Mitosis and cell division in Cryptomonas (Cryptophyceae) Deprrrtmenr ofBiology, York University. 4700 Keele Street, Downsview, Ot~t., Cannrln M3J 1P3 AND Deprrrrtnetlt ofBolrrtly, The Utzivemity ofBritis11 Colrrtnbin, Vnncorn~er, B . C . , Cnnndrr V6T lW5 Received May 26, 1977 OAKLEY, B. R.. and T. BISALPUTRA. 1977. Mitosis and cell division in Cryptotnot~rr.r (Cryp- tophyceae). Can. J. Bot. 55: 2789-2800. Mitosis and cytokinesis are examined in Ctyptomonrrs sp., a member of the Cryptophyceae. The beginning of prophase is signalled by the replication of the flagellar bases which are at the anterior of the cell and a proliferation of the microtubules which run from them to the nucleus at the posterior. The microtubules continue to proliferate as the flagellar bases migrate apart and the nucleus migrates to the anterior. They dissociate from the flagellar bases and enter the nucleus as the nuclear envelope breaks down. A rectangular spindle forms and at prometaphase chromatin is scattered along the spindle. From this stage until late telophase microtubules are found attached singly to the chromatin. In metaphase the chromatin forms a solid plate penetrated by tunnels through which microtubules pass and in anaphase the chromatin separates in two masses which move toward the poles as the spindle elongates. In telophase the nuclear envelope re-forms while a number of microtubules remain between the daughter nuclei. The cytokinetic furrow forms during metaphase and constricts gradually until cytokinesis is complete at telophase. A thin ring of amorphous material is seen under the plasma membrane in the cytokinetic furrow. These results suggest that on the basis of mitotic criteria there is little similarity between the cryp- tophytes and either the dinoflagellates or red algae to which they have been previously linked. Introduction genetic position of the cryptophytes is reflected The Cryptophyceae is a small, distinct class of by the different positions they occupy in various unicellular, flagellated algae which has aroused phylogenetic schemes. Lee (1972) suggests that considerable interest in recent years because of its possibly primitive but highly controversial phylogenetic position. Traditionally the crypto- phytes have been placed in the Pyrrophyta along with the dinoflagellates but a number of authors in recent years have linked them to the red algae on the basis that both groups have phyco- bilin accessory pigments (e.g. Raven 1970). Several authors have suggested that the dino- flagellates are among the most primitive extant eukaryotes on the basis of nuclear and mitotic characters (Dodge 1965; Kubai and Ris 1969; Dupraw 1970; Kubai 1975). The red algae have been regarded as the most primitive eukaryotes by many others on the basis of their lack of flagella and on the similarity of their pigments to those of the blue-green algae. If the crypto- phytes are related to either the dinoflagellates or to the red algae they should be very primitive and should occupy a very interesting, possibly key, evolutionary position. The extent of disagreement over the phylo- 'Present address: Department of Pharmacology, Rutgers Medical School, CMDNJ, P.O. Box 101, Piscataway, NJ, U.S.A. 08854. all the algae evolved from various cryptophytes although only the red algae evolved from those with chloroplasts. Scagel et al. (1965), however, place them well out on the same evolutionary line as the dinoflagellates, diatoms, brown algae, and chrysophytes and quite far away from the red algae. Cavalier-Smith (1975) has them evolving directly from a primordial "chromo-amoebo- flagellate" and giving rise to nothing. Various authors have recently pointed out that mitotic structures may be useful phylogenetic criteria (Pickett-Heaps 1969, 1 9 7 2 ~ ; and Kubai 1975 among others). With this in mind Oakley and Dodge (1973, 1976) studied mitosis in Chroomonas salina and suggested that crypto- phyte mitosis has similarities to that of a number of algae of various classes but little similarity to that of either the red algae or dinoflagellates. This study was undertaken with three chief aims in mind. The first was to begin to obtain data on the variability of mitosis in the Crypto- phyceae to see if conclusions based on Chroomo- nus salina are applicable to the rest of the class. The second was to extend light microscopic ob- servations of division in the class and correlate Can. J. Bot. Downloaded from www.nrcresearchpress.com by YORK UNIV on 06/29/14 For personal use only.
Transcript
Mitosis and cell division in Cryptomonas (Cryptophyceae)
Deprrrtmenr ofBiology, York University. 4700 Keele Street, Downsview, O t ~ t . , Cannrln M3J 1P3
OAKLEY, B . R.. and T. BISALPUTRA. 1977. Mitosis and cell division in Cryptotnot~rr.r (Cryp- tophyceae). Can. J. Bot. 55: 2789-2800.
