Masters of Applied Research HL84 Dominic Orth BAppSc (HMS) with Honours School of Human Movement Studies 2011 INTERACTING CONSTRAINTS OF DEFENSIVE PRESSURE AND BALL DISPLACEMENT TRAJECTORIES SHAPE LOCOMOTOR POINTING BEHAVIOURS IN ASSOCIATION FOOTBALL
Transcript
Masters of Applied Research HL84
Dominic Orth BAppSc (HMS) with Honours
School of Human Movement Studies
2011
INTERACTING CONSTRAINTS OF DEFENSIVE PRESSURE AND BALL
James Gibson (1979), founder of the ecological psychology approach to the
study of animal-environment relations, provided a theoretical rationale of how
animals are able to guide their actions relative to key surfaces, objects and
events in their environment. His account of emergent behaviour was predicated
on a theory of direct perception (Gibson, 1979). Direct perception proposed an
explanation of how properties in the environment are perceived to support
actions. From a Gibsonian perspective, the information available in the
environment does not need to be disambiguated by the Central Nervous System
(CNS) (Warren, 1998). Instead information is specific enough to be directly
percieved by an organism’s functionally adapted sensory-movement systems (for
alternative perspectives see, Norman, 2002; Zago, McIntyre, Senot, &
Lacquaniti, 2009).
Gibson argued that physical laws and properties of the environment, coupled
with the need to locomote for survival, served a role in the evolution of integrated
perceptual and action systems (Warren, 1998). Because lawful relationships 1 The key term, locomotor pointing, is defined as goal-directed gait that requires positioning of a body segment or held implement at a location in space and time. Locmotor pointing is provided a detailed definition in section 1.4 below.
19
exist between environmental properties and the structure of the surrounding
energy flows (Warren, 1998), the environment can unambiguously inform the
animal about the motion and location of objects and their own self-motion
(Williams, et al., 1999).
Gibson (1979) also proposed mechanisms for how humans perceived
information in the environment in order to coordinate contextual, role specific
actions. In the theory of Direct Perception, variants and invariants from the
surrounding energy flows are related to affordances for intentionally driven
behaviour.
1.2.2 Affordances and Intentionality
Variants and invariants in the energy flows surrounding the organism are
generated by movement. As the organism moves, at a superficial level,
structures in the informational array change relative to key surfaces or events
and are considered variant (Davids, et al., 2008). On the other hand, invariants
in the underlying structure remain (Davids, et al., 2008). Invariants represent
higher order properties constantly available to inform actors what behaviours are
occurring and are required in order to achieve performance goals (Montagne,
2005). Optical energy picked-up by visual perception systems may specify the
timing for an action such as positioning of a foot on a target at a specific location
in space (Warren, Kay, Zosh, Duchon, & Sahuc, 2001). For example, in the long
jump run-up the orderly nature of the expansion of the retinal image of the take-
off board2 might contain an optical invariant to support when, where and how to
begin preparing to position the feet for the take-off footfall (Fajen, Riley, &
From an ecological psychology perspective, the spatial-temporal specification of
an invariant when coupled with intentions of an animal, would be considered an
affordance for an animal to undertake related actions in preparation of carrying
2 In long jump, athletes attempt to, in the jump footfall, position their toe relative to the edge of a take-off board. In this footfall athletes generate a jump where the objective is to achieve as mush distance from the edge of the take-off board as possible. Athletes are penalised for positioning the toe of the jump footfall beyond the edge of the take-off board.
20
out an interaction with an affordance (Davids, et al., 2008; Montagne, 2005). By
definition, an affordance is an opportunity for action specified in the environment
relative to the organism’s personal constraints (including intentions) and current
state of movement (Gibson, 1979). Objects in the environment can afford certain
actions; balls on the ground are kick-able, balls in the air are catch-able or head-
able, and implements in the hands are throw-able (Fajen, et al., 2009). For
example, how kick-able a ball is, will be related to its mass relative to the force
an athlete is able to generate whilst this action is driven by the intent to meet
environmental challenges (Fajen, 2005). The implications of the use of
affordances in movement coordination indicate that organisms do not generate
actions according to an arbitrary measurement such as grams, rather, they do so
in a manner scaled from the reference point of the performer relative to the
environment (Turvey, 1992).
The possibility that affordances provide can help explain goal directed behaviour
was tested in a seminal study by Warren (1984) who undertook a stair climbing
experiment into whether actions could be informed by the limb lengths of
participants relative to the environmental properties of the stair raiser height.
Warren (1984) demonstrated key influences on decisional behaviour in a stair
climbing task at the performer-environment scale of analysis. He observed that
performance of participants was supported by the metrics of both the individual’s
leg length and the stair raiser height. He found a dimensionless ratio between a
participant’s leg length and the height of the stair. As the height of the stair was
systematically increased, a decision that the stair was no longer ‘step-up-able’
emerged at a critical ratio value (0.88), with participants spontaneously adopting
a four-limbed climbing style instead beyond this value. The scaling of actions
relative to body dimensions is an example of how movement solutions are ‘body-
scaled’ and are likely to be individualised.
In addition, decisions are also influenced by the action capabilities of the
individual performer whereby, action capabilities scale perception of the capacity
to produce a response to an environmental challenge (Ramenzoni, Riley, Davis,
Shockley, & Armstrong, 2008). For example, in football during penalty kicks,
research has shown that a goalkeeper’s movement time, constrains when they
21
initiate movement to intercept the ball relative to his/her own action capabilities
(Dicks, Davids, & Button., 2010). Movement initiation relative to action-
capabilities was shown in that goalkeepers with slower movement times tended
to move earlier and base their decisions on the perception of information
exclusively from the run-up of the penalty kickers. Whereas goalkeepers with
fast movement times delayed their movements, giving them the capacity to
observe information from the shooter’s run up and kicking action (Dicks, et al.,
2010). This tendency to delay actions relative to one’s own action-capabilities
was described by Dicks et al. (2010) as a reflection of an interaction between the
performer and environment, limited by a critical action boundary. The critical
action boundary biased performers’ actions toward remaining within a stable
performance region (or safety margin), ensuring that a performer retained access
to functional (and achievable) actions (Fajen & Devaney, 2006).
Although previous research on stair climbing and goalkeeping strategies in the
penalty kick have demonstrated how perception-action coupling supports
coordinated actions in dynamic performance environments, intentionality also
has an important role in how performers interact with affordances (Turvey,
1992). Intentionality refers to the specific intentions that each individual has at
any moment during performance, for example, to run as fast as possible or to run
with accuracy in order to prepare to place a foot on a target area in the
environment (Montagne, 2005).
The research discussed so far indicates that each individual will generate a
relatively unique coordinated movement pattern for the ‘same’ problem because,
movement solutions are both scaled to body dimensions and action capabilities
(Dicks, et al., 2010; Ramenzoni, et al., 2008; Warren, 1984) and interactions
with affordances can change depending on the intentions of the individual in a
given performance context (Montagne, 2005). Affordances have been shown to
be dynamic in the behavioural opportunities they provide (Dicks, et al., 2010).
Despite a location in space being a static surface, the intentions of a performer
can change what they afford (Maraj, 2002; Newell & Ranganathan, 2010).
Perception and action relative to key surfaces unfolds over the process of
carrying out a performance objective (Montagne, 2005). The moment-to-moment
22
nature of perception and action implicates an ongoing coupling between
perceptions of important information with subsequent movements. Control of
action by a perception-action coupling implies that adjustments will not be
produced unless the requirement is perceived as functionally necessary for
performance (Montagne, Cornus, Glize, Quaine, & Laurent, 2000). According to a
prospective control model, adjusting movements occur continuously throughout
the process of coordinating actions with the environment (Montagne, 2005). The
next section details theoretical arguments and research supporting a prospective
control of performance behaviour.
1.2.3 Prospective Control
Evidence for prospective control is fundamental to ecological psychology’s theory
of Direct Perception (Turvey, 1992). This is because if performers can be shown
to generate adaptive behaviour that is required and based on their current
behaviour, it provides evidence of a continuous and circular relationship between
perception and action, rather than a process interrupted by the need to recall
and initiate a specific movement plan (Montagne, 2005; Warren, 1998). Strong
evidence for prospective control has been observed in activities that involve
interception during tasks with severe spatio-temporal constraints, such as those
found in many sport contexts involving interceptive action (for a review see
Davids, Savelsbergh, Bennett, & van der Kamp, 2002).
For example, the study by Bootsma and van Wieringen (1990) exemplified the
role of prospective control in interceptive actions. In their work, highly
experienced table-tennis players performed fore-hand returns to a target location
on the other side of a competition table (Bootsma & van Wieringen, 1990). The
researchers found that despite variability observed at the initiation of
movements, these skilled players were able to modify the acceleration and the
orientation of the bat to achieve high level of timing and placement at the all-
important point of bat/ball contact (Bootsma & van Wieringen, 1990). Terming
the phenomena as ‘kinematic convergence’ (also termed funnel shaped control,
Montagne, et al., 2000), the largest variability in bat position occurred at onset of
stroke initiation and the lowest bat position variability occurred at the point of
bat/ball contact (Bootsma & van Wieringen, 1990). This finding of a pattern of
23
high to low amounts of variability in bat/ball positioning indicated that the
athletes were not just producing a routine action; they were adapting their
movements to achieve the task goal of hitting the ball. Through further individual
analysis, Bootsma and van Wieringen (1990) found that 2 of the 5 players
displayed the lowest amount of variability during the middle to final part of their
movement (Bootsma & van Wieringen, 1990). The individual analysis suggested
that some athletes were adapting their movements relative to the ball
throughout the entire action, implicating a prospective control strategy (Bootsma
& van Wieringen, 1990).
1.2.4 Theoretical Summary
1.2.4.1 Ecological Psychology
To summarise so far, the environment is considered to contain sources of
unambiguous information that has influenced the evolution of perceptual and
action systems (Warren, 1998). Evolutionary scale adaptations has subsequently
been shaped to directly perceive a variety of continuously available sources of
energy to support perception-action cycles and for humans include vision, haptic,
auditory, olfactory and proprioception (Davids, et al., 2008). Coordinated
solutions to task specific and environmental affordances for action have been
shown to be individualised (Dicks, et al., 2010; Warren, 1984). Behaviour that
reflects individualisation of movement patterns has been observed in the
influence of body-scaling on actions (Dicks, et al., 2010; Ramenzoni, et al.,
2008), implying that unique coordination patterns are scaled to body dimensions
(Warren, 1984) and action capabilities of individual performers (Dicks, et al.,
2010). The ability to adapt movements continuously throughout interceptive
actions by use of a prospective control strategy has been outlined (Bootsma &
van Wieringen, 1990; Montagne, 2005; Turvey, 1992). In summary, an argument
has been proposed to account for how information on a variety of factors is
detected to control action, and vice versa, in terms of a perception-action cycle.
Control using a continuous perception-action cycle relies on:
an unambiguous link between the status of the environment and that
which is perceived (Jacobs & Michaels, 2007; Turvey, 1992);
24
the availability of higher order invariants that can specify current and
required actions (Williams, et al., 1999);
coordinated actions being scaled by body dimensions and action
capabilities relative to the environment (Dicks, et al., 2010; Turvey, 1992;
Warren, 1984),
a cyclic, interdependent process of movement for perception and
perception for movement (Bootsma & van Wieringen, 1990; Montagne,
2005), and;
the adaptation of movement based on the current and required state of
the movement system relative to environmental challenges (Turvey,
1992).
The ideas given coverage from ecological psychology’s theory of Direct
Perception describe how information can help regulate action in a manner
spread over the performer-environment system. Clearly Gibson (1979) did not
argue that performer-environment interactions were regulated in a manner akin
to the use of a central executive or decision maker (Warren, 2006). However,
additional perspectives are needed to understand other constraints on the
emergence of performance behavior, such as instructions and intentions. For
instance, just because an action is possible according to affordances, does not
mean it will emerge. What constrains and drives perception and action and how
is this related to emergent performance? In the following section, how
constraints of performance contexts impinge on an individual’s perceptual-action
cycles is considered (Glazier & Davids, 2009). It is argued that this dynamic
process results in the emergence of self-organised patterns of movement
coordination (Glazier & Davids, 2009).
