JPM Vol 27. No. 2 April 1992:79-83 Liver Microsomes Contain Muitiple Forms of Inositoi 1,4,STrisphosphate Binding Proteins: identification by Nitroceluiose Blot Overlay Nawab Afi and Devendra K. Agrawal Deparrment of Internal Medicine, Chighton Universify School qf’ Medicine, Omaha, Nebraska, U.S.A. A group of proteins binding to inositol ~,4,~~trisphosphate (lP3) has been identi~ed in rat liver microsomes by a nitrocellulose blot-overlay technique. Proteins were resolved by SDS-PAGE, blotted on nitrocellulose and incubated with [32P]IP3 followed by autoradiogra- phy. Approximately eight OPT-binding polypeptides ranging Mr 23-50 kDA were present exclusively in microsomes; these were absent from ptasma membrane and mitochondria~ fractions. Binding of [32PlIP3 to these proteins was displaceable to a great extent by 5 PM unlabeled XPabut not by 10 PM IPI, IP2, IPd, ATP, or GTPyS. These results suggest that liver microsomes contain multiple forms of OPT-binding proteins that can be detected by this new method. Keywords: IP3 binding proteins; Liver microsomes; Nitrocellulose blot overlay: Signal transduction, PI turnover The hydrolysis of phosphatidyIin~sit~l4,5bisphos- phate by phosphatidyli~~sitof-speci~c phosphofipase C is a major signal transduction pathway during stimu- fation of many cells by hormones and other regulatory figands, Inositof I ,4,5-t~sphosphate (IPJ) and diacyf- giycerol generated during phosphoinositide hydrolysis act as second messengers to increase i~tracelfuIar Ca2+ and to activate protein kinase C, respectively, The role of IP3 in calcium mobilization from nonmito- chondrial intracellular stores has been well established (Dawson and Irvine, 1984). It has been postulated that IP3 is released from the inner leaflet of the plasma membrane and binds to IPs receptors present on endo- plasmic reticulum which has a high capacity for cai- cium accumdation. The existence of IP:, receptors has been demonst~ted in many cells (Guiliemette et al., 1988; Sh~shan-Barmatz et al., 1990) by radioiigand binding experiments. The receptor has subsequently Address reprint requests to: Dr. Devendra K. Agrawal, Allergic Disease Center, Creighton University School of Medicine, Omaha, NE 68178, U.S.A. Received I August 1991; revised and accepted 4 November HI. Journal of ~~~acologi~al and Toxicological Methods 27, 79-83 (19923 0 1992Eiscvier Science Publishing Co., inc., 655 Avenue of the Americas. New York, NY 10010 been puri~ed (Suppatopo~e et at., 1988) and cloned {Furuichi et al., 1989). Although the molecular identity of only one IPs- binding protein corresponding to 260 kDA has been demonstrated (Suppatopone et al., 1988), heterogene- ity with regard to the affinity of IP, binding in various tissues has been described (~halfiss et al., 1990; Var- ney et af., 1990). It is not clear whether variation in apparent IP~-birding affinity is due to tissue-specific factors such as the state of phosphoryiation of the re- ceptors or the presence of more than one type of IP3- binding protein. Also, it is not known whether subtypes of IP3 receptors exist (Varney et al., 1990; Wiltcocks and Nahorski, 1989). Recently, inositol 1. ,3,4,5-tetra- kisphosphat~ (Theibert et al., 1990) and inositol hexa- kisphosphate (Nicoletti et al., 1990) binding sites that are distinct from IP3 receptors have been demonstrated in the brain. It is also possible that other protein com- ponents, such as inositol phosphate phosphatases and kinases which might interact with IP3 and other inositof po~yphosphates, are present in the cell and that these components may account far the observed heterogene- ity in the IP~-binding affinities (Challiss et al., 1990). In this study, we explored the possibifity that a group of proteins that binds to IP, exists in rat liver
Transcript
JPM Vol 27. No. 2 April 1992:79-83
Liver Microsomes Contain Muitiple Forms of Inositoi 1,4,STrisphosphate Binding Proteins: identification by Nitroceluiose
Blot Overlay
Nawab Afi and Devendra K. Agrawal
Deparrment of Internal Medicine, Chighton Universify School qf’ Medicine, Omaha, Nebraska, U.S.A.