Mitosis and cytokinesis are examined in Ctyptomonrrs sp., a member of the Cryptophyceae. The beginning of prophase is signalled by the replication of the flagellar bases which are at the anterior of the cell and a proliferation of the microtubules which run from them to the nucleus at the posterior. The microtubules continue to proliferate as the flagellar bases migrate apart and the nucleus migrates to the anterior. They dissociate from the flagellar bases and enter the nucleus as the nuclear envelope breaks down. A rectangular spindle forms and at prometaphase chromatin is scattered along the spindle. From this stage until late telophase microtubules are found attached singly to the chromatin. In metaphase the chromatin forms a solid plate penetrated by tunnels through which microtubules pass and in anaphase the chromatin separates in two masses which move toward the poles a s the spindle elongates. In telophase the nuclear envelope re-forms while a number of microtubules remain between the daughter nuclei. The cytokinetic furrow forms during metaphase and constricts gradually until cytokinesis is complete at telophase. A thin ring of amorphous material is seen under the plasma membrane in the cytokinetic furrow. These results suggest that on the basis of mitotic criteria there is little similarity between the cryp- tophytes and either the dinoflagellates or red algae to which they have been previously linked.
Introduction genetic position of the cryptophytes is reflected The Cryptophyceae is a small, distinct class of by the different positions they occupy in various
unicellular, flagellated algae which has aroused phylogenetic schemes. Lee (1972) suggests that considerable interest in recent years because of its possibly primitive but highly controversial phylogenetic position. Traditionally the crypto- phytes have been placed in the Pyrrophyta along with the dinoflagellates but a number of authors in recent years have linked them to the red algae on the basis that both groups have phyco- bilin accessory pigments (e.g. Raven 1970). Several authors have suggested that the dino- flagellates are among the most primitive extant eukaryotes on the basis of nuclear and mitotic characters (Dodge 1965; Kubai and Ris 1969; Dupraw 1970; Kubai 1975). The red algae have been regarded as the most primitive eukaryotes by many others on the basis of their lack of flagella and on the similarity of their pigments to those of the blue-green algae. If the crypto- phytes are related to either the dinoflagellates or to the red algae they should be very primitive and should occupy a very interesting, possibly key, evolutionary position.
The extent of disagreement over the phylo-
'Present address: Department of Pharmacology, Rutgers Medical School, C M D N J , P.O. Box 101, Piscataway, NJ, U.S.A. 08854.
all the algae evolved from various cryptophytes although only the red algae evolved from those with chloroplasts. Scagel et al. (1965), however, place them well out on the same evolutionary line as the dinoflagellates, diatoms, brown algae, and chrysophytes and quite far away from the red algae. Cavalier-Smith (1975) has them evolving directly from a primordial "chromo-amoebo- flagellate" and giving rise to nothing.
Various authors have recently pointed out that mitotic structures may be useful phylogenetic criteria (Pickett-Heaps 1969, 1 9 7 2 ~ ; and Kubai 1975 among others). With this in mind Oakley and Dodge (1973, 1976) studied mitosis in Chroomonas salina and suggested that crypto- phyte mitosis has similarities to that of a number of algae of various classes but little similarity to that of either the red algae or dinoflagellates.
This study was undertaken with three chief aims in mind. The first was to begin to obtain data on the variability of mitosis in the Crypto- phyceae to see if conclusions based on Chroomo- nus salina are applicable to the rest of the class. The second was to extend light microscopic ob- servations of division in the class and correlate
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
2790 C A N . J. BOT. \
our electron microscopy with light microscopy. The third was to extend the very limited observa- tions on the cytokinetic process in the class.
Materials and Methods Cr)'p/ott~otms sp. was obtained from Dr. Naval Antia
(Fisheries Research Board, North Vancouver). The original culture was started by Dr. Luigi Provasoli (Haskins Laboratories, New Haven) from a collection made in the gulf stream and identified a s Rhodotno~ms letls Pascher et Ruttner. We reject this designation on two grounds. The first is that the genus R/~odot?~ot~as is probably not valid (Butcher 1967). The second is that the organism does not resemble very closely the original description of Rhodomo~~as Iet~s which was a freshwater organism collected from alpine lakes (Pascher 1913). In fact we have not been able to find any description which matches the organism very closely and it is probably a new species. If one accepts the genus Rhodotnonos it would clearly be in that genus on the basis of its "bor- deaux-red" color. We tend to agree with Butcher's view that this genus is meaningless, however, and place it in the genus Crypton7011os. By all present taxonomies it is clearly not in the genus C/~root~~otm.s because it has four rows of gullet trichocysts.
Cultures were grown in F/2 medium (Guillard and Ryther 1962) or NS medium (Loeblich 1975). Cir- cumstances necessitated growing the organisms in diverse culture conditions where they proved to grow quite rapidly under various light regimes between 20 and 30°C.
For phalloidin treatment cells were grown in con- tinuous light in small test tubes containing N S medium. Thirty minutes before fixation phalloidin in the samc ~nedium was added to make final concentrations of to M. Phalloidin was a generous donation of Dr. Theodor Wieland (Abteilung Naturstoff-Chernie, Heidel- berg).
Fixation for the determination of cell division rates and for light microscopy was carried out in the following manner. Since the cells adhere to the bottom and sides of glass flasks the flasks were shaken gently before sam- pling. Two-millilitre samples were taken and fixed im- mediately by the addition of one o r two drops of IOZ glutaraldehyde solution. Cells were examined using inter- ference contrast optics.
