Markers of Exidative Stress in Hepatopancreas of Crayfish (Orconectes limosus, Raf) Experimentally Exposed to Nanosilver Witold Struz ˙ yn ´ ski, 1 Beata Da ˛ browska-Bouta, 2 Tomasz Grygorowicz, 2 Elz ˙ bieta Ziemin ´ ska, 3 Lidia Struz ˙yn ´ ska 2* 1 Department of Animal Environment Biology, Unit of Zoology, Faculty of Animal Sciences, Warsaw University of Life Science, 8 Ciszewskiego str., 02-786 Warsaw, Poland 2 Department of Neurochemistry, Laboratory of Pathoneurochemistry, Mossakowski Medical Research Center, Polish Academy of Sciences, 5 Pawinskiego str., 02-106 Warsaw, Poland 3 Department of Neurochemistry, Laboratory of Pharmaconeurochemistry, Mossakowski Medical Research Center, Polish Academy of Sciences, 5 Pawinskiego str., 02-106 Warsaw, Poland Received 12 April 2012; revised 1 February 2013; accepted 2 February 2013 ABSTRACT: Silver nanoparticles, chemically neutral particles in the size range of 1–100 nm, express strong antimicrobial activity and therefore have a broad range of applications. The increased use of consumer prod- ucts with nanosilver (nanoAg) may result in its release into the environment, and may particularly affect aquatic systems. The mechanisms of the harmful effects of nanoAg against aquatic organisms are unclear. Therefore, in the present study we investigate the pro-oxidative potential of these nanoparticles in experimentally exposed crayfish Orconectes limosus. Markers of oxidative stress and parameters of the antioxidant cell defense system such as total glutathione, glutathione reductase and the level of sulfhydryl groups were examined in the hepato- pancreas of both sexes of O. limosus collected seasonally from Biale Lake (Poland) and subsequently exposed to nanoAg particles for 2 weeks. Exposure to nanoAg led to a high concentration-dependent increase in the rate of lipid peroxidation and a decrease of protein-bound SH groups which indicates protein oxidation. These markers of oxidative stress were accompanied by decreased levels of thiols and reduced activity of glutathione reductase. These results indicate a deficiency of reduced glutathione and suggest that the exposed organisms have less efficient antioxidative mechanisms available to counter ROS-mediated cellular stress. Furthermore, we find that confocal microscopy is of limited utility in monitoring the presence of silver nanoparticles in tissues of exposed crayfish. # 2013 Wiley Periodicals, Inc. Environ Toxicol 00: 000–000, 2013. Keywords: NanoAg; glutathione; lipid peroxidation; glutathione reductase; SH groups INTRODUCTION Silver in its ionic form has well known antibacterial proper- ties. Advances in nanotechnology have resulted in the abil- ity to produce silver nanoparticles. In the ‘‘nano’’ form (chemically neutral particles typically in the size range of 1–100 nm) this metal is much more effective as an antimi- crobial compound. Because of their extremely high antibac- terial potential, silver nanoparticles are incorporated into an increasing number of consumer products and during the last few years have become one of the most commonly used nanomaterials (Ahamed et al., 2010a). These particles are used in food packaging, odor-resistant textiles, liquids for laundering, and in medical products (Kubik et al., 2005; Chen and Schluesener, 2008). The adverse effects of nanoAg against bacteria are of particular concern as they Correspondence to: L. Struz ˙yn ´ska; e-mail: [email protected]Contract grant sponsor: Polish Ministry of Science and Higher Education Published online in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/tox.21859 C 2013 Wiley Periodicals, Inc. 1
Transcript
Markers of Exidative Stress in Hepatopancreas ofCrayfish (Orconectes limosus, Raf)Experimentally Exposed to Nanosilver
Witold Struzynski,1 Beata Dabrowska-Bouta,2 Tomasz Grygorowicz,2
Elzbieta Zieminska,3 Lidia Struzynska2*
1Department of Animal Environment Biology, Unit of Zoology, Faculty of Animal Sciences,Warsaw University of Life Science, 8 Ciszewskiego str., 02-786 Warsaw, Poland
2Department of Neurochemistry, Laboratory of Pathoneurochemistry, Mossakowski MedicalResearch Center, Polish Academy of Sciences, 5 Pawinskiego str., 02-106 Warsaw, Poland
3Department of Neurochemistry, Laboratory of Pharmaconeurochemistry, MossakowskiMedical Research Center, Polish Academy of Sciences, 5 Pawinskiego str., 02-106Warsaw, Poland
Received 12 April 2012; revised 1 February 2013; accepted 2 February 2013
ABSTRACT: Silver nanoparticles, chemically neutral particles in the size range of 1–100 nm, express strongantimicrobial activity and therefore have a broad range of applications. The increased use of consumer prod-ucts with nanosilver (nanoAg) may result in its release into the environment, and may particularly affect aquaticsystems. The mechanisms of the harmful effects of nanoAg against aquatic organisms are unclear. Therefore,in the present study we investigate the pro-oxidative potential of these nanoparticles in experimentally exposedcrayfish Orconectes limosus. Markers of oxidative stress and parameters of the antioxidant cell defense systemsuch as total glutathione, glutathione reductase and the level of sulfhydryl groups were examined in the hepato-pancreas of both sexes of O. limosus collected seasonally from Białe Lake (Poland) and subsequently exposedto nanoAg particles for 2 weeks. Exposure to nanoAg led to a high concentration-dependent increase in therate of lipid peroxidation and a decrease of protein-bound SH groups which indicates protein oxidation. Thesemarkers of oxidative stress were accompanied by decreased levels of thiols and reduced activity of glutathionereductase. These results indicate a deficiency of reduced glutathione and suggest that the exposed organismshave less efficient antioxidative mechanisms available to counter ROS-mediated cellular stress. Furthermore,we find that confocal microscopy is of limited utility in monitoring the presence of silver nanoparticles in tissuesof exposed crayfish. # 2013 Wiley Periodicals, Inc. Environ Toxicol 00: 000–000, 2013.