1.3 COMPLEX DYNAMIC SYSTEMS
Studies of complex systems occur across various disciplines including for
example, sport (Gréhaigne, Bouthier, & David, 1997; Hristovski, Davids, Araújo, &
Button, 2006; McGarry, 2005), skill acquisition (Chow, et al., 2011; Davids, et
al., 2008), human movement systems (Kelso, 1995; Schöner & Kelso, 1988),
Normand, 1980), and social networks (Miller & Page, 2007). Generally, complex
systems are defined as systems made up of interacting and interdependent units
that constrain each other’s behaviours across multiple levels and timescales
(Davids, et al., 2008; Newell & Liu, 2001). Despite apparent differences, such as
being a chemical or biological system, complex systems share key characteristics
that extend across the discipline of observation. These characteristics include
self organisation, local interaction rules which can lead to large scale system
changes and emergence. In this section, characteristics of complex systems are
defined alongside empirical examples in biological3 and sport performance
systems.
1.3.1 Complex Systems in Nature
Self-organisation is defined as the process of pattern-formation that occurs in
complex systems solely from the numerous local interactions within the system
and without intervention from external directing forces (Camazine, et al., 2001;
Kauffman, 1993). Rather than requiring an external component to direct
perceptions and actions of organisms, local information rules specify the
interactions among the components that make up the system (Camazine, et al.,
2001). Complex system principles (i.e. self organisation, local interaction rules,
emergence, stability, instability, large scale changes, heterogenous responses)
can underpin the coordination tendencies shown by interacting components that
make up complex systems. Considering pattern formation as a coordination
tendency reflects how individual components can function independently but can
come together and coordinate actions as they become informationally-coupled
(Kelso, 1995; Kelso & Engstrom, 2006).
The study and measurement of complex sysems begins with the observation of
patterns that occur among a systems components (Kelso, 1995). The patterns
displayed by a complex system can show emergent properties, and it is largely
these properties that generate the research interest into complex systems
3 The choice to focus on how complex system properties relate to biological systems in particular was made because, biological systems differ fundamentally to physical systems in that: (i) sub-units exhibit greater complexity in biological systems, and; (ii) the interactions are between inanimate objects in physical systems (Camazine et al., 2001).
26
(Camazine, et al., 2001; Kelso, 1995). This is because the patterns of behaviour
appear to equal much more than the sum of the individual parts that contribute
to their occurance (Camazine, et al., 2001). The local interactions can
sometimes be identified to follow simple rules of thumb which paradoxically
generate highly complex, functional behaviours (Camazine, et al., 2001). A key
objective of complex systems research is to identify the rules that regulate the
interactions between the elements in a system responsible for generating
emergent properties (Camazine, et al., 2001). Consider for example the building
of the large African termite mounds of the genus Macrotermes depicted below in
Figure 1.1. These mounds reflect sophistocated structures that regulate heat
critical to the survival and reproduction of the inhabitants (Bristow & Holt, 1987),
contain a variety of purpose built chambers (Rouland, Lenoir, & Lepage, 1991),
generate a self-sufficient food supply (Rouland, et al., 1991) and are immense in
size relative to the inhabitants who build them.
27
Figure 1.1 An image of a termite mound of the genus Macrotermes
28
Millions of fairly homogenous units (termites) interact and build these structures
over numerous lifetimes from the intial beginnings of a queen-king combination
(Camazine, et al., 2001). How do termites know where to begin, or when to stop?
Do each follow an individualised template?; are they guided by the queen or, are
actions influenced by self-organising principles, such as through information
governed interactions with local concentrations of pheromone laid down by other
ants undertaking nest builing activities (Camazine, et al., 2001)? Understanding
what governs emergent behaviours can reveal how natural systems, that cen be
made up to millions of degrees of freedom, as in termite colonies, exploit
surrounding physical and informational resources in efficient, functional and
creative ways (Camazine, et al., 2001). Whether self-organising features of
natural biological systems can inform sport performance is predicated on the
notion of whether human movement systems also display similar emergent self-
organised qualities.
1.3.2 Self-Organisation in Sport Performance
To consider the feasability of a complex systems approach in sport performance
contexts, research has imported systemic theoretical perspectives and methods
to consider human movement systems as a complex system in their own right
(Davids, Glazier, Araújo, & Bartlett, 2003). For example the human body can be
considered as made up of numerous heterogenous parts, with some 102 joints,
103 muscles, 103 cell types and 1014 neurons and neruonal connections, whose
interactions contribute to a functional, neurobiological movement system (Kelso,
1995). Extending this within individual systems approach, has been the
conceptualisation of between human interactions as a complex system (for
reviews see Marsh, Richardson, Baron, & Schmidt, 2006; Oullier, de Guzman,
Jantzen, Lagarde, & Kelso, 2008). From a systems perspective, the interactions
between team members on sport fields provide a context rich with emergent
patterns of interpersonal coordination tendencies (McGarry, 2005). As athletes
support their decisions on the local information available (i.e. distance to the
nearest team mate, an area of empty space, the stumble of a defender), a global
pattern often emerges, with no one player responsible for orchestrating the
29
functional structure of an offensive or defensive system (Gréhaigne, et al.,
1997).
Important questions that systems theorists are interested in posing regarding
sport performance in team games include: how do humans exploit the available
physical and informational resources in performance contexs in such efficient,
functional and creative ways? Do these performance behaviours display
emergent properties and if so, what local interaction rules govern coordination
between humans, their environments and tasks? Can such insights be
harnessed to promote emergence in performance behaviour through
theoretically driven design of practice settings? (Chow, et al., 2011; Davids, et
al., 2008; Renshaw, 2010).
Early work applying complex systems perspectives to sport have used a dynamic
systems approach to consider sport performance. Outlined for example by
McGarry et al. (2002), the dynamic systems approach involves an emphasis on
mapping the possible states of complex systems in terms of a numerical phase
space (Davids, et al., 2008). By definition a dynamic system is any system that
evolves in time under the action of a deterministic or stochastic (random) law or
rule (Kelso, 1995). Two key components give a prediction of a systems dynamics
(Kelso, 1995). First, a state vector (a collection of state variables) describes the
state of the system at any instant of time (Kelso, 1995; Newell & Liu, 2001).
Second, a rule or function predicts/controls where the system will be in a future
instant of time, given that the current state is known (Kelso, 1995; Newell & Liu,
2001). By observing a system as it responds to changing circumstances, the
constraints important in shaping the organisation of functional behaviours of the
system can be identified (Kelso, 1995; Kelso & Engstrom, 2006). Coordination
tendencies revealed this way are considered functional in that they allow an
organism to function more effectively in its particular performance environment
(Davids, et al., 2008).
Due to the difficulty of mapping a numerical phase space in sport contexts,
dynamic systems approaches in team sport have tended to be qualitative
(McGarry, Anderson, Wallace, Hughes, & Franks, 2002). For example, McGarry et
al. (2002) characterised football as a dynamic system made up of two sub-
30
systems (i.e. each team). The system as a whole displays periods of stability
(Gréhaigne, et al., 1997), transient periods of disorder (Hughes, Dawkins, David,
& Mills, 1998) and complete break downs in structural organisation (Gréhaigne,
et al., 1997). McGarry et al. (2002) speculated that simple local interaction rules
might govern these states. For instance to generate team structural order and
fulfill a defensive role players might tend to keep the distance to team members
relatively consistent or reduce distance to near opponents (McGarry, et al.,
2002). While on the other hand to fulfill an attacking role and generate
opportunities for a goal, team members might attempt to fill available space and
undertake actions that generate distance between themselves and a defensive
player (McGarry, et al., 2002).
Passos, et al. (2009) quantitatively extended understanding in the field of
dynamic systems theory research in sport performance settings provided by
qualitative analysis. Using a dynamic systems approach to understand try scoring
in rugby, Passos et al. (2009) analysed coordination tendencies between players
as it emerged from rugby 1v1 situations at the defensive try-line. In their
experiment, an attacker was positioned 10 m from the try-line where a defender
was positioned to protect it. The attacker had an area of 5 m width in which to
attempt to run past the defender to score a try. In this situation, although the
attacker’s desired to have a large distance between themselves and the
defender, they were pressured by the task objective to reduce this distance
(Passos, Araújo, Davids, Serpa, et al., 2009). At a specific distance to the
defender (a value of 4 m), the mutual actions of the players became coordinated
and very instrumental in determining the performance outcome (Passos, Araújo,
Davids, Serpa, et al., 2009). That is, within close proximity actions became highly
correlated and if the attacker could generate a high enough relative velocity with
the defender inside a 4 m proximity, then the attacker could take advantage of
the space surrounding the defender and move past (Passos, Araújo, Davids,
Serpa, et al., 2009). If the defender could keep the relative velocity low, there
was a greater probability of success in intercepting the attacker and maintaining
stability in the dyadic system (Passos, Araújo, Davids, Serpa, et al., 2009). The
relatively simple and nested rules Passos et al. (2009) uncovered for attackers
to get past a defender in 1v1 sub-phases of team sports, suggested that an
31
attacker should aim to achieve a velocity that is higher relative to an approximate
defender when within 4 m in distance. Behaviours that might reduce this relative
velocity within this critical 4 m region should be avoided. Any action the defender
shows that might be exploited to increase the relative velocity should be
attended to. These straightforward pedagogical instructions can direct an
individual performer’s attention to important task specific information and
actions. Importantly, information governed instructions enables the performer to
generate prospectively controlled, creative and emergent behaviours to achieve
task objectives (Chow et al., 2007; Passos & Araújo, 2008).
Passos et al. (2009) showed how examining a situation that commonly occurs in
sport (known as a representative sub-phase, for a review of these methods see
Davids, Button, Araújo, Renshaw, & Hristovski, 2006), can reveal insights into
how constraints interact to influence coordination tendencies and ultimately
sport performance outcomes. The interaction between the task and the
performers constrained the behaviours that emerged. The task constraints
established a boundary to the performance context that resulted in emergent
coordination tendencies (Passos, Araújo, Davids, Serpa, et al., 2009). As
behaviours emerged under the pressure of constraints, functional aspects of the
performance context became apparent (i.e. interpersonal distance was critically
nested with relative velocity). The constraints that influence movement
performance were first described by Newell (1986) and provide a model for
understanding how coordinated behaviour emerges from complex dynamical
systems. The important role and characteristics of constraints are defined and
described in the following section.
1.3.3 Constraints on Movement Coordination
Newell (1986) provided a model summarising the categories of constraints on
behavior (see below in Figure 1.2). It has since been adapted to show how
constraints interact to impinge on circular information-movement couplings
(Glazier & Davids, 2009). There are three categories of constraints to consider
32
including: personal, environmental and task constraints (Newell, 1986).
Figure 1.2 Newell’s adapted Model of Constraints on Performance
Personal constraints refer to existing structural and functional characteristics of
an individual (Chow et al., 2006). Structural characteristics include body
composition, height and limb lengths. Functional characteristics refer to
connective strength of synapses in the brain, motivations, emotions, intentions
and cognitions (Chow, et al., 2006). Environmental constraints may be physical
(mechanical), informational or social in nature (Chow, et al., 2006). Physical
constraints might include forces generated through the body such as ground
reaction forces. Informational constraints might include those such as ambient
light, temperature, sound, wind and moisture. Social constraints might be factors
such as peer groups, parents and socioeconomic status that act on performance
(albeit over a somewhat larger timescale) (Chow, et al., 2006). Finally, there are
task constraints. These tend to be much more specific to the performance
context and include rules, equipment, opponents or field dimensions (Chow, et
al., 2006).
