1 Department of Medical Oncology, Shuguang Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai 201203, China,
2 Department of Colorectal Surgery, Fudan University Shanghai Cancer Center, Shanghai 200032, China,
3 Shanghai University of Traditional Chinese Medicine, Shanghai 201210, China,
4 Oncology Department I of Traditional Chinese Medical Hospital affiliated Xinjiang Medical University, Xinjiang 830000, China,
5 Department of Pharmaceutical Sciences, College of Pharmacy and Health Sciences, St. John’s University, Queens, NY 11439, USA #These authors contributed equally to this work. Correspondence to: Prof./Dr. Qi Li, Department of Medical Oncology, Shuguang Hospital, Shanghai University of Traditional Chinese Medicine. 528 Zhangheng Rd, Shanghai 201203, P.R.China Tel:+8621 20256517; Fax:+8621 20256533, E-mail of Qi Li: [email protected] Running title: miR-200c induce MDR and metastasis Keywords: Multidrug resistance; ABCB1/P-glycoprotein; miR-200c; JNK signaling pathway; metastasis
Abbreviations list: CRC: colorectal cancers; MDR: multidrug resistance; ABCB1: ATP-binding cassette (ABC) transporter-subfamily B member 1; miRNAs: microRNAs; VCR: vincristine; L-OHP: Oxaliplatin; FOLFOX or CapeOX: the standard chemotherapy regimen for treatment of colorectal cancer
Grant support: National Natural Science Foundation of China (81202812, H Sui; 81373862, HR Zhu; 81001055, GX Cai), Shanghai Pujiang Program (13PJD008, GX Cai), Program of Shanghai Committee of Science and Technology (13140902500, Q Li), Shanghai Municipal Education Commission (2011JW57, H Sui; 12ZZ118 Q Li), Shanghai Municipal Health Bureau (20114Y013, H Sui; 2010019, Q Li) and National High Technology Research and Development Program (2012AA02A506, SJ Cai)
Conflict of Interest Statement: Authors have no conflicts of interest.
Total number of figures and tables: Six figures, four supplementary figures, seven supplementary tables
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
reference was performed in parallel. The relative level of miRNA expression was calculated by the
change in cycle threshold method. RNA U6 levels were used as internal reference. 2.5. Transfection of plasmids, siRNAs, miRNA mimics, miRNA inhibitors, and lentivirus production
On-target siRNAs were used to knock down JNK2 expression (Sequences of all the primers are
shown in Supplementary Table S8). miRNA mimics and inhibitors used in this study were
previously described (25). Transfection procedures were performed according to manufacturers'
instructions, with Lipofectamin 2000 as transfection reagent (Invitrogen). Briefly, 2×104 cells were
plated in each well of a 6-well plate and incubated overnight. A mixture of Lipofectamine 2000
with siRNA (50 nM), MiRNA mimic or inhibitor (50 nM) was added onto the cells, followed by a
48-h incubation in regular medium. miR-200c and miR-control lentiviral particles used to transfect
HCT8/V cells were generated by using viral packaging 293T cells. The GFP positive cells,
transfected with miR-200c-GFP-Lentivirus, were sorted and the stable clones were cultured as
previously described (25). 2.6. 3’UTR luciferase reporter assay
Full-length JNK2 (2787 nt) and ABCB1 (379 nt) 3’UTR were synthesized and cloned into the
pmiR-GLO vector (Applied Biosystems, Foster City, CA) containing a luciferase reporter gene
(JNK2 or ABCB1 3’UTR wt). The putative miR-200c recognition sites in JNK2 or ABCB1 3’UTR
were subjected to site-directed mutagenesis (JNK2 or ABCB1 3’UTR mut), and the mutated
sequences were validated by DNA sequencing. To determine the effects of miR-200c on the activity
of ABCB1 3’UTR, pmiR- ABCB1-3’UTR, pmiR- ABCB1-3’UTR-mut or negative control vector,
along with a normalized construct TK-Renilla, were co-transfected into HCT8/V cells using
TurboFectTM. Fourty-eight hours later, the transfected cells were lysed for detection of luciferase
activity. HCT8/V cells co-transfected with pmiR-JNK2-3’UTR, pmiR-JNK2-3’UTR-mut or
negative control vector were similarly examined. 2.7. Western blot analysis
Whole cell lysates for Western blot analysis of ABCB1, ABCC-1/2, BCRP, JNK1, JNK2, p-JNK,
c-Jun, p-c-Jun, ATF-2, p-ATF-2, Elk-1 and �-actin expression were prepared as previously reported
(10). Briefly, the cells were lysed on ice in immunoprecipitation assay buffer for 2 h before being
homogenized using a mortar and pestle. The homogenized sample was centrifuged, and the
supernatant was collected and stored at -80°C. Densitometric analysis was done using the Scion
Imaging software (Scion Corporation), with �-actin as internal reference.