Keywords: NanoAg; glutathione; lipid peroxidation; glutathione reductase; SH groups
INTRODUCTION
Silver in its ionic form has well known antibacterial proper-ties. Advances in nanotechnology have resulted in the abil-ity to produce silver nanoparticles. In the ‘‘nano’’ form
(chemically neutral particles typically in the size range of1–100 nm) this metal is much more effective as an antimi-crobial compound. Because of their extremely high antibac-terial potential, silver nanoparticles are incorporated into anincreasing number of consumer products and during the lastfew years have become one of the most commonly usednanomaterials (Ahamed et al., 2010a). These particles areused in food packaging, odor-resistant textiles, liquids forlaundering, and in medical products (Kubik et al., 2005;Chen and Schluesener, 2008). The adverse effects ofnanoAg against bacteria are of particular concern as they
Contract grant sponsor: Polish Ministry of Science and Higher
Education
Published online in Wiley Online Library (wileyonlinelibrary.com).
DOI 10.1002/tox.21859
�C 2013 Wiley Periodicals, Inc.
1
are now widely used as component of medical productssuch as antibacterial fluids, wound dressings, implants, andcatheters (Chaloupka et al., 2010). However, there is also arisk of toxic effects that it may exert towards many otherorganisms. The surface area of small nanoparticles (NPs) isvery large compared to their volume what means that thissurface is an important component of the material, has adifferent composition from the core structure and is one ofthe reasons for some of unusual properties of nanoparticles,including increased reactivity and enhanced toxicity(Christian et al., 2008).
If released into surface waters, nanoAg may threaten
aquatic ecosystems. There is an increasing risk that due to
its widespread use, nanosilver will contaminate the water
system (Blaser et al., 2008) and by polluting rivers and
lakes may lead to poisoning of a variety of aquatic organ-
isms (Fabrega et al., 2011). Crustaceans, including crayfish,
are frequently used as bioindicators of water pollution and
are excellent model organisms for researching the ecotoxi-
cological relevance of toxin-induced changes at the molec-
ular level. Moreover, as a component of the human diet,
crayfish may transfer accumulated toxins, including
nanoAg, through the food chain into humans.
Orconectes limosus, a spiny-cheek crayfish, is a species
native to the east coast of North America. It was introduced
into Europe in 1890 and became widespread across almost
all European countries, including Poland (Struzynski and
Smietana, 1999). Ecological preferences of O. limosusinclude silty and muddy lowland waters, such as large riv-
ers, streams, canals, ponds and lakes, including organically
rich and polluted bodies of water (Struzynski et al., 2001;
Holdich and Black, 2007). It survives in water of poor qual-
ity and is therefore suitable for conducting ecotoxicological
studies as a specific endpoint in determination of toxin
interactions (Borkovic et al., 2008), including those medi-
ated by silver nanoparticles.
The mechanisms of nanoAg toxicity in living organisms
are poorly understood, although in vitro studies have
pointed out that oxidative stress may be one of the most
important mechanisms involved (Hussain et al., 2005).
NanoAg was reported to induce generation of reactive oxy-
gen species (ROS) and suppress level of reduced glutathi-
one (GSH) in human Chang liver cells (Piao et al., 2011).
Upregulation of oxidative stress-related genes has been
demonstrated in nanoAg-exposed fish (Chae et al., 2009;
Scown et al., 2010) and rodents (Rahman et al., 2009).
The characteristics of these mechanisms in different spe-
cies and different routes of exposure may be useful in
predicting ecological risks associated with nanoAg contami-
nation. Aquatic crustaceans have not been tested thoroughly
for susceptibility to nanoAg toxicity. There are data pointed
out the acute toxicity of nanosilver in Daphnia magna (Jo
et al., 2011) and its ability to inhibit growth and reproduction
under condition of chronic exposure (Zhao and Wang, 2011).
