ORIGINAL RESEARCH Mechanism of immunotoxicological effects of tributyltin chloride on murine thymocytes Neelima Sharma & Anoop Kumar Received: 3 November 2013 /Accepted: 14 February 2014 # Springer Science+Business Media Dordrecht 2014 Abstract Tributyltin-chloride, a well-known organotin compound, is a widespread environmental toxicant. The immunotoxic effects of tributyltin-chloride on mamma- lian system and its mechanism is still unclear. This study is designed to explore the mode of action of tributyltin- induced apoptosis and other parallel apoptotic pathways in murine thymocytes. The earliest response in oxidative stress followed by mitochondrial membrane depolariza- tion and caspase-3 activation has been observed. Pre- treatment with N-acetyl cysteine and buthionine sulfoximine effectively inhibited the tributyltin- induced apoptotic DNA and elevated the sub G 1 popu- lation, respectively. Caspase inhibitors pretreatment pre- vent tributyltin-induced apoptosis. Western blot and flow cytometry indicate no translocation of apoptosis- inducing factor and endonuclease G in the nuclear frac- tion from mitochondria. Intracellular Ca 2+ levels are significantly raised by tributyltin chloride. These results clearly demonstrate caspase-dependent apoptotic path- way and support the role of oxidative stress, mitochon- drial membrane depolarization, caspase-3 activation, and calcium during tributyltin-chloride (TBTC)-induced thymic apoptosis. Keywords Apoptosis . Apoptosis-inducing factor . Calcium . Caspase . Endonuclease G . Tributyltin Introduction Tributyltin-chloride (TBTC), the well-known organotin compound, is a widespread environmental toxicant. It has several uses, such as in wood preservation and in anti- fouling paints used for marine vessels, quays, and buoys. It is also used as a biocide in cooling systems and as an organometallic chemical in diverse applications (Isomura et al. 2013). TBTC is extremely stable and resistant to natural degradation in water (Dimitriou et al. 2003). Several studies have reported toxic effects of TBTC in a variety of cell types, including thymocytes, neuronal cells, Jurkat lymphoma cells, natural killer cells, and keratinocytes (Vivani and MarinovichGalli 1997; Stridh et al. 1999; Gennari et al. 2003; Grote et al. 2004; Inadera 2006). Increased production of reactive oxygen species and oxidative damage (changes in activity of superoxide dismutase and increased lipid peroxidation) are detected in liver tissues of rats by TBTC exposure (Liu et al. 2006). The immune system is the major target of TBTC toxicity. It is widely accepted that exposure to TBTC can result in suppression of both innate and acquired immunity, leading to an increased susceptibility to in- fection. In rats, TBTC causes thymic involution due to depletion of cortical thymocytes (Bressa et al. 1991; Dewaal et al. 1997). TBTC also affects non-specific immune functions like natural killer (NK) cells (Whalen et al. 2002; Wilson et al. 2004; Aluoch et al. 2006) and macrophage activity (Kergosien and Rice 1998). Furthermore, TBTC is found to produce T- helper T H 2 polarization in vitro and in vivo, as indicated by suppression of production of T H 1-related cytokines Cell Biol Toxicol DOI 10.1007/s10565-014-9272-7 N. Sharma (*) : A. Kumar Department of Pharmaceutical Sciences, Birla Institute of Technology, Mesra, Ranchi 835215 Jharkhand, India e-mail: [email protected]
Transcript
ORIGINAL RESEARCH
Mechanism of immunotoxicological effects of tributyltinchloride on murine thymocytes
Neelima Sharma & Anoop Kumar
Received: 3 November 2013 /Accepted: 14 February 2014# Springer Science+Business Media Dordrecht 2014
Abstract Tributyltin-chloride, a well-known organotincompound, is a widespread environmental toxicant. Theimmunotoxic effects of tributyltin-chloride on mamma-lian system and its mechanism is still unclear. This studyis designed to explore the mode of action of tributyltin-induced apoptosis and other parallel apoptotic pathwaysinmurine thymocytes. The earliest response in oxidativestress followed by mitochondrial membrane depolariza-tion and caspase-3 activation has been observed. Pre-treatment with N-acetyl cysteine and buthioninesulfoximine effectively inhibited the tributyltin-induced apoptotic DNA and elevated the sub G1 popu-lation, respectively. Caspase inhibitors pretreatment pre-vent tributyltin-induced apoptosis. Western blot andflow cytometry indicate no translocation of apoptosis-inducing factor and endonuclease G in the nuclear frac-tion from mitochondria. Intracellular Ca2+ levels aresignificantly raised by tributyltin chloride. These resultsclearly demonstrate caspase-dependent apoptotic path-way and support the role of oxidative stress, mitochon-drial membrane depolarization, caspase-3 activation,and calcium during tributyltin-chloride (TBTC)-inducedthymic apoptosis.
Keywords Apoptosis . Apoptosis-inducing factor .
Calcium . Caspase . Endonuclease G . Tributyltin
Introduction
Tributyltin-chloride (TBTC), the well-known organotincompound, is a widespread environmental toxicant. It hasseveral uses, such as in wood preservation and in anti-fouling paints used for marine vessels, quays, and buoys.It is also used as a biocide in cooling systems and as anorganometallic chemical in diverse applications (Isomuraet al. 2013). TBTC is extremely stable and resistant tonatural degradation in water (Dimitriou et al. 2003).
Several studies have reported toxic effects of TBTC ina variety of cell types, including thymocytes, neuronalcells, Jurkat lymphoma cells, natural killer cells, andkeratinocytes (Vivani and MarinovichGalli 1997; Stridhet al. 1999; Gennari et al. 2003; Grote et al. 2004; Inadera2006). Increased production of reactive oxygen speciesand oxidative damage (changes in activity of superoxidedismutase and increased lipid peroxidation) are detectedin liver tissues of rats by TBTC exposure (Liu et al.2006).
The immune system is the major target of TBTCtoxicity. It is widely accepted that exposure to TBTCcan result in suppression of both innate and acquiredimmunity, leading to an increased susceptibility to in-fection. In rats, TBTC causes thymic involution due todepletion of cortical thymocytes (Bressa et al. 1991;Dewaal et al. 1997). TBTC also affects non-specificimmune functions like natural killer (NK) cells(Whalen et al. 2002; Wilson et al. 2004; Aluoch et al.2006) and macrophage activity (Kergosien and Rice1998). Furthermore, TBTC is found to produce T-helper TH2 polarization in vitro and in vivo, as indicatedby suppression of production of TH1-related cytokines
Cell Biol ToxicolDOI 10.1007/s10565-014-9272-7
N. Sharma (*) :A. KumarDepartment of Pharmaceutical Sciences, Birla Institute ofTechnology, Mesra,Ranchi 835215 Jharkhand, Indiae-mail: [email protected]
verses stimulation of TH2 cytokine production (Katoet al. 2004).
