric oxide (NO) Abbreviations: DPI, diphenyleneiodonium; H 2 DCF-DA, 2 0 ,7 0 -dichlorodihydro- fluoresceindiacetate; H 2 O 2 , hydrogen peroxide; LPS, lipopolysaccharide; NADPH, nicotinamide adenine dinucleotide phosphate; PAMP, pathogen-associated molec- ular pattern; ROS, reactive oxygen species; RBOH, respiratory burst oxidase ho- molog; SAR, systemic acquired resistance; SSA, serum-seawater agar. * Corresponding author. Tel.: þ1 904 620 2830; fax: þ1 904 620 3885. E-mail address: [email protected](C. Ross). Contents lists available at SciVerse ScienceDirect Plant Physiology and Biochemistry journal homepage: www.elsevier.com/locate/plaphy 0981-9428/$ e see front matter Ó 2013 Elsevier Masson SAS. All rights reserved. http://dx.doi.org/10.1016/j.plaphy.2013.05.023 Plant Physiology and Biochemistry 70 (2013) 295e303
Transcript
at SciVerse ScienceDirect
Plant Physiology and Biochemistry 70 (2013) 295e303
Contents lists available
Plant Physiology and Biochemistry
journal homepage: www.elsevier .com/locate/plaphy
Research article
Lipopolysaccharides elicit an oxidative burst as a component of theinnate immune system in the seagrass Thalassia testudinum
Kyle Loucks, David Waddell, Cliff Ross*
Department of Biology, University of North Florida, 1 UNF Drive, Jacksonville, FL 32224, USA
a r t i c l e i n f o
Article history:Received 17 April 2013Accepted 20 May 2013Available online 5 June 2013
0981-9428/$ e see front matter � 2013 Elsevier Mashttp://dx.doi.org/10.1016/j.plaphy.2013.05.023
a b s t r a c t
This study represents the first report characterizing the biological effects of a lipopolysaccharide (LPS)immune modulator on a marine vascular plant. LPS was shown to serve as a strong elicitor of the earlydefense response in the subtropical seagrass Thalassia testudinum Banks ex König and was capable ofinducing an oxidative burst identified at the single cell level. The formation of reactive oxygen species(ROS), detected by a redox-sensitive fluorescent probe and luminol-based chemiluminescence, includeda diphenyleneiodonium sensitive response, suggesting the involvement of an NADPH oxidase. A 900 bpcDNA fragment coding for this enzyme was sequenced and found to encode a NAD binding pocketdomain with extensive homology to the Arabidopsis thaliana rbohF (respiratory burst oxidase homolog)gene. The triggered release of ROS occurred at 20 min post-elicitation and was dose-dependent, requiringa minimal threshold of 50 mg/mL LPS. Pharmacological dissection of the early events preceding ROSemission indicated that the signal transduction chain of events involved extracellular alkalinization,G-proteins, phospholipase A2, as well as Kþ, Ca2þ, and anion channels. Despite exclusively thriving in amarine environment, seagrasses contain ROS-generating machinery and signal transduction componentsthat appear to be evolutionarily conserved with the well-characterized defense response systems foundin terrestrial plants.
� 2013 Elsevier Masson SAS. All rights reserved.
1. Introduction
Seagrasses are widely distributed marine vascular plants thatnot only serve as critical sources of coastal primary production andhabitat, but also contribute significantly toward biogeochemicalcycling and near shore sediment stabilization [1]. Seagrassmeadows have experienced significant declines in recent decadesdue to persistent threats from both local and global scale sources.One such threat is the incidence of disease in which pathogenicprotists of the genus Labyrinthula spp. (Division Stramenopila)cause local and regional-scale seagrass die-off events; a phenom-enology termed wasting disease [2]. Wasting disease outbreakshave had long-lasting effects on dominant seagrass species(e.g. Thalassia testudinum Banks ex König, which is found in tropicsand subtropics and Zostera marina Linnaeus, which is found in
temperate regions) as some populations affected by the diseasehave not fully recovered, and other populations have been driven tolocal extinction [3]. While there is a recent increasing awareness ofseagrass wasting disease in the marine environment [4,5] thebiochemical basis of pathogen recognition and defense responsesin seagrasses are virtually unexplored. Considering seagrasses aredescendants of terrestrial plants that evolved into a distinct lineagew75 million years ago [6], it would be anticipated that there wouldbe a selected degree of homology in the principles of activatedinnate immunity.
A substantial amount of evidence has accumulated highlightingthe importance of pathogen-associated molecular patterns(PAMPs) in plant defense systems [7]. PAMPs are expressed by awide range of microorganisms and include well-characterizedelicitors such as chitin, eubacterial flagellin, peptidoglycans, lip-oteichoic acid (LTA) of Gram-positive bacteria and the lipopoly-saccharides (LPS) of Gram-negative bacteria, which often serve inmany experimental systems as the prototypical model PAMP [8].LPS is a structurally conserved lipoglycan capable of inducing bothsystemic acquired resistance (SAR) and induced systemic resistance(ISR) in terrestrial plant studies [9]. In addition, LPS has beenimplicated in the elicitation of immune responses modulatingextracellular alkalinization (Kþ/Hþ exchange), nitric oxide (NO)
Fig. 1. (A) Kinetics of ROS release in response to LPS elicitation in T. testudinum. Datapoints are the mean of 5 replicates (�1 SEM). (B) H2O2 produced at the oxidative burstmaxima (20 min time point of Fig. 1A) in response to selected concentrations of LPS.Data points are the mean of five replicates (�1 SEM). Letters above bars indicatesignificantly different groups (p ¼ 0.000).
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303296
production, calcium influx, the induction of pathogenesis related(PR) gene expression and cell-wall alterations that include thedeposition of callose and phenolics [10].
The production of reactive oxygen species (ROS) in response toLPS elicitation is another early innate immune response that hasbeen well documented chiefly in plant cell suspension cultures[11,12]. Since the discovery of the elicited production of superoxide(O2
�) in potato in response to an avirulent isolate of Phytophthorainfestans [13], superoxide and other ROS such as hydrogen peroxide(H2O2), singlet oxygen (1O2) and hydroxyl radical (OH$) have beenimplicated as key elements in eukaryotic defense [14]. While theenzymatic sources of ROS may include polyamine, oxalate or otherapoplastic oxidases [15,16], plasma membrane-bound NADPH oxi-dases (EC 1.6.3.1.; Rbohs, for respiratory burst oxidase homologs)and cell-wall peroxidases are considered to be the main sources interms of plant innate immunity [17e19].