It is important to note that constraints are interrelated and interdependent in
how they impinge on behaviour (Davids, et al., 2008). This can make it
somewhat difficult to separate one type of constraint from another because as
one constraint changes, this will influence another, often in non-linear ways
(Davids, et al., 2008; Newell & Liu, 2001). For example how a performer might
interact with task constraints might change based on contextual constraints
33
previously not available to the performers. Guerin and Kunkle (2004) highlighted
how task constraints are dynamic and can emerge and decay over time. Consider
a goal-keeper’s task to kick a ball from a goal-kick situation. The goal-keeper
might kick the ball long in order that the ball is contested in the other team’s half
or alternatively, kick the ball short in order that the receiving player might work
the ball up the field with a succession of passes between team-members. Early
in the game, the goal-keeper might be more inclined to pass short in order that
his/her team retains possession of the ball and have the opportunity to generate
goal scoring opportunities. If, later in the game the goal-keeper’s team is holding
on to a game winning lead by points, new task constraints have emerged, he/she
might consider it too risky to chance a short pass, and instead play the ball long.
This is an example of how constraints can emerge (i.e. need to score goals) and
decay to be replaced by a different one (i.e. prevent goal scoring opportunities for
the other team) through the outcomes of interactions that occur over time (i.e.
point scoring).
Another important feature of constraints on behaviour is that they can act over
different, shorter and longer time-scales relative to the time that a performance
occurs over (Newell & Liu, 2001). For example in basketball, tallness carries
certain performance advantages and could lead to institutions choosing players
on the basis of their height (Phillips, Davids, Renshaw, & Portus, 2010). This in
turn would change the environment for players train and perform under. As a
consequence perceptual and action learning and coordination would be shaped
differently over time to reflect the homogenisation of opposition characteristics
(Cordovil et al., 2009).
1.3.4 Theoretical Summary
1.3.4.1 Complex Dynamic Systems
To summarise, complex systems theoretical principles have been shown to play
an important role in emergent performer-environment coordination (Camazine, et
al., 2001; Cordovil, et al., 2009; Davids, et al., 2008; Newell & Liu, 2001;
Passos, Araújo, Davids, Serpa, et al., 2009). The value and potential
mechanisms of emergent properties in biological systems have been outlined.
34
Complex systems can show functional properties of self-organised behaviour
based on simple rules governing local information based interactions (Camazine,
et al., 2001). The different types of constraints (personal, environmental and
task) impinging on the self-organisation of behaviour have been outlined (Chow,
et al., 2006; Newell, 1986). Their interrelated and interdependent
characteristics, ability to emerge and decay, and to act over different time-scales
have been described (Cordovil, et al., 2009; Guerin & Kunkle, 2004; Newell &
Liu, 2001; Phillips, et al., 2010).
In sport performance, distinctive patterns emerge from the interdependence and
interactions among players, their environments and tasks (Davids, et al., 2008).
Despite the apparent order in performance behaviour, there is no single dictating
source responsible for imposing it, rather, human performance systems are self-
organising systems under constraint (Kauffman, 1993). The role of complex
systems theory in understanding how organism-environment interactions are
constrained can be summarised in that functional coordination tendencies
emerge:
from interdependent and interrelated interactions of task, environmental
and personal constraints that act across multiple timescales (Davids, et
al., 2008; Newell & Liu, 2001);
displaying characteristics of dynamic, self-organising systems (Passos et
change depending on the contextual focus that instructions provide
the performers (Maraj, et al., 1998).
57
1.5 CONCLUSIONS
1.5.1 Research Issues Examined in this Thesis
This thesis will take an ecological dynamics approach in considering how
constraints interact to shape the coordination process of performer-environment
relations. The extant locomotor pointing research has focused on single
constraint based manipulations with a bias toward manipulation of visual
information. This research bias toward manipulating visual information has
distinct limitations in that visual information in typical sport performance and
day-to-day contexts is normally not withdrawn. In fact to the contrary, most urban
environments and organised sports carefully monitor ambient light and take
measures to ensure its availability. This provides a strong case for ensuring
visual information be maintained, rather than occluded in an experimental
testing environment.
Previous research in ecological dynamics in sport performance has shown how
interpersonal constraints generate coordination tendencies when the constraints
are representative of the performance context (Passos et al, 2009). In
performance settings, constraints provide a drive to undertake goal directed
locomotion, often these constraints are dynamic in nature and capable of
being fluid. For example in sport, perceptions and actions emerge relative to
the actions (and perceptions) of team-mates and/or opponents (Fajen, 2009).
How do dynamic task constraints influence emergent coordination tendencies
when compared to task instructions?
As a task vehicle, team games like football can provide a performance context to
empirically evaluate the influence of interpersonal constraints on emergent
locomotor pointing behaviour. Football is abundant with examples of goal
directed gait under the influence of dynamic, interpersonal constraints. For
example, attacking players often run towards or dribble with a ball in order to
make a cross pass to team-member inside the penalty area. This emergent sub-
phase of the game often occurs whilst an opponent pursues the attacker.
58
By using a representative sub-phase from football a number of important
theoretical and practical questions can be considered. This thesis will consider
the following questions:
Does running to make a cross pass in football involve the same funnel-
type control strategy as observed in other locomotor pointing tasks with
nested actions at the end?
How do interpersonal constraints differently influence movement
coordination in locomotor pointing tasks when compared to instructional
constraints alone?
The influence of instructions in comparison to interpersonal and task constraints
will be evaluated by measures on the following dependent variables: (i) an
attacking player’s patterns of foot/ball distance variability when running to cross
a static ball, (ii) successive step length patterns of the attacking player, and (iii)
ongoing horizontal displacement velocity of the attacking player. If instructional
constraints influence coordination in the same manner as interpersonal
constraints then, these dependent variables should not significantly differ
between conditions. On the other hand, if there are significant differences in the
attacking player’s patterns of foot/ball variability, step length control patterns
and displacement velocity, this will reflect a coordination tendency that emerges
under the flux of sub-phase specific interpersonal and task constraints.
Additionally, if coordination is scaled relative to affordances then, if a defender is
closer to the attacker, a spatial constraint should emerge to change behaviour in
a functional manner.
In the chapters that follow two studies are presented to address the questions
raised. In Chapter 2, a study is presented which examines the influence of
instructional constraints when compared to interpersonal constraints during a
football run up and cross. In Chapter 3, to consider how the task constraint of
dribbling a football ball might influence coordination is then considered under
the same manipulations of Study 1. In the final chapter, Chapter 4, a reflections
section will be presented detailing the contributions and considerations for
future research generated by this Master’s thesis.
59
Chapter 2: Study 1
2. EFFECTS OF PRESENCE AND PROXIMITY OF THE NEAREST DEFENDER ON A
PERFORMER’S GAIT PATTERN WHEN RUNNING TO CROSS A STATIONARY BALL
2.1 INTRODUCTION
Locomotor pointing research typically has relied on instructions (Bradshaw &
Sparrow, 2001; Maraj, et al., 1998), or natural task constraints (Lee, et al.,
1982; Montagne, et al., 2000) to make tasks meaningful to participants during
coordination of goal directed gait. In many day-to-day and sport performance
activities, goal directed gait is under the pressure of constraints that are
dynamic. Dynamic constraints are not commonly observed in the extant
locomotor pointing research (Fajen, et al., 2009; Montagne, 2005; Zago, et al.,
2009). However, during goal directed gait, constraints can change over the time
taken to perform the task. For example, a human aiming to cross a road may be
driven by multiple constraints. Some may be static, such as invariants (Williams,
et al., 1999) in the environmental surface layout (for example the target curb on
the opposite side of the road). Others may be dynamic (Fajen, et al., 2009), and
alter gait patterns significantly, such as avoiding collision with an approaching
vehicle.
Fajen (2005) outlined a basis for affordance-based control that included
perceptual calibration to a critical action threshold. The critical action threshold
presumably might be scaled to the action capabilities (Ramenzoni, et al., 2008)
and body dimensions (Warren, 1984) of individuals, to separate those actions
which are possible from those that are impossible (Fajen, 2005). This critical
region has been shown to influence behaviours in drivers’ braking to avoid a
collision (Fajen, 2005) and in interceptive actions in sport (Dicks, et al., 2010). In
sport, performance tasks often involve locomotion toward key objects and events
(Fajen, et al., 2009). If performers are under influence of an affordance-based
60
control mechanism, performers should perceptually engage with this critical
action threshold (Fajen, 2005). Evidence of the perception of a critical region
(either accurately or inaccurately by the person) would be implicated via its role
in constraining decisional behaviours (i.e. emergent performance actions) (Dicks,
et al., 2010). For example, in football as a player approaches a ball under
defensive pressure, perception of whether he/she can reach the ball prior to the
defender would be shown in the action of attempting to reach the ball first.
Should the attacker perceive he/she has the time to carry out an additional task
(for example he/she may attempt a cross pass), this would be shown by the
attempt at carrying out the goal directed kicking action. Therefore, the decision
by the attacker to attempt to cross the ball would be due to: (i) the distance and
behaviour of the defender affording the attacker to reach the ball first, and (ii)
that the affordance is perceived (Fajen, 2005). Affordance-based control would
suggest that decisions are based on both what oneself is capable of achieving
and on the basis of what the defender is capable of achieving. The relative action
capabilities of an attacker and defender would together influence a critical
boundary. An attacking player who is perceptually calibrated to the critical region
would therefore scale his/her possible actions relative to the defender’s possible
actions (Fajen, 2005). It is likely that locomotion behviours in sport performance
contexts such as a football match emerge scaled relative to interpersonal
affordances, although, this idea has largely gone unexplored in the extant
research (for an exception see Dicks et al, 2010).
An experiment was designed with the aim of considering the difference between
static task constraints and dynamic task constraints that accommodated the
possibility of affordance-based control into the experimental design. An emergent
sub-phase from football was chosen whereby an attacking player runs down the
sideline in order to make a cross pass back toward the penalty area in order to
generate a goal scoring opportunity. This situation commonly emerges in football
matches.
Players were asked to undertake this task with and without the presence of
defensive pressure. It was expected that instructional task constraints would
generate functionally different player-ball coordination tendencies when
61
compared to coordination generated when dynamic interpersonal information
was present pressurizing the attacker whilst undertaking the cross. By increasing
the defensive pressure the aim was to observe whether an attacker would attend
to a critical action threshold by moving faster. Whether players perceive and act
according to a critical threshold informed by both, their own and a defender’s
action capabilities was also considered by scaling a defender’s interpersonal
starting position to two different distances. If the players scale their actions
relative to a perceived critical threshold, it was anticipated that players would
attempt to give themselves a certain amount of time to carry out the task. This
would be reflected in the attacking player’s displacement velocity being higher,
the closer that the defender is initially positioned.
Furthermore, how the defender might influence the player/ball coordination
tendencies was also of interest because it might be that attackers quantitatively
change how they manage the position of their footfalls when approaching a ball
under defensive pressure. Montagne et al. (2000) demonstrated that there is a
link between the amount of variability in a given trial and the amount of
adjustment that a person undergoes when undertaking a locomotor pointing task
at maximal velocity. Specifically Montagne et al. (2000) showed that the greater
the variability in footfall placements, the greater the amount of adjustments
people make when approaching the nested task. Scott et al, (1997) provided
evidence that expert long jumpers functionally spread adjustments between foot
placements over as many footfalls as available in the zeroing in phase of the
long-jump. If football players run faster under defensive pressure, then it might
be expected that the management of foot positions may become much more
important for success in the kick. Therefore, football players would be expected
to more functionally position their feet relative to the ball throughout the entire
run-up in a manner important for beating a defensive player, as opposed to
merely accurately pass a ball. A reduced level of foot/ball6 distance variability
during the run-up would be important so that the amount of adjustments at the
zeroing in phase would be manageable whilst running at the higher velocity
required to complete the task before the defender could intercept the ball. This
hypothesis would be confirmed if the magnitude of the standard deviations
6 The term foot/ball distance signifies the foot/ball distance value at each footfall placement.
62
around the mean foot/ball positions were reduced when attackers are under
defensive pressure due to players running faster during the task under increased
defensive pressure.