Therefore, the present study was undertaken to
explore the possibility of biochemical changes occurring
in tissues of an aquatic organism (the crayfish O. limo-sus) as a result of experimental chronic exposure to
nanoAg and to gain insight into mechanisms involved in
the changes. The pro-oxidative ability of nanoAg was
investigated in the hepatopancreas of crayfish and lipid
peroxidation of membranes was used as a marker of this
process. The glutathione antioxidant defense system in
tissue was monitored.
The crustacean hepatopancreas, also called the digestive
gland, is an organ responsible for most of the major
metabolic events in the organism including digestion,
absorption and storage of nutrients (Fingerman et al.,
1967), and molting (Gorell and Gilbert, 1969). As metabol-
ically active, displays comparatively higher levels and
activity of the oxyradical generating enzymes than other
tissues (Malik et al., 1987; Kandemir et al., 2011) and is
therefore more susceptible for oxidative stress under toxic
conditions. For this reason it was used in the present analy-
sis to effectively reflect the biochemical changes connected
with exposure to nanoAg.
The method used to monitor the accumulation of
nanoAg in crayfish was confocal microscopy. Until now
this simple method, which exploits the natural capability
of nanoAg to autofluorescence, has been used to detect
silver nanoparticles in bacteria treated with nanoAg
(Choi et al., 2010) but had not been used to examine
tissues. Thus, the secondary objective of this study was
to assess the utility of confocal microscopy for detecting
the presence and distribution of nanoAg in tissues of
exposed organisms.
EXPERIMENTAL METHODS
Animals
American freshwater crayfish, Orconectes limosus, were
collected by hand from Lake Białe near Włodawa (Poland)
GPS: 51829048.1800 N, 23832005.0200 in the autumn. A total
of 35 specimens of crayfish of each sex were kept alive and
immediately transferred to the laboratory. The crayfish
were then measured and weighed (mean total length 76 6
11 mm; mean weight 10.5 6 1.9 g), and placed individually
in 10 L aquaria with fresh water where they were allowed
to acclimatize for 1 day prior to the initiation of the study.
ABBREVIATIONS
GR glutathione reductaseGSH reduced form of glutathioneGSSG oxidized form of glutathionenanoAg silver nanoparticlesNPs nanoparticlesNRS nitrogen reactive speciesROS reactive oxygen speciesSH guoups sulfhydryl groupsTBARS thiobarbituric acid reactive substances
2 STRU _ZYNSKI ET AL.
Environmental Toxicology DOI 10.1002/tox
Exposure to Nanosilver
Silver nanopowder was purchased from Sigma–Aldrich, St.
Louis, MO. The commercially available Ag nanoparticles
used in the present study were defined as a mixture of poly-
vinylpyrrolidone-coated nanoparticles (PVP-coated
AgNPs) \100 nm in diameter and characterized by the
ated with the nanoparticle surface establishes a double layer
of charge that stabilizes the particles and prevents aggrega-
tion. This material was used in a previous toxicological
study (Park et al., 2010).
Nanoparticles, including nanoAg, have the tendency to
aggregate easily in aqueous solutions with biological media
where changes in aggregation state are caused by pH, ionic
strength and composition (Bradford et al., 2009; Zhang
et al., 2009). Thus, a stock solution of 10 g L21 of nanoAg
was prepared by dispersing the nanoparticles in ultrapure
water (Millipore) and subsequent sonication for 1 h prior to
dosing (BANDELIN sonoplus HD 70). To characterize the
degree of dispersion and particle size distribution, electron
microscopic (TEM) analysis of the solution was performed
according to a standard method.
Animals were divided into five groups. The control
group was not exposed to nanoAg and was retained in fresh
water. The four experimental groups were consistently sub-
jected to different concentrations of nanoAg over the course
of 2 weeks. The final concentrations of silver nanoparticles
in the experimental groups were 1, 5, 10, and 50 ppm.
These concentrations were chosen for consistency with the
report of Ahamed et al. (2010) which examined the results
of experiments with different doses of nanoAg.
Crayfish were exposed to nanoAg concentrations at a
temperature of 158C under a light and dark cycle of 8 h/16 h.
Collection and Preparation of Tissue forBiomarker Assays
After the exposure period, animals were rinsed in water,
anesthetized with ether and sectioned. The hepatopancreas
was isolated from each specimen and rinsed with ice-cold
saline to remove nanoparticles adhering to the surface. The
tissues were then frozen immediately in liquid nitrogen and
stored at 2708C until use.
The collected tissues were then homogenized in Krebs-
Ringer buffer, pH 7.4 before use in biomarker assays.