For electron microscopy the technique of Oakley and Dodge (1976) was used with two modifications. The first was that 0.4 M sucrose was used for osmotic balance in the primary fixative solution. The second was that in- stead of concentrating the cells by centrifugation before fixation the medium was gently decanted from the growth flask and those cells adhering to the flask were fixed by adding the primary fixative solution directly to the flask.
Detailed methodology for the serial sectioning will be given later (Oakley and Heath, in preparation). Briefly, however, after flat embedding on slides, dividing cells were selected and photographed using interference con- trast optics, marked, cut out, reembedded, and serial sectioned. Serial sections were picked up using single- slot, formvar-coated I x 2 mm grids.
Observations Syncl~rony
It proved difficult to determine growth rates accurately in cultures of Cryptomonas because many cells adhered tenaciously to the culture flasks thus making accurate cell counts very difficult. Observations on cultures growing in a 13 h light-1 1 h dark cycle at 25°C showed that average cell size increased in the light period and decreased in the dark period. This suggested that most cell division occurred in the dark so at- tempts were made to ascertain exactly when the divisions occurred. Initial attempts using living samples failed but, when samples were taken and immediately fixed, dividing cells were found from the 8th to the 10th h of darkness. In young, rapidly growing cultures there were very few or no divisions at other times but in older cultures occasional divisions were found outside of this period. Initial failures were probably due to the fact that the period in which the cells are recognizably dividing (i.e. cytokinesis) only lasts about 10 min (see observations following) and cells do not seem to initiate division after they are agitated and taken into the light. Thus, in living samples most divisions have finished be- fore accurate counts can be made and no new divisions begin. The greatest percentage of cytokinetic cells we have found during the maximal division period is about 4%. This seems a small figure but if the short cytokinetic time is taken into consideration it means that about 25% of the cells divide in an hour. Electron micros- copy of material fixed in this period showed more than adequate numbers of dividing cells for observation.
Light Microscopy Careful comparison of light and electron
micrographs of the same cell shows that it is possible in some instances to discern the chro- matin mass with the light inicroscope within living cells. Light microscopy proves in general to be of little use in following the movement of the chromatin because the cells are small and many refractile bodies complicate the image. It is quite useful, though, in following flagellar replication and the changes in cell shape which occur during cell division. It is also necessary, obviously, to use light microscopy to determine the time course of division.
As revealed by both light and electron micros-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
OAKLEY A N D RISAL.PU1'KA 2791
FIGS. 1-6. A time sequence of division in Crypfomo~ms. Figure 1 was taken at time 0, Fig. 2 at 20 min, Fig. 3 at 30 min, Fig. 4 at 40 min, Fig. 5 at 50min, and Fig. 6 at 60min. Observatio~l a t various planes of focus shows four flagella at time 0. The cell rounds LIP gradually but the nucleolus is still present in Fig. 4 (arrowhead). Mitosis and cytokinesis proceed very rapidly, the entire sequence occurring between Fig. 4 and Fig. 6. The cytokinetic furrow is apparent in Fig. 5. All figures Nomarski interference contrast optics. All x 2800.
copy, cells before division are about 12 pm long, tually flagella of the half-cells extend in opposite - -
7 pm wide, and 5 pm thick and are ovoid with a directions. gullet at the anterior end. The gullet is lined with We can not be sure of the exact duration of cell four rows of trichocysts which are visible with division because there is an obvious effect from the light microscope and two flagella extend from illumination under the microscope. If cells are the base of the gullet. The beginning of the observed continuously under even the minimum mitotic sequence is signalled by an increase in possible light intensity cytokinesis may take an the number of trichocysts surrounding the gullet hour or more and is often never completed. If and by the replication of the flagella: Evidently cells are observed intermittently with illumina- the old flagella remain intact because cells with tion for picture taking a t 10-min intervals cyto- two short and two long flagella are often seen kinesis is much quicker, being completed in whereas cells with four short flagella are not roughly 10 min. Figures 1-6 show a series of seen.
As division proceeds (Figs. 1-6) the cell rounds up and the flagella move apart. Cyto- kinesis occurs as the result of a ring of con- striction passing around the cell in the anterior- posterior plane (Fig. 5). As cytokinesis occurs daughter 'half-cells' twist in relation to each other around the axis of division so that even-
pictures of a dividing cell, from flagellar rep- lication through cytokinesis, taken in this manner. They almost certainly do not divide any faster than normal in these conditions and may divide somewhat slower. These figures also show that the time from flagellar base replication to the beginning of cytokinesis is considerably longer than the time of cytokinesis itself.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
2792 CAN. J . BOT. VOL. 55, 1977
Electron Microscopy Interphase Interphase cells of Cryptomonas sp. (Fig. 7)
are typically cryptophycean in appearance re- sembling very closely cryptophytes previously studied (Dodge 1969; Lucas 1970a, 19706; Hibberd et al. 1971 ; Taylor and Lee 1971). The periplast of the cell is very similar to that described by Hibberd et al. (1971) for Crypto- monas and by Gantt (1971) for Chroomonas with a plasma membrane beneath which are small trichocysts interspaced by very thin plates. There is a single parietal chloroplast running from posterior to anterior. It is 'U' shaped in cross section and encompasses about three- quarters of the cell. From the pair of flagellar bases at the base of the gullet a few micro- tubules run back to the area of the nucleus which is at or near the extreme posterior of the cell. The nucleus is typically cryptophycean, possessing a large nucleolus and abundant heterochromatin. Occasionally there is a lipid globule posterior to the nucleus but this is not common in young cultures.
