Page 1
BRAINA JOURNAL OF NEUROLOGY
Melatonin augments hypothermic neuroprotectionin a perinatal asphyxia modelNicola J. Robertson,1 Stuart Faulkner,1 Bobbi Fleiss,2,3,4 Alan Bainbridge,5
Csilla Andorka,1 David Price,5 Elizabeth Powell,1 Lucy Lecky-Thompson,1 Laura Thei,1
Manigandan Chandrasekaran,1 Mariya Hristova,1 Ernest B. Cady,5 Pierre Gressens,2,3,4
Xavier Golay6 and Gennadij Raivich1
1 Institute for Women’s Health, University College London, London WC1E 6AU, UK
2 Department of Surgery and Cancer, Imperial College, Centre for the Developing Brain, Institute of Reproductive and Developmental Biology,
London W12 ONN, UK
3 Inserm U676, Hopital Robert-Debre, 75019 Paris, France
4 Faculte de Medecine, Universite Paris Diderot, 75205 Paris, France
5 Department of Medical Physics and Bio-engineering, University College London Hospitals NHS Foundation Trust, London WC1E 6DB, UK
6 UCL Institute of Neurology, London WC1N 3BG, UK
Correspondence to: Nicola J. Robertson,
Institute for Women’s Health,
University College London,
74 Huntley Street,
London WC1E 6AU, UK
E-mail: [email protected]
Despite treatment with therapeutic hypothermia, almost 50% of infants with neonatal encephalopathy still have adverse out-
comes. Additional treatments are required to maximize neuroprotection. Melatonin is a naturally occurring hormone involved
in physiological processes that also has neuroprotective actions against hypoxic–ischaemic brain injury in animal models.
The objective of this study was to assess neuroprotective effects of combining melatonin with therapeutic hypothermia after
transient hypoxia–ischaemia in a piglet model of perinatal asphyxia using clinically relevant magnetic resonance spectroscopy
biomarkers supported by immunohistochemistry. After a quantified global hypoxic–ischaemic insult, 17 newborn piglets were
randomized to the following: (i) therapeutic hypothermia (33.5�C from 2 to 26 h after resuscitation, n = 8) and (ii) therapeutic
hypothermia plus intravenous melatonin (5 mg/kg/h over 6 h started at 10 min after resuscitation and repeated at 24 h, n = 9).
Cortical white matter and deep grey matter voxel proton and whole brain 31P magnetic resonance spectroscopy were acquired
before and during hypoxia–ischaemia, at 24 and 48 h after resuscitation. There was no difference in baseline variables, insult
severity or any physiological or biochemical measure, including mean arterial blood pressure and inotrope use during the 48 h
after hypoxia–ischaemia. Plasma levels of melatonin were 10 000 times higher in the hypothermia plus melatonin than hypo-
thermia alone group. Melatonin-augmented hypothermia significantly reduced the hypoxic–ischaemic-induced increase in the
area under the curve for proton magnetic resonance spectroscopy lactate/N-acetyl aspartate and lactate/total creatine ratios in
the deep grey matter. Melatonin-augmented hypothermia increased levels of whole brain 31P magnetic resonance spectroscopy
nucleotide triphosphate/exchangeable phosphate pool. Correlating with improved cerebral energy metabolism, TUNEL-positive
nuclei were reduced in the hypothermia plus melatonin group compared with hypothermia alone in the thalamus, internal
capsule, putamen and caudate, and there was reduced cleaved caspase 3 in the thalamus. Although total numbers of microglia
were not decreased in grey or white matter, expression of the prototypical cytotoxic microglial activation marker CD86 was
decreased in the cortex at 48 h after hypoxia–ischaemia. The safety and improved neuroprotection with a combination of
melatonin with cooling support phase II clinical trials in infants with moderate and severe neonatal encephalopathy.
doi:10.1093/brain/aws285 Brain 2013: 136; 90–105 | 90
Received June 29, 2012. Revised August 31, 2012. Accepted September 3, 2012. Advance Access publication November 26, 2012
� The Author (2012). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved.
For Permissions, please email: [email protected]
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 2
Keywords: hypoxia–ischaemia; therapeutic hypothermia; neuroprotection; melatonin; neonatal encephalopathy
Abbreviations: HypoT = hypothermia alone group; HypoT + Mel = hypothermia plus melatonin group; IBA1 = ionized calciumbinding adaptor molecule 1; MRS = magnetic resonance spectroscopy; NTP = nucleotide triphosphate; TUNEL = transferase-mediated deoxyuridine triphophate nick-end labelling
IntroductionPerinatal hypoxic–ischaemic brain injury of the term infant remains
a significant problem throughout the world. After 2 decades of
laboratory studies (Thoresen et al., 1995; Bona et al., 1998), clin-
ical trials (Edwards et al., 2010) and recent endorsement from
regulatory bodies (NICE, 2010), therapeutic hypothermia is
becoming standard clinical care for moderate to severe neonatal
encephalopathy in the developed world. Therapeutic hypothermia,
however, is only partially effective, with almost 50% of treated
infants having adverse outcomes (Edwards et al., 2010). Previous
studies have shown that it may be possible to augment neuropro-
tection of the developing brain using a combination of adjunct
drugs with cooling, and thus reduce the number of affected in-
fants (Liu et al., 2004; Jatana et al., 2006).
Melatonin (N-acetyl-5-methoxytryptamine) is a naturally occur-
ring hormone secreted by the pineal gland in response to environ-
mental light–dark cycles (Lynch et al., 1975; Lewy et al., 1980).
Melatonin has many other biological functions in different organs
and tissues (Reiter et al., 2010) that include defence against oxidative
stress (Tan et al., 2007), balancing energy metabolism (Leon et al.,
2004), enhancement of immune function (Altun and Ugur-Altun,
2007) and retarding ageing (Bubenik and Konturek, 2011).
Melatonin acts through several different pathways to modulate
the physiology and molecular biology of cells. Many actions are
mediated through the well-characterized G protein-coupled mela-
tonin receptors in cellular membranes (Brzezinski, 1997; Luchetti
et al., 2010); other actions of melatonin involve its interaction with
orphan nuclear receptors and other molecules such as calmodulin
in the cytosol (Benıtez-King and Anton-Tay, 1993). Non–receptor-
mediated actions of melatonin and its metabolites relate to its ability
to detoxify reactive oxygen species (Tan et al., 2002). Intriguingly,
the synthesis of melatonin may be inducible as a result of oxidative
stress after brain injury (Seifman et al., 2008).
Melatonin has neuroprotective actions against hypoxic–ischae-
mic brain injury in animal models. In adult stroke models, delayed
intravenous administration of melatonin enhances electrophysiolo-
gical and neurobehavioural recoveries (Lee et al., 2004), reduces
cortical and striatal infarct sizes (Pei et al., 2002, 2003; Lee et al.,
2005) and reduces the intracerebral inflammatory response (Lee
et al., 2007). Neuroprotection was seen with both single and mul-
tiple 5-mg/kg injections when commenced within 2 h of middle
cerebral artery occlusion in adult rats (Pei et al., 2003). In foetal
sheep, direct infusion of melatonin reduced inflammation and cell
death in white matter after umbilical cord occlusion (Welin et al.,
2007); neuroprotection was also seen in sheep foetus when mela-
tonin was administered to the mother at the time of asphyxia
(Miller et al., 2005).
As therapeutic hypothermia is already a routine therapy for neo-
natal encephalopathy, it was important to determine whether
melatonin augments hypothermic neuroprotection. Therefore,
the aim of this study was to assess neuroprotective effects of
melatonin-augmented hypothermia compared with hypothermia
alone after a global hypoxic–ischaemic insult in a piglet model of
perinatal asphyxia, using intravenous melatonin administered
10 min after resuscitation combined with cooling. The treatment
effect was explored using the following clinically relevant bio-
markers: in vivo proton (1H) magnetic resonance spectroscopy
(MRS) for lactate, N-acetyl aspartate, creatine (Cheong et al.,
2006; Thayyil et al., 2010), 31P-MRS for inorganic phosphate,
phosphocreatine and nucleotide triphosphate (NTP) (Azzopardi
et al., 1989) and serial EEG and amplitude-integrated EEG (Toet
et al., 1999). At the end of the 48-h survival period, histological
assessment to quantify cell death was performed using transferase-
mediated deoxyuridine triphophate nick-end labelling (TUNEL)
and cleaved caspase 3-positive cells. Microglial ionized calcium-bind-
ing adaptor molecule 1 (IBA1) antibody staining for immunohisto-
chemistry and quantitative PCR for inflammatory markers were
performed to reveal the extent of microglial activation.
Materials and methods
Animal experiments and surgicalpreparationAll animal experiments were performed under UK Home Office
Guidelines [Animals (Scientific procedures) Act, 1986]. Seventeen
male piglets aged 524 h, with a mean (SD) weight of 1.71
(0.12) kg (Table 1), were anaesthetized and surgically prepared as
described previously (Lorek et al., 1994). Briefly, piglets were sedated
with intramuscular midazolam (0.2 mg/kg), and arterial O2 saturation
was monitored (Nonin Medical). Isoflurane anaesthesia (4% v/v) was
initially given through a face mask to facilitate tracheostomy and in-
tubation and was maintained (3% during surgery and 2% otherwise).
Piglets were mechanically ventilated; ventilator settings were adjusted
to maintain partial pressure of oxygen (PaO2) and carbon dioxide
(PaCO2) at 8–13 kPa and 4.5–6.5 kPa, respectively, allowing for tem-
perature correction of the arterial blood sample.
An umbilical venous catheter was inserted for infusion of mainten-
ance fluids (10% dextrose, 60 ml/kg/day), fentanyl (3–6 mg/kg/h) and
antibiotics (benzylpenicillin 50 mg/kg and gentamicin 2.5 mg/kg, every
12 h). An umbilical arterial catheter was inserted for continuous moni-
toring of heart rate and arterial blood pressure, and intermittent blood
sampling was used to measure PaO2, PaCO2, pH, electrolytes, glucose
and lactate (Abbot Laboratories). Bolus infusions of colloid (Gelofusin,
B Braun Medical Ltd.) and inotropes maintained mean arterial blood
pressure 440 mm Hg. All animals received continuous physiological
monitoring (SA instruments) and intensive life support throughout
experimentation. Arterial lines were maintained by infusing 0.9%
saline solution (Baxter; 1 ml/h); heparin sodium was added at a con-
centration of 1 IU/ml to prevent line blockage.
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 91
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 3
Both common carotid arteries were surgically isolated at the level
of the fourth cervical vertebra and encircled by remotely controlled
vascular occluders (OC2A, In Vivo Metric). After surgery, piglets
were positioned prone in a plastic pod, and the head immobilized
securely in a stereotactic frame.
Cerebral hypoxia–ischaemiaTwo MRI surface coils were secured to the head, and the animal was
positioned into the bore of a 9.4-T Varian spectrometer for serial 1H
and 31P MRS data acquisition. Whilst in the MRS system, transient
hypoxia–ischaemia was induced by remote occlusion of both
common carotid arteries, using inflatable vascular occluders; the
fractional inspired oxygen was also reduced to 12%. During transient
hypoxia–ischaemia, cerebral energetic changes were observed every
2 min by 31P-MRS, and the b-nucleotide triphosphate (NTP; mainly
ATP) peak height was continuously monitored. Once the b-NTP
peak height had fallen to 40% of baseline, fraction of inspired O2
was titrated to maintain the b-NTP peak height at 40% baseline
value for 12.5 min. At the end of this 12.5-min period, the occluders
were deflated and fraction of inspired O2 was normalized. 31P mag-
netic resonance spectra were acquired for a further 1 h to monitor
recovery from hypoxia–ischaemia.
