REGULAR ARTICLE Maize seedling phosphorus nutrition: Allocation of remobilized seed phosphorus reserves and external phosphorus uptake to seedling roots and shoots during early growth stages Muhammad Nadeem & Alain Mollier & Christian Morel & Muhammad Shahid & Muhammad Aslam & Muhammad Zia-ur-Rehman & Muhammad Ashfaq Wahid & Sylvain Pellerin Received: 5 December 2012 / Accepted: 13 March 2013 # Springer Science+Business Media Dordrecht 2013 Abstract Background and aims The growth of green plants depends not only on photosynthesis, but also on the successful remobilization and translocation of seed phos- phorus (P) reserves to the vegetative parts of the devel- oping seedling during early growth. Remobilization and photosynthesis are therefore two parallel and co- coinciding processes involved in better seedling establishment and early growth. Methods A study was conducted to evaluate the pri- ority of developing maize seedlings to translocate the remobilized seed P reserves and external P uptake to seedling root and shoot sinks during 4 weeks of early growth. Two fluxes of P in growing seedlings, one from seed remobilized P reserves and one from exter- nal P uptake, were distinguished by labelling external nutrient solution P with 32 P. Results The seedling phytomass was equally distrib- uted between seedling roots and shoots for 530 cumu- lated degree days after sowing. Seedlings partitioned up to 71 % of P from seed reserves and up to 68 % of P acquired from the nutrient solution, to the shoots, depending on the seed P content and P concentration in the nutrient solution. It appears that accumulation of P slows down in seedling roots corresponds to the translocative functions of root P towards shoots for start of photosynthesis. Conclusions Our results suggest that the major part of seed P reserves and external P uptake were used in early development of the seedling and the preferred sink was seedling shoots. Keywords Maize . Seed . Phosphorus . Phytomass . Isotope . P uptake . Early growth Plant Soil DOI 10.1007/s11104-013-1689-x Responsible Editor: Philip John White. M. Nadeem : A. Mollier : C. Morel : S. Pellerin INRA, UMR 1220 TCEM, 71 avenue Bourlaux, CS 20032, 33882 Villenave d’Ornon Cedex, France M. Nadeem : A. Mollier : C. Morel : S. Pellerin Bordeaux Sciences Agro, UMR 1220 TCEM, 33175 Gradignan, France M. Nadeem (*) : M. Shahid : M. Aslam Department of Environmental Sciences, COMSATS Institute of Information Technology, Vehari 61100, Pakistan e-mail: [email protected]M. Zia-ur-Rehman Institute of Soil and Environmental Sciences, University of Agriculture, Faisalabad 38000, Pakistan M. A. Wahid Department of Agronomy, University of Agriculture, Faisalabad 38000, Pakistan
Transcript
REGULAR ARTICLE
Maize seedling phosphorus nutrition: Allocationof remobilized seed phosphorus reserves and externalphosphorus uptake to seedling roots and shootsduring early growth stages
Muhammad Nadeem & Alain Mollier &
Christian Morel & Muhammad Shahid &
Muhammad Aslam & Muhammad Zia-ur-Rehman &
Muhammad Ashfaq Wahid & Sylvain Pellerin
Received: 5 December 2012 /Accepted: 13 March 2013# Springer Science+Business Media Dordrecht 2013
AbstractBackground and aims The growth of green plantsdepends not only on photosynthesis, but also on thesuccessful remobilization and translocation of seed phos-phorus (P) reserves to the vegetative parts of the devel-oping seedling during early growth. Remobilization and
photosynthesis are therefore two parallel and co-coinciding processes involved in better seedlingestablishment and early growth.Methods A study was conducted to evaluate the pri-ority of developing maize seedlings to translocate theremobilized seed P reserves and external P uptake toseedling root and shoot sinks during 4 weeks of earlygrowth. Two fluxes of P in growing seedlings, onefrom seed remobilized P reserves and one from exter-nal P uptake, were distinguished by labelling externalnutrient solution P with 32P.Results The seedling phytomass was equally distrib-uted between seedling roots and shoots for 530 cumu-lated degree days after sowing. Seedlings partitionedup to 71 % of P from seed reserves and up to 68 % of Pacquired from the nutrient solution, to the shoots,depending on the seed P content and P concentrationin the nutrient solution. It appears that accumulation ofP slows down in seedling roots corresponds to thetranslocative functions of root P towards shoots forstart of photosynthesis.Conclusions Our results suggest that the major part ofseed P reserves and external P uptake were used inearly development of the seedling and the preferredsink was seedling shoots.
Keywords Maize . Seed . Phosphorus . Phytomass .
Isotope . P uptake . Early growth
Plant SoilDOI 10.1007/s11104-013-1689-x
Responsible Editor: Philip John White.