Prophase The beginning of the mitotic sequence is
signalled by the replication of the flagellar bases and the growth of two new flagella. Figure 8 shows four flagellar bases close together in a diamond with the small but complex flagellar root system present. We can not be sure of the exact sequence of events in flagellar replication but it is obvious the new flagellar bases form as part of a complex with the old pair and later separate rather than being formed separately.
Beginning with the replication of the flagellar bases there is a proliferation of microtubules extending from the flagellar roots and from amorphous material around the flagellar bases. Many but not all of these microtubules extend down to the nucleus. The nucleus then migrates
toward the anterior of the cell as the cell rounds UP.
The pairs of flagellar bases begin to separate as the number of microtubules associated with the flagellar bases continues to increase. This process results in a pair of flagellar bases with great numbers of associated microtubules on each side of the nucleus (Fig. 9). The nuclear envelope then breaks down as the spindle begins to form. Microtubules lose their association with the flagellar bases and reorient to form a spindle running across the cell. As the spindle forms the chromatin condenses and prometaphase begins (Figs. 10-12). In this stage the chromatin is not found in one tight mass as it is in metaphase (Fig. 15) or two tight masses as it is in anaphase (Fig. 17) but rather in clumps along the spindle. Serial sectioning reveals that these clumps are interconnected. This stage is not found often in sections and thus is probably very short in duration. It ends as the chromatin forms a tight metaphase plate.
Metaphase The spindle at metaphase (Figs. 13, 15) is
shaped like a short, stubby barrel. The exact dimensions are somewhat variable from cell to cell but the diameter at the metaphase plate averages about 3.5 pm and the pole-to-pole distance averages about 2.5 pm. There are flagellar base pairs on opposite sides of the chromatin mass but there is no connection between the spindle microtubules and the bases.
A detailed analysis of the configuration of the microtubules in the spindle based on serial sectioning is in progress (Oakley and Heath 1976, and in preparation) and will be given elsewhere. We will, however, summarize briefly the types of microtubules found. There are a very small number ( I 1%) of microtubules which go from pole to pole. There are greater
FIG. 7. An interphase cell showing the general shape of the cell and the arrangement of organelles. The darkly staining organelles in rows at the upper left are g~lllet trichocysts. Smaller trichocysts are seen beneath the periplast. The nucleus with abundant heterochromatin and prominent nucleolus are in the posterior half of the cell anterior to a small lipid globule. x 11 000. FIG. 8. Flagellar bases soon after replication. The four bases are close together in a diamond pattern. Portions of the flagellar root system are visible as are several microtubules. x 65 000. Frc. 9. Microtubule proliferation shortly be- fore nuclear envelope breakdown. At this stage the two pairs of flagellar bases are anterior to and on either side of the nucleus. The enormous numbers of microtubules running from the bases are visible. One flagellar base is visible and the others are just out of the plane of section. The nuclear envelope appears to be deformed by the microtubules. x 44 000.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
OAKLEY A N D BISALPUTRA
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
2794 CAN. J. BOT. VOL. 55. 1977