In previous studies, it has been reported that TBTCinduces apoptosis in isolated thymocytes at concentra-tions which are relevant to those causing thymus atro-phy in vivo. It can also induce apoptosis in PC12 cells(Nakatsu et al. 2007), hepatocytes (Grondin et al. 2007),and human amnion cells (Zhu et al. 2007). While themechanism of TBTC-induced apoptosis is still unex-plored, it could be related to changes in intracellularcalcium (Reader et al. 1999; Gennari et al. 2000). Itinduces pro-apoptotic signalling in rat hepatocytesthrough the pathways which involve endoplasmic re-ticulum and mitochondria (Grondin et al. 2007).Toxicogenomics studies for in vitro immunotoxicityof bis (tri-n-butyltin) oxide shows that TBTC may exertits toxic effects on the thymus primarily by affectingapoptosis (Baken et al. 2007). Considering theimmunotoxic effects of TBTC on mammalian systemand its mechanism still unclear, it is necessary to inves-tigate all the possible pathways involved during TBTC-induced apoptosis in immune cells which will help inthe amelioration of its toxic effects.
Thus, the aim of present investigation is to explorethe multiple pathways activated by TBTC during apo-ptosis in primary murine thymocytes, taking into ac-count the role of oxidative stress [reactive oxygen spe-cies (ROS) and reduced glutathione (GSH)], mitochon-drial membrane potential, pro-apoptotic molecules suchas caspase-3, apoptosis-inducing factor (AIF), endonu-clease G (Endo G) as well as calcium.
Materials and methods
Chemicals
All the chemicals were of highest grade purity available.Tributyltin-chloride (TBTC), RNase A, RPMI 1640,antibiotic–antimycotic solution, Dulbecco’s phosphatebuffered saline (PBS), fetal bovine serum (FBS),3-(4,5-dimethyl-2-yl)-2,5-diphenyl tetrazolium bromide(MTT), 2′,7′-dichlorofluorescein diacetate (DCFH-DA),caspase inhibitor, DEVD-AFC substrate, FITC conjugat-ed anti-rabbit IgG antibody, and all other chemicals werepurchased from Sigma Aldrich, USA. Rhodamine 123(Rh 123) and 5′-chloromethylfluorescein diacetate(CMF - DA) from Molecular Probes, Propidium Iodide(PI) from Calbiochem, Rabbit anti AIF polyclonal
antibody and rabbit anti Endo G polyclonal antibodyfrom Biovision, [3H] thymidine from BRIT, Bombay.
Preparation of thymocyte suspension
Male BALB/cmice (4–6 weeks old), weight between 20and 25 g were selected for the study. Thymus wasdissected from mice and single cell suspension wasprepared under aseptic conditions. The suspension waspassed through 100-μM stainless steel mesh andsuspended in complete cell culture medium (RPMI1640 containing HEPES and 2 mM glutamine, supple-mented with 10 % FBS and 1 % antibiotic–antimycoticsolution). The cell density was adjusted to 1.5×106
cells/ml and the viability of the freshly isolated cellswas always over 95 % (trypan blue exclusion test).
Assessment of cell viability
The cell viability was measured by the Mosmann meth-od (1983). Cells in RPMI 1640 was seeded at a densityof 1.0×104 cells in 96-well plate. TBTC (0.1, 1, 3, 10,and 30 nM) and DMSO (solvent control) was added for6 and 18 h at 37 °C in a CO2 incubator. Ten microlitresof MTT (5 mg/ml PBS) was added to the wells 4 h priorto the completion of incubation time. The plate wascentrifuged at 1,200×g for 10 min and 100 μl ofDMSO was added after removing the supernatant, todissolve the formazan formed. The absorbance was readat 530 nm after 5 min, in a microplate reader (SynergyHT of BIO-TEK International, USA).
Measurement of caspase-3 activity
Caspases, a family of cysteine proteases, are the centralregulators of apoptosis. Caspase-3 executes apoptosisby cleaving cellular proteins following specific aspartateresidues. The release of cytochome c is also reported toactivate caspase-3 and -9 in several cell types (Gennariet al. 2003). A population of 3.0×106 cells/ml wasincubated with TBTC (0.1, 1, 3, and 10 nM) for 1 and3 h at 37 °C in a CO2 incubator. The cells were scrapedand lysed on ice for 10 min using cell lysis buffer(20 mM EDTA, 10 mM tris HCL, and triton X-100).The reaction buffer (10 mM Tris HCl, 1 mM EDTA,10 mM DTT, 5 % glycerol, pH 7.4) and DEVD-AFCsubstrate (50 μM final concentration) was then addedand further incubated at 37 ºC in dark for 1 h.Fluorescence was measured at excitation and emission
Cell Biol Toxicol
wavelengths of 400 and 505 nm, respectively, on amicroplate reader.
To confirm the role of caspase-3 in TBTC-inducedapoptosis, cells were pretreated for 2 h with 50 μMcaspase-3 inhibitor (Z-DEVD-fmk) followed by TBTCfor 3 h.
Flow cytometry analysis
All the following assays were carried out on thymocytestreated with TBTC (0.1, 1, 3, 10 nM) for different timeintervals (60 min, 1.5, 3, 6, and 18 h) at 37 °C in a CO2
incubator. The flow cytometric analysis was done onBD-LSR flow cytometer (Ormerod 2008). Cell debris,characterized by a low FSC/SSC was excluded fromanalysis. The data was analysed by Cell Quest software(BD Biosciences) and mean fluorescence intensity wasobtained by histogram statistics.
Apoptotic DNA analysis
The cells with hypodiploid DNA were determined bycell cycle studies. After the treatment period, the har-vested cells were washed with PBS and fixed by drop-by-drop addition of ice cold 70 % ethanol and stored at4 °C overnight. The fixed cells were harvested, washedwith PBS, and suspended in 1 ml PBS. Phosphate citratebuffer (200 μl, pH 7.8) was added and the cells incu-bated for 60 min at room temp. After centrifugation, thecells were resuspended in 0.5 ml of PI stain (10 mg PI,0.1 ml Triton-X 100 and 3.7 mg EDTA in 100 ml PBS)and 0.5 ml of RNase A (50 μg/ml) and further incubatedfor 30 min in dark at room temperature. The PI fluores-cence was measured though a FL-2 filter (585 nm) and10,000 events were acquired (Darzynkiewicz et al.1992).
Mitochondrial membrane potential
Rh (Rhodamine) 123, a lipophilic cationic fluorescentdye, is selectively taken up by mitochondria and itsuptake is directly proportional to mitochondrial mem-brane potential of cells (Scaduto and Grotyohann 1999).The cells were incubated with Rh 123 (5 μg/ml finalconcentration) for 60 min in dark at 37 °C, harvested,and suspended in PBS. The mitochondrial membranepotential was measured by the fluorescence intensity(FL-1, 530 nm) of 10,000 cells (Bai et al. 1999).