The time integral of the change in b-NTP peak area relative to
the exchangeable phosphate pool [exchangeable phosphate pool =
inorganic phosphate + phosphocreatine + (2� + b) � NTP] during
Table 1 Physiological variables for the piglets in each group
Measurements Hypothermia (33.5�C)mean (SD)
Hypothermia + melatonin (5 mg/kg/hduring 6 h every 24 h) mean (SD)
P-value
Post-natal age (h) 24.1 (0.4) 24.7 (0.6) 0.06
Body weight (g) 1713 (138) 1722 (135) 0.89
Heart rate (bpm)
Baseline 158 (25) 151 (19) 0.48
End of insult 186 (22) 164 (21) 0.05
2–3.5 h after time zero 121 (13) 125 (18) 0.69
3.5–26 h after time zero 114 (22) 124 (26) 0.38
26–48 h after time zero 145 (24) 146 (23) 0.97
Mean arterial blood pressure (mm Hg)
Baseline 55 (5) 55 (10) 0.99
End of insult 54 (7) 50 (9) 0.38
2–3.5 h after time zero 43 (9) 41 (8) 0.62
3.5–26 h after time zero 47 (3) 44 (8) 0.42
26–48 h after time zero 50 (6) 45 (8) 0.23
Rectal temperature (�C)
Baseline 38.4 (0.6) 38.5 (0.2) 0.61
End of insult 38.5 (0.5) 38.6 (0.3) 0.69
2–3.5 h after time zero 35.3 (0.7) 35.2 (0.7) 0.62
6–26 h after time zero 33.5 (0.1) 33.5 (0.1) 0.84
26–48 h after time zero 37.1 (0.3) 37.1 (0.3) 0.92
PaO2 (kPa)
Baseline 8.8 (2.7) 8.4 (3.4) 0.79
Nadir of the insult 4.1 (2.2) 3.0 (0.6) 0.27
12 h after time zero 10.1 (2.2) 8.0 (1.7) 0.07
24 h after time zero 12.0 (7.2) 10.2 (2.3) 0.54
48 h after time zero
PaCO2 (kPa)
Baseline 5.0 (0.9) 5.2 (1.1) 0.67
Nadir of the insult 3.9 (1.4) 4.6 (0.7) 0.31
12 h after time zero 4.4 (1.5) 4.8 (1.3) 0.60
24 h after time zero 5.6 (1.3) 5.6 (2.4) 0.95
48 h after time zero 5.7 (2.9) 4.8 (1.1) 0.47
Haematocrit (mmol/l)
Baseline 22.9 (5.0) 22.4 (5.1) 0.86
Nadir of the insult 22.4 (6.6) 21.7 (4.6) 0.83
12 h after time zero 23.3 (6.2) 23.4 (4.8) 0.98
24 h after time zero 22.9 (6.6) 24.9 (7.4) 0.56
48 h after time zero 22.3 (9.2) 15.2 (5.8) 0.14
Time zero was the time of resuscitation and the end of the baseline measurements. Mean (SD) values are presented for the two groups: (i) HypoT (n = 8) and(ii) HypoT + Mel (n = 9). Heart rate and blood pressure (mean arterial blood pressure) data are omitted for 30 min after a saline/Geloplasma bolus was administered or aftercardiac arrest.Analysis using a Mann-Whitney test indicated that there was no evidence of a difference between the two groups for any of the outcomes examined at any of the time
points.
92 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 4
hypoxia–ischaemia and the first 60 min of resuscitation gave the mag-
nitude of acute energy depletion, as described previously (Faulkner
et al., 2011).
Pilot studies exploring the effect ofmelatonin doses on mean arterial bloodpressure during therapeutichypothermiaBefore the main randomized study, we explored the effect of three
different doses of melatonin on mean arterial blood pressure in the
piglet after a hypoxic–ischaemic insult. Intravenous melatonin (20, 10
and 5 mg/kg/h over 6 h) was administered 10 min after hypoxia–
ischaemia over 6 h, with cooling started at 2 h after hypoxia–ischaemia
(as described later in the text).
Experimental groupsBaseline data were acquired before transient hypoxia–ischaemia but
after stabilization of the animal in the MRS system. After resuscitation,
piglets were randomized into two groups with intervention from 2 to
26 h: (i) hypothermia (33.5�C) n = 8 (HypoT) and (ii) hypothermia
(33.5�C) + melatonin infusion of 5 mg/kg over 6 h starting 10 min
and 24 h after hypoxia–ischaemia, n = 9 (HypoT + Mel). Cooling was
achieved using a water mattress; piglets were cooled for 90 min to
reach the target rectal temperature of 33.5�C, which was maintained
for between 2 and 26 h after hypoxia–ischaemia.
Melatonin (Sigma) was dissolved in ethanol (2.5% final v/v) and
saline in the dark shortly before administration to HypoT + Mel
animals. The HypoT animals received vehicle ethanol (2.5% final
v/v) and saline during the same period. At 26 h after hypoxia-ischae-
mia, piglets were rewarmed to normothermia at 0.5�C/h. Blood was
sampled at 6-h intervals after the hypoxic–ischaemic insult.
Magnetic resonance spectroscopyMagnetic resonance spectra were acquired as described previously
(Faulkner et al., 2011), using a combination of a 6.5 � 5.5-cm ellip-
tical receive surface coil tuneable to 1H and a 150-mm (Rapid Biospec)
transmit volume coil with a separate elliptical transmit–receive coil
tuned to 31P resonance frequency. Whole brain 31P magnetic reso-
nance spectra were acquired using single-pulse acquisition with a repe-
tition time of 10 s. Baseline spectra were acquired before, during and
for the first 60 min of recovery after hypoxia–ischaemia (31P only), and
thereafter at 24 and 48 h (Fig. 1). Point-resolved (PRESS) hypoxia–
ischaemia MRS was acquired from two positions: deep grey matter
centred on both lateral thalami and hypothalami (14 � 14 � 7-mm
voxel) (Fig. 1A-F); and dorsal right subcortical white matter at the
centrum semiovale level (white matter; 9 � 4 � 20-mm voxel)
(Supplementary Fig. 2A-F, H, J). 1H magnetic resonance spectra
were acquired with a repetition time of 5 s, 128 averages and echo
time of 25, 144 and 288 ms.
Magnetic resonance spectroscopyanalysis1H magnetic resonance spectra were analysed using the LC Model
(Provencher, 1993). For each spectrum, the relative signal amplitudes
of choline, total creatine, N-acetyl aspartate and lactate were mea-
sured. The primary 1H MRS outcome measures nominated a priori
were lactate/N-acetyl aspartate and lactate/creatine. Metabolite
peak/area under the curve ratios were plotted from baseline to 48 h
after hypoxia–ischaemia for each voxel in each subject, on a logarith-
mic scale and analysed according to the randomized group.31P magnetic resonance spectra were analysed by AMARES
(advanced method for accurate, robust and efficient spectral fitting
with the use of prior knowledge) (Vanhamme et al., 1997). Signal
amplitudes and chemical shifts were measured for phosphocreatine,
inorganic phosphate (three independent components fitted) and �-,
b- and �-NTP; the equivalent signal amplitude for exchangeable phos-
phate pool was also calculated. Metabolite peak-area under the curve
ratios were plotted from baseline to 48 h after hypoxia–ischaemia for
each voxel in each subject on a logarithmic scale and analysed accord-
ing to the randomized group.
Electroencephalography and amplitude-integrated electroencephalographyAfter surgical preparation, multichannel EEG (Nicolet) monitoring
(six-lead) was acquired at baseline and during the period between
the MRS data acquisitions i.e. between 2–20, 25–40 and at 48 h
after hypoxia–ischaemia. Filtered amplitude-integrated EEG recordings
were classified according to the pattern classification (de Vries and
Hellstrom-Westas, 2005). A score of 0 was flat trace; 1, continuous
low voltage; 2, burst suppression; 3, discontinuous normal voltage
and 4, continuous normal voltage, at baseline, 3, 6, 12 and then
every 12 h after hypoxia-ischaemia, thereafter noting seizure activity
and duration. Classification was omitted within 6 h of phenobarbitone
treatment. The area under the curve of the amplitude-integrated EEG
category was used for statistical comparison using a 2-tailed t-test.
To further compare groups, a discrete threshold of 0.35 was used in
a �2 test.
Sample sizeThe primary outcome, the change in area under the curve for lactate/
N-acetyl aspartate from baseline to 48 h, was used to calculate the
sample size. Previous work with our model suggested that the change
in lactate/N-acetyl aspartate during 48 h varied between normo- and
hypothermic groups by 1.0 U, with a standard deviation of 0.75 U
(both log scale). Assuming similar magnitude of additional effect for
melatonin-augmented hypothermia (versus hypothermia alone) and
with 5% significance and 80% power, nine subjects would be required
in each group.
Brain histologyAt 48 h after hypoxia–ischaemia, piglets were euthanized with
pentobarbital; the brain was fixed through cardiac perfusion with cold
4% paraformaldehyde in PBS, dissected out and post-fixed at 4�C in
2% paraformaldehyde for 7 days. Coronal slices (5 mm thick) of the
right hemisphere, starting from anterior to the optic chiasma, were
embedded in paraffin wax and sectioned to 5-mm thickness and stained
for haematoxylin and eosin. To assess cell death and glial activation,
additional brain sections were stained for nuclear DNA fragmentation
using histochemistry with TUNEL, and the appearance of activated
caspase 3 and microglial ionized calcium-binding adaptor molecule 1
(IBA1) immunoreactivity (Ito et al., 2001). Two sections per animal,
placed 5 mm apart, were examined with each stain.
For all histochemical and immunohistochemical stains, brain sections
were dehydrated in xylene (3 � 10 min) and rehydrated in graded
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 93
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 5
Figure 1 1H and 31P MRS shows that melatonin reduces metabolic changes after a hypoxic–ischaemic insult in the neonatal piglet.
