Microbiological processes in banded iron formation deposition
NICOLE R. POSTH*1, KURT O. KONHAUSER† and ANDREAS KAPPLER**Center for Applied Geosciences, University of Tuebingen, 72076, Tuebingen, Germany(E-mail: [email protected])†Department of Earth and Atmospheric Sciences, University of Alberta, Edmonton, Alberta, CanadaT6G 2E3
ABSTRACT
Banded iron formations have been studied for decades, particularly regard-
ing their potential as archives of the Precambrian environment. In spite of
this effort, the mechanism of their deposition and, specifically, the role that
microbes played in the precipitation of banded iron formation minerals,
remains unresolved. Evidence of an anoxic Earth with only localized oxic
areas until the Great Oxidation Event ca 2�45 to 2�32 Ga makes the investi-
gation of O2-independent mechanisms for banded iron formation deposition
relevant. Recent studies have explored the long-standing proposition that
Archean banded iron formations may have been formed, and diagenetically
modified, by anaerobic microbial metabolisms. These efforts encompass a
wide array of approaches including isotope, ecophysiological and phylog-
eny studies, molecular and mineral marker analysis, and sedimentological
reconstructions. Herein, the current theories of microbial processes in
banded iron formation mineral deposition with particular regard to the
mechanisms of chemical sedimentation and post-depositional alteration are
described. The main findings of recent years are summarized and compared
here, and suggestions are made regarding cross-disciplinary information
still required to constrain the role of the biosphere in banded iron forma-
tion deposition.
Keywords Aerobic and anaerobic Fe(II) oxidation, anoxygenic phototrophs,atmospheric evolution, banded iron formations, cell-mineral aggregates,mineral diagenesis.
INTRODUCTION
Precambrian banded iron formation (BIF) depo-sition continues to be a topic of research interestbecause their mineral and chemical composi-tions provide direct evidence of the environ-mental conditions that existed at the time oftheir deposition. In particular, a number ofrecent studies have focused on how BIFs can beused as proxies for the emergence of oxygen,from its initial production in the oceans by atleast 2�7 Ga (Eigenbrode & Freeman, 2006;Godfrey & Falkowski, 2009; Kendall et al.,
2010), to its spread throughout the atmospherebetween 2�45 Ga and 2�32 Ga, the so-calledGreat Oxidation Event (GOE) (Rye & Holland,1998; Pavlov & Kasting, 2002; Bekker et al.,2004; Buick, 2008; Farquhar et al., 2011; Konha-user et al., 2011a,b). In this regard, BIFs offer anarchive of both early microbial life and thetransition to an oxygen-dominated Earth system.At the heart of understanding the formation of
these deposits lies the potential interaction ofabiotic and biological processes in these ancientsystems; both abiotic and biotic models of for-mation exist and continue to be explored. The
1Nordic Centre for Earth Evolution (NordCEE), Institute for Biology, University of Southern Denmark, 5230,Odense, Denmark
relevant question to the efficacy of the microbialmodels in terms of BIF precipitation is whetherthe dominant processes of Fe(II) oxidation andmineral precipitation can be deduced from their:(i) microfossils and fabrics; (ii) stable isotopecomposition; (iii) biosignature identity; (iv) geo-chemical composition; and (v) sedimentologicalfeatures. Moreover, if a biological mechanismwas important in the initial process of Fe(II) oxi-dation in the ancient ocean water column, it isexpected that biomass would have settled to thesea floor along with the Fe(III) minerals. Thisorganic carbon would subsequently have servedas an oxidizable substrate for sedimentarymicrobial communities during diagenesis andmetamorphism. These post-depositional pro-cesses would have modified the initial BIF pre-cursor sediment into the mineral assemblagesobserved today. Herein, the existing evidencefor microbiological involvement in both BIFprecipitation and burial is summarized.
BANDED IRON FORMATION SETTINGAND DEPOSITIONAL THEORIES
Banded iron formations are iron-rich (ca 20 to40% Fe) and siliceous (ca 40 to 50% SiO2) sedi-mentary deposits that precipitated throughoutmuch of the late Archean (2�7 to 2�5 Ga) and Pal-aeoproterozoic (2�5 to 1�8 Ga). The ‘Superior’ typeBIFs, including those in the Hamersley Group,Western Australia, and the Transvaal Super-group, South Africa, are hundreds of metresthick, over 105 km2 in areal extent and contain>1013 tons of iron (Beukes, 1984; Trendall, 2002).These formations are characteristically laminated(James, 1954; Fig. 1), with banding observed overa wide range of scales, from coarse macrobands(metres in thickness) to mesobands (centimetre-thick units) to millimetre and sub-millimetre lay-ers (Trendall & Blockley, 1970). Among the latterare the various varve-like repetitive laminae,known as microbands. Although it is not resolvedthat these microbands are seasonal, much of thework on these structures is based on that premise.The mineralogy of the least metamorphosed
BIFs consists of chert, magnetite, hematite, car-bonates (siderite and dolomite-ankerite), green-alite, stilpnomelane and riebeckite (Klein, 2005;Bekker et al., 2010): the presence of both ferricand ferrous minerals gives BIFs an averageoxidation state of Fe2�4+ (Klein & Beukes, 1992).It is generally agreed that none of the mineralsin BIFs are primary, but that instead, the mine-
rals reflect both diagenetic and metamorphicoverprinting. For instance, the primary ironminerals were most probably ferric hydroxide[Fe(OH)3], greenalite [(Fe)3Si2O5(OH)4] andsiderite (FeCO3) (Han, 1966, 1978; Dimroth &Chauvel, 1973; Perry et al., 1973; Klein, 2005). Ithas also been suggested that the Archean oceanhad significantly elevated concentrations ofdissolved silica, at least as high as at saturationwith cristobalite (0�67 mM at 40°C in sea water),and possibly even amorphous silica (2�20 mM)(Maliva et al., 2005; Konhauser et al., 2007a).Under such silica-rich conditions, the precipita-tion of amorphous silica could have taken placedirectly on the sea floor (Krape�z et al., 2003).Although BIFs are generally believed to have
been deposited on the continental shelf at waterdepths >200 m due to the lack of obvious cur-rent and wave-generated structures (Beukes,1973; Klein & Beukes, 1992; Morris & Horwitz,1983; for review see Klein, 2005), it has beenargued that some were deposited off the conti-nental slope in the deep ocean (Krape�z et al.,2003). The dominant source of Fe(II) into theArchean ocean was hydrothermal (Jacobsen &Pimentel-Klose, 1988; Bau & M€oller, 1993;Hamade et al., 2003), but the proximity of theFe(II) source to the site of deposition is stillunclear. It is believed that the concentration ofFe(II) in these basins ranged from 0�05 to 0�5 mM
(Holland, 1973; Morris, 1993). On the one hand,Fe could have been delivered from the deepocean to the outer continental shelf by upwell-ing currents from a mid-ocean ridge system(Holland, 1973; Morris & Horwitz, 1983).Accordingly, BIFs would sediment from belowthe wave base (without the influence of waveand storm-induced currents) onto partially sub-merged platforms of the continental shelves. Onthe other hand, the direct supply of Fe(II) intothe photic zone by hydrothermal plumes associ-ated with shallow seamount-type systems (Isley,1995; Isley & Abbot, 1999) would curtail the dif-ficulties introduced by the high upwelling ratesneeded to bring sufficient iron from the deep seaonto the continental shelf (Konhauser, 2007b).The mineralogy of BIFs dictates that some oxi-
dation of Fe(II) was necessary for formation,although the dominant mechanism(s) is uncertain(Fig. 2). Prior to the rise of atmospheric oxygenand the development of a protective ozone layer,the Earth’s surface was subjected to high levels ofultraviolet (UV) radiation. Bulk ocean waters thatwere anoxic at this time could have supportedhigh concentrations of dissolved Fe(II). Under
such conditions, dissolved ferrous iron species,such as Fe2+ or Fe(OH)+, would have absorbedradiation in the 200 to 400 nm range, leading tothe formation of dissolved ferric iron [reaction 1],which, in turn, hydrolysed to form ferric hydrox-ide at circumneutral pH (Cairns-Smith, 1978;Braterman et al., 1983; Fig. 2A).