In contrast to the growing body of terrestrial plant literaturedescribing the onset of oxidative bursts and the preceding coordi-nated signal transduction events, there is only limited knowledgeon the biochemical-based induced defense responses of aquaticphotoautotrophs. Furthermore, the available data is restricted toalgal taxa, as there have been no reports on aquatic vascular plants.An oxidative burst was described in the brown algal kelp Laminariadigitata and was shown to play a critical role in the regulation ofepiphytic, putatively pathogenic, bacteria [20,21]. In addition, thesame algae could recognize host-derived oligoguluronate frag-ments as well as exogenous prostaglandins, both capable of trig-gering the sudden release of ROS [20,22]. Weinberger et al. [23,24]demonstrated that the Rhodophyte (red algae) Gracilaria confertacould respond to microbial degradation of its agar cell-wall matrixwith a DPI-sensitive oxidative burst, suggesting the involvement ofan NADPH oxidase. Despite these efforts demonstrating the capa-bility for marine algae to respond to both endogenous and exoge-nous cues, there has only been one report highlighting theinvolvement of an LPS-induced oxidative burst in a marine mac-roalgae. This was observed in the Phaeophyceae (brown algae)member L. digitata [25]. The lack of evidence characterizing theeffects of exogenous elicitors, such as LPS, on the innate immunityof marine photoautotrophs is probably not due to the lack ofbiochemical machinery in these taxa, but more so due to the lack ofinvestigations on marine species overall [26]. In order to explorethe early defense responses inmarine vascular photoautotrophs weused the tropical/subtropical seagrass T. testudinum as a represen-tative model species. In light of the historical die-off events asso-ciated with wasting disease, we undertook this study to betterunderstand the cellular factors that regulate seagrass innate im-munity. The objectives of this work were to determine: (1) ifT. testudinum has the capability of responding to exogenous elici-tation, using purified LPS, and engage in a signaling cascade thatresults in an oxidative burst; and (2) if T. testudinum has the abilityto recognize biochemical extracts of Labyrinthula sp. to offer insightinto this important yet understudied hostepathogen interactionsystem.
2. Results
2.1. Induction of an oxidative burst in T. testudinum
The addition of LPS-induced an oxidative burst in T. testudinumwith a maximal release of H2O2 observed at 20 min post-elicitation(Fig. 1A). The magnitude of the H2O2 burst varied significantly as afunction of LPS concentration (Fig. 1B, one-way ANOVA, F ¼ 14.891,p ¼ 0.000). The addition of 25 mg/mL LPS caused a 16% increase inthe ROS produced when compared to non-elicited controls yet thiseffect was not statistically significant. The minimum threshold of
LPS required to elicit a response was 50 mg/mL (Fig. 1B). The addi-tion of two elevated concentrations of LPS (100 and 200 mg/mL)resulted in a 56 and 86 percent increase, respectively, in the H2O2released into the surrounding media when compared to controls.Hydrogen peroxide levels did not reach saturation in the range ofconcentrations tested. Extracts of Labyrinthula sp. failed to induceany oxidative burst in T. testudinum, using concentrations up to5 mg/mL (data not shown).
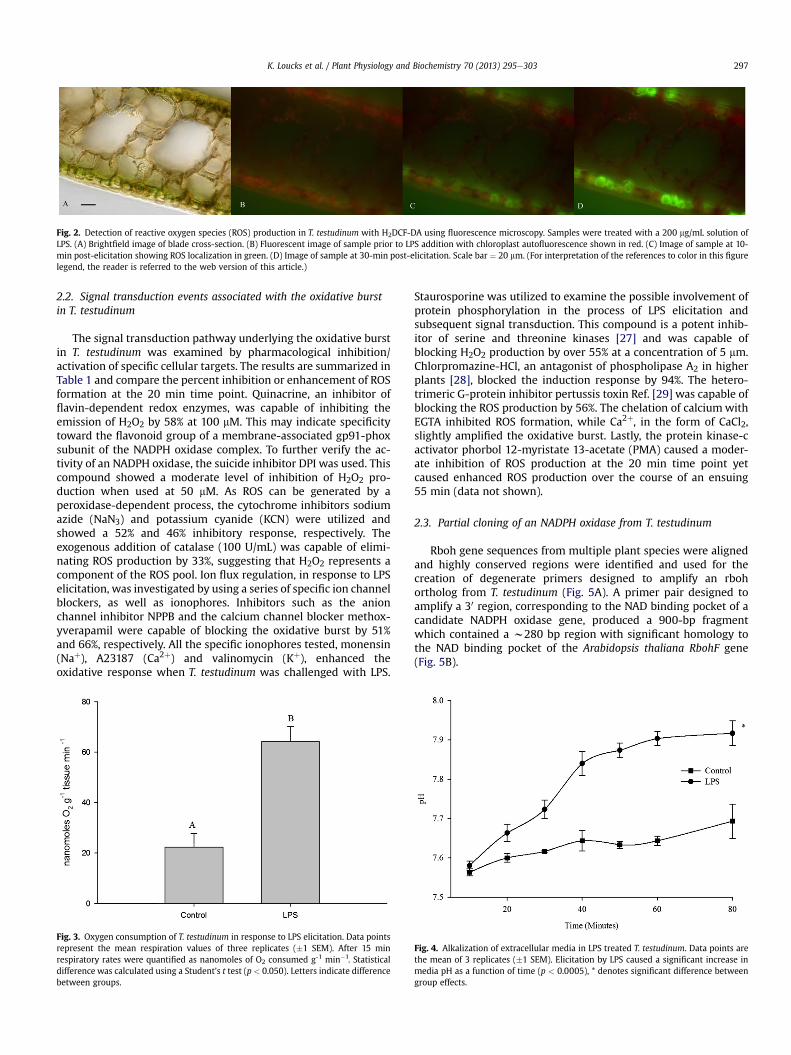
Sections of T. testudinum were challenged with LPS and moni-tored for ROS release using fluorescence microscopy in conjunctionwith the redox-sensitive probe H2DCF-DA. The production of ROScould be localized to epidermal cells and was not detected inmesophyll cells bordering the air lacunae (Fig. 2). The ROS signalcould be detected as early as 10 min post-elicitation and continuedto intensify up through 30 min.
Following LPS addition, specimens of T. testudinum underwent asignificant increase in oxygen consumption compared to untreatedcontrols (Fig. 3, Student’s t test, p < 0.050). While a distinct respi-ratory burst was not evident, there was a steady increase in oxygenconsumption as a function of time. By the end of the experimentchallenged plants had an average oxygen consumption rate of 64.2(�6.0) nanomoles of O2 g�1 min�1 while control plants had valuesroughly three times lower (22.4 [�5.4]). Extracellular pH alsoshowed a steady increase following LPS addition and was signifi-cantly different than the controls (Fig. 4, repeated measuresANOVA, F ¼ 155.046, p � 0.005). The seawater underwent analkalinization by approximately 0.3 pH units over the course of80 min. There was no significant interaction between time and LPStreatment (Wilks’ Lambda: F ¼ 88.695, p ¼ 0.079).
