Macro- and microstructural diversity of sea urchin teeth revealed by large-scale micro-computed tomography survey
Alexander Ziegler*a, Stuart R. Stockb, Björn H. Menzec, Andrew B. Smithd
aMuseum of Comparative Zoology, Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA; bFeinberg School of Medicine, Northwestern University, 710 N Lake Shore Drive, Chicago, IL 60611, USA; cInstitut für
Bildverarbeitung, Eidgenössische Technische Hochschule, Sternwartstraße 7, 8092 Zürich, Switzerland; dDepartment of Palaeontology, The Natural History Museum, Cromwell Road, London
SW7 5BD, UK
ABSTRACT
Sea urchins (Echinodermata: Echinoidea) generally possess an intricate jaw apparatus that incorporates five teeth. Although echinoid teeth consist of calcite, their complex internal design results in biomechanical properties far superior to those of inorganic forms of the constituent material. While the individual elements (or microstructure) of echinoid teeth provide general insight into processes of biomineralization, the cross-sectional shape (or macrostructure) of echinoid teeth is useful for phylogenetic and biomechanical inferences. However, studies of sea urchin tooth macro- and microstructure have traditionally been limited to a few readily available species, effectively disregarding a potentially high degree of structural diversity that could be informative in a number of ways. Having scanned numerous sea urchin species using micro-computed tomography (µCT) and synchrotron µCT, we report a large variation in macro- and microstructure of sea urchin teeth. In addition, we describe aberrant tooth shapes and apply 3D visualization protocols that permit accelerated visual access to the complex microstructure of sea urchin teeth. Our broad survey identifies key taxa for further in-depth study and integrates previously assembled data on fossil species into a more comprehensive systematic analysis of sea urchin teeth. In order to circumvent the imprecise, word-based description of tooth shape, we introduce shape analysis algorithms that will permit the numerical and therefore more objective description of tooth macrostructure. Finally, we discuss how synchrotron µCT datasets permit virtual models of tooth microstructure to be generated as well as the simulation of tooth mechanics based on finite element modeling.
1. INTRODUCTION Most sea urchin species (Echinodermata: Echinoidea) possess an internal jaw apparatus commonly known as the Aristotle’s lantern. This complex organ consists of 40 skeletal and numerous soft tissues elements that are arranged with pentameral symmetry. The Aristotle's lantern lies inside the sea urchin's test (Fig. 1A), with only the tips of its five teeth protruding externally. Each tooth is an elongate, slightly curved rod-like structure (Fig. 1B) that is constantly being worn down at its tip as it is used to rasp and bite with a vice-like action. Because sea urchin teeth have evolved to withstand deformation encountered during feeding, they are arguably the most complex skeletal structure in echinoderms and have been the focus of studies involving biomineralization, biomechanics, and systematics for over 150 years [1-13].
When viewed in transverse section (Fig. 1C, D), echinoid teeth are highly variable in shape, which provides important information for establishing phylogenetic relationships [5, 14, 15]. Traditionally, four types of teeth are distinguished: keeled teeth which are T-shaped in cross-section (Fig. 1E), grooved teeth which are U-shaped in cross-section (Fig. 1F), diamond-shaped teeth (Fig. 1G), and wedge-shaped teeth (Fig. 1H). In addition to variation in macrostructure, sea urchin teeth also have a complex microstructure that has been documented in the past using light microscopy, scanning electron microscopy, and micro-computed tomography (µCT) [16-24]. Each tooth is constructed of parallel arrays of curved plates stacked within each other like ice-cream cones as well as long prisms and fine needles all bound together by secondary calcite growth. There is an abaxial zone of primary plates, a narrow core of needles connected to the primary
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
V.4r1 > 40,
Atirely 4_ e 7
44>,,e1 0\ v.
D-'E' E 4%1
plates by lamellae known as the stone zone, and an adaxial zone of secondary plates and prisms [6, 25, 26]. When fully mineralized, each tooth represents a single crystal of high-magnesium calcite. The precise arrangement of tooth plates and prisms varies considerably amongst echinoids, yet few studies have tried to synthesize this information [5, 15, 27]. In addition, tooth macrostructure has so far been described only in qualitative ways using ambiguous and vague terminology. A better understanding of how tooth morphology varies across sea urchins would therefore not only improve our understanding of their evolutionary relationships, but also of rates and spatial distribution of biomineralization in echinoid teeth, and the mechanisms producing mechanical properties superior to those of inorganic calcite.
Figure 1. The location of teeth within Aristotle's lantern is illustrated by cut volume renderings (A, C) and by longitudinal (B) as well as transverse (D) sections through a µCT dataset of Echinus esculentus (Echinidae). This species has five keeled teeth (E). Apart from keeled teeth, three other forms of sea urchin teeth have traditionally been differentiated: grooved teeth (F, Histocidaris elegans), diamond-shaped teeth (G, Apatopygus recens), and wedge-shaped teeth (H, Echinarachnius parma). Table 1 provides further specimen and dataset information. Not to scale.
Unfortunately, large-scale morphological surveys of internal structures like the Aristotle's lantern have been difficult to achieve in the past because they require the destruction of specimens in order to isolate the elements of interest. While common shallow-water echinoid species such as Arbacia lixula, Paracentrotus lividus, and Lytechinus variegatus can easily be obtained for destructive sampling, rarer species housed in museum collections usually cannot. The application of tomographic imaging techniques that reveal soft or hard parts non-invasively is therefore a major step forward as it permits the use of museum specimens on a large scale and therefore results in much wider taxonomic sampling [28]. One of these imaging techniques, µCT, is particularly suitable for studies on sea urchins and various other echinoderms because of the presence of large amounts of calcified tissue in this group of marine organisms [29]. We here present first results from a broad survey of tooth macro- and microstructure based on image data derived from µCT and synchrotron µCT scanning experiments.