M. Nadeem :A. Mollier :C. Morel : S. PellerinINRA, UMR 1220 TCEM,71 avenue Bourlaux, CS 20032,33882 Villenave d’Ornon Cedex, France
M. Nadeem :A. Mollier :C. Morel : S. PellerinBordeaux Sciences Agro, UMR 1220 TCEM,33175 Gradignan, France
M. Nadeem (*) :M. Shahid :M. AslamDepartment of Environmental Sciences, COMSATSInstitute of Information Technology,Vehari 61100, Pakistane-mail: [email protected]
M. Zia-ur-RehmanInstitute of Soil and Environmental Sciences,University of Agriculture,Faisalabad 38000, Pakistan
M. A. WahidDepartment of Agronomy, University of Agriculture,Faisalabad 38000, Pakistan
AbbreviationsLS seeds Seeds with low seed P reservesHS seeds Seeds with high seed P reservesLS seedlings Seedling grown from LS seedsHS seedlings Seedlings grown from HS seeds0P Zero external P availabilityLP Low external P availabilityHP High external P availability°C DAS Cumulated degree days after
sowingLPLS Seedlings grown from LS seeds
in low external P availabilityHPLS Seedlings grown from LS seeds
in high external P availabilityLPHS Seedlings grown from HS seeds
in low external P availabilityHPHS Seedlings grown from HS seeds
in high external P availability
Introduction
The seed occupies a central position in the life cycle ofangiosperms (Bewley and Black 1994). Stored reservesin the endosperm and scutellum of seeds are the maindriving force behind germination and seedling establish-ment during early growth before autotrophic growth isattained (Allen 2003; Finch-Savage et al. 2005; Miller2003; Modi and Asanzi 2008). Seed germination andearly seedling establishment thus depend on the amountand availability of seed endosperm and scutellum re-serves, in combination with sowing depth, temperature,water and soil health conditions (Sikder et al. 2009).During germination, the embryo develops into visibleroots and shoots by using stored energy in the form ofseed carbon (C) and other nutrient reserves (Liu et al.2010). Seed C is the main source used by developingseedling and 7-day-old maize seedlings are entirelymade up of carbon remobilized from seed C reserves,whereas seedling root growth results from C remobilizedfrom seed C reserves until the ninth day after germina-tion (Deleens et al. 1984). Phosphorus is one of themajorcomponents of all fundamental cell structures, includingphospholipids, DNA, RNA and enzymes. In maize, seedP is mainly stored in the seed scutellum whereas seedphytomass is mainly concentrated in endosperm (Harveyand Oaks 1974; Nadeem et al. 2011; Nadeem et al.2012a). Earlier studies have shown that seed reserves(C, N, P) start to remobilize soon after imbibition and are
translocated towards growing seedling sinks (Guardiolaand Sutcliffe 1971; Covell et al. 1986; Deleens et al.1984; Nadeem et al. 2011; Nadeem et al. 2012a; Squire1999). Seeds of crop plants generally contain sufficient Pto support maximum seedling growth for several weeksafter germination (White and Veneklaas 2012). Theremobilization of seed P reserves is not a limiting stepfor seedling P nutrition during the early growth of maizeseedlings (Nadeem et al. 2012b). However, the alloca-tion of P from seed P reserves and from external P uptaketowards the developing roots and shoots of young seed-lings is one of the main factors determining seedlingestablishment and final crop harvest.
Germination ends when seedlings have exhaustedthe food reserves stored in the seed endosperm andscutellum. Thereafter, seedling growth rate is deter-mined by the nutrient supply from the surroundingenvironment. Early growth is a key factor for the finalcrop harvest (Grant et al. 2001; Sekiya and Yano2010) and several studies have identified P nutritionas being critical during the early stages of crop growth(Mollier and Pellerin 1999; Colomb et al. 2000; Plenetet al. 2000; Assuero et al. 2004; Hammond et al. 2009;Martinefsky et al. 2010). This raises the question ofthe identity of the major sinks for the remobilized seedP reserves and external P uptake during early seedlinggrowth. The objective of this study was thus to ana-lyze the priority of growing maize seedlings in allo-cating the remobilized seed P reserves and external Puptake among developing roots and shoots and toidentify the main source of P for the developing sinks(roots and shoots) during early growth.
Material and methods
The study was conducted in controlled environmentalconditions in a growth chamber at INRA Bordeaux,France, in November-December 2010. Homogenousmaize seeds with low (LS; 506 μg P seed−1) and high(HS; 952 μg P seed−1) seed P reserves were used. LSand HS seeds were harvested as part of a long term Pfertilization experiment with irrigated maize at the ex-perimental site of Cestas-Pierroton in southwest France.Uniform seeds from LS and HS seed lots with thehomogenous seed weight (0.33 g±0.001) were sownin small opaque polypropylene pots with three concen-trations of available external P, supplied as NaH2PO4, inthe nutrient solution: 0, 100 μmol P L−1 and 1,000 μmol
Plant Soil
P L−1 for the zero P (0P), low (LP) and high P (HP)treatments, respectively. The nutrient solution (Bhadoriaet al. 2004) consisted of 1 mMNO3 as Ca(NO3)2.4H2O,0.2 mMK asKCl, 0.1 mMMg asMgSO4.7H2O, 46μMB as H3BO3, 9.1 μMMn asMnCl2.4H2O, 0.8 μMZn asZnSO4.7H2O, 0.3 μMCu as CuSO4.5H2O, 0.5 μMMoas (NH4)6Mo7O24.4H2O, and 2 mg L−1 Fe asSequestrene-138 Fe. The pH of the all three nutrientsolutions was adjusted to 6.4. The external P was la-belled with radioactive P (32P) to monitor thepartitioning of external P uptake between seedling rootsand shoots. An initial measured quantity (Rt0),1,561 kBq and 1,669 kBq of 32P was added in 16 L ofthe 0P and LP solutions, respectively, while 1,667 kBqwas added in 15 l of the HP solution for labelling ofexternal P. The initial specific activity (SAt0) was1.043 kBq (μmol P)−1 and 0.111 kBq (μmol P)−1 inLP and HP labelled nutrient solutions, respectively.