2Fe2þðaqÞ þ 2Hþ þ hv ! 2Fe3þðaqÞ þH2 " ð1Þ
However, these experiments focused on deter-mining the specific rates of Fe(II) photochemicaloxidation, and did not simulate the complex,disequilibrium water chemistry characteristic ofan ocean where Fe(II)-rich hydrothermal watersreacted with ambient Si-saturated sea water thatalso contained high concentrations of HCO3
�.Indeed, in fluids with high dissolved Fe(II),Si(OH)4 and HCO3
�, the oxidation effects of
either UVA or UVC were found to be negligiblecompared with the precipitation of ferrous-ironsilicates (Konhauser et al., 2007b).As an alternative to the abiological model, the
presence of ferric iron minerals in BIFs has alsobeen ascribed to the metabolic activity of plank-tonic bacteria in the ocean’s photic zone. Chemi-cal oxidation of Fe(II) by photosyntheticallyproduced O2 is one possibility, allowing for theindirect biogenic precipitation of ferric hydrox-ide (Fig. 2B and C). Under an anoxic atmosphere,this O2 could have been confined to localized‘oxygen oases’ associated with cyanobacterialblooms in coastal settings (Cloud, 1965, 1973).Cloud (1965, 1973) further proposed that suchprimitive O2-producing photosynthetic bacteria,which lacked suitably advanced oxygen-mediat-ing enzymes, required ferrous iron to detoxifyoxygen. If so, these micro-organisms would have
A B C
Fig. 1. Overview of the laminated BIF from the Joffre iron formation, Pilbara Craton, north-west Australia. Imagecourtesy of Stefan Lalonde (A). Core consists of red microbands (<1 mm) of chert-hematite-riebeckite (bluishbands in upper core) alternating with lighter chert–dolomite–siderite–crocidolite mesobands (� 1 cm) and denser,dark magnetite mesobands. A slumped chert band due to post-depositional compaction is visible in core top (B).A scanned thin section of the Joffre iron formation in plane-polarized light (C) shows dense blue riebeckite micro-bands interbedded with brownish hematite/goethite microbands and chert microbands and mesobands. The chertmesobands contain wavy, fine-grained carbonate and hematite/goethite microbands (image scale: 2�6 9 4�6 cm).Images (B) and (C) courtesy of Rasmus Haugaard.
Microbiological processes in banded iron formation deposition 1735
flourished specifically when Fe(II) and nutrientswere made episodically available (Cloud, 1969;Fralick & Pufahl, 2006). Once oxygen was pre-sent, microaerophilic Fe(II) oxidizers could havecontributed to the direct precipitation of ferrichydroxide (Holm, 1989).Garrels et al. (1973) and Hartman (1984) later
suggested that light, not O2, may have coupledthe carbon and iron cycles, via photosynthesisthat used Fe(II) rather than H2O as an electrondonor, producing Fe(III) rather than O2 (Fig. 2D).With the evolution of this photoferrotrophy, bio-logical Fe(II) oxidation would have supersededphotochemical oxidation because the bacteriacould grow deeper in the water column whereUV radiation would be effectively absorbed (Ko-nhauser et al., 2002). Modelling of experimen-tally determined photosynthetic Fe(II) oxidationrates even suggests that such micro-organismscould have accounted for all of the Fe(III) initiallydeposited in primary BIF sediment (Kappleret al., 2005).Although the average oxidation state of the
iron in BIFs (Fe2�4+) can be attributed to thesimultaneous deposition of Fe(II) and Fe(III), thepartial microbial or abiotic reduction of Fe(III) to
Fe(II) can also explain this mixed oxidation state.The presence of magnetite as: (i) disseminatedgrains within but obscuring sedimentary lami-nae; (ii) laminated beds that clearly truncate sedi-mentary layering; and (iii) cleavage fills, suggestsa secondary origin to magnetite in BIFs (Ewers &Morris, 1981; Krape�z et al., 2003). MicrobialFe(III) reduction is carried out by anaerobic het-erotrophic bacteria that link this process toorganic matter oxidation (Lovley & Phillips,1988). Fe(III) oxyhydroxides can also be abioti-cally reduced after sedimentation with a reduc-tant, such as the organic carbon associated withplanktonic bacteria necromass, and can producesiderite (FeCO3) and magnetite (Fe3O4) (Schae-dler et al., 2009; Posth et al., 2010). Recentexperiments point to the production of sideritespherulites, also found in many BIFs, by theexperimental diagenesis of ferrihydrite and glu-cose at relevant pressure and temperature. Threelines of evidence point to the antiquity of suchan anaerobic respiratory pathway. Firstly, extanthyperthermophilic Bacteria and Archaea branchdeeply in the universal phylogenetic tree and canreduce Fe(III) (Vargas et al., 1998). Secondly,highly negative d56Fe values in magnetite-rich
A B
C D
Fig. 2. Models of BIF deposition: For an anoxic water column, the proposed abiotic mechanism of Fe(II) photooxida-tion by UV light (A). The traditional model of BIF deposition involves production of oxygen by cyanobacteria, whichthen chemically reacts with hydrothermal dissolved Fe(II). The restriction of cyanobacterial mats to the near-shorewould physically separate Fe(III) oxyhydroxides from organic carbon precipitates (B), which would not be the casein a system where cyanobacteria also populated off-shore regions (C). A biotic mechanism of deposition proposed foran anoxic setting is the direct microbial oxidation of Fe(II) via anoxygenic Fe(II)-oxidizing phototrophy (D).
BIF samples as old as 2�9 Ga have been observed(Johnson et al., 2003; Yamaguchi et al., 2005) andare comparable to the negative fractionationsmeasured in experimental culture with dissimila-tory Fe(III)-reducing bacteria (Johnson et al.,2005). Thirdly, Li et al. (2011) recently con-ducted high-resolution mineral analyses of the2�48 Ga Dales Gorge BIF in Western Australia andreported the detection of an Fe(III)-acetate salt, aswell as nanocrystals of apatite in association withmagnetite; the latter had a crystal chemistry iden-tical to modern biogenic magnetite. This combi-nation of features points to the original presenceof biomass in the BIF sediments, and also indi-cates that the organic carbon served as an electrondonor during bacterial Fe(III) reduction.Crucial to understanding of the biological role
in BIF formation is an assessment of the relationbetween biomass production and the BIF deposi-tional setting. To this end, results of physiologi-cal experiments give a sense of the viable rangeof initial phototrophic productivity in theocean’s photic zone and of precipitated cell-Fe(OH)3 aggregate composition. Investigation of theredox properties and the cycling of elementshave also proven valuable in the reconstructionof the microbial role at the time of BIF deposi-tion. The utilization of H2O by cyanobacteria andthe direct oxidation of Fe(II) by photoferrotrophsare both coupled to CO2 reduction and biomassproduction. Ferric hydroxide particles andorganic biomass could be deposited in eithercase. In the photoferrotroph model, direct Fe(II)oxidation by the cells would lead to a joint sedi-mentation of precipitated Fe(III) and microbiallyproduced biomass. In the cyanobacteria model,the production of O2 by the cells, and the oxida-tion of the Fe(II) by this O2 are independent. Con-sequently, the potential association of the Fe(III)minerals with the cyanobacterial biomassdepends on the local distribution of the cells rel-ative to the location of Fe(III) mineral deposition(open ocean or shore, Fig. 2C).