2. MATERIALS AND METHODS 2.1 Micro-computed tomography
Whole dry and wet sea urchin specimens were obtained from natural history museum collections. Table 1 mentions all samples analyzed by µCT that were used in the present contribution, while [29] provides a full list. Depending on their original state of preservation, the animals were scanned either in air or in 75% ethanol. In the course of our studies on sea urchin teeth, a total of 182 extant sea urchin species were scanned as whole specimens. Two µCT systems were used: a
Proc. of SPIE Vol. 8506 85061G-2
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
Phoenix Nanotom X-ray tube tomography system equipped with a tungsten target and a 2,300 x 2,300 pixel detector (GE Sensing & Inspection Technologies GmbH, Germany), and a HMX ST 225 X-ray tube tomography system equipped with a tungsten target, and a 2,000 x 2,000 pixel detector (Nikon Metrology, Leuven, Belgium). Projections were recorded over 360°. Dataset voxel sizes ranged from 3.3 µm to 36.5 µm isotropic, depending on the size of the specimen. Dataset reconstruction was accomplished using the software provided with each individual scanner, i.e., Phoenix DatosX Reconstruction 1.5 for the GE system and Metris XT 2.2 for the Nikon system. Reconstruction was performed without noise reduction or binning.
Table 1. Analysis of tooth macrostructure. List of whole specimens analyzed for the present contribution using µCT. BMNH = Natural History Museum, London; CASIZ = California Academy of Sciences Invertebrate Zoology; MCZ = Museum of Comparative Zoology, Cambridge, MA, USA; NHMW = Naturhistorisches Museum Wien; USNM = National Museum of Natural History; ZMB = Museum für Naturkunde Berlin; ZMH = Zoologisches Museum Hamburg; ZMK = Zoologisk Museum Købnhavn; ZSM = Zoologische Staatssammlung München.
Family Species Specimen number Voxel size Tooth width Histocidaridae Histocidaris elegans ZMH E307 16.0 µm 1.6 mmCidaridae Chondrocidaris gigantea MCZ 4074 18.3 µm 2.4 mm Cidaris cidaris BMNH 1925.10.30.103-113 18.9 µm 1.3 mm Compsocidaris pyrsacantha MCZ 7861 29.6 µm 1.5 mm Eucidaris metularia BMNH 1969.5.1.15-40 13.9 µm 0.7 mm Phormosomatidae Phormosoma placenta USNM E17633 13.9 µm 0.7 mm Echinothuriidae Araesoma belli MCZ 7765 15.1 µm 1.1 mm Sperosoma obscurum MCZ 903 20.0 µm 1.7 mm Micropygidae Micropyga tuberculata BMNH 98.8.8.45/6 13.9 µm 0.9 mm Diadematidae Centrostephanus longispinus BMNH 1952.3.26.64-8 13.9 µm 0.6 mm Diadema ascensionis BMNH 1972.8.22.50-52 13.4 µm 1.1 mm Aspidodiadematidae Aspidodiadema arcitum USNM 27568 9.8 µm 0.7 mm Plesiodiadema indicum ZMB Ech 7232 13.9 µm 1.2 mm Pedinidae Caenopedina mirabilis USNM 31182 13.9 µm 0.6 mm Saleniidae Salenia goesiana USNM 14581 8.9 µm 0.5 mm Salenocidaris hastigera ZMB Ech 5816 13.9 µm 0.7 mm Salenocidaris varispina MCZ 4883 7.4 µm 0.4 mm Arbaciidae Arbacia lixula BMNH 1966.5.6.57-65 13.9 µm 0.6 mm Dialithocidaris gemmifera MCZ 8317 10.9 µm 0.6 mm Pygmaeocidaris prionigera MCZ 8741 6.0 µm 0.3 mm Stomopneustidae Stomopneustes variolaris USNM E45930 13.9 µm 0.7 mm Parasaleniidae Parasalenia gratiosa BMNH 1983.2.15.7 14.0 µm 0.8 mm Parechinidae Paracentrotus lividus ZMB Ech 7406 13.4 µm 0.8 mm Echinidae Echinus esculentus ZMB Ech 3826 13.9 µm 0.9 mm Toxopneustidae Lytechinus variegatus ZMB Ech 7408 13.9 µm 0.5 mm Echinometridae Echinometra mathaei BMNH 1969.5.1.61-75 13.9 µm 1.0 mm Evechinus chloroticus NHMW 10898 30.0 µm 1.6 mm Heliocidaris erythrogramma ZMB Ech 5745 13.9 µm 0.8 mm Strongylocentrotidae Strongylocentrotus purpuratus CASIZ 5724 13.9 µm 0.8 mm Temnopleuridae Amblypneustes pallidus ZMB Ech 6334 13.9 µm 0.9 mm Holopneustes inflatus ZMB Ech 2639 13.9 µm 0.8 mm Temnopleurus toreumaticus ZMB Ech 2802 13.9 µm 0.7 mm Trigonocidaridae Trigonocidaris albida ZSM 20012468 10.0 µm 0.3 mm Echinoneidae Echinoneus cyclostomus MCZ 2747 5.5 µm 0.1 mm Apatopygidae Apatopygus recens ZMK Mortensen collection 6.5 µm 0.2 mm Echinolampadidae Echinolampas depressa MCZ 8990 3.3 µm 0.2 mm Clypeasteridae Clypeaster reticulatus USNM 34282 13.9 µm 0.6 mm Laganidae Laganum joubini BMNH 1979.1.25.52-60 13.9 µm 0.6 mm Echinarachniidae Echinarachnius parma ZSM 20011676 13.9 µm 0.3 mm Astriclypeidae Astriclypeus manni MCZ 7300 36.5 µm 0.6 mm
Proc. of SPIE Vol. 8506 85061G-3
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
2.2 Synchrotron micro-computed tomography
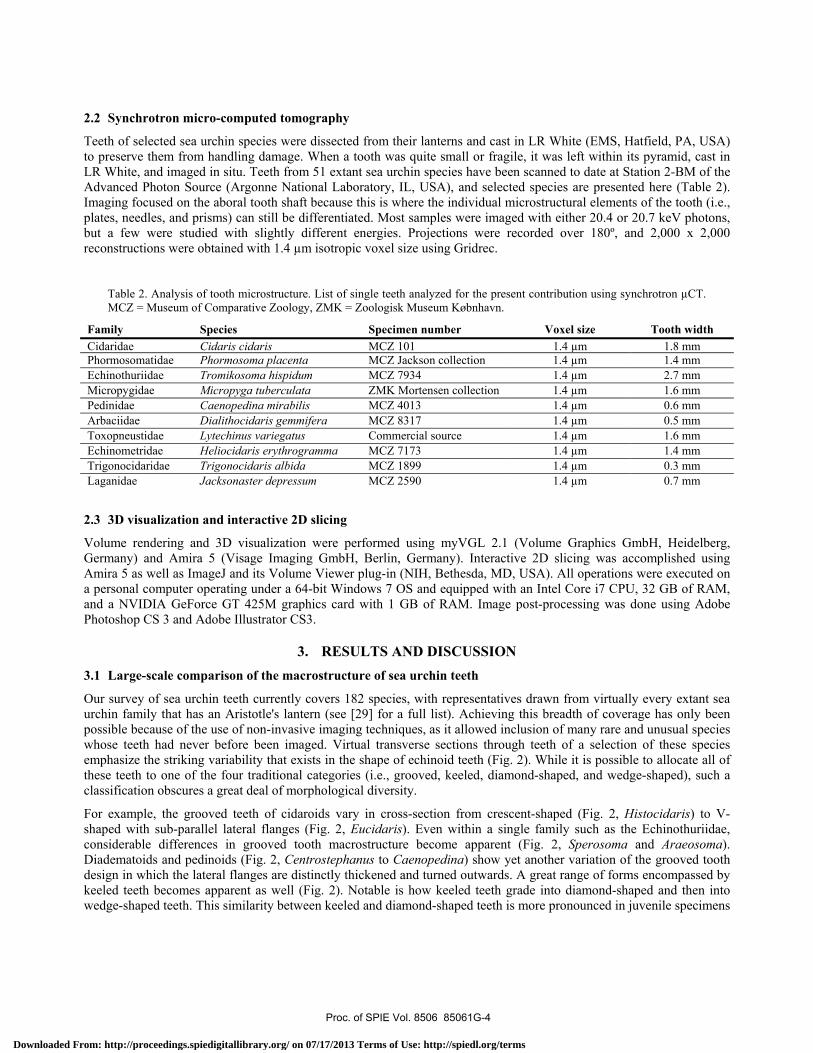