Fig. 2. Detection of reactive oxygen species (ROS) production in T. testudinum with H2DCF-DA using fluorescence microscopy. Samples were treated with a 200 mg/mL solution ofLPS. (A) Brightfield image of blade cross-section. (B) Fluorescent image of sample prior to LPS addition with chloroplast autofluorescence shown in red. (C) Image of sample at 10-min post-elicitation showing ROS localization in green. (D) Image of sample at 30-min post-elicitation. Scale bar ¼ 20 mm. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303 297
2.2. Signal transduction events associated with the oxidative burstin T. testudinum
The signal transduction pathway underlying the oxidative burstin T. testudinum was examined by pharmacological inhibition/activation of specific cellular targets. The results are summarized inTable 1 and compare the percent inhibition or enhancement of ROSformation at the 20 min time point. Quinacrine, an inhibitor offlavin-dependent redox enzymes, was capable of inhibiting theemission of H2O2 by 58% at 100 mM. This may indicate specificitytoward the flavonoid group of a membrane-associated gp91-phoxsubunit of the NADPH oxidase complex. To further verify the ac-tivity of an NADPH oxidase, the suicide inhibitor DPI was used. Thiscompound showed a moderate level of inhibition of H2O2 pro-duction when used at 50 mM. As ROS can be generated by aperoxidase-dependent process, the cytochrome inhibitors sodiumazide (NaN3) and potassium cyanide (KCN) were utilized andshowed a 52% and 46% inhibitory response, respectively. Theexogenous addition of catalase (100 U/mL) was capable of elimi-nating ROS production by 33%, suggesting that H2O2 represents acomponent of the ROS pool. Ion flux regulation, in response to LPSelicitation, was investigated by using a series of specific ion channelblockers, as well as ionophores. Inhibitors such as the anionchannel inhibitor NPPB and the calcium channel blocker methox-yverapamil were capable of blocking the oxidative burst by 51%and 66%, respectively. All the specific ionophores tested, monensin(Naþ), A23187 (Ca2þ) and valinomycin (Kþ), enhanced theoxidative response when T. testudinum was challenged with LPS.
Fig. 3. Oxygen consumption of T. testudinum in response to LPS elicitation. Data pointsrepresent the mean respiration values of three replicates (�1 SEM). After 15 minrespiratory rates were quantified as nanomoles of O2 consumed g-1 min�1. Statisticaldifference was calculated using a Student’s t test (p < 0.050). Letters indicate differencebetween groups.
Staurosporine was utilized to examine the possible involvement ofprotein phosphorylation in the process of LPS elicitation andsubsequent signal transduction. This compound is a potent inhib-itor of serine and threonine kinases [27] and was capable ofblocking H2O2 production by over 55% at a concentration of 5 mm.Chlorpromazine-HCl, an antagonist of phospholipase A2 in higherplants [28], blocked the induction response by 94%. The hetero-trimeric G-protein inhibitor pertussis toxin Ref. [29] was capable ofblocking the ROS production by 56%. The chelation of calciumwithEGTA inhibited ROS formation, while Ca2þ, in the form of CaCl2,slightly amplified the oxidative burst. Lastly, the protein kinase-cactivator phorbol 12-myristate 13-acetate (PMA) caused a moder-ate inhibition of ROS production at the 20 min time point yetcaused enhanced ROS production over the course of an ensuing55 min (data not shown).
2.3. Partial cloning of an NADPH oxidase from T. testudinum
Rboh gene sequences from multiple plant species were alignedand highly conserved regions were identified and used for thecreation of degenerate primers designed to amplify an rbohortholog from T. testudinum (Fig. 5A). A primer pair designed toamplify a 30 region, corresponding to the NAD binding pocket of acandidate NADPH oxidase gene, produced a 900-bp fragmentwhich contained a w280 bp region with significant homology tothe NAD binding pocket of the Arabidopsis thaliana RbohF gene(Fig. 5B).
Fig. 4. Alkalization of extracellular media in LPS treated T. testudinum. Data points arethe mean of 3 replicates (�1 SEM). Elicitation by LPS caused a significant increase inmedia pH as a function of time (p < 0.0005), * denotes significant difference betweengroup effects.
Table 1Effect of modulators on the oxidative burst in Thalassia testudinum. Data shown are the average value obtained from five independent experiments (�1 SE). Details of theexperiment are provided in Section 4.
Modulator Concentration H2O2 formed (% control) Mechanism of action
ElicitorLipopolysaccharide (E. coli) 200 mg/mL 100 N/A
InhibitorsQuinacrine 100 mM 42 (�5.7) Flavoprotein inhibitorDPI 50 mM 67 (�7.6) NADPH oxidase inhibitorKCN 1 mM 54 (�3.1) Peroxidase inhibitorNaN3 1 mM 48 (�11.1) Peroxidase inhibitorCatalase 100 U/mL 67 (�5.8) H2O2 decompositionNPPB 100 mM 49 (�3.8) Anion channel blockerStaurosporine 5 mM 56 (�2.4) Protein kinase inhibitorChlorpromazine-HCl 100 mM 6 (�1.3) Phospholipase A2 inhibitorMethoxyverapamil 200 mM 34 (�13) Ca2þ channel blockerPertussis toxin 1 mg/mL 44 (�6.7) G-protein inhibitorEGTA 1 mM 64 (þ3.7) Calcium chelator
IonophoresMonensin 100 mM 104 (�1.9) Naþ ionophoreA23187 100 mM 145 (�7.9) Ca2þ ionophoreValinomycin 100 mM 155 (�8.2) Kþ ionophore
ActivatorsPMA 15 mM 59 (�15) Protein kinase activatorCaCl2 2 mM 102 (�5.4) Calcium supplement
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303298
3. Discussion
3.1. Induction of an oxidative burst in T. testudinum
Despite the principal roles of ROS in terrestrial plant physiology,reports of its occurrence in marine vascular plants, especially in thecontext of disease resistance, are extremely limited. To ourknowledge, this study represents the first finding of an LPS-inducedoxidative burst in a seagrass and more broadly demonstrates thatpathogen perception in aquatic angiosperms parallels the innateimmune systems found in terrestrial plants and lower algae.
The concentration of LPS required to elicit an oxidative burst inT. testudinumwas 50 mg/mL, which is comparable to results found inother plant studies where the LPS threshold ranged from 5 to100 mg/mL [10]. While plants can detect other prokaryotic PAMPssuch as flagellin and Ef-Tu elongation factor at subnanomolar levels,it has been suggested that plants only possess low-affinity mem-brane-associated receptors to detect LPS [10,30]. The time frame ofmaximal H2O2 emission was similar to results obtained in tobaccocell suspensions when challenged with LPS from the phytopathicbacteria Xanthomonas campestris. In both studies, a delayedoxidative burst occurredwhich could be attributed to the decreasedavailability of LPS in aqueous solutions, as LPS can form micellarvesicles [31]. Alternatively, the lag phase could have been due to thebiochemical conversion of metabolites or induction of enzymesupstream of hydrogen peroxide production as suggested by Küpperet al. [25].