EVIDENCE FOR ANCIENTCYANOBACTERIA
Microfossils and biological fabrics
The timing of the origins of cyanobacteriaremains highly controversial. For many years,the oldest morphological evidence for cyanobac-teria was believed to have come from weatheredclasts in the 3�45 Ga Apex cherts of the
Warrawoona Group, Western Australia. Struc-tures reminiscent of unbranched, partitionedtrichomes, with dimensions and morphologiesindicative of extant cyanobacteria, led to thesuggestion that the Archean micro-organismswere capable of gliding and, possibly, phototac-tic motility (Schopf, 1993). Moreover, confocallaser scanning microscopy of the filaments andRaman spectroscopy were used to infer the pres-ence of biogenic kerogen in higher concentra-tions than the surrounding matrix (Schopf et al.,2002; Schopf & Kudryavtsev, 2009). Additionalsupport for the early evolution of cyanobacteriacame from the discovery of large spheroidal,sheath-like structures (up to 20 lm in diameter)in cherts from the underlying Towers Formation(Schopf & Packer, 1987), as well as from thepresence of conical and pseudocolumnar stro-matolites in the 3�4 Ga Strelley Pool Chert, Wes-tern Australia (Van Kranendonk, 2006).A re-examination of the Apex chert by Brasier
et al. (2002, 2005), however, called into questionthe biogenicity of the filamentous structures andthe sedimentary origins of the earliest ‘fossilife-rous’ deposits, citing Fischer–Tropsch-type reac-tions associated with sea floor hydrothermalsystems. Buick (1988) has maintained that theactual chert units from which these microfossilsderive are secondary hydrothermal deposits ofmuch younger age that cross-cut the primarybedding. The Warrawoona samples also led toexperiments which demonstrate that kerogen ina microfossiliferous sample does not in itselfindicate biogenicity. Simple organic hydrocar-bons, whose sources are abiological (formalde-hyde), readily condense onto silica-carbonateinorganic filaments and subsequently polymer-ize under gentle heating to yield kerogenousproducts (Garcia-Ruiz et al., 2003). Indeed, thesame kind of Raman spectral signature isobtained from kerogen as stems from many otherpoorly ordered and abiogenic carbonaceousmaterials (Pasteris & Wopenka, 2003). Further-more, a recent study focusing on similar struc-tures from the Apex chert using Ramanspectroscopy showed that the microstructureswere a mixture of hematite and quartz and thatthe matrix did in fact consist of carbonaceousmaterial, but the interpretation of the originalstructures as biogenic was a result of image com-pilation (Marshall et al., 2011). Thus, the discus-sion surrounding the Warrawoona samplesdemonstrates that the presence of carbonaceousmaterial is not sufficient proof of biogenicity.Importantly, it highlights the stringent criteria
Microbiological processes in banded iron formation deposition 1737
needed to determine bona fide biogenicity inancient rocks (for further discussion on thesecriteria, see Schopf et al., 2007). A firmer bench-mark for the presence of cyanobacteria comesfrom the 2�7 Ga stromatolitic assemblages of theTumbiana Formation, Western Australia. Basedon their habitat in a sulphate-deficient evapora-tive lake, it was suggested that the originalmicrobial mat contained cells that were metabo-lized by oxygenic photosynthesis (Buick, 1992).This view is supported by the earliest recog-nized fossil assemblage of filamentous and coc-coid cells colonies, from the 2�6 Ga CampbellGroup, South Africa, which appears to includeoscillatoriacean cyanobacterial genera, such asPhormidium or Lyngbya. These micro-organismscontributed to the formation of stromatoliticreefs in shallow subtidal to intertidal settings(Altermann & Schopf, 1995). Other possibleexamples of ancient cyanobacteria occur inlaminates and oolites of the 2�5 Ga GamohaanFormation, South Africa, where Wright &Altermann (2000) reported dolomicrites formedon the outer margins of the sheaths of filamen-tous cyanobacterial remains. More putativeevidence for cyanobacteria stems from the2�9 Ga Pongola Supergroup in South Africa,where Noffke et al. (2008) observed microbiallyinfluenced sedimentary structures that areconsistent with similar features constructedtoday by benthic cyanobacteria.The link between oxygenic photosynthesis
and stromatolite formation is supported by mod-ern studies which show that the trapping andbinding of carbonate grains by cyanobacterial fil-aments and their extracellular polymers (such asextracellular polymeric substances) are integralto the structure of intertidal stromatolites (Burnset al., 2004, 2009; Bosak et al., 2009). However,a series of studies show how similar structurescan be formed via abiological processes (Grotzin-ger & Rothman, 1996; Grotzinger & Knoll, 1999;McLoughlin et al., 2008). In addition, sequenceanalysis of small sub-unit rRNA genes amplifiedwith PCR from genomic DNA of modern stro-matolite communities of Hamelin Pool, SharkBay, Australia, showed that anoxygenic photo-trophs represent a considerable fraction of thebiomass (Papineau et al., 2005). Along the samelines, Bosak et al. (2007) later demonstrated thatthe anoxygenic phototroph, Rhodopseudomonaspalustris, stimulates the precipitation of calcitein saturated solutions and builds stromatolite-like structures. It has been suggested that beforecyanobacteria evolved, ferrous iron was the
main reductant in ancient oceans; only withdepletion in ferrous iron supply to the oceansdid H2O2 and then water replace ferrous iron asthe electron donor in the cyanobacterial line ofevolution (Olson & Blankenship, 2004). It seemspossible that prior to the rise of cyanobacteria,predecessor anaerobes, such as anoxygenicphototrophs, were dominant stromatolite-build-ing organisms, which could account for the stro-matolitic record that pre-dates strong evidencefor oxygenic phototrophs. Further studies intothe mechanisms of stromatolite formation prom-ise to help parse the dominant processes to usethese structures more accurately in ecosystemreconstruction.
Stable isotope composition
One persuasive argument in favour of early CO2
fixation via photosynthesis, and the potentialpresence of cyanobacteria, comes from thehighly negative carbon isotopic values of kero-gen residues in Archean strata. The negative val-ues are seemingly indicative of life because thetransformation of inorganic carbon (for example,CO2 or HCO3
�) via autotrophic pathways intoorganic carbon involves the preferential incorpo-ration of the lighter isotope, 12C, into the organicmaterial, leaving behind a reservoir enriched inthe heavier isotope, 13C. Consequently, organiccompounds produced by autotrophic pathwaysdisplay a marked preference for the light isotope(for example, d13C in cyanobacteria range from�4 to �35&), whereas the heavy carbon isretained in the surface reservoir of oxidized car-bon, mostly as dissolved bicarbonate, and laterincorporated into precipitated carbonate miner-als, for example, calcite and dolomite, or asatmospheric CO2 (Schidlowski, 2000).Analysis of kerogens in microfossiliferous
units in Archean formations from Western Aus-tralia and South Africa contain negative carbonisotopic values that range from �20 to �35&(Altermann & Kazmierczak, 2003; Tice & Lowe,2004), whereas the organic carbon in early Prote-rozoic cherts similarly averages ca �31�0 � 4�7(Strauss et al., 1992). More recently, a study ofthe C-isotope signatures in the Transvaal Super-group showed the organic d13C to range from�40 to �25& (Fischer et al., 2009). Collectively,these results are an indication of early photosyn-thesis. Unfortunately, a number of carbon fixa-tion pathways (Calvin cycle and the reductiveacetyl-CoA, reductive citric acid and hydroxy-propionate pathways) utilized by phototrophic
bacteria seem to have overlapping degrees ofcarbon isotope fractionation, which hinders con-clusive interpretation.Strong evidence for oxygen in the Archean
oceans comes from extremely isotopicallydepleted kerogens from 2�72 to 2�59 Ga carbo-nates and shales in the Hamersley Province inWestern Australia. Organic carbon 13C valuesrange from as low as �57 to �28& (Eigenbrode& Freeman, 2006). The 13C enrichment has beenascribed to the assimilation of methane by met-hanotrophic bacteria that utilize electronacceptors such as O2, SO4
2� or NO3�; both
sulphate and nitrate require oxygen for theirformation (Hayes, 1983). Potentially, althoughnot yet conclusively demonstrated, methane oxi-dation could be linked to the reduction of Fe(III)(Konhauser et al., 2005). Interestingly, there is a13C enrichment of 10& in kerogen in post-2�7 Ga shallow-water facies relative to the deep-water facies, suggesting that the shallow settingbecame oxygenated, marginalizing the methano-gens and methanotrophs to deeper waters,whereas the shallow waters became dominatedby cyanobacteria (Eigenbrode & Freeman, 2006).
Biomarker identity
An additional tool for the identification of a spe-cific group of organisms in ancient rocks is theanalysis of organic biomarkers. These organiccompounds, derived from more complex precur-sors, retain some resemblance to the originalbiological molecules, even after a long history ofdecomposition and alteration that accompaniesburial and diagenesis. Bitumens from the 2�6 GaMarra Mamba Iron Formation and the 2�5 GaMount McRae Shale of the Hamersley Group,Western Australia, for example yield abundant2a-methylhopanes, derivatives of prominentlipids in cyanobacteria (methyl-bacteriohopane-polyols) that serve to improve cell membranerigidity (Brocks et al., 1999; Summons et al.,1999; Eigenbrode et al., 2008).These compounds were first strictly found in
cyanobacteria and, therefore, were interpreted asdiagnostic of oxygenic photosynthetic organismsin ancient rocks. The belief that they onlyserved a structural function, however, made itdifficult to connect the presence of these com-pounds to a specific metabolism. Genomic data-bases revealed that several facultative andobligate anaerobes possess the appropriate genesfor hopanoid biosynthesis. In fact, Geobactersulfurreducens actually produces a wide variety
of complex hopanoids structurally related to 2a-methylhopane under strictly anoxic conditionsin pure culture (Fischer et al., 2005; H€artneret al., 2005). It has been demonstrated that ananoxygenic Fe(II)-oxidizing phototroph, R. pa-lustris strain TIE-1, generates substantial quanti-ties of 2-methylhopanoids in the absence ofoxygen (Rashby et al., 2007). Utilizing this samestrain, a recent study now shows that C-2methylated hopanoids are preferentially synthe-sized by the R. palustris strain TIE-1 underconditions of elevated Fe(II) concentrations(Eickhoff et al., 2013). Accordingly, their pres-ence in the rock record may also signal anoxicferrous conditions. Although the full physiologi-cal roles of 2-methylhopanoids are still notdefined, it seems that their origins are morediverse than first believed.Hopanes carrying 3-methyl substituents have
also been recovered from 2�72 to 2�56 Ga carbo-nates and shales from the Hamersley Province(Eigenbrode et al., 2008). The only known extantbacteria to produce these particular hopanes areaerobic methanotrophs. Their presence, andstrong correlation with the carbon isotope com-positions in kerogen (as discussed in the previ-ous section), may thus confirm the presence ofoxygen in the photic zone by the late Archean.A third suite of biomarkers, specific steranes of
28 to 30 carbon isomers, are unique alterationproducts of the sterols used in extant eukaryoticcell membranes. The only prokaryotes known tosynthesize sterols have biosynthetic pathwaysleading to different structural isomers. BecauseO2 is required for the biosynthesis of sterols, theirextraction from Archean rocks suggests that atleast some dissolved oxygen (ca 0�002 ml O2 l�1)was present at that time. These steranes werereported in bitumens of the 2�7 Ga shales of theJeerinah Formation, Hamersley Group (Brockset al., 1999). This finding was later questioned asa subsequent study suggested that the sampleswere contaminated by younger fluids (Rasmussenet al., 2008). In recent experiments using theyeast Saccharomyces cerevisiae as a test organ-ism, steroid biosynthesis was observed at dis-solved O2 concentrations ranging from 6�5 lM to7 nM (Waldbauer et al., 2011), suggesting thatvery low O2 concentrations were sufficient forsteroid biosynthesis. There is still no experimen-tal evidence to support the alternative theory thatsteranes may have been produced via an anaero-bic pathway utilizing hydrogen peroxide ororganic peroxides (Raymond & Blankenship,2004; Fischer & Pearson, 2007).