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
in CRC MDR, probably through modulating JNK signaling dependent P-gp expression.
Next, we assessed the expression levels of Bcl-2, Bcl-xl, Survivin, MMP2/9 and TIMP-1/2, the
important proteins regulating invasion and migration, in HCT8/V cells treated by miR-200c
inhibition. Western blot analysis determined that the protein levels of Bcl-2, Bcl-xl, and Survivin
did not significantly change, but the protein levels of MMP2/9 were increased, while the levels of
TIMP-1/2 (suppressors of MMPs) were lowered upon miR-200c inhibition, in HCT8/V cells (Fig.
3H&I). 3.4. miR-200c modulates MDR and cancer cell invasion/migration in vitro
Next we determined if miR-200c reversed the P-gp mediated MDR. We first established that
either increasing the expression of miR-200c (miR-200cover) or decreasing JNK2 protein levels
(sh-JNK2) reduced cell growth in CRC cell lines HCT8/V and HCT116/MDR treated with
chemotherapy agents such as cDDP, 5-FU, MMC and THP (Fig. 4 A&B and Supplementary Fig.
4). It is important to note, the overexpression of JNK2 (JNK2over) rescued the cells from
miR-200cover induced growth inhibition. Flow cytometry analyses then demonstrated an increased
apoptosis accompanying miR-200cover or sh-JNK2 expression in CRC cells (Fig. 4C). Employing
HPLC assay, we next discovered that the intracellular accumulation of chemotherapeutic agents
such as VCR in HCT8/V cells was increased by miR-200cover in a dose-dependent manner (Fig. 4D).
These data suggested that miR-200c significantly decreased the efflux of chemotherapeutic drugs in
a P-gp-overexpressing MDR cancer cell, resulting in higher intracellular drug concentration and
longer retaining time, enhancing the tumoritoxic effect of the drug.
Previous studies have shown that transfection with miR-200c inhibits cell invasion and migration
in human cancer cell lines (30). Herein, we investigated the effect of miR-200c reconstitution on the
invasive and migratory capabilities of MDR CRC cell lines. As shown in Fig. 4E, miR-200cover
significantly reduced HCT8/V cells’ capability of invading through the Matrigel-coated transwell.
Similar results were seen in wound-healing assays, in which miR-200cover in MDR cell lines
markedly reduced cell migration (Fig. 4F). In conclusion, these data support a role for
miR-200c/JNK2 axis in regulating the MDR and invasion/migration.
3.5. miR-200c reverses P-gp mediated MDR and metastasis in vivo To further test our hypothesis of miR-200c’s role in MDR in vivo, we first established a
xenograft tumor model of fluorescence-labeled MDR CRC cell line, control vs miR-200cover. When
we treated these tumor bearing mice with oxaliplatin, a clinically used chemotherapy agent, the
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
miR-200cover tumors are smaller than the controls (Fig. 5A). We next tested the in vivo metastatic
potential of miR-200cover CRC cells HCT116/L in a colonic orthotopic xenograft tumor model, and
found fewer colonic metastatic sites in livers and lungs of the miR-200cover group (Fig. 5B and
Supplementary Table S9). It is important to note that our procedures to set up xenograftic tumors
using lentivirus infected cells had minimal detrimental effect to the animals. Immunohistochemistry
analyses of these tumor confirmed that the levels of JNK2, p-JNK, p-c-Jun, P-gp and MMP-2/9
were decreased in miR-200cover tumors, compared controls (Fig. 5C). In summary, our data
purported that in vivo tumor growth and metastasis of MDR CRC were inhibited by miR-200c
reconstitution via repressing JNK2/c-Jun/P-gp signaling.