Teeth of selected sea urchin species were dissected from their lanterns and cast in LR White (EMS, Hatfield, PA, USA) to preserve them from handling damage. When a tooth was quite small or fragile, it was left within its pyramid, cast in LR White, and imaged in situ. Teeth from 51 extant sea urchin species have been scanned to date at Station 2-BM of the Advanced Photon Source (Argonne National Laboratory, IL, USA), and selected species are presented here (Table 2). Imaging focused on the aboral tooth shaft because this is where the individual microstructural elements of the tooth (i.e., plates, needles, and prisms) can still be differentiated. Most samples were imaged with either 20.4 or 20.7 keV photons, but a few were studied with slightly different energies. Projections were recorded over 180º, and 2,000 x 2,000 reconstructions were obtained with 1.4 µm isotropic voxel size using Gridrec.
Table 2. Analysis of tooth microstructure. List of single teeth analyzed for the present contribution using synchrotron µCT. MCZ = Museum of Comparative Zoology, ZMK = Zoologisk Museum Købnhavn.
Family Species Specimen number Voxel size Tooth width Cidaridae Cidaris cidaris MCZ 101 1.4 µm 1.8 mmPhormosomatidae Phormosoma placenta MCZ Jackson collection 1.4 µm 1.4 mm Echinothuriidae Tromikosoma hispidum MCZ 7934 1.4 µm 2.7 mm Micropygidae Micropyga tuberculata ZMK Mortensen collection 1.4 µm 1.6 mm Pedinidae Caenopedina mirabilis MCZ 4013 1.4 µm 0.6 mm Arbaciidae Dialithocidaris gemmifera MCZ 8317 1.4 µm 0.5 mm Toxopneustidae Lytechinus variegatus Commercial source 1.4 µm 1.6 mm Echinometridae Heliocidaris erythrogramma MCZ 7173 1.4 µm 1.4 mm Trigonocidaridae Trigonocidaris albida MCZ 1899 1.4 µm 0.3 mm Laganidae Jacksonaster depressum MCZ 2590 1.4 µm 0.7 mm
2.3 3D visualization and interactive 2D slicing
Volume rendering and 3D visualization were performed using myVGL 2.1 (Volume Graphics GmbH, Heidelberg, Germany) and Amira 5 (Visage Imaging GmbH, Berlin, Germany). Interactive 2D slicing was accomplished using Amira 5 as well as ImageJ and its Volume Viewer plug-in (NIH, Bethesda, MD, USA). All operations were executed on a personal computer operating under a 64-bit Windows 7 OS and equipped with an Intel Core i7 CPU, 32 GB of RAM, and a NVIDIA GeForce GT 425M graphics card with 1 GB of RAM. Image post-processing was done using Adobe Photoshop CS 3 and Adobe Illustrator CS3.
3. RESULTS AND DISCUSSION 3.1 Large-scale comparison of the macrostructure of sea urchin teeth
Our survey of sea urchin teeth currently covers 182 species, with representatives drawn from virtually every extant sea urchin family that has an Aristotle's lantern (see [29] for a full list). Achieving this breadth of coverage has only been possible because of the use of non-invasive imaging techniques, as it allowed inclusion of many rare and unusual species whose teeth had never before been imaged. Virtual transverse sections through teeth of a selection of these species emphasize the striking variability that exists in the shape of echinoid teeth (Fig. 2). While it is possible to allocate all of these teeth to one of the four traditional categories (i.e., grooved, keeled, diamond-shaped, and wedge-shaped), such a classification obscures a great deal of morphological diversity.
For example, the grooved teeth of cidaroids vary in cross-section from crescent-shaped (Fig. 2, Histocidaris) to V-shaped with sub-parallel lateral flanges (Fig. 2, Eucidaris). Even within a single family such as the Echinothuriidae, considerable differences in grooved tooth macrostructure become apparent (Fig. 2, Sperosoma and Araeosoma). Diadematoids and pedinoids (Fig. 2, Centrostephanus to Caenopedina) show yet another variation of the grooved tooth design in which the lateral flanges are distinctly thickened and turned outwards. A great range of forms encompassed by keeled teeth becomes apparent as well (Fig. 2). Notable is how keeled teeth grade into diamond-shaped and then into wedge-shaped teeth. This similarity between keeled and diamond-shaped teeth is more pronounced in juvenile specimens
Proc. of SPIE Vol. 8506 85061G-4
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
\I/V%1 Nir Ner NileHistocidaris Cidaris Eucidaris Phormosoma Sperosoma Araeosoma
(data not shown) and is evidence for a sister-group relationship uniting the so-called "regular" echinoids with keeled teeth and the irregular echinoids with their diamond- or wedge-shaped teeth.