In summary, the aims of this experiment were to consider: (i) whether the
running to complete a football kick is under similar funnel shaped control as
other locomotor pointing tasks, (ii) how movement coordination is influenced by
instructional task constraints compared to dynamic interpersonal constraints,
and (iii) whether players attend to a critical threshold informed by the action
capabilities from themselves and their opponent.
2.2 METHODS
2.2.1 Participants
Participants (n = 8) were members of a football school of excellence program
(average age = 15.25 yrs, SD = 0.46 yrs) and had on average of 8.25 years (SD =
2.12 yrs) of competition season experience. The demographic details of
participants are summarised below in Table 2.1. The study was approved by a
University Ethics Committee. Participants along with parent guardians signed
information and consent forms prior to undertaking the experiment.
Table 2.1 Participant age, competition experience and relevant anthropometric details
Participant (ID)
Age (yrs)
Competitive Seasons (yrs)
Height (m)
Hip-to-Foot length (m)
Weight (kg)
1 16 5 1.84 0.81 80.5 2 15 8 1.81 0.98 68.6
3 15 9 1.84 1.07 60.3
4 16 10 1.82 0.93 80.0
5 15 10 1.82 0.95 59.0
6 15 5 1.73 0.86 68.4
7 15 9 1.80 0.94 63.9
8 15 10 1.60 0.87 53.5
Mean 15.25 8.25 1.78 0.93 66.78
63
2.2.2 Task
Below Figure 2.1 depicts the nature of the task and the roles of each player
involved in the experiment. Broadly speaking the task involved an attacking
player sprinting down the side-line and toward the opposition by-line where a
stationary ball was positioned in order to cross it back toward a team-member
positioned at the opposition penalty spot. The experimental manipulations
involved changing the level of defensive pressure by having the attacker
undertake the task with: (i) no defensive pressure, (ii) with a defender initially
positioned at a large distance (with a 20% disadvantage, refer below for an
explanation of how this value was arrived at), and (iii) with a defender initially
positioned at a close distance (with a 10% disadvantage). The defender was
always positioned at a disadvantage relative to the attacker and the position of
the ball. That is, the attacker was always able to get to the ball first based on the
action scaling procedure used.
Figure 2.1 Schematic of the experimental task in Study 1. B = Ball, A = Attacker, GK = Goal-keeper, R = Receiver, D = Defender. Only the defender’s involvement was changed across the different levels of defensive pressure (either absent, far or near), all other variables were held constant. The numbered scale reflects units of distance in metres.
64
To precisely scale the defender’s starting distance to the ball, pilot work
established: (i) the time it took each attacker to sprint 20 m and cross a ball, and
(ii) the time it took each defender to sprint 20 m. Referring to Appendix A for
formulae, the pilot data provided the information necessary to action-scale the
defender’s initial starting distance from the ball relative to the attacker. Because
the defender was scheduled to arrive after the attacker by a precise amount, the
attacker-defender dyads should not have been mismatched due to the action
capabilities of either individual and it could be predicted the attacker would
arrive at the ball first and with enough time to carry out the kicking task.
Task instructions for each player in the experiment were as follows. The attacker
was instructed to sprint to the ball (denoted B below in figure 2.1) and cross it
back towards the penalty spot. A receiving player (denoted R) was the target to
receive the crossed ball at the penalty spot. The receiver was instructed to time
the run to receive the ball at the penalty spot and attack the goal. A goal-keeper
(denoted GK) was instructed to protect the goal. In conditions where one was
present, the defender (denoted D) was instructed to meet the attacker at the
earliest point and prevent the cross within the laws of the game. To enforce the
laws of the game a referee was positioned in the field. The task was initiated at
the attacker’s discretion after a signal from the referee.
In interpreting the data, it is important to note that participants were not given
specific instructions on how to regulate footfall placement and velocity during the
run up. The behaviours of participants observed in this study were emergent
under the interacting constraints of performance.
2.2.3 Apparatus
The experiment was undertaken on the participants’ regular training field to
ensure familiarity (this was a grassed outdoor football field), at a similar time of
the day between sessions. Observations between sessions were recorded within
a one week time frame. A regular competition approved size 5 football ball was
used. This was positioned 3 m from the by-line and 4 m from the side-line (to
65
ensure consistent ball placement a spot was marked on the grass). Referring
below to Figure 2.2 for dimensions of the task environment, the attacker was
asked to pass the ball a distance of approximately 28 m perpendicular to their
starting position. The target was the penalty spot which was marked with white
spot paint. As shown in Figure 2.2, the position of the defender was manipulated
along a line taken from the ball that intersected with the corner of the 18-yard
box. For health and safety reasons, trials were only undertaken and observed in
dry weather conditions. Participants wore the same clothes and equipment as
they normally would for competition (including studded boots and shin pads).
Figure 2.2 Schematic of the experimental task and apparatus dimensions. A = Attacker, D = Defender, B = Ball, m = metres.
66
2.2.4 Data Capture
Two cameras were positioned perpendicular to the running direction of the
attacker (Sony HDR-XR520V and Sony HVR-V1P). The cameras were positioned
10 m apart, at 15 m from the side of the run and at a 5 m elevation relative to
the running surface (similar to methods by Maraj et al, 1998). The cameras were
zoomed to collectively capture an area of 25 m by 5 m. An overlap in the visual
angle of each camera allowed the entire run-up and cross action to be captured.
Post processing cut the footage at 25 Hz and synchronized it using an LED light
in the shared visual angle (files were saved in *.AVI format using Final Cut Pro 7
software (Final Cut Pro, Apple inc, Cupertino, CA)). Although no explicit
instructions were given, this was the area in which the attackers were expected
to remain within during the performance of the task. This area was surrounded
by high visibility markers that provided control points needed for two-dimensional
direct linear transformation (2D-DLT; these procedures first described by Abdel-
Aziz & Karara, 1971). This was performed using custom-built and pre-validated
software for these purposes (Digipan software).
High visibility markers (2.5 cm wide x 5 cm height) were placed on three
positions of each foot of the attacker. These were: the center of the heel, 2.5 cm
from the heel, and 5 cm from the heel. The position of the marker 2.5 cm from
the heel was the reference point of the foot used to calculate the distance of the
foot to the ball. The distance from the foot to the ball was calculated for each
footfall taken by attackers across all 96 trials. The foot/ball distance value at
each footfall placement was then used to obtain the standard deviations (SD) of
each foot position for each participant in each condition.
Measures of the attacker’s displacement velocity was recorded using the known
distance travelled and the number of frames between which the attacker was in
mid-swing of their gait cycle (as per Berg & Mark, 2005; and Lee, et al., 1982).
The central moving differences method was used to derive the velocity of the
attacker at each footfall (subsequently the velocity at the first and final footfall
were unknown) (Hamill & Knutzen, 2003). Pilot work established accuracy using
the method proposed by Bradshaw and Sparrow (2001). Shoes positioned at
known locations on a tape measure inside a number of locations throughout the
67
capture field were digitized and the digitized coordinates were compared to the
tape measured coordinates. The mean differences (n = 13) of this procedure
between the real and digitized points were found at 1.53 ±.63 cm. These levels
are similar across studies of this type (e.g. Lee et al, 1982, Berg et al, 2005).
Since the approximations of the foot positions were made using 2D-DLT
procedures some key assumptions needed to be met (Duarte et al., 2010).
2D-DLT is a valid method of reconstructing the locations of objects in a digital
image. To define the location of a point of interest in an image using 2D-DLT a
minimum of four points in the image and the distances between them must be
visible and known. These points must be collectively non-collinear to each other
and co-planar to the point of interest. The establishment of comprehensive
experimental design protocols regarding control points, the point/s of interest
and operation of the camera can maximise the accuracy and reliability of the
technique. These requirements for accurate and reliable application of 2D-DLT is
summarised below in
Figure 2.3.
68
Figure 2.3 The assumptions regarding linearity and planarity of two-dimensional direct linear transformation (2D-DLT). The left column shows conditions that violate assumptions of 2D-DLT whilst, the right column shows conditions that meet the assumption. The point of interest is marked as I. The four control points are also shown, denoted c1, c2, c3 and c4. Note how the relationship between the point of interest and the control points are critical to accurate 2D-DLT.
Additionally in order to collect data on the outcomes of the cross a radar gun was
positioned in line with the anticipated kicking direction of the cross. This allowed
the collection ball peak velocities post kick. Finally, the accuracy of the pass was
assessed by notational analysis whereby if the centrally positioned receiving
player touched the ball a score of 1 as coded for the trial whilst, if the receiving
player failed to touch the ball the pass was coded a 0.
2.2.5 Experimental Design
After verifying that all the assumptions were met with due corrections for
violation of the sphericity assumption (Schultz & Gessaroli, 1987) a one-way
69
analysis of variance (ANOVA) with repeated measures was used to investigate
three levels of defensive pressure (defender absent (control), far and near
positioning of defender) on the means of the gait parameters and cross
outcomes (ball speed and accuracy). Planned contrasts were to follow up the
main effects. A detailed analysis of at each footfall was then undertaken to
compare the gait parameters at successive footfalls across the three conditions.
Participants performed four trials in each condition for a total of 12 trials per
participant (i.e. a total of 32 trials per condition and a grand total of 96 trials
across the three conditions). These were administered using a quasi-Latin
square design to counterbalance order of treatment effects (Thomas, Nelson, &
Silverman, 2005).
To calculate the F-ratio main effect using the ANOVA with repeated measures
design note that:
F = MSM/MSR
Since:
MSM = MSM/dfM
And that:
MSR = MSR/dfR
We note that:
dfM = (k - 1)
Noting also that:
dfR = (dfw – dfM)
Considering also:
dfW = n × (k – 1)
70
Therefore the degrees of freedom for the variation explained by the model (MSM)
which is denoted dfM, equals the number of conditions (k) minus 1 which gives
dfM of 2.
The degrees of freedom for the variation explained by extraneous variables (i.e.
mean square of the residual error, denoted MSR) which is denoted dfR, is equal to
the degrees of freedom of the within participant sums of squares (i.e. dfW =
number of participants (n) multiplied by the number of conditions (k) subtracted
by 1) subtracted by the degrees of freedom of the model (dfM, see above).
Consequently this results in the sum, 16 minus 2 which results in dfR = 14 (i.e.
df(2, 14))
Since planned contrasts at each footfall were also intended it is noted the
degrees of freedom for the planned contrast F-ratio are: dfM = k -1, which is the
sum 2 -1 resulting in dfM = 1; and, dfR = dfM - (n × (k – 1)), which is the sum, 1 -
The experiment was administered over two separate testing sessions with one
week between sessions. Participants undertook 4 trials in succession before
being scheduled a minimum rest period of 8 minutes. Between successive trials,
rest periods were scheduled for 2-minutes or participants took as long as they
needed to recover between trials. Attacker-defender dyads were formed by
random assignment. SPSS version 15.0 (SPSS inc, Chicago, IL) was used for
analysis.
2.3 RESULTS
Table 2.2 shows the main effects on mean gait parameters and peak ball speed
and pass accuracy outcomes in the different conditions: defender-absent,
defender-far and defender-near. Significance was declared at the traditional
alpha level of .05.
The following table summarises the main effects of the three conditions
defensive pressure and provides overall mean values on each of the dependent
variables on these conditions.