Attempts to detect an oxidative burst in T. testudinum inresponse to extracts of Labyrinthula sp. were unsuccessful. Oneexplanation for the lack of ROS production could be due toT. testudinum’s inability to recognize Labyrinthula-based PAMPs.Lipopolysaccharides are not cell-wall constituents in members ofthe Labyrinthulomycetes. The cell walls of Labyrinthula sp. arecharacterized by the presence of dictyosome-derived carbohydratescales [32], of which the precise composition is not known. Evi-dence for the presence of sulfated polysaccharides (predominantlygalactose or fucose-based) and proteins in the cell walls of closelyrelated taxa, such as thraustochytrids has been reported [32,33];thus, it seems plausible that similar constituents may be foundin Labyrinthula sp. External to the cell wall, Labyrinthula sp. possessan ectoplasmic network (EN) that has been characterized as
plasma-membrane-bound cytoplasm rich in actin and myosin Ref.[34]. It is possible that T. testudinum does not have the capability torecognize any of these cell wall or EN-based components whenpresented in the form of an aqueous crude extract. However, itshould be noted that the use of crude extracts has been successfulin eliciting defense responses in A. thaliana (at much lower con-centrations used in our study) with fungal-derived elicitors(200 mg/mL; [35]).
Our O2 consumption data for T. testudinum indicated that therewas a notable uptake of O2 from the ambient seawater. This uptakewould clearly provide an adequate level of oxygen to use as asubstrate for the enzymatically based turnover of O2 to H2O2. Wedid not detect an immediate respiratory burst as has been charac-terized in other plant systems [14]. However, there was a steady-state increase in O2 consumption as a function of time followingLPS elicitation. A similar response has been detected in potato usingheat killed Pseudomonas syringae as the elicitor [36].
Extracellular alkalinization is one of the earliest events occur-ring in elicitor-treated plant cell suspension cultures. This alkalin-ization results from elicitor-induced depolarization of the plasmamembrane and subsequent Kþ/Hþ exchange, with Ca2þ influx/Cl�
efflux [37]. There is typically a simultaneous acidification of thecytoplasm that is believed to be an essential component of thesignal transduction pathway leading to an oxidative burst [38]. Theintensity of the pH shift in plants is clearly dependent upon thetype of host cells used and the structure of the elicitor provided[39]. The ability for T. testudinum to undergo extracellular alkalin-ization in response to LPS elicitation suggests this phenomenologyis conserved in marine vascular plants.
3.2. Elicitor signal transduction
Results obtained from our pharmacological studies using DPIsupport the premise that the oxidative burst in T. testudinum mayinvolve the activity of a flavoprotein-containing NADPH oxidasecomplex that is analogous to what is involved in ROS production interrestrial plants [40]. The ability for quinacrine to inhibit the burstprovides additional evidence for the possible involvement of aflavonoid component of a membrane-associated gp91-phox sub-unit. As NaN3 and KCN could partially inhibit the production ofH2O2, this prompted speculation as to exactly where ROS were
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303 299
being produced. Both azide and cyanide can block heme-basedperoxidases, yet NADPH oxidases, which fall under the flavopro-tein family, are typically insensitive to these inhibitors [39]. Itshould be noted that DPI has been reported to serve as an effectiveinhibitor of heme-based peroxidases, suggesting that it may not bevery selective when distinguishing between NADPH oxidases andperoxidases [18]. However, it is possible that both enzymaticsources are contributing toward the ROS pool or functioningsequentially as NADPH oxidases could be activated by ROS origi-nating from cell-wall bound peroxidases [39,41].
Research identifying the signaling pathway components inaquatic vascular plants is most certainly lagging behind the ever-expanding state of knowledge found in the terrestrial plant litera-ture. To add to the complexity, the activation of different signalingsystems may vary based upon the origin of the sensor and on thenature of the stress event. In both the NADPH oxidase and apo-plastic peroxidase ROS-generating models, an elicitor molecule isdetected by a receptor located on the plasma membrane. In turn, asignal transduction cascade is initiated that activates downstreammolecular responses. Data available from terrestrial plants supportthe involvement of G-proteins, ion channels (especially Ca 2þ),protein kinases, protein phosphatases, phospholipases A and C, andcyclic AMP. This pathway ultimately gives rise to the activation ofeither an NADPH oxidase complex or a pH-dependent cell-wallperoxidase [14]. Our data show that a variety of identifiable signaltransduction steps (homologous to what has been reported interrestrial plants) are involved in ROS production in T. testudinumwhen elicited with LPS.
The phosphorylating activities of kinases are considered to bean integral component of numerous metabolic regulatory mecha-nisms. Staurosporine was reported to block protein phosphoryla-tion events involved in induction of defense responses incryptogein-elicited tobacco cells [42] and in fungal-elicitedspruce cells [43]. In our study, staurosporine proved to be aneffective inhibitor in T. testudinum, suggesting that serine andthreonine kinases are involved in an upstream stage of signaltransduction. The involvement of protein kinase-C (PKC) in respi-ratory burst signaling cascades, by use of the activator phorbol 12-myristate 13-acetate (PMA), has been demonstrated in fishphagocytes [44] and in chlorophytic algae [45]. In both cases, PMAcaused a dose-dependent activation of the oxidative burst.Conversely, in T. testudinum, there was a moderate inhibition ofROS produced at the 20 min time point. PMA has been previouslyshown to have an inhibitory effect on Naþ/Hþ exchange in carci-noma cells [46] as well as on the phosphorylation-dependent in-hibition of anaplastic thyroid cancer cell line proliferation [47].Chlorpromazine-HCl, a selective inhibitor of phospholipase A2,caused a strong inhibition of the oxidative burst in T. testudinum.The production of ROS, involving the activation of phospholipases,has also been demonstrated in soybean [48,49] and in brownalgae [20].
Bacterial toxins, such as pertussis toxin, have proven to be usefulprobes for studying the roles of heterotrimeric G-proteins in signaltransduction pathways [50]. The participation of G-proteins wasanalyzed using cholera toxin in French beans [51] and was alsoshown to be involved in the signal transduction cascade associatedwith the fungal-elicited synthesis of alkaloids in the bloodrootflower Sanguinaria canadensis [52]. Pertussis toxin proved to inhibitROS production in T. testudinum indicating that G-proteins areinvolved in this elicitor-triggered oxidative burst.
The regulation of ion flow, mainly Ca2þ and Hþ influx and Kþ
and Cl- efflux, is one of the most rapid responses of plant cells toelicitation [53]. The multifunctional roles of ion fluxes in plantsystems prompted our investigation into the utilization of ionchannels as part of the signal transduction process in T. testudinum.
The use of the Kþ ionophore valinomycin in conjunction with LPScaused a very large enhancement of ROS production. A similarobservation was reported by Küpper et al. [20] using oligoguluro-nate elicitation in the brown algae L. digitata. Conversely, Bolwellet al. [17] noted that the pretreatment of valinomycin in Frenchbean cells triggered ion leakage which subsequently caused anegative impact on the efficiency of pH-dependent cell-wallperoxidase(s).