Confocal Microscopy Analysis
Three animals of each group (control and 1, 5, 10, 50 ppm
nanoAg) were anesthetized with a lethal dose of ether and
sectioned. Hepatopancreas and green glands were taken
and postfixed in fixative solution for 1.5 h at room tempera-
ture (4% paraformaldehyde; Sigma–Aldrich, St. Louis,
MO; in PBS). The organs were then cryoprotected in
sucrose in a three-step procedure: 10% sucrose (overnight),
20% (2 days) and 30% (4–5 days). The frozen tissues were
then cut into 40-mm sections and retained free-floating in
PBS, pH 7.4 with 0.1% sodium azide. Sections were
mounted on silane slides and coverslipped under Vecta-
shield Mounting Medium (Vector). The hepatopancreas
and green gland tissues of exposed crayfish were analyzed
to determine the presence of nanoAg. In some sections,
Hoechst stain (1.3 lg mL21) (Sigma–Aldrich, St. Louis,
MO) was used to stain the nuclei of the cells.
A two-photon laser scanning confocal microscope was
used as a previously defined method for nanoAg fluores-
cence imaging (Choi et al., 2010). NanoAg powder was
mounted on a silane slide and coverslipped under
Vectashield Mounting Medium (Vector) to analyze the
autofluorescence of nanoparticles.
Images (of nanoAg powder and organs) were obtained
using a two-photon laser scanning confocal microscope
(Zeiss LSM 510). An argon laser (488 nm) and a helium-
neon laser (543 nm) were used for excitation. A UV diode
was used for excitation when Hoechst stain was used.
Images were processed using the Zeiss LSM 510 software
package v. 3.2.
Measurement of Lipid Peroxidation
Lipid peroxidation was measured using the thiobarbituric
acid (TBAR) test according to Asakawa and Matsushita
(1980). Hepatopancreas homogenates were suspended in
Krebs-Ringer buffer, pH 4.0 since acidic pH gives optimal
conditions for high color formation and good color purity
of the reaction. To estimate the effect of nanoparticles of
silver on Fe31-dependent peroxidation, the samples were
preincubated with 25 lM Fe31, 800 lM adenosine diphos-
phate (ADP) and 200 lM ascorbate at 308C in a water bath.
After incubation, 1 mL of 30% TCA, 0.1 mL of 5 M HCl
and 1 mL of 0.75% TBAR were added. The tube was
capped and the mixture was heated at 1008C for 15 min in
boiling water. After centrifugation, the optical density of
the supernatant was determined at 535 nm. The molar
extinction coefficient used to calculate of malondialdehyde
(MDA) was 1.56 3 105 M21 cm21.
Total Glutathione Assay
Measurement of total glutathione (GSH 1 GSSG) content
was performed using the method of Tietze (1969) which
employs the glutathione disulfide reductase/DTNB recy-
cling assay. Briefly, 5% PCA was added to the samples of
hepatopancreas homogenates before centrifugation at
10,000 3 g for 15 min. The supernatant was added to a
mixture of 0.3 mM NADPH and 6 mM DTNB in stock
buffer (125 mM Na2HPO4, 6.3 mM NaEDTA, pH 7.5) and
3NANOSILVER-EVOKED OXIDATIVE STRESS IN CRAYFISH
Environmental Toxicology DOI 10.1002/tox
the absorbance was determined at 412 nm. Thereafter 5 mL
of glutathione reductase (130 U mg21 protein) was added
and the absorbance was monitored continuously until it
reached a value of 2.000. The total glutathione concentra-
tion was calculated from the difference between absor-
bances as mg mg21 protein using a standard curve.
Glutathione Reductase (GR) Assay
Glutathione reductase activity was measured spectrophoto-
metrically according to the method of Carlberg and Man-
nervik (1985). The reaction mixture contained 1 mM oxi-
dized glutathione (GSSG) as a substrate, 2 mM NADPH
and protein sample (hepatopancreas homogenate) in phos-
phate buffer (pH 7.0). The decrease in absorbance at 340
nm which indicates NADPH oxidation was measured. One
unit of enzyme activity is defined as 1 nmol of NADPH
oxidized in 1 h per mg of protein.
Measurements of the Level of SulfhydrylGroups
The level of sulfhydryl (SH) groups was determined by the
method of Sedlak and Lindsay (1968). For the determination
of total SH groups, samples of homogenates were mixed with
0.2 M Tris buffer, pH 8.2 and 0.1 M dithionitrobenzoic acid
(DTNB). Nonprotein SH groups were estimated after the
addition of 50% TCA to each sample. The tubes were shaken
and centrifuged at 3000 3 g for 10 min. The absorbance of
the supernatants was read within 5 min at 412 nm after the
addition of 0.4 M Tris buffer, pH 8.9 and 0.1 M DTNB
against a reagent blank. The protein-bound SH group content
was calculated from the total and nonprotein SH groups.