A group of proteins binding to inositol ~,4,~~trisphosphate (lP3) has been identi~ed in rat liver microsomes by a nitrocellulose blot-overlay technique. Proteins were resolved by SDS-PAGE, blotted on nitrocellulose and incubated with [32P]IP3 followed by autoradiogra- phy. Approximately eight OPT-binding polypeptides ranging Mr 23-50 kDA were present exclusively in microsomes; these were absent from ptasma membrane and mitochondria~ fractions. Binding of [32PlIP3 to these proteins was displaceable to a great extent by 5 PM unlabeled XPa but not by 10 PM IPI, IP2, IPd, ATP, or GTPyS. These results suggest that liver microsomes contain multiple forms of OPT-binding proteins that can be detected by this new method.
Keywords: IP3 binding proteins; Liver microsomes; Nitrocellulose blot overlay: Signal transduction, PI turnover
The hydrolysis of phosphatidyIin~sit~l4,5bisphos- phate by phosphatidyli~~sitof-speci~c phosphofipase C is a major signal transduction pathway during stimu- fation of many cells by hormones and other regulatory figands, Inositof I ,4,5-t~sphosphate (IPJ) and diacyf- giycerol generated during phosphoinositide hydrolysis act as second messengers to increase i~tracelfuIar Ca2+ and to activate protein kinase C, respectively, The role of IP3 in calcium mobilization from nonmito- chondrial intracellular stores has been well established (Dawson and Irvine, 1984). It has been postulated that IP3 is released from the inner leaflet of the plasma membrane and binds to IPs receptors present on endo- plasmic reticulum which has a high capacity for cai- cium accumdation. The existence of IP:, receptors has been demonst~ted in many cells (Guiliemette et al., 1988; Sh~shan-Barmatz et al., 1990) by radioiigand binding experiments. The receptor has subsequently
Address reprint requests to: Dr. Devendra K. Agrawal, Allergic Disease Center, Creighton University School of Medicine, Omaha, NE 68178, U.S.A.
Received I August 1991; revised and accepted 4 November HI.
Journal of ~~~acologi~al and Toxicological Methods 27, 79-83 (19923 0 1992 Eiscvier Science Publishing Co., inc., 655 Avenue of the Americas. New York, NY 10010
been puri~ed (Suppatopo~e et at., 1988) and cloned {Furuichi et al., 1989).
Although the molecular identity of only one IPs- binding protein corresponding to 260 kDA has been demonstrated (Suppatopone et al., 1988), heterogene- ity with regard to the affinity of IP, binding in various tissues has been described (~halfiss et al., 1990; Var- ney et af., 1990). It is not clear whether variation in apparent IP~-birding affinity is due to tissue-specific factors such as the state of phosphoryiation of the re- ceptors or the presence of more than one type of IP3- binding protein. Also, it is not known whether subtypes of IP3 receptors exist (Varney et al., 1990; Wiltcocks and Nahorski, 1989). Recently, inositol 1. ,3,4,5-tetra- kisphosphat~ (Theibert et al., 1990) and inositol hexa- kisphosphate (Nicoletti et al., 1990) binding sites that are distinct from IP3 receptors have been demonstrated in the brain. It is also possible that other protein com- ponents, such as inositol phosphate phosphatases and kinases which might interact with IP3 and other inositof po~yphosphates, are present in the cell and that these components may account far the observed heterogene- ity in the IP~-binding affinities (Challiss et al., 1990).