The plastic pots were filled with perlite and nineseeds from each LS and HS treatment were sown at2 cm sowing depth. There were 183 pots in total ofwhich 90 pots were planted with LS seeds and theremaining 90 pots with HS seeds. The three extra potsplanted with HS seed followed the HP treatment todetermine background radioactivity. A measured vol-ume of labelled nutrient solution (210 mL pot−1) wasadded to perlite (120 g pot−1) in each pot to reach90 % saturation capacity of perlite. Out of total 90 potssown with LS seeds, three sets of 30 pots each wereirrigated with a labelled nutrient solution of 0P, LP,and HP, respectively. Similarly, out of 90 pots plantedwith HS seeds, three sets of 30 pots each were irrigat-ed with a labelled nutrient solution of 0P, LP, and HP,respectively. Throughout the experiment, the potswere irrigated with water on the basis of water lostfrom the pots by evapotranspiration. Pots destined forthe final seedling harvest were weighed daily to cal-culate average weight loss due to evapotranspiration.
All the pots were placed in plastic trays on fixed ironstands. The photoperiod was 18/6 h of light/darknessthroughout the seedling growth period. Air temperature,relative humidity and light intensity in the growth cham-ber were measured throughout the experiment. Theaverage air temperature, relative humidity and lightintensity were 28 °C, 49 % and 657 μmol m−2 s−1,respectively. Thermal time was calculated in cumulateddegree days after sowing (°C DAS) using 10 °C as basetemperature (Eagles and Hardacre 1979) from hourlyrecorded air temperature data.
Seedling harvest and chemical analysis
The seedlings were harvested 10 times during thegrowth period of 530 °C DAS. The seedlings wereharvested at 16, 34, 52, 71, 89, 126, 163, 199, 309 and530 °C DAS corresponding to 1, 2, 3, 4, 5, 7, 9, 11, 17and 29 days after sowing. Three pots were selected ateach seedling harvest and six homogenous seedlingswere selected for each sampling. The six selected seed-lings were first washed with a 400 mL solution of0.5 mM CaSO4 at 4 °C for 1 min to remove all theperlite from the seedling roots. The seedlings were thenwashed in a 400 mL solution containing P (0, 100 μmolP L−1 or 1,000 μmol P L−1, depending upon the externalP treatment) and CaSO4 (0.5 mM) for desorption of 32Pfrom the seedling roots (4 min at 4 °C) (Rubio et al.2004). After washing, the seedlings were placed on aglass plate and the roots and shoot separated with aspatula. The dry phytomass (g seedling−1) of the seed-ling roots and shoots was determined at each seedlingharvest on samples lyophilized for 24 h.
Phosphorus concentrations in seedling roots andshoots were determined by adapting the malachite greencolorimetric technique (Van Veldhoven and Mannaerts1987). Each tissue (root or shoot) was ground separatelyin a Retsch MM 400 mixer mill, (Retsch GmbH, Haan,Germany). Each ground sample was weighed and thenreduced to ashes at 550 °C for 5 h. The resulting ash wasdissolved in 5 mL distilled water and left on the hotplateto evaporate until only a few drops remained. After Pmineralization with HNO3, P contents were measuredcolorimetrically (Van Veldhoven and Mannaerts 1987).Total seedling P represents the sum of seedling root Pand shoot P from seed remobilized P and external Puptake at each seedling harvest.
The external P uptake was estimated assuming thatno 32P/31P-fractionation occurred during P uptake fromthe external solution (Pext) by seedling roots (Fardeau1993; Schjørring and Jensén 1984) as follows,
Rt
Pns
� �¼ rt
Pext
� �) Pext ¼ rt
RtPns
� �
where RtPns
� �is specific radioactivity SAt measured at
harvest time t in the nutrient solution, rt is32P-radioac-
tivity measured in the mineralized seedling roots andshoots at harvest time t. Rt and Pns are the amount ofcarrier-free 32P and 31P measured in the nutrient
Plant Soil
solution, respectively. The Rt and rt values were countedusing a scintillation cocktail (Insta-gel Plus Packard,PerkinElmer) using a Packard TR 2100 (CanberraIndustries, Meriden, CT). The standard counting timewas 20 min. Since no significant 32P activity wasdetected in roots or shoots of seedlings grown in theHP nutrient solution without labelling, backgroundnoise caused by radioactivity was considered to be zero.
Experimental design and statistical analysis
The treatments were defined as the factorial combina-tion of two seed P reserves (LS, HS) and three rates ofavailable external P (0P, LP, HP) in the nutrient solu-tions. The treatments were applied in a completelyrandomized block design with three replicates and 10seedling harvest times. Data were analyzed byANOVA using the R environment for statistical com-puting and graphics, version 2.9.1 (R DevelopmentCore Team 2009). Means were compared usingTukey’s test at the 0.05 probability level.
Results
Seedling phytomass partitioning among rootsand shoots
The growing maize seedlings started to accumulateroot and shoot phytomass at 34 °C DAS with theemergence of the coleoptiles and radicle, respectively,from the imbibed seeds. All the seedlings continued toaccumulate phytomass up to the final harvest (530 °CDAS) with equal C partitioning between the roots andthe shoot, with no significant differences observedbetween treatments (Fig. 1).
Seedling phosphorus partitioning among rootsand shoots
With the emergence of seedling radicle and coleoptilesat 34 °C DAS, the seedling roots and shoots started toaccumulate P. The partitioning of P between the rootsand the shoot of the seedlings was similar during theearly stages of all the treatments, but by 126 °C DASseedlings began to partition significantly more P to-wards their shoot than to their roots in all the LStreatments and the HS treatment with zero external Pavailability (Fig. 2a, b, c, d). HS seedling increased P
partitioning to shoots at 163 and 199 °C DAS in theLP and HP treatments, respectively (Fig. 2e, f).