(A–F) Time course changes for the lactate/creatine (A and D), N-acetyl aspartate/creatine (B and E) and lactate/N-acetyl aspartate
(C and F) ratios in the thalamic voxel, in animals treated with hypothermia alone (HypoT, A–C), and those cotreated with melatonin
(HypoT + Mel, D–F) in the 48 h after a hypoxic–ischaemic insult (0 h is end of insult). The data points are plotted on a semi-logarithmic
scale, and each trace of data points represents an individual animal. Note in the HypoT group the increase of lactate/creatine (A) and
lactate/N-acetyl aspartate (C) ratios in four of eight animals, and a decrease in N-acetyl aspartate in three of eight animals (B). These
changes were abrogated in the HypoT + Mel group. (G–L) 31P-MRS time course for inorganic phosphate/exchangeable phosphate pool
(G and J), phosphocreatine/exchangeable phosphate pool (H and K) and NTP/exchangeable phosphate pool (I and L) in the whole
forebrain voxel in HypoT (G–I), and HypoT + Mel piglets (J–L). In 31P-MRS, the HypoT group shows three of six animals had increasing
inorganic phosphate/exchangeable phosphate pool (G) and reductions in phosphocreatine/exchangeable phosphate pool (H) and NTP/
exchangeable phosphate pool (I). These changes were abrogated in the HypoT + Mel group. (M–O) Summary of the area under the curve
data (mean � SEM) for the thalamic 1H (M), the white matter 1H (N) and the 31P whole forebrain (O) metabolite ratio time courses. In the
case of decreasing ratios (N-acetyl aspartate/creatine, phosphocreatine/exchangeable phosphate pool and NTP/exchangeable phosphate
pool), the bars represent the area over the curve algorithm. Red bars show data for the HypoT group, blue bars for the HypoT and Mel
group, mean � SEM. *P50.05 in t-test. Cr = creatine; ePP = exchangeable phosphate pool; HI = hypoxia–ischaemia; Lac = lactate;
NAA = N-acetyl aspartate; PCr = phosphocreatine; Pi = inorganic phosphate; TH = thalamic; WM = white matter.
94 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 6
ethanol solutions (100–70%), followed by double-distilled water. For
TUNEL, the sections were pretreated for 15 min in 3% H2O2 in metha-
nol to remove endogenous peroxidase, followed by a 15-min pepti-
dase predigestion with 20mg/ml proteinase K (Promega) at 65�C, and
then incubated at 37�C for 2 h with the TUNEL solution (Roche) con-
taining biotinylated dUTP. For immunohistochemistry, the sections
were processed for antigen retrieval (800-mW microwave irradiation
in 0.1 M citrate buffer, 10 min), followed by overnight incubation with
primary rabbit antibody against activated caspase 3 (1:500) (Abcam)
or IBA1 (1:1000) (Wako), and then 2-h incubation with a biotinylated
secondary goat anti-rabbit immunoglobulin antibody (1:100, Jackson
Laboratory). The biotin residues were detected with the avidin-bioti-
nylated horseradish peroxidase complex (ABC, Vector Laboratories)
and visualized with diaminobenzidine/H2O2 (Sigma), with CoCl2 and
NiCl2 included to intensify TUNEL histochemistry. The sections were
counterstained with haematoxylin, dehydrated in graded alcohol and
xylene and mounted with Depex (VWR), or alternatively, mounted
with Vectashield + 4’,6-diamidino-2-phenylindole (DAPI) aqueous
mounting media (Vector Labs), to facilitate total cell number counts
during analysis of Iba1 and activated caspase 3.
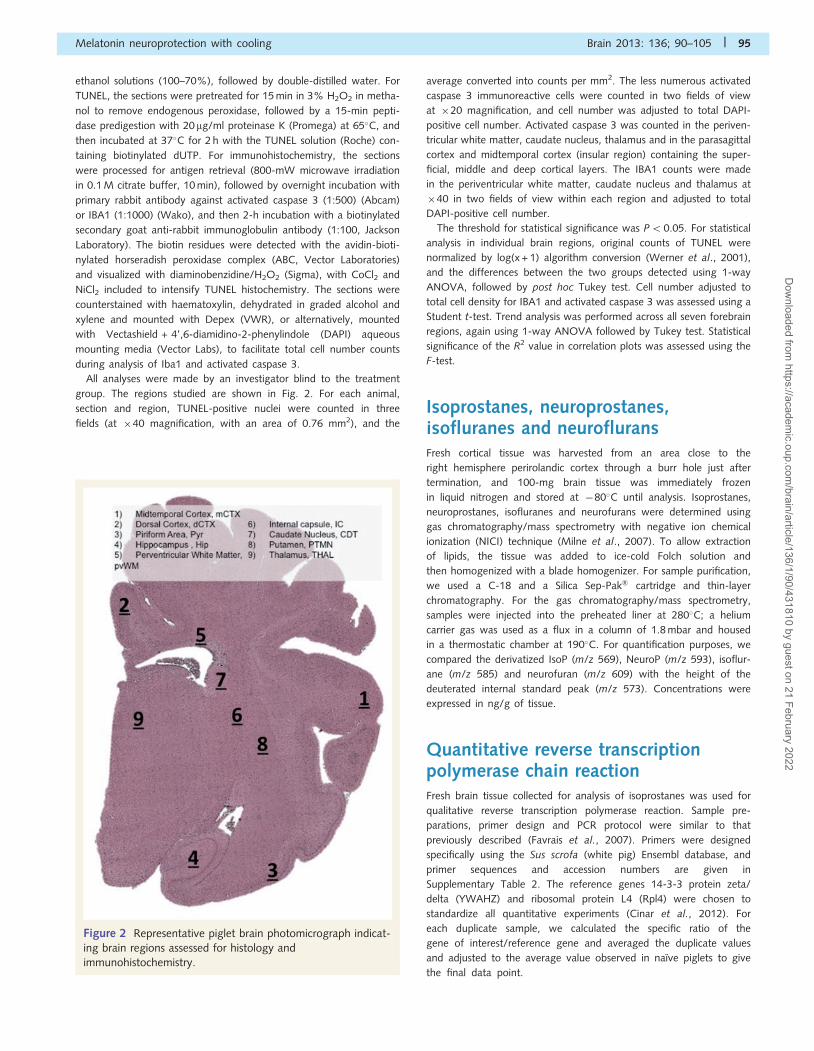
All analyses were made by an investigator blind to the treatment
group. The regions studied are shown in Fig. 2. For each animal,
section and region, TUNEL-positive nuclei were counted in three
fields (at �40 magnification, with an area of 0.76 mm2), and the
average converted into counts per mm2. The less numerous activated
caspase 3 immunoreactive cells were counted in two fields of view
at �20 magnification, and cell number was adjusted to total DAPI-
positive cell number. Activated caspase 3 was counted in the periven-
tricular white matter, caudate nucleus, thalamus and in the parasagittal
cortex and midtemporal cortex (insular region) containing the super-
ficial, middle and deep cortical layers. The IBA1 counts were made
in the periventricular white matter, caudate nucleus and thalamus at
�40 in two fields of view within each region and adjusted to total
DAPI-positive cell number.
The threshold for statistical significance was P5 0.05. For statistical
analysis in individual brain regions, original counts of TUNEL were
normalized by log(x + 1) algorithm conversion (Werner et al., 2001),
and the differences between the two groups detected using 1-way
ANOVA, followed by post hoc Tukey test. Cell number adjusted to
total cell density for IBA1 and activated caspase 3 was assessed using a
Student t-test. Trend analysis was performed across all seven forebrain
regions, again using 1-way ANOVA followed by Tukey test. Statistical
significance of the R2 value in correlation plots was assessed using the
F-test.
Isoprostanes, neuroprostanes,isofluranes and neurofluransFresh cortical tissue was harvested from an area close to the
right hemisphere perirolandic cortex through a burr hole just after
termination, and 100-mg brain tissue was immediately frozen
in liquid nitrogen and stored at �80�C until analysis. Isoprostanes,
neuroprostanes, isofluranes and neurofurans were determined using
gas chromatography/mass spectrometry with negative ion chemical
ionization (NICI) technique (Milne et al., 2007). To allow extraction
of lipids, the tissue was added to ice-cold Folch solution and
then homogenized with a blade homogenizer. For sample purification,
we used a C-18 and a Silica Sep-Pak� cartridge and thin-layer
chromatography. For the gas chromatography/mass spectrometry,
samples were injected into the preheated liner at 280�C; a helium
carrier gas was used as a flux in a column of 1.8 mbar and housed
in a thermostatic chamber at 190�C. For quantification purposes, we
compared the derivatized IsoP (m/z 569), NeuroP (m/z 593), isoflur-
ane (m/z 585) and neurofuran (m/z 609) with the height of the
deuterated internal standard peak (m/z 573). Concentrations were
expressed in ng/g of tissue.
Quantitative reverse transcriptionpolymerase chain reactionFresh brain tissue collected for analysis of isoprostanes was used for
qualitative reverse transcription polymerase reaction. Sample pre-
parations, primer design and PCR protocol were similar to that
previously described (Favrais et al., 2007). Primers were designed
specifically using the Sus scrofa (white pig) Ensembl database, and
primer sequences and accession numbers are given in
Supplementary Table 2. The reference genes 14-3-3 protein zeta/
delta (YWAHZ) and ribosomal protein L4 (Rpl4) were chosen to
standardize all quantitative experiments (Cinar et al., 2012). For
each duplicate sample, we calculated the specific ratio of the
gene of interest/reference gene and averaged the duplicate values
and adjusted to the average value observed in naıve piglets to give
the final data point.
Figure 2 Representative piglet brain photomicrograph indicat-
ing brain regions assessed for histology and
immunohistochemistry.
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 95
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 7
Plasma melatonin assayPlasma melatonin concentrations were determined by radioimmunoas-
say (Claustrat et al., 1984). Melatonin was isolated from plasma using
diethyl-ether extraction. Because the standard curve ranged from 3 to
2400 ng/l, the highest melatonin concentrations required dilution in a
phosphate albumin buffer solution. The intra-assay coefficients of var-
iation were 57% between 30 and 200 pg/ml, and the inter-assay
coefficients of variation were 8.7 and 7.9% (n = 18) for melatonin
concentrations of 55 and 115 pg/ml, respectively.
Results
Pilot study of melatonin dosescombined with hypothermiaA dose of 5 mg/kg/h over 6 h was safe in a pilot study of three
doses of melatonin combined with hypothermia. As the clinical
use of therapeutic hypothermia in babies is associated with
increased use of inotropes (Battin et al., 2009) and can potentially
lead to hypotension in some animals (Kerenyi et al., 2012) we
wanted to assess any interaction between the combination of
hypothermia with melatonin. A melatonin dose of 10 mg/kg/h
or higher was associated with hypotension and a requirement
for saline boli and inotropic support to maintain the mean arterial
blood pressure normal; 5 mg/kg/h melatonin was tolerated well
in our model (see Supplementary material and Supplementary
Fig. 1).
Physiological measuresThere were no intergroup differences in body weight, post-natal
age, baseline physiological (heart rate and mean arterial blood
pressure) and biochemical measures (blood lactate, base excess
and glucose) (Table 1 and Supplementary Table 1). Insult severity
qualified with triphosphate levels and PaO2 and PaCO2 were simi-
lar between groups during hypoxia–ischaemia, and the time taken
to reach target rectal temperature after hypoxia–ischaemia
hypothermic induction was similar between the groups. Heart
rate at the end of the insult was lower in the HypoT + Mel
group than in the HypoT group (P = 0.05).
During the cooling induction (2–3.5 h), maintenance (3.5–26 h)
and rewarming/normothermia (26–48 h) periods, mean heart rate,
mean arterial blood pressure, blood chemistry and electrolytes
(pH, lactate, base excess and glucose) were similar between
the groups (Table 1). Median total (saline and Geloplasma)
volume (ml/kg) and dopamine use (mg/kg/min) were similar
between groups. Inotropic support was required in both groups
(dobutamine: 1/8 versus 2/9; adrenaline: 1/8 versus 3/9 9
for HypoT and HypoT + Mel groups, respectively); however,
there were no significant differences between the groups
(Table 2).