Microbiological processes in banded iron formation deposition 1739
Geochemical composition
At present, there is a temporal gap in the rockrecord with regard to the geochemical evidencefor the evolution of cyanobacteria and the effectsof their metabolism on atmospheric oxygenation.In addition to the features described above thatsupport cyanobacterial evolution ca 2�7 Ga,there are geochemical signatures in variouslithologies that point towards the presence ofshallow oxygenated sea water by the Neoar-chean. Firstly, nitrogen isotopic composition ofkerogens in minimally altered shales from theCampbellrand-Malmani platform in South Africashowed a significant rise in the d15N values by2�67 Ga (Godfrey & Falkowski, 2009). Thisincrease has been proposed as evidence for theonset of nitrification–denitrification reactions inthe surface oceans; importantly, these microbialprocesses require the presence of oxygen.Secondly, a recent study of the trace elementcomposition of those same shales showed a highabundance of Re but a low abundance of Mo,which together with the speciation of sedimen-tary iron, confirms the presence of oxygen in thewater column, arguably to depths of severalhundred metres (Kendall et al., 2010). In con-trast to the above, a number of features suggestthat the atmosphere did not contain significantamounts of oxygen until ca 200 Myr later at ca2�5 Ga, the so-called Great Oxidation Event.These include:1 Detrital uraninite, pyrite and siderite that are
easily oxidized by O2 have been recovered influvial siliciclastic sediments of the 3�2 to 2�7Gyr Pilbara craton (Rasmussen & Buick, 1999).They are not abundant in fluvial systems youn-ger than 2�3 Ga.2 High-resolution chemostratigraphy reveals an
episode of enrichment of the redox-sensitive tran-sition metals molybdenum and rhenium in the2�5 Ga Mount McRae Shale in Western Australia(Anbar et al., 2007). These findings suggest thatthe metals were supplied to the oceans by oxida-tive weathering of crustal sulphide minerals.Similarly, a recent compilation of Cr concentra-tions in BIFs shows an enrichment beginning at2�45 Ga (Frei et al., 2009; Konhauser et al.,2011b). Given the insolubility of Cr minerals, itsmobilization and incorporation into BIFs indi-cates enhanced chemical weathering at that time,likely associated with the evolution of aerobiccontinental pyrite oxidation.3 Iron speciation and sulphur isotope data
from the Mount McRae Shale provide evidence
for euxinic (anoxic and sulphidic) layers in thewater column by 2�5 Ga (Reinhard et al., 2009).These conditions were probably stimulated byan increase in oceanic sulphate concentrationsarising from aerobic continental pyrite oxida-tion. Correspondingly, previous low levels ofmarine sulphate suggest minimal oxidativeweathering of sulphide minerals (Canfield et al.,2000).4 Mass-independent fractionation (MIF) of sul-phur isotopes (Δ33S and Δ36S) in sulphide andsulphate-containing rocks deposited prior to2�45 Ga, but not after 2�32 Ga. This observationindicates that the sulphur cycle changed fromone governed by gas-phase photochemical reac-tions in an O2-deficient atmosphere to onedominated by oxidative weathering, where thedifferent sulphur species lost their MIF signaldue to microbial SO4
2� reduction (Farquharet al., 2000; Mojzsis et al., 2003; Bekker et al.,2004). To preserve the MIF in Archean andearly Palaeoproterozoic sediments, the atmos-pheric oxygen concentration must have been<10�5 present atmospheric levels (PAL). Bycontrast, in atmospheres with O2 concentrations>10�5 PAL, reduced sulphur-bearing specieswould typically have been oxidized to sulphatebefore becoming incorporated into the sedi-ment, so any MIF signature would have beenlost (Pavlov & Kasting, 2002). A more recentand comparative study utilizing investigationsof the Mount McRae Shale and correlating it tothe Gamohaan and Kuruman iron formations ofSouth Africa dates the progressive rise of oxy-gen at 2�5 Ga and implies that thisphenomenon was certainly on a widespread, ifnot global, scale (Kaufman et al., 2007).
EVIDENCE FOR PHOTOFERROTROPHS
Given that pre-2�7 Ga BIFs probably wereprecipitated from anoxic sea water, and thevarious uncertainties regarding the presence ofcyanobacteria in the rock record, it is relevant toconsider BIF precipitation mechanisms based onthe direct biological oxidation of Fe(II) viaanoxygenic photosynthesis (Cloud, 1965, 1973;Garrels et al., 1973; Hartman, 1984). Althoughthere is still no actual physical or chemicalevidence for the existence of Fe(II)-oxidizingphototrophs in the Archean, six independentlines do suggest their presence on the earlyEarth.
Of the seven known strains of anoxygenic Fe(II)-oxidizing phototrophs known to date, six ofthem have been classified as Proteobacteria; theseventh is a green sulphur bacterium (Table 1).The Proteobacteria are a large and diverse phylaof bacteria consisting of five major classes; alpha[including Fe(II)-oxidizing purple non-sulphurbacteria], beta, gamma [including Fe(II)-oxidiz-ing purple sulphur bacteria], delta and epsilon,all of which may have diversified from oneancestral phototroph (Woese, 1987). Purple sul-phur and purple non-sulphur bacteria use bacte-riochlorophyll (Bchl a or b) to absorb light inthe near-infrared (800 to 1040 nm). Althoughcontroversy has arisen from phylogenetic studiesof anoxygenic and oxygenic photosynthesis dueto partial gene loss and fast lateral transfer ofBchl biosynthesis genes, molecular phylogeneticanalyses of a number of enzymes involved inBchl biosynthesis suggests that anoxygenic pho-tosynthetic lineages are almost certain to bemore deeply rooted than the oxygenic cyanobac-terial lineages (Xiong, 2006).
Ecophysiological studies with extant speciesof anoxygenic Fe(II)-oxidizing phototrophs
Although the plausibility of an anoxygenic Fe(II)-oxidizing metabolism had long been speculatedbased on thermodynamic considerations (Garrelset al., 1973; Hartman, 1984), such organismswere discovered more recently and subsequentlyisolated (Widdel et al., 1993). A number ofexperimental studies have since confirmed thatvarious purple and green phototrophic bacteria,both fresh water and marine (Table 1), can useFe(II) as a reductant for CO2 fixation [reaction 2](Widdel et al., 1993; Heising et al., 1999; Straub
et al., 1999). This fact has made it possible tostudy these organisms as potential modernanalogues of ancient organisms.