The origin of keeled teeth has long remained a mystery, as no intermediate shapes are known that bridge the gap between grooved and keeled teeth. Furthermore, molecular and morphological studies have failed to resolve the immediate sister group to echinoids with keeled teeth with any degree of confidence [30]. Interestingly, amongst the species with grooved teeth shown here, there is one that has a dense keel-like axial growth zone at its core (Fig. 2, Micropyga). Differences in density within the tooth point to significant differences in the structural arrangement of plates and prisms, which is confirmed by synchrotron µCT analysis (see section 3.3). In the case of Micropyga, the dense core would seem to be a prismatic outgrowth - a significant finding, as such an outgrowth of prisms also forms the core of the keel in keeled teeth. If this inference is correct, it may provide key evidence in resolving Micropygidae as sister group to echinoids with keeled teeth.
Figure 2. High degree of macrostructural diversity of sea urchin teeth revealed by µCT. These selected virtual transverse sections illustrate a greater diversity of sea urchin tooth shapes than previously reported. Table 1 provides further specimen and dataset information. Not to scale.
Although the traditional four-fold terminology for echinoid tooth shapes will still be useful at an informal level, a full understanding of sea urchin tooth evolution will clearly benefit from a more rigorous (i.e., numerical) approach to
Proc. of SPIE Vol. 8506 85061G-5
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
defining tooth macrostructure as well as a better knowledge of tooth microstructure gained through synchrotron µCT imaging. This work is currently underway (see sections 3.3, 4.1, and 4.2).
3.2 Discovery of unusual tooth shapes in sea urchins
Of particular interest both from a phylogenetic as well as a biomechanical point of view is the discovery of aberrant tooth shapes in several distantly related sea urchin groups. We here provide a number of examples that further illustrate the usefulness of a large-scale exploratory approach.
As outlined above, cidaroid teeth come in various forms from crescent-shaped to V-shaped structures (Fig. 2, Histocidaris to Eucidaris). Fossil specimens show that an open, crescent-shaped tooth is the primitive condition for echinoids [12]. However, some cidaroid teeth have a broad, almost cubic tooth shape (Fig. 3A), and this shape difference must have an impact on the mechanical properties of the tooth. In contrast to the predominantly shallow-water cidaroids, aspidodiadematids are a small group of deep-sea sea urchins related to micropygids and diadematids (Fig. 2, Micropyga to Diadema). Using µCT, the highly asymmetrical tooth shape of a single aspidodiadematid species, Plesiodiadema indicum, had recently been reported [31]. We included additional aspidodiadematid species in the present study in order to verify this unexpected finding. Our data suggest that all aspidodiadematids possess teeth with varying degrees of asymmetry (Fig. 3B). The functional significance of this asymmetry is unknown.
Figure 3. Unusual tooth shapes among sea urchins revealed by µCT. (A) Broad, almost cubic cidaroid teeth: Chondrocidaris gigantea and Compsocidaris pyrsacantha. Left and right, respectively, here and below. (B) Asymmetrical aspidodiadematid teeth: Aspidodiadema arcitum and Plesiodiadema indicum. (C) Rhombic salenioid teeth: Salenocidaris hastigera and Salenocidaris varispina. (D) Rhombic arbacioid teeth: Pygmaeocidaris prionigera and Dialithocidaris gemmifera. (E) Anvil-shaped echinometrid teeth: Heliocidaris erythrogramma and Evechinus chloroticus. (F) Wide-flanged temnopleurid teeth with truncated keel: Amblypneustes pallidus and Holopneustes inflatus. Table 1 provides further specimen and dataset information. Not to scale.
As stirodont taxa, salenioids and arbacioids typically possess keeled teeth (Fig. 2, Salenia and Arbacia). Interestingly, our data reveal that some species within these two groups possess rhombic teeth as adults (Fig. 3C, D). Whether this rhombic shape is a consequence of the sometimes minute size of these animals, and therefore a likely retention of juvenile characters (pedomorphism), will require further study. Contrary to stirodonts, the teeth of camarodont sea urchin species such as Paracentrotus lividus or Lytechinus variegatus have been extensively studied and are considered the archetype of the keeled tooth (Fig. 2, Parasalenia to Strongylocentrotus). Relatively little macrostructural variation was thought to occur among camarodont teeth [2, 5, 14, 27]. However, we found the teeth of two closely related camarodont genera, Heliocidaris and Evechinus, to possess an anvil-shaped keel of yet unknown function (Fig. 3E). This shape had recently been reported for further specimens of Evechinus [32]. Finally, the teeth of most temnopleurids are T-shaped in form with a rounded aspect and a widened keel (Fig. 2, Temnopleurus). In contrast to this prevalent shape among temnopleurids, the teeth of two derived genera, Amblypneustes and Holopneustes, possess a truncated keel and widened lateral flanges (Fig. 3F). The functional significance of this particular shape remains yet to be studied.
Proc. of SPIE Vol. 8506 85061G-6
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
3.3 Large-scale comparison of the microstructure of sea urchin teeth
In broad terms, the biomineralization of sea urchin teeth consists of two stages. The first-stage calcite forms in well-defined syncytial spaces, has a composition of Ca1-xMgxCO3 with 0.1<x<0.15 (i.e., high Mg calcite), and consists of different types of plates (primary plates, secondary plates, and carinar process plates in some species) as well as a transitional lamellar needle complex and prisms [1-4, 15]. Each plate or prism is part of a parallel array and is separate from its neighbors until the second-stage mineral - also calcite, but with 0.3<x<0.4 (i.e., very high Mg calcite) - forms and cements the neighbors together. All of these structures have one or more minimum dimensions and spacings ranging from 1-25 µm. µCT scans with voxel size >5 µm (Figs. 1-3) can resolve regions with low mineral density, but not the organization of individual structures within the different tooth regions. In contrast, as a comparison of teeth shown in Figs. 2 and 4 reveals, synchrotron µCT with 1.4 µm isotropic voxels can resolve tooth microstructure. Although some synchrotron µCT experiments on sea urchin teeth had been performed previously [8, 24, 33], this imaging modality has so far not been exploited to systematically quantify tooth microstructure across echinoid species.