The protein concentration in tissue samples was meas-
ured by the method of Lowry et al. (1951) and used to cal-
culate the concentration of each of the markers per mg
protein.
Statistical Analysis
The determined biochemical parameters are expressed as
the mean 6 SD from the number of experiments stated
under the figures. The statistically significant differences
among the groups were evaluated using the one-way analy-
sis of variance (ANOVA) followed by Dunnett’s multiple
comparison test at P level \ 0.01 or 0.05 (as stated below
the figures) using PRISM, Graph Pad as a software.
RESULTS
TEM Characteristic of nanoAg and AqueousExposure of Crayfish
TEM analysis revealed distinct particles dispersed in solu-
tion (Fig. 1). Nanoparticles size distribution was calculated
semistatistically using 10 electron micrographs and was
expressed as percentages of a total number of particles
(Fig. 1). Size of counted nanoAg particles ranged mostly
between 10 and 40 nm (more than 70%) and only 27%
exceeded 40 nm.
Aqueous exposure to silver nanoparticles did not affect
body weight of crayfish and did not cause mortality. Only
one crayfish from the control group was lost during the ex-
perimental period and this probably was a result of molting.
No unusual behavior was observed.
Imaging of Intracellular nanoAg Deposits WithFluorescence Analysis
NanoAg was excited at two wavelengths, 488 and 543 nm,
and fluorescence emissions were detected. At both excita-
tion wavelengths nanosilver powder produced emission
spectra in the visible range observable as an autofluores-
cence of nanoAg. However, the argon laser (green light,
Fig. 1. Representative TEM micrographs of dispersednanoAg particles after sonication. Graph shows percentagedistribution of nanoparticle size.
4 STRU _ZYNSKI ET AL.
Environmental Toxicology DOI 10.1002/tox
488 nm) excited less fluorescence than the helium-neon
laser (red light, 543 nm) (Fig. 2). Thus, we selected the he-
lium-neon laser to estimate the fluorescence in tissues.
We attempted to detect the presence and distribution of
nanoAg in hepatopancreas of treated crayfish based on the
confirmed autoflourescence ability exhibited by the
nanoparticles (Choi et al., 2010). The results indicated the
presence of different-sized particles. Small fluorescent par-
ticles were observed in samples obtained from each of the
exposed groups. They were numerous in samples taken
from animals exposed to 10–50 ppm concentrations
[Fig. 3(A), exposed, arrows] whereas they were sparse in
1–5 ppm concentrations. A tendency toward particle aggre-
gation was observed. In some regions, particle aggregations
of increased size were visible [Fig. 3(B), exposed, arrow].
Surprisingly, in hepatopancreas obtained from control cray-
fish, we also noticed large (about 15 mm) fluorescent mate-
rial sparsely distributed throughout the tissue [Fig. 3(B),
control, asterisk], structurally different from nanoAg par-
ticles aggregations. Thus, we decided to compare another
type of tissue and isolated the antennal gland (green gland)
which functions as an excretory and osmoregulatory organ
in crayfish. Because it participates in the process of excre-
tion, the presence of nanoAg could be expected in this
organ of exposed crayfish. Indeed, fluorescent particles
were visible in the treated samples whereas they were not
present in gland tissue obtained from controls [Fig. 2(C),
Fig. 2. Autofluorescence of nanoAg powder. Confocal images of nanoAg obtained using different excitation wavelengths: (A)488 nm (an argon laser) and (B) 543 nm (a helium-neon laser), (C) merged image.
Fig. 3. Confocal images of hepatopancreas (A, B) and antennal gland (C) of control and nanoAg-exposed crayfish taken at543 nm. Arrows indicate fluorescent particles of different sizes. Hoechst stain was used as a blue marker of nuclei.
5NANOSILVER-EVOKED OXIDATIVE STRESS IN CRAYFISH
Environmental Toxicology DOI 10.1002/tox
control], [Fig. 3(C), arrows]. Staining of cell nuclei with
Hoechst stain indicated that the particles are mainly in the
cell cytoplasm.
Biomarkers of Oxidative Stress
Lipid Peroxidation and Oxidation of Proteins
Exposure to nanoAg was found to generally cause a statisti-
cally significant increase in the level of TBARS in homoge-
nates of crayfish hepatopancreas in all experimental groups
exposed to nanoAg when compared to controls. This sug-
gests a contribution of oxidative stress in mechanisms of its
peroxidation in a concentration-dependent pattern (Fig. 4).
Exposure to 50 ppm of nanoAg was found to cause the
greatest increase in TBARS (by about 600% over control).
The level of protein-bound SH groups had a tendency to
decrease, indicating the conversion of ��SH into oxidized
S��S bridges. In the case of the group treated with 1 ppm of
nanoAg, no changes were visible but exposure to 5, 10, and
50 ppm nanoAg produces a statistically significant decrease
of protein-bound SH groups below the level of the control by
about 33, 25, and 35%, respectively (Fig. 5).