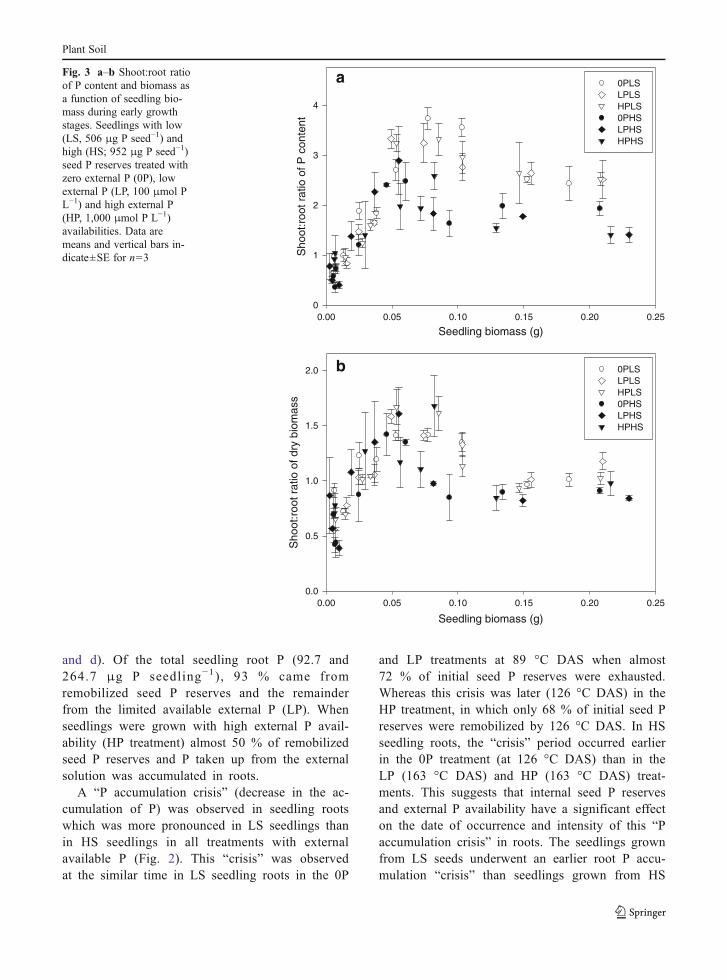
As seedling biomass increased, the P shoot:root ratiorose steeply in all treatments and then decreased butremained above 1.0 (Fig. 3a). These results indicate thatP remobilized from seed reserves and external P waspreferentially allocated toward the seedling shoot.Whenseedling biomass was higher than 0.06 g, the P shoot:root ratio was higher in the LS than in the HS treatmentand a similar trend was observed in root:shoot biomassratio but was less pronounced (Fig. 3b). This trendprevailed until 530 °C DAS indicating that when seed-lings were grown from seeds with low (506μg P seed−1)P reserves, seedling P was preferentially translocatedtowards the shoot. The P shoot:root ratio was not affect-ed by the availability of external P.
External phosphorus uptake partitioning between rootsand shoots
Although in the 0P treatment, the nutrient solution waslabelled with 32P, no significant P uptake from the exter-nal solution was observed by LS or HS seedlings in the0P treatment. This result confirmed that in the 0P treat-ment, the external P concentration was close to zero andlower than the minimum concentration required for Puptake. Significant 32P activity was observed in roots ofLS and HS seedlings provided with external P at 71 °CDAS (Fig. 4). As soon as the seedling leaves were devel-oped, from 71 °C DAS, the external P taken up by theroots was translocated towards the seedling shoot (Fig. 4).Translocation of external P to the seedling shoot wasgreater than that to the roots in LS seedlings from163 °C DAS on and this trend continued until the finalseedling harvest (Fig. 4a and c). In HS seedlings, the trendwas observed only at the last seedling harvest, possiblydue to the greater supply of seed P reserves to the seedlingroots and shoots (Fig. 4b and d). Comparatively moreexternal P was translocated to the seedling shoot in LSseedlings than in HS seedlings at 530 °C DAS.
Shoot P originated from both remobilized seedP reserves and external P uptake
Root and shoot P found in LS and HS seedlingsoriginated from seed P reserves in the 0P treatment.The increasing trend of seedling shoot P originatedfrom two different sources: remobilized seed P re-serves and P uptake from the available external P
Plant Soil
source (for LP and HP treatments) up to 530 °C DAS(Fig. 5). Shoot P contents were lower in LPLS(Fig. 5a) after 163 °C DAS because all the seed Preserves were exhausted before this date, but as soonas the seedling roots received a significant quantity ofexternal P, shoot P contents again started to increase asobserved at the last seedling harvest at 530 °C DAS.Quantitatively more P was accumulated in seedlingshoots in HS seedlings than in LS seedlings due tomore remobilized seed P reserves and their transfer to
the shoot (Fig. 5b and d). An equal portion ofremobilized seed P reserves (94 % representing a totalamount of 216.9 and 350.0 μg P seedling−1) wastranslocated to seedling shoots in both LS and HSseedlings in the treatment with low external P avail-ability. The greater availability of external P for LSand HS seedlings changed the partitioning ofremobilized seed P reserves and external P uptake tothe shoot. Of the total P in the HPLS seedling shoots(416.1 μg P seedling−1), 56 % came from remobilized
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
Shoot Root
0PLS
Sh
oo
t an
d r
oo
t p
hyt
om
ass
(g s
eed
ling
-1)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0PHS
LPLS
0 100 200 300 400 500 600
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
LPHS
0 100 200 300 400 500 600
HPLS HPHS
Thermal time (cumulated degree days after sowing)
a
b
c
d
e
f
Fig. 1 a–f Translocation ofphytomass (g seedling−1) tothe maize seedling shoot(black circle) and roots(white circle) during earlygrowth stages. Seedlingswith low (LS, 506 μg Pseed−1) and high (HS;952 μg P seed−1) seed P re-serves treated with zero ex-ternal P (0P, 0 μmol P L−1),low external P (LP,100 μmol P L−1) and highexternal P (HP, 1,000 μmolP L−1) availabilities, weregrown for 530 cumulateddegree days after sowing.Data are means and verticalbars indicate±SE for n=3
Plant Soil
seed P reserves and remainder was supplemented byexternal P uptake, whereas this trend was more prom-inent in HPHS seedlings in which 63 % (355.4 μg Pseedling−1) came from remobilized seed P reserves.