One piglet in the melatonin group suffered a cardiac arrest due
to mechanical obstruction of the tracheal tube at 46 h but was
resuscitated successfully; no piglets died prematurely.
1H and 31P magnetic resonancespectroscopyTo standardize for the MRS geometry and detection sensitivity,
metabolite peak/area under the curve ratios were calculated for
lactate/creatine, N-acetyl aspartate/creatine, choline/creatine and
lactate/N-acetyl aspartate for 1H-MRS in the dorsal subcortical
white matter and deep grey matter voxels, and the ratios for
inorganic phosphate/exchangeable phosphate pool, phosphocrea-
tine/exchangeable phosphate pool and NTP/exchangeable phos-
phate pool for the 31P-MRS in the whole brain. The data were
plotted on a semi-logarithmic scale. Data from all 17 piglets (n = 8
HypoT, n = 9 HypoT + Mel) were available for analysis of 1H-MRS
in the thalamic voxel (Fig. 1A–F) and white matter voxel
(Supplementary Fig. 2A–F, H, J); data from 12 piglets (n = 6
HypoT, n = 6 HypoT + Mel) were available for 31P-MRS
(Fig. 1G–L).
In four of the eight piglets (piglets 148, 153, 161 and 162) that
received hypothermia alone after hypoxia–ischaemia, there was a
marked increase in lactate/creatine and lactate/N-acetyl aspartate
in the thalamic voxel—these metabolite ratios increased to 41
over the following 48 h (Fig. 1A and C). In the other four piglets,
there was little change, with ratios �0.1–0.2. Similar differences
were also observed for N-acetyl aspartate/creatine (Fig. 1B), with
three of the animals (piglets 153, 161 and 162) dropping to a ratio
of 50.8, and the other five staying in a narrow range of �1.5.
The choline/creatine ratios were generally unchanged after
hypoxia–ischaemia insult (Supplementary Fig. 2G and H).
Strikingly, in animals additionally treated with melatonin, none
showed an increase in lactate/creatine and lactate/N-acetyl
Table 2 Average total fluid replacement and inotrope infusion for the piglets in each group during 48 h after insult
Measurement Hypothermia (33.5�C)median (IQR)
Hypothermia + melatonin (5 mg/kg/hduring 6 h every 24 h) median (IQR)
P-value
Saline bolus ml/kg 0 (0, 17) 0 (0, 16) 0.65
Geloplasma bolus ml/kg 0 (0, 28) 30 (0, 34) 0.20
Total ml/kg 17 (0, 37) 30 (17, 51) 0.26
Dopamine (mg/kg/min) 6.9 (2.5, 9.6) 5.3 (4.9, 6.0) 0.74
Dobutamine (mg/kg/min) 0 (0, 0) 0 (0, 0) 0.52
Adrenaline (ng/kg/min) 0 (0, 0) 0 (0, 0) 0.52
Median (IQR) values for saline and Geloplasma boluses and inotrope infusion are presented for the two groups. Analysis using a Mann-Whitney test revealed no evidence of
a statistically significant difference between groups for fluid replacement or inotrope usage.
96 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 8
aspartate or a decrease in N-acetyl aspartate/creatine. All nine
animals showed lactate/creatine and lactate/N-acetyl aspartate
ratios of 51 (Fig. 1D and F, P50.05 in �2 test comparison
with the HypoT group), and N-acetyl aspartate/creatine ratios
40.8 (generally �1.5) (Fig. 1E, P = 0.055 in �2 test). The cho-
line/creatine ratios were unchanged (Supplementary Fig. 2I and J).
A similar divergence in the appearance of animals with lactate
overshoot or drop in N-acetyl aspartate was also present in the1H-white matter voxel (Supplementary Fig. 2A–F) and 31P-whole
brain data (Fig. 1G–L). In the HypoT group, four of eight animals
(piglets 148, 153, 161 and 162) showed an increase of 41 for
lactate/creatine (Supplementary Fig. 2A) and lactate/N-acetyl
aspartate (Supplementary Fig. 2C) and a drop 50.8 for N-acetyl
aspartate/creatine (Supplementary Fig. 2B). In the HypoT + Mel
group, none of the nine had an increase in lactate/creatine
(Supplementary Fig. 2D), and one of nine had an increase in
lactate/N-acetyl aspartate (Supplementary Fig. 2F) and a drop in
N-acetyl aspartate/creatine (Supplementary Fig. 2E). The differ-
ence between the numbers for lactate/creatine and lactate/N-
acetyl aspartate in the HypoT + Mel group was because of piglet
155, which showed a subthreshold increase in lactate/creatine but
also a pronounced additional drop in N-acetyl aspartate/creatine
levels in its white matter voxel.
In the case of 31P-MRS, three of six HypoT animals showed
an increase of inorganic phosphate/exchangeable phosphate pool
475% (Fig. 1G), and a drop of phosphocreatine and NTP 515%
(Fig. 1H and I, respectively), compared with none of eight in the
HypoT + Mel group (Fig. 1J–L, P = 0.024 in �2 test). Piglet 155
again showed an increase in inorganic phosphate/exchangeable
phosphate pool and a drop in phosphocreatine/exchangeable phos-
phate pool compared with its peer group, but these changes did
not reach the thresholds described earlier in the text.
As a next step, the overall changes in biomarker outcome over
48 h were assessed using the area under the curve algorithm for
increasing (lactate/creatine, lactate/N-acetyl aspartate, inorganic
phosphate/exchangeable phosphate pool) and decreasing
(N-acetyl aspartate/creatine, phosphocreatine/exchangeable phos-
phate pool, inorganic phosphate/exchangeable phosphate pool)
biomarker ratios. In both cases, these outcome parameters
increased with more pronounced brain metabolite change and
showed a strong positive correlation with histological evidence
of brain injury (Faulkner et al., 2011). A summary of all outcome
area under the curve and area over the curve parameters is shown
for 1H-MRS in the thalamic (Fig. 1M) and white matter (Fig. 1N)
voxels and for 31P-MRS in the whole forebrain voxel (Fig. 1O).
Compared with the HypoT group, combined treatment with
HypoT + Mel was associated with a significant overall decrease in
thalamic lactate/creatine and lactate/N-acetyl aspartate area
under the curve (P50.05, t-test). All other hypoxia–ischaemia-
affected outcome parameters (inorganic phosphate/exchangeable
phosphate pool area under the curve, N-acetyl aspartate/
creatine area over the curve, phosphocreatine/exchangeable
phosphate pool area over the curve, NTP/exchangeable
phosphate pool area over the curve) showed smaller values in
the HypoT + Mel group, but these did not reach statistical
significance.
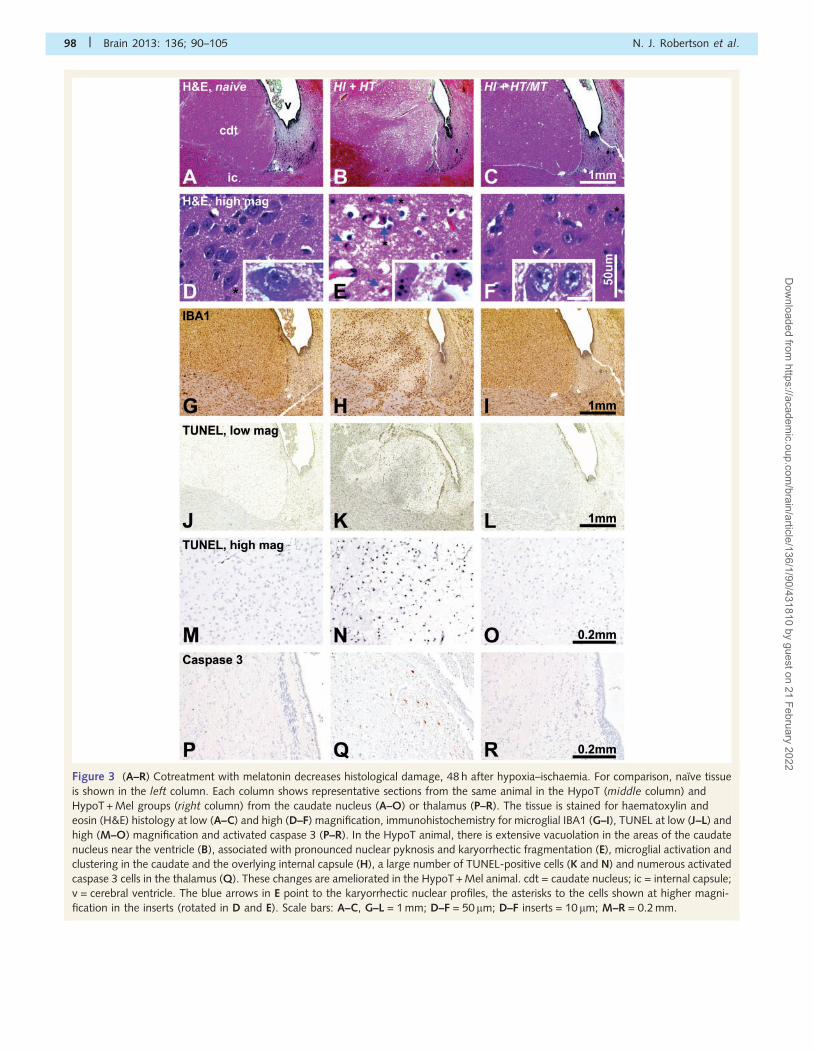
HistologyThe brain regions assessed are shown in Fig. 2. Representative
haematoxylin and eosin histology, microglial IBA1 immunohisto-
chemistry, TUNEL at low and high magnification and activated cas-
pase-3 are shown in Fig 3. In routine haematoxylin and eosin stains,
compared with the naıve animals (Fig. 3A and D), the HypoT group
revealed neural injury that was particularly pronounced in the
caudatoputamen and thalamus, with extensive vacuolation of the
neuropil (Fig. 3B) and, at high magnification, frequent pyknotic
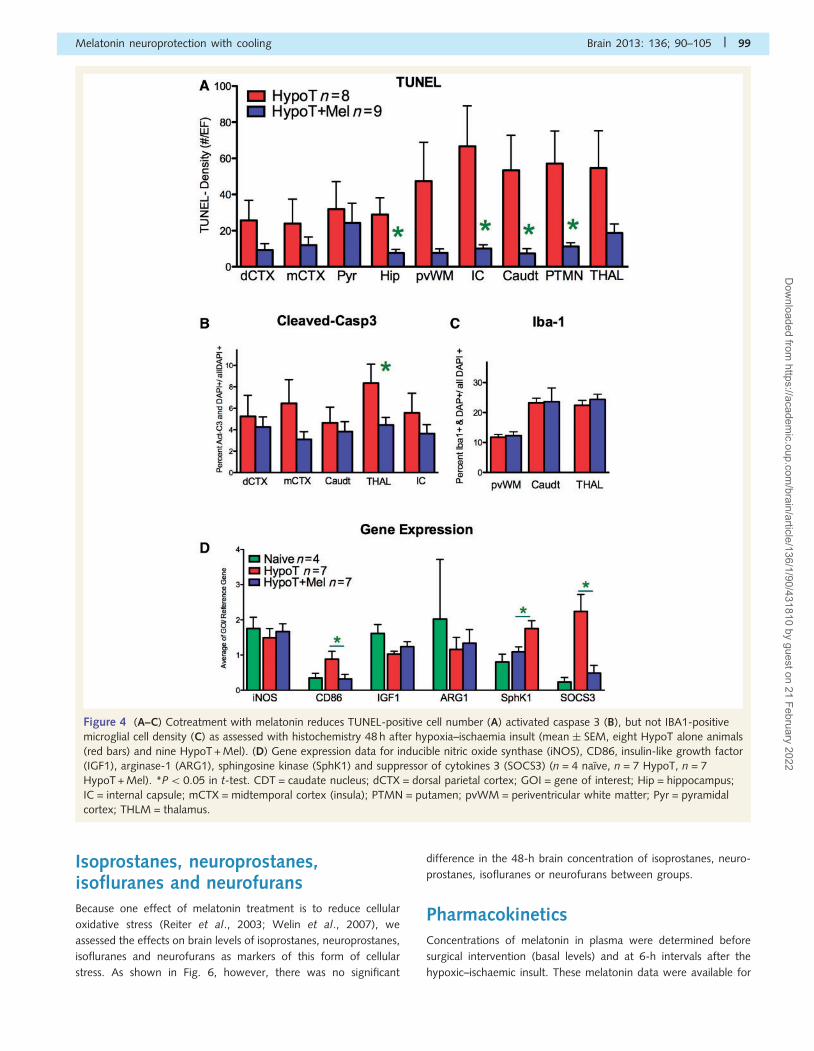
and karyorrhectic nuclear profiles (Fig. 3E). Quantification of the
TUNEL-positive cells showed significant decreases in the HypoT +
Mel group compared with the HypoT group for four forebrain
regions—periventricular white matter, internal capsule, caudate
nucleus and putamen (P50.05) (Fig. 4A). Quantification of the
number of activated caspase 3-positive cells per total (DAPI-
positive) cell number revealed a reduced density of cells expressing
this cell death marker in the thalamus (Fig. 4B). Immunostaining for
IBA1 in brain microglia revealed a trend towards more microglial
clustering (e.g. in the caudate nucleus in Fig. 3H), but there was no
significant difference in the overall density of the IBA1-positive cell
counts (Fig. 4C).