Reactive oxygen species (ROS) measurement
DCFH-DA, a permeable dye, is cleaved to form nonflu-orescent dichlorofluorescein (DCFH) in the cells, whichgets oxidised to fluorescent dichlorofluorescein (DCF)by ROS. The generation of ROS was detected by DCFfluorescence. The esterified form DCFH-DA can per-meate cell membranes before being deacetylated byintracellular esterases. The resulting compound,dichlorodihydrofluorescein (DCFH), is oxidised to fluo-rescence DCF by ROS. Thymocytes were incubatedwith TBTC and DCFH-DA (100 μM final concentra-tion) was added simultaneously to the medium. Thecells were harvested, suspended in PBS and ROS gen-eration was measured by the fluorescence intensity (FL-1, 530 nm) of 10,000 cells (Wang et al. 1996).
Glutathione (GSH) measurement
The intensity of CMF fluorescence was well correlatedwith biochemically estimated content of GSH(Chikahisa et al. 1996). The cellular level of GSH inthymocytes was monitored by CMF-DA. After treat-ment, the cells were incubated with CMF-DA (1 μMfinal concentration) for 30 min in dark at 37 °C. Afterharvesting, the cells were suspended in PBS and GSHwas measured by the fluorescence intensity (FL-1,530 nm) of 10,000 cells (Okada et al. 1996).
To further confirm the role of oxidative stress inTBTC-induced apoptosis, cells were pretreated withthiol modulators. N-acetyl cysteine (NAC) can raiseintracellular glutathione levels and thereby protect thecells from the effects of ROS. In addition, the –SHgroup of the reagent can react directly with radicals.Cells were pretreated with NAC (10 mM), 10 min priorto the addition to TBTC (3 nM) for 6 h. For GSHdepletion, buthionine sulfoximine (BSO) was included,which acts by inhibiting ν-glutamyl cysteine synthetaseactivity. The cells were pretreated with BSO (500 μM)for 20 h, after which the medium was replaced, TBTC(3 nM) was added and the cells were further incubatedfor 6 h.
Proteins expression
AIF and Endo G expression were observed in the mito-chondrial and nuclear fractions of TBTC-treated thymo-cytes. To prepare the fractions, cells were allowed toswell in ice cold mitochondria buffer (250 mM sucrose,
Cell Biol Toxicol
20 mM HEPES, 10 mM KCl, 1.5 mM MgCl2, 1 mMEDTA, 1 mM EGTA, 1 mM dithiotheitol, 1 mMphenylmethylsulfonyl fluoride, and protease inhibitormixture, pH 7.4) for 20 min on ice. The cells werehomogenized by syringe and centrifuged at 800×g for10 min to collect nuclear fraction. The supernatant wasagain centrifuged at 10,000×g for 30 min for mitochon-drial fraction.
Both the fractions were suspended in 1 ml RPMIbuffer (containing 2 % FBS and 0.1 % sodium azide).An aliquot of 200 μl was incubated with 2 μl of0.5 mg/ml stock solution of rabbit anti AIF polyclonalantibody/rabbit anti Endo G polyclonal antibody for20 min at 2–8 °C. The excess antibody was removedbywashing cells twice with RPMI buffer. The cells werethen resuspended in 200 μl RPMI buffer and binding ofunlabelled antibodies was visualized by adding 5 μl of50 μg/ml stock solution of FITC conjugated anti-rabbitIg G antibody for 20 min at 2–8 °C. After incubation,cells were washed with RPMI buffer, then with PBS and
resuspended in 400 μl PBS. The FITC fluorescence wasmeasured though FL-1 filter (530 nm) and 10,000events were acquired.