3.6. Relationship of miR-200c and JNK2, MMPs after chemotherapy As blood corpuscle has been reported to contain high levels of RNA activity (24), we screened
miR-200c expression in plasma samples of the patients with poor response to chemotherapy. We
found that the patients with poor response to chemotherapy had lower amount of miR-200c, but
higher levels of ABCB1, JNK2 and MMP-2/-9 in their plasma samples. And more interestingly, the
levels of miR-200c progressively decreased, while the expression levels of ABCB1, JNK2 and
MMP-9 increased, as more rounds of chemotherapy were administrated (Fig. 6A&B&C&D).
However, from Fig. 6E we could observe that level of MMP-2 was not changed significantly in the
colon cancer patients during the chemotherapy process.
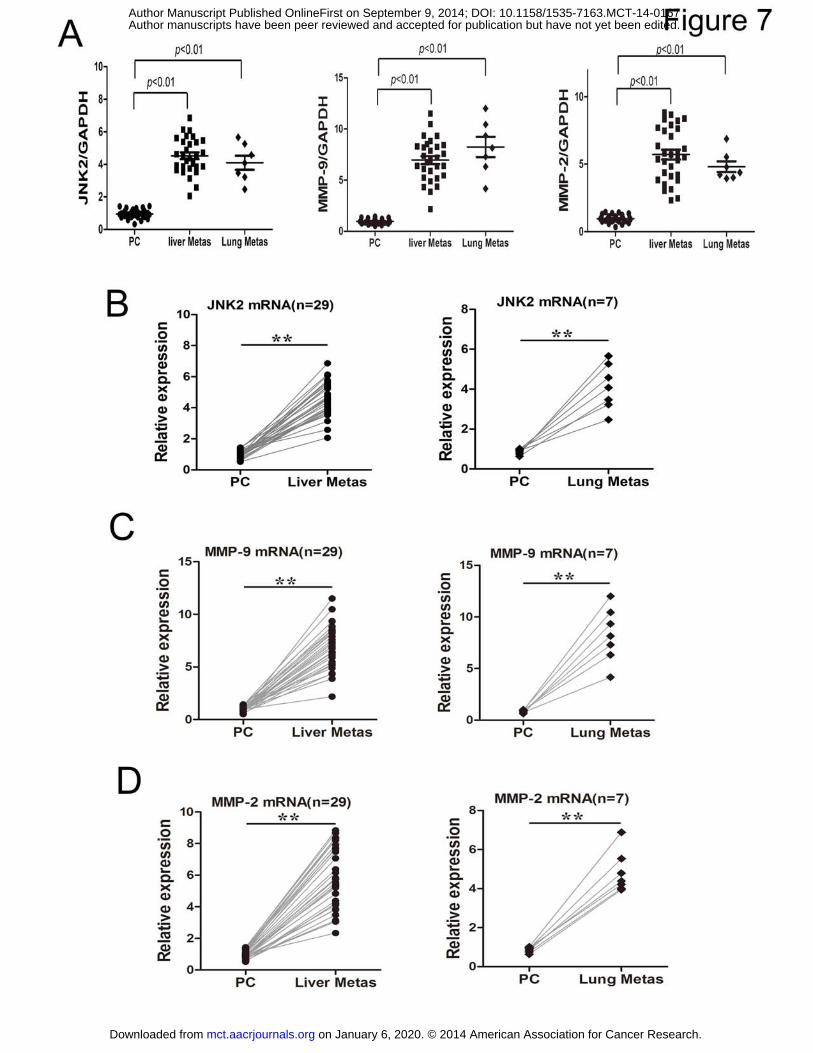
Lastly, we examined the levels of JNK2 and MMP-2/-9 mRNA in 30 pairs of primary CRCs and
their matched liver and lung metastases. The results indicated that the expression of JNK2 and
MMP-2/-9 were significantly higher, inversely correlated with the low levels of miR-200c in
metastasized CRC sites compared to the primary tumors (Fig. 7A&B&C&D). To summarize, these
data suggest that the attenuated expression of miR-200c correlated with up-regulated JNK2
expression, promoting P-gp function, MDR phenotype and metastasis in CRC.