In this study, we explored the possibifity that a group of proteins that binds to IP, exists in rat liver
80 JPM Vol 27. No. 2 April 1992:79-83
microsomes in addition to the known IP3 receptors. We have developed a new technique to identify IP3- binding proteins by incubation of nitrocellulose blots containing immobilized proteins in the presence of [=P]IP3.
Materials and Methods
Materials
D-[5-32P]-inositol-1 ,4,5trisphosphate (specific ac- tivity = 111 Ci/mmol) was obtained from New England Nuclear, DuPont. GTPyS, Tris-ATP and unlabeled IPi, IP2, IP3, and IP4 were purchased from Sigma Chemical Co., St. Louis, MO. Nitrocellulose mem- brane papers (0.1 pm) were obtained from Schleicher and Schuell, Inc., Keene, NH. Other materials were either from Sigma or Fisher Chemical Co.
Preparation of Subcellular Fractions
Subcellular fractions originating from the sinusoidal and bile canalicular domains of the hepatocyte plasma membrane, and microsomes were prepared from the livers of Sprague-Dawley male rats (150-200 g) and characterized, as previously described (Wisher and Evans, 1975; Evans, 1980; Ali et al., 1989, 1990). Liv- ers were first perfused with ice-cold 150 mM NaCl and then homogenized in 250 mM sucrose containing 1 mM phenylmethy-sulfonyl fluoride (PMSF), and 5 pg/ml each of leupeptin, antipain, and aprotinin. A bile can- alicular plasma membrane fraction from the nuclear pellet and sinusoidal plasma membrane and microso- ma1 fractions from the postnuclear supernatant were collected (Ali et al., 1990).
Microsomal fractions were also prepared and char- acterized as described by Dawson and Irvine (Dawson and Irvine, 1984). Briefly, postnuclear supernatant was first centrifuged at 8000 g for 10 min to remove mito- chondria and then at 35,000 g for 20 min. The latter pellet was resuspended in 250 mM sucrose and centri- fuged at 35,000 g for 20 min to obtain the microsomal fraction. A third method for the preparation of micro- somes was also used (Casals et al., 1985). In this method, tissue homogenate in 50 mM potassium phos- phate buffer, pH 7.4, containing 150 mM KCl, 1 mM EDTA, 1 mM DTE, and 10% glycerol was first centri- fuged for 5 min at 2500 g and then twice for 15 min at 18,000 g in a Sorvall centrifuge. Postmitochondrial supernatants were then centrifuged at 105,000 g in a Beckman ultracentrifuge. The microsomal pellet was resuspended in the same buffer and centrifuged at 150,000 g. The pellets containing microsomes were re- suspended in the same buffer and stored at -70°C for further analysis. Protein was measured by the Bradford
method (Bradford, 1976) using bovine serum albumin as the standard. Relative purity of microsomal and mi- tochondrial fractions were assessed using rotenone-in- sensitive NADH-cytochrome C reductase (Fleischer and Fleischer, 1970) and succinate-cytochrome C re- ductase (Fleischer and Fleischer, 1967) as their respec- tive markers.
Electrophoresis, Electroblotting, and Staining by [ 32P]IP3
Proteins (100 kg) were solubilized in Laemmli buffer containing 75 mM Tris-HCl, pH 6.8, 2% SDS, 0.55 M B-mercaptoethanol, 10% glycerol, and 0.003% Bromo- phenol Blue, boiled for 2-3 min and resolved by poly- acrylamide-gel electrophoresis in SDS-containing buff- ers. The resolved proteins were transferred to nitrocel- lulose papers by electrophoresis for 16 hr at 20 V as described (Ali et al., 1989). Nitrocellulose papers were washed 3 x 10 min with buffer A (50 mM Tris-HCl, pH 7.5, 10 mM MgC12, 1 mM DTT, and 0.3% Tween- 20) and then incubated for 30 min at room temperature with buffer A containing [32P]IP3 (1 nM) with and with- out unlabeled inositol phosphates, GTPyS or ATP. Ni- trocellulose papers were washed 3 x 10 min with buffer A, dried, and autoradiographed.