Root P originated from remobilized seed P reservesand external P uptake
Phosphorus accumulated in seedling roots in LSand HS seedlings up to 530 °C DAS (Fig. 6).
There was a linear increase in P accumulation inLPLS seedling roots mainly originating fromremobilized seed P reserves up to 89 °C DAS(Fig. 6a). Thereafter the observed increase wasonly due to the uptake of external P from thenutrient solution. A similar trend was observed inP accumulation in seedling roots in HPLS seed-lings (Fig. 6c), whereas the seedling P accumula-tion was higher in HS seedlings grown under low(LP) and high (HP) external P availability (Fig. 6b
0
100
200
300
400
500
600
700
Shoot Root
0PLS
Sh
oo
t an
d r
oo
t P
co
nte
nts
(µ
g P
see
dlin
g-1
)
0
100
200
300
400
500
600
700
0PHS
LPLS
0 100 200 300 400 500 600
0
100
200
300
400
500
600
700
LPHS
0 100 200 300 400 500 600
HPLS HPHS
Thermal time (cumulated degree days after sowing)
a
b
c
d
e
f
Fig. 2 a–f Translocation ofphosphorus (μg seedling−1)to the maize seedling shoot(black circle) and roots(white circle) during earlygrowth stages. Seedlingswith low (LS, 506 μg Pseed−1) and high (HS;952 μg P seed−1) seed P re-serves treated with zero ex-ternal P (0P), low external P(LP, 100 μmol P L−1) andhigh external P (HP,1,000 μmol P L−1) avail-abilities, were grown for 530cumulated degree days aftersowing. Data are means andvertical bars indicate±SEfor n=3
Plant Soil
and d). Of the total seedling root P (92.7 and264.7 μg P seedling−1), 93 % came fromremobilized seed P reserves and the remainderfrom the limited available external P (LP). Whenseedlings were grown with high external P avail-ability (HP treatment) almost 50 % of remobilizedseed P reserves and P taken up from the externalsolution was accumulated in roots.
A “P accumulation crisis” (decrease in the ac-cumulation of P) was observed in seedling rootswhich was more pronounced in LS seedlings thanin HS seedlings in all treatments with externalavailable P (Fig. 2). This “crisis” was observedat the similar time in LS seedling roots in the 0P
and LP treatments at 89 °C DAS when almost72 % of initial seed P reserves were exhausted.Whereas this crisis was later (126 °C DAS) in theHP treatment, in which only 68 % of initial seed Preserves were remobilized by 126 °C DAS. In HSseedling roots, the “crisis” period occurred earlierin the 0P treatment (at 126 °C DAS) than in theLP (163 °C DAS) and HP (163 °C DAS) treat-ments. This suggests that internal seed P reservesand external P availability have a significant effecton the date of occurrence and intensity of this “Paccumulation crisis” in roots. The seedlings grownfrom LS seeds underwent an earlier root P accu-mulation “crisis” than seedlings grown from HS
Seedling biomass (g)0.00 0.05 0.10 0.15 0.20 0.25
Sho
ot:r
oot r
atio
of P
con
tent
0
1
2
3
4
0PLS LPLS HPLS 0PHS LPHS HPHS
Seedling biomass (g)
0.00 0.05 0.10 0.15 0.20 0.25
Sho
ot:r
oot r
atio
of d
ry b
iom
ass
0.0
0.5
1.0
1.5
2.0 0PLS LPLS HPLS 0PHS LPHS HPHS
b
aFig. 3 a–b Shoot:root ratioof P content and biomass asa function of seedling bio-mass during early growthstages. Seedlings with low(LS, 506 μg P seed−1) andhigh (HS; 952 μg P seed−1)seed P reserves treated withzero external P (0P), lowexternal P (LP, 100 μmol PL−1) and high external P(HP, 1,000 μmol P L−1)availabilities. Data aremeans and vertical bars in-dicate±SE for n=3
Plant Soil
seeds. Similarly, the availability of external P alsohad a significant effect on the P accumulation“crisis” in the roots. Seedlings grown in the 0Ptreatment underwent a root P accumulation “crisis”earlier than seedlings grown in the LP or HP treatments.In seedling roots, the “P accumulation crisis” had recov-ered at the following harvests only in the seedlingsgrown with available external P. Thereafter there was aregular increase in seedling root P accumulation due toexternal P uptake (in LS seedlings) and P uptake andremobilized seed P reserves in HS seedlings (due to theavailability of seed P reserves up to the last seedlingharvest≈160 μg P seed−1 in HS seeds).