numbers of microtubules which extend from the 0.7 pm thick and has a number of chromatin- poles and some of these extend past the equator free tunnels of variable diameter (usually 0.1 pm and overlap with microtubules from the other or less) running through it. These tunnels con- pole. There are chromosomal microtubules tain the pole-to-pole and interdigitating micro- (Fig. 14) which extend poleward from protru- tubules. sions of the chromatin. We will term the point Anaphase of attachment of the microtubule to the chro- At anaphase (Fig. 17) the chromatin separates matin a kinetochore even though there is no in two disk-shaped masses each about 0.4 pm obvious differentiation of the chromatin at thick. The masses move to opposite poles as this point. There are also free microtubules the spindle elongates and narrows. The pole- which are scattered through the spindle with to-pole distance reaches about 4 pm, the no obvious connection to anything. chromosome-to-pole distance reduces to nothing
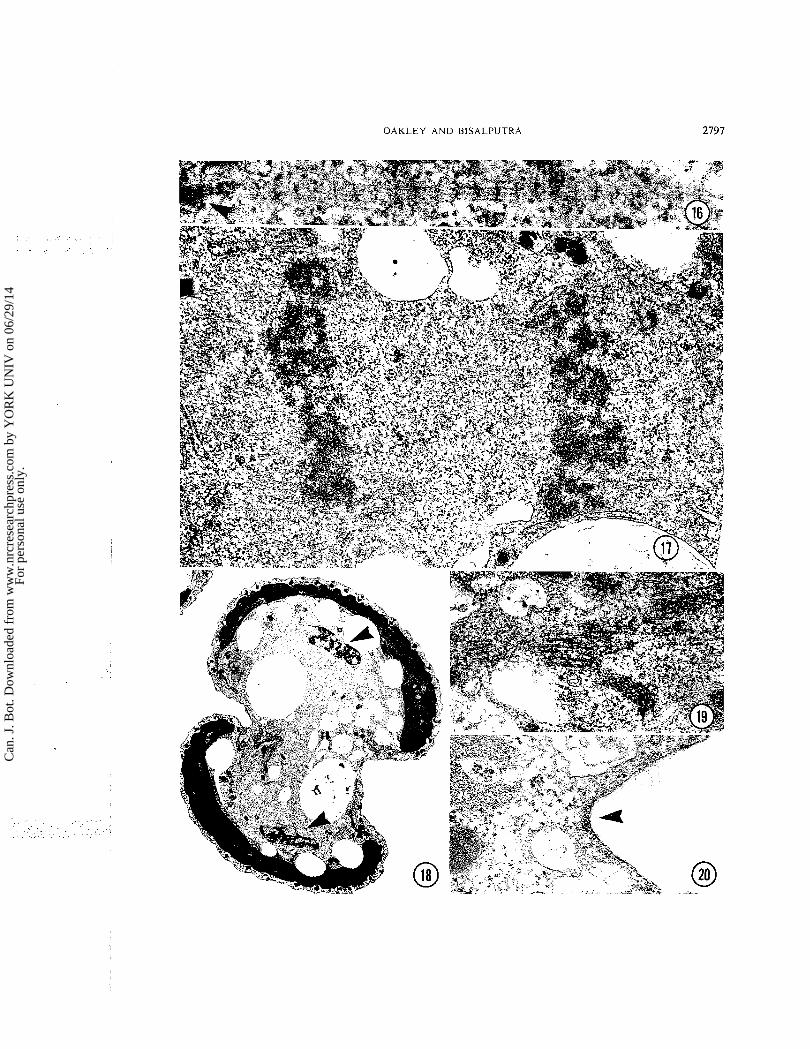
In addition to the microtubules there is a and the diameter of the spindle reduces to about rhizoplast in each polar region (Fig. 16). This 1.3 pm. The flagellar base pairs move along structure is seldom seen if the cells are fixed anterior to and just poleward of the moving normally. It has been seen in only 3 or 4 cells chromatin masses. out of the 100 or more cells we have examined in detail and even when it is present it is barely Telophase
visible. In cells treated with phalloidin, however, After the two plates of chromatin stop moving
it is almost always present and is much better (Fig. they round and the preserved. It is striated like the flagellar roots begins to re-form. This formation be-
and one extends from each of the pairs of gins before all the microtubules break down. In
flagellar bases along the polar region of the particular a number of microtubules remain in
metaphase spindle. Although the rhizoplasts may the region between the daughter "'lei and a'-
be quite long they are only about b0 nm wide though they are diminished greatly in number in
and 20 nm thick. They are never seen in direct cOm~ar i son with ana~hase may be quite
contact with microtubules and are often some long and even pack together to form a structure
distance away from them. They remain in the to degree a midbody (Fig. 19).
polar areas through telophase so they must move This is probably a highly transient because
along as the spindle elongates. We are unable to cells slightly further along in the cytokinetic
ascribe any probable function to them. process show few microtubules in this area.
The chromatin itself is in a disk-shaped mass Cytokinesis at metaphase. It occupies the entire equatorial The formation of the cytokinetic ring almost area of the spindle and so its diameter is roughly always begins by the time the metaphase plate the same as that of the spindle. [t is about has formed (Fig. 13). It pinches in steadily and
FIGS. 10-12. Light and electron n~icrographs of the same prometaphase cell. Fig. 10. Interference contrast micrograph of the cell embedded in resin showing the cell shape. x 2700. Fig. 11. Low-power electron micrograph showing the position and orientation of the spindle within the cell and the posi- tion of the other organelles. x 13 000. Fig. 12. A higher power micrograph showing the shape of the spindle and the distribution of the chromatin. Note the pair of flagellar bases above and slightly to the left of the chromatin. There is another pair to the right and out of the plane of section. x 34 000.
FIG. 13. A low power of a metaphase cell showing the cell shape and the fact that the cytokinetic furrow has formed (arrowhead). x 7800. FIG. 14. A portion of a metaphase spindle showing the point of attachment (arrowhead) of a microtubule to the chromatin (kinetochore). x 69 000. FIG. 15. A metaphase spindle showing the shape of the spindle and the tight mass of chromatin penetrated by regions of nucleoplasm containing microtubules. Serial sectioning reveals these regions to be tunnels. Note also the position of the pair of flagellar bases anterior to and slightly to the right of the chromatin. x 35 000.