Western blotting
The expression of AIF and endo G (nuclear and mito-chondrial fractions) was determined by western immu-noblotting. Both the fractions were suspended in 30 μlloading buffer, heated for 5 min and the lyophilized
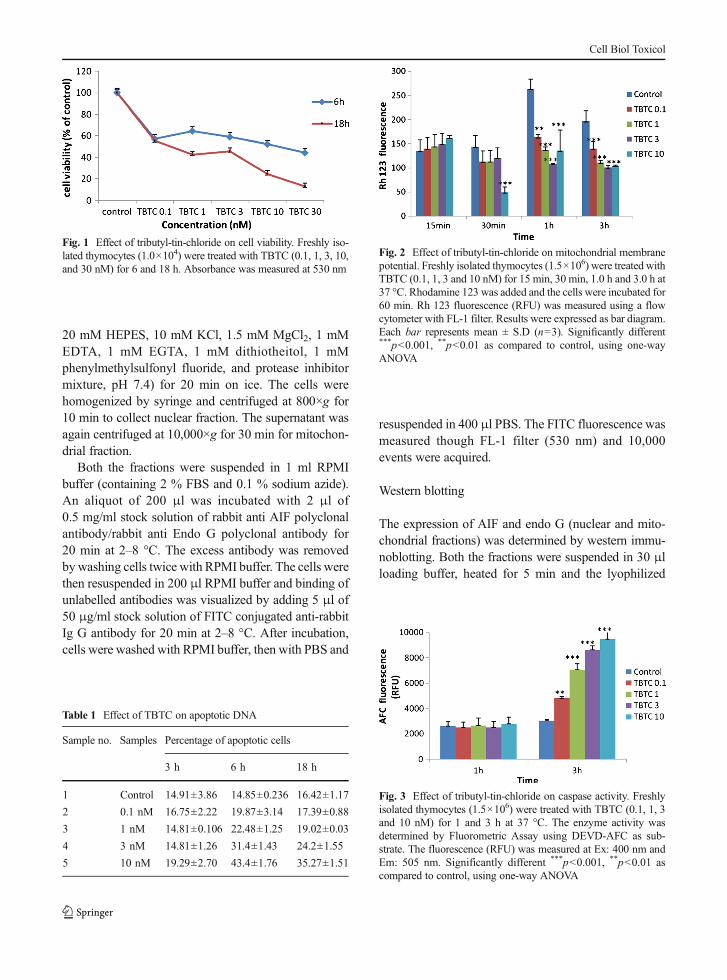
Fig. 1 Effect of tributyl-tin-chloride on cell viability. Freshly iso-lated thymocytes (1.0×104) were treated with TBTC (0.1, 1, 3, 10,and 30 nM) for 6 and 18 h. Absorbance was measured at 530 nm
Table 1 Effect of TBTC on apoptotic DNA
Sample no. Samples Percentage of apoptotic cells
3 h 6 h 18 h
1 Control 14.91±3.86 14.85±0.236 16.42±1.17
2 0.1 nM 16.75±2.22 19.87±3.14 17.39±0.88
3 1 nM 14.81±0.106 22.48±1.25 19.02±0.03
4 3 nM 14.81±1.26 31.4±1.43 24.2±1.55
5 10 nM 19.29±2.70 43.4±1.76 35.27±1.51
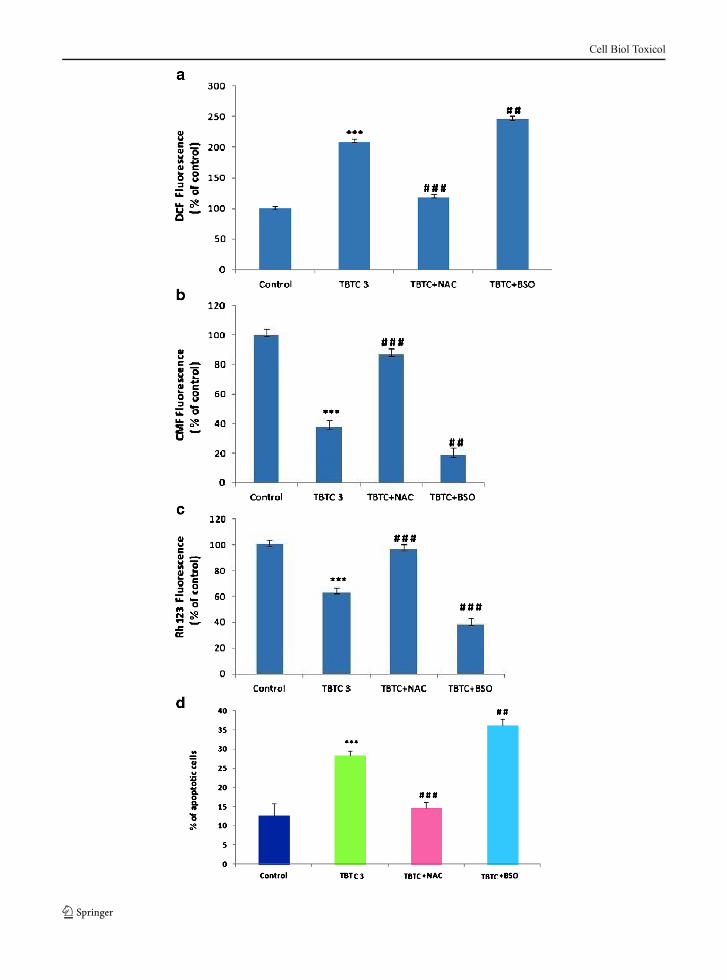
Fig. 2 Effect of tributyl-tin-chloride on mitochondrial membranepotential. Freshly isolated thymocytes (1.5×106) were treated withTBTC (0.1, 1, 3 and 10 nM) for 15 min, 30 min, 1.0 h and 3.0 h at37 °C. Rhodamine 123 was added and the cells were incubated for60 min. Rh 123 fluorescence (RFU) was measured using a flowcytometer with FL-1 filter. Results were expressed as bar diagram.Each bar represents mean ± S.D (n=3). Significantly different***p<0.001, **p<0.01 as compared to control, using one-wayANOVA
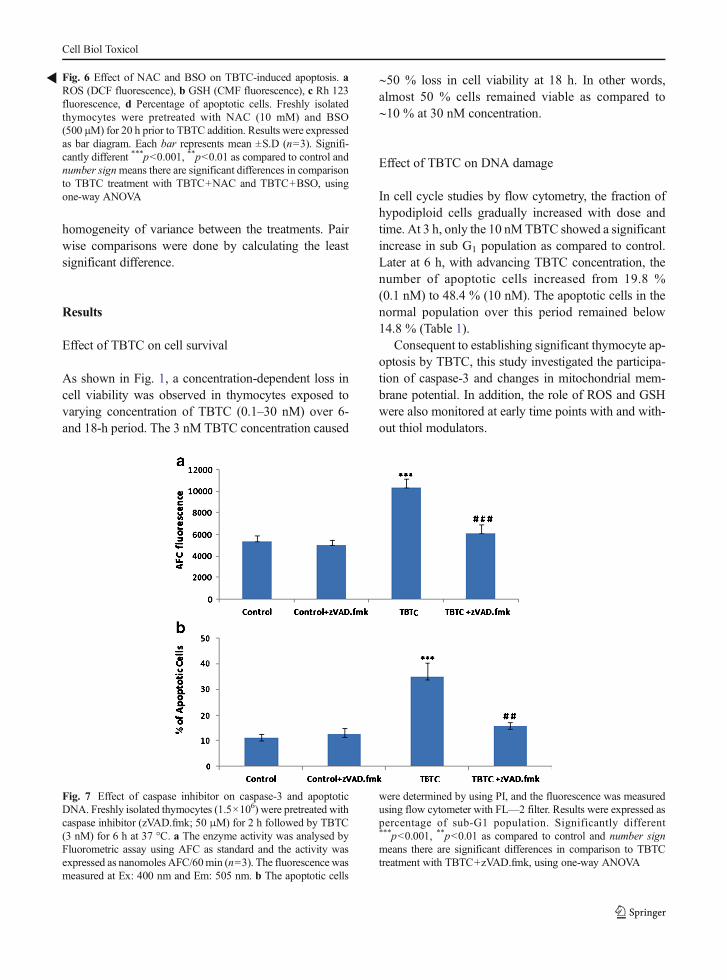
Fig. 3 Effect of tributyl-tin-chloride on caspase activity. Freshlyisolated thymocytes (1.5×106) were treated with TBTC (0.1, 1, 3and 10 nM) for 1 and 3 h at 37 °C. The enzyme activity wasdetermined by Fluorometric Assay using DEVD-AFC as sub-strate. The fluorescence (RFU) was measured at Ex: 400 nm andEm: 505 nm. Significantly different ***p<0.001, **p<0.01 ascompared to control, using one-way ANOVA
Cell Biol Toxicol
nuclear and mitochondrial proteins (50 μg) were sepa-rated by SDS-PAGE on 10 % acrylamide gel. Beforeloading, amount of protein was estimated by Bradfordreagent. The protein bands were transferred onto nitro-cellulose membranes. Blots were blocked with 5 %nonfat dry milk for 1 h and incubated overnight at4 °C with primary anti-AIF (1:500) or anti-Endo G(1:500) antibodies in blocking solution. After three to
five washings of 5 min each with wash buffer (PBS and0.05 % Tween-20), blots were incubated with HP-conjugated secondary antibody (1:1,000) in blockingsolution for 2 h at room temperature. The colour wasdeveloped by DAB system (Bangalore Genei) afterthree to five washings.
Measurement of intracellular calcium
The intracellular calcium level was determined by usinga fluorescent dye FURA-2 AM. Thymocytes were in-cubated with TBTC (3 nM) for 15 min, 30 min, 1, 2, 4,and 6 h at 37 °C and FURA-2 AM (1.0 μM finalconcentration) was added simultaneously to the medi-um. This fluorescent probe has been found suitable forthe detection of intracellular free Ca2+. The cells wereharvested, suspended in PBS and fluorescence wasmeasured at excitation and emission wavelengths of340 and 505 nm, respectively, on a microplate reader.
To inhibit Ca2+, the cells were pretreated withRuthenium red (RR) (5.0 μM), a potent inhibitor ofmitochondrial calcium uniporter (Shih et al. 2005) for1.0 h followed by TBTC.