71
Table 2.2 Main effect and planned contrasts of the three levels of defensive pressure on the dependent variables during the locomotor pointing task
Gait Characteristics
Main Effect,
F (dfM, dfR)
Defender Condition (±95% CI)
Absent Far Near
Mean Footfall Variability (m) 3.75* (2,14) 0.922
±0.28 0.461,3 ±0.28
0.592 ±0.29
Mean Step Length (m) 1.20 (2,14) 1.50
±0.04 1.52
±0.07 1.52
±0.06
Mean Velocity (m•s-1) 8.69* (2,14) 5.403
±0.21 5.413
±0.15 5.601,2
±0.21
Mean Foot/Ball Distance (m) 3.43 (2, 14) 9.443
±0.62 9.84
±1.26 9.941 ±1.00
Outcome Characteristics
Accuracy (1 = Received, 0 = Not Received)
1.36 (2,14) 0.78 ±0.23
0.78 ±0.17
0.59 ±0.19
Peak Ball Velocity (km•hr-
1)
21.18* (2,14)
83.632,3 ±2.61
74.421,3 ±2.57
69.581,2 ±2.57
Significant main effects starred (*0.05). Pair-wise significant effects numbered where different to 0.05 level (1Absent/2Far/3Near different at 0.05 level). CI = confidence intervals. dfM = Degrees of freedom of the model. dfR = Degrees of freedom of the residual. F = critical value for F-ratio
Presented in detail below are the planned contrasts of each of the dependant
variables across the final 13 footfalls across the three conditions. In interpreting
the following data, it is important to note that: (i) the final 13 footfalls were
evaluated across all because this number of steps was common to all
participants, and (ii) footfalls are reported by number from the planting phase.
For example footfall number 0 indicates this is the planting phase footfall that
supported the kick. Footfall number 1 indicates that this is one footfall from the
planting phase footfall that supported the kick and so on.
72
2.3.1 Outcomes
There were no significant differences between conditions in terms of pass
accuracy (Table 2.2, Figure 2.4). Analysis of peak ball velocities showed that
participants kicked the ball fastest in the defender-absent condition compared to
the defender-far condition (F(1, 7) = 14.12 (p ≤ .05) and the defender-near
condition (F(1, 7) = 28.85 (p ≤ .05)). This trend continued with participants
kicking the ball with less velocity in the defender-near condition compared with
the defender-far condition (F(1, 7) = 14.89 (p ≤ .05)).
Figure 2.4 Mean outcomes of accuracy (primary axis) and ball velocity (secondary axis) of the three conditions of defensive pressure. km/hr = kilometres per hour. Error bars = 95% confidence intervals.
73
2.3.2 Foot/Ball Distance Variability and Step Length Analysis
2.3.2.1 Foot/Ball Distance Variability
A one way ANOVA with repeated measures was undertaken to evaluate the effect
of the three levels of defensive pressure on the overall mean foot/ball distance
(m) variability averaged over the final 13 footfalls leading into the kick. Footfall
variability was significantly affected by the level of defensive pressure, F(1.11,
The pair-wise comparisons of the means for each level of defensive pressure
revealed that the defender-absent condition was more variable than the
defender-far condition (F(1, 7) = 5.14 (p ≤ .05)) and the defender-far condition
was significantly less variable than the defender-near condition (F(1, 7) = 5.54 (p
≤ .05)). No other significant differences between levels of defensive pressure
were found.
Planned contrasts of the mean variability between each condition and at each
footfall leading into the cross were also undertaken. These values with 95%
confidence intervals are shown below in Figure 2.5 for visual inspection (see also
Table2.3).
74
Figure 2.5 Mean foot/ball distance variability patterns of the eight participants for each of the three levels of defensive pressure for the final 13 footfalls of the run-up to cross. Error bars = 95% Confidence Intervals.
Planned contrasts revealed that the defender-absent condition was significantly
more variable compared to the defender-far condition on: footfall 12 (F(1, 7) =
Table 2.3 Planned contrasts of the mean foot/ball distance (m) standard deviations at the final 13 footfalls across the three levels of defensive pressure.
Footfall Defender Condition
Absent Far Near
12 1.012 0.441 0.61
11 1.122 0.481 0.64
10 1.202 0.541 0.68
9 1.212 0.531, 3 0.722
8 1.252 0.551 0.71
7 1.282 0.611 0.74
6 1.34 0.69 0.73
5 1.212 0.571 0.69
4 1.002 0.471 0.65
3 0.66 0.38 0.59
2 0.37 0.31 0.48
1 0.22 0.213 0.292
0 0.082 0.131 0.14
Planned contrast significant effects numbered where different to 0.05 level (1absent/2far/3near at 0.05 level)
2.3.2.2 Absolute Step Lengths
The patterns of step length modifications were analysed for the final successive
13 footfalls for each three conditions of defensive pressure. Defensive Pressure
revealed no main effect on the overall mean step lengths (m) adopted by the
significant comparisons between conditions on overall mean step length (Error!
Reference source not found.).
The mean step lengths over the final 13 footfalls for each condition with 95%
confidence intervals are shown below in Figure 2.6 for visual inspection.
76
Figure 2.6 The mean step lengths for the eight participants across the three levels of defensive pressure, calculated between the final 13 successive footfalls of the run-up to cross. Error bars = 95% confidence intervals.
The mean step lengths at each footfall were then compared between each level
of defensive pressure. The mean values and significant planned contrasts are
summarised below in Table 2.4.
Planned contrasts between the defender-absent and defender-far conditions
showed that early in the run up the players adopted significantly larger distances
between footfalls in the defender-absent condition between footfalls 12-11 (F(1,
7) = 14.07 (p ≤ .05)). This trend was reversed with the defender-absent condition
showing a significantly reduced step length compared to the defender-far
Despite trends in which the attacking players adopted larger step lengths
particularly over footfalls 6-3, there were no significant planned contrasts in the
step lengths adopted between the defender-far and defender-near conditions
over final 13 footfalls leading into the cross.
78
Table 2.4 Planned contrast of the mean distances (m) between the final 13 successive footfalls across the three levels of defensive pressure.
Step Defender Condition
Absent Far Near
13-12 1.24 1.16 1.18
12-11 1.402,3 1.311 1.281
11-10 1.46 1.42 1.41
10-9 1.553 1.51 1.491
9-8 1.58 1.57 1.55
8-7 1.65 1.62 1.61
7-6 1.75 1.71 1.72
6-5 1.66 1.76 1.68
5-4 1.522,3 1.731 1.711
4-3 1.283 1.41 1.46 1
3-2 1.363 1.50 1.631
2-1 1.25 1.25 1.31
1-0 1.77 1.76 1.71 Pair-wise significant effects numbered where different to 0.05 level (1absent/2far/3near at 0.05 level)
2.3.2.3. Absolute Foot/Ball Distance
The mean foot/ball distance (m) observed over the final 13 footfall in the run-up
were analysed in a one way ANOVA with repeated measures with three levels of
Defensive Pressure. The results indicated no significant main effect of Defensive
Pressure on the mean foot/ball distance of the attacking players (F(1, 14) =
3.43).
Planned contrasts of the average foot/ball distance between the different levels
of defensive pressure however, showed that the players in the defender-absent
condition covered significantly more ground over the final 13 footfalls than the
defender-near condition (F(1, 7) = 8.27 (p ≤ .05). No other significant differences
between conditions emerged. These findings are summarised above in Error!
Reference source not found..
The mean foot/ball distance over the final 13 footfalls for each condition with
95% confidence intervals are shown below in Figure 2.7 for visual inspection.
79
Figure 2.7 Mean foot/ball distances across the three levels of defensive pressure calculated over the final 13 footfalls. m = metres. Error bars = 95% confidence intervals.
The mean foot/ball distance at each footfall (12-0) were then compared between
each level of defensive pressure. The mean values and significant planned
contrasts are summarised below in Table 2.5.
Planned contrasts of the mean foot/ball distance at each footfall between the
defender-absent and defender-far condition revealed that in the defender-far
condition, players were further from the ball at footfall 0 (F(1, 7) = 9.99 (p ≤
.05)). No other significant differences were found between these groups at any
other footfall.
Planned contrasts of the mean foot/ball distance at each footfall between the
defender-absent and defender-near condition revealed that in the defender-near
condition, players had covered significantly less distance at: footfall 12 (F(1, 7) =
Planned contrasts of the mean foot/ball distance at each footfall between the
defender-far and defender-near condition revealed that there were no footfalls
where these two conditions were significantly different.
Table 2.5 Planned contratsts of the mean foot/ball distance (m) at the final successive 13 footfalls for the three levels of defensive pressure.
Footfall Defender Condition
Absent Far Near 12 18.603 18.99 19.041
11 17.213 17.69 17.761
10 15.743 16.26 16.351
9 14.203 14.76 14.861
8 12.623 13.19 13.311
7 10.963 11.57 11.701
6 9.213 9.86 9.981
5 7.55,3 8.10 8.301
4 6.033 6.37 6.591
3 4.75 4.96 5.13
2 3.39 3.46 3.50 1 2.14 2.21 2.19
0 0.372,3 0.451 0.481
Pair-wise significant effects numbered where different to 0.05 level (1absent/2far/3near at 0.05 level).
2.3.3 Displacement Velocity
The mean horizontal displacement velocities observed over the final 13 footfall
(minus footfall 0) in the run-up were analysed in a one way ANOVA with repeated
measures with three levels of Defensive Pressure. The results indicated a
significant main effect of Defensive Pressure on the mean horizontal
displacement velocity (m•s-1) of the attacking players (F(1.22, 8.58) = 8.69 (p ≤
.05)).
Planned contrasts of the average velocity between the different levels of
defensive pressure showed that the players in the defender-absent condition ran
significantly slower than when undertaking the cross compared to the defender-
81
near condition (F(1, 7) = 57.12 (p ≤ .05). Additionally, players in the defender-far
condition ran significantly slower than the defender-near condition (F(1, 7) =
7.41 (p ≤ .05). No other significant differences emerged between groups (Error!
Reference source not found.).
The mean horizontal displacement velocity (m•s-1) over the final 13 footfalls
(minus footfall 0) for each condition of defensive pressure with 95% confidence
intervals are shown below in Figure 2.8 for visual inspection.
Figure 2.8 Mean horizontal displacement velocity across the three levels of defensive pressure calculated at each footfall. Note that because the central moving differences method was used, the final footfall (0) is unknown. m•s-1 = metres per second. Error bars = 95% confidence intervals.
The mean horizontal displacement velocity at each footfall (12 to 1) were then
compared between each level of defensive pressure. The mean values and
significant planned contrasts are summarised below in Table 2.6.
82
Planned contrasts of the mean velocity at each footfall between the defender-
absent and defender-far condition revealed that in the defender-far condition,
also examined in this study, where stride rate was found to increase and stride
lengths decreased compared with running free at the same relative speed. Key
92
limitations of physiological research is that: (i) in the case of player tracking
studies, the descriptive methods used mean each action of the player is
decontextualised from the performance behaviour of dribbling; and (ii), the
measurements of football players whilst dribbling on a treadmill is likely to
represent a very different task constraint than that required in the performance
environment. When players run and dribble with a ball during games, they do so
under the specific constraints of the performance context, such as the presence
of defenders and team mates. The presence and positioning of defenders, for
example, in Study 1 of this thesis have been shown to influence how players
coordinate movements relative to the ball and should be a feature of
experimental designs that consider the movement coordination of football
players.
The aims of this study were to: (i) test whether dribbling the ball reduces the
variability in foot/ball distance between trials during the acceleration phase
when under defensive pressure, and (ii) test whether football players gait
patterns and displacement velocity remains stable under defensive pressure
when free to chose when and where to kick from.
3.2 METHODS
The same eight participants were involved in this study as Study 1 (see Table
2.1). The same apparatus and data capture procedures were also reapplied in
this experiment from Study 1 (see section 2.2.3 Apparatus, and section 2.2.4
Data Capture above). In addition, the same dependent variables, foot/ball
distance SD, step length patterns and the horizontal displacement velocity of the
players were used to evaluate the independent variables in this study (i.e.
dribbling under no defensive pressure, and increased levels of defensive
pressure, detailed below in section 3.2.1 Task). Because of the nature of the
dribbling task constraints in this study, which did not pre-specify where and when
to generate the cross (i.e. the participants could cross the ball at any time and
place), the additional dependent variable, ‘player-to-by-line distance’ was
consequently evaluated. This measure indicated how far down the field the
players dribbled the ball before making the cross. The closer to the by-line that
93
they got, the further they had dribbled the ball. Finally, rather than evaluate the
foot/ball absolute distance at each footfall (as in Study 1), only the final plant
phase foot/ball distance was considered as this value is of interest to evaluate
whether kicking strategy changed between levels of defensive pressure.