FIG. 16. A rhizoplast in a phalloidin treated cell. The fuzzy area on the left (arrowhead) is associated with a flagellar base. x 93 000. FIG. 17. Anaphase. Note the two tight masses of chromatin. x 41 000. FIG. 18. Telophase. Note the shape of the cell and the position of the re-forming nuclei (arrowheads). Cytokinesis is quite advanced at this stage. x 8300. FIG. 19. The midbody-like arrangement of micro- tubules found between the re-forming nuclei of the cell shown in Fig. 18. x 39 000. FIG. 20. The amorphous fuzz (arrowhead) beneath the plasmalemma in a cytokinetic fi~rrow. x 70 000.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
O A K L E Y A N D B l S A L P U T R A 2795
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
CAN. J. BOT. VOL. 5 5 . 1977
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
O A K L E Y A N D RISALI'UTRA 2797
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
2798 C A N . J . DOT.
Pascher 1913; Butcher 1967). Our results show
FIG. 21. A lower power of the rhizoplast (arrowhead) seen in Fig. 16 showing its position in relation to the chromatin mass (c). The spindle axis runs from left to right and a few spindle microtubules are visible in as- sociation with the chromatin although they are not as clear in this phalloidin-treated material as in normally fixed material. It is clear that the rhizoplast is in the general polar region but does not contact the spindle microtubules. x 43 000.
cytokinesis is completed a t late telophase as the daughter nuclei begin to regain their interphase appearance. No microtubules appear to be directly involved but there is a thin layer of amorphous material (Fig. 20) just under the plasma membrane in the ring.
Discussion The structures and sequence of events in
mitosis in Cryptomonas sp. have some fairly important implications for the mechanisms of mitosis per se. These, however, will be discussed a t length elsewhere along with the fine structure of the spindle (Oakley and Heath 1976, and in preparation). We will confine our remarks to the taxonomic and phylogenetic implications of these findings.
It is worth noting that various authors use numbers of rows of gullet trichocysts and cell size as diagnostic taxonomic criteria (e.g.
that these two parameters change significantly before and during the course of cell division. When this is coupled with the observation that Cryptomonas sp. synchronizes itself in condi- tions very much like those which must be en- countered in its natural habitat (25°C and 13 h light and 1 1 h dark must be fairly typical of late spring in the gulf stream) it calls into ques- tion the applicability of these criteria. A collec- tion made a t dawn in the wild would presumably have a large percentage of 6-pm cells and collections made later in the day would have larger cells. Cells collected late a t night might likewise have more rows of trichocysts than those collected during the day. Thus the same organism might be identified as three or- ganisms. In view of the fact that a naturaI division synchrony is not unusual in marine planktonic organisms (Hastings and Sweeney 1964) perhaps it is advisable to use narrow size ranges as diagnostic criteria only with extreme caution.
Cytokinesis in Cryptomonas sp. appears to be like that of Cl7roornonas salina (Oakley and Dodge 1976) but since cytokinesis was not studied very completely in CIzroomonas we will go into some detail here. Cytokinesis in C~.ypto- rnonas appears to involve a contractile ring and thus would appear to resemble that of animal cells. It is possible that the fuzzy material found in the cytokinetic ring of Cryptomowas is actin as in higher animal cells (reviewed by Schroeder 1975). Attempts to verify this have been in- conclusive because normal glycerination pro- cedures used for heavy meromyosin labelling experiments (Forer and Behnke 1972) damage Cryptonlonas so severely that it is very difficult to select dividing cells and even if dividing cells were found it would be very difficult to rule out the possibility that actin was moved from some other part of the cell to the cytokinetic furrow during the glycerination process.
Cytokinesis in Cryptomonas is rather unusual among the algae. In most types of algae which have been studied there is an evident involve- ment of microtubules either in orienting or motivating cytokinesis (reviewed by Pickett- Heaps 1972a). Neither role is likely in Crypto- monas because microtubules are absent from the area of the cytokinetic ring except for a brief period in telophase. Moreover, the cyto- kinetic furrow is formed at early metaphase and cleavage is finished after most if not all mitotic
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
O A K L E Y A N D
microtubules have broken down. Since cyto- kinesis proceeds while microtubules are in various configurations it is unlikely that they are directly involved. Perhaps the alga which has a cytokinetic process most similar to that of Cryptomonas is the unicellular red alga Por- phyridium (Gantt and Conti 1965). Cytokinesis has been studied very incompletely in this organism, however, and most conclusions would be premature. Moreover, cytokinesis would ap- pear to be somewhat different in the filamentous red alga Membranoptera (McDonald 1972) and so phylogenetic affinities between the crypto- phytes and the red algae on the basis of cyto- kinetic similarity would be very tenuous.
The structures and events in the mitosis of Cryptomonas sp. resemble very closely those of Clzroomonas .salina. There are several features which tend to show their similarity while dis- tinguishing them from other algae. Among these are spindle shape, chromatin configura- tion, prophase nuclear movement, and prolifera- tion of microtubules from, and positioning of, flagellar bases. There are some differences between this account and the previous one on Clzroomonas but these are due in general to a change in methodology (e.g. the rhizoplast found after phalloidin treatment), a change in ter- minology (e.g. the connections between chro- matin and chromosomal microtubules are called kinetochores in this study but not in the previous studies although they are identical in appearance) or to the fact that we simply looked at more cells in this study and found structures we may have missed previously (a possible example being the midbody-like structure). The similarity in mitosis in Chroomonas and Cryptomonas cer- tainly suggests but does not prove that mitosis is similar throughout the class.