Figure 4. High degree of microstructural diversity of selected sea urchin teeth revealed by synchrotron µCT. (A) Cidaris cidaris (Cidaridae), (B) Phormosoma placenta (Phormosomatidae), (C) Tromikosoma hispidum (Echinothuriidae), (D) Micropyga tuberculata (Micropygidae), (E) Caenopedina mirabilis (Pedinidae), (F) Dialithocidaris gemmifera (Arbaciidae), (G) Lytechinus variegatus (Toxopneustidae), (H) Heliocidaris erythrogramma (Echinometridae), (I) Trigonocidaris albida (Trigonocidaridae), and (J) Jacksonaster depressum (Laganidae). Table 2 provides further specimen and dataset information. Not to scale.
Virtual transverse sections from teeth of selected species are shown in Fig. 4. The microstructure found within the teeth of 51 species imaged with synchrotron µCT to date varies considerably and does not always reflect the macrostructural pattern observed for the respective species. For example, despite their similar crescent-shaped macrostructure, the teeth
Proc. of SPIE Vol. 8506 85061G-7
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
of Araeosoma (Fig. 4C) and Micropyga (Fig. 4D) possess quite different plate morphologies in their adaxial part. The plates in the specimen shown in Fig. 4C are short and thin and tend to lie end-to-end along the abaxial-adaxial axis. In the specimen shown in Fig. 4D, the plates curve considerably and are much longer, while in addition three open zones (termed fenestrae) are present in each lateral flange of the tooth. A central prism zone can be discerned between the two medial fenestrae of this specimen, but no such prisms are visible in the specimen shown in Fig. 4C.
Tooth microstructure can be imaged at similar or even better resolution using optical or scanning electron microscopy, and structures in different species have previously been described in detail using these techniques [15-21]. In contrast, synchrotron µCT offers simple and efficient access to the third dimension. Although serial polishing of mounted samples followed by scanning electron microscopy or optical imaging can also provide access to the third dimension, this approach is so labor-intensive and fraught with other difficulties that it is rarely employed. The possibility of visualizing the complex internal microstructure of sea urchin teeth in 3D is one significant advantage of synchrotron µCT datasets (see section 3.4), but equally important is the ability to quantify, for example, prism trajectories, plate thicknesses, or distribution of a plate’s surface normals relative to the tooth axis. Such quantities may prove to be important numerical characters for phylogenetic inferences and are certainly important for an understanding of the biomechanics of sea urchin teeth, both qualitatively and quantitatively. With the availability of 3D datasets of entire teeth, our understanding of tooth mineralization across species will improve, for example, through a comparison of the amount of second-stage mineral within selected single transverse sections or even along the length of the entire tooth across species.
3.4 2D and 3D visualization of the complex microstructure of sea urchin teeth
Because an isotropic voxel size <2 µm is usually necessary in order to reveal the microstructure of sea urchin teeth, datasets derived from synchrotron µCT experiments were employed for 3D visualization and interactive 2D slicing. The HDF-4 image files were loaded in ImageJ, cropped, and converted to 8-bit contrast. The Volume Viewer plug-in was used to obtain oblique virtual sections through the volume (Fig. 5A, B). 3D visualization was performed by loading the transformed image stacks into Amira (Fig. 5C). Using a corner-cut view, internal elements of the volume were revealed as well (Fig. 5D). Segmentation and isolation of individual structures are key advantages of tomographic datasets (see, e.g., [31, 34]). In addition to permitting finite element modeling (FEM) studies, this approach allows for a better communication of complex structures such as the tooth's primary plates, prisms, and needles as fully interactive 3D models using, for example, 3D PDF technology [35].
Figure 5. 2D and 3D visualization of a synchrotron µCT dataset of part of the tooth of Caenopedina mirabilis (Pedinidae). (A) Virtual transverse section. (B) Virtual longitudinal section along the axis indicated in (A). (C) Volume rendering of an image stack composed of 300 virtual sections. Oblique axial view. (D) Same view as in the previous image, but with part of the tooth virtually removed. Table 2 provides further specimen and dataset information. Not to scale.
Proc. of SPIE Vol. 8506 85061G-8
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
V VvYV vV V
uW
vuA
ae
4 4440
4
0
. ., 11 ..
1 41,
b
4. OUTLOOK 4.1 Application of shape analysis algorithms to sea urchin teeth
As shown above, the traditional four-fold subdivision into keeled, grooved, diamond-shaped, and wedge-shaped teeth only captures a portion of the existing variation, and much information would be lost if this classification were further applied. A numerical approach using shape analysis algorithms circumvents the use of word-based descriptions and provides an unbiased, reproducible characterization of tooth shape.
After binary segmentation, transverse sections of selected sea urchin teeth were smoothed using erosion, dilation, and hole-filling operators and reduced to 64 x 64 pixel dimensions in ImageJ. Using the whole images as the inputs, an image vector with 64 x 64 binary pixel values as entries from the images was calculated by concatenating all images into one vector with a length of 4096 pixels. The 4096-dimensional image feature vector was then compressed using a principal component analysis (PCA). Out of the 4096 features, only about 2% (i.e., 80) were informative. Among these 2%, the first 10-20 features contained nearly all of the information present in the dataset. Pair-wise scatter plots of the first four PCA features were used to differentiate the groupings of tooth shapes. Finally, a projection of the transverse sections through this four-component space was calculated using hierarchical clustering methods (Fig. 6). Our metric illustrates how the different tooth shapes group in low-dimensional space. Further improvements of the analysis are bound to resolve the more subtle shape differences within the present groupings.