4Fe2þþHCO�3 þ10H2O!4FeðOHÞ3þðCH2OÞþ7Hþ
ð2ÞAnoxygenic Fe(II) oxidizing phototrophs are
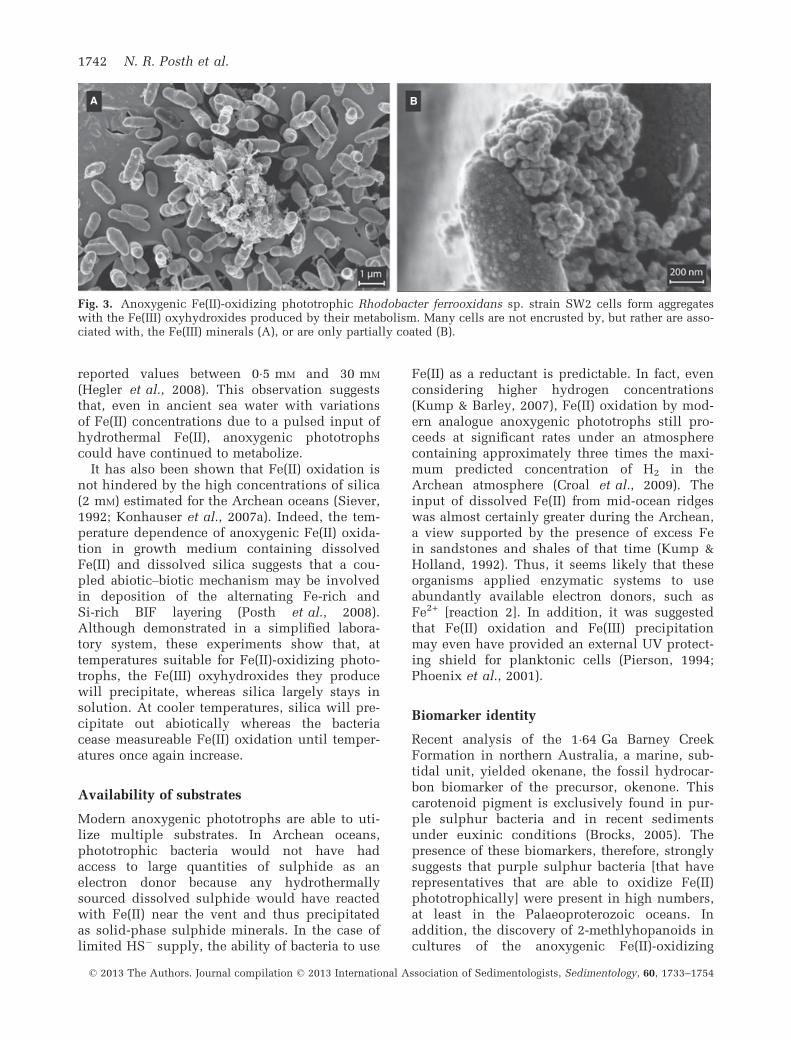
diverse and broadly distributed in the environ-ment (Table 1). None of these organisms areunique in form as all are rod-shaped (Fig. 3);however, morphological variety is apparent, forexample, the presence of vacuoles in Thiodict-yon sp. strain F4 (Croal et al., 2004). Further-more, these strains are capable of utilizing avariety of substrates, such as acetate, H2 andeven FeS, and were shown to oxidize Fe(II) evenin the presence of H2 (Croal et al., 2009). Thisdiversity suggests that these organisms have thepotential to be ubiquitous in both ancient andmodern anoxic, Fe-rich environments.Importantly, although the Fe(II) oxidation rate
of these strains is dependent on light intensity,anoxygenic phototrophs are capable of oxidizingFe(II) in low light regimes befitting the photiczone of ocean water (Kappler et al., 2005; Hegleret al., 2008). Considering the highly efficientlight harvesting mechanism employed by anoxy-genic phototrophs, it was estimated that, even inthe presence of an overlying layer of cyanobacte-ria, sufficient light for the metabolism of anoxy-genic phototrophs could penetrate up to 100 mocean depth (Kappler et al., 2005). Similar lightharvesting has been seen in the Black Sea, withanoxygenic phototrophs detected at a depth of80 to 100 m (Overmann et al., 1992; Manskeet al., 2005). These strains also continue to carryout the oxidation of Fe(II) in the presence of awide range of Fe(II) concentrations, with
Table 1. The anoxygenic Fe(II)-oxidizing phototrophs isolated to date include both marine and fresh waterstrains.
Phylum Strain Setting/location
a-Proteobacteria(purple non-sulphur)
Rhodobacter ferrooxidans sp. strain SW2 Fresh water ponds, Hannover Germany1
Microbiological processes in banded iron formation deposition 1741
reported values between 0�5 mM and 30 mM
(Hegler et al., 2008). This observation suggeststhat, even in ancient sea water with variationsof Fe(II) concentrations due to a pulsed input ofhydrothermal Fe(II), anoxygenic phototrophscould have continued to metabolize.It has also been shown that Fe(II) oxidation is
not hindered by the high concentrations of silica(2 mM) estimated for the Archean oceans (Siever,1992; Konhauser et al., 2007a). Indeed, the tem-perature dependence of anoxygenic Fe(II) oxida-tion in growth medium containing dissolvedFe(II) and dissolved silica suggests that a cou-pled abiotic–biotic mechanism may be involvedin deposition of the alternating Fe-rich andSi-rich BIF layering (Posth et al., 2008).Although demonstrated in a simplified labora-tory system, these experiments show that, attemperatures suitable for Fe(II)-oxidizing photo-trophs, the Fe(III) oxyhydroxides they producewill precipitate, whereas silica largely stays insolution. At cooler temperatures, silica will pre-cipitate out abiotically whereas the bacteriacease measureable Fe(II) oxidation until temper-atures once again increase.
Availability of substrates
Modern anoxygenic phototrophs are able to uti-lize multiple substrates. In Archean oceans,phototrophic bacteria would not have hadaccess to large quantities of sulphide as anelectron donor because any hydrothermallysourced dissolved sulphide would have reactedwith Fe(II) near the vent and thus precipitatedas solid-phase sulphide minerals. In the case oflimited HS� supply, the ability of bacteria to use
Fe(II) as a reductant is predictable. In fact, evenconsidering higher hydrogen concentrations(Kump & Barley, 2007), Fe(II) oxidation by mod-ern analogue anoxygenic phototrophs still pro-ceeds at significant rates under an atmospherecontaining approximately three times the maxi-mum predicted concentration of H2 in theArchean atmosphere (Croal et al., 2009). Theinput of dissolved Fe(II) from mid-ocean ridgeswas almost certainly greater during the Archean,a view supported by the presence of excess Fein sandstones and shales of that time (Kump &Holland, 1992). Thus, it seems likely that theseorganisms applied enzymatic systems to useabundantly available electron donors, such asFe2+ [reaction 2]. In addition, it was suggestedthat Fe(II) oxidation and Fe(III) precipitationmay even have provided an external UV protect-ing shield for planktonic cells (Pierson, 1994;Phoenix et al., 2001).
Biomarker identity
Recent analysis of the 1�64 Ga Barney CreekFormation in northern Australia, a marine, sub-tidal unit, yielded okenane, the fossil hydrocar-bon biomarker of the precursor, okenone. Thiscarotenoid pigment is exclusively found in pur-ple sulphur bacteria and in recent sedimentsunder euxinic conditions (Brocks, 2005). Thepresence of these biomarkers, therefore, stronglysuggests that purple sulphur bacteria [that haverepresentatives that are able to oxidize Fe(II)phototrophically] were present in high numbers,at least in the Palaeoproterozoic oceans. Inaddition, the discovery of 2-methlyhopanoids incultures of the anoxygenic Fe(II)-oxidizing
A B
Fig. 3. Anoxygenic Fe(II)-oxidizing phototrophic Rhodobacter ferrooxidans sp. strain SW2 cells form aggregateswith the Fe(III) oxyhydroxides produced by their metabolism. Many cells are not encrusted by, but rather are asso-ciated with, the Fe(III) minerals (A), or are only partially coated (B).
phototroph, R. palustris strain TIE-1 stimulatedby environmental Fe(II) in the absence of oxygen(Rashby et al., 2007) demonstrates the utility ofthis biomarker in reconstructing anoxygenicphototrophy in ancient sediment.
Stable isotope composition
Large variations of d56Fe (�2�5 to 1�0& relativeto the bulk Earth) in late Archean and early Pal-aeoproterozoic BIFs from the Transvaal Super-group, South Africa, incorporate the entire rangeof values measured on Earth and are interpretedto highlight the role of fluid-mineral isotopefractionation and, possibly, microbial processes(Johnson et al., 2003). Although Fe isotopevariations have the potential to define ancientsedimentary environments and could help dis-tinguish between biotic and abiotic processes, aswell as whether post-depositional processes mayhave altered the Fe redox state in BIFs, the com-plexity of the depositional environments andinfiltration of external fluids during diagenesisand/or low temperature metamorphism mayerase the primary isotope record (Hoefs, 1997;Johnson et al., 2003). Yet, the Fe isotope compo-sition of iron minerals in BIFs are markedly dif-ferent from the homogenous values seen inigneous rocks and many modern marine sedi-ments (d56Fe 0�00 � 0�05&) and the Fe isotopecompositions of the major minerals found inBIFs (hematite, magnetite, Fe-carbonate and pyr-ite) reflect equilibrium fractionation of minerals,a variation in the fluid isotope composition fromwhich the minerals precipitated, and microbialprocesses (Johnson et al., 2003, 2004; Johnson &Beard, 2005; Steinhofel et al., 2010).A recent study focused on the rare earth
element and Fe isotope archive in the iron andmicrofossil-rich stromatolites of the 1�89 GaGunflint and Biwabik iron formations (USA)with the aim of determining the dominantmicrobes involved in Fe(II) oxidation (Planavskyet al., 2009). These authors point to the predom-inantly positive d56Fe values in this formation(ranging from �0�66 to +0�82&) as evidence ofFe(II)-oxidizing bacteria, whereas Ce anomaliesin these same formations suggest low oxygenconditions during deposition. As a comparison,laboratory experiments designed to define isoto-pic fractionation by anoxygenic Fe(II)-oxidizingphototrophs in pure culture, as well as enrich-ment cultures, show that ferric hydroxides areenriched in the heavy isotope by 1�5 � 0�2&relative to Fe(II) (Croal et al., 2004). This frac-
tionation was independent of the rate of theFe(II) oxidation by these organisms. HeavierFe(III) oxyhydroxides were also shown to beproduced by microbially catalysed aerobic Fe(II)oxidation at low pH (Balci et al., 2006), nitrate-reducing anaerobic Fe(II) oxidizers (Kappleret al., 2010), aerobic chemical oxidation (Bullenet al., 2001) and UV Fe(II) oxidation (Stratonet al., 2006), suggesting that all major processesproduce similar Fe isotope fractionations. Onthe one hand, the isotopic fractionationsreported from modern experiments correlatewell with values measured in Archean to earlyProterozoic BIFs from the Transvaal Supergroup,South Africa; they have 56Fe/54Fe values inhematite as high as +0�75 to +1�0& (Johnsonet al., 2003) compared with the predicted valuesfor Fe effused from hydrothermal vents (�0&).Consequently, in the case of anoxic Archeanoceans, the positive fractionation appears to beconsistent with (although not proof of) photo-trophic Fe(II) oxidation in the Archean oceans.On the other hand, the similarity in values fromanoxygenic phototrophs with those of Fe(III) oxy-hydroxides formed by chemical oxidation makesit difficult to distinguish between biotic and abi-otic processes (Bullen et al., 2001). Iron isotopefractionation during the oxidation itself is obvi-ously independent of the oxidation mechanism(chemical or biological) because of the very rapidisotopic exchange between aqueous Fe2+ andFe3+. Therefore, isotopic equilibrium will bereached between reduced and oxidized aqueousspecies.