Calcium flux or local fluctuations of internal calcium concen-trations are among one of the earliest signaling events associatedwith the oxidative burst in terrestrial plants [49]. In addition,positive feedback loops have been identified where the binding ofCa2þ promotes ROS formation via NADPH oxidases. This subse-quently causes calcium channels to become activated, resulting in aself-amplifying signal [54]. The involvement of elevated cytosolicCa2þ levels also appears to be integrated into the signaling cascadeof T. testudinum. The combined effects of EGTA chelation, CaCl2supplementation, and the use of methoxyverapamil and ionophoreA23187 all strongly implicate the role of intracellular Ca2þ in ROSformation. Similar effects have been noted in French bean [55],Arabidopsis [39], spruce [43] in addition to brown and green algae[20,28]. This suggests that the ROS-Ca2þ relationship represents amore widely used signal codependence that spans the majority ofphotoautotrophic taxa.
3.3. Molecular characterization of an NADPH oxidase fromT. testudinum
Plant Rbohs are a multigenic family of enzymes with a C-ter-minal region containing cytosolic FAD- and NADPH-binding do-mains, responsible for oxidase activity, in addition to sixconserved transmembrane helices and an N-terminal regulatorydomain Ref. [19]. Rboh homologs are unique among ROS-producing enzymes in that they integrate different signal trans-duction pathways such as calcium, protein phosphorylation andlipid signaling with ROS production. The partial cloning of a genecontaining a NAD binding pocket, in conjunction with the signaltransduction and pharmacological data presented above, suggestthat seagrass may possess at least one functional respiratory burstoxidase enzyme gene and this would represent the first time thisclass of enzyme has been identified in a seagrass species. Whileour evidence suggests that it is possible that NADPH oxidases andperoxidases could both be serving as enzymatic sources of ROS inT. testudinum, it is clear that seagrasses have the capability ofresponding to elicitors derived from Gram-negative bacteria.Furthermore, the response components appear to be evolution-arily conserved with the well-characterized defense responsesystems found in terrestrial plants.
4. Materials and methods
4.1. Seagrass collection and maintenance
T. testudinum shoots were collected from the Indian RiverLagoon in Ft. Pierce, Florida (27�47’N, 80�31’W and 27�58’N,80�31’W) and cleaned of epiphytes. Short shoots included theblades, sheath and a 2-inch section of rhizome. Following collec-tion, all specimens were allowed to acclimate for no less than 1week under greenhouse conditions at the University of NorthFlorida, Jacksonville, FL prior to use in any experiment. Plants weremaintained in aquaria at a salinity of 30 under shade cloth allowinga maximal penetration of 15% photosynthetic active radiation(<300 mmol m�2 s�1). Diel temperature values ranged between 25and 27 �C.
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303300
Fig. 5. (continued).
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303 301
4.2. Labyrinthula culture
A known virulent strain of Labyrinthula spp., previously isolatedfrom T. testudinum, was used for all Labyrinthula elicitation exper-iments. Labyrinthula spp. cultures were maintained in serum-seawater agar (SSA) as previously described in Trevathan et al. [4].
4.3. Preparation of elicitors
Labyrinthula extract was obtained by removing 5 mm diameterplugs of Labyrinthula spp. from serum-seawater agar plates (SSA)and transferred to 800 � 1000 acrylic sheets covered in 1 L Serum-Seawater liquid media for 48 h. The liquid media was the same asthe SSA Labyrinthula culture media minus the agar. After 48 h ofgrowth all Labyrinthula that covered the surface of the plate wasremoved via razor blade, placed in microcentrifuge tubes and pel-leted by centrifugation (3000 � g for 5 min). The pelleted sampleswere subsequently lyophilized and the resulting material wasresuspended and vortexed in 0.22 mm filtered seawater (salinity of30) to create known concentrations of Labyrinthula elicitor.
Lipopolysaccharides extracted from Escherichia coli serotypeO127:B8 (Sigma Aldrich, St. Louis, MO, USA) were used for all LPSinduction assays. Stock solutions of 200 mg/mL LPS were preparedin DI water.
4.4. Hydrogen peroxide detection
The concentrationofhydrogenperoxide released into themediumsurrounding the plants was quantified using a chemiluminescence
Fig. 5. Partial sequence of an NADPH oxidase from T. testudinum. (A) Alignment of the nuclO. sativa, M. truncatula, N. tabacum, H. vulgare, and A. thaliana. The locations of the sequenlabeled and underlined. (B) Alignment of a segment of the cloned T. testudinum NADPH oxidregion of the rbohF gene from A. thaliana. The region corresponding to the aligned sequenreferences to color in this figure legend, the reader is referred to the web version of this ar
luminol assay [20] with minor modification using a Synergy 2 SLLuminescence Microplate Reader (BioTek, Winooski, VT, USA).Selected volumes of Labyrinthula extract or LPS stock solution werebrought up to a final volume of 10 mL using filtered seawater toachieve the desired final concentrations of each elicitor. Every 10 mLreplicate contained 0.5 g freshweight of second rank plant leaf tissue.At each timepoint 144 mL of samplewas combinedwith the followingreaction components directly into the microwell plate: 20 mL of248UmL�1 horseradish peroxidase (HRP) (Sigma, dissolved inpH7.8phosphate buffer); 36 mL of 300 mM luminol (5-amino-2, 3-dihydro-1,4-phthalazinedione, Sigma). Both the HRP and luminol were auto-injected and chemiluminescence was recorded with a signal inte-gration time of 10 s. Calibration with a standard curve of H2O2 wasconducted at least once during a set of any experiments. All controlswere run using the experimental setup described above without theaddition of any elicitor.
4.5. Fluorescence microscopy
To identify the cellular origin of ROS production in challengedT. testudinum sections, ROS accumulation was qualitatively moni-tored using the probe 20,70-dichlorodihydrofluoresceindiacetate(H2DCF-DA; Invitrogen, Carlsbad, CA, USA), a nonfluorescing,nonpolar compound. When H2DCF-DA reacts with cellular ester-ases, the diacetate group is cleaved off to yield the polar compoundH2DCF. Oxidation of H2DCF by ROS yields the fluorescent productDCF. Cross sections were made from second rank leaf tissue ofT. testudinum using a razor blade and incubated in 5 mL of filteredseawater (0.22 mm) containing 5 mL of 10 mM H2DCF-DA (dissolved
eotide sequence corresponding to the NAD binding pocket domain of rboh genes fromce used to design the forward and reverse degenerate primers are highlighted (red),ase fragment that shows significant sequence homology with the NAD binding pocketce is indicated on the graphical depiction of the rbohF gene. (For interpretation of theticle.)