Figure 6. Low-dimensional projection of transverse binarized virtual sections of sea urchin teeth obtained by µCT. Using a non-linear measure, the distances between tooth shapes were calculated and teeth were grouped according to their anatomy. Grooved (A) and wedge-shaped (B) teeth cluster together, but diamond-shaped teeth (C = Echinolampas and Apatopygus + Echinoneus) group with keeled teeth. However, consideration of the third principal component dimension (data not shown) reveals that this present discrepancy is not actually present.
Proc. of SPIE Vol. 8506 85061G-9
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
4.2 Advanced analysis of sea urchin tooth microstructure
Sea urchin teeth are very complex 3D structures, and section 3.3 lists prism trajectories, plate thicknesses, and the distribution of a plate’s surface normals relative to the tooth axis as examples of quantities that could be used on the one hand to trace phylogenetic relationships and on the other to better understand tooth mechanics. In analogy to the shape descriptors for tooth macrostructure proposed in section 4.1, it should be possible to develop numerical shape descriptors for microstructural elements within sea urchin teeth. Because calcite is a wretched structural material, the high degree of functionality of sea urchin teeth - which exceeds that of inorganic calcite crystals - is of extraordinary interest.
From a materials science perspective, the various tooth shapes represent a sampling of the design space for calcite-based structures that require a measure of resistance to bending moments and to wear. Understanding systems with dozens of parameters is intrinsically difficult, but evolution obviously has selected combinations that are effective. Many materials scientists, however, assume that these and other natural structures are optimized for function and base their studies on this. Such a perspective may be useful in focusing analysis, but it misses the point that the existing solutions of the design problem are not the endpoints of evolution - and may even represent evolutionary accidents that are good enough to survive or have been preserved for other reasons. Understanding complex materials systems as compromises between many competing requirements is still a discipline in its infancy, but 3D quantification as proposed here will add much to this endeavor.
From a structural engineering perspective, 3D datasets of sea urchin teeth with a voxel size <2 µm allow the microstructure to be imported into FEM analyses. The effect of variations of microstructure can then be simulated: the performance of intermediates between structures in extant species can be compared virtually with those of the endpoints.
From a biomineralization perspective, sea urchin teeth are important models in part because each tooth contains all stages of the mineralization process. That is not to say that tooth macro- and microstructure are identical between juveniles and adults, but rather that mineral is added continuously to replace material worn from the tip and develops over time into the mature structures. Use of 3D datasets of entire teeth allows quantification of the mineral added as the tooth matures, both the material's location and its precise amount. These data will address outstanding fundamental questions of what triggers the formation of very high Mg calcite between plates and prisms and of why the composition changes from high to very high Mg calcite. As higher resolution µCT and nanoCT as well as techniques such as local tomographic reconstruction are applied, in situ interrogation of intact teeth will reveal much more about internal structures and the underlying mineralization process.
4.3 Impact of µCT data on our understanding of sea urchin biology
There is no doubt that large-scale surveys across a wide spectrum of species are the best way to provide a broad perspective on the evolution of form and function. Using non-invasive imaging techniques for such surveys has two major advantages over previous approaches: (i) they allow the wealth of biodiversity held in museum collections to be sampled, and (ii) they allow for rigorous quantification of shape differences and structural organization through the availability of purely digital data. Because virtual sections of sea urchin teeth can be generated at standardized orientations, morphometric techniques can now be applied to these datasets, allowing similarity to be measured objectively for the first time. This will allow a move away from the simplistic four-fold subdivision of echinoid tooth shapes that is currently employed to document and understand the existing diversity. Direct comparisons can now be made of transverse sections at standardized positions along the length of the tooth, providing insight into how shape and skeletal organization change during growth. This should give important insight into the evolution of tooth form, highlighting for example the similarities that link diamond-shaped and keeled teeth. By optimizing the tooth data onto firmly established phylogenetic trees derived from morphological as well as molecular datasets [30, 36], the evolution of tooth design will be determined in greater detail than previously possible.
All the work reported here pertains to extant species, but a full history of tooth evolution requires the incorporation of fossil data. While tomographic imaging of fossil echinoid microstructure is possible [37], suitable fossil specimens with preserved lanterns will always remain rare. There is, however, a substantial amount of literature describing teeth of extant and fossil sea urchin species [5, 12, 14, 22]. While this approach does not provide data that would match tomographic imagery in its precision, it should still be possible to integrate this information with high-quality morphometric data. The focus of our studies is documenting tooth shape at the macro- and microstructural level, but there is also a strong biomechanical control to tooth structure that clearly constrains the way in which the animal can
Proc. of SPIE Vol. 8506 85061G-10
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
feed. It has long been recognized that keeled teeth are biomechanically superior to grooved teeth [3, 25], an observation that correlates with differences in diet between the carnivorous cidaroids with their grooved teeth and the algae-grazing euechinoids with their keeled teeth [5]. However, the present survey has highlighted that there are also striking structural differences that exist amongst species with grooved teeth as well as amongst those with keeled teeth. A better understanding of the biomechanical properties of the different tooth shapes will undoubtedly help to shed new light on echinoid ecology and the evolution of feeding strategies.
Finally, while sea urchin teeth are fascinatingly complex structures to try to understand, there are many other aspects of the morphology of Aristotle's lantern that would benefit from a similar large-scale, non-invasive imaging approach. For example, using magnetic resonance imaging, Ziegler et al. [38] have recently demonstrated that there is important variation in lantern muscle organization among sea urchins, which has phylogenetic and functional implications. The demipyramids of Aristotle's lantern in Lytechinus variegatus have been studied using µCT at resolutions similar to those shown in Figs. 1-3, and differences in attenuation with position were related to local calcite volume fraction [39]. Furthermore, a demipyramid was successfully studied using synchrotron µCT [40], and one expects that phylogenetic studies employing local tomography techniques will add important information. Further lantern soft and hard parts will no doubt be studied in the same comprehensive manner, but a comparative study of lantern biomechanics integrating all of the different skeletal elements and muscles must surely be the ultimate goal. Any such future work can build on the pioneering biomechanical work performed in the last decades [41-46]. Only by integrating the various strands of information will we truly start to understand the evolution of the complex echinoid lantern and how its form and function may have directed echinoid ecology and behavior.