Statistical analysis
Significance of mean of different parameters betweenthe treatment groups were analysed using one-way anal-ysis of variance (ANOVA) after ascertaining the
Fig. 4 Effect of tributyl-tin-chloride on ROS generation. Freshlyisolated thymocytes (1.5×106) were incubated with DCFH-DA(100 μM) and TBTC (0.1, 1, 3 and 10 nM) for 15 min, 1, 3, 6, and18 h at 37 °C. DCF fluorescence was measured using a flowcytometer with FL-1 filter. Fluorescence (RFU) results wereexpressed as bar diagram. Each bar represents mean ± S.D(n=3). Significantly different ***p<0.001, **p<0.01, *p<0.05 ascompared to control, using one-way ANOVA
Fig. 5 Effect of tributyl-tin-chloride on GSH levels. Freshly iso-lated thymocytes (1.5×106) were treated with TBTC (0.1, 1, 3 and10 nM) for 15 min and 1, 3, 6, and 18 h at 37 °C. CMF-DAwasadded and the cells were incubated for 30 min. CMF fluorescence(RFU) was measured using a flow cytometer with FL—1 filter.
Fluorescence results were expressed as representative histogram(a, 6 h) and bar diagram (b). Each bar represents mean ± S.D (n=3). Significantly different ***p<0.001, **p<0.01, *p<0.05 as com-pared to control
Cell Biol Toxicol
Cell Biol Toxicol
homogeneity of variance between the treatments. Pairwise comparisons were done by calculating the leastsignificant difference.
Results
Effect of TBTC on cell survival
As shown in Fig. 1, a concentration-dependent loss incell viability was observed in thymocytes exposed tovarying concentration of TBTC (0.1–30 nM) over 6-and 18-h period. The 3 nM TBTC concentration caused
∼50 % loss in cell viability at 18 h. In other words,almost 50 % cells remained viable as compared to∼10 % at 30 nM concentration.
Effect of TBTC on DNA damage
In cell cycle studies by flow cytometry, the fraction ofhypodiploid cells gradually increased with dose andtime. At 3 h, only the 10 nMTBTC showed a significantincrease in sub G1 population as compared to control.Later at 6 h, with advancing TBTC concentration, thenumber of apoptotic cells increased from 19.8 %(0.1 nM) to 48.4 % (10 nM). The apoptotic cells in thenormal population over this period remained below14.8 % (Table 1).
Consequent to establishing significant thymocyte ap-optosis by TBTC, this study investigated the participa-tion of caspase-3 and changes in mitochondrial mem-brane potential. In addition, the role of ROS and GSHwere also monitored at early time points with and with-out thiol modulators.
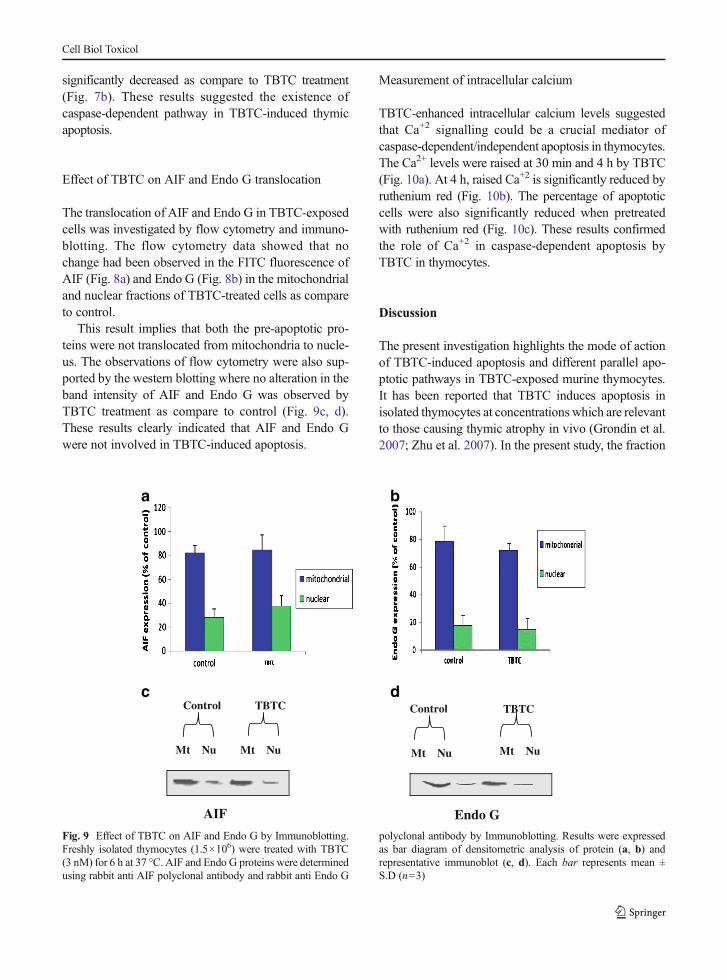
Fig. 6 Effect of NAC and BSO on TBTC-induced apoptosis. aROS (DCF fluorescence), b GSH (CMF fluorescence), c Rh 123fluorescence, d Percentage of apoptotic cells. Freshly isolatedthymocytes were pretreated with NAC (10 mM) and BSO(500 μM) for 20 h prior to TBTC addition. Results were expressedas bar diagram. Each bar represents mean ±S.D (n=3). Signifi-cantly different ***p<0.001, **p<0.01 as compared to control andnumber signmeans there are significant differences in comparisonto TBTC treatment with TBTC+NAC and TBTC+BSO, usingone-way ANOVA
R
Fig. 7 Effect of caspase inhibitor on caspase-3 and apoptoticDNA. Freshly isolated thymocytes (1.5×106) were pretreated withcaspase inhibitor (zVAD.fmk; 50 μM) for 2 h followed by TBTC(3 nM) for 6 h at 37 °C. a The enzyme activity was analysed byFluorometric assay using AFC as standard and the activity wasexpressed as nanomoles AFC/60min (n=3). The fluorescence wasmeasured at Ex: 400 nm and Em: 505 nm. b The apoptotic cells
were determined by using PI, and the fluorescence was measuredusing flow cytometer with FL—2 filter. Results were expressed aspercentage of sub-G1 population. Significantly different***p<0.001, **p<0.01 as compared to control and number signmeans there are significant differences in comparison to TBTCtreatment with TBTC+zVAD.fmk, using one-way ANOVA
Cell Biol Toxicol
Effect of TBTC on mitochondrial membrane potential
Being an early marker of apoptosis, a significant de-crease in mitochondrial membrane potential occurred asearly as 30 min, only at 10 nM TBTC concentration(p<0.001) (Fig. 2). Later on, at 1 and 3 h, the loss inmembrane potential became more prominent and wasconcentration related.