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303302
in DMSO) for 30 min. Following the incubation, samples werewashed three times in 5 mL filtered seawater to remove any un-bound probe. Individual plant sections were placed on 35mm glassbottom microwell dishes (MatTek Corp., Ashland, MA, USA) in adrop of filtered seawater. To start the experiment, seawater waswicked off and replaced with a 200 mg/mL solution of LPS. Fluo-rescent imaging (FITC: ex 488, em 525 nm; Texas Red: ex 496, em615 nm) was taken using an Olympus IX 81 inverted epifluorescentmicroscope (Olympus Center Valley, PA, USA) in conjunctionwith aDP21 digital camera (Olympus).
4.6. Respiration and pH measurements
To evaluate the impact of LPS addition on dark-adapted respi-ration, 2-cm sections of T. testudinum (total of 0.5 g per replicate),were analyzed for oxygen uptake. Respiration studies were con-ducted using an Oxygraph system outfitted with a DW3 liquid-phase electrode chamber (Hansatech Instruments�, Norfolk, UK).Samples were dark-adapted for at least 1 h prior to any measure-ment. The system was calibrated by eliminating oxygen with theaddition of sodium hydrosulfite. An O2 maximum was created bybubbling air into filtered seawater. In the reaction chamber, the leafsections were submerged in 10mL of O2-saturated filtered seawater(0.22 mm). One mL of 5000 mg mL�1 LPS was injected into the re-action chamber to achieve a final concentration of 500 mgmL�1 LPS.Following injection, oxygen uptake was measured over the courseof 15 min for each experiment. Respiration was calculated asnanomoles of oxygen consumed per minute per gram of tissue. Toassay for alkalization of extracellular media in response to LPSaddition, 0.5 g of plant tissue was combined with 500 mg mL�1 LPSin 10mL of seawater. pHwasmonitored using an Oaktron pHmeterand probe (Oakton Instruments, Vernon Hills, IL, USA) over thecourse of 80 min.
4.7. Pharmacological inhibitors and activators of ROS production
Pharmacological inhibitors and activators of select cellulartargets were utilized to characterize the signaling pathwaysinvolved in ROS generation in T. testudinum following LPS elici-tation. Concentrations used were based upon previous studiesreported in the literature. Compounds, final concentrations usedand mechanisms of action/target were as follows: KCN (1 mM;target, heme-dependent enzymes; 39); NaN3 (1 mM; target,redox enzymes; 39); Catalase (100 U/mL; mechanism, H2O2decomposition; 39); EGTA (1 mM; mechanism, calcium chelator;43); CaCl2 (2 mM; mechanism, calcium supplement; 39); andPertussis toxin (1 mg/mL; target, G-protein inhibitor specificallyused in the study of adenylate cyclase regulation and the role of Giproteins; 45) were dissolved in H2O. Diphenyliodonium (DPI,50 mM; target, suicide inhibitor of NADPH oxidases that bindsirreversibly to the flavonoid group of the membrane-associatedgp91phox subunit; 39); and Phorbol 12-myristate 13-acetate(PMA, 15 mM; mechanism, protein kinase-c activator; 45) weredissolved in DMSO. Quinacrine (500 mM; target, flavin-dependentredox enzymes; 20); 5-nitro-2-[3-phenylpropylamino]-benzoicacid (NPPB, 100 mM; target, anion channel blocker; 20); staur-osporine (5 mM; target, protein kinases; 20); chlorpromazine-HCl(100 mM; target, phospholipase A2; 20); methoxyverapamil(100 mM; target, Ca2þ channel blocker; 45); monensin (100 mM;mechanism, Naþ ionophore; 45); A23187 (100 mM; mechanism,Ca2þ ionophore; 45) and valinomycin (100 mM; mechanism, Kþ
ionophore; 45) were dissolved in ethanol. Seagrass samples werepreincubated with selected compounds for 20 min prior to theintroduction of 200 mg mL�1 LPS. All chemicals were purchasedfrom SigmaeAldrich.
4.8. Cloning of an NADPH oxidase from T. testudinum
4.8.1. Primer designDegenerate primers were designed by aligning respiratory burst
oxidase homolog (rboh) gene sequences downloaded from thePubmed database for Hordeum vulgare (Accession numberAM265370),Oryza sativa (Accession number AY603975), A. thaliana(Accession number NM_105079), Nicotiana tabacum (Accessionnumber AJ309006), and Medicago truncatula (Accession numberAM494844) using the ClustalW2 sequence alignment tool availableon the European Bioinformatics Institute webpage (http://www.ebi.ac.uk/). Highly conserved sequences in the 50 and 30 regions ofthe rboh alignment were identified and two pairs of degenerateprimers were designed. The primer sequences are:
4.8.2. Amplification and cloningRNA was extracted using an RNeasy Plant Mini Kit (Qiagen,
Germantown, MD, USA) from T. testudinum according the manu-facturer’s protocol. Reverse transcription was performed withMoloney Murine Leukemia Virus (M-MLV) reverse transcriptase(Invitrogen) according the manufacturer’s protocol. Briefly, 100 ngof oligo dT primer was mixed with 500 ng of total RNA and dNTPsand heated to 65 �C for 5 min and quick chilled on ice. First-StrandBuffer, DTT, RNase Inhibitor and M-MLV RT was then added to thereaction and incubated at 37 �C for 50 min, followed by a 15 minincubation at 70 �C to inactivate the reaction. Polymerase chainreaction (PCR) was performed using 10% for the RT reaction, mixedwith PCR buffer, MgCl2, dNTP mix, Rboh degenerate forward andreverse primers and Taq DNA polymerase (Invitrogen). The PCRreaction was performed in an Eppendorf Mastercycler Pro at 95 �Cfor 30 s, 55 �C for 45 s, 72 �C for 60 s and cycled 35 times. PCRproducts were agarose gel purified using a QIAquick Gel ExtractionKit (Qiagen), ligated into the pGEMT-Easy plasmid (Promega,Fitchburg, WI, USA) according the manufacturer’s protocol andtransformed into DH5a competent E. coli cells (Invitrogen). Col-onies were then selected, mini-prepped using the QIAprep SpinMiniprep Kit (Qiagen), restriction digested with EcoRI (New En-gland Biolabs, Ipswich, MA, USA) to confirm the presence of aninsert and sequenced in both directions with T7 and SP6sequencing primers using ABI BigDye Terminator v3.1 cyclesequencing chemistry (EurofinsMWGOperon, Huntsville, AL, USA).Finally, the sequence from a PCR fragment generated using theRboh 30 region degenerate primer pair was blasted against theA. thaliana genome database using the nucleotide BLAST (blastn)tool available on the NCBI website optimized for a discontiguousmegablast and was found to share sequence homology with theRbohF gene.
4.9. Statistical analysis
Data for hydrogen peroxide accumulationwere analyzed using aone-way ANOVA. A Tukey’s post hoc test was performed to deter-mine significant groupings. Data for respiration were analyzedusing a student’s t-test. Data for media alkalizationwere natural logtransformed and subsequently analyzed using a repeated-mea-sures ANOVA with time as the repeated factor. The data werenormally distributed as was determined by the ShapiroeWilk test.All statistical analyses were conducted using IBM SPSS Statistics 19(IBM Corp., Armonk, NY, USA).