Plausibility of anoxygenic photoferrotrophybased on Fe and C budget
Ecophysiological studies of these modern strainsof anoxygenic Fe(II)-oxidizing phototrophs havebeen utilized to test their plausibility as a BIFdepositional mechanism, with specific regard tocarbon and Fe(III) oxyhydroxide productivity.For example, it has been suggested that commu-nities of Fe(II)-oxidizing phototrophs in theearly Archean could have generated up to1�9 9 1013 mol C year�1 throughout all oceans(Kharecha et al., 2005). To put this number intocontext, Canfield (2005) has estimated that thetotal net primary productivity in the lateArchean–Palaeoproterozoic oceans was 1�8 to5�6 9 1014 mol C year�1.By modelling the photosynthetic Fe(II) oxida-
tion rates, it has been suggested that such micro-organisms could have accounted for all of the
Microbiological processes in banded iron formation deposition 1743
Fe(III) initially deposited in primary BIFsediment (Konhauser et al., 2002; Kappler &Newman, 2004; Kappler et al., 2005). Due to thepigments they utilize (Bchl and carotenoids), an-oxygenic phototrophs absorb light over specificwavelengths (Bchla 800 to 880 nm, Bchlb1020 nm, Bchlc ca 750 nm, carotenoids 360 to517 nm), many of which (>300 nm and<600 nm) would be filtered out in the first fewmetres of ocean water. By filtering out the wave-lengths >650 nm, experimental studies haveshown that the oxidation rate of these strainswas reduced to 20% of that of the full spectrum,yet that rate still allowed for the oxidation of0�08 mm Fe(II) per day. Based on this reductionin oxidation efficiency due to light filtering, theaverage cell density in a planktonic population,assumptions of Archean ocean Fe(II) concentra-tions (ca 0�5 mM; Holland, 1973; Morris, 1993)and the flux of dissolved Fe(II) from the deepoceans, a conservative estimate for the layerthickness of a planktonic population of anoxy-genic Fe(II)-oxidizing phototrophs required tooxidize all dissolved Fe(II) present was 17�6 m(Kappler et al., 2005). With this information, atheoretical amount of Fe(III) oxyhydroxide pre-cipitated by planktonic anoxygenic phototrophswas estimated to be ca 9�0 9 1012 mol Fe yr�1
for large basins such the Hamersley in WesternAustralia (which is ca 1011 m2); this value isconsistent with calculations based on maximumBIF deposition rates of 1 mm yr�1 where theamount of iron precipitated annually is ca4�5 9 1012 mol Fe yr�1 (Konhauser et al., 2002).Considering the parameters for a model of Fe(II)oxidation rate, light intensity, substrate concen-tration and cell numbers, even with reducedefficiency in Fe(II) oxidation at conservative val-ues of nutrients and light, anoxygenic photo-trophs had the potential to play a significantrole in BIF deposition.
ROLE OF CELLULAR CARBON INPOST-DEPOSITIONAL ALTERATION
If a direct biological mechanism was importantin the initial process of Fe(II) oxidation in anArchean ocean water column, it is thenexpected that biomass settled to the sea flooralong with the Fe(III) minerals (Konhauseret al., 2011a). Yet, BIFs contain very littleorganic carbon, meaning that the process waseither fully abiotic, or biomass was cycled inthe water column or bottom sediment via the
combined metabolic processes of fermentationand chemoheterotrophy (Konhauser et al.,2005). In the case of the latter, any buriedorganic carbon would be oxidized and, impor-tantly, any cellular biomass would bedestroyed, i.e. no microfossils would be pre-served in the Fe-rich layers.If, as today, the organic carbon was oxidized
during burial by either diagenesis or metamor-phism, the relevant question is which terminalelectron acceptor was present at the sea floorduring times of BIF deposition, and at what con-centrations? Despite the possibility of a surfacewater oxic zone generated by cyanobacterialactivity by 2�7 Ga (see discussion above), deepwaters remained anoxic (Canfield, 1998). In theabsence of O2, the fermentation products in thebottom waters and/or shallow sediments wouldhave been oxidized via some other form ofanaerobic respiratory process (Rothman et al.,2003). In terms of such pathways, the paucity ofO2 would have meant minimal nitrate and sul-phate availability; the latter being evident fromnegligible sulphur isotopic fractionationsbetween sulphide and sulphate minerals duringthe Archean (Strauss, 2003), and the absence ofpyrite in BIFs, except in association with inter-layered shaley units (Ewers & Morris, 1981). Thesupply of MnO2 was probably also not signifi-cant because the concentration of Mn(II)released in hydrothermal effluent is up to fivetimes lower than that of iron (Campbell et al.,1988). Furthermore, there are presently noknown phototrophic Mn(II)-oxidizing bacteriathat would allow the formation of significantamounts of Mn(IV) oxides in the absence of O2.By contrast, however, there was abundant Fe(III)oxyhydroxide deposited as BIFs, and given thepresence of partially reduced phases such asmagnetite and siderite, a microbial process cou-pling the oxidation of organic carbon to thereduction of ferric iron producing such reducediron mineral phases seems very likely (Nealson& Myers, 1990).Supporting evidence for an ancient Fe(III)
reduction pathway comes from the experimentalobservations that many deeply branchingArchaea (i.e. some of the oldest purportedspecies) are capable of using H2 to reduce Fe(III)to support chemolithoautotrophic growth(Vargas et al., 1998). These organisms can evenuse quinone moieties as electron shuttlesbetween solid-phase ferric iron-containing min-erals and H2, thereby alleviating the need fordirect contact between the cell and mineral
surface (Lovley et al., 2000). Moreover, Fe(III)reduction has been shown to be broadly distrib-uted among several known Proteobacteria gen-era, suggesting that this form of metabolismbecame widespread over the course of evolution(Barns & Nierzwicki-Bauer, 1997).Evidence of ancient microbial Fe(III) reduction
comes from Fe isotopic ratios in Fe-bearingminerals from Archean sedimentary rocks thatclosely mimic those observed during moderndissimilatory Fe(III) reduction. For instance,analyses of magnetite-rich rocks from the 2�9 GaRietkuil Formation, Witwatersrand Supergroup,South Africa, revealed d56Fe values as low as�2�3& (Yamaguchi et al., 2005), whereas side-rite in the 2�5 Ga Dales Gorge Member, BrockmanIron Formation, Western Australia, had d56Fevalues as low as �2�3& (Steinhofel et al., 2010).These negative fractionations are very similar towhat might be expected from multiple stages ofFe(III) reduction (Johnson et al., 2004), whereeach single step during bacterial Fe(III) reductionleads to isotopically light (d56Fe < 1�2&) aque-ous Fe(II) relative to the initial ferric hydroxidesubstrate (Icopini et al., 2004; Crosby et al.,2007). The importance of Fe(III) reduction iniron fractionation has, however, been challengedby Rouxel et al. (2005). These authors suggestedthat highly variable, but negative, values in py-rite from black shales (0�5 to �3�5&) between2�8 Ga and 2�3 Ga is more likely to reflect theinitial deposition of Fe(III) oxides (for example,BIFs) that preferentially removed isotopicallyheavy 56Fe, driving the ocean waters to the nega-tive d56Fe values recorded in pyrite. This inter-pretation is consistent with the notion thatpartial biological and abiological processes oxi-dized the dissolved Fe(II) brought into the shal-low waters, but is problematic in light of recentstudies showing significant isotopic variationsfor minerals within close proximity, and thustime of deposition (Johnson et al., 2008). Ashighlighted by Johnson et al. (2008), the mostplausible way of explaining this variation is ifthe Fe isotopes reflect diagenetic pathways, andnot the Fe composition of the bulk sea water.The fate of cell biomass (organic carbon)
deposited with Fe(III) minerals has not yet beenshown experimentally. Theoretically, the rela-tion between Fe(III) and Corg in the primary sed-iment would be stoichiometric if the Fe(II) wasoxidized to Fe(III) at the location where biomasswas produced, and if the Corg and Fe(III) mine-rals were deposited together, such as in theanoxygenic photoferrotroph model. In this sce-
nario, Fe(III) could be completely re-reduced toFe(II). This could be the case involving anoxy-genic photoferrotrophs. However, from experi-mental studies it is known that some planktoniccells remain in the water column (Konhauseret al., 2005; Miot et al., 2009; Posth et al., 2010)and only a portion of the cells form aggregateswith the Fe(III) hydroxide (Fig. 3), which thensettle to the sea floor. The reduction of theFe(III) mineral and the formation of secondaryiron minerals, such as magnetite or siderite,would follow. In such a case, Fe(II) in BIFs canbe used as a proxy for deposited Corg if theFe(II) formed by Fe(III) reduction remains in theBIFs.In contrast to the anoxygenic phototroph
model, if the dissolved Fe(II) was oxidized byan abiotic mechanism, either via the UV-photo-oxidation mechanism or chemical oxidation bycyanobacterially generated O2 at some distancefrom the cells (Fig. 2A and B), then the Fe(III)hydroxides produced would be spatially sepa-rated from the biomass. This Fe(III) couldfurther react with excess dissolved Fe(II) in thewater column forming magnetite [reaction 3],while the cells could be transported away fromthe shore (Fig. 2B) or be continuously present inthe open ocean (Fig. 2C).