Since mitosis in Cryptomonas is essentially similar to that of Chroomonas the phylogenetic implications are virtually the same. For this reason we refer readers to the analysis of phylogenetic relationships made by Oakley and Dodge (1976) and will only summarize it here. There are a number of unicells of various classes which have mitotic apparatuses similar but not identical to that of Cryptomonas. Some examples of these are Pyramimonas (Pearson and Norris 1975), Prymnesium (Manton 1964), Cyanophora (Pickett-Heaps 19726) and Ocliromonas (Slankis and Gibbs 1972). It is probably not valid, how- ever, to postulate exact phylogenetic relation- ships between Cryptomonas and any of these
because it has not been established that the rather subtle differences in mitotic structures among these organisms are very useful phylo- genetic criteria. What we can argue with more certainty is that there is a great deal of dif- ference between cryptophycean mitosis and that of the red algae (McDonald 1972) and dino- flagellates (Leadbeater and Dodge 1967 ; Kubai and Ris 1969; Soyer 1969; Oakley and Dodge 1974) which have been studied. These dif- ferences, in turn, suggest that the cryptophytes are not closely related to either the dinoflagellates or the red algae.
The mitotic system of Cryptomonas sp. also shows few obviously primitive characteristics. One can make a case that extranuclear mitotic apparatuses such as are found in dinoflagellates are primitive (Kubai and Ris 1969 ; Kubai 1975) or that completely intranuclear spindles such as are found in yeasts are primitive (Pickett-Heaps 1969) but it would be very difficult to construct an internally consistent argument that crypto- phycean mitosis is primitive because it shares a number of characteristics with that of higher organisms, particularly higher plants. Some of these shared characteristics are as follows: break- down of the nuclear envelope in prophase, similar spindle geometry, a lengthening of the pole-to-pole distance as well as a shortening of the chromosome-to-pole distance in anaphase, and similar types of spindle microtubules.
Maltaux and Massart (1906) observed that the entire mitotic process from flagellar replication to the end of cytokinesis took only 15 min at 22°C in Chilomonas, another cryptophyte. Their result is consistent with ours and together these results point out that cryptophyte mitosis is very quick. In fact Maltaux and Massart point out that in Chilomonas at 35°C the entire mitotic process takes only 5 min, which is probably the fastest recorded time for mitosis in any organism. Thus if cryptophyte mitosis is primitive it is certainly not inefficient.
Cryptophycean mitosis differs from that of higher organisms in a t least two ways. The first is that cryptophytes lack the elaborate, mor- phologically differentiated kinetochores found in higher organisms. The second is that the chromatin in the cryptophytes always appears to be interconnected so that it is impossible to distinguish individual chromosomes. It is dif- ficult to determine whether these characteristics are primitive. Tightly packed chromatin plates are not uncommon in algae, particularly small
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/2
9/14
For
pers
onal
use
onl
y.
2800 CAN. 1. BOT. VOL. 5 5 . 1977
flagellates such as Chlamydomonas (Johnson and and Porter 1968), Ochromonas (Slankis and Gibbs 1972), and Pyramimonas (Pearson and Norris 1975), so this feature may not be partic- ularly significant. Perhaps it is fair to say that cryptophyte mitosis might be primitive in rela- tion to higher organisms but that there is little evidence to suggest that it is primitive in relation to other algae.
Acknowledgements The authors acknowledge the excellent sec-
retarial assistance of Mrs. Dorothy Gunning. This work was supported by National Research Council of Canada grants to the second author and to I.B. Heath. BUTCHER, R. W. 1967. An introductory account of the
smaller algae of British coastal waters. IV. Cryp- tophyceae. Fish. Invest. Minist. Agric. Fish. Food (G.B.) Ser. IV.
CAVALIER-SMITH, T . 1975. The origin of nuclei and of eukaryotic cells. Nature (London). 256: 463-468.
DODGE. J . D. 1965. Chromosome structure in the di- noflagellates and the problem of the mesocaryotic cell. Prog. Protozool. Proc. Int. Congr. Protozool. pp. 264-265.
1969. The ultrastructure of Cl7roornonrrs tneso- srig~rrrrrcr Butcher (Cryptophyceae). Arch. Mikrobiol. 69: 266-280.
DUPRAW, E. J . 1970. DNA and chromosomes. Chap. 6. Holt, Rinehart and Winston, Inc., New York.
FORER, A., and 0 . BEHNKE. 1972. An actin-like compo- nent in spermatocytes of a cranefly (Nephrororna slrr1rrn1i.s Loew). I . The spindle. Chrornosoma. 39: 175-190.
GANTT, E. 1971. Micromorphology of the periplast of Chrootnotlcls sp. (Cryptophyceae). J . Phycol. 7: 177- 184.
GANTT, E., and S. F. CONTI. 1965. The ultrastructure of Porphyridi~~tn ulrenrlrrn. J. Cell Biol. 26: 365-38 I.
GUILLARD. R. R. L., and J. H. RYTHER. 1962. Studies on marine planktonic diatoms. 1. Cyclorello nono Hustedt and Deron~rlo confervrrcer~ (Cleve) Gran. Can. J. Mi- crobiol. 8: 229-239.