Gene expression analysisIn addition to direct counts of microglia (IBA1-positive), inflammatory
mediators were assessed using quantitative PCR in seven Hypo and
seven Hypo + Mel animals; expression of inducible nitric oxide
synthase, CD86, insulin-like growth factor (IGF1), arginase 1
(ARG1), sphingosine kinase (SphK1) and suppressor of cytokines 3
(SOCS3) is shown in Fig. 4D. There was an increase in SphK1 gene
expression in the HypoT + Mel group compared with the HypoT-
only group (P5 0.05). Expression of CD86 was decreased
(P5 0.05), as was expression of SOCS3 (P50.05), but expression
of IGF1, ARG1 or inducible nitric oxide synthase was not altered.
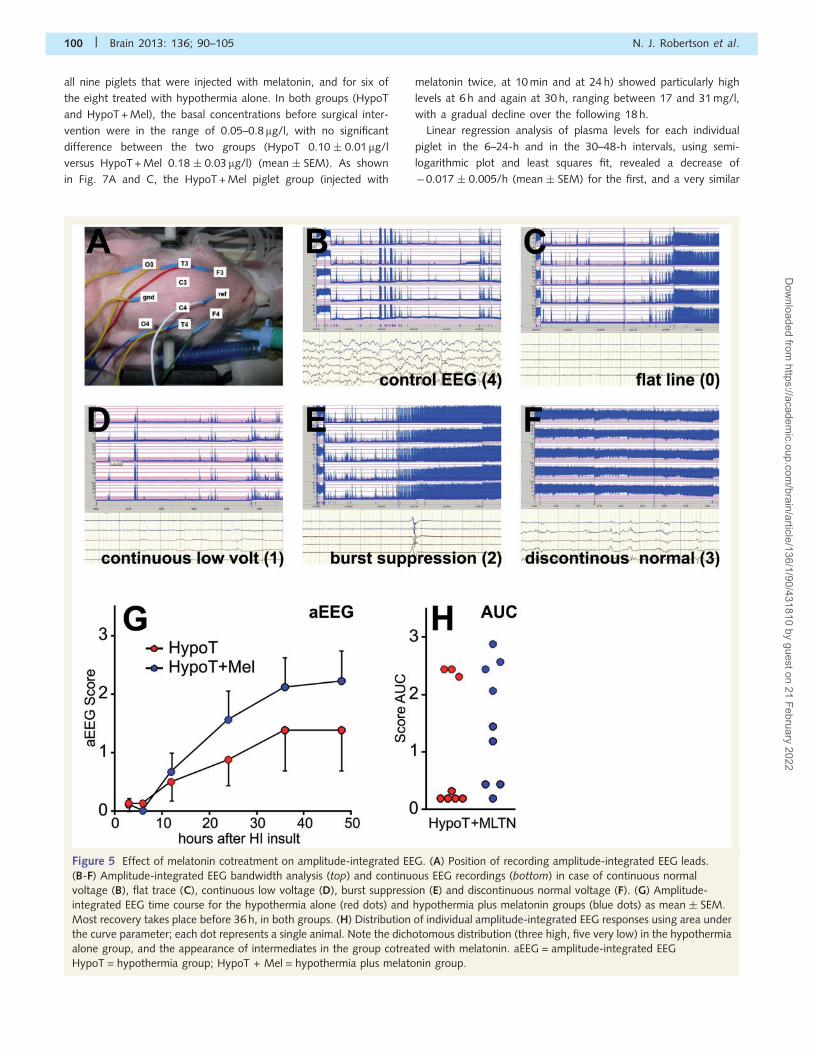
ElectroencephalographyAt baseline, the raw EEG trace was active with an amplitude-
integrated EEG bandwidth of 10–50mV. The placement of EEG
leads is shown in Fig. 5A. Typically, 3 h after hypoxia–ischaemia,
cortical activity was suppressed (55mV) in all but a few animals
(compare Fig. 5C at 3 h with Fig. 5B before insult). These were
the same animals as those with adverse MRS bioenergetics (piglets
148, 153, 161 and 162). A score of 0 was given for a flat trace,
1 for continuous low voltage, 2 for burst suppression, 3 for dis-
continuous normal voltage and 4 for continuous normal voltage
(Fig. 5B–E). As shown in Fig. 5G, most of the recovery took place
before 36 h, with half maximal return occurring at �20 h. The
HypoT + Mel subgroup appeared to recover amplitude-integrated
EEG somewhat more rapidly than the cooling-only group, but this
did not reach statistical significance for any of the individual time
points. Further analysis using the area under the curve parameter
(Fig. 5H) for the HypoT group showed a dichotomy, with three
animals revealing ‘good’ recovery (i.e. in the range of 2–3) (piglets
149, 150 and 160) and five with very poor recovery of 50.35
(piglets 145, 148, 153, 161 and 162). The HypoT + Mel subgroup
demonstrated a more even distribution (Fig. 5H).
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 97
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 9
Figure 3 (A–R) Cotreatment with melatonin decreases histological damage, 48 h after hypoxia–ischaemia. For comparison, naıve tissue
is shown in the left column. Each column shows representative sections from the same animal in the HypoT (middle column) and
HypoT + Mel groups (right column) from the caudate nucleus (A–O) or thalamus (P–R). The tissue is stained for haematoxylin and
eosin (H&E) histology at low (A–C) and high (D–F) magnification, immunohistochemistry for microglial IBA1 (G–I), TUNEL at low (J–L) and
high (M–O) magnification and activated caspase 3 (P–R). In the HypoT animal, there is extensive vacuolation in the areas of the caudate
nucleus near the ventricle (B), associated with pronounced nuclear pyknosis and karyorrhectic fragmentation (E), microglial activation and
clustering in the caudate and the overlying internal capsule (H), a large number of TUNEL-positive cells (K and N) and numerous activated
caspase 3 cells in the thalamus (Q). These changes are ameliorated in the HypoT + Mel animal. cdt = caudate nucleus; ic = internal capsule;
v = cerebral ventricle. The blue arrows in E point to the karyorrhectic nuclear profiles, the asterisks to the cells shown at higher magni-
fication in the inserts (rotated in D and E). Scale bars: A–C, G–L = 1 mm; D–F = 50 mm; D–F inserts = 10 mm; M–R = 0.2 mm.
98 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 10
Isoprostanes, neuroprostanes,isofluranes and neurofuransBecause one effect of melatonin treatment is to reduce cellular
oxidative stress (Reiter et al., 2003; Welin et al., 2007), we
assessed the effects on brain levels of isoprostanes, neuroprostanes,
isofluranes and neurofurans as markers of this form of cellular
stress. As shown in Fig. 6, however, there was no significant
difference in the 48-h brain concentration of isoprostanes, neuro-
prostanes, isofluranes or neurofurans between groups.
PharmacokineticsConcentrations of melatonin in plasma were determined before
surgical intervention (basal levels) and at 6-h intervals after the
hypoxic–ischaemic insult. These melatonin data were available for
Figure 4 (A–C) Cotreatment with melatonin reduces TUNEL-positive cell number (A) activated caspase 3 (B), but not IBA1-positive
microglial cell density (C) as assessed with histochemistry 48 h after hypoxia–ischaemia insult (mean � SEM, eight HypoT alone animals
(red bars) and nine HypoT + Mel). (D) Gene expression data for inducible nitric oxide synthase (iNOS), CD86, insulin-like growth factor
(IGF1), arginase-1 (ARG1), sphingosine kinase (SphK1) and suppressor of cytokines 3 (SOCS3) (n = 4 naıve, n = 7 HypoT, n = 7
HypoT + Mel). *P5 0.05 in t-test. CDT = caudate nucleus; dCTX = dorsal parietal cortex; GOI = gene of interest; Hip = hippocampus;
IC = internal capsule; mCTX = midtemporal cortex (insula); PTMN = putamen; pvWM = periventricular white matter; Pyr = pyramidal
cortex; THLM = thalamus.
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 99
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 11
all nine piglets that were injected with melatonin, and for six of
the eight treated with hypothermia alone. In both groups (HypoT
and HypoT + Mel), the basal concentrations before surgical inter-
vention were in the range of 0.05–0.8mg/l, with no significant
difference between the two groups (HypoT 0.10 � 0.01mg/l
versus HypoT + Mel 0.18 � 0.03 mg/l) (mean � SEM). As shown
in Fig. 7A and C, the HypoT + Mel piglet group (injected with
melatonin twice, at 10 min and at 24 h) showed particularly high
levels at 6 h and again at 30 h, ranging between 17 and 31 mg/l,
with a gradual decline over the following 18 h.
Linear regression analysis of plasma levels for each individual
piglet in the 6–24-h and in the 30–48-h intervals, using semi-
logarithmic plot and least squares fit, revealed a decrease of
�0.017 � 0.005/h (mean � SEM) for the first, and a very similar
Figure 5 Effect of melatonin cotreatment on amplitude-integrated EEG. (A) Position of recording amplitude-integrated EEG leads.