K. Loucks et al. / Plant Physiology and Biochemistry 70 (2013) 295e303 303
Acknowledgments
This work was conducted under permit no. SAL-109-1172A-SR is-sued through the Florida Fish andWildlife Conservation Commissionandwassupportedby theUniversityofNorth Florida’sCoastalBiologyProgram and Academic Affairs Scholarship Grant awarded to CR.
References
[1] M. Waycott, C.M. Duarte, T.J.B. Carruthers, R.J. Orth, W.C. Dennison,S. Olyarnik, A. Calladine, J.W. Fourqurean, K.L. Heck, A.R. Hughes,G.A. Kendrick, W.J. Kenworthy, F.T. Short, S.L. Williams, Accelerating loss ofseagrasses across the global threatens coastal ecosystems, Proc. Natl. Acad. Sci.U.S.A. 106 (2009) 12377e12381.
[2] L. Muehlstein, Perspectives on the wasting disease of eelgrass, Zostera marina,Dis. Aquat. Organ. 7 (1989) 211e221.
[3] E.L. Young, Studies on Labyrinthula. The etiologic agent of the wasting diseaseof eelgrass, Am. J. Bot. 30 (1943) 586e593.
[4] S.M. Trevathan, A. Kahn, C. Ross, Effects of short-term hypersalinity exposureon the susceptibility to wasting disease in the subtropical seagrass Thalassiatestudinum, Plant Physiol. Biochem. 49 (2011) 1051e1058.
[5] A.C. Bockelmann, K. Beining, T.B.H. Reusch, Widespread occurance of endo-phytic Labyrinthula spp. in northern European eelgrass Zostera marina beds,Mar. Ecol. Prog. Ser. 445 (2012) 109e116.
[6] T. Janssen, B. Bremer, The age of major monocot groups inferred from 800þrbcL sequences, J. Linn. Soc. Bot. 146 (2004) 385e398.
[7] J.D.G. Jones, J.L. Dangl, The plant immune system, Nature 444 (2006) 323e329.[8] M. Dow, M.A. Newman, E. von Roepenack, The induction and modulation of
[9] H.S. Coventry, I.A. Dubery, Lipopolysaccharides from Burkholderia cepaciacontribute to an enhanced defensive capacity and the induction of pathogenesis-related proteins in Nicotiana tabacum, Physiol. Mol. Plant 58 (2001) 149e158.
[10] G. Erbs, M. Newman, The role of lipopolysaccharide and peptidoglycan, twoglycosylated bacterial microbe-associated molecular patterns (MAMPs), inplant innate immunity, Mol. Plant Pathol. 13 (2012) 95e104.
[11] I.B. Gerber, D. Zeidler, J. Durner, I.A. Duber, Early perception responses ofNicotiana tabacum cells in response to lipopolysaccharides from Burkholderiacepacia, Planta 218 (2004) 647e657.
[12] Y. Desaki, A. Miya, B. Venkatesh, B. Tsuyumu, H. Yamane, H. Kaku, E. Minami,N. Shibuya, Bacterial lipopolysaccharides induce defense responses associatedwithprogrammedcell death in rice cells, Plant Cell Physiol. 47 (2006) 1530e1540.
[13] N. Doke, Generation of superoxide anion by potato tuber protoplasts duringthe hypersensitive response to hyphal wall components of Phytophthorainfestans and specific inhibition of the reaction by suppressors of hypersen-sitivity, Physiol. Plant Pathol. 23 (1983) 359e367.
[14] P. Wojtaszek, Oxidative burst: an early plant response to pathogen infection,Biochem. J. 322 (1997) 681e692.
[15] Z. Zhang, D.B. Collinge, H. Thordal-Christensen, Germin-like oxalate oxidase, aH2O2-producing enzyme, accumulates in barley attacked by the powderymildew fungus, Plant J. 8 (1995) 139e145.
[16] A.C. Allan, R. Fluhr, Two distinct sources of elicited reactive oxygen species intobacco epidermal cells, Plant Cell 9 (1997) 1559e1572.
[17] G.P. Bolwell, V.S. Butt, D.R. Davies, A. Zimmerlin, The origin of the oxidativeburst in plants, Free Radic. Res. 23 (1995) 517e532.
[18] G.P. Bolwell, D.R. Davies, C. Gerrish, C.K. Auh, T.M. Murphy, Comparativebiochemistry of the oxidative burst produced by rose and french bean cellsreveals two distinct mechanisms, Plant Physiol. 116 (1998) 1379e1385.
[19] M. Sagi, R. Fluhr, Production of reactive oxygen species by plant NADPH ox-idases, Plant Physiol. 141 (2006) 336e340.
[20] F.C.Küpper, B.Kloareg, J. Guern, P.Potin,Oligoguluronateselicit anoxidativeburstin the brown algal kelp Laminaria digitata, Plant Physiol. 125 (2001) 278e291.
[21] F.C. Küpper, D.G. Müller, A.F. Peters, B. Kloareg, P. Potin, Oligoalginate recog-nition and oxidative burst play a key role in natural and induced resistance ofsporophytes of Laminariales, J. Chem. Ecol. 28 (2002) 2057e2081.
[22] A.G. Zambounis, M.S. Kalamaki, E.E. Tani, E.J. Paplomatas, A.S. Tsaftaris,Expression anlaysis of defense related genes in cotton (Gossypium hirsutum)after Fusarium oxysporum f. sp. vasinfectum infection and following chemicalelicitation using salicylic acid analog and methyl jasmonate, Plant Mol. Biol.Rep. 30 (2012) 225e234.
[23] F. Weinberger, M. Friedlander, H.G. Hoppe, Oligoagars elicit a physiologicalresponse in Gracilaria conferta (Rhodophyta), J. Phycol. 35 (1999) 747e755.
[24] F. Weinberger, P. Leonardi, A. Miravalles, J.A. Correa, U. Lion, B. Kloareg,P. Potin, Dissection of two distinct defense-related response to agar oligo-saccharides in Gracilaria chilensis (Rhodophyta) and Gracilaria conferta (Rho-dophyta), J. Phycol. 41 (2005) 863e873.
[25] F.C.K. üpper, E. Gaquerel, E.M. Boneberg, S. Morath, J.P. Salaün, P. Potin, Earlyevents in the perception of lipopolysaccharides in the brown alga Laminariadigitata include an oxidative burst and activation of fatty acid oxidation cas-cades, J. Exp. Bot. 57 (2006) 1991e1999.
[26] F. Weinberger, Pathogen-induced defense and innate immunity in macro-algae, Biol. Bull. 213 (2007) 290e302.