Discussion
The maize seedlings showed equal preference forphytomass translocation to roots and shoots duringearly growth stages of maize in all the P treatments
up to 530 °C DAS. The root:shoot ratios also showedequal distribution of seedling phytomass among rootsand shoots. Deleens et al. (1984) demonstrated that 7-day-old maize seedling leaves were entirely made upof remobilized seed C reserves and that thereafter,external C was incorporated through photosynthesis,whereas root growth continued until the ninth dayusing remobilized seed C reserves. The similar alloca-tion of phytomass towards shoots and roots indicatesthat the C was utilized in the early development ofseedling roots and shoots. These findings are in agree-ment with those of Deleens et al. (1984) and Liu et al.(2010). More seedling phytomass was allocated toseedling roots than to shoots in P deficient plants asreported by Cakmak et al. (1994), whereas in ourstudy we did not observe any preferred sink forphytomass allocation during first four weeks of seed-ling growth. Despite very low P availability in theseedling root zone, the increase in seedling biomasswas not reduced. This was due to the remobilization of
0 100 200 300 400 500 600
0
5
10
15
20
25
30
Shoot Root
0 100 200 300 400 500 600
0 100 200 300 400 500 600
0
50
100
150
200
250
300
0 100 200 300 400 500 600
Sh
oo
t an
d r
oo
t P
co
nte
nts
ori
gin
ated
fro
m e
xter
nal
P u
pta
ke (
µg
P s
eed
ling
-1)
Thermal time (cumulated degree days after sowing)
a b
c d
LPLS LPHS
HPLS HPHS
Fig. 4 a–d External P up-take (μg seedling−1) andtranslocation to seedlingroots (white circle) and tothe shoot (black circle) dur-ing early growth stages.Seedlings with low (LS,506 μg P seed−1) and high(HS; 952 μg P seed−1) seedP reserves treated with lowexternal P (LP, 100 μmol PL−1) and high external P(HP, 1,000 μmol P L−1)availabilities, were grownfor 530 cumulated degreedays after sowing. Data aremeans and vertical bars in-dicate±SE for n=3
Plant Soil
stored P reserves in the seeds. In both LS and HSseedlings, this remobilization was sufficient to meetthe P needs of the shoot. The P concentration in theshoot thus remained above the critical tissue P con-centration at which the shoot growth is retarded. Thisshows that when temperature and soil moisture condi-tions are satisfactory, seed P reserves are sufficient forseedling emergence in the field even under low P avail-ability. This is possibly due to P concentrations in theleaf which were higher than the critical tissue P concen-tration (Assuero et al. 2004; Plenet et al. 2000;Martinefsky et al. 2010; White and Brown 2010) atwhich seedling growth is retarded and seedlings startto allocate more phytomass towards roots than to shootsto enable the roots to explore a larger area to improveacquisition of soil P (Lynch and Brown 2008; Shen et al.2011). Under low external P availability, shoot growth isreduced whereas root growth is not affected or is evenenhanced in the short term (Mollier and Pellerin 1999;Wissuwa et al. 2005). Our results show that P is prefer-entially allocated to shoots when available P is low,implying that root P demand is lower than shoot P
demand (Nadeem et al. 2011; Nadeem et al. 2012a).Under low P availability, the allocation of P towardsroots is enhanced but this effect is greater when thegrowth period exceeds 6 weeks, whereas under high Pavailability, partitioning is higher to shoots than roots(Wissuwa et al. 2005).
In seeds, P is stored in the form of phytate. Duringgermination, the hydrolysis of phytate provides P forthe developing seedlings (Nadeem et al. 2011;Nadeem et al. 2012b). Phosphorus is the main com-ponent of nucleic acid, phospholipids and metabolitesand is involved in energy production and photosyn-thesis (Veneklaas et al. 2012). As soon as the seedlingsdeveloped their root system, they started to take upexternal P from the root zone to fill seedling P re-quirements. The efficiency of the leaf as an assimila-tory organ depends on a wide range of biochemicaland physiological processes are in a dynamic relation-ship with leaf ontogeny (Silvius et al. 1978). Seedlingsare undergoing intense and rapid cell division and theleaves of seedlings require large amounts of phospho-rus. We observed that both seed P reserves and
0
100
200
300
400
500
600
Total shoot PRemobilized seed PExternal P uptake
0 100 200 300 400 500 600
0
100
200
300
400
500
600
0 100 200 300 400 500 600
Sh
oo
t P
co
nte
nt
(µg
P s
eed
ling
-1)
Thermal time (cumulated degree days after sowing)
a b
c d
LPLS LPHS
HPLS HPHS
Fig. 5 a–d P (black circle)contents of seedling shoots(μg seedling−1) originatedfrom remobilized seed P re-serves (white circle) and ex-ternal P uptake (black down-pointing triangle) duringearly growth stages. Seed-lings with low (LS, 506 μgP seed−1) and high (HS;952 μg P seed−1) seed P re-serves treated with low ex-ternal P (LP, 100 μmol PL−1) and high external P(HP, 1,000 μmol P L−1)availabilities, were grownfor 530 cumulated degreedays after sowing. Data aremeans and vertical bars in-dicate±SE for n=3
Plant Soil
external P supplied roots and leaves, but a slightlyhigher proportion of P from seed reserves went toleaves rather than to roots. Allocation of a higherproportion of remobilized P from the seed and ofexternal P towards the seedling shoot indicates ahigher seedling shoot P demand during this periodand any P limitation affects photosynthesis and carbonpartitioning differently when applied over a long peri-od than over a short period (Rao and Terry 1995). Lowavailability of P to sugar beet plants reduced photo-synthesis whereas the supply of P to sugar beet seed-lings caused an increase in sugar phosphate in plantleaves that most likely increased photosynthesis (Raoand Terry 1995). This indicates that leaf P require-ments are higher than root requirements during earlygrowth because the leaves are the active sites ofphotosynthesis.