4. Discussion
Resistance to anticancer drug therapies and tumor metastases are the main causes of morbidity
and mortality of cancer patients. MDR is a complicated multifaceted phenomenon, which is
mediated by a spectrum of integral membrane proteins, including ABCB1/P-gp, ABCC-1/2 and
ABCG2/BCRP. To date, the mechanisms of regulating the expression levels of these proteins
remain largely unexplored. Recently, several lines of evidence have purported to study the drug
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
We thank Drs. Qing Ji and Jian Sun for technical assistance. We thank Dr. S. Paul Gao for
manuscript copyedited assistance.
References 1. Lu L, Zhou D, Jiang X, Song K, Li K, Ding W. Loss of E-cadherin in multidrug resistant breast cancer cell
line MCF-7/Adr: possible implication in the enhanced invasive ability. Eur Rev Med Pharmacol Sci 2012;16:1271-9.
2. Qiang F, Guangguo R, Yongtao H, Dandan D, Hong Y. Multidrug resistance in primary tumors and metastases in patients with esophageal squamous cell carcinoma. Pathol Oncol Res 2013;19:641-8.
3. Liang Z, Wu H, Xia J, Li Y, Zhang Y, Huang K, et al. Involvement of miR-326 in chemotherapy resistance of breast cancer through modulating expression of multidrug resistance-associated protein 1. Biochem Pharmacol 2010;79:817-24.
4. Ward AB, Szewczyk P, Grimard V, Lee CW, Martinez L, Doshi R, et al. Structures of P-glycoprotein reveal its conformational flexibility and an epitope on the nucleotide-binding domain. Proc Natl Acad Sci U S A 2013;110:13386-91.
5. Sui H, Fan ZZ, Li Q. Signal transduction pathways and transcriptional mechanisms of ABCB1/Pgp-mediated multiple drug resistance in human cancer cells. The Journal of International Medical Research 2012;40:426-35.
6. Bankovi� J, Andrä J, Todorovi� N, Podolski-Reni� A, Miloševi� Z, Miljkovi� D, et al. The elimination of P-glycoprotein over-expressing cancer cells by antimicrobial cationic peptide NK-2: the unique way of multi-drug resistance modulation. Exp Cell Res 2013;319:1013-27.
7. Raciti M, Lotti LV, Valia S, Pulcinelli FM, Di Renzo L. JNK2 is activated during ER stress and promotes cell survival. Cell Death Dis 2012;3:e 429.
8. Sabapathy K, Hochedlinger K, Nam SY, Bauer A, Karin M, Wagner EF. Distinct Roles for JNK1 and JNK2 in Regulating JNK Activity and c-Jun-Dependent Cell Proliferation. Mol Cell 2004;15:713-25.
9. Zhan X, Feng X, Kong Y, Chen Y, Tan W. JNK signaling maintains the mesenchymal properties of multi-drug resistant human epidermoid carcinoma KB cells through snail and twist1. BMC Cancer 2013;13:180.
10. Sui H, Zhou S, Wang Y, Liu X, Zhou L, Yin P, et al. COX-2 contributes to P-glycoprotein-mediated multidrug resistance via phosphorylation of c-Jun at Ser63/73 in colorectal cancer. Carcinogenesis 2011;32:667-75.