(B-F) Amplitude-integrated EEG bandwidth analysis (top) and continuous EEG recordings (bottom) in case of continuous normal
voltage (B), flat trace (C), continuous low voltage (D), burst suppression (E) and discontinuous normal voltage (F). (G) Amplitude-
integrated EEG time course for the hypothermia alone (red dots) and hypothermia plus melatonin groups (blue dots) as mean � SEM.
Most recovery takes place before 36 h, in both groups. (H) Distribution of individual amplitude-integrated EEG responses using area under
the curve parameter; each dot represents a single animal. Note the dichotomous distribution (three high, five very low) in the hypothermia
alone group, and the appearance of intermediates in the group cotreated with melatonin. aEEG = amplitude-integrated EEG
HypoT = hypothermia group; HypoT + Mel = hypothermia plus melatonin group.
100 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 12
value of �0.018 � 0.003/h for the second, later interval.
The average decrease (first and second interval) was �0.017 �
0.004/h, corresponding to an average half-life for plasma melato-
nin of 21.6 � 3.3 h.
In neonatal piglets treated with hypothermia alone (Fig. 7B),
plasma melatonin values at every time point 6–48 h after hypoxia–
ischaemia were on average 6–15 times higher than the basal levels,
before surgical intervention (P5 0.01, paired t-test).
DiscussionThis study in newborn piglets demonstrates that 5 mg/kg/h mela-
tonin administered intravenously 10 min after the end of transient
hypoxia–ischaemia over 6 h and repeated at 24 h augments
hypothermic neuroprotection based on improved cerebral energy
metabolism using 1H MRS biomarkers (deep grey matter lactate/
N-acetyl aspartate and lactate/creatine). Melatonin-augmented
hypothermia was also associated with increased levels of cerebral
ATP (whole brain 31P MRS NTP/exchangeable phosphate pool).
Correlating with improved cerebral energy metabolism, TUNEL-
positive nuclei were reduced in the melatonin-augmented hypo-
thermia group compared with hypothermia alone in the thalamus,
white matter, internal capsule, putamen and caudate, and there was
reduced activated caspase 3 in the thalamus. The addition of 5 mg/
kg/h over 6 h of intravenous melatonin to hypothermia at 10 min
and 24 h after hypoxia–ischaemia did not alter the heart rate or
mean arterial blood pressure, need for inotropes, blood electrolytes,
or glucose and lactate levels during or after treatment.
We monitored brain metabolism using MRS biomarkers that are
known to change after hypoxia–ischaemia in the piglet (Lorek
et al., 1994; Penrice et al., 1997), serve as linking biomarkers in
infants with neonatal encephalopathy (Robertson et al., 1999;
Cheong et al., 2006), and are currently used as a surrogate out-
come measure in clinical neuroprotection trials for neonatal ence-
phalopathy (Azzopardi, 2011). Specifically, high levels of lactate/
N-acetyl aspartate on thalamic MRS in neonates at 48 h are pre-
dictive of poorer 12–18-month neurodevelopmental outcome
(Robertson et al., 1999; Cheong et al., 2006; Thayyil et al.,
2010). As shown in Fig. 1, the addition of melatonin to hypother-
mia abrogated the MRS lactate/N-acetyl aspartate and lactate/
creatine increases in the forebrain/thalamic voxel. This increase
in MRS lactate/N-acetyl aspartate occurred in only 50% of
HypoT-treated piglets; this is not surprising as therapeutic
hypothermia itself compared with normothermia preserves
energy metabolism after transient hypoxia-ischaemia in piglets
(Penrice et al., 1997). We also observed a higher ATP level in
the melatonin-augmented cooling group compared with the cool-
ing-only group using 31P MRS. Higher ATP on 31P MRS in infants
with hypoxic–ischaemic brain injury is associated with better long-
term outcome in clinical studies (Azzopardi et al., 1989). We also
used continuous EEG monitoring as another measure of cell bioe-
nergetics and as this is also a predictor of outcome after hypoxia–
ischaemia (Toet et al., 1999; Thoresen et al., 2010). We observed
that melatonin decreased the number of piglets with a poor recov-
ery after hypoxia–ischaemia (area under the curve 50.35), and
increased those with an intermediate outcome (area under the
Figure 7 Melatonin pharmacokinetics. (A–C) Concentrations in blood plasma at the basal level (bsl) and 6–48 h after hypoxic–ischaemic
insult. (A) Mean � SEM levels for the HypoT alone (red circles, n = 6 animals) and HypoT + Mel (blue circles, n = 9 animals). (B and C)
Time course data for individual HypoT (D) and HypoT + Mel (E) animals. Green arrows in A and C indicate intravenous melatonin injection.
In B, the interruptions in individual traces were due to missing time points. Note that in B the y-axis is in mg/l and in C the y-axis is in mg/l.
bsl = baseline; HI = hypoxia–ischaemia.
Figure 6 (A) Effect of melatonin treatment on isoprostane (IP),
neuroprostane (NP), neurofurans (NF) and isoflurane (IF) brain
levels 48 h after insult. Mean � SEM for naıve (green bars,
n = 2), HypoT (red bars, n = 8) and HypoT + Mel (blue bars,
n = 7) groups, respectively. None of the four neurochemicals
affected by oxidative stress showed a significant difference
between groups.
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 101
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 13
curve between 0.35 and 2.0). Other studies have also shown the
benefit of delayed intravenous administration of melatonin of elec-
trophysiological recovery after cerebral hypoxia–ischaemia (Lee
et al., 2004).
Melatonin treatment led to elevated plasma levels of melatonin
for 48 h compared with baseline, and the average half-life for
plasma melatonin in the piglets was 21.6 � 3.3 h. This is consider-
ably longer than reported in normothermic rodent studies (20–
30 min) (Cheung et al., 2006); this is likely to be due to the effects
of hypothermia to slow pharmacokinetic clearance of melatonin
(van den Broek et al., 2010). An endogenous increase in plasma
melatonin levels (6–15 times higher) was observed in the
hypothermia-only group after hypoxia–ischaemia. This response
to injury has also been reported in humans after adult traumatic
brain injury (Seifman et al., 2008). It has been demonstrated that
this melatonin release is a critical endogenous neuroprotective
mechanism against hypoxia–ischaemia brain injury using pinealec-
tomized and melatonin-supplemented adult rats (Manev et al.,
1996; Joo et al., 1998).
The precise level of melatonin needed for neuroprotection is
unknown; previous studies in other acute global or focal
hypoxia–ischaemia models have shown neuroprotection with a
wide range of melatonin doses (1.5–50 mg/kg) and timing
(hours or days before or after hypoxia–ischaemia) (Reiter et al.,
2004). We chose 5 mg/kg/h melatonin for safety reasons based
on our pilot study using 5, 10 or 20 mg/kg/h over 6-h melatonin
combined with cooling in the piglet hypoxia–ischaemia insult
model (Supplementary material and Supplementary Fig. 1). A mel-
atonin dose of 10 mg/kg/h or higher was associated with hypo-
tension and a requirement for saline boli and inotropic support to
maintain a normal mean arterial blood pressure; 5-mg/kg/h over
6 h melatonin did not alter the need for support or any physiolo-
gical variable. This dose (5 mg/kg/h over 6 h repeated at 24 h) is
similar to that already used safely in sick newborn infants, where
up to 100 mg of melatonin during 72 h has been administered
safely (Gitto et al., 2001, 2004; Fulia et al., 2005). We adminis-
tered melatonin at 10 min after hypoxia–ischaemia; further studies
are needed to determine the therapeutic window for melatonin
neuroprotection. However, rodent studies, which are more suited
to exploring therapeutic time windows, have shown that the ther-
apeutic window for a single melatonin dose is 4 h (Husson et al.,
2002), and this increases to 24 h with five melatonin doses 24 h
apart (P. Gressens, unpublished).
Receptor and non-receptor pathways underpin the multiple
neuroprotective effects of melatonin that include supporting mito-
chondrial function and post-lesional plasticity and its anti-oxidant,
anti-apoptotic and anti-inflammatory actions (Husson et al., 2002;
Leon et al., 2004; Kilic et al., 2005; Tan et al., 2007; Welin et al.,
2007; Gressens et al., 2008). This study was designed to assess
the preclinical safety and efficacy of melatonin to augment
hypothermia and not specifically its mechanism of action.
However, aspects of this study suggest melatonin may act to
maintain cellular energetics and modulate neuroinflammation.
In the present study, cerebral ATP levels are protected by mel-
atonin treatment, indicating a non-receptor-mediated effect of
melatonin directly on the electron transport chain (Leon et al.,
2004). Melatonin also prevented the increase in lactate/N-acetyl
aspartate in the reperfusion phase (increased lactate/N-acetyl
aspartate is indicative of compromised or damaged mitochondria)
(Penrice et al., 1997; Kristian, 2004; Moffett et al., 2007); this
effect may be due to melatonin’s actions to prevent compromise
of the mitochondrial membranes (Andrabi et al., 2004; Wang,
2009). Such positive effects on the mitochondria and melatonin’s
ability to increase expression of anti-apoptotic proteins (Ling et al.,
1999) likely play a role in reducing the TUNEL and activated
caspase 3 cell death in the melatonin-augmented hypothermia
group.
Melatonin treatment in this study failed to reduce microglial cell
number but decreased (to levels seen in naıve animals) expression
of CD86, a prototypical marker of cytotoxic microglial activation,
due to its co-stimulatory role with major histocompatibility com-
plex Class II (Allison, 1994). Inducible nitric oxide synthase is also
a marker of a cytotoxic microglial phenotype (Block et al., 2007),
but we observed no change in its expression due to melatonin, as
levels were already possibly reduced by hypothermia to levels seen
in the naıve subjects. This is in agreement with the anti-inflamma-
tory effects of hypothermia that have been previously reported
(Lynch et al., 1975; Yenari and Han, 2012). Previous observations
in normothermia hypoxia–ischaemia injury models have shown
that melatonin treatment is anti-inflammatory (Welin et al.,
2007; Koh, 2008; Villapol et al., 2011). Melatonin treatment
increased expression of SphK1, which is produced (at least in
part) by microglia suggested to be in a deactivated state
(Colton, 2009). SphK1 activation is capable of decreasing
proinflammatory cytokine production, increasing proliferation and
decreasing cell death (Hughes et al., 2008). Melatonin-augmented
hypothermia also altered expression of the immunomodulatory
cytokine SOCS3 produced by microglia (Qin et al., 2007). In
rodents, SOCS3 is increased after brain injury (Raghavendra Rao
et al., 2003), and melatonin has previously been demonstrated to
reduce SOCS3 expression (Tsai et al., 2011). Altogether these data
suggest that melatonin-mediated neuroprotection may act in part
by modulating microglial phenotype.
Melatonin-augmented hypothermia had no effect on brain iso-
prostanes at 48 h after hypoxia–ischaemia compared with hypother-
mia alone. This is in contrast to previous observations after
melatonin treatment in normothermic hypoxia–ischaemia injury
models where serum measurements of 8-isoprostane were reduced
at 6 h after hypoxia–ischaemia in those sheep foetuses administered
melatonin (Welin et al., 2007). The absence of any effect of mela-
tonin on isoprostanes in our study may be due to the rise in reactive
oxygen species occurring at time points well before 48 h when our
cerebral tissue was harvested or due to hypothermia already maxi-
mally reducing reactive oxygen species (Yenari and Han, 2012).