Results
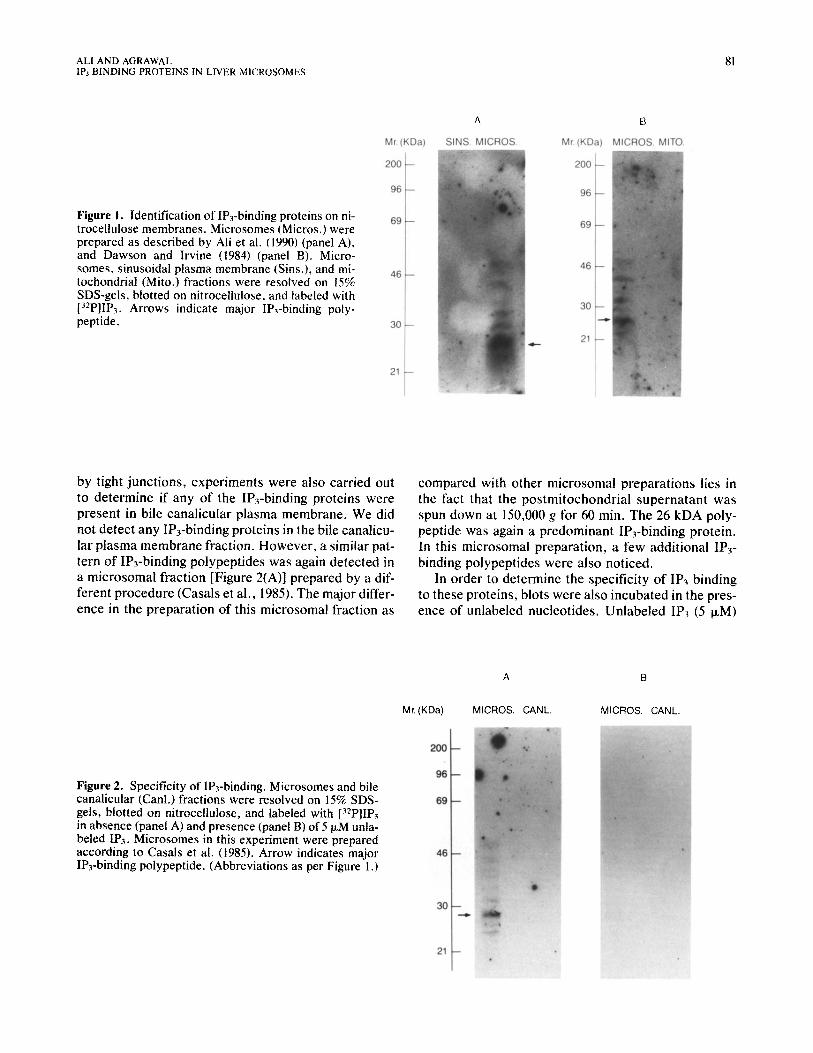
Subcellular fractions were resolved by SDS-PAGE and electrophoretically transferred to nitrocellulose paper. IP3-binding proteins were identified by incuba- tion of the blot with 1 nM [32P]IP3 followed by autoradi- ography. We routinely observed about eight polypep- tides exclusively in the microsomal fraction. They were bound to 1P3 and ranged in molecular weight be- tween 23 and 50 kDA [Figure l(A)]. The labeling inten- sity of various polypeptides varied; the most intensely labeled polypeptide migrated at approximately 26 kDA on SDS-gel. This microsomal fraction was obtained during subcellular fractionation of sinusoidal plasma membrane (Ali et al., 1989). None of the IP3-binding polypeptides were detected in sinusoidal plasma mem- brane. For IP3-binding proteins, we have also used the microsomal and mitochondrial fractions prepared si- multaneously as previously described (Dawson and Ir- vine, 1984). The pattern of IP3-binding polypeptides found in the microsomal fractions [Figure l(B)] was similar to that in Figure l(A). None of the IP3-binding polypeptides were detected in the mitochondrial frac- tion [Figure l(B)].