ACKNOWLEDGEMENTS
We are grateful to F. Kosar and M. Ogurreck for their assistance with µCT. X. Xiao is acknowledged for his help with synchrotron µCT. A. Baldinger, A. Cabrinovic, T. Ebert, D. Eibye-Jacobsen, C. Lüter, R. Mooi, D. Pawson, B. Ruthensteiner, and A. Schmidt-Rhaesa provided us with specimens from their collections. We would like to thank F. Beckmann and G. Giribet for their generous support of our µCT scanning experiments. SRS acknowledges support from the National Institutes of Health through NIDCR Grant No. DE-001374 (to Arthur Veis). AZ was supported by the Deutsche Forschungsgemeinschaft through Grant No. ZI-1274/1-1. Use of the Advanced Photon Source was made possible by the U.S. Department of Energy, Office of Science, Office of Basic Energy Sciences under Contract No. DE-AC02-06CH11357. This work was performed in part at the Center for Nanoscale Systems, a member of the National Nanotechnology Infrastructure Network, which is supported by the National Science Foundation under Award No. ECS-0335765.
REFERENCES
[1] Salter, S. J. A., "On the structure and growth of the tooth of Echinus," Phil. Trans. R. Soc. Lond. 151, 387-407 (1861).
[2] Giesbrecht, W., "Der feinere Bau der Seeigelzähne," Morph. Jb. 6, 79-105 (1880). [3] Märkel, K., Gorny, P., and Abraham, K., "Microarchitecture of sea urchin teeth," Fortschr. Zool. 24, 103-114
(1977). [4] Jensen, M., "Primary plates of sea urchin teeth (Echinoidea)," Vidensk. Meddr. Dansk Naturh. Foren. 141, 7-27
(1979). [5] Smith, A. B., "Implications of lantern morphology for the phylogeny of post-Palaeozoic echinoids," Palaeontology
24, 779-801 (1981). [6] Wang, R. Z., Addadi, L., and Weiner, S., "Design strategies of sea urchin teeth: structure, composition and
micromechanical relations to function," Phil. Trans. R. Soc. Lond. B 352, 469-480 (1997). [7] Stock, S. R., Barss, J., Dahl, T., Veis, A., and Aimer, J. D., "X-ray absorption microtomography (microCT) and
small beam diffraction mapping of sea urchin teeth," J. Struct. Biol. 139, 1–12 (2002). [8] Stock, S. R., Ignatiev, K. I., Dahl, T., Veis, A., and De Carlo, F., "Three-dimensional microarchitecture of the plates
(primary, secondary and carinar process) in the developing tooth of Lytechinus variegatus revealed by synchrotron X-ray absorption microtomography (microCT)," J. Struct. Biol. 144, 282-300 (2003).
Proc. of SPIE Vol. 8506 85061G-11
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
[9] Robach, J. S., Stock, S. R., and Veis, A. "Transmission electron microscopy characterization of macromolecular domain cavities and microstructure of single-crystal calcite tooth plates of the sea urchin Lytechinus variegatus," J. Struct. Biol. 151, 18-29 (2005).
[10] Ma, Y., Cohen, S. R., Addadi, L., and Weiner, S., "Sea urchin tooth design: an 'all-calcite' polycrystalline reinforced fiber composite for grinding rocks," Adv. Mater. 20, 1555-1559 (2008).
[11] Killian, C. E., Metzler, R. A., Gong, Y. U. T., Olson, I. C., Aizenberg, J. A., Politi, Y., Wilt, F. H., Scholl, A., Young, A., Doran, A., Kunz, M., Tamura, N., Coppersmith, S. N., and Gilbert, P. U. P. A., "Mechanism of calcite co-orientation in the sea urchin tooth," J. Am. Chem. Soc. 131, 18404-18409 (2009).
[12] Reich, M., and Smith. A. B., "Origins and biomechanical evolution of teeth in echinoids and their relatives," Palaeontology 52, 1149-1168 (2009).
[13] Killian, C. E., Metzler, R. A., Gong, Y., Churchill, T. H., Olson, I. C., Trubetskoy, V., Christensen, M. B. Fournelle, J. H., De Carlo, F., Cohen, S., Mahamid, J., Scholl, A., Young, A., Doran, A., Wilt, F. H., Coppersmith, S. N., and Gilbert, P. U. P. A., "Self-sharpening mechanism of the sea urchin tooth," Adv. Funct. Mater. 21, 682-690 (2011).
[14] Jackson, R. T., "Phylogeny of the Echini, with a revision of Palaeozoic species," Mem. Boston Soc. Nat. Hist. 7, 1-491 (1912).
[15] Jensen, M., "Morphology and classification of Euechinoidea Bronn, 1860 - a cladistic analysis," Vidensk. Meddr. Dansk Naturh. Foren. 143, 7-99 (1981).
[16] Märkel, K., and Titschack, H., "Morphologie der Seeigelzähne. I. Der Zahn von Stylocidaris affinis (Phil.) (Echinodermata, Echinoidea)," Z. Morph. Tiere 64, 179-200 (1969).
[17] Märkel, K., "Morphologie der Seeigelzähne. II. Die gekielten Zähne der Echinacea (Echinodermata, Echinoidea)," Z. Morph. Tiere 66, 1-50 (1969).
[18] Märkel, K., "Morphologie der Seeigelzähne. III. Die Zähne der Diadematoida und Echinothuroida (Echinodermata, Echinoidea)," Z. Morph. Tiere 66, 189-211 (1970).
[19] Märkel, K., "Morphologie der Seeigelzähne. IV. Die Zähne von Laganum und Clypeaster (Echinodermata, Echinoidea)," Z. Morph. Tiere 68, 370-389 (1970).
[20] Märkel, K., "The tooth skeleton of Echinometra mathaei (Blainville) (Echinodermata, Echinoidea)," Annot. Zool. Japon. 43, 188-199 (1970).
[21] Märkel, K., "Morphologie der Seeigelzähne. V. Die Zähne der Clypeasteroida (Echinodermata, Echinoidea)," Z. Morph. Tiere 78, 221-256 (1974).
[22] Märkel, K., "On the teeth of the recent cassiduloid Echinolampas depressa Gray, and on some Liassic fossil teeth nearly identical in structure (Echinodermata, Echinoidea)," Zoomorphology 89, 125-144 (1978).