Indeed, when comparing the hypothermia-only group with naıve
brain, there was no difference (Fig. 6).
The piglet model of hypoxia–ischaemia used in the current study
has strong preclinical value, as it allows hypothermia to be applied
as it would in a clinical setting, and an intensive care level that is
on par with a neonatal intensive care unit. This includes supported
ventilation, maintenance of cardiac output, blood volume and
electrolytes, cardiac resuscitation (if required) and continuous
EEG monitoring. These studies have been designed in such a
way that they are ‘proof of principle’, i.e. constructed to maximize
102 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 14
the demonstrable therapeutic effect. Thus, in the current study,
piglets were subjected to a severe hypoxia–ischaemia insult, and
intervention with melatonin was started at 10 min of completing
hypoxia–ischaemia; this would be difficult to perform in the clinic.
We initiated cooling 2 h after hypoxia–ischaemia, which is in line
with recent suggested protocols for hypothermia (Kendall et al.,
2010), although sooner than in randomized clinical trials
(Gluckman et al., 2005; Shankaran et al., 2005; Azzopardi,
2010). Cooling for 24 h is shorter than the current clinical protocol
of 72 h (Azzopardi et al., 2009), but the neuroprotective efficacy
of this regime has previously been validated in this piglet model
(O’Brien et al., 2006).
Melatonin’s multiple neuroprotective actions, its ability to pene-
trate the brain and organelles (Acuna-Castroviejo et al., 2001) and
lack of toxicity in humans (Rybakowski et al., 1995) and animals
(Jahnke et al., 1999) make it an attractive neuroprotective agent.
Melatonin is relatively easy to administer by intravenous (Cheung
et al., 2006), oral (Mistraletti et al., 2010) and rectal (Weishaupt
et al., 2006) routes. In our study, melatonin-augmented hypother-
mia was safe and provided significant neuroprotection compared
with hypothermia alone in this piglet perinatal asphyxia model.
Augmented neuroprotection was demonstrated by improved cere-
bral energy metabolism on MRS biomarkers and reduced cell
death across the brain. These data along with the safety profile
of melatonin suggest the consideration of phase I and II clinical
studies of melatonin-augmented therapeutic hypothermia for neo-
natal encephalopathy.
AcknowledgementsWe thank Aron Kerenyi and Dorottya Kelen for their help with the
experimental series, Kevin Broad for data analysis, Fabrizio Proietti,
Mariangela Longini and Giuseppe Buonocore (Sienna, Italy) for the
isoprostane study, Mrs Odile Blanchard (INSERM, France) for her
helpful technical assistance with the melatonin pharmacokinetic
study and Sophie Lebon (INSERM, Paris) for the work on quanti-
tative PCR for the inflammatory markers.
FundingThe study was funded by Wellbeing of Women, UK. This work
was undertaken at University College Hospital/University College
London, which received a proportion of funding from the UK
Department of Health’s National Institute for Health Research
Biomedical Research Centres funding scheme.
Supplementary materialSupplementary material is available at Brain online.
ReferencesAcuna-Castroviejo D, Martın M, Macıas M, Escames G, Leon J,
Khaldy H, et al. Melatonin, mitochondria, and cellular bioenergetics.
J Pineal Res 2001; 30: 65–72.
Allison J. CD28-B7 interactions in T-cell activation. Curr Opin Immunol
1994; 6: 414–19.
Altun A, Ugur-Altun B. Melatonin: therapeutic and clinical utilization. Int
J Clin Pract 2007; 61: 835–45.
Andrabi S, Sayeed I, Siemen D, Wolf G, Horn T. Direct inhibition of the
mitochondrial permeability transition pore: a possible mechanism
responsible for anti-apoptotic effects of melatonin. FASEB J 2004;
18: 869–71.Azzopardi D. Clinical management of the baby with hypoxic ischaemic
encephalopathy. Semin Fetal Neonatal Med 2010; 86: 345–50.Azzopardi D. Neuroprotective effects of hypothermia combined with
inhaled xenon following perinatal asphyxia (TOBYXe) 2011.
Available from: http://clinicaltrialsgov/ct2/results?term=00934700
(19 September 2011, date last accessed).
Azzopardi D, Strohm B, Edwards A, Dyet L, Halliday H, Juszczak E, et al.
Moderate hypothermia to treat perinatal asphyxial encephalopathy.
N Engl J Med 2009; 361: 1349–58.Azzopardi D, Wyatt J, Cady E, Delpy D, Baudin J, Stewart A, et al.
Prognosis of newborn infants with hypoxic-ischemic brain injury
assessed by phosphorus magnetic resonance spectroscopy. Pediatr
Res 1989; 25: 445–51.
Battin M, Thoresen M, Robinson E, Polin R, Edwards A, Gunn A. Cool
Cap Trial Group. Does head cooling with mild systemic hypothermia
affect requirement for blood pressure support? Pediatrics 2009; 123:
1031–6.
Benıtez-King G, Anton-Tay F. Calmodulin mediates melatonin cytoskel-
etal effects. Experimentia 1993; 49: 635–41.
Block M, Zecca L, Hong J. Microglia-mediated neurotoxicity: uncovering
the molecular mechanisms. Nat Rev Neurosci 2007; 8: 57–69.
Bona E, Hagberg H, Løberg E, Bagenholm R, Thoresen M. Protective
effects of moderate hypothermia after neonatal hypoxia-ischemia:
short- and long-term outcome. Pediatr Res 1998; 43: 738–45.
Brzezinski A. Melatonin in humans. N Engl J Med 1997; 336: 186–95.
Bubenik G, Konturek S. Melatonin and aging: prospects for human treat-
ment. J Physiol Pharmacol 2011; 62: 13–9.
Cheong J, Cady E, Penrice J, Wyatt J, Cox I, Robertson NJ. Proton MR
spectroscopy in neonates with perinatal cerebral hypoxic-ischemic
injury: metabolite peak-area ratios, relaxation times, and absolute con-
centrations. Am J Neuroradiol 2006; 27: 1546–54.
Cheung R, Tipoe G, Tam S, Ma E, Zou L, Chan P. Preclinical evaluation
of pharmacokinetics and safety of melatonin in propylene glycol for
intravenous administration. J Pineal Res 2006; 41: 337–43.
Cinar M, Islam M, Uddin M, Tholen E, Tesfaye D, Looft C, et al.
Evaluation of suitable reference genes for gene expression studies in
porcine alveolar macrophages in response to LPS and LTA. BMC Res
Notes 2012; 5: 107.
Claustrat B, Chazot G, Brun J, Jordan D, Sassolas G. A chronobiological
study of melatonin and cortisol secretion in depressed subjects: plasma
melatonin, a biochemical marker in major depression. Biol Psychiatry
1984; 19: 1215–28.
Colton C. Heterogeneity of microglial activation in the innate immune
response in the brain. J Neuroimmune Pharmacol 2009; 4: 399–418.
de Vries L, Hellstrom-Westas L. Role of cerebral function monitoring in
the newborn. Arch Dis Child Fetal Neonatal Ed 2005; 90: F201–7.
Edwards A, Brocklehurst P, Gunn A, Halliday H, Juszczak E, Levene M,
et al. Neurological outcomes at 18 months of age after moderate
hypothermia for perinatal hypoxic ischaemic encephalopathy: synthesis
and meta-analysis of trial data. BMJ 2010; 340: C363.Faulkner S, Bainbridge A, Kato T, Chandrasekaran M, Hristova M, Liu M,
et al. Xenon augmented hypothermia reduces early lactate/NAA and
cell death in Perinatal Asphyxia. Ann Neurol 2011; 70: 133–50.
Favrais G, Schwendimann L, Gressens P, Lelievre V. Cyclooxygenase-2
mediates the sensitizing effects of systemic IL-1-beta on excitotoxic
brain lesions in newborn mice. Neurobiol Dis 2007; 25: 496–505.
Fulia F, Gitto E, Cuzzocrea S, Reiter R, Dugo L, Gitto P, et al. Increased
levels of malondialdehyde and nitrite/nitrate in the blood of
asphyxiated newborns: reduction by melatonin. J Pineal Res 2005;
31: 343–9.
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 103
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 15
Gitto E, Karbownik M, Reiter R, Tan D, Cuzzocrea S, Chiurazzi P, et al.
Effects of melatonin treatment in septic newborns. Pediatr Res 2001;
50: 756–60.
Gitto E, Romeo C, Reiter R, Impellizzeri P, Pesce S, Basile M, et al.
Melatonin reduces oxidative stress in surgical neonates. J Pediatr
Surg 2004; 39: 184–9.
Gluckman P, Wyatt J, Azzopardi D, Ballard R, Edwards A, Ferriero D,
et al. Selective head cooling with mild systemic hypothermia after
neonatal encephalopathy: multicentre randomised trial. Lancet 2005;
365: 663–70.
Gressens P, Schwendimann L, Husson I, Sarkozy G, Mocaer E, Vamecq J,
et al. Agomelatine, a melatonin receptor agonist with 5-HT(2C) recep-
tor antagonist properties, protects the developing murine white matter
against excitotoxicity. Eur J Pharmacol 2008; 588: 58–63.
Hughes J, Srinivasan S, Lynch K, Proia R, Ferdek P, Hedrick C.
Sphingosine-1-phosphate induces an antiinflammatory phenotype in
macrophages. Circ Res 2008; 102: 950–8.Husson I, Mesples B, Bac P, Vamecq J, Evrard P, Gressens P.
Melatoninergic neuroprotection of the murine periventricular white
matter against neonatal excitotoxic challenge. Ann Neurol 2002; 51:
82–92.
Ito D, Tanaka K, Suzuki S, Dembo T, Fukuuchi Y. Enhanced expression
of Iba1, ionized calcium-binding adapter molecule 1, after transient
focal cerebral ischemia in rat brain. Stroke 2001; 32: 1208–15.
Jahnke G, Marr M, Myers C, Wilson R, Travlos G, Price C. Maternal and
developmental toxicity evaluation of melatonin administered orally to
pregnant Sprague-Dawley rats. Toxicol Sci 1999; 50: 271–9.Jatana M, Singh I, Singh A, Jenkins D. Combination of systemic hypo-
thermia and N-acetylcysteine attenuates hypoxic-ischemic brain injury
in neonatal rats. Pediatr Res 2006; 59: 684–9.
Joo J, Uz T, Manev H. Opposite effects of pinealectomy and melatonin
administration on brain damage following cerebral focal ischemia in
rat. Restor Neurol Neurosci 1998; 13: 185–91.
Kendall G, Kapetanakis A, Ratnavel N, Azzopardi D, Robertson NJ.
Cooling on Retrieval Study Group. Passive cooling for initiation of
therapeutic hypothermia in neonatal encephalopathy. Arch Dis Child
Fetal Neonatal Ed 2010; 95: F408–12.Kerenyi A, Kelen D, Faulkner S, Bainbridge A, Chandrasekaran M,
Cady E, et al. Systemic effects of whole-body cooling to 35�C,
33.5�C, and 30�C in a piglet model of perinatal asphyxia: implications
for therapeutic hypothermia. Pediatr Res 2012; 71: 573–82.