11. Wang N, Li Z, Tian F, Feng Y, Huang J, Li C, et al. PKC� inhibited apoptosis by decreasing the activity of JNK in MCF-7/ADR cells. Exp Toxicol Pathol 2012;64:459-64.
12. Takeyama H, Yamamoto H, Yamashita S, Wu X, Takahashi H, Nishimura J, et al. Decreased miR-340 expression in bone marrow is associated with liver metastasis of colorectal cancer. Mol Cancer Ther 2014;13:976-85.
13. Jia W, Eneh JO, Ratnaparkhe S, Altman MK, Murph MM. MicroRNA-30c-2* expressed in ovarian cancer cells suppresses growth factor-induced cellular proliferation and downregulates the oncogene BCL9. Mol Cancer Res 2011;9:1732-45.
14. Wei J, Wang F, Kong LY, Xu S, Doucette T, Ferguson SD, et al. miR-124 inhibits STAT3 signaling to enhance T cell-mediated immune clearance of glioma. Cancer Res. 2013;73:3913-26.
15. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009;136:215-33. 16. Xia L, Zhang D, Du R, Pan Y, Zhao L, Sun S, et al. miR-15b and miR-16 modulate multidrug resistance by
targeting BCL2 in human gastric cancer cells. Int J Cancer 2008;123:372-9. 17. Wang M, Zhang P, Li Y, Liu G, Zhou B, Zhan L, et al. The quantitative analysis by stem-loop real-time PCR
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
revealed the microRNA-34a, microRNA-155 and microRNA-200c overexpression in human colorectal cancer. Med Oncol 2012;29:3113-8.
18. Chen J, Wang W, Zhang Y, Chen Y, Hu T. Predicting distant metastasis and chemoresistance using plasma miRNAs. Med Oncol 2014;31:799.
19. Lu H, Sun J, Xu JH, Fan ZZ. Establishment of an oxaliplatin-resistant human colon carcinoma HCT116/L-OHP cell and preliminary exploration for the mechanism of resistance. Tumor 2011;31:675-81.
20. Sui H, Zhu HR, Wu J, Nikitin AY, Cai JF, Fan ZZ, Li Q. Effets of Jianpi Jiedu Recipe on Reversion of P-glycoprotein–mediated Multidrug Resistance through COX-2 pathway in Colorectal Cancer. Chin J Integr Med 2013;doi: 1111117/s11655-112-1257-x.
21. Li CL, Liu JW, Sa XY, Deng WL, Xu JH, Fan ZZ. Preliminary study on establishment of a VCR resistant colon carcinoma cell line and its underlyingmechan ism of drug resistance. Tumor 2009;29:31-4.
22. Gu W, Zhang YN, Li B, Han J, Cheng BB, Ling CQ. Establishment of a multidrug- resistant cell line BEL-7402/5-FU of human hepatocellular carcinoma and its biological characteristics. Journ al of Chinese Integrative Medicine 2006;4:265-70.
23. Pan HM, Fei HX, Du JP, Chen ZH, Zhang T. Construction of a cisplatin-induced human gastric cancer drug resistant cell line World Chinese Journal of Digestology 2007;15:2009-13.
24. Huang Z, Huang D, Ni S, Peng Z, Sheng W, Du X. Plasma microRNAs are promising novel biomarkers for early detection of colorectal cancer. Int J Cancer 2010;127:118-26.
25. Lin PC, Chiu YL, Banerjee S, Park K, Mosquera JM, Giannopoulou E, et al. Epigenetic repression of miR-31 disrupts androgen receptor homeostasis and contributes to prostate cancer progression. Cancer Res 2013;73:1232-44.
26. Sui H, Liu X, Jin BH, Pan SF, Zhou LH, Yu NA, et al. Zuo Jin Wan, a Traditional Chinese Herbal Formula, reverses P-gp mediate MDR in vitro and in vivo. ECAM 2013;doi.org/10.1155/2013/957078.