3.2.1 Task
Below, Figure 3.1 depicts the nature the task and roles of each player. The
attacker (denoted A) was instructed to sprint with the ball (denoted B) to the by-
line and cross it back towards the penalty spot. A receiving player (denoted R)
was instructed to receive the ball at the penalty spot and attack the goal. A goal-
keeper (denoted GK) was instructed to protect the goal. In conditions where one
was present, the defender (denoted D) was instructed to meet the attacker at the
earliest point and stop the cross. In order to ensure health and safety,
participants were instructed to perform within the laws of the game with an
emphasis on the non-contact requirements of the sport. To enforce the laws of
the game a referee was positioned in the field. The task was initiated at the
attacker’s discretion after a signal from the referee.
94
Figure 3.1 Schematic of the experimental task in Study 2. B = Ball, A = Attacker, GK = Goal-keeper, R = Receiver, D = Defender. Only the defender was changed across conditions of defensive pressure (i.e. absent, far or near), all other variables were held constant. The numbered scale reflects meter units. The arrow indicates that the target for the attacker was the penalty spot. The brackets indicate that the attacker was free to pass at any location in the approach toward the by-line.
A one way analysis of variance (ANOVA) with repeated measures was used to
investigate the effect of the three levels of defensive pressure (Defender Absent
(control), Defender Far and Defender Near) on the overall means of the
dependent variables and also the means at each footfall leading into the cross.
Participants performed four trials for each of the three levels of defensive
pressure for a total of 12 trials per participant (i.e. a total of 32 trials per
condition and a grand total of 96 trials across the three conditions). Trials were
completed over two separate testing sessions. Participants were scheduled 2-
minute rest periods between trials or took as long as required. Treatments were
counterbalanced using a quasi-Latin square design.
Direction
of play
95
Defenders were positioned with a 20% and a 10% disadvantage relative to the
attacker. This was achieved by recording each defender’s sprint time over 20 m
and each attacker’s time to dribble a ball as quickly as possible over 20 m and
cross the ball. The same procedure described in Study 1 to position the
defenders was used (refer to Appendix A: for Action-Scaled Adversary Distance
for formulas and to section 2.2.5 Experimental Design for a more detailed
description). Note also that the degrees of freedom were derived in the same
way as Study 1.
3.3 RESULTS
The results are presented in two sub-sections below. The first sub-section is
concerned with the foot/ball position effects across the three conditions of
defensive pressure. These include the foot/ball variability and the successive
mean step lengths. The second section is concerned with the horizontal
displacement velocities at each footfall prior to the planting phase footfall and
the distance from the by-line that the pass emerged between the different levels
of defensive pressure. As with Study 1 because of the multiple levels of
independent variables, where the assumptions of sphericity were violated
according to Mauchly’s test, effects were reported with the degrees of freedom of
the error term of the ANOVAs corrected using the Greenhouse-Geisser estimates
of sphericity (Shutz & Gessaroli, 1987). Note also that the degrees of freedom
were determined using the same method described in section 2.2.5
Experimental Design.
In interpreting the following data, it is critical to note that, the final 11 footfalls
are evaluated across the three conditions of defensive pressure because this
number of footfalls was common to all participants across the three conditions.
In addition, for ease of interpretation, also note that the footfalls are reported by
number from the plant phase footfall. For example footfall number 0 indicates
this is the final planted foot position used to support the kick. Footfall number 1
indicates that this is one footfall from the plant phase footfall and so on. Error!
Reference source not found. shows the main effects of the defensive pressure
96
conditions and provides overall mean values on each of the dependent variables
under the three levels of defensive pressure.
Table 3.1 Main effects of Study 2.
Gait Characteristic
Main Effect,
F (dfM, dfR)
Player
Absent Far Near
By-Line Distance (m) at Footfall 0
3.02 (2,12) 3.50 ±0.74
4.81 ±1.27
4.62 ±1.01
Mean Footfall Variability (m)
0.57 (2, 12) 0.72 ±0.42
0.63 ±0.32
0.82 ±0.81
Mean Step Length (m)
0.53 (2, 12) 1.45 ±0.05
1.48 ±0.09
1.48 ±0.10
Mean Horizontal Displacement Velocity (m•s-1)
2.86 (2, 14) 4.81 ±0.69
5.05 ±0.55
5.02 ±0.64
Mean Plant Phase Foot/ Ball Distance (m)
3.03 (2, 14) 3.51 ± 1.48
4.81 ±2.55
4.62 ±2.02
Outcome Variables
Accuracy (1 = received, 0 = not received)
1.36 (2,14) 0.59 ±0.19
0.53 ±0.21
0.56 ±0.21
Peak Ball Velocity (km•hr-1)
92.58 (2,14)*
80.62 ±2.78
67.89 ±2.57
65.81 ±2.56
Significant main effects starred (*0.05). Pair-wise significant effects numbered where different to 0.05 level (1Absent/2Far/3Near different at 0.05 level). CI = confidence intervals. dfM = Degrees of freedom of the model. dfR = Degrees of freedom of the residual. F = critical value for F-ratio
3.3.1 Outcomes
Two outcome measures were collected in order to provide an indication of level
of success performers were capable of achieving between levels of Defensive
97
Pressure. The mean values with 95% confidence intervals are shown below in
Figure 3.2 for visual inspection. The main effect of Defensive Pressure on Pass
Accuracy (1 = Received, 0 = Not Received) revealed no significant effect of
Defensive Pressure, F(2, 14) = 0.13. Additionally there were no significant
differences between conditions.
With regard to the peak ball velocities (m•s-1) generated by players, similar to
study 1 there was a significant main effect of Defensive Pressure, F(2, 14) =
53.28 (p ≤ .05). Planned contrasts showed that players kicked the ball fastest in
the defender-absent condition compared to the defender-far condition (F(1, 7) =
10.69 (p ≤ .05) and the defender-near condition (F(1, 7) = 10.87 (p ≤ .05)).
However there were no significant differences in ball speeds between the
defender-near condition compared with the defender-far condition.
Figure 3.2 Mean outcomes of accuracy (primary axis) and ball velocity (secondary axis) when players were required to dribble the ball under different levels of defensive pressure. m/s = metres per second. Error bars = 95% confidence intervals.
98
3.3.2 Foot/Ball Distance and Step Length Control Analysis
3.3.2.1 Foot/Ball Distance Variability
The mean foot/ball distance standard deviations (m) for the final 11 footfalls
were examined across each of the three levels of defensive pressure. A one way
ANOVA with repeated measures was conducted on the effect of the three levels
of defensive pressure on overall mean foot/ball distance variability. There was no
significant main effect of level of Defensive Pressure found, F(1.46, 10.22) =
0.40.
The mean standard deviations of the foot/ball distances were plotted below with
95% confidence intervals in Figure 3.3 for visual examination.
Figure 3.3 Mean foot/ball distance variability patterns of the eight participants for each of the three levels of defensive pressure for the final 11 footfalls of the dribble to cross. Error bars = 95% confidence intervals.
The plot showed an unexpected trend in the foot/ball distance SDs at the plant
phase footfall in the defender-absent condition. Instead of reducing
99
systematically during the zeroing in phase as expected, the foot/ball distance
variability increased. An individual analysis was undertaken to explore the data
for potential outliers. An outlier beyond three standard deviations from the mean
was shown by participant number 2 (shown below in Figure 3.4). The data of
Participant Two was subsequently removed from future data analyses regarding
foot/ball coordination variables.
Figure 3.4 Participant Two’s foot/ball distance variability patterns for the three levels of defensive pressure. Note the large and non-declining nature of the Defender Absent condition.
The ANOVA with repeated measures was repeated on the foot/ball distance
variability between the three levels of defensive pressure to the exclusion of
Participant Two. The main effect of Defensive Pressure again did not show a
significant effect, F(1.55, 9.31) = 0.57. However, there was a large change in the
defender-absent final foot/ball distance SD pattern which can be seen below in
100
Figure 3.5. No planned contrasts showed significant differences between
conditions at any footfall (the mean values for each condition are summarised
above in Error! Reference source not found.).
Figure 3.5 Mean foot/ball distance variability patterns of seven participants (to the exclusion of Participant Two) for each of the three levels of defensive pressure for the final 11 footfalls of the dribble to cross. Note the large reduction in the mean foot/ball distance SDs in the No Defender conditions final few footfalls. Error bars = 95% confidence intervals.
3.3.2.2 Absolute Step Lengths
The step length patterns were analysed across the final 11 footfalls for each
three conditions of defensive pressure. A one way ANOVA with repeated
measures with 3 levels of Defensive Pressure was conducted on the overall
mean step length adopted by the attacking players for each condition. The main
effect of Defensive Pressure was not statistically significant (F(1.52, 11.57) =
0.53, nor were there any significant planned contrasts found between groups.
The overall mean values across the three levels of defensive pressure are
summarised above in Error! Reference source not found..
101
The mean values and 95% confidence intervals of the distance between each
successive footfall for each condition of defensive pressure are summarised
below in Figure 3.6 for visual inspection.
Figure 3.6 The mean step lengths for the seven participants (participant number 2 excluded) across the three defensive pressure conditions for Study 2. Step lengths were calculated by taking the difference between the final 11 successive footfalls of the dribble to cross.
The mean step lengths at each footfall were then compared between each level
of defensive pressure. The mean values and significant planned contrasts are
summarised below in Table 3.2.
Planned contrasts between the defender-absent and defender-far conditions
showed that early in the run up the players adopted significantly larger step
lengths when undertaking the cross in the defender-absent condition between
footfalls 7-6 F(1, 6) = 8.63 (p ≤ .05). This trend was reversed however later in the
run-up with the players adopting a significantly larger step length when
undertaking the cross under the defender-far condition between footfalls 5-4,
102
F(1, 6) = 12.49 (p ≤ .05). No other significant differences between the two
conditions emerged.
Planned contrasts between the defender-absent and defender-near conditions
showed that early in the run up the players adopted shorter step lengths under
the defender-near condition between: footfalls 10-9 F(1, 6) = 6.07 (p ≤ .05) and
footfalls 8-7 F(1, 6) = 7.53 (p ≤ .05). Despite trends of an increase in step length
under the defender-near condition, specifically between footfalls 5-4, no other
differences between the two conditions emerged.
There were no significant planned contrasts in the step lengths adopted between
the defender-far and defender-near conditions over final 11 footfalls leading into
the cross.
103
Table 3.2 Planned contrasts of the distance between the final 11 successive footfalls across the three conditions of defensive pressure when players dribbled the ball.
Step Defender Condition
Absent Far Near
11-10 1.38 1.39 1.34
10-9 1.553 1.43 1.391
9-8 1.433 1.41 1.491
8-7 1.64 1.54 1.56
7-6 1.452 1.561 1.58
6-5 1.54 1.60 1.67
5-4 1.462 1.601 1.59
4-3 1.32 1.37 1.42
3-2 1.44 1.39 1.37
2-1 1.20 1.20 1.22
0-1 1.70 1.65 1.73
Pairwise significant effects numbered where different to 0.05 level (1absent/2far/3near at 0.05 level)
3.3.3 Velocity Change during the Dribble to Cross
The mean horizontal displacement velocities (m•s-1) observed over the final 11
footfall (minus footfall 0) in the run-up were analysed in a one way ANOVA with
repeated measures with three levels of Defensive Pressure. The results indicated
a non-significant main effect of Defensive Pressure on the mean horizontal
displacement velocity of the attacking players (F(2, 14) = 2.86).
Pair-wise comparisons of the average velocity between the different levels of
defensive pressure showed no significant differences between conditions. These
findings are summarised above in Error! Reference source not found..
The mean horizontal displacement velocity (m•s-1) over the final 11 footfalls
(minus footfall 0) for each condition with 95% confidence intervals are shown
below in Figure 3.7 for visual inspection.