Antioxidant Defense
Total glutathione levels (GSH 1 GSSG) measured in ho-
mogenates of hepatopancreas show a tendency to increase
in a concentration-dependent manner (Fig. 6). In the case of
crayfish treated with 1 or 5 ppm of nanoAg, changes in the
level of total glutathione were not observed relative to the
control. However, the level of glutathione doubled in cray-
fish exposed to 10 or 50 ppm of nanoAg suggesting that
stimulation of de novo synthesis of glutathione occurs in
response to existing oxidative stress.
Although the level of total glutathione was enhanced, at
least in higher nanoAg concentrations, the levels of nonpro-
tein sulfhydryl groups (which are mainly of GSH origin
and may reflect indirectly the amount of reduced form of
glutathione) were found to be practically the same in
control and nanoAg-treated animals (Fig. 7). Thus, a defi-
ciency of the reduced form of glutathione which is of im-
portance in scavenging the reactive oxygen\nitrogen spe-
cies is highly possible.
This possibility is supported by the observation of a sig-
nificant decrease in the specific GR activity of exposed
crayfish. The ability of glutathione reductase (GR) to
reduce oxidized glutathione to recover GSH is very impor-
tant for the cellular antioxidant defense system. Lowering
of GR activity was observed to occur in all of the treated
groups of animals and was highly concentration-dependent
Fig. 4. Lipid peroxidation measured by the TBARS-test andexpressed as nmoles of malondialdehyde (MDA) in homoge-nates of hepatopancreas obtained from control and chroni-cally nanoAg-treated crayfish. The results are presented asthe means 6 SD for five replicates. *P \ 0.01 vs. control(Dunnett’s Multiple Comparison test).
Fig. 5. The content of protein-bound ��SH groups in ho-mogenates of hepatopancreas obtained from control andnanoAg-treated crayfish. The values represent means 6 SDfrom four replicates. *P\0.05 vs. control (Dunnett’s MultipleComparison test).
Fig. 6. Total glutathione levels in homogenates of hepato-pancreas obtained from control and nanoAg-treatedcrayfish. The results are expressed as means 6 SD from fivereplicates, each conducted using tissue obtained from onedissected crayfish. *P\ 0.01 vs. control (Dunnett’s MultipleComparison test).
6 STRU _ZYNSKI ET AL.
Environmental Toxicology DOI 10.1002/tox
(Fig. 8). After exposure to 1 ppm of nanoAg, GR activity
was half of the control value and the most extreme effect
was noted in the crayfish treated with the 50 ppm dose. In
this group, the GR activity reached only about 20% of the
control activity.
DISCUSSION
Silver NPs are widely used in many consumer goods in
efforts to take advantage of their antimicrobial properties.
Hence, the possibility of environmental release of nanoAg
and potential adverse effects has been widely discussed
(Benn et al., 2010).
As nanoAg tends to precipitate and accumulate in the
sediment layer of aquatic systems, organisms such as cray-
fish that live at the bottom of water reservoirs and feed on
sediments, would be constantly exposed. In addition, aggre-
gation of certain NPs may enhance their ingestion and
diminish their retention thereby increasing their bioavaila-
bility (Ward and Kach, 2009). In view of this, filter-feeding
species such as crayfish may represent valuable models for
characterizing nanoAg bioavailability and its degree of
bioaccumulation. It was previously shown that aqueous
exposure of rainbow trout to silver nanoAg resulted in the
highest accumulation of nanoparticles taken up via the gut
as a function of drinking and/or feeding on sediment mate-
rial in the liver tissue (Scown et al., 2010).
Our results have clearly identified the induction of
oxidative stress in hepatopancreas (the analogue of liver in
fish) of nanoAg-exposed crayfish. Parameters used in the
present study to assess the presence of oxidative stress were
previously shown to be specific biomarkers changing under
the influence of different environmental pollutants in a vari-
ety of water organisms (Cossu et al., 1997; Borkovic et al.,
2005), including crayfish (Kovacevic et al., 2008).
A few investigations of nanoAg toxicity in aquatic
organisms have been published (for review see: Fabrega
et al., 2011), especially in invertebrates. However, previous
in vitro (Piao et al., 2011) and in vivo (Federici et al., 2007)
studies have shown that different types of nanoparticles can
lead to peroxidation of cellular membrane lipids. An
enhanced TBARS reaction in the hepatopancreas of
exposed crayfish identified in the current study confirms the
previous observations. Our results are also in agreement
with those of Ahamed et al. (2010b) who observed
enhanced lipid peroxidation in Drosophila melanogasterexposed to nanoAg. This suggests that the reaction is a spe-
cies-independent general reaction caused by NPs. More-
over, we also noticed that the level of protein-based ��SH
groups is lowered. This indicates oxidation of the ��SH
groups to S��S bridges. Mechanisms of lipid peroxidation
and protein oxidation under conditions of stress are the
result of oxidative stress induced by the imbalance between
production of ROS/RNS and cellular regulatory mecha-
nisms. NanoAg toxicity toward fish has been shown to be
connected with reduced oxygen uptake and induction of
oxidative stress (Bilberg et al., 2010). Moreover, overex-
pression of genes related to carcinogenic and oxidative
stress was observed (Chae et al., 2009).