The slowdown in seedling root P accumulation (Paccumulation crisis) was observed earlier in LS seed-ling roots than in HS seedling roots. In the 0P treat-ment, this P accumulation crisis was observed earlierthan in treatments with available external P in both LS
and HS seedlings (Fig. 2). Root P accumulation crisisindicates the translocation of root P towards leavesthat are the main centers of energy production throughphotosynthesis for which P is one of the main nutrientsrequired (Usuda and Shimogawara 1991; Xu et al.2007). Deleens et al. (1984) observed a similar carbonaccumulation crisis in maize seedlings root andreported that this crisis was the driving force behindthe shift of seedlings from the transitional stage to theautotrophic stage for seedling C demand. The root Paccumulation crisis was also observed in maize seed-lings during a similar period as reported by Deleens etal. (1984) suggesting a close relationship between Pand C demand for the shift of developing maize seed-lings towards the autotrophic stage for nutrition, dur-ing which the seedlings absorb C from theatmosphere, whereas P is taken up from the seedlingroot zone by the roots. This raises the question of therelationship between C and P remobilization in germi-nating maize seeds and the ability of developing seed-lings to reach the autotrophic stage for C and P duringthe early growth stages.
0
100
200
300
400
Total root PRemobilized seed PExternal P uptake
0 100 200 300 400 500 600
0
100
200
300
400
0 100 200 300 400 500 600
Ro
ot
P c
on
ten
t (µ
g P
see
dlin
g-1
)
Thermal time (cumulated degree days after sowing)
a b
c d
LPLS LPHS
HPLS HPHS
Fig. 6 a–d Phosphorus(black circle) contents ofseedling roots (μg seed-ling−1) originated fromremobilized seed P reserves(white circle) and external Puptake (black down-pointingtriangle) during earlygrowth stages. Seedlingswith low (LS, 506 μg Pseed−1) and high (HS;952 μg P seed−1) seed P re-serves treated with low ex-ternal P (LP, 100 μmol PL−1) and high external P(HP, 1,000 μmol P L−1)treatments, were grown for530 cumulated degree daysafter sowing. Data aremeans and vertical bars in-dicate±SE for n=3
Plant Soil
Conclusion
The aim of this research was to understand the priorityof translocating the phytomass, remobilized seed P re-serves and external P uptake to seedling roots and to theshoot during early growth stages of maize. Our resultssuggest that most of the stored seed reserves ofphytomass and P were used in the early developmentof the growing maize seedlings and that the majority ofremobilized seed P reserves and external P uptake wastranslocated to shoots rather than to roots. This indicatesthe preference of developing seedlings to allocate P tothe shoot which is the active site for photosynthesis. Thesecond aim was to distinguish the P reserves allocated toroots and shoots according to their origin. Up to 530 °CDAS, the most P in roots and shoots originated fromremobilized P seed reserves and the secondary sourcewas external P uptake. The translocation of remobilizedseed P reserves and external P uptake to the shoots ofseedlings revealed the high P requirement of this organ,which might be related to P demand for leaf elongationand photosynthesis. This high P demand by leavesmight explain why maize seedlings often suffer from Pdeficiencies and reduced leaf growth during earlygrowth stages.
Acknowledgments This study was funded by a project grantby the Higher Education Commission (HEC), Pakistan andbenefited from financial support from INRA (French NationalInstitute for Agricultural Research). The authors acknowledgetechnical help and advice from Alain Vives, Loïc Prud’homme,Sylvie Milin and Sylvie Bussiere. We also thank DaphneGoodfellow for revising the English. Thanks are also due totwo referees their useful comments.
References
Allen P (2003) When and how many? Hydrothermal models andthe prediction of seed germination. New Phytol 158(1):1–3
Assuero SG, Mollier A, Pellerin S (2004) The decrease ingrowth of phosphorus-deficient maize leaves is related toa lower cell production. Plant Cell Environ 27(7):887–895
Bewley JD, Black M (1994) Seeds: physiology of developmentand germination. Plenum Press, New York, pp 1–31
Bhadoria PS, El Dessougi H, Liebersbach H, Claassen N (2004)Phosphorus uptake kinetics, size of root system and growthof maize and groundnut in solution culture. Plant Soil262(1–2):327–336
Cakmak I, Hengeler C, Marschner H (1994) Partitioning ofshoot and root dry matter and carbohydrates in bean plantssuffering from phosphorus, potassium and magnesium de-ficiency. J Exp Bot 45(278):1245–1250
Colomb B, Kiniry JR, Debaeke P (2000) Effect of soil phos-phorus on leaf development and senescence dynamics offield-grown maize. Agron J 92(3):428–435
Covell S, Ellis RH, Roberts EH, Summerfield RJ (1986) Theinfluence of temperature on seed germination rate in grainlegumes. 1: a comparaison of chickpea, lentil, soybeanand cowpea at constant temperatures. J Exp Bot 37:705–715
Deleens E, Gregory N, Bourdu R (1984) Transition betweenseed reserve use and photosynthetic supply during devel-opment of maize seedlings. Plant Sci Lett 37(1–2):35–39
Eagles HA, Hardacre AK (1979) Genetic variation in maize(Zea mays L.) for germination and emergence at 10 °C.Euphytica 28(2):287–295
Fardeau JC (1993) Available soil phosphate: its representationby a functional multiple compartment model. Agronomie13(4):317–331