Because hepatocytes are polarized cells having si- nusoidal (basolateral) and bile canalicular (apical) plasma membrane domains at opposite ends separated
ALI AND AGRAWAL 81 IF’, BINDING PROTEINS IN LIVER MICROSOMES
Figure 1. Identification of IPj-binding proteins on ni- trocellulose membranes. Microsomes (Micros.) were prepared as described by Ali et al. (1990) (panel A), and Dawson and Irvine (1984) (panel B). Micro- somes, sinusoidal plasma membrane (Sins.), and mi- tochondrial (Mito.) fractions were resotved on 15% SDS-gels, blotted on nitrocellulose, and labeled with [32PlIP~. Arrows indicate major IPj-binding poly- peptide.
Mr. (KDa)
36 t
69
46 I-
21 - /
by tight junctions, experiments were also carried out to determine if any of the IP3-binding proteins were present in bile canalicular plasma membrane. We did not detect any IP3-binding proteins in the bile canahcu- lar plasma membrane fraction. However, a similar pat- tern of IPrbinding polypeptides was again detected in a microsomal fraction [Figure 2(A)] prepared by a dif- ferent procedure (Casals et al., 1985). The major differ- ence in the preparation of this microsomal fraction as
Figure 2. Specificity of IP3-binding. Microsomes and bile canalicular (Canl.) fractions were resolved on 15% SDS- gels, blotted on nitrocellulose, and labeled with [32P]IP3 in absence (panel A) and presence (panel B) of 5 pM unla- beled IP3. Microsomes in this experiment were prepared according to Casals et al. (1985). Arrow indicates major IP3-binding polypeptide. (Abbreviations as per Figure 1.)
A
SINS. MICROS.
0
Mr. (KDa) MICROS. MITO.
200 ’ t-
96 I-
30
F 21 I--
compared with other microsomal preparations lies in the fact that the postmitochondrial supernatant was spun down at 150,000 g for 60 min. The 26 kDA poly- peptide was again a predominant IP3-binding protein. In this microsomal preparation, a few additional IP3- binding polypeptides were also noticed.
In order to determine the specificity of IP3 binding to these proteins, blots were also incubated in the pres- ence of unlabeled nudeotides. Unlabeled IP3 (5 FM)
A
Mr. (KDa) MICROS. CANL.
96
69
46
0
MICROS. CANL.
82 JPM Vol 27. No. 2 April 1992:79-83
Mr. (KDa) A 8 c
Figure 3. Effect of IP2 and IP4 on the IP3 binding. Micro- somes prepared by the method of Casals et al. (1985) were resolved on 17% SDS-gels, blotted on nitrocellulose, and la- beled with [‘2P]IP~ in the absence (panel A) and presence of IP2 (panel B) or IP4 (panel C). Arrows indicate major IP,- binding polypeptide.
reduced the [32PlIP3 binding to a great extent [Figure 2(B)]. However, complete abolition of the IP3 binding was not observed even at 10 FM unlabeled IP3 (results not shown). We next considered whether IP3 binding was displaceable by other inositol phosphates and nu- cleotides. Figure 3 shows that incubation of the blots in the presence of 10 FM of IP2 and IP4 had no effect on the IP3 binding. Also, IPi, ATP, and GTPyS at a concentration of 10 p.M did not reduce the IP3 binding (not shown), suggesting that these proteins specifically bind to IP3.
Discussion
This paper describes a method for identification of a group of IP3-binding proteins that can be detected by radiolabeled IP3 overlay on nitrocellulose blots. These proteins represent a group of about eight polypeptides migrating on SDS-gels between Mr 23 and 50 kDA and have the ability to bind with IP3 even after electropho- resis and electroblotting. Although in a competition ex- periment binding of radioactive IP3 was not displaced completely by unlabeled IP3, it was greatly reduced. Specificity of IP3 binding was also evident in that the binding was not displaceable by other inositol phos- phates and nucleotides. The present method for identi- fication of IP3-binding proteins is analogous to the methods used for identification of a number of other proteins on nitrocellulose membrane, for example, Ca-, calmodulin-, and GTP-binding proteins (Maruy- ama et al., 1984; Bhullar and Haslam, 1987).