27. Chen D, Zhang Y, Wang J, Chen J, Yang C, Cai K, et al. MicroRNA-200c overexpression inhibits tumorigenicity and metastasis of CD117+CD44+ovarian cancer stem cells by regulating epithelial-mesenchymal transition. J Ovarian Res 2013;6:50.
28. Kolinsky K, Shen BQ, Zhang YE, Kohles J, Dugan U, Zioncheck TF, et al. In vivo activity of novel capecitabine regimens alone and with bevacizumab and oxaliplatin in colorectal cancer xenograft models. Mol Cancer Ther 2009;8:75–82.
29. Rokavec M, Wu W, Luo JL. IL6-mediated suppression of miR-200c directs constitutive activation of inflammatory signaling circuit driving transformation and tumorigenesis. Mol Cell 2012;45:777-89.
30. Hur K, Toiyama Y, Takahashi M, Balaguer F, Nagasaka T, Koike J, et al. MicroRNA-200c modulates epithelial-tomesenchymal transition (EMT) in human colorectal cancer metastasis. Gut 2013;62:1315-26.
31. Feng DD, Zhang H, Zhang P, Zheng YS, Zhang XJ, Han BW, et al. Down-regulated miR-331-5p and miR-27a are associated with chemotherapy resistance and relapse in leukaemia. J Cell Mol Med 2011;15:2164-75.
32. Zhang P, Bill K, Liu J, Young E, Peng T, Bolshakov S, et al. MiR-155 is a liposarcoma oncogene that targets casein kinase-1� and enhances �-catenin signaling. Cancer Res 2012;72:1751-62.
33. Yu CC, Tsai LL, Wang ML, Yu CH, Lo WL, Chang YC, et al. miR145 targets the SOX9/ADAM17 axis to inhibit tumor-initiating cells and IL-6-mediated paracrine effects in head and neck cancer. Cancer Res 2013;73:3425-40.
34. Lu L, Zhou D, Jiang X, Song K, Li K, Ding W. Loss of E-cadherin in multidrug resistant breast cancer cell line MCF-7/Adr: possible implication in the enhanced invasive ability. Eur Rev Med Pharmacol Sci 2012;16:1271-9.
35. Zhu W, Xu H, Zhu D, Zhi H, Wang T, Wang J, et al. miR-200bc/429 cluster modulates multidrug resistance of human cancer cell lines by targeting BCL2 and XIAP. Cancer Chemother Pharmacol 2012;69:723-31.
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
36. Rokavec M, Wu W, Luo JL. IL6-Mediated Suppression of miR-200c Directs Constitutive Activation of Inflammatory Signaling Circuit Driving Transformation and Tumorigenesis. Mol Cell 2012;45:777-89.
37. Davies C, Tournier C. Exploring the function of the JNK (c-Jun N-terminal kinase) signalling pathway in physiological and pathological processes to design novel therapeutic strategies. Biochem Soc Trans 2012;40:85-9
38. Zhang W, Chen BA, Jin JF, He YJ, Niu YQ. Involvement of c-Jun N-terminal kinase in reversal of multidrug resistance of human leukemia cells in hypoxia by 5-bromotetrandrine. Leuk Lymphoma 2013;54:2506-16.
39. Li Z, Min W, Gou J. Knockdown of cyclophilin A reverses paclitaxel resistance in human endometrial cancer cells via suppression of MAPK kinase pathways. Cancer Chemother Pharmacol 2013;72:1001-11.
40. Tang PM, Zhang DM, Xuan NH, Tsui SK, Waye MM, Kong SK, et al. Photodynamic therapy inhibits P-glycoprotein mediated multidrug resistance via JNK activation in human hepatocellular carcinoma using the photosensitizer pheophorbide a. Mol Cancer 2009;8:56.
41. Yan F, Wang XM, Liu ZC, Pan C, Yuan SB, Ma QM. JNK1, JNK2, and JNK3 are involved in P-glycoprotein-mediated multidrug resistance of hepatocellular carcinoma cells. Hepatobiliary Pancreat Dis Int 2010;9:287-95.