[27] A.M. Lawrie, E.M. Noble, P. Tunnah, N.R. Brown, J.A. Endicott, Protein kinaseinhibition by staurosporine revealed in details the molecular interaction withCDK2, Nat. Struct. Mol. Biol. 4 (1997) 796e800.
[28] S. Chandra, P.F. Heinstein, P.S. Low, Activation of phospholipase A by plantdefense elicitors, Plant Physiol. 110 (1996) 979e986.
[29] Y. Fujisawa, H. Kato, Y. Iwasaki, Structure and function of heterotrimeric Gproteins in plants, Plant Cell Physiol. 42 (2001) 789e794.
[30] D. Zeidler, U. Zähringer, I. Gerber, I. Dubrey, T. Hartung, W. Bors, P. Hutzler,J. Durner, Innate immunity in Arabidopsis thaliana: lipopolysaccharides acti-vate nitric oxide synthase (NOS) and induce defense genes, Proc. Natl. Acad.Sci. U.S.A. 101 (2004) 15811e15816.
[31] U. Seydel, K. Labischinki, M. Kastowsky, K. Brandenburg, Phase behavior, su-pramolecular structure, and molecular conformation of lipopolysaccharide,Immunobiology 187 (1993) 191e211.
[32] G. Bahnweg, I. Jäckle, A new approach to taxonomy of the thraustochytrialesand labyrinthulales, in: S.T. Moss (Ed.), The Biology of Marine Fungi, Cam-bridge Publishing, New York, 1986, pp. 131e140.
[33] S. Raghukumar, Ecology of the marine protists, the labyrinthulomycetes(thraustochytrids and labyrinthulids), Eur. J. Protistol. 38 (2002) 127e145.
[34] T.M. Preston, C.A. King, Actin-based motility in the net slime mould Laby-rinthula: evidence for the role of myosin in gliding movement, J. Eukaryot.Microbiol. 52 (2005) 461e475.
[35] B.K. Ndimba, S. Chivasa, J.M. Hamilton, W.J. Simon, A.R. Slabas, Proteomicanalysis of changes in the extracellular matrix of Arabidopsis cell suspensioncultures induced by fungal elicitors, Proteomics 3 (2003) 1047e1059.
[36] J.C. Baker, E.W. Orlandi, K.L. Deahl, Oxidative metabolism in plant/bacteriainteractions: characterization of a unique oxygen uptake response of potatosuspension cells, Physiol. Mol. Plant 59 (2001) 25e32.
[37] T. Boller, Chemoperception of microbial signals in plant cells, Annu. Rev. PlantPhys. 46 (1995) 189e214.
[38] J. Zhao, L.C. Davis, R. Verpoote, Elicitor signal transduction leading to pro-duction of plant secondary metabolites, Biotechnol. Adv. 23 (2005) 283e333.
[39] D.R. Davies, L.V. Bindschedler, T.S. Strickland, P.G. Bolwell, Production ofreactive oxygen species in Arabidopsis thaliana cell suspension cultures inresponse to an elicitor from Fusarium oxysporum: implications for basalresistance, J. Exp. Bot. 57 (2006) 1817e1827.
[40] P. Liu, L. Zhang, Q.L. Wang, K. Niehaus, F. Balu�ska, J. �Samaj, J.X. Lin, Lipidmicrodomain polarization is required for NADPH oxidase-dependent ROSsignaling in Picea meyeri pollen tube tip growth, Plant J. 60 (2009) 303e313.
[41] A. Daudi, Z. Cheng, J.A. O’Brien, N. Mammarella, S. Khan, F.M. Ausubel,G.P. Bolwell, The apoplastic oxidative burst peroxidase in Arabidopsisis a major component of pattern-triggered immunity, Plant Cell 24 (2012)275e287.
[42] M.P. Viard, F. Martin, A. Pugin, P. Ricci, J.P. Blein, Protein phosphorylation isinduced in tobacco cells by the elicitor cryptogein, Plant Physiol. 104 (1994)1245e1249.
[43] R. Schwacke, A. Hagar, Fungal elicitors induce a transient release of activeoxygen species from cultured spruce cells that is dependent on Ca2þ andprotein-kinase activity, Planta 187 (1992) 136e141.
[44] R. Castro, I. Zarra, J. Lamas, Water soluble seaweed extracts modulate the res-piratory burst activity of turbot phagocytes, Aquaculture 229 (2004) 67e78.
[45] C. Ross, F.C. Küpper, R.S. Jacobs, Involvement of reactive oxygen species andreactive nitrogen species in the wound response of Dasycladus vermicularis,Chem. Biol. 13 (2006) 353e364.
[46] B. Whiteley, D. Cassel, Y.X. Zhuang, L. Glaser, Tumor promoter phorbol 12-myristate 13-acetate inhibits mitogen-stimulated Naþ/Hþ exchange in hu-man epidermoid carcinoma A431 cells, J. Cell Biol. 99 (1984) 1162e1166.
[47] E. Afrasiabi, J. Ahlgren, N. Bergelin, K. Törnquist, Phorbol 12-myristate 13-acetate inhibits FRO anaplastic human thyroid cancer cell proliferation byinducing cell cycle arrest in G1/S phase: evidence for an effect mediated byPKCd, Mol. Cell. Endocrinol. 292 (2008) 26e35.
[48] L. Legendre, Y.G. Yueh, R. Crain, N. Haddock, P.F. Heinstein, P.S. Low, Phos-pholipase C activation during elication of the oxidative burst in cultured plantcells, J. Biol. Chem. 268, 24559e24563.
[49] S. Chandra, P.S. Low, Measurement of Ca2þ fluxes during elicitation of theoxidative burst in aequorin-transformed tobacco cells, J. Biol. Chem. 272(1997) 28274e28280.
[50] C.E. Moya, R.S. Jacobs, Pseudopterosin A inhibits phagocytosis and altersintracellular calcium turnover in a pertussis toxin sensitive site in Tetrahy-mena thermophile, Comp. Biochem. Physiol. C. 143 (2006) 436e443.
[51] G.P. Bolwell, M.W. Rodgers, D.L. Murphy, D. Jones, Modulation of the elicita-tion response in cultured French bean cells its implication for the mechanismof signal transduction, Phytochemistry 30 (1991) 397e405.
[52] G.B. Mahady, C. Liu, C.W.W. Beecher, Involvement of protein kinase and Gproteins in the signal transduction of benzophenanthridine alkaloid biosyn-thesis, Phytochemistry 48 (1998) 93e102.
[54] Y. Ogasawara, H. Kaya, G. Hiraoka, et al., Synergistic activation of the Arabi-dopsis NADPH oxidase Atrboh by Ca2þ and phosphorylation, J. Biol. Chem. 283(2008) 8885e8892.
[55] L.V. Bindschedler, F. Minibayeva, S.L. Gardner, C. Gerrish, D.R. Davies,G.P. Bolwell, Early signaling events in the apoplastic oxidative burst insuspension-cultured French bean cells involve cAMP and Ca2þ, New Phytol.151 (2001) 185e194.