In order to demonstrate exclusive localization of
IP3-binding proteins in microsomes, we have used three different methods of isolation of microsomes and checked the purity with respect to contamination of mitochondria (succinate-cytochrome C reductase ac- tivity in microsomes was <IO% of that in mitochon- dria). Microsomal fractions having NADH-cyto- chrome C reductase activity more than 60-fold com- pared with the homogenate were used for IP3-binding proteins. We observed virtually identical IP3-binding polypeptide patterns in all microsomal preparations. A slight variation in IP3-binding polypeptide pattern was apparent in microsomes (Figure 2) obtained after high- speed centrifugation (150,000 g for 30 min). In this mi- crosomal fraction, although a few additional IP3-bind- ing polypeptides were present, the predominant poly- peptide was still recovered at 26 kDA. We have also used two plasma membrane fractions and a mitochon- drial fraction of established purity to examine the pres- ence of IP3-binding proteins. We found no detectable IP3-binding polypeptides in the fractions other than mi- crosomes.
Because an IP3-binding protein of 260 kDA which serves as a receptor for IP,-induced calcium mobiliza- tion is known (Suppatopone et al., 1988), one would expect the binding of IP3 at a higher molecular weight range (260 kDA). However, this method did not detect an IP3 receptor at 260 kDA. It is, however, possible that this method may not recognize all IP3-binding pro- teins on nitrocellulose membrane.
The possibility of the newly identified IP3-binding proteins being proteolytic fragments of IP3 receptor is highly unlikely; first, because there was no IP3 binding at 260 kDA, and second, proteolytic degradation was avoided by including a mixture of protease inhibitors. It may be possible that there are several IPj-binding proteins present in microsomes that can now be de- tected by IP3 binding on nitrocellulose. In fact, hetero- geneity with regard to the affinity of IP3-binding has been observed (Challiss et al., 1990; Varney et al., 1990), and it has recently been shown that IP3 may interact with more than one type of IP3-binding site (Challiss et al., 1991). Furthermore, the IP3-binding proteins detected on the blots may correspond, in part, to some of the protein components such as inositol polyphosphate phosphatases and/or kinases that are involved in the metabolism of IP3 and other inositol polyphosphates (Berridge and Irvine, 1989; Stephens and Irvine, 1990) and may have sites inter- acting with IP3.
In conclusion, we have identified a group of IP3- binding proteins by a nitrocellulose blot-overlay tech- nique. Although the function(s) of these IP3-binding proteins is not clear at present, their exclusive location in microsomes suggests possible involvement in the regulation of intracellular calcium.
ALI AND AGRAWAL IPx BINDING PROTEINS IN LIVER Mt~ROSOMES
83
References Ali N, Milligan G, Evans WH (1989) Distribution of G-proteins in
rat liver plasma membrane domains and endocytic pathways. Biorhem J 261:905-912.
Furuichi T, Yoshikawa S, Miyawaki A, Wada K, Maeda N, Mikos- hiba K (1989) Primary structure and functional expression of the inositol 1,4,5-trisphosphate-binding protein P~oo. Nnrttre 342:32-38.
Guillemette G. Balla T. Baukal AJ. Catt KJ (1988) Characterization
Ali N, Aligue R, Evans WH (1990) Highly purified bile-canalicular vesicles and lateral plasma membranes isolated from rat liver on Nycodenz gradients. ~~0~~~~ J 271: 185-192.
Bhullar RP, Haslam RJ (1987) Detection of 23-27 KDa GTP-binding proteins in platelets and other cells. Biothem J 245:6i7-620.