104
Figure 3.7 Mean horizontal displacement velocity across the three conditions calculated for the final 11 footfalls (minus footfall 0). Note that because the central moving differences method was used, the final footfall (0) is unknown. m/s = metres per second. Error bars = 95% confidence intervals.
The mean horizontal displacement velocity at each footfall (10 to 1) were then
compared between each level of defensive pressure. The mean values and
significant planned contrasts are summarised below in Table 3.3.
Planned contrasts of the mean velocity at each footfall between the defender-
absent and defender-far condition revealed that in the defender-far condition,
Finally, there were no significant differences between the defender-near
condition compared to the defender-far conditions.
Table 3.3 Planned contrasts of the mean horizontal displacement velocity (m•s-1) at each of the final 11 (minus footfall 0) footfalls for each level of defensive pressure.
Footfall Defender Condition
Absent Far Near
10 5.13 4.91 4.82
9 5.21 5.39 5.30 8 5.25 5.41 5.40
7 5.142,3 5.431 5.401
6 5.112,3 5.521 5.531
5 4.842,3 5.321 5.371
4 4.75 5.17 5.12
3 4.57 4.81 4.75
2 4.23 4.39 4.38
1 3.52 4.02 4.08
Pair‐wise significant effects numbered where different to 0.05 level (1absent/2far/3near at 0.05 level)
3.3.4 Final Player-Byline Distance at the Cross
The foot-to-by-line distance at the plant phase footfall was analysed in a 3 level
(Defensive Pressure) one-way ANOVA with repeated measures. The analysis
revealed no significant main effect between levels of defensive pressure, F(2,
14) = 3.03. Though there was a trending significance for the players to kick the
ball further from the by-line when under defensive pressure, post hoc pair-wise
comparisons between the different combinations of the 3 levels of defensive
pressure showed no significant differences between conditions. The mean
values, main effects and outcomes of the planned contrasts of the plant phase
footfall-to-by-line distance are displayed above in Error! Reference source not
found..
106
3.4 DISCUSSION
The aim of this experiment was to consider: (i) are the player-to-ball coordination
tendencies when dribbling under the same funnel shaped control as shown when
players run to kick a stationary ball?; and (ii) how are gait parameters influenced
by different levels of defensive pressure when football players are able to dribble
the ball before crossing it?
3.4.1 Funnel-Shaped Control During Dribbling
Referring above to Figure 3.5 (see section 3.3.2.1 Foot/Ball Distance Variability),
it can be seen that across all levels of defensive pressure, there is a
characteristic trend showing initially high amounts of foot/ball distance variability
and that at a given footfall, the variability systematically reduces to low levels
(beginning approximately at footfall 6 in the Defender Absent and at footfall 4 in
the defender-far and defender-near conditions). This overall player/ball distance
pattern of variability is similar to previous locomotor pointing research on long
jump and cricket bowler run-ups (Montagne, 2005) and to Study 1 of this thesis.
The similarities appear to confirm the presence of a funnel shaped control even
under conditions where the location of the position of the kick is not pre-
specified at the initiation of the task. The finding that funnel shaped control
occurs even under conditions when the location of the pass is not pre-specified
might indicate that players at some point in the dribbling process perceive and
act toward an emergent location area to perform the nested action, determined
in some part by the future arrival of the moving ball. Indeed, ubiquitous to all
players was the strategy of running with ball toward the by-line and at some
point, beginning a curved approach around the moving ball, intercepting it to
cross it whilst it was still moving.
Previous research into the volley ball pass has found that in intrinsically timed
tasks, or self-paced tasks, that funnel shaped control emerges despite the
location of the object for interception not held to a specific location in space
(Davids, Lees, & Burwitz, 2000). Players appear to generate a consistent relation
to objects for interception despite their variability in terms of a global coordinate
system. That is, players adapt readily to the position of a moving ball by
107
repositioning their own body and segmental orientations relative to the object for
interception (Davids, et al., 2000).
The lack of any distinct trends in the data sets between conditions raises doubts
as to whether this variability is indeed an indication of any specific control
strategy as was suggested in Study 1. There are limitations in the current
methods for calculating foot/ball variability when a player is dribbling a ball. It
might be for example, that the task should be reconsidered in that with each
successive touch with the ball the player is undergoing a locomotor pointing
process (i.e. running to the ball in order to kick it ahead themselves in order to
again be able to kick the ball). Future research should account of the specific
process of dribbling and undertake an individual, trial-by-trial analysis in order to
calculate each footfall relative to the balls next touch location rather than relative
to the balls final kicked position. Despite the limitations raised by the grouped
analysis used that may mask locomotor pointing strategies, the analysis in this
study can confirm that dribbling appears to undergo an acceleration phase,
characterised by high amounts of foot/kick-location variability, followed by a
zeroing-in phase, characterised by a systematic reduction to relatively low
amounts of foot/ball position variability, despite the location not being pre-
specified at the initiation of the task.
3.4.2 Effect of Defensive Pressure on Gait Parameters While
Dribbling
The effect of defensive pressure on gait parameters analysed in this study
showed some interesting results in contrast to those revealed in Study 1. The
step lengths (shown above in Figure 3.6, section 3.3.1.2) of the football players
displayed more shortening and lengthening in the acceleration phase of the
dribble process when compared with the Stationary Ball conditions in Study 1.
This ‘sawtooth’ pattern of step lengthening followed by step shortening during
the initial dribbling period (from footfalls 11 to 6 in the defender-absent
condition) occurs for several cycles of footfalls. However the pattern is
systematically reduced (or flattened) when defensive pressure was present (i.e.
the players appeared to systematically increase their step lengths without
108
intervening step length shortenings). The ‘flattening’ process under defensive
pressure might be associated with increased running velocity (see Figure 3.7 and
Table 3.3 for a summary of the mean velocity at each footfall across the three
levels of defensive pressure). This interpretation of a link between the step
length flattening and increased running velocity is supported in that the peak
running velocity significantly increased when under defensive pressure (see
Table 3.3).
The same trend in Study 1 where players appeared to delay the footfall at which
step lengths were systematically reduced under defensive pressure also
appeared to occur when players dribble a ball. This was shown by the
significantly larger step length between footfalls 5-4 in the defender-far
conditions when compared to the defender-absent condition (see Table 3.2).
Again as in Study 1, it would appear the presence of a defender results in
player’s sacrificing the number of footfalls over which they regulate the final
approach into the cross pass for the gain of an increased peak running velocity.
Delay in step length shortening occurred despite players having the ability to
pass at any point in the approach toward the by-line. Defensive pressure appears
to influence movement coordination of ball-carriers.
Whilst similar to Study 1 there was an increase in the peak running velocities of
the defender present as opposed the defender absent condition however, when
dribbling there was no dose response relationship between increased defensive
pressure and increased running velocity (i.e. there were no pair-wise significant
differences between the defender-far and defender-near conditions at any
footfall leading into the approach). As the defender was positioned closer, the
attacker did not run significantly faster. An explanation for the lack of an increase
in velocity relative to a closer defender is that the football players chose to pass
the ball earlier. Indeed, players under no defensive pressure trended toward
passing the ball significantly closer to the by-line compared to the defender
present conditions. An explanation of why there was no significant difference in
the distance of the kick at the plant phase might be that players in the defender-
near and defender-far conditions either passed early on some occasions, whilst
driving closer to the by-line on others. Future research interested in examining
109
how defensive pressure influences emergence of a pass location when players
are dribbling should take a trial-by-trial method of analysis.
The question raised in Study 1, whether players were adopting an undesirable
kicking strategy due to the requirement to run faster when under defensive
pressure, was evaluated in this study by considering the variability and absolute
distance between the plant phase foot and ball (data in Error! Reference source
not found.). Contrary to Study 1, when football players dribble the ball, there were
no significant differences in foot/ball distance variability or the absolute foot/ball
distance at the plant phase footfall. The non-significant differences suggest that
when football players have the ability to choose the pass location and have
control of the ball; they use a preferred foot/ball position at plant phase footfall
despite different levels of defensive pressure. This would suggest players are
capable of sacrificing speed without the consequences of not getting the ball
first.
Of the two hypotheses that might explain why football players do not change
plant phase strategies when dribbling (as they appeared to do when approaching
a stationary ball in Study 1) it is likely that this is related to the overall running
velocity being much lower by comparison to the stationary ball condition. Football
players’ plant phase foot positions become more variable when they are required
to run at high velocities due to needing to reach a pre-specified location prior to a
defender. However, the football players managed to be consistent between trials
in the plant phase footfall when they are able to dribble the ball prior to kicking it.
There appears to be a change in plant phase strategy when football players
travel at near maximal velocities up to 4 footfalls prior to preparing for the kick.
3.5 CONCLUSIONS AND FUTURE RESEARCH
This study has added to the current understanding of how football players control
and coordinate gait under conditions relevant to their performance context.
Methods from previous locomotor pointing research were shown to be useful in
analysing coordination during dribbling. Further analysis and research is
recommended to better capture how football players coordinate relative to each
touch of the football ball, rather than observing movement coordination relative
110
to the final ball position prior to a pass. Additionally, this study has also shown
that coordination of gait changes under the presence of defensive pressure:
football players reduce the number of footfalls over which they prepare for the
pass and increase their peak running velocity. However, in contrast to Study 1,
there appeared no evidence of a dose-response relationship in the running
velocities of the players when the initial position of the nearest defender was
scaled closer. The only explanation found for this was that players are able to
pass the ball at a self chosen position and manage velocity differently. Future
research is warranted to understand what information informs this choice of
passing position when defenders are pressuring the ball-carrier. The
interpersonal scale of analysis is needed if the influence of the defender is to be
more comprehensively understood.
111
Chapter 4: Conclusion
4. REFLECTIONS The two studies presented in this Master’s thesis have a number of theoretical
and practical implications. The contributions to the two interrelated areas of
sport science and human performance research will be summarised in the
following reflections section.
4.1 SUMMARY OF THE PRACTICAL IMPLICATIONS
This Master’s thesis has shown that there is a strong link between perception
and action during a representative passing task in football. This is evident in the
changes of intrapersonal coordination tendencies that emerge when a defender
is present to pressurise the time and space of the passing player. It is hoped that
the implications will influence future methods in sports science research that
hope to measure performance and influence its pedagogy by considering the
effect of constraints on perception and action couplings and subsequent
generalisability to the performance context.
4.2 SUMMARY OF THE THEORETICAL IMPLICATIONS
Locomotor pointing research provides an understanding into how humans are
influenced by information to support interactions with environments. This field of
research can be influenced by theoretical concepts such as affordances and
complex systems theory to provide an insight into explaining behavioural
emergence in sport and in day-to-day activities. This study goes some way in
showing that affordances can be included in generating experimental designs
that contain highly dynamic task constraints (i.e. such as other humans).
The importance of measuring both inter-personal and intra-personal variables in
this study showed how key objects and events are interacted with in a dynamic
performance context. Intrapersonal dynamics showed that lengthening and
112
shortening of footfalls is context dependent. Humans respond quantitatively
differently in how foot/object interactions are regulated when under dynamic
constraints. Differences in regulation reflect a change in how context specific
information modified movement coordination. Because of the strong link
between perception and action, the different velocities that players generated
suggest different information was being acted on. Additionally, because actions
were generated differently, for example the delay in when the players began to
decelerate and prepare for the kick, the opportunities available to undertake
regulatory behaviour become highly constrained and the room for error reduced.
Strikingly, this reduction in the room for error was shown to take effect over the
entire process of running to and approaching the football ball in Study 1. Under
dynamic constraints, representative of a participant’s performance tasks and
environment, perceptions and actions are quantitatively different experiences for
performers. Future experimental designs should consider the implications of the
environments and tasks performers are expected to reflect their perceptual-
action capabilities under.