We also observed dose-dependent activation of cellular
defense mechanisms in response to increased levels of
nanoAg. The increase in the total glutathione level is one
example. This may reflect the induction of de novo synthe-
sis of this main antioxidant. However, considering that
mechanisms of cellular membrane lipid peroxidation and
protein oxidation were activated, the level of de novo syn-
thesized glutathione appears to be insufficient to prevent
oxidative stress. Increase in total glutathione was not paral-
leled by the enhancement of its reduced form exhibiting
antioxidant activity, presumably due to the disturbed mech-
anism of its regeneration by GR.
Glutathione is the most important molecule in defense
against oxidative stress and cellular damage. Interactions
of the ��SH groups of glutathione with free radicals gener-
ated in cells under conditions of stress leads to the forma-
tion of disulfide bridges (GSSG). Glutathione together with
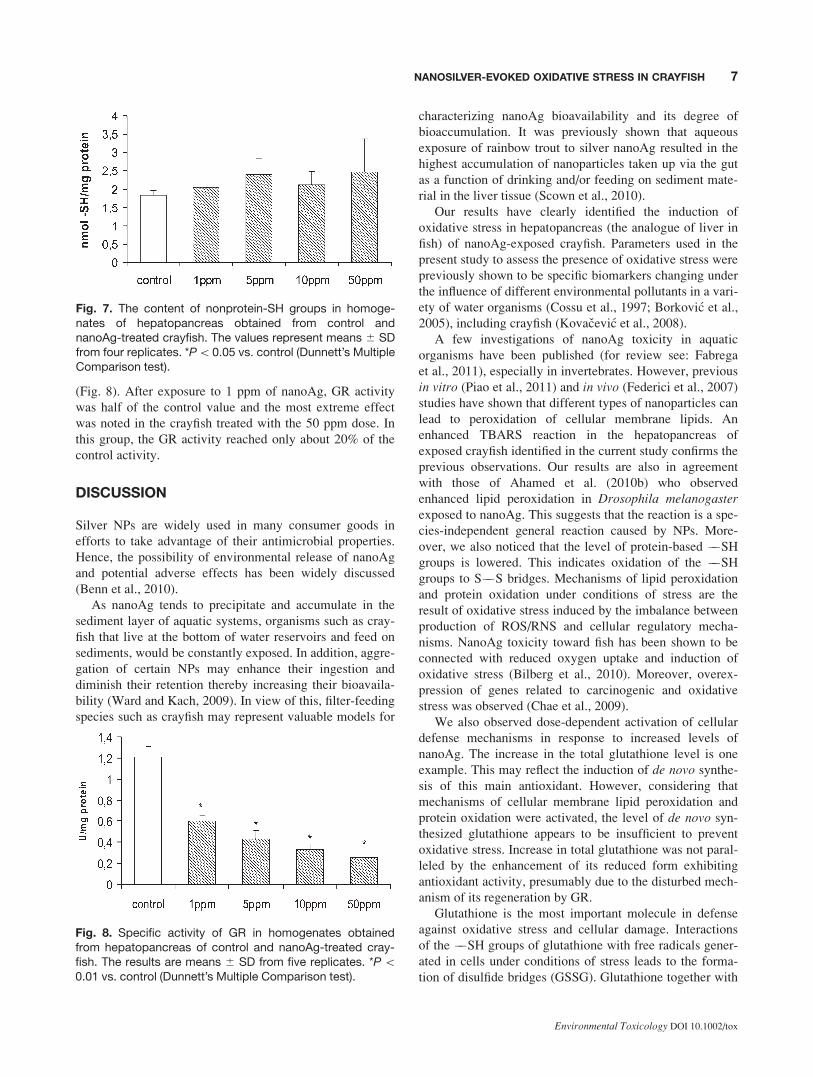
Fig. 7. The content of nonprotein-SH groups in homoge-nates of hepatopancreas obtained from control andnanoAg-treated crayfish. The values represent means 6 SDfrom four replicates. *P\0.05 vs. control (Dunnett’s MultipleComparison test).
Fig. 8. Specific activity of GR in homogenates obtainedfrom hepatopancreas of control and nanoAg-treated cray-fish. The results are means 6 SD from five replicates. *P\0.01 vs. control (Dunnett’s Multiple Comparison test).