Bradford M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of pro- tein-dye binding. Anal Biochem 72~248-254.
Casals C. Herrero LM, Soriano Y, Garcia-Barren0 P, Municio AM (1985) Membrane regulation of liver and lung microsomes under low oxygen tension. Biochem Bi~p~y~ Res Common 126:551-558.
Challiss RAJ, Chilvers ER, Willcocks AL, Nahorski SR (1990) Het- erogeneity of [3H]inositol I ,4,5_trisphosphate binding sites in ad- renal-cortical membranes: Characterization and validation of ra- dioreceptor assay. Biochem .I 26.5:421-427.
Challiss RAJ, Smith SM, Potter BVL, Nahorski SR (1991) D-
[35S9U)]inosito1 1,4,5-trisphosphorothioate, a novel radioligand for the inositol t ,4,5_trisphosphate receptor: Complex binding to rat cerebellar membranes. FEBS Lerr 281: 101-104.
Dawson AP, Irvine RF (1984) Lnositol (1,4,S)trisphosphate-pro- moted Ca’ + release from microsomal fractions of rat liver. Bio- them Biophys Res Commun 120:858-864.
Evans WH (1980) A biochemical dissection of the functional polarity
of inositol 1.4,5-trisphosphate receptor and calcium mobilization in hepatic plasma membrane fraction. J Biol Chem 263:4541-4548.
Maruyama K, Mikawa T, Ebashi S f 1984) Detection of calcium bind- ing proteins by 45Ca autoradiography on nitrocellulose membrane after sodium dodecyl sulfate gel electrophoresis. J Biochem 96~511-519.
Nicoletti F, Bruno V, Cavallaro S, Copani A, Sortino MA, Canonico PL (1990) Specific binding sites for inositol hexakisphosphate in brain and anterior pituitary. Molec f’harmaco/37:689-693.
Shoshan-Barmatz V, Zhang GH. Garretson L, Kraus-Friedmann N (1990) Distinct rynodine and inositol I ,4,5-trisphosphate-binding sites in hepatic microsames. B&hem J 268:699-705.
Stephens LR, Irvine RF (19901 Stepwise phosphorylation of myo- inositol leading to myoinositol hexakisphosphate in dictyostel- ium. Nurure 346:580-583.
Suppatopone S, Worley PF, Baraban JM, Snyder SH (1988) Solubili- zation, purification, and characterization of an inositol trisphos- phate receptor. f Biol Chem 263: 1530- 1534.
Theibert AM, Supattapone S, Ferris CD, Danoff SK, Evans RK, Snyder SH (1990) Solubilization and separation of inositol 1,3,4,5tetrakisphosphate- and inositol I ,4,5-trisphosphate-bind- ing proteins and metabolizing enzymes in rat brain. Biochem 3 268:441-445.
Vamey MA, Rivera J, LobezBemal A, Watson SP (1990) Are there subtypes of the inositol I ,4,5trisphosphate receptor? Biochem J 269:21 I-216.
of the plasma membrane of the hepatocyte. Bioc’hem biophys Actn 604:27-64.
Fleischer S, Fleischer B (1967) Removal and binding of polar lipids in mitochond~a and other membrane systems. methods E~~y~~~~ 10:405-433.
Fleischer B, Fleischer S (1970) Preparation and characterization of Golgi membranes from rat liver. Biochem Biophys Acfa 219:301-319.
Willcocks AL, Nahorski SR (1989) Stereospecific binding of myo- inositol (I ,4,5) trisphosphate to rat cerebellum and bovine adrenal cortex: Evidence for heterogeneity. Br J Phurm~c~l96:364P.
Wisher MN, Evans WH (1975) Functional polarity of the rat hepato- cyte surface membrane: Isolation and characterization of plasma membrane subfractions from the blood sinusoidal, bile canalicu- lar and contiguous surfaces of the hepatocyte. Biochem J 146:375-388.