[23] Kniprath, E., "Ultrastructure and growth of the sea urchin tooth," Calc. Tiss. Res. 14, 211-228 (1974). [24] Stock, S. R., Barss, J., Dahl, T., Veis, A., Almer, J. D., and De Carlo, F., "Synchrotron X-ray studies of the keel of
the short-spined sea urchin Lytechinus variegatus: absorption microtomography (microCT) and small beam diffraction mapping," Calcif. Tissue Int. 72, 555-566 (2003).
[25] Märkel, K., and Gorny, P., "Zur funktionellen Anatomie der Seeigelzähne (Echinodermata, Echinoidea)," Z. Morph. Tiere 75, 223-242 (1973).
[26] Jensen, M., "The Strongylocentrotidae (Echinoidea), a morphologic and systematic study," Sarsia 57, 113-148 (1974).
[27] Smith, A. B., [Echinoid palaeobiology], George Allen & Unwin, London, 190 pp. (1984). [28] Ziegler, A., and Menze, B.H., "Accelerated acquisition, visualization, and analysis of zoo-anatomical data,"
Computation for Humanity, CRC Press, Boca Raton (in press). [29] Ziegler, A., "Broad application of non-invasive imaging techniques to echinoids and other echinoderm taxa," Proc.
7th ECE, Springer, Heidelberg (in press). [30] Kroh, A., and Smith, A. B., "The phylogeny and classification of post-Palaeozoic echinoids," J. Syst. Pal. 8, 147-212
(2010). [31] Ziegler, A., Ogurreck, M., Steinke, T., Beckmann, F., Prohaska, S., and Ziegler, A., "Opportunities and challenges
for digital morphology," Biol. Direct 5, 45 (2010). [32] Laxminarayana, R., Rodrigues, S., and Dickinson, M., "Characterization of E. chloroticus sea urchin tooth using
nanoindentation and SEM," Intern. J. Mod. Phys. Conf. Ser. 6, 705-708 (2012). [33] Stock, S. R., Ignatiev, K. I., Veis, A., De Carlo, F., and Almer, J. D., "MicroCT of sea urchin ossicles supplemented
with microbeam diffraction,” Proc. SPIE 5535, 11-20 (2004). [34] Ziegler, A., Faber, C., Mueller, S., and Bartolomaeus, T., "Systematic comparison and reconstruction of sea urchin
(Echinoidea) internal anatomy: a novel approach using magnetic resonance imaging," BMC Biol. 6, 33 (2008).
Proc. of SPIE Vol. 8506 85061G-12
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms
[35] Ziegler, A., Mietchen, D., Faber, C., von Hausen, W., Schöbel, C., Sellerer, M., and Ziegler, A., "Effectively incorporating selected multimedia content into medical publications," BMC Med. 9, 17 (2011).
[36] Smith, A. B., Pisani, D., Mackenzie-Dodds, J. A., Stockley, B., Webster, B. L., and Littlewood, D. T. J., "Testing the molecular clock: molecular and paleontological estimates of divergence times in the Echinoidea (Echinodermata)," Mol. Biol. Evol. 23, 1832-1851 (2006).
[37] Stock, S. R., and Veis, A., "Preliminary microfocus X-ray computed tomography survey of echinoid fossil microstructure," Geol. Soc. Lond. Spec. Publ. 215, 225-235 (2003).
[38] Ziegler, A., Schröder, L., Ogurreck, M., Faber, C., and Stach, T., "Evolution of a novel muscle design in sea urchins (Echinodermata: Echinoidea)," PLoS ONE 7, e37520 (2012).
[39] Stock, S. R., Nagaraja, S., Barss, T., Dahl, T., and Veis, A., "X-ray microCT study of pyramids of the sea urchin Lytechinus variegatus," J. Struct. Biol. 141, 9-21 (2003).
[40] Stock, S. R., Ignatiev, K., De Carlo, F., "Very high resolution synchrotron microCT of sea urchin ossicle structure," Echinoderms: München, Balkema, Leiden, 353-358 (2004).
[41] Andrietti, F., Candia Carnevali, M. D., Wilkie, I. C., Lanzavecchia, G., Melone, G., and Celentano, F. C., "Mechanical analysis of the sea-urchin lantern: the overall system in Paracentrotus lividus," J. Zool. 220, 345-366 (1990).
[42] Andrietti, F., Candia Carnevali, M. D., and Wilkie, I. C., "Mechanical analysis of the lantern movement in the cidarid sea-urchin Stylocidaris affinis (Phil.): comparison with a typical camarodont lantern," Echinoderm Research 1991, Balkema, Rotterdam, 95-98 (1992).
[43] Bonasoro, F., Candia Carnevali, M. D., Wilkie, I. C., "The peristomial membrane of regular sea-urchins: functional morphology of the epidermis and coleomic lining in Paracentrotus lividus (Lamarck)," Boll. Zool. 62, 121-135 (1995).
[44] Candia Carnevali, M. D., Lanzavecchia, G., Melone, G., Celentano, F. C., and Andrietti, F., "Aristotle's lantern in the regular sea urchin Paracentrotus lividus. II. Biomechanical approach to the interpretation of movement," Echinoderm Biology, Balkema, Rotterdam, 663-672 (1988).
[45] Lanzavecchia, G., Candia Carnevali, M. D., Melone, G., Celentano, F. C., and Andrietti, F., "Aristotle's lantern in the regular sea urchin Paracentrotus lividus. I. Functional morphology and significance of bones, muscles and ligaments," Echinoderm Biology, Balkema, Rotterdam, 649-662 (1998).
[46] Wilkie, I. C., Candia Carnevali, M. D., and Andrietti, F., "Mechanical properties of sea-urchin lantern muscles: a comparative investigation of intact muscle groups in Paracentrotus lividus (Lam.) and Stylocidaris affinis (Phil.) (Echinodermata, Echinoidea)," J Comp. Physiol. B 168, 204-212 (1998).
Proc. of SPIE Vol. 8506 85061G-13
Downloaded From: http://proceedings.spiedigitallibrary.org/ on 07/17/2013 Terms of Use: http://spiedl.org/terms