7NANOSILVER-EVOKED OXIDATIVE STRESS IN CRAYFISH
Environmental Toxicology DOI 10.1002/tox
glutathione reductase which regenerates its reduced form,
provide a strong antioxidant defense system. Interestingly,
in the hepatopancreas of exposed crayfish, we identified a
change in GR activity which decreases in a concentration-
dependent manner. The induction of GR activity is used as
a biomarker of oxidative stress in aquatic organisms such
as crayfish (Borkovic et al., 2005, 2008). In our study, the
activity of this enzyme is strongly inhibited under condi-
tions of exposure to nanoAg. Although mechanisms of denovo synthesis of glutathione run properly and the total cel-
lular content of this antioxidant increases, lowered GR ac-
tivity leads to the impairment of the regeneration of its
reduced form (GSH) from oxidized disulfide (GSSG).
Thus, cells are not supplemented with enough reduced glu-
tathione. Moreover, depleted levels of protein-bound SH
groups and a lack of a parallel increase in nonprotein SH
groups, indicates that the GSH/GSSG system is not effi-
cient enough to buffer the reactive oxygen species gener-
ated by nanoAg.
Current tools for tracing NPs in biological systems are
based on measurements of light absorption, fluorescence
(e.g., autofluorescence or labeling with fluorescent probes)
or magnetic properties (Oughton et al., 2008). In the current
study, we employed confocal microscopy to take advantage
of the autofluorescence of nanoAg. However, it was shown
that complex organic matter may interfere with nanoAg
detection due to its own fluorescence (Cherukuri et al.,
2006; Mureau et al., 2007). The hepatopancreas contain
large quantities of organic matter since it is involved in the
processes of absorption and storage as well as metabolism
of such nutrients like lipids and glycogen (Fingerman et al.,
1967). This may explain the presence of fluorescent mate-
rial of relatively large size (oversizing Ag particles many
times and distinct in structure) in the hepatopancreas of
control crayfish (not experimentally exposed to nanoAg).
Presumably, these particles represent complexes of organic
compounds present in this organ. Exploiting the nanosilver
with red fluorescence was shown to be a useful method for
investigating the fate and transport of nanosilver in plank-
tonic and biofilm bacterial cultures (Choi et al., 2010).
However, current study indicates that the results obtained
by this method are not specific. Undoubtedly, the resulting
image of fluorescence in both control and exposed organ-
isms depends upon the type of tissue examined. Therefore,
in our opinion, this method has the limited utility for detect-
ing nanosilver in tissues of exposed organisms.
In conclusion, the results of the present study show that
silver NPs exhibit pro-oxidative potential in the hepatopan-
creas of exposed crayfish as evidenced by lipid peroxida-
tion and oxidation of free ��SH groups of proteins. We
suggest that this pro-oxidative activity is the result of inhi-
bition of GR activity. Decreased GR activity together with
the lack of simultaneous enhancement of non-protein SH
groups indicate insufficient level of reduced glutathione
available and needed for interactions with excessive ROS
production under conditions of nanoAg exposure. As very
little research has been done on nanoAg toxicity among in-
vertebrate aquatic organisms, our results may be valuable
from this point of view as well as in terms of considering
the increasing need to develop methods for detection and
quantification of the accumulation of NPs in exposed
organisms. The present study reveals the toxicity of silver
NPs to aquatic organisms and identifies nanoAg as a poten-
tial source of environmental risks. Thus, further assess-
ments of the harmful effects on aquatic organisms are
needed to better understand a realistic scenario of environ-
mental exposure to silver NPs and related health effects
(Fabrega et al., 2011).
The authors thank Dr. Małgorzata Frontczak-Baniewicz for
TEM assistance.
REFERENCES
Ahamed M, Alsalhi MS, Siddiqui MKJ. 2010a. Silver nanoparticle
applications and human health. Clin Chim Acta 411:1841–
1848.
Ahamed M, Posgai R, Gorey TJ, Nielsen M, Hussain SM, Rowe
JJ. 2010b. Silver nanoparticles induced heat shock protein 70,
oxidative stress and apoptosis in Drosophila melanogaster. Tox
Appl Pharmacol 242:263–269.
Asakawa T, Matsushita S. 1980. Coloring conditions of thiobarbi-
turic acid test for detecting lipid hydroperoxidas. Neurol
Toxicol 14:137–148.
Benn T, Cavanagh B, Hristovski K, Posner JD, Westerhoff P.
2010. The release of nonosilver from consumer products used
in the home. J Environ Qual 39:1875–1882.
Bilberg K, Malte H, Wang T, Baatrup E. 2010. Silver nanopar-
ticles and silver nitrate cause respiratory stress in Eurasian
























![[RAF CAREERS] What Careers are there in the RAF?](https://static.documents.pub/doc/80x56/55cf8f58bb61ebe4598b4842/raf-careers-what-careers-are-there-in-the-raf.jpg)