Fe2þ þ 2FeðOHÞ3 þ 2OH� ! Fe3O4 þ 4H2O ð3Þ
As was described for the photoferrotrophs, thisscenario could also lead to a partial associationof biomass with Fe(III). In this regard, the pres-ence of cyanobacteria or of photoferrotrophswould be difficult to distinguish based on min-eralogical or iron isotope studies, but possiblywith the identification of metabolism-specificbiomarkers.To estimate the likelihood and plausible
impact of the anoxygenic phototroph scenario,Konhauser et al. (2005) modelled the Archeanmarine Fe cycle by making two assumptions.Firstly, it was assumed that the bulk of theFe(II) component in Fe-rich BIF-type macro-bands formed diagenetically through biologicalFe(III) reduction, i.e. the magnetite is not pri-mary. Based on a predicted rate of Fe(III) deposi-tion annually (1 mM yr�1), Konhauser et al.(2005) then quantified how many electrons wereneeded to generate that amount of magnetitereported in BIFs (one-third of the ferric oxideminerals; Morris, 1993). Secondly, these authorsquantified the amount of photosynthetic Fe(II)-
Microbiological processes in banded iron formation deposition 1745
oxidizer biomass that may have been generatedin the photic zone of the water column (basedon Kappler et al., 2005) to estimate the amountof Fe recycled prior to burial. The results dem-onstrated that, under ideal growth conditions, asmuch as 70% of the biologically formed Fe(III)could have been recycled back into the watercolumn via fermentation and organic carbon oxi-dation coupled to microbial Fe(III) reduction.Konhauser et al. (2005) also suggested that someof the biomass may have been ultimately con-sumed via methanogenesis, i.e. coupling the oxi-dation of acetate or H2 to methane formation.That hypothesis is to some extent corroboratedby the analyses of kerogens (extracted fromrocks 2�8 Ga and 2�6 Ga) with highly negatived13C signatures (between �40& and �60&) thatpossibly formed as the result of methanogenic12C-rich gas production, the incorporation ofthis methane into the biomass of methanotroph-ic bacteria and inevitably the preservation of12C-enriched organic matter (Hayes, 1983; Ei-genbrode & Freeman, 2006).Recent laboratory experiments by the present
authors have focused on the influence of tem-perature (170°C) and pressure (1�2 kbar) on bio-genic Fe(III) minerals associated with biomassby utilizing chemically synthesized ferrihydrite(as a proxy for biogenic ferric hydroxide) andglucose (as a proxy for biomass) mixtures inlieu of biogenic minerals. When chemicallysynthesized ferrihydrite was treated alone, itwas transformed to hematite via dehydration.Mixtures of ferrihydrite and glucose treatedunder the same temperature and pressure con-ditions for just a few days produced not onlyhematite, but also magnetite and siderite. Thepresence of a cell biomass proxy in lowamounts resulted in the production of hematiteand small amounts of magnetite and siderite.When the amount of initial biomass proxy wasincreased, considerable quantities of magnetiteand siderite formed while hematite was nolonger detectable. These experiments demon-strate that the amount of biomass deposited tothe ocean floor in association with the Fe(III)minerals controls the type of Fe minerals pre-served in BIFs over geological time frames atelevated temperature and pressure. This showsthat the key minerals found in BIF depositstoday can be produced, but that there is also adifference in mineral transformation in abioticand biotic systems (Walker, 1984; Johnsonet al., 2003, 2008; Konhauser et al., 2005; Koeh-ler et al., 2013).
BIOLOGICALLY GENERATEDSEDIMENTOLOGICAL FEATURES
As described above, many BIFs comprise layerscontaining magnetite and/or hematite, whichalternate on the scale of several millimetres andharbour bands of microcrystalline silica. Onaverage, the microbands range from 0�2 to1�6 mm in thickness, whereas the total Fecontent of the microbands is inversely related tothe microband thickness; varying from 30% Fewhen the microband is 0�3 mm to 5% when thethickness is 1�5 mm (Trendall & Blockley, 1970).This correlation indicates that the iron contentin microbands is constant, whereas variations inthe silica content control both the layer thick-ness and relative iron content (Trendall, 2002).Well-banded iron formations, the typical BIFs,are mostly restricted to Archean and earlyPalaeoproterozoic sequences (for example, theTransvaal Supergroup, South Africa; Fig. 4Aand B). By contrast, large portions of late Palaeo-proterozoic iron formations from the Superiorand Slave cratons in North America are com-prised of sand-sized grains that commonly arecross-bedded and lack the finely laminated tex-tures of BIFs; these shallow-water deposits aregenerally referred to as granular iron formations(GIFs). These GIFs are typically intercalatedwith well-laminated BIFs, Fe-rich mudstones,mafic and felsic volcanic rocks, carbonate andsandstone (for example, the Rapitan Formation,Canada; Fig. 4C and D).The origin of the banding in BIFs remains
enigmatic, with two potential end member pro-cesses being responsible for their presence. Thefirst assumes that the banding is a primary fea-ture caused by episodic precipitation of the ironlayers. The second assumes that the bandingwas caused by some combination of secondaryphysical and post-depositional processes. In thecase of the former, this can include, for example,BIF precursor sediment being transported to theBIF depositional basin via physical processes,such as density currents (Krape�z et al., 2003). Inthe case of the latter, both allochemical andauthigenic BIF sediment would then be subjectto chemical and mineralogical transformationsarising during burial. The banding thus resultswhen either the Fe(III) was reduced to Fe(II) andthe latter diffused out of the sediments, or thesilica layers were dissolved and reprecipitated.Although a combination of these processes islikely to have arisen, there still exists a lack ofconsensus as to which processes were more
important, and how these processes may havevaried from one BIF depositional basin toanother.
Primary banding
Based on microbanding in the 2�48 Ga DalesGorge Member of Western Australia, that waspresumably correlated over 100 km (Trendall &Blockley, 1970; Ewers & Morris, 1981), it wasproposed that deposition must have been uni-form across the basin. Moreover, given the rela-tively regular spacing between microbandswithin some larger mesobands, Trendall &Blockley (1970) further proposed that each ironand silica-rich microband couplet resulted fromone year of chemical precipitation. The term‘varving’ is often used to describe these cou-plets. It should, however, be pointed out thatthe iron-rich and silica-rich microbandsthemselves are often comprised of multiple
internal laminae, which possibly reflects someform of seasonal variations in terms of deposi-tion. For instance, Morris (1993) suggested thatthe internal complexity may follow four mainstages: (i) during the summer, iron was precipi-tated in the photic zone, producing the iron-richlaminae; (ii) the now depleted photic zone wasreplenished several times from the subsurfaceby storm mixing, leading to additional iron-richlaminae; (iii) in the winter, fewer storms meantless Fe replenishment, while cooling led to sil-ica precipitation from sea water that was alreadysaturated with respect to amorphous silica; and(iv) with the return of the summer, iron wasonce again precipitated.The link between BIF deposition and seaso-
nality was later advanced by Posth et al. (2008)who demonstrated how temperature fluctuationsin the surface oceans could link the biotic pre-cipitation of Fe(III) hydroxides by anoxygenicFe(II)-oxidizing phototrophic bacteria to the
A B
C D
Fig. 4. Thin sections of BIF fromthe Mamatwan Manganese Mine,Hotazel, Northern Cape Province,South Africa, show jasper andmagnetite microbanding. Thisexample demonstrates the variationfrom thin, alternating lensstructures (A) at 83�4 m depth towell-developed, uninterrupted andalternating fine layers (B) at 87�5 mdepth. In contrast, thin sectionsfrom the Rapitan Formation,Canada GIF depict the typicalsand-sized grains anddiscontinuous, uneven layeringthat suggests stormwave or currentinfluence (C) and (D).