Effect of TBTC on caspase-3 activity
There was no increase in caspase activity at 1 h but at3 h, there was a dose and time-dependent enhancementin caspase-3 activity (Fig. 3). These results indicate thatthere is mitochondrial membrane depolarization,coupled with caspase-3 activation during TBTC-induced thymic apoptosis.
Effect of TBTC on ROS generation
The DCF fluorescence, proportionate to the ROS levelsin the cells, was monitored on flow cytometer. The doseand time-dependent generation of ROS continued to risetill 6 h. However at 18 h, the DCF fluorescence dimin-ished (Fig. 4). Significant ROS generation in the thy-mocytes was observed as early as 15 min with the twohigher TBTC concentrations (3 and 10 nM), prior to thelowering in mitochondrial membrane potential whichwas observed only at 30 min with 10 nM TBTC.
Effect of TBTC on intracellular GSH
GSH depletion started as early as 15 min and at latertime periods, there was a dose-dependent depletion ofGSH till 18 h. The 10 nM TBTC concentration caused asignificant reduction in GSH levels at 15 min, 1, 3, 6,and 18 h (Fig. 5). It appears that GSH and ROS may actas intracellular signals causing mitochondrial membranepermeabilization at 30 min and the triggering effectbeing translated to DNA damage evident at 3 h with10 nM TBTC and at 18 h with all the concentrations.
Effect of thiol modulators on TBTC-induced apoptosis
To further investigate whether ROS generation is acrucial mediator in TBTC-induced apoptosis, thiol mod-ulators were incorporated. The antioxidant (NAC)lowered the DCF fluorescence and increased the CMFfluorescence while on other hand, BSO enhanced the
DCF fluorescence and reduced the CMF fluorescence(Fig. 6a, b).
The lowered Rh 123 fluorescence in the TBTC-exposed cells was enhanced drastically in the presenceof NAC and further inhibited with BSO. Regarding theapoptotic DNA (sub G1 population), NAC effectivelyinhibited the TBTC-induced apoptotic DNA. In con-trast, BSO caused an elevation in the sub G1 populationas shown in (Fig. 6d). These results confirmed the roleof oxidative stress in TBTC-induced apoptosis.
Effect of Caspase inhibitor on TBTC-induced apoptosis
The release of cytochome c is reported to activatecaspase-3 and -9 in several cell types. TBTC treatmentcaused almost 3.0-fold activation of caspase-3. To ana-lyse further the mechanisms responsible for TBTC-induced apoptosis, caspase-3 activity was inhibited byZ-DEVD-fmk followed by TBTC for 3 h (Fig. 7a). Inthe presence of inhibitor, the resultant protease (caspase-3)activity and the percentage of apoptotic cells were
Fig. 8 Effect of TBTC on AIF and Endo G by flow cytometry.Freshly isolated thymocytes (1.5×106) were treated with TBTC(3 nM) for 6 h at 37 °C. AIF and Endo G proteins were determinedusing rabbit anti AIF polyclonal antibody and rabbit anti Endo Gpolyclonal antibody by flow cytometry. Results were expressed asbar diagram (a, b). Each bar represents mean ± S.D (n=3)
Cell Biol Toxicol
significantly decreased as compare to TBTC treatment(Fig. 7b). These results suggested the existence ofcaspase-dependent pathway in TBTC-induced thymicapoptosis.
Effect of TBTC on AIF and Endo G translocation
The translocation of AIF and Endo G in TBTC-exposedcells was investigated by flow cytometry and immuno-blotting. The flow cytometry data showed that nochange had been observed in the FITC fluorescence ofAIF (Fig. 8a) and Endo G (Fig. 8b) in the mitochondrialand nuclear fractions of TBTC-treated cells as compareto control.
This result implies that both the pre-apoptotic pro-teins were not translocated from mitochondria to nucle-us. The observations of flow cytometry were also sup-ported by the western blotting where no alteration in theband intensity of AIF and Endo G was observed byTBTC treatment as compare to control (Fig. 9c, d).These results clearly indicated that AIF and Endo Gwere not involved in TBTC-induced apoptosis.
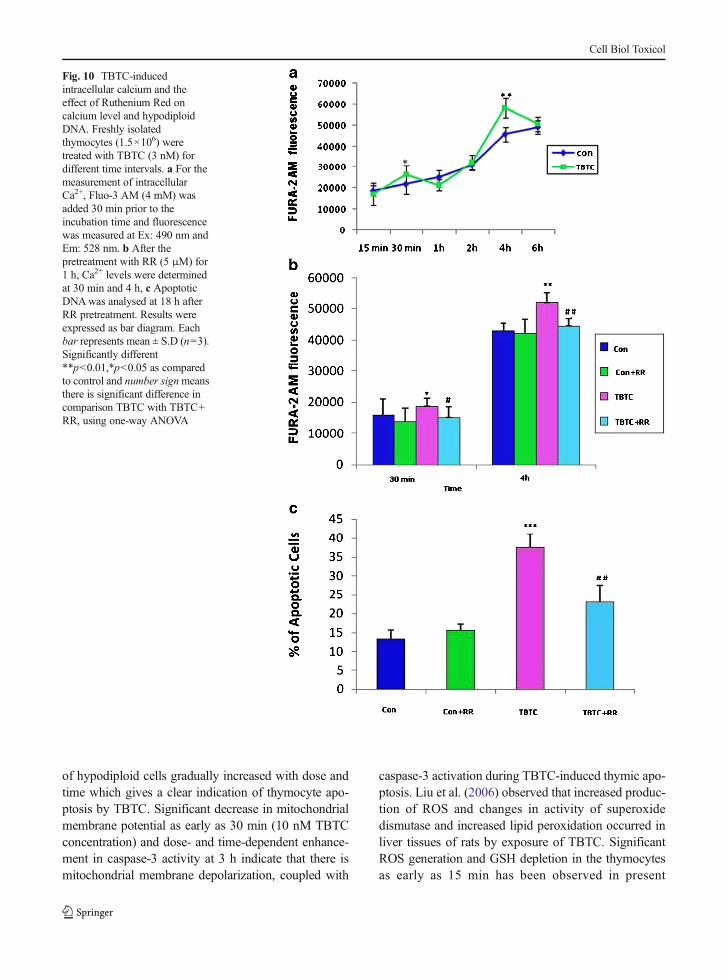
Measurement of intracellular calcium
TBTC-enhanced intracellular calcium levels suggestedthat Ca+2 signalling could be a crucial mediator ofcaspase-dependent/independent apoptosis in thymocytes.The Ca2+ levels were raised at 30 min and 4 h by TBTC(Fig. 10a). At 4 h, raised Ca+2 is significantly reduced byruthenium red (Fig. 10b). The percentage of apoptoticcells were also significantly reduced when pretreatedwith ruthenium red (Fig. 10c). These results confirmedthe role of Ca+2 in caspase-dependent apoptosis byTBTC in thymocytes.