Finch-Savage WE, Rowse HR, Dent KC (2005) Developmentof combined imbibition and hydrothermal thresholdmodels to simulate maize (Zea mays) and chickpea (Cicerarietinum) seed germination in variable environments.New Phytol 165(3):825–838
Grant CA, Flaten DN, Tomasiewicz DJ, Sheppard SC (2001)The importance of early season phosphorus nutrition. Can JPlant Sci 81(2):211–224
Guardiola JL, Sutcliffe JF (1971) Mobilization of phosphorus inthe cotyledons of young seedlings of the garden pea (Pisumsativum L.). Ann Bot 35(4):809–823
Hammond JP, Broadley MR, White PJ, King GJ, Bowen HC,Hayden R, Meacham MC, Mead A, Overs T, SpracklenWP, Greenwood DJ (2009) Shoot yield drives phosphorususe efficiency in Brassica oleracea and correlates with rootarchitecture traits. J Exp Bot 60(7):1953–1968
Harvey BMR, Oaks A (1974) The hydrolysis of endospermprotein in Zea mays. Plant Physiol 53(3):453–457
Liu SL, Siao W, Wang SJ (2010) Changing sink demand ofdeveloping shoot affects transitory starch biosynthesis inembryonic tissues of germinating rice seeds. Seed Sci Res20(3):137–144
Lynch J, Brown K (2008) Root strategies for phosphorus acquisi-tion. In: White P, Hammond J (eds) The Ecophysiology ofPlant-Phosphorus Interactions, vol 7, Plant Ecophysiology.Springer, Netherlands, pp 83–116
Martinefsky MJ, Assuero SG, Mollier A, Pellerin S (2010)Analysis of the response of two tall fescue cultivars ofdifferent origin to P deficiency. Environ Exp Bot 69:250–258
Miller BM (2003) Seed germination. In: Miller BM, LawrenceOC (eds) Principles of seed science and technology.Kluwer Academic Publishers, Norwell, pp 72–124
Modi AT, Asanzi NM (2008) Seed performance of maize inresponse to phosphorus application and growth tempera-ture is related to phytate-phosphorus occurrence. Crop Sci48(1):286–297
Mollier A, Pellerin S (1999) Maize root system growth anddevelopment as influenced by phosphorus deficiency. JExp Bot 50(333):487–497
Nadeem M, Mollier A, Morel C, Vives A, Prud’homme L,Pellerin S (2011) Relative contribution of seed phosphorusreserves and exogenous phosphorus uptake to maize (Zeamays L.) nutrition during early growth stages. Plant Soil346(1–2):231–244
Plant Soil
Nadeem M, Mollier A, Morel C, Vives A, Prud’homme L,Pellerin S (2012a) Maize (Zea mays L.) endogenous seedphosphorus remobilization is not influenced by exogenousphosphorus availability during germination and earlygrowth stages. Plant Soil 357:13–24
Nadeem M, Mollier A, Morel C, Vives A, Prud’homme L,Pellerin S (2012b) Seed phosphorus remobilization is nota major limiting step for phosphorus nutrition during earlygrowth of maize. J Plant Nutr Soil Sci 175:805–809
Plenet D, Etchebest S,Mollier A, Pellerin S (2000) Growth analysisof maize field crops under phosphorus deficiency—I. Leafgrowth. Plant Soil 223(1–2):117–130
R Development Core Team (2009) R: a language and environ-ment for statistical computing. R Foundation for StatisticalComputing, Vienna, Austria. http://www.r-project.org/.Accessed 4 Feb 2012
Rao IM, Terry N (1995) Leaf phosphate status, photosynthesis,and carbon partitioning in sugar beet. IV. Changes withtime following increased supply of phosphate to low-phosphate plants. Plant Physiol 107(4):1313–1321
Rubio G, Sorgona A, Lynch JP (2004) Spatial mapping ofphosphorus influx in bean root systems using digital auto-radiography. J Exp Bot 55(406):2269–2280
Schjørring JK, Jensén P (1984) Phosphorus nutrition of barley,buckwheat and rape seedlings. II. Influx and efflux ofphosphorous by intact roots of different P status. PhysiolPlant 61(4):584–590
Sekiya N, Yano K (2010) Seed P-enrichment as an effective Psupply to wheat. Plant Soil 327(1–2):347–354
Shen J, Yuan L, Zhang J, Li H, Bai Z, Chen X, Zhang W, ZhangF (2011) Phosphorus dynamics: from soil to plant. PlantPhysiol 156(3):997–1005
Sikder S, Hasan MA, Hossain MS (2009) Germination charac-teristics and mobilization of seed reserves in maize varie-ties as influenced by temperature regimes. J Agric RuralDev 7(1&2):51–56
Silvius JE, Kremer DF, Lee DR (1978) Carbon assimilation andtranslocation in soybean leaves at different stages of devel-opment. Plant Physiol 62:54–58
Squire GR (1999) Temperature and heterogeneity of emergencetime in oilseed rape. Ann Appl Biol 135:439–447
Usuda H, Shimogawara K (1991) Phosphate deficiency inmaize. I. Leaf phosphate status, growth, photosynthesisand carbon partitioning. Plant Cell Physiol 32(4):497–504
Van Veldhoven PP, Mannaerts GP (1987) Inorganic and organicphosphate measurements in the nanomolar range. AnalBiochem 161(1):45–48
Veneklaas EJ, Lambers H, Bragg J, Finnegan PM, Lovelock CE,Plaxton WC, Price CA, Scheible W-R, Shane MW, WhitePJ, Raven JA (2012) Opportunities for improvingphosphorus-use efficiency in crop plants. New Phytol195(2):306–320
White PJ, Brown PH (2010) Plant nutrition for sustainabledevelopment and global health. Ann Bot 105(7):1073–1080
White P, Veneklaas E (2012) Nature and nurture: the importanceof seed phosphorus content. Plant Soil 357(1):1–8
Wissuwa M, Gamat G, Ismail AM (2005) Is root growth underphosphorus deficiency affected by source or sink limita-tions? J Exp Bot 56(417):1943–1950
Xu HX, Weng XY, Yang Y (2007) Effect of phosphorus defi-ciency on the photosynthetic characteristics of rice plants.Russ J Plant Physiol 54(6):741–748