Microbiological processes in banded iron formation deposition 1747
abiotic precipitation of silica. The precipitationof ferric oxyhydroxides by these bacteria islinked directly to their physiological tempera-ture dependence; within a range, these organ-isms can precipitate more or less ferricoxyhydroxides. For instance, the bacteria grewmost profusely in the temperature range of 20 to25°C, with a decrease in growth rates at lowertemperatures and a strict upper temperaturelimit (Posth et al., 2008). By contrast, silica pre-cipitation is abiotically induced by loweringtemperature, and increased solubilization ofsolid-phase silica at higher temperatures. Fur-thermore, the extent of temperature fluctuations,for which these processes were demonstratedwith model organisms, are within the rangeexperienced in the world oceans today over ayearly cycle, meaning that temperature couldhave been a unifying trigger for Fe and Sibanding.Other depositional models for the Dales Gorge
Member advocate that BIF deposition took placeon the ocean basin floor, beyond the slope envi-ronment (Krape�z et al., 2003). In this regard, itremains unclear what, if any, role microbes mayhave played in the initial Fe(III) hydroxide pre-cipitation. The model proposes that the Fe(III)-rich sediments accumulated during periods ofrising and high sea-level, and presumably weresourced from a submarine hydrothermal systemand transported to the BIF depositional basinvia deep-sea density currents. The chert layers,by contrast, represent in situ silicification of theexisting sediment. The chert defines the top ofevery depositional sequence by sharp erodedcontacts with overlying dolostone or mudrockand gradational contacts into underlying BIFs.The chert units also contain multiple shalerelicts, 5 to 10 cm thick, which indicates thatambient suspension sediments during sedimentstarvation (i.e. a drop in sea-level) were notBIFs, and that BIF deposition ceased prior to theend of depositional cycles. Similar features arenot only preserved throughout all of the Hamers-ley Province, but they also have been docu-mented in the Transvaal Province (Beukes &Gutzmer, 2008); it appears to be the commonarchitecture of all deep-water BIFs (Bekkeret al., 2010).
Diagenetic banding
A number of studies have suggested that thefine-scale banding in BIFs is a secondary featurederived during burial diagenesis. One of the first
such studies was based largely on the observa-tion of compressed lateral terminations of chertpods (Trendall & Blockley, 1970). These authorsdemonstrated that the compressed areas areenriched in iron minerals at the expense mainlyof silica and, using this evidence, concludedthat all high iron bands, podded or not, repre-sented compression of the primary BIF sedimentwith vertical escape of silica. The remobilizedsilica then reprecipitated evenly into overlyingpods that are rich in silica. Along similar lines,Simonson (1987) proposed that the chert podswere concretions that formed early via localcementation, followed by differential compac-tion of the surrounding, less indurated sedi-ments during burial.More recently, and based instead on facies
and sequence-stratigraphic analyses, Krape�zet al. (2003) and Pickard et al. (2004) concludedthat all chert in BIFs is diagenetic in origin.Moreover, these authors concluded that chertmesobands are siliceous equivalents of modern-day sea floor hardgrounds, in which silicareplaced precursor sediment at or below the sed-iment–water interface. Three-dimensional andmicro-scale lenticularity of chert and relics ofprecursor sediment within lamina sets and dis-continuous bands, as well as erosion surfaces onbedded cherts, show that chert has a replace-ment origin and formed during early diagenesis,prior to compaction. As discussed above, theiron layers, by contrast, represent re-suspendedFe(III) hydroxide-rich sediments that werecarried into the depositional basin via densitycurrents and then subject to post-depositionalprocesses (Krape�z et al., 2003).Other studies have put forward the possibility
that it was the iron that was selectively mobi-lized during diagenesis. Based on the earlier dis-covery that some anaerobic heterotrophs couldgrow by coupling the oxidation of organic matterto the reduction of Fe(III) hydroxides (Lovley &Phillips, 1988), Nealson & Myers (1990) subse-quently proposed that such bacteria could pro-vide a mechanism by which iron-poor layerscould be generated during diagenesis; the iron-rich layers represent sediment buried in whichmolar Fe : C ratios exceeded unity and all thebiomass was oxidized. Coupling the reductionof Fe(III) minerals to the oxidation of organicmatter not only explains the low content oforganic carbon in the BIFs via the consumptionof carbon (<0�5%; Gole & Klein, 1981), but alsoexplains the abundance of light carbon isotopicsignatures associated with the interlayered
carbonate minerals (Perry et al., 1973; Walker,1984; Baur et al., 1985).
CONCLUSIONS
Understanding the mechanism(s) underpinningbanded iron formation (BIF) deposition wouldoffer significant insights into some of the biogeo-chemical processes that took place on the earlyEarth. The first suggestions of a microbial rolewere postulated over 40 years ago (Cloud, 1965).At present, there is still indirect evidence of amicrobial mechanism driven by either anoxygen-ic Fe(II)-oxidizing photosynthesis or cyanobacte-rially driven O2 production. Moreover, thecomplexity of BIF deposition, on temporal andspatial scales, makes it possible that multiplemechanisms aided BIF deposition, dependingon the ocean and atmospheric redox state, aswell as on the local geochemical and microbio-logical conditions. One could imagine a scenariowith anoxygenic phototrophs being mainlyresponsible for the deposition of the BIFs in lowO2 oceans prior to the Great Oxidation Event(GOE). After the emergence of cyanobacteria, ithas been suggested that O2 producing cyanobac-teria could have helped drive the abiotic oxida-tion of Fe(II), whereas anoxygenic phototrophscarried out Fe(II) oxidation in an anoxic niche(Kappler et al., 2005). In order to understandBIFs and the biogeosphere in which they weredeposited, as well as to be able to interpret themas an archive of Earth history, it is necessary totake a holistic approach – integrating new meth-ods for biomarker and stable isotope analyses,and also keeping this in context with mineralo-gical examinations and ecophysiological studieswith modern analogue micro-organisms.
ACKNOWLEDGEMENTS
We thank Nicolas Beukes for his advice onSouth African BIFs, as well as his guidance insample collection. Gert van der Linde of theHotazel Manganese Mines, Hotazel, SouthAfrica, provided BIF samples. We also thankClaus Burkhardt and Sebastian Sch€adler for thepreparation of SEM images, which was carriedout at the Natural and Medical Sciences Insti-tute at the University of Tuebingen (NMI). Thiswork was supported by an Emmy-Noether fel-lowship and a research grant from the GermanResearch Foundation (DFG) made to AK (KA
1736/2-1, 2-2 and 4-1), a research grant from theGerman Research Foundation (DFG) made toN.P. (PO-1624/1-1, 2-1), and a Natural Sciencesand Engineering Research Council award toKOK. We thank Clark Johnson, Phil Fralick,Nora Noffke and an anonymous reviewer fortheir helpful comments, which greatly improvedthe quality of the manuscript.
REFERENCES
Altermann, W. and Kazmierczak, J. (2003) Archean
microfossils: a reappraisal of early life on Earth. Res.
Microbiol., 154, 611–617.Altermann, W. and Schopf, J.W. (1995) Microfossils from the
Neoarchean Campbell Group, Griqualand West Sequence
of the Transvaal Supergroup, and their paleoenvi-
ronmental and evolutionary implications. Precambrian
T.S., Edmond, J.M., Lawrence, J.R., Casey, J.F.,Thompson, G., Humphris, S., Rona, P. and Karson, J.A.(1988) Chemistry of hot springs on the Mid-Atlantic Ridge.
Nature, 335, 514–519.Canfield, D.E. (1998) A new model for Proterozoic ocean
chemistry. Nature, 396, 450–453.Canfield, D.E. (2005) The early history of atmospheric
oxygen: homage to Robert M. Garrels. Annu. Rev. Earth
Planet. Sci., 33, 1–36.Canfield, D.E., Habicht, K.S. and Thamdrup, B. (2000) The
Archean sulfur cycle and the early history of atmospheric
oxygen. Science, 288, 658–661.
Cloud, P. (1965) Significance of the Gunflint (Precambrian)
microflora. Science, 148, 27–35.Cloud, P. (1969) Atmospheric and hydrospheric evolution on
the primitive Earth. Science, 160, 729–736.Cloud, P. (1973) Paleoecological significance of the banded
Microbiological processes in banded iron formation deposition 1751
mineralogy, metamorphism, geochemistry, and origin. Am.
Mineral., 90, 1473–1499.Klein, C. and Beukes, N.J. (1992) Time Distribution,
Stratigraphy, and Sedimentologic Setting, and
Geochemistry of Precambrian Iron-Formations. In: The
Proterozoic Biosphere: A Multidisciplinary Study (Eds J.W.
Schop and C. Klein), pp. 139–146. Press syndicate of the
University of Cambridge, Cambridge.
Koehler, I., Papineau, D., Konhauser, K.O. and Kappler, A.
(2013) Biological carbon precursor to diagenetic siderite
spherulites in banded iron formations. Nature Commun.,
doi:10.1038/ncomms2770.
Konhauser, K.O. (2007) Introduction to Geomicrobiology.
Blackwell, Oxford, 425 pp.
Konhauser, K., Hamade, T., Raiswell, R., Morris, R.C.,Ferris, F.G., Southam, G. and Canfield, D.E. (2002) Couldbacteria have formed the Precambrian banded iron
formations? Geology, 30, 1079–1082.Konhauser, K., Newman, D.K. and Kappler, A. (2005) The
potential significance of microbial Fe(III) reduction during