42. Bark H, Xu HD, Kim SH, Yun J, Choi CH. P-glycoprotein down-regulates expression of breast cancer resistance protein in a drug-free state. FEBS Lett 2008;582:2595-600.
43. Tajitsu Y, Ikeda R, Nishizawa Y, Mataki H, Che XF, Sumizawa T, et al. Molecular basis for the expression of major vault protein induced by hyperosmotic stress in SW620 human colon cancer cells. Int J Mol Med 2013;32:703-8.
44. Tang H, Deng M, Tang Y, Xie X, Guo J, Kong Y, et al. miR-200b and miR-200c as prognostic factors and mediators of gastric cancer cell progression. Clin Cancer Res 2013;19:5602-12.
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Figure 5. miR-200c reverses P-gp mediated MDR and metastasis in vivo (A) Left panel: Luciferase imaging of the mice with control or miR-200c overexpressing xenografts day 10 and day
60 after tumor cell implantation. These tumor bearing mice were treated with oxaliplatin for 30 days. Upper-right
panel: The quantification of luciferase intensities in tumors of Group 2 and 3 mice. Low-right panel: Tumor samples
dissected from the indicated mice and photographed on the 60th day post-tumor implantation. (B) Upper: mice were
colonic-orthotopically implanted with control and miR-200c overexpressing CRC cells. Luciferase imaging revealing
primary tumor growth and distant metastasis. Low-left panel: body weight of tumor bearing mice. Low-right panel:
The survival of mice implanted with HCT116/L-OHP (NC), lentivirus control and lentivirus miR-200c expressing
cells. (C) The orthotopic transplantation tumor samples in mice of Groups 5 and 6 were analyzed by
immunohistochemistry for P-gp, JNK2, p-c-Jun, p- JNK and MMP2/9. Magnification 400x. Positive cells were
stained brown.
Figure 6. Relationship of miR-200c and JNK2 after chemotherapy (A, B, C) Large-scale validation of miR-200c JNK2 and ABCB1 levels in patient plasma samples. Scatter plots of
plasma levels of miR-200c in healthy subjects (n=30), before chemotherapy (n=30), after second (n=30), fourth
(n=30) and sixth (n=27) cycle of chemotherapy. Expression levels of the miRNAs (fold change at Y-axis) are
normalized to U6. The line represents the median value. Mann-Whitney U test was used to determine statistical
significance. (D) RNA was extracted from 30 primary CRC samples and paired liver and lung metastasis tissues (as
Fig. 1B described). The mRNA levels of JNK2 were quantified by qRT-PCR in primary (N = 30), hepatic metastasis
(N = 29), and lung metastasis (N = 7). (E) Expression of JNK2 in matched primary CRC (PC) and their
corresponding liver and lung metastasis. The bold horizontal bar represents mean expression levels; *p<0.05,
**p<0.01, t test.
Figure 7. Expression of JNK2 and MMP-2/9 in matched primary CRC (PC) and their corresponding liver and lung metastasis samples (A) RNA was extracted from 30 primary CRC samples and paired liver and lung metastasis tissues (as Fig. 1B
described). The mRNA levels of JNK2 and MMP-2/9 were quantified by qRT-PCR in primary (N = 30), hepatic
metastasis (N = 29), and lung metastasis (N = 7). (B, C, D) Expression of JNK2, MMP-2 and MMP-9 in matched
primary CRC (PC) and their corresponding liver and lung metastasis. The bold horizontal bar represents mean
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167
Published OnlineFirst September 9, 2014.Mol Cancer Ther Hua Sui, Guo-Xiang Cai, Shu-Fang Pan, et al. targeting JNK2/c-Jun signaling pathway in colorectal cancermiR-200c attenuates P-gp mediated MDR and metastasis by
Updated version
10.1158/1535-7163.MCT-14-0167doi:
Access the most recent version of this article at:
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on September 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0167