Discussion
The present investigation highlights the mode of actionof TBTC-induced apoptosis and different parallel apo-ptotic pathways in TBTC-exposed murine thymocytes.It has been reported that TBTC induces apoptosis inisolated thymocytes at concentrations which are relevantto those causing thymic atrophy in vivo (Grondin et al.2007; Zhu et al. 2007). In the present study, the fraction
Control TBTC
Mt Nu Mt Nu
Control TBTC
Mt Nu Mt Nu
AIF Endo G
a b
c d
Fig. 9 Effect of TBTC on AIF and Endo G by Immunoblotting.Freshly isolated thymocytes (1.5×106) were treated with TBTC(3 nM) for 6 h at 37 °C. AIF and Endo G proteins were determinedusing rabbit anti AIF polyclonal antibody and rabbit anti Endo G
polyclonal antibody by Immunoblotting. Results were expressedas bar diagram of densitometric analysis of protein (a, b) andrepresentative immunoblot (c, d). Each bar represents mean ±S.D (n=3)
Cell Biol Toxicol
of hypodiploid cells gradually increased with dose andtime which gives a clear indication of thymocyte apo-ptosis by TBTC. Significant decrease in mitochondrialmembrane potential as early as 30 min (10 nM TBTCconcentration) and dose- and time-dependent enhance-ment in caspase-3 activity at 3 h indicate that there ismitochondrial membrane depolarization, coupled with
caspase-3 activation during TBTC-induced thymic apo-ptosis. Liu et al. (2006) observed that increased produc-tion of ROS and changes in activity of superoxidedismutase and increased lipid peroxidation occurred inliver tissues of rats by exposure of TBTC. SignificantROS generation and GSH depletion in the thymocytesas early as 15 min has been observed in present
Fig. 10 TBTC-inducedintracellular calcium and theeffect of Ruthenium Red oncalcium level and hypodiploidDNA. Freshly isolatedthymocytes (1.5×106) weretreated with TBTC (3 nM) fordifferent time intervals. a For themeasurement of intracellularCa2+, Fluo-3 AM (4 mM) wasadded 30 min prior to theincubation time and fluorescencewas measured at Ex: 490 nm andEm: 528 nm. b After thepretreatment with RR (5 μM) for1 h, Ca2+ levels were determinedat 30 min and 4 h, c ApoptoticDNAwas analysed at 18 h afterRR pretreatment. Results wereexpressed as bar diagram. Eachbar represents mean ± S.D (n=3).Significantly different**p<0.01,*p<0.05 as comparedto control and number signmeansthere is significant difference incomparison TBTC with TBTC+RR, using one-way ANOVA
Cell Biol Toxicol
investigation. This earliest response in oxidative stress(ROS production and GSH depletion) followed by thelowering of mitochondrial membrane potential and ac-tivation of caspase-3 would then lead the cells toapoptose.
Mitra et al. (2013) demonstrated that TBTC candisrupt blood brain barrier, induce oxidative stress,cause cell death, and initiate neurodegeneration in ratbrain. Various studies demonstrate that oxidative stressplays the key role in the process of apoptosis (Pathakand Khandelwal 2007; Kumar et al. 2013; Mitra et al.2013). Thus, to further investigate the role of oxidativestress, thymocytes are pretreated with NAC and BSOfollowed by TBTC. The percentage of apoptotic cellsare significantly reduced as NAC effectively minimizedthe enhanced ROS and raised the intracellular GSHlevels. On the other hand, BSO significantly enhancedapoptotic cells when the cells were depleted of GSH.These results imply that oxidative stress (ROS increaseand GSH decrease) play major role in TBTC-inducedapoptosis.
Nakatsu et al. (2006) observed that caspase-3 inhib-itor had no effect on tributyltin toxicity, and tributyltindid not change the nuclear morphology of cultured ratcortical neurons. In the present investigation, the resul-tant protease (caspase-3) activity, enhanced almost 3.0-fold by TBTC, was significantly inhibited in the pres-ence of caspase inhibitor. This result suggested theexistence of caspase-dependent pathway in TBTC-induced thymic apoptosis in murine thymocytes.
The release of mitochondrial proapoptotic proteinssuch as cytochome c (caspase dependent), AIF, andendo G (caspase independent) have been reported dur-ing the cytotoxicity induced by several organotin com-pounds (Stridh et al. 1998; Grondin et al. 2007). DNAfragmentation by endonucleases was observed as anearly event in tri-n-butylstannyl benzoate (TBSB)-treat-ed lymphocytic cells of human leukemic K562 lineage(Ray et al. 2000). In this study, flow cytometric data andimmunoblotting studies revealed that there is no trans-location of AIF and Endo G from mitochondria tonucleus. Thus, there is no involvement of AIF andEndo G in TBTC-induced apoptosis.
The possibility of mitochondrial–calcium pathwayalso exists, since calcium signals have been identifiedas one of the major signals that converge on mitochon-dria to trigger apoptosis. (Ray et al. 2000), has beenreported that TBTC-induced apoptosis by causing ex-tracellular Ca2+ influx. In the present investigation,
intracellular calcium levels were significantly enhancedby the treatment of TBTC. Ruthenium Red completelyabolished the raised calcium levels and influenced thecaspase activity as well as apoptosis, implying that theCa2+-dependent-mitochondria–caspase pathway isprevalent.
In summary, this study clearly demonstrates the roleof oxidative stress (ROS production and GSH depletion)followed by the lowering of mitochondrial membranepotential in TBTC-induced caspase-dependent apopto-sis pathway in murine thymocytes. Regarding caspase-independent pathway, there is no involvement AIF andEndo G preapoptotic proteins. Calcium signalling alsoacts as a crucial mediator of TBTC-induced apoptoticeffects. Thus, the present investigation provides a betterunderstanding of the mechanism of TBTC-induced ap-optosis in murine thymocytes.
Acknowledgments The authors acknowledge DST for finicalsupport in the form of DST-INSPIRE fellowship and are alsograteful to Dr. Shashi khandelwal, Head, Immunotoxicity labora-tory, IITR, India, for her valuable suggestions and guidance forconducting this work.
References
Aluoch AO, Odman-Ghazi SO, Whalen MM. Alteration of anessential NK cell signaling pathway by low doses of tribu-tyltin in human natural killer cells. Toxicol. 2006;224:229–37.
Bai J, Rodriguez AM, Malendez JA, Cederbaum AI.Overexpression of catalase in cytosolic or mitochondrialcompartment protects HepG2 cells against oxidative injury.J Biol Chem. 1999;274:26217–24.
Baken KA, Arkusz J, Pennings J, Vandebriel LA, Van Loveren H.In vitro immunotoxicity of bis(tri-n-butyltin)oxide (TBTO)studied by toxicogenomics. Toxicol. 2007;237:35–48.
Bressa G, Hinton RH, Price SC, Isbir M, Ahmed RS, Grasso P.Immunotoxicity of tri-n-butyltin oxide (TBTO) and tri-n-butyltin chloride (TBTC) in the rat. J Appl Toxicol.1991;11:397–402.