Kilic U, Kilic E, Reiter R, Bassetti C, Hermann D. Signal transduction
pathways involved in melatonin-induced neuroprotection after focal
cerebral ischemia in mice. J Pineal Res 2005; 38: 67–71.
Koh P. Melatonin regulates nitric oxide synthase expression in ischemic
brain injury. J Vet Med Sci 2008; 70: 747–50.
Kristian T. Metabolic stages, mitochondria and calcium in hypoxic/
ischemic brain damage. Cell Calcium 2004; 36: 221–33.
Lee E, Lee M, Chen H, Hsu Y, Wu T, Chen S, et al. Melatonin attenuates
gray and white matter damage in a mouse model of transient focal
cerebral ischemia. J Pineal Res 2005; 38: 42–52.Lee E, Wu T, Lee M, Chen T, Tsai Y, Chuang J, et al. Delayed treatment
with melatonin enhances electrophysiological recovery following tran-
sient focal cerebral ischemia in rats. J Pineal Res 2004; 36: 33–42.
Lee M, Kuan Y, Chen H, Chen T, Chen S, Huang C, et al. Intravenous
administration of melatonin reduces the intracerebral cellular inflam-
matory response following transient focal cerebral ischemia in rats. J
Pineal Res 2007; 42: 297–309.
Leon J, Acuna-Castroviejo D, Sainz R, Mayo J, Tan D, Reiter R.
Melatonin and mitochondrial function. Life Sci 2004; 75: 765–90.
Lewy A, Wehr T, Goodwin F, Newsome D, Markey S. Light suppresses
melatonin secretion in humans. Science 1980; 210: 1267–9.
Ling X, Zhang L, Lu S, Li X, Sun F. Protective effect of melatonin on
injuried cerebral neurons is associated with bcl-2 protein
over-expression. Zhongguo Yao Li Xue Bao 1999; 20: 409–14.Liu Y, Barks J, Xu G, Silverstein F. Topiramate extends the therapeutic
window for hypothermia-mediated neuroprotection after stroke in
neonatal rats. Stroke 2004; 35: 1460–5.
Lorek A, Takei Y, Cady E, Wyatt J, Penrice J, Edwards A, et al.
Delayed (“secondary”) cerebral energy failure after acute
hypoxia-ischemia in the newborn piglet: continuous 48-hour studies
by phosphorus magnetic resonance spectroscopy. Pediatr Res 1994;
36: 699–706.
Luchetti F, Canonico B, Betti M, Arcangeletti M, Pilolli F, Piroddi M, et al.
Melatonin signaling and cell protection function. FASAB J 2010; 24:
3603–24.Lynch H, Wurtman R, Moskowitz M, Archer M, Ho M. Daily rhythm in
human urinary melatonin. Science 1975; 187: 169–71.Manev H, Uz T, Kharlamov A, Joo J. Increased brain damage after stroke
or excitotoxic seizures in melatonin-deficient rats. FASEB J 1996; 10:
1546–51.Miller S, Yan E, Castillo-Melendez M, Jenkin G, Walker D. Melatonin
provides neuroprotection in the late-gestation fetal sheep brain
in response to umbilical cord occlusion. Dev Neurosci 2005; 27:
200–10.
Milne G, Yin H, Brooks J, Sanchez S, Jackson Roberts L, Morrow J.
Quantification of F2-isoprostanes in biological fluids and tissues as a
measure of oxidant stress. Methods Enzymol 2007; 72: 1419–24.
Mistraletti G, Sabbatini G, Taverna M, Figini M, Umbrello M, Magni P,
et al. Pharmacokinetics of orally administered melatonin in critically ill
patients. J Pineal Res 2010; 48: 142–7.
Moffett JR, Ross B, Arun P, Madhavarao CN, Namboodiri AM.
N-Acetylaspartate in the CNS: from neurodiagnostics to neurobiology.
Prog Neurobiol 2007; 2: 89–131.
National Institute for Clinical Excellence: Therapeutic hypothermia with
intracorporeal temperature monitoring for hypoxic perinatal brain
injury: guidance NICE guidelines: interventional procedures 2010.
Available from: http://www.nice.org.uk/nicemedia/live/11315/
48809/48809.pdf.O’Brien F, Iwata O, Thornton J, De Vita E, Sellwood M, Iwata S, et al.
Delayed whole-body cooling to 33 or 35 degrees C and the develop-
ment of impaired energy generation consequential to transient cerebral
hypoxia-ischemia in the newborn piglet. Pediatrics 2006; 117:
1549–59.
Pei Z, Pang S, Cheung R. Administration of melatonin after onset of
ischemia reduces the volume of cerebral infarction in a rat middle
cerebral artery occlusion stroke model. Stroke 2003; 34: 770–5.
Pei Z, Pang SF, Cheung RT. Pretreatment with melatonin reduces volume
of cerebral infarction in a rat middle cerebral artery occlusion stroke
model. J Pineal Res 2002; 32: 168–72.
Penrice J, Lorek A, Cady EB, Amess PN, Wylezinska M, Cooper CE, et al.
Proton magnetic resonance spectroscopy of the brain during acute
hypoxia-ischemia and delayed cerebral energy failure in the newborn
piglet. Pediatr Res 1997; 41: 795–802.Provencher S. Estimation of metabolite concentrations from localized in
in vivo proton NMR spectra. Magn Reson Med 1993; 30: 672–9.Qin H, Roberts K, Niyongere S, Cong Y, Elson C, Benveniste E.
Molecular mechanism of lipopolysaccharide-induced SOCS-3 gene ex-
pression in macrophages and microglia. J Immunol 2007; 179:
5966–76.
Raghavendra Rao V, Dhodda V, Song G, Bowen K, Dempsey R.
Traumatic brain injury-induced acute gene expression changes in rat
cerebral cortex identified by GeneChip analysis. J Neurosci Res 2003;
71: 208–19.
Reiter R, Tan D, Fuentes-Broto L. Melatonin: a multitasking molecule.
Prog Brain Res 2010; 181: 127–51.
Reiter R, Tan D, Mayo J, Sainz R, Leon J, Czarnocki Z. Melatonin as an
antioxidant: biochemical mechanisms and pathophysiological implica-
tions in humans. Acta Biochim Pol 2003; 50: 1129–46.
Reiter R, Tan D, Pappolla M. Melatonin relieves the neural oxidative
burden that contributes to dementias. Ann N Y Acad Sci 2004;
1035: 179–96.
Robertson N, Cox I, Cowan F, Counsell S, Azzopardi D, Edwards A.
Cerebral intracellular lactic alkalosis persisting months after neonatal
encephalopathy measured by magnetic resonance spectroscopy.
Pediatr Res 1999; 46: 287–96.
104 | Brain 2013: 136; 90–105 N. J. Robertson et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022
Page 16
Rybakowski C, Mohar B, Wohlers S, Leichtweiss H, Schroder H. Thetransport of vitamin C in the isolated human near-term placenta. Eur
J Obstet Gynecol Reprod Biol 1995; 62: 107–14.
Seifman M, Adamides A, Nguyen P, Vallance S, Cooper D, Kossmann T,
et al. Endogenous melatonin increases in cerebrospinal fluid of patientsafter severe traumatic brain injury and correlates with oxidative stress
and metabolic disarray. J Cereb Blood Flow Metab 2008; 28: 684–96.
Shankaran S, Laptook A, Ehrenkranz R, Tyson J, McDonald S,
Donovan E, et al. Whole-body hypothermia for neonates withhypoxic-ischemic encephalopathy. N Engl J Med 2005; 353: 1574–84.
Tan D, Manchester L, Terron M, Flores L, Reiter R. One molecule, many
derivatives: a never-ending interaction of melatonin with reactiveoxygen and nitrogen species? J Pineal Res 2007; 42: 28–42.
Tan D, Reiter R, Manchester L, Yan M, El-Sawi M, Sainz R, et al.
Chemical and physical properties and potential mechanisms: melatonin
as a broad spectrum antioxidant and free radical scavenger. Curr TopMed Chem 2002; 2: 181–97.
Thayyil S, Chandrasekaran M, Taylor A, Bainbridge A, Cady E, Chong K,
et al. Cerebral magnetic resonance biomarkers in neonatal encephal-
opathy: a meta-analysis. Pediatrics 2010; 125: e382–95.Thoresen M, Hellstrom-Westas L, Liu X, de Vries L. Effect of hypother-
mia on amplitude-integrated electroencephalogram in infants with
asphyxia. Pediatrics 2010; 126: e131–39.
Thoresen M, Penrice J, Lorek A, Cady E, Wylezinska M, Kirkbride V,et al. Mild hypothermia after severe transient hypoxia-ischemia ameli-
orates delayed cerebral energy failure in the newborn piglet. Pediatr
Res 1995; 37: 667–70.Toet M, Hellstrom-Westas L, Groenendaal F, Eken P, de Vries L.
Amplitude integrated EEG 3 and 6 hours after birth in full term neo-
nates with hypoxic-ischaemic encephalopathy. Arch Dis Child Fetal
Neonatal Ed 1999; 81: F19–23.
Tsai M, Chen W, Tsai M, Ching C, Chuang J. Melatonin attenuates
brain contusion-induced oxidative insult, inactivation of signal
transducers and activators of transcription 1, and upregulation of
suppressor of cytokine signaling-3 in rats. J Pineal Res 2011; 51:
233–45.
van den Broek M, Groenendaal F, Egberts A, Rademaker C. Effects of
hypothermia on pharmacokinetics and pharmacodynamics: a system-
atic review of preclinical and clinical studies. Clin Pharmacokinet 2010;
49: 277–94.
Vanhamme L, van den Boogaart A, van Huffel S. Improved method for
accurate and efficient quantification of MRS data with use of prior
knowledge. J Magn Reson 1997; 129: 35–43.
Villapol S, Fau S, Renolleau S, Biran V, Charriaut-Marlangue C, Baud O.
Melatonin promotes myelination by decreasing white matter inflam-
mation after neonatal stroke. Pediatr Res 2011; 69: 51–5.
Wang X. The antiapoptotic activity of melatonin in neurodegenerative
diseases. CNS Neurosci Ther 2009; 15: 345–57.
Weishaupt J, Bartels C, Polking E, Dietrich J, Rohde G, Poeggeler B, et al.
Reduced oxidative damage in ALS by high-dose enteral melatonin
treatment. J Pineal Res 2006; 41: 313–23.
Welin AK, Svedin P, Lapatto R, Sultan B, Hagberg H, Gressens P, et al.
Melatonin reduces inflammation and cell death in white matter in the
mid-gestation fetal sheep following umbilical cord occlusion. Pediatr
Res 2007; 61: 153–8.
Werner A, Martin S, Gutierrez-Ramos J, Raivich G. Leukocyte recruit-
ment and neuroglial activation during facial nerve regeneration in
ICAM-1-deficient mice: effects of breeding strategy. Cell Tissue Res
2001; 305: 25–41.
Yenari M, Han H. Neuroprotective mechanisms of hypothermia in brain
ischaemia. Nat Rev Neurosci 2012; 13: 267–78.
Melatonin neuroprotection with cooling Brain 2013: 136; 90–105 | 105
Dow
nloaded from https://academ
ic.oup.com/brain/article/136/1/90/431810 by guest on 21 February 2022