HAS~INGS, J . W., and B. M. SWEENY. 1964. Phased cell division in the marine dinoflagellates. 117 Synchrony in cell division and growth. Erlired by E. Zeuthen. Intersci- ence Publishers, New York.
H~BBERD,D.J . , A. D. GREENVJOOD, and H. B.GRIFFITHS. 1971. Observations on the ultrastructure of the flagella and periplast in the Cryptophyceae. Br. Phycol. J . 6: 61-72.
JOHNSON, U . G., and K. R. PORTER. 1968. Fine structure of cell division in Cl~ln~nyrlornonns reinhordi, basal bodies and microtubules. J. Cell Biol. 38: 403-425.
KUBAI , D. F. 1975. The evolution of the mitotic spindle. Int. Rev. Cytol. 43: 167-227.
KUBAI , D. F., and H. RIS. 1969. Division in the dinoflagel- late Gyrodinillm cohnii (Schiller), a new type of nuclear reproduction. J . Cell Biol. 40: 508-528.
LEADBEATER, B., and J. D. DODGE. 1967. An electron microscope study of nuclear and cell division in a di- noflagellate. Arch. Mikrobiol. 57: 239-254.
LEE. R. E. 1972. Origin of plastids and the phylogeny ofthe algae. Nature (London), 237: 44-46.
LOEBLICH. A. R. 111. 1975. A seawater medium for di- noflagellates and the nutrition of Crrchoninn niei. J. Phycol. 11: 80-86.
L u c ~ s , I . A. N. 19700. Observations on the fine structure of the Cryptophyceae. I. The genus Cryptornonos. J. Phycol. 6: 30-38.
1970b. Observations on the ultrastructure of rep- resentatives of the genus Hetniseltnis and Chrootnotlns (Cryptophyceae). Br. Phycol. J. 5: 29-37.
MCDONALD, K. 1972. The ultrastructure of mitosis in the marine red alga Mernbrrrtzoprerrrploryphyllrr. J. Phycol. 8: 156-166.
MALTAUX, M.. and J. MASSAR.~. 1906. Sur les excitants de la division cellulaire. Rec. Inst. Bot. Leo Errera. Brux. 6: 369-42 1.
MANTON. I. 1964. Observations with the electron micro- scope on the division cycle in the flagellate Prymtlesillm prrrvlrrn Carter. J. R. Microsc. Soc. 83: 317-325.
OAKLEY. B. R.. and J . D. DODGE. 1973. Mitosis in the Cryptophyceae. Nature (London), 244: 521-522.
1974. Kinetochores associated with the nuclear envelope in the mitosis of a dinoflagellate. J. Cell Biol. 63: 322-325.
1976. The ultrastructure of mitosis in Cllroornot~os scrlit~a (Cryptophyceae). Protoplasma, 88: 24 1-254.
OAKLEY, B. R., and I. B. HEATH. 1976. An analysis of the metaphase spindle of Cryptotnonrrs. Can. Fed. Biol. SOC. 19: 129.
PASCHER, A. 1913. Cryptomonadineae. It1 Die Siiss- wasserflora Deutschlands. Osterreichs und der Schweiz. 11-Flngello~oe. Ediredhy A. Pascher. G. Fischer, Jena. pp96-114.
PEARSON, B. R., and R. E. NORRIS. 1975. Fine structure of cell division in Pyrotnbnotzns prrr.kenc. Norris and Pear- son (Chlorophyta, Prasinophyceae). J . Phycol. 11: 113-124.
PICKETT-HEAPS. J. D. 1969. The evolution of the mitotic apparatus: an attempt at comparative u~trastructural cytology in dividing plant cells. Cytobios, 3: 257-280.
19720. Variation in mitosis and cytokinesis in plant cells: its significance in the phylogeny and evolution of ultrastructural systems. Cytobios, 5: 59-77.
19726. Cell division in Cynnophorclpnrr~cloxtr. New Phytol. 71: 561-567.
RAVEN. P. H. 1970. A multiple origin for plastids and mitochondria. Science, 169: 641-646.
SCAGEL, R. F., R. J . BANDONI, G. E. ROUSE, W. B. SCHOFIELD, J. R. STEIN. and T . M. C. TAYLOR. 1965. An evolutionary survey of the plant kingdom. Wadsworth Publishing Company, Inc., Belmont, California. p. 502.
SCHROEDER, T. E. 1975. Dynamics of the contractile ring. In Molecules and cell movement. Ediredby S. Inoue and R. E. Stevens. Raven Press. New York. pp 305-334.
SLANKIS, T., and S. P. GIBBS. 1972. The fine structure of mitosis and cell division in the chrysophycean alga Och- ronloncrs donica. J. Phycol. 8: 243-256.
SOYER. M.-0. 1969. Rapports existant entre chromosomes et membrane nucleaire chez un dinoflagelle parasite du genre Blosrodinilr~r~ Chatton. C.R. Acad. Sci. Ser. D, 268: 2082-2084.
TAYLOR, D. L., and C. C. LEE. 1971. A new cryptomonad from Antarctica: Crypro~rlo~lns oyophylo sp.nov. Arch. Mikrobiol. 75: 269-280.