The observation of a progressively increased velocity as the initial distances of
defender’s were scaled closer to the attacking players leave unanswered
questions for future research. The fact that players ran faster when the ball was
stationary and when dribbling supports the view that decisions in sport can occur
relative to an action-scaled boundary. However, because player interpersonal
distances were not analysed, the exact characteristics of this boundary or critical
action threshold remains unknown. The importance of measuring the
interpersonal distance of interacting players over time and throughout the
performance process might show important interrelated processes such as how
affordances are interacted with over the performance period.
The presence of critical thresholds for decision making in sport raises many
questions for future research. What might have happened to the attackers’
behaviours if, by action-scaling, reaching the ball before a defender was equi-
probable or if they were unable to reach the ball before the defender? Would the
attacking player change their approach trajectory once they no longer have time
to undergo the process of preparing for the cross. Might they instead begin to run
113
toward and shield the ball from the defender to ensure they at least gain
possession of the ball? What would the threshold be where a spontaneous
change in behaviour occur? Would it be relative to an action scaled value for
each attacker-defender dyad?
Theoretically, emergent decisional behaviour on the sporting field might be
conceptualised under a dynamical perception of affordances. The recording of
movement data from the defender was beyond the scope of this thesis, and
represents an area for further research. Future research should combine
methods of understanding affordances with measures of interpersonal dynamics
to consider whether this relatively simple rule (perception of affordances)
generates functional decisional behaviour in apparently highly complex
performance contexts. Theoretically, this idea would fulfil a gap between two
topics that have been researched separately: perceiving the action possibilities
of significant others and coordinating one’s own actions with dynamic objects.
114
Appendix A: Formulae for Action-Scaled Adversary Distance To clarify, the method of determining the initial position of the defender was
based on the manipulation of an action scaling7 equation. Whereby to calculate
the defender distance from the scheduled pass location two unknowns needed
to be established beforehand: (1) a measure of the velocity and associated
derivatives (time and displacement) of the attacking player under conditions
where they carry out the run and pass task; and (2) a measure of the velocity,
and associated derivatives, of the defender carrying out a straight run over the
same distance (that is in this case, a 20 m straight run without an additional task
nested to the end).
With these known, we may determine an action scaled distance (dDA.S.D.) (see
Dicks, et al., 2010) from the ball to position the defender which acts to equalize
the arrival time of the attacker and defender over a fixed attacker distance (in
this case 20 m). In other words both players should arrive at the ball at the same
time, despite having to cover separate distances. The following equation
exemplifies these variables:
VD.T. = dDA.S.D. / dTA.T.
Equation 4.1
Therefore to find dDA.S.D:
7 Action scaling in this case is used to cancel out the advantage that another player might have over functional and structural constraints.
115
dDA.S.D. = (dDD.T. / dTD.T.) (dDA.T. / VA.T.)
Equation 4.2
Where the attacker terms are defined as: overall attacker task velocity is VA.T.,
Attacker task time is dTA.T.,
and attacker task displacement is dDA.T.
And the defender terms are defined as: overall defender task velocity, VD.T.,
defender task time, dTD.T.,
and, defender task displacement, dDD.T..
With the action scaled distance known this may then be modified exactly by
percent variation.
116
Appendix B: Example Participant Information and Consent Forms
PARTICIPANT INFORMATION FOR QUT RESEARCH PROJECT
Pattern‐Forming Dynamics in Performer‐Ball Systems in Team Sports
Research Team Contacts
Dominic Orth — Masters Student Professor Keith Davids — Supervisor
QUT School of Human Movement Studies QUT School of Human Movement Studies
This project is being undertaken as part of a Masters project for Dominic Orth. The
project is funded by the Queensland University of Technology. The funding body will
not have access to the data obtained during the project.
117
The purpose of the project is to investigate influences on a football player’s ability
to maintain possession of a ball whilst running and how this might affect successful
passing and decision making when under defensive pressure.
The research team requests your assistance because you are a skilled football
player, between the ages of 14 to 17 years and currently involved in ongoing
training to improve your performance in competitive football.
PARTICIPATION
Your participation in this project is voluntary. If you do agree to participate, you can
withdraw from participation at any time during the project without comment or
penalty. Your decision to participate will in no way impact upon your current or
future relationship with QUT or with Kelvin Grove State College.
Your participation will involve information being gathered about your age, leg
lengths, height and practice history. You will then be filmed on your football field
whilst wearing small sensors that can measure your legs movements (positioned o
of each of your thighs and back and with a total additional weight of 450 grams) as
you undertake tasks similar to those you would normally perform during training or
match situations. Specifically, these tasks will ask you to take the role of an
attacking and defending player. As the attacker your role will be to run as fast as
you can in order to beat a defender and make a pass toward another player. The
experiments will take place during your teams existing training session under the
supervision of your coach and will vary in terms of how far you will be expected to
run (which will be between 30 to 40 metres each time). They will occur over five
118
separate occasions and require you to perform a total four to six sprints, taking
approximately 10‐15 minutes of your time on each occasion.
EXPECTED BENEFITS
It is expected that this project will benefit you by providing you information that can
lead to better decision making on a task that often occurs during football matches.
Also, the information you make available through participation will indirectly
benefit the football community – helping to progress our ability to improve learning
environments and player performance analysis in the future.
RISKS
There are no risks beyond your normal day‐to‐day football skills based training
requirements associated with your participation in this project. You will be required
to wear shin protection and studded football boots to minimise leg‐to‐leg contact
injuries and the incidence of slips or falls respectively. A group warm up routine
including stretching and games based activities will be completed prior to any
testing under the supervision of your usual coach team. Also, you will also be
ensured a warm up period to adjust to the added weight of the sensors.
Appropriate first aid precautions are in place.
CONFIDENTIALITY All comments and responses are anonymous and will be treated confidentially. The names of individual persons are not required in any of the responses.
CONSENT TO PARTICIPATE We would like to ask you to sign a written consent form (enclosed) to confirm your agreement to participate.
QUESTIONS / FURTHER INFORMATION ABOUT THE PROJECT Please contact the researcher team members named above to have any questions answered or if you require further information about the project.
CONCERNS / COMPLAINTS REGARDING THE CONDUCT OF THE PROJECT QUT is committed to researcher integrity and the ethical conduct of research projects. However, if you do have any concerns or complaints about the ethical conduct of the project you may contact the QUT
119
Research Ethics Unit on +61 7 3138 5123 or email [email protected]. The Research Ethics Unit is not connected with the research project and can facilitate a resolution to your concern in an impartial manner.
Thank you for helping with this research project. Please keep this sheet for your information.
CONSENT FORM FOR QUT RESEARCH PROJECT
120
Pattern‐Forming Dynamics in Performer‐Ball Systems in Team Sports
Research Team Contacts
Dominic Orth — Masters Student Professor Keith Davids — Supervisor
QUT School of Human Movement Studies QUT School of Human Movement Studies
have read and understood the information document regarding this project
have had any questions answered to your satisfaction
understand that if you have any additional questions you can contact the research team
understand that you are free to withdraw at any time, without comment or penalty
understand that you can contact the Research Ethics Unit on +61 7 3138 5123 or email [email protected] if you have concerns about the ethical conduct of the project
for projects involving minors: have discussed the project with your child and their requirements if participating
understand that the project will include audio and/or video recording
agree to participate in the project
Name
121
Signature
Date / /
STATEMENT OF CHILD CONSENT
Your parent or guardian has given their permission for you to be involved in
this research project.
This form is to seek your agreement to be involved.
By signing below, you are indicating that the project has been discussed with you and you agree to participate in the project.
















































































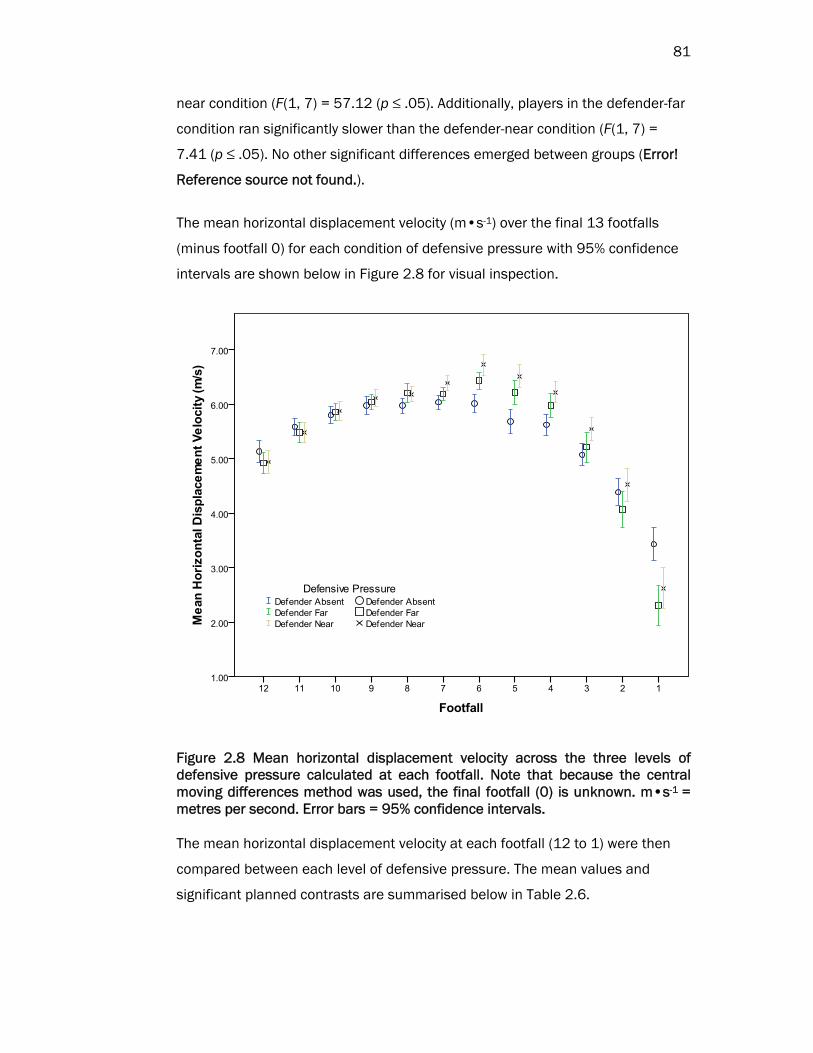


















































![LPV hard constraints IJCAS resubmit3 · nored, such as space telescope pointing [21] and machine tool control. For both interpretations, a solution to the OCC control problem results](https://static.documents.pub/doc/80x56/5e78dde83dbdfd0f59138701/lpv-hard-constraints-ijcas-resubmit3-nored-such-as-space-telescope-pointing-21.jpg)

![2014 Pointing Task Evaluation of Leap Motion Controller in ...chi-sparks.nl/2014/wp-content/uploads/2014/03/... · constraints of each interaction device itself [7]. A few of the](https://static.documents.pub/doc/80x56/5f0ea4687e708231d44038de/2014-pointing-task-evaluation-of-leap-motion-controller-in-chi-constraints-of.jpg)

![Optimal LPV Control with Hard Constraints · nored, such as space telescope pointing [21] and machine tool control. For both interpretations, a solution to the OCC control problem](https://static.documents.pub/doc/80x56/5e78cba8069f9b55f40223ba/optimal-lpv-control-with-hard-constraints-nored-such-as-space-telescope-pointing.jpg)


![LPV hard constraints IJCAS resubmit3jchoi/files/papers/WhiteZhuChoiIJCAS2016.pdf · nored, such as space telescope pointing [21] and machine tool control. For both interpretations,](https://static.documents.pub/doc/80x56/5e78cba7069f9b55f40223b9/lpv-hard-constraints-ijcas-jchoifilespaperswhitezhuchoiijcas2016pdf-nored.jpg)