Chikahisa L, Oyama Y, Okazaki E, Noda K. Fluorescent estima-tion of H2 O2-induced changes in cell viability and cellularnonprotein thiol level of dissociated rat thymocytes. JPharmacol. 1996;71:299–305.
Darzynkiewicz Z, Bruno S, Del Bino G, Gorczyca W, Hotz MA,Lassota P, et al. Features of apoptosis cellsmeasured by flowcytometry. Cytometry. 1992;13:795–808.
Dewaal EJ, Schuurman HJ, Van Loveren H, Vos JG. Differentialeffects of 2,3,7,8-tetrachlorodibenzo-p-dioxin, bis(tri-n-butyltin) oxide and cyclosporine on thymus histophysiology.Crit Rev Toxicol. 1997;27:381–430.
Cell Biol Toxicol
Dimitriou P, Castritsi-Catharios J, Miliou H. Acute toxicity effectsof tributyltin chloride and triphenyltin chloride on giltheadseabream, Sparus aurata L., embryos. Ecotoxicol EnvironSaf. 2003;54:30–5.
Gennari A, Cortese E, Boveri M, Casado J, Prieto P. Sensitiveendpoints for evaluating cadmium-induced acute toxicity inLLC-PK1 cells. Toxicol. 2003;183:211–20.
Gennari A, Viviani B, Galli CL, Marinovich M, Pieters R, CorsiniE. Organotins induce apoptosis by disturbance of [Ca2+] andmitochondrial activity, causing oxidative stress and activationof caspases in rat thymocytes. Toxicol Appl Pharmacol.2000;169:185–90.
Grondin M, Marion M, Denizeau F, Averill-Bates DA. Tributyltininduces apoptotic signaling in hepatocytes though pathwaysinvolving the endoplasmic reticulum and mitochondria.Toxicol App Pharmacol. 2007;222:57–68.
Grote K, Stahlschmidt B, Talsness CE, Gericke C, Appel KE,Chahoud I. Effects of organotin compounds on pubertal malerats. Toxicol. 2004;202:145–58.
Inadera H. The immune system as a target for environmentalchemicals: xenoestrogens and other compounds. ToxicolLett. 2006;164:191–206.
Isomura M, Kotake Y, Masuda K, Miyara M, Okuda K, Samizo S,et al. Tributyltin-induced endoplasmic reticulum stress and itsCa2+-mediated mechanism. Toxicol Appl Pharmacol.2013;272:137–46.
Kato T, Uchikawa R, Yamada M, Arizono N, Oikawa S,Kawanishi S. Environmental pollutant tributyltin promotesTh2 polarization and exacerbates airway inflammation. Eur JImmunol. 2004;34:1312–21.
Kergosien DH, Rice CD. Macrophage secretory function is en-hanced by low doses of tributyltin-oxide (TBTO), but nottributyltin-chloride (TBTC). Arch Environ Contam Toxicol.1998;34:223–8.
Kumar A, Sasmal D and Sharma N. Deltamethin induced anapoptogenic signalling pathway in murine thymocytes: ex-ploring themolecular mechanism. 2013: doi:10.1002/jat.2948.
Liu SM, Hsia MP, Huang CM. Accumulation of butylin com-pounds in cobia rachycentron canadum raised in off shoreaquaculture sites. Sci Total Environ. 2006;355:167–75.
Mitra S, Gera R, Siddiqui W, Khandelwal S. Tributyltin inducesoxidative damage, inflammation and apoptosis via distur-bance in blood–brain barrier and metal homeostasis in cere-bral cortex of rat brain: an in vivo and in vitro study. Toxicol.2013;310:39–52.
Mosmann T. Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity assays.J Immunol Methods. 1983;65:55–63.
Nakatsu Y, Kotake Y, Ohta S. Concentration dependence of themechanisms of tributyltin-induced apoptosis. Toxicol Sci.2007;97:438–47.
Nakatsu Y, Kotake Y, Komasaka K, Hakozaki H, Taguchi R,Kume T, et al. Glutamate excitotoxicity is involved in celldeath caused by tributyltin in cultured rat cortical neurons.Toxicol Sci. 2006;89:235–42.
Okada S, Takehara Y, Yabuki M, Yoshioka T, Yasuda T, Inoue M,et al. Nitric oxide, a physiological modulator of mitochon-drial function. Physiol Chem Phys Med. 1996;28:69–82.
Ormerod MG. Flow cytometry—a basic introduction. USA DeNovo; 2008: 2–4.
Pathak N, Khandelwal S. Role of oxidative stress and apoptosis incadmium induced thymic atrophy and splenomegaly in mice.Toxicol Lett. 2007;169:95–108.
Ray D, Sarma KD, Antony A. Differential effects of tri-n-butylstannyl benzoates on induction of apoptosis in K562and MCF-7 cells. IUBMB Life. 2000;49:519–25.
Reader SV, Moutardier, Denizeau F. Tributyltin triggers apoptosisin trout hepatocytes: the role of Ca2+, protein kinase Cand proteases. Biochim Biophys Acta. 1999;1448:473–85.
Scaduto Jr, Grotyohann LW.Measurement of mitochondrial mem-brane potential using fluorescent rhodamine derivatives. JBiophys. 1999;76:469–77.
Shih YL, Lin CJ, Hsu SW. Cadmium toxicity toward caspase-independent apoptosis though the mitochondrial-calciumpathway in mtDNA-depleted cells. Ann N Y Acad Sci.2005;1042:497–505.
Stridh H, Fava E, Single B, Nicotera P, Orrenius S. Tributyltin-induced apoptosis requires glycolytic adenosine trisphos-phate production. Res Toxicol. 1999;12:874–82.
Stridh H, Kimland M, Jones DP, Orrenius S, Hampton MB.Cytochrome c release and caspase activation in hydrogenperoxide- and tributyltin-induced apoptosis. FEBS Lett.1998;429:351–5.
Vivani B, MarinovichGalli CL. Action modification and calciumhomoeostasis in neurotoxicity: the case of organotin salts. JToxicol In Vitro. 1997;11:499–503.
Wang JF, Jerrells TR, Spitzer JH. Decreased production of reactiveoxygen intermediates is an early event during in vitro apo-ptosis of rat thymocytes. Free Radic Biol Med. 1996;20:533–42.
Whalen MM, Green SA, Loganathan BG. Brief butyltin exposureinduces irreversible inhibition of the cytotoxic function onhuman natural killer cells. In vitro Environ Res. 2002;88:19–29.
Wilson S, Dzon L, Reed A, Pruitt M, Whalen MM. Effects ofin vitro exposure to low levels of organotin and carbamatepesticides on human natural killer cell cytotoxic function.Environ Toxicol. 2004;19:554–63.
Zhu X, Xing M, Lou J, Wang X, Fu W, Xu L. Apoptotic relatedbiochemical changes in human amnion cells induced bytributyltin. Toxicol. 